
Submitted:
05 September 2023
Posted:
07 September 2023
You are already at the latest version
Abstract
The genome sequencing technologies reveal the molecular mechanisms of differentiated thyroid cancer (DTC). Unlike somatic mutation analysis from thyroidectomy samples, germline mutations showing genetic susceptibility to DTC are less understood. The study aimed to assess the prevalence of germline mutations predisposing to DTC in a cohort of Polish individuals based on their whole genome sequencing (WGS) data. We analyzed sequencing data from 1076 unrelated individuals, released openly for academic and clinical research as The Thousand Polish Genomes database (https://1000polishgenomes.com). The list of genes chosen for further analysis was based on the review of previous studies. The cohort contained 104 variants located within the coding and noncoding DNA sequences of 90 genes selected by ClinVar classification as pathogenic and potentially pathogenic. The frequency of variants in the Polish cohort (our study) was compared to the frequency estimated for the non-Finnish European population, obtained from the gnomAD database (gnomad.broadinstitute.org). Statistically significant variants included 23 genes. Even though the Polish population is genetically similar to other European populations, there are significant differences in variant frequencies contributing to the disease development and progression, such as RET, CHECK2, BRCA1, SLC26A4 or TERT. Further studies are needed to identify genomic variants associated directly with DTC.
Keywords:
thyroid cancer
; whole-genome sequencing
; non-medullary thyroid cancer
; germline
; genetics
1. Introduction
Thyroid cancer is the most common malignant endocrine tumor [1]. It accounts for 1-2% of all neoplasms [2]. Its prevalence is steadily and the most rapidly rising among all cancers [3]. This increasing trend may be partly linked to an improved detection of smaller (< 2 cm) tumors thanks to more frequent and better ultrasound detection, fine-needle aspiration biopsies and the increased pathological reporting of incidental microcarcinomas [4,5]. In 2020, the World Health Organization (Global Cancer Statistics 2020: GLOBOCAN) reported 586202 new cases of thyroid cancer, of which 43646 resulted in death [6].
Thyroid cancer types are classified according to their histological characteristics (Figure 1) [7]. Differentiated thyroid cancer (DTC) is also known as non-medullary thyroid cancer (NMTC) and represents approximately 90% of all thyroid cancers [8]. Over 90% of thyroid cancer is sporadic, due to somatic genetic alterations [9]. Only 3-9% of all thyroid cancers are familial non-medullary thyroid cancer (FNMTC) cases, defined by the presence of thyroid cancer in 2 or more first-degree relatives, in the absence of predisposing environmental factors [10]. FNMTC can be divided into syndromic or non-syndromic FNMTC. It depends on whether the thyroid cancer is a part of one of many constellations of tumors (syndromic) or is the primary cancer (non-syndromic FNMTC) [11]. Out of all FNMTCs, only 5% in the syndromic form have well-defined driver germline mutations. On the contrary, 95% of FNMTC is non-syndromic with less well-defined genetic susceptibility [11].
In the last 30 years, the availability of the genome sequence has enabled progress in elucidating the molecular mechanisms underlying thyroid cancer. However, most studies involved somatic mutations analysis from fresh frozen paraffin embedded samples from thyroidectomy [13]. Driver mutations that promote cancer development were identified in over 90% of TC [14], but germline mutations showing genetic susceptibility to differentiated thyroid cancer are less studied. For the latter, genetic studies are vital. Despite the completion of the Human Genome Project [15], the information on the full spectrum of the human genetic variation remains incomplete [16]. Open, population-scale databases of human genetic variation are important for clinical genetics, biomedical research, prioritizing and tailoring genetic screening programs or improving guidelines for genetic counselling [17].
The high-quality of the sequencing data enabled the building of a unique repository of genetic variation in the Polish population, released publicly as the Thousand Polish Genomes database [17]. This database includes small and structural variants, runs of homozygosity, mitochondrial haplogroups, and novel variants identified in genomes of 1076 Poles.
The genetic background predisposing to differentiated thyroid cancer may differ among populations. To our best knowledge, there is no analysis including the Slavic population - accounting for over 4.5% of the world’s inhabitants. The Polish population, which is homogenous and sedentary in its nature but influenced by many migrations of the past, is not unique and very similar to many other European populations, therefore can serve as a genetic reference for the Slavic populations as long as there are no broader studies in other countries [17].
The risk of thyroid cancer inheritance may be 8- to 12-fold higher for first-degree relatives compared to the general population [18,19]. It makes thyroid cancer one of the most heritable cancers, displaying Mendelian inheritance [12]. The comparison of the aggressiveness of the disease in FNMTC patients compared to sporadic cases brings ambiguous results [12]. It may result in a more aggressive disease course at a younger age, with larger tumors and more lymph node involvement [20,21,22]. FNMTC may also express clinical anticipation with presentation at a younger age, with more severe symptoms at the second generation (genetic anticipation) [23]. Early diagnosis based on screening enables identification of thyroid cancer tumors of smaller size with less lymph node metastases and thus requiring less extensive treatments, potentially improving the treatment outcome [24]. The penetrant mutations in susceptibility genes to FMNTC could be vital for identifying at-risk individuals, thereby making early diagnosis, and selecting appropriate treatment possible [12]. If there exists genetic heterogeneity in the risk assigned to particular susceptibility genes or/and significant differences in risk allele frequencies, genetic screening should be matched to the relevant population. Therefore, the aim of the study was to assess the prevalence of germline mutations predisposing to the differentiated thyroid cancer (FMNTC) in a cohort of Polish individuals and compare prevalence of these mutations to the non-Finnish European population.
2. Materials and Methods
2.1. Gene Search Strategy
The first step was to define the list of genes, which germline mutations have been already linked to increased risk of developing differentiated thyroid cancer. The search strategy included Medical Subject Headings terms and keywords: “familial” OR “hereditary” AND “non-medullary thyroid cancer”. Reference lists of all the selected articles, previous meta-analyses, and reviews were hand-searched for any additional articles. We included studies, regardless of their sample size, with the investigation of the association between germline mutations and differentiated thyroid cancer occurrence. We carried out the systematic review following the guidelines formulated in the Cochrane Handbook for Systematic Reviews of Interventions [25] and the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) guidelines [26]. We searched PubMed, MEDLINE, Academic Search Complete, CINAHL Complete, CINAHL, Scopus, Cochrane, Health Source: Nursing/Academic Edition, Web of Knowledge, MasterFILE Premier, Health Source-Consumer Edition, Agricola, Dentistry and Oral Science Source databases from January 2006 up to January 2023 to find all relevant, full-text journal articles written in English.
2.2. Data Extraction and methodology assessment
Two authors (MBo and MBr) independently selected publications, which fulfilled the inclusion criteria specified above and extracted data for the outcomes using a standardized data extraction form. The risk of bias in the included studies was independently assessed based on the Cochrane risk of bias tool [27]. All included studies were assessed using the Newcastle-Ottawa Scale [28]. Only studies rated with at least seven stars were included for further consideration. Abstracts/papers focused on somatic mutations were excluded. The final list of NMTC susceptibility genes was identified and divided into genes related to syndromic and nonsyndromic FNMTC.
2.3. Genetic study
The cohort analyzed in this study consisted of 1076 unrelated individuals of Polish origin. The median age of participants was 45.4 years, with predominance of males (697 vs. 525). The analysis of clinical data showed that the most common chronic diseases reported by the participants were hypertension (13.0%), cancer (4.6%), diabetes (4.0%), and hypothyroidism or Hashimoto’s disease (3.0%). No health problems (excluding COVID-19 infection) were reported by 86% of participants. The sequence data encompassed over 1018 billion read pairs, yielding an average 35.26× read depth per genome. In every sample, over 91% of the reference genome was covered with at least 10 reads. The allelic frequencies of small and structural variants identified in this cohort were released openly for academic and clinical research as the Thousand Polish Genomes database (POL) [17].
The list of genes selected for further analysis (Figure 2 and Table 1) was based on the review of previous studies and included 90 genes involved in the development of DTC. The variants were annotated using the following resources: Ensemble Variant Effect Predictor v.108 [29], including references to databases of genomic variants from ClinVar v. 20221224 [30], and dbSNP build 154 [31], variant population frequencies from the 1000 Genomes Project, and gnomAD v2.0.1 and v3.0, as well as pathogenicity scores, such as Polyphen-2, SIFT and CADD [32]. All gene coordinates were padded with variants in the 10 kb range at both ends of the genes. The analyzed set contained 165736 variants in 90 genes. Following the ClinVar classification, we filtered pathogenic and potentially pathogenic variants for further analysis. The frequency of variants in the Polish cohort (our study) was compared to the frequency estimated for the non-Finnish European (NFE) population, obtained from the gnomAD database (accessed on 2023-01-19). Per gene cumulative allele frequencies were calculated and defined as the relative frequency of an allele (variant of a gene) at a particular locus in a particular gene in a population, expressed as a fraction or percentage [33].
Statistical significance of odds ratios for each variant was estimated with Fisher’s exact test and corrected for false discovery rate (FDR), where only variants with q-value < 0.05 were considered significant.
2.4. Rechecking data in genetic resources
Variants with significant differences in allele frequency among the Polish population (Thousand Polish Genomes database) and the non-Finnish European population (gnomAD) were clinically annotated using resources such as: ClinGen, NCCN guidelines, OMIM, Genetics Home Reference, GeneCards, ClinVar, and Gene-NCBI to describe their role and possible impact on DTC occurrence.
3. Results
Among the 90 genes considered, 19 were related to syndromic FNMTC (Table 1) and 71 were related to non-syndromic FNMTC. Following the ClinVar classification, out of the 165736 variants located in those genes, we selected pathogenic and potentially pathogenic ones for further analysis. The alleles frequencies of variants in the Polish cohort (our study) were compared to the frequencies estimated for the non-Finnish European population obtained from the gnomAD database. Among them, 23 had significantly different frequencies in the Polish population than in non-Finnish European: APC, ARSB, ATM, BRCA1, CHEK2, DICER1, GPD1L, INSR, KCNJ10, MYH9, PALB2, PLCB1, PLEKHG5, PTEN, RET, SEC23B, SERPINA1, SLC26A4, SMAD3, STK11, TERT, TOE1, and WRN. Most of those genetic variants are more frequent among Polish population, except from DICER1 and KCNJ10.
Among genes related to syndromic FNMTC (Table 1) those significantly characteristic for Polish population and with the highest OR were (Table 2, Table S1) APC (rs201375478; OR = 22.99), ATM (rs3092859; OR = 13.55), DICER1 (rs117358479; OR = 4.93), KCNJ10 (rs145947380; OR = 11.76), SEC23B (rs138198461; OR = 4.77), SLC26A4 (rs17154362; OR = 15.55), STK11 (rs587782259; OR = 24.31), WRN (rs4987238; OR = 5.65). Among genes related to nonsyndromic FNMTC (Table 1) the ones significantly characteristic for the Polish population and with the highest OR were ARSB (rs200040980; OR = 8.62), BRCA1 (rs80357087; OR = 17.02), CHEK2 (rs121908698; OR = 47,42), MYH9 (rs762239398; OR = 126,00), PALB2 (rs377085677; OR = 21.08), PTEN (rs180953647; OR = 14.47), RET (rs377767388) (OR = 50.59), TERT (rs377216965) (OR = 42.15). The genetic variants with an impact as modifiers according to ClinVar were: ARSB (rs72764913), ATM (rs879796523), DICER1 (rs1555368535), KCNJ10 (rs116418256, rs192835895, rs56656397), PALB2 (rs138200248), PCLB1 (rs532302075), PTEN (rs180953647), SERPINA1 (rs11558258), SLC26A4 (rs17154362), SMAD3 (rs958007552), STK11 (rs587782259), TOE1 (rs3219466).
4. Discussion
Between 5% and 15% of NMTC cases occur due to germline mutations [20]. Despite the mounting evidence for the heritability of thyroid cancer, at least partially, to date only a handful of genetic variants have been convincingly associated with a higher risk of this cancer [12]. As the studies have shown, the high heritability of thyroid cancer may occur likely due to the contributions of many rare but high-penetrance genetic variants in some cases, or common, low-penetrance variants in the other [66]. The first attempt to identify germline mutations with the risk for cancer was already done about ten years ago using GWAS technique, finding five single nucleotide polymorphisms (SNPs; rs965513, rs944289, rs966423, rs2439302, and rs116909374) to be associated with papillary thyroid carcinoma (PTC) in the cohort of Polish individuals [67]. Each of the variants showed highly significant but moderate to low disease risk, therefore the cumulative risk has been assessed and concluded that it is less significant than previously suggested, thus, the clinical use may not be feasible [67].
Among genes analyzed in our study, 19 were related to syndromic FNMTC and 71 were genes related to non-syndromic FNMTC. Within this group statistically significant variants included APC, ARSB, ATM, BRCA1, CHEK2, DICER1, GPD1L, INSR, KCNJ10, MYH9, PALB2, PLCB1, PLEKHG5, PTEN, RET, SEC23B, SERPINA1, SLC26A4, SMAD3, STK11, TERT, TOE1, WRN and indeed many of them have been already described as important in the initiation or progression and development of thyroid cancer. However, not many of them have ever been described in the context of population frequency. Most of those genetic variants are more frequent among Polish population (Thousand Polish Genomes database) than in non-Finnish European (gnomAD), except from DICER1 and KCNJ10. Below, we briefly discuss each of the genes with the perspective of thyroid cancer contribution and known prevalence in other populations.
APC gene variants are well known for their contribution to the familial adenomatous polyposis (FAP), Gardner syndrome, however papillary thyroid carcinoma (PTC) associated with FAP is very rare [68]. Even though in most cases papillary thyroid carcinoma associated with FAP occurs in females in their 30s and rarely in the elderly, there are several cases described where the disease occurred among patients in other age groups, as well as among members of the same family. As a rare extracolonic manifestation of the FAP, thyroid cancer, not solely PTC, is diagnosed in about 2.6% of all FAP patients globally, however the molecular background of this malignancy remains unknown [69,70]. The most significant variants occurring more frequently among Polish population (see Table S1) are missense variants of conflicting interpretation of pathogenicity. All of them have been described before and known to be cancer related.
The expression level of ARSB in the thyroid cancer tissue is low [71], however its significance in thyroid cancer development and the prevalence of its variants has been already reported among the Italian population [35,72,73]. In our cohort three variants reached statistical significance, however their prevalence is not very different from other analyzed populations in gnomAD. This gene has not been described profoundly in the context of thyroid cancer and may be an interesting novel candidate for further studies. The protein encoded by ARSB gene is located in the lysosome and it is known to participate in the regulation of cell adhesion, cell migration and invasion for example in colonic epithelium [74]. In the central nervous system, is a regulator of neurite outgrowth, as well as neuronal plasticity, acting through the control of sulphate glycosaminoglycans and neuronal levels (ARSB_HUMAN, P15848).
ATM gene is crucial when it comes to the response to ionizing radiation-induced DNA damage, and therefore it has been long suspected it might be involved in thyroid cancer [75]. The cBioPortal lists somatic ATM mutations in 2.4% of thyroid cancers [76]. Mutations of the ATM gene are responsible for ataxia telangiectasia, a rare inherited disorder characterized by progressive ataxia, but also radiosensitivity, many cell-cycle checkpoint defects, genome instability, and a predisposition to cancer of all types [75]. The expression of the gene in the thyroid is high, especially in glandular cells [77]. It has been reported that the individual susceptibility to DTC may be attributable to the genetic variants of the ATM gene [75]. However, little is known about the population prevalence of the ATM mutations among thyroid cancer patients, even if there are several interesting case reports describing this association [78]. In our cohort several variants of the ATM gene have been discovered as significantly more frequent, many of them with clearly pathogenic status (see Table S1).
BRCA1, similarly to the CHEK2 and PALB2 genes, are engaged in the DNA repair mechanisms, therefore absolutely crucial for cancer protection. In our cohort only 3 variants of this gene have been significantly more frequent than in gnomAD cohort, two of them are of pathogenic status and one missense variant of conflicting interpretation status. However, among CHEK2 gene variants all significant mutations are of conflicting interpretation of pathogenicity status and all of them appear to be more frequent among analyzed cohort. It has been suggested that CHEK2 mutations generally predispose to thyroid cancer, together with familial aggregations of breast and thyroid cancer and even to double primary cancers of the breast and thyroid, with several cases already described among the Polish population [42]. It plays a role in maintaining genomic stability, and it also acts as a tumor suppressor [79]. However, in the recent work of Cieszyńska et al. of all women with breast cancer analyzed in the study, only 0.49% developed a second case of primary thyroid cancer [80]. Interestingly, the ten-year risk of thyroid cancer development was higher in women who also carried a CHEK2 mutation (1.5%) than among women who carried no other mutations (0.9%) [80]. Among Polish females, according to the study, following a diagnosis of breast cancer, the age-specific annual incidence rates of thyroid cancer increased at least 4 times [80]. Some genes have already had extensive research done in the thyroid cancer perspective within the Polish population, just like aforementioned CHEK2. Pathogenic variants in this gene were reported in both PTC and FTC, although not with the same prevalence: about 15.6% of patients with PTC had mutations in CHEK2 gene [42,81]. Interestingly, CHEK2 mutation carriers reported a family history of breast cancer 2.2 times more frequently than non-carriers [42].
DICER1 gene mutations are crucial for DICER1 syndrome associated with familial pleuropulmonary blastoma), cystic nephroma and ovarian Sertoli-Leydig cell tumors [22]. Co-occurrence of DTC with Sertoli-Leydig cell tumor is highly suggestive of DICER1 syndrome [22]. Most patients with DICER1 syndrome diagnosed with DTC (PTC and FTC) had prior exposure to radiation and chemotherapy for the treatment of associated malignancy [22]. However, recently PTC in a patient without past medical history of oncological treatment was reported [43] and the risk of DTC was 16- to 24-times increased over a DICER1 patient’s lifetime [43]. In our study the frequency of DICER1 variants were like the comparator, except for one variant with slightly higher prevalence among Polish population, but not clearly pathogenic (see Table S1).
GPD1L mutations in patients with DTC were described in the Italian, Polish and Spanish populations [35], whereas INSR only in the Korean population [44]. In our cohort only one variant of the GPD1L gene occurs statistically more frequently than in the gnomAD control group, possessing the status of conflicting interpretation of pathogenicity. Similarly, INSR variants discovered in our study are more frequent among Polish cohort than in the gnomAD control group, all of which also possess the status of conflicting interpretation of pathogenicity (see Table S1).
KCNJ10 mutations were described in patients with Pendred syndrome, which is an autosomal recessive syndrome linked to sensorineural deafness/hearing impairment, goiter, and an abnormal organification of iodide with or without hypothyroidism, as well as to DTC [12]. Little is known about the frequency of its variants among other population, however in our cohort at least eight variants seem to be important (see Table S1). Interestingly, 5 of them are in the 3′UTR region of the gene.
PLEKHG5 germline variants in familial non-syndromic PTC (classical and follicular variant) were identified in a Brazilian population [50]. In our study one missense variant was found to be significantly more frequent among Polish population. Its protein product regulates autophagy of synaptic vesicles in axon terminal of motoneurons (by similarity). Involved in the control of neuronal cell differentiation [82]. Also plays a role in angiogenesis through regulation of endothelial cells chemotaxis [83].
MYH9 encodes myosin-9 and potentially impacts the risk of PTC by interacting with a long noncoding RNA (encoded by the PTCS2 gene) and FOXE1 gene [47]. Only one missense variant has been found to be relevantly more frequent among Polish cohort, even though it has the status of conflicting interpretation of pathogenicity. It may also be an interesting candidate gene for further studies on DTC. This gene is a novel cancer stem cell marker and claimed as a prognostic indicator in esophageal cancer that promotes oncogenesis through the PI3K/AKT/mTOR axis [84].
InDel intronic variant within PLCB1 was the first mutation identified in familial multiple papilloid adenomata-type DTC patients and in a subset of patients with sporadic DTC [85]. In patients who were carriers of this mutation, multinodular goiter progressed to PTC (follicular variant) through overexpression of phospholipase C beta 1 (PLCB1) [86]. Among Polish population three variants gained statistical significance with higher prevalence but having conflicting interpretation of pathogenicity.
PTEN hamartoma tumor syndrome (PHTS) embodies a group of diseases caused by germline mutations in the phosphatase and tensin homolog (PTEN) gene located at 10q23.31. They include Cowden syndrome, PTEN-related Proteus syndrome, Proteus-like syndrome, Bannayan-Riley-Ruvalcaba syndrome (BRRS), and adult Lhermitte-Duclos disease (LDD) [12]. Approximately 6 to 38% of PHTS patients develop thyroid cancer with a median age of diagnosis between 31 and 37 years [12]. A risk of DTC is 26 to 39 times higher than for individuals without a PTEN mutation [87]. In our study three variants located in the noncoding regions have been found as appearing more frequently among Polish population, however little is known about them in the context of DTC cancer.
Although RET gene mutations are crucial for medullary thyroid cancer, a germline RET oncogene mutation may be also linked to NMTC, especially as a part of FAP syndrome [34]. In our study several genetic variants of this gene have been found as being significantly more frequent among Poles, most of them are missense variants or located in the 3′UTR or 5′UTR regions.
SEC23B mutation was identified in patients with FTC as a component of the Cowden syndrome in a WGS study [56]. In a SEC23B pathogenic variant carriers, Yehia et al. found a significantly increased age-adjusted standardized incidence rate of DTC when compared with the general population [56]. Apart from DTC, the Cowden syndrome may cause hamartomas, breast, kidney, colon, endometrium, and brain tumors, mucocutaneous lesions and macrocephaly [12]. Again, in our population discovered variants are more frequent, one of them possessing status of being pathogenic (see Table S1). SEC23B encodes Sec23 Homolog B, a component of coat protein complex II (COPII). It is involved in vesicle trafficking, responsible for transporting proteins from the endoplasmic reticulum to the Golgi apparatus [56].
SERPINA1 (α1-AntiTrypsin) located at 14q32.13, is highly expressed in colorectal cancer, cutaneous squamous cell carcinoma, and papillary thyroid carcinoma [50]. The role of this gene in thyroid tumorigenesis is not fully understood. However, in colorectal cancer cell invasion and migration have been associated with SERPINA1 upregulation of fibronectin. Mutations in this gene may also present a mitogenic action, stimulating malignant cell proliferation [50]. One variant of this gene located in the 5′UTR region has been found as statistically more prevalent among Polish population.
Among the Korean population, it has been reported that SLC24A6 gene, which encodes a mitochondrial sodium and calcium ion exchanger, is important for the risk of developing thyroid cancer [44,88]. However, its relationship to DTC pathogenesis is not established yet, even though an interesting finding was the stronger genetic variant association with FTC rather than PTC in that Korean study [44,88]. This striking finding suggested that at least some genetic markers of susceptibility to DTC are different between thyroid cancer types. In our study many variants seem to be more frequent among analyzed population, most of which are missense variants of conflicting interpretation of pathogenicity, whereas one discovered variant is likely pathogenic as a frameshift mutation.
The SMAD family member 3 gene (SMAD3) show higher expression in the thyroid than in most other tissues. It supports a potential role for this factor in predisposition to thyroid cancer [8]. SMAD3 is an important transcriptional mediator of transforming growth factor-b (TGF-b) signaling associated with PTC [12,59]. Wang et al. investigated the downstream mechanisms by which alterations of SMAD3 contribute to thyroid cancer susceptibility [89]. Only two variants of conflicting interpretation of pathogenicity have been found in this study as being slightly more prevalent among Polish cohort.
Mutation in serine/threonine kinase 11 (STK11) gene is a causative agent for Peutz-Jeghers Syndrome (PJS). It is an autosomal dominant syndrome characterized by hamartomatous polyps and mucocutaneous hyperpigmentation with a 4-fold increase in cancer risk (regardless of a primary side) as compared to the general population [12]. The increased cancer risk has been connected to the P53 pathway [90]. PJS has been associated with multiple cases of thyroid cancer of PTC, FTC, tall cell variant PTC, and follicular variant PTC subtypes [12,62]. Only one intronic variant has been significantly more frequent among Polish cohort and it is in the intronic region of the gene, thus, may possess a regulatory function.
TOE1 gene product inhibits cell growth rate and cell cycle: it induces CDKN1A gene expression as well as TGF-beta expression. Moreover, it also mediates the inhibitory growth effect of EGR1 [91]. TOE1 mutations were associated with PTC occurring in patients with FAP. They also suffered from colorectal carcinoma, ampullary carcinoma, hepatoblastoma, medulloblastoma, PTC did not affect their survival [92]. In our study one intronic variant seems to be slightly more prevalent among Polish population, compared with gnomAD cohort, although it has a conflicting interpretation of pathogenicity at this moment.
TERT gene encodes a protein which is an essential component of the telomere length maintenance complex and is weakly expressed in normal thyroid tissue. However, it is known to be reactivated in many human cancers, including thyroid cancer through the transcriptional regulation [88,93]. It is correlated with a more severe form of the cancer disease, and mutations in the TERT promoter in thyroid cancer were suggested as having a prognostic potential if coexisting with the BRAFV600E mutation [88,94]. In our study two variants of TERT gene have been found as occurring more frequently among Polish population, both with conflicting interpretation of pathogenicity.
Mutations in the WRN gene are associated with Werner syndrome [65,73]. It is a rare autosomal recessive disorder classified as one of the progeria syndromes, thus associated with premature ageing and increased risk for several malignancies, especially melanoma, meningioma, soft tissue sarcoma, leukemia, osteosarcoma, but also differentiated thyroid cancer, usually follicular carcinoma [65,73,95]. It may increase the cancer risk irrespective of their location. Several missense variants have been found to occur more frequently among our cohort than in a control gnomAD group, however all of them with conflicting interpretation of pathogenicity.
It is important to notice the significance of the population studies, since in every cohort there might be several founder variants typical for the population, absent or very rare in other groups. Due to the historical influences in many populations some genetic variants may be more prevalent, and this information is very useful for clinical geneticists and other healthcare professionals, since they may provide us with additional hints at the point of diagnosis: facing financial constraints in many countries genetic testing is still not very popular and analysis of selected variants or genes, instead of huge panels or WGS, is simply cheaper and faster, thus, more available.
Despite our best efforts, there are several limitations to the study. First, we decided to focus only on the protein-coding genes, since the knowledge about other elements of the genome, especially being involved in the thyroid cancer risk, is still in its infancy. In our study we decided to focus on protein-coding genes, since the knowledge on non-coding elements of the genome, being involved in the thyroid cancer risk, is still in its infancy. Only two studies indicate association of PTC with mutations within miRNA genes in the Polish population [73,96]. Secondly, a reliable comparison of allele frequency between populations suffers from a low number of available data. Even the gnomAD cohort contains collected data for participants from many different populations, but usually of a small sample size that does not allow for an accurate allele frequency estimation. Especially when it comes to the WGS data, there are still only a few large, populational databases. Despite interesting findings, it is important to stress that only carefully selected genes have been analyzed in our research. Even if the entire gene sequence was taken into consideration, still there might be other elements of the genome with equally important significance, or even contributing more to the regulation of gene expression. Also, epigenetic factors are known to be crucial in many cancer stages of development, therefore further studies are crucial to fully understand the thyroid cancer intricacies.
5. Conclusions
Despite the progress in elucidating the molecular mechanisms underlying differentiated thyroid cancer (DTC) thanks to genome sequencing, germline mutations responsible for genetic susceptibility to DTC are still poorly recognized. The aim of this study was to assess the prevalence of genetic germline mutations predisposing to the differentiated thyroid cancer in a cohort of Polish individuals using whole genome analysis to make a basis for further tailored prophylaxis. Undoubtedly more research is needed to provide a more extensive background on the penetrance, molecular function and functional consequences of the genetic variants presented here, which can further clarify the etiology of thyroid cancer and probably also support the identification of disease risk in family members of NMTC patients, not solely among Polish population, but serving as a hint for further populational studies. Finally, many studies describing the genetic predisposition to NMTC are case-control designs. What is needed are genome-wide association studies (GWAS), preferably based on next generation sequencing, and family-based exome sequencing. Genomic studies should be followed by transcriptome (e.g., Single-Cell RNA Sequencing) and/or molecular (droplet digital polymerase chain reaction (ddPCR) or Sanger sequencing) analyses, and linkage studies to identify and confirm the new susceptibility loci associated with NMTC [97].
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1: Genes with significant (q-value <0.05) differences in pathogenic variant burden among Polish population (Thousand Polish Genomes database) compared with non-Finnish European (gnomAD)
Author Contributions
Conceptualization, M.Bo. and P.D.; methodology, M.Bo., M.S., and P.D.; software, M.S.; validation, M.Bo., J.S., M.Br., M.O., and P.D.; formal analysis, M.Bo., M.S., and P.D.; investigation, M.Bo., M.S., and P.D.; resources, M.Bo., M.S., and P.D.; data curation, M.Bo., M.S., M.Br., M.O., and P.D.; writing—original draft preparation, M.Bo., M.S., and P.D.; writing—review and editing, J.S., M.Br., K.Z., M.R., M.O., Z.K.; visualization, M.Bo., M.S., and M.Br.; supervision, P.D.; project administration, M.B and P.D.; funding acquisition, M.R., Z.K, and P.D.
Funding
The dataset of the repository has been collected during the research partially funded by the Polish National Science Centre grant No. SZPITALE JEDNOIMIENNE/2/2020 and by the Medical Research Agency grant No 2020/ABM /COVID19/0022.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki, the highest data security standards 140 of ISO 27001 and the General Data Protection Regulation (GDPR) act and approved by the Ethics Committee of the Central Clinical Hospital of the Ministry of Interior and Administration in Warsaw (decision nr: 41/2020 from 03.04.2020 and 125/2020 from 1.07.2020).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
The idea for this study was raised in August 2022, bioinformatics analysis performed in autumn 2022 and the manuscript written in the first half of 2023. The datasets presented in this study can be found in an online repository: https://1000polishgenomes.com [access date: October 2022]. Full cohort description can be found in the following paper: https://doi.org/10.3390/ijms23094532
Acknowledgments
The Authors would like to thank all sample donors that participated in the study, as well as the medical personnel of the Central Clinical Hospital of the Ministry of the Interior and Administration in Warsaw for their active support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Wells, S.A. Progress in Endocrine Neoplasia. Clin Cancer Res 2016, 22, 4981–4988. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Li, H.; Wang, M.; Li, N.; Tian, T.; Wu, Y.; Xu, P.; Yang, S.; Zhai, Z.; Zhou, L.; et al. Global Burden of Thyroid Cancer From 1990 to 2017. JAMA Netw Open 2020, 3, e208759. [Google Scholar] [CrossRef] [PubMed]
- Davies, L.; Morris, L.G.T.; Haymart, M.; Chen, A.Y.; Goldenberg, D.; Morris, J.; Ogilvie, J.B.; Terris, D.J.; Netterville, J.; Wong, R.J.; et al. American Association of Clinical Endocrinologists and American College of Endocrinology Disease State Clinical Review: The Increasing Incidence of Thyroid Cancer. Endocrine Practice 2015, 21, 686–696. [Google Scholar] [CrossRef] [PubMed]
- Megwalu, U.C.; Moon, P.K. Thyroid Cancer Incidence and Mortality Trends in the United States: 2000-2018. Thyroid 2022, 32, 560–570. [Google Scholar] [CrossRef] [PubMed]
- Tofe, S.; Argüelles, I.; Forteza, A.; Alvarez, C.; Repetto, A.; Masmiquel, L.; Rodríguez, I.; Losada, E.; Sukunza, N.; Cabrer, M.; et al. Age-Standardized Incidence, Mortality Rate and Trend Changes of Thyroid Cancer in the Balearic Islands during the 2000-2020 Period: A Population-Based Study. Eur Thyroid J 2023, ETJ-22-0183. [Google Scholar] [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J Clin 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Vriens, M.R.; Suh, I.; Moses, W.; Kebebew, E. Clinical Features and Genetic Predisposition to Hereditary Nonmedullary Thyroid Cancer. Thyroid 2009, 19, 1343–1349. [Google Scholar] [CrossRef]
- Hińcza, K.; Kowalik, A.; Kowalska, A. Current Knowledge of Germline Genetic Risk Factors for the Development of Non-Medullary Thyroid Cancer. Genes 2019, 10, 482. [Google Scholar] [CrossRef]
- Borowczyk, M.; Szczepanek-Parulska, E.; Dębicki, S.; Budny, B.; Janicka-Jedyńska, M.; Gil, L.; Verburg, F.A.; Filipowicz, D.; Wrotkowska, E.; Majchrzycka, B.; et al. High Incidence of FLT3 Mutations in Follicular Thyroid Cancer: Potential Therapeutic Target in Patients with Advanced Disease Stage. Ther Adv Med Oncol 2020, 12, 1758835920907534. [Google Scholar] [CrossRef]
- Capezzone, M.; Robenshtok, E.; Cantara, S.; Castagna, M.G. Familial Non-Medullary Thyroid Cancer: A Critical Review. J Endocrinol Invest 2021, 44, 943–950. [Google Scholar] [CrossRef]
- Guilmette, J.; Nosé, V. Hereditary and Familial Thyroid Tumours. Histopathology 2018, 72, 70–81. [Google Scholar] [CrossRef]
- Kamani, T.; Charkhchi, P.; Zahedi, A.; Akbari, M.R. Genetic Susceptibility to Hereditary Non-Medullary Thyroid Cancer. Hered Cancer Clin Pract 2022, 20, 9. [Google Scholar] [CrossRef] [PubMed]
- Cancer Genome Atlas Research Network Integrated Genomic Characterization of Papillary Thyroid Carcinoma. Cell 2014, 159, 676–690. [CrossRef]
- Prete, A.; Borges de Souza, P.; Censi, S.; Muzza, M.; Nucci, N.; Sponziello, M. Update on Fundamental Mechanisms of Thyroid Cancer. Front Endocrinol (Lausanne) 2020, 11, 102. [Google Scholar] [CrossRef] [PubMed]
- 1000 Genomes Project Consortium; Auton, A.; Brooks, L.D.; Durbin, R.M.; Garrison, E.P.; Kang, H.M.; Korbel, J.O.; Marchini, J.L.; McCarthy, S.; McVean, G.A.; et al. A Global Reference for Human Genetic Variation. Nature 2015, 526, 68–74. [Google Scholar] [CrossRef]
- Popejoy, A.B.; Fullerton, S.M. Genomics Is Failing on Diversity. Nature 2016, 538, 161–164. [Google Scholar] [CrossRef]
- Kaja, E.; Lejman, A.; Sielski, D.; Sypniewski, M.; Gambin, T.; Dawidziuk, M.; Suchocki, T.; Golik, P.; Wojtaszewska, M.; Mroczek, M.; et al. The Thousand Polish Genomes-A Database of Polish Variant Allele Frequencies. Int J Mol Sci 2022, 23, 4532. [Google Scholar] [CrossRef]
- Bonora, E.; Tallini, G.; Romeo, G. Genetic Predisposition to Familial Nonmedullary Thyroid Cancer: An Update of Molecular Findings and State-of-the-Art Studies. J Oncol 2010, 2010, 385206. [Google Scholar] [CrossRef]
- Goldgar, D.E.; Easton, D.F.; Cannon-Albright, L.A.; Skolnick, M.H. Systematic Population-Based Assessment of Cancer Risk in First-Degree Relatives of Cancer Probands. J Natl Cancer Inst 1994, 86, 1600–1608. [Google Scholar] [CrossRef]
- Moses, W.; Weng, J.; Kebebew, E. Prevalence, Clinicopathologic Features, and Somatic Genetic Mutation Profile in Familial versus Sporadic Nonmedullary Thyroid Cancer. Thyroid 2011, 21, 367–371. [Google Scholar] [CrossRef]
- Mazeh, H.; Benavidez, J.; Poehls, J.L.; Youngwirth, L.; Chen, H.; Sippel, R.S. In Patients with Thyroid Cancer of Follicular Cell Origin, a Family History of Nonmedullary Thyroid Cancer in One First-Degree Relative Is Associated with More Aggressive Disease. Thyroid 2012, 22, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Peiling Yang, S.; Ngeow, J. Familial Non-Medullary Thyroid Cancer: Unraveling the Genetic Maze. Endocrine-Related Cancer 2016, 23, R577–R595. [Google Scholar] [CrossRef]
- Park, Y.J.; Ahn, H.Y.; Choi, H.S.; Kim, K.W.; Park, D.J.; Cho, B.Y. The Long-Term Outcomes of the Second Generation of Familial Nonmedullary Thyroid Carcinoma Are More Aggressive than Sporadic Cases. Thyroid 2012, 22, 356–362. [Google Scholar] [CrossRef]
- Klubo-Gwiezdzinska, J.; Yang, L.; Merkel, R.; Patel, D.; Nilubol, N.; Merino, M.J.; Skarulis, M.; Sadowski, S.M.; Kebebew, E. Results of Screening in Familial Non-Medullary Thyroid Cancer. Thyroid 2017, 27, 1017–1024. [Google Scholar] [CrossRef]
- Cumpston, M.; Li, T.; Page, M.J.; Chandler, J.; Welch, V.A.; Higgins, J.P.; Thomas, J. Updated Guidance for Trusted Systematic Reviews: A New Edition of the Cochrane Handbook for Systematic Reviews of Interventions. Cochrane Database Syst Rev 2019, 10, ED000142. [Google Scholar] [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 Statement: An Updated Guideline for Reporting Systematic Reviews. BMJ 2021, 372, n71. [Google Scholar] [CrossRef] [PubMed]
- Higgins, J.P.T.; Altman, D.G.; Gøtzsche, P.C.; Jüni, P.; Moher, D.; Oxman, A.D.; Savović, J.; Schulz, K.F.; Weeks, L.; Sterne, J.A.C. The Cochrane Collaboration’s Tool for Assessing Risk of Bias in Randomised Trials. BMJ 2011, 343, d5928. [Google Scholar] [CrossRef] [PubMed]
- Lo, C.K.-L.; Mertz, D.; Loeb, M. Newcastle-Ottawa Scale: Comparing Reviewers’ to Authors’ Assessments. BMC Med Res Methodol 2014, 14, 45. [Google Scholar] [CrossRef] [PubMed]
- McLaren, W.; Gil, L.; Hunt, S.E.; Riat, H.S.; Ritchie, G.R.S.; Thormann, A.; Flicek, P.; Cunningham, F. The Ensembl Variant Effect Predictor. Genome Biol 2016, 17, 122. [Google Scholar] [CrossRef]
- Landrum, M.J.; Lee, J.M.; Benson, M.; Brown, G.; Chao, C.; Chitipiralla, S.; Gu, B.; Hart, J.; Hoffman, D.; Hoover, J.; et al. ClinVar: Public Archive of Interpretations of Clinically Relevant Variants. Nucleic Acids Res 2016, 44, D862–868. [Google Scholar] [CrossRef]
- Sherry, S.T.; Ward, M.H.; Kholodov, M.; Baker, J.; Phan, L.; Smigielski, E.M.; Sirotkin, K. DbSNP: The NCBI Database of Genetic Variation. Nucleic Acids Res 2001, 29, 308–311. [Google Scholar] [CrossRef]
- Rentzsch, P.; Witten, D.; Cooper, G.M.; Shendure, J.; Kircher, M. CADD: Predicting the Deleteriousness of Variants throughout the Human Genome. Nucleic Acids Res 2019, 47, D886–D894. [Google Scholar] [CrossRef]
- Middleton, D.; Menchaca, L.; Rood, H.; Komerofsky, R. New Allele Frequency Database: Http://Www.Allelefrequencies.Net. Tissue Antigens 2003, 61, 403–407. [Google Scholar] [CrossRef]
- Cetta, F. FAP Associated Papillary Thyroid Carcinoma: A Peculiar Subtype of Familial Nonmedullary Thyroid Cancer. Patholog Res Int 2015, 2015, 309348. [Google Scholar] [CrossRef]
- Figlioli, G.; Köhler, A.; Chen, B.; Elisei, R.; Romei, C.; Cipollini, M.; Cristaudo, A.; Bambi, F.; Paolicchi, E.; Hoffmann, P.; et al. Novel Genome-Wide Association Study–Based Candidate Loci for Differentiated Thyroid Cancer Risk. The Journal of Clinical Endocrinology & Metabolism 2014, 99, E2084–E2092. [Google Scholar] [CrossRef]
- Dombernowsky, S.L.; Weischer, M.; Allin, K.H.; Bojesen, S.E.; Tybjaerg-Hansen, A.; Nordestgaard, B.G. Risk of Cancer by ATM Missense Mutations in the General Population. J Clin Oncol 2008, 26, 3057–3062. [Google Scholar] [CrossRef]
- Akulevich, N.M.; Saenko, V.A.; Rogounovitch, T.I.; Drozd, V.M.; Lushnikov, E.F.; Ivanov, V.K.; Mitsutake, N.; Kominami, R.; Yamashita, S. Polymorphisms of DNA Damage Response Genes in Radiation-Related and Sporadic Papillary Thyroid Carcinoma. Endocr Relat Cancer 2009, 16, 491–503. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Yu, Y.; Ai, L.; Shi, J.; Liu, X.; Sun, H.; Liu, Y. Association of the ATM Gene Polymorphisms with Papillary Thyroid Cancer. Endocrine 2014, 45, 454–461. [Google Scholar] [CrossRef]
- Wójcicka, A.; Czetwertyńska, M.; Świerniak, M.; Długosińska, J.; Maciąg, M.; Czajka, A.; Dymecka, K.; Kubiak, A.; Kot, A.; Płoski, R.; et al. Variants in the ATM-CHEK2-BRCA1 Axis Determine Genetic Predisposition and Clinical Presentation of Papillary Thyroid Carcinoma. Genes Chromosomes Cancer 2014, 53, 516–523. [Google Scholar] [CrossRef]
- Wang, G.; Li, Z.; Li, X.; Zhang, C.; Peng, L. RASAL1 Induces to Downregulate the SCD1, Leading to Suppression of Cell Proliferation in Colon Cancer via LXRα/SREBP1c Pathway. Biol Res 2019, 52, 60. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Singh, P.; Yin, K.; Wang, J.; Bao, Y.; Wu, M.; Pathak, K.; McKinley, S.K.; Braun, D.; Lubitz, C.C.; et al. Non-Medullary Thyroid Cancer Susceptibility Genes: Evidence and Disease Spectrum. Ann Surg Oncol 2021, 28, 6590–6600. [Google Scholar] [CrossRef]
- Siołek, M.; Cybulski, C.; Gąsior-Perczak, D.; Kowalik, A.; Kozak-Klonowska, B.; Kowalska, A.; Chłopek, M.; Kluźniak, W.; Wokołorczyk, D.; Pałyga, I.; et al. CHEK2 Mutations and the Risk of Papillary Thyroid Cancer. International Journal of Cancer 2015, 137, 548–552. [Google Scholar] [CrossRef]
- Rutter, M.M.; Jha, P.; Schultz, K.A.P.; Sheil, A.; Harris, A.K.; Bauer, A.J.; Field, A.L.; Geller, J.; Hill, D.A. DICER1 Mutations and Differentiated Thyroid Carcinoma: Evidence of a Direct Association. J Clin Endocrinol Metab 2016, 101, 1–5. [Google Scholar] [CrossRef]
- Son, H.-Y.; Hwangbo, Y.; Yoo, S.-K.; Im, S.-W.; Yang, S.D.; Kwak, S.-J.; Park, M.S.; Kwak, S.H.; Cho, S.W.; Ryu, J.S.; et al. Genome-Wide Association and Expression Quantitative Trait Loci Studies Identify Multiple Susceptibility Loci for Thyroid Cancer. Nat Commun 2017, 8, 15966. [Google Scholar] [CrossRef]
- Lai, X.; Umbricht, C.B.; Fisher, K.; Bishop, J.; Shi, Q.; Chen, S. Identification of Novel Biomarker and Therapeutic Target Candidates for Diagnosis and Treatment of Follicular Carcinoma. J Proteomics 2017, 166, 59–67. [Google Scholar] [CrossRef]
- Yang, X.; Wu, Y.; Xu, S.; Li, H.; Peng, C.; Cui, X.; Dhoomun, D.K.; Wang, G.; Xu, T.; Dong, M.; et al. Targeting the Inward Rectifier Potassium Channel 5.1 in Thyroid Cancer: Artificial Intelligence-Facilitated Molecular Docking for Drug Discovery. BMC Endocrine Disorders 2023, 23, 113. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; He, H.; Li, W.; Phay, J.; Shen, R.; Yu, L.; Hancioglu, B.; de la Chapelle, A. MYH9 Binds to LncRNA Gene PTCSC2 and Regulates FOXE1 in the 9q22 Thyroid Cancer Risk Locus. Proc Natl Acad Sci U S A 2017, 114, 474–479. [Google Scholar] [CrossRef] [PubMed]
- Kamihara, J.; Zhou, J.; LaDuca, H.; Wassner, A.J.; Dalton, E.; Garber, J.E.; Black, M.H. Germline Pathogenic Variants in Cancer Risk Genes among Patients with Thyroid Cancer and Suspected Predisposition. Cancer Medicine 2022, 11, 1745–1752. [Google Scholar] [CrossRef] [PubMed]
- Bakhsh, A.D.; Ladas, I.; Hamshere, M.L.; Bullock, M.; Kirov, G.; Zhang, L.; Taylor, P.N.; Gregory, J.W.; Scott-Coombes, D.; Völzke, H.; et al. An InDel in Phospholipase-C-B-1 Is Linked with Euthyroid Multinodular Goiter. Thyroid 2018, 28, 891–901. [Google Scholar] [CrossRef] [PubMed]
- Sarquis, M.; Moraes, D.C.; Bastos-Rodrigues, L.; Azevedo, P.G.; Ramos, A.V.; Reis, F.V.; Dande, P.V.; Paim, I.; Friedman, E.; De Marco, L. Germline Mutations in Familial Papillary Thyroid Cancer. Endocr Pathol 2020, 31, 14–20. [Google Scholar] [CrossRef]
- Ngeow, J.; Eng, C. PTEN in Hereditary and Sporadic Cancer. Cold Spring Harb Perspect Med 2020, 10, a036087. [Google Scholar] [CrossRef]
- Pilarski, R.; Burt, R.; Kohlman, W.; Pho, L.; Shannon, K.M.; Swisher, E. Cowden Syndrome and the PTEN Hamartoma Tumor Syndrome: Systematic Review and Revised Diagnostic Criteria. J Natl Cancer Inst 2013, 105, 1607–1616. [Google Scholar] [CrossRef]
- Hendricks, L.A.J.; Hoogerbrugge, N.; Schuurs-Hoeijmakers, J.H.M.; Vos, J.R. A Review on Age-Related Cancer Risks in PTEN Hamartoma Tumor Syndrome. Clin Genet 2021, 99, 219–225. [Google Scholar] [CrossRef]
- Bubien, V.; Bonnet, F.; Brouste, V.; Hoppe, S.; Barouk-Simonet, E.; David, A.; Edery, P.; Bottani, A.; Layet, V.; Caron, O.; et al. High Cumulative Risks of Cancer in Patients with PTEN Hamartoma Tumour Syndrome. J Med Genet 2013, 50, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Jonker, L.A.; Lebbink, C.A.; Jongmans, M.C.J.; Nievelstein, R. a. J.; Merks, J.H.M.; Nieveen van Dijkum, E.J.M.; Links, T.P.; Hoogerbrugge, N.; van Trotsenburg, A.S.P.; van Santen, H.M. Recommendations on Surveillance for Differentiated Thyroid Carcinoma in Children with PTEN Hamartoma Tumor Syndrome. Eur Thyroid J 2020, 9, 234–242. [Google Scholar] [CrossRef]
- Yehia, L.; Niazi, F.; Ni, Y.; Ngeow, J.; Sankunny, M.; Liu, Z.; Wei, W.; Mester, J.L.; Keri, R.A.; Zhang, B.; et al. Germline Heterozygous Variants in SEC23B Are Associated with Cowden Syndrome and Enriched in Apparently Sporadic Thyroid Cancer. Am J Hum Genet 2015, 97, 661–676. [Google Scholar] [CrossRef]
- Vierlinger, K.; Mansfeld, M.H.; Koperek, O.; Nöhammer, C.; Kaserer, K.; Leisch, F. Identification of SERPINA1 as Single Marker for Papillary Thyroid Carcinoma through Microarray Meta Analysis and Quantification of Its Discriminatory Power in Independent Validation. BMC Med Genomics 2011, 4, 30. [Google Scholar] [CrossRef] [PubMed]
- Makhlouf, A.-M.; Chitikova, Z.; Pusztaszeri, M.; Berczy, M.; Delucinge-Vivier, C.; Triponez, F.; Meyer, P.; Philippe, J.; Dibner, C. Identification of CHEK1, SLC26A4, c-KIT, TPO and TG as New Biomarkers for Human Follicular Thyroid Carcinoma. Oncotarget 2016, 7, 45776–45788. [Google Scholar] [CrossRef] [PubMed]
- Gudmundsson, J.; Thorleifsson, G.; Sigurdsson, J.K.; Stefansdottir, L.; Jonasson, J.G.; Gudjonsson, S.A.; Gudbjartsson, D.F.; Masson, G.; Johannsdottir, H.; Halldorsson, G.H.; et al. A Genome-Wide Association Study Yields Five Novel Thyroid Cancer Risk Loci. Nat Commun 2017, 8, 14517. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wang, Y.; Li, D.; Jing, S. Notch and TGF-β/Smad3 Pathways Are Involved in the Interaction between Cancer Cells and Cancer-Associated Fibroblasts in Papillary Thyroid Carcinoma. Tumour Biol 2014, 35, 379–385. [Google Scholar] [CrossRef]
- Buryk, M.A.; Picarsic, J.L.; Creary, S.E.; Shaw, P.H.; Simons, J.P.; Deutsch, M.; Monaco, S.E.; Nikiforov, Y.E.; Witchel, S.F. Identification of Unique, Heterozygous Germline Mutation, STK11 (p.F354L), in a Child with an Encapsulated Follicular Variant of Papillary Thyroid Carcinoma within Six Months of Completing Treatment for Neuroblastoma. Pediatr Dev Pathol 2015, 18, 318–323. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.; LiVolsi, V.A.; Brose, M.S.; Montone, K.T.; Morrissette, J.J.D.; Baloch, Z.W. STK11 Mutation Identified in Thyroid Carcinoma. Endocr Pathol 2016, 27, 65–69. [Google Scholar] [CrossRef]
- Alzahrani, A.S.; Alswailem, M.; Murugan, A.K.; Alghamdi, B.; Al-Hindi, H. Papillary Thyroid Cancer and a TERT Promotor Mutation-Positive Paraganglioma in a Patient With a Germline SDHB Mutation. J Endocr Soc 2022, 6, bvac076. [Google Scholar] [CrossRef] [PubMed]
- Abe, I.; Lam, A.K.-Y. Anaplastic Thyroid Carcinoma: Current Issues in Genomics and Therapeutics. Curr Oncol Rep 2021, 23, 31. [Google Scholar] [CrossRef]
- Lauper, J.M.; Krause, A.; Vaughan, T.L.; Monnat, R.J. Spectrum and Risk of Neoplasia in Werner Syndrome: A Systematic Review. PLoS One 2013, 8, e59709. [Google Scholar] [CrossRef] [PubMed]
- Risch, N. The Genetic Epidemiology of Cancer: Interpreting Family and Twin Studies and Their Implications for Molecular Genetic Approaches. Cancer Epidemiol Biomarkers Prev 2001, 10, 733–741. [Google Scholar]
- Liyanarachchi, S.; Gudmundsson, J.; Ferkingstad, E.; He, H.; Jonasson, J.G.; Tragante, V.; Asselbergs, F.W.; Xu, L.; Kiemeney, L.A.; Netea-Maier, R.T.; et al. Assessing Thyroid Cancer Risk Using Polygenic Risk Scores. Proc. Natl. Acad. Sci. U.S.A. 2020, 117, 5997–6002. [Google Scholar] [CrossRef]
- Abdullah Suhaimi, S.N.; Nazri, N.; Nani Harlina, M.L.; Md Isa, N.; Muhammad, R. Familial Adenomatous Polyposis-Associated Papillary Thyroid Cancer. Malays J Med Sci 2015, 22, 69–72. [Google Scholar]
- Xu, M.; Zheng, Y.; Zuo, Z.; Zhou, Q.; Deng, Q.; Wang, J.; Wang, D. De Novo Familial Adenomatous Polyposis Associated Thyroid Cancer with a c.2929delG Frameshift Deletion Mutation in APC: A Case Report and Literature Review. World Journal of Surgical Oncology 2023, 21, 73. [Google Scholar] [CrossRef]
- Chenbhanich, J.; Atsawarungruangkit, A.; Korpaisarn, S.; Phupitakphol, T.; Osataphan, S.; Phowthongkum, P. Prevalence of Thyroid Diseases in Familial Adenomatous Polyposis: A Systematic Review and Meta-Analysis. Familial Cancer 2019, 18, 53–62. [Google Scholar] [CrossRef]
- Expression of ARSB in Cancer - Summary - The Human Protein Atlas. Available online: https://www.proteinatlas.org/ENSG00000113273-ARSB/pathology (accessed on 15 August 2023).
- Pstrąg, N.; Ziemnicka, K.; Bluyssen, H.; Wesoły, J. Thyroid Cancers of Follicular Origin in a Genomic Light: In-Depth Overview of Common and Unique Molecular Marker Candidates. Molecular Cancer 2018, 17, 116. [Google Scholar] [CrossRef] [PubMed]
- Nagy, R.; Ringel, M.D. Genetic Predisposition for Nonmedullary Thyroid Cancer. HORM CANC 2015, 6, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, S.; Tobacman, J.K. Arylsulfatase B Regulates Colonic Epithelial Cell Migration by Effects on MMP9 Expression and RhoA Activation. Clin Exp Metastasis 2009, 26, 535–545. [Google Scholar] [CrossRef]
- Xu, L.; Morari, E.C.; Wei, Q.; Sturgis, E.M.; Ward, L.S. Functional Variations in the ATM Gene and Susceptibility to Differentiated Thyroid Carcinoma. The Journal of Clinical Endocrinology & Metabolism 2012, 97, 1913–1921. [Google Scholar] [CrossRef]
- CBioPortal for Cancer Genomics. Available online: http://www.cbioportal.org/ (accessed on 15 August 2023).
- Tissue Expression of ATM - Staining in Thyroid Gland - The Human Protein Atlas. Available online: https://www.proteinatlas.org/ENSG00000149311-ATM/tissue/thyroid+gland (accessed on 15 August 2023).
- Miasaki, F.Y.; Saito, K.C.; Yamamoto, G.L.; Boguszewski, C.L.; de Carvalho, G.A.; Kimura, E.T.; Kopp, P.A. Thyroid and Breast Cancer in 2 Sisters With Monoallelic Mutations in the Ataxia Telangiectasia Mutated (ATM) Gene. Journal of the Endocrine Society 2022, 6, bvac026. [Google Scholar] [CrossRef]
- PubChem BRCA1 - BRCA1 DNA Repair Associated (Human). Available online: https://pubchem.ncbi.nlm.nih.gov/gene/BRCA1/human (accessed on 15 August 2023).
- Cieszyńska, M.; Kluźniak, W.; Wokołorczyk, D.; Cybulski, C.; Huzarski, T.; Gronwald, J.; Falco, M.; Dębniak, T.; Jakubowska, A.; Derkacz, R.; et al. Risk of Second Primary Thyroid Cancer in Women with Breast Cancer. Cancers 2022, 14, 957. [Google Scholar] [CrossRef]
- Swierniak, M.; Pfeifer, A.; Stokowy, T.; Rusinek, D.; Chekan, M.; Lange, D.; Krajewska, J.; Oczko-Wojciechowska, M.; Czarniecka, A.; Jarzab, M.; et al. Somatic Mutation Profiling of Follicular Thyroid Cancer by next Generation Sequencing. Mol Cell Endocrinol 2016, 433, 130–137. [Google Scholar] [CrossRef]
- De Toledo, M.; Coulon, V.; Schmidt, S.; Fort, P.; Blangy, A. The Gene for a New Brain Specific RhoA Exchange Factor Maps to the Highly Unstable Chromosomal Region 1p36.2-1p36.3. Oncogene 2001, 20, 7307–7317. [Google Scholar] [CrossRef]
- Azzedine, H.; Zavadakova, P.; Planté-Bordeneuve, V.; Vaz Pato, M.; Pinto, N.; Bartesaghi, L.; Zenker, J.; Poirot, O.; Bernard-Marissal, N.; Arnaud Gouttenoire, E.; et al. PLEKHG5 Deficiency Leads to an Intermediate Form of Autosomal-Recessive Charcot-Marie-Tooth Disease. Hum Mol Genet 2013, 22, 4224–4232. [Google Scholar] [CrossRef]
- Kai, J.-D.; Cheng, L.-H.; Li, B.-F.; Kang, K.; Xiong, F.; Fu, J.-C.; Wang, S. MYH9 Is a Novel Cancer Stem Cell Marker and Prognostic Indicator in Esophageal Cancer That Promotes Oncogenesis through the PI3K/AKT/MTOR Axis. Cell Biology International 2022, 46, 2085–2094. [Google Scholar] [CrossRef]
- Cameselle-Teijeiro, J.M.; Mete, O.; Asa, S.L.; LiVolsi, V. Inherited Follicular Epithelial-Derived Thyroid Carcinomas: From Molecular Biology to Histological Correlates. Endocr Pathol 2021, 32, 77–101. [Google Scholar] [CrossRef] [PubMed]
- Bakhsh, A.; Kirov, G.; Gregory, J.W.; Williams, E.D.; Ludgate, M. A New Form of Familial Multi-Nodular Goitre with Progression to Differentiated Thyroid Cancer. Endocr Relat Cancer 2006, 13, 475–483. [Google Scholar] [CrossRef] [PubMed]
- Yehia, L.; Plitt, G.; Tushar, A.M.; Joo, J.; Burke, C.A.; Campbell, S.C.; Heiden, K.; Jin, J.; Macaron, C.; Michener, C.M.; et al. Longitudinal Analysis of Cancer Risk in Children and Adults With Germline PTEN Variants. JAMA Netw Open 2023, 6, e239705. [Google Scholar] [CrossRef] [PubMed]
- Saenko, V.A.; Rogounovitch, T.I. Genetic Polymorphism Predisposing to Differentiated Thyroid Cancer: A Review of Major Findings of the Genome-Wide Association Studies. Endocrinol Metab (Seoul) 2018, 33, 164–174. [Google Scholar] [CrossRef]
- Wang, Q.; Shen, Y.; Ye, B.; Hu, H.; Fan, C.; Wang, T.; Zheng, Y.; Lv, J.; Ma, Y.; Xiang, M. Gene Expression Differences between Thyroid Carcinoma, Thyroid Adenoma and Normal Thyroid Tissue. Oncol Rep 2018. [Google Scholar] [CrossRef]
- Jiang, Y.-L.; Zhao, Z.-Y.; Li, B.-R.; Yang, F.; Li, J.; Jin, X.-W.; Wang, H.; Yu, E.-D.; Sun, S.-H.; Ning, S.-B. The Altered Activity of P53 Signaling Pathway by STK11 Gene Mutations and Its Cancer Phenotype in Peutz-Jeghers Syndrome. BMC Medical Genetics 2018, 19, 141. [Google Scholar] [CrossRef]
- Lardelli, R.M.; Schaffer, A.E.; Eggens, V.R.C.; Zaki, M.S.; Grainger, S.; Sathe, S.; Van Nostrand, E.L.; Schlachetzki, Z.; Rosti, B.; Akizu, N.; et al. Biallelic Mutations in the 3′ Exonuclease TOE1 Cause Pontocerebellar Hypoplasia and Uncover a Role in SnRNA Processing. Nat Genet 2017, 49, 457–464. [Google Scholar] [CrossRef]
- Expression of TOE1 in Thyroid Cancer - The Human Protein Atlas. Available online: https://www.proteinatlas.org/ENSG00000132773-TOE1/pathology/thyroid+cancer (accessed on 15 August 2023).
- Liu, X.; Bishop, J.; Shan, Y.; Pai, S.; Liu, D.; Murugan, A.K.; Sun, H.; El-Naggar, A.K.; Xing, M. Highly Prevalent TERT Promoter Mutations in Aggressive Thyroid Cancers. Endocr Relat Cancer 2013, 20, 603–610. [Google Scholar] [CrossRef]
- Matsuse, M.; Yabuta, T.; Saenko, V.; Hirokawa, M.; Nishihara, E.; Suzuki, K.; Yamashita, S.; Miyauchi, A.; Mitsutake, N. TERT Promoter Mutations and Ki-67 Labeling Index as a Prognostic Marker of Papillary Thyroid Carcinomas: Combination of Two Independent Factors. Sci Rep 2017, 7, 41752. [Google Scholar] [CrossRef]
- Muftuoglu, M.; Oshima, J.; von Kobbe, C.; Cheng, W.-H.; Leistritz, D.F.; Bohr, V.A. The Clinical Characteristics of Werner Syndrome: Molecular and Biochemical Diagnosis. Hum Genet 2008, 124, 369–377. [Google Scholar] [CrossRef]
- Jazdzewski, K.; Murray, E.L.; Franssila, K.; Jarzab, B.; Schoenberg, D.R.; de la Chapelle, A. Common SNP in Pre-MiR-146a Decreases Mature MiR Expression and Predisposes to Papillary Thyroid Carcinoma. Proceedings of the National Academy of Sciences 2008, 105, 7269–7274. [Google Scholar] [CrossRef] [PubMed]
- Zuk, O.; Hechter, E.; Sunyaev, S.R.; Lander, E.S. The Mystery of Missing Heritability: Genetic Interactions Create Phantom Heritability. Proceedings of the National Academy of Sciences 2012, 109, 1193–1198. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Thyroid cancer classification (modified from Vriens et al. 2009 [7], Hińcza et al. 2019 [8] and Kamani et al. 2022 [12].
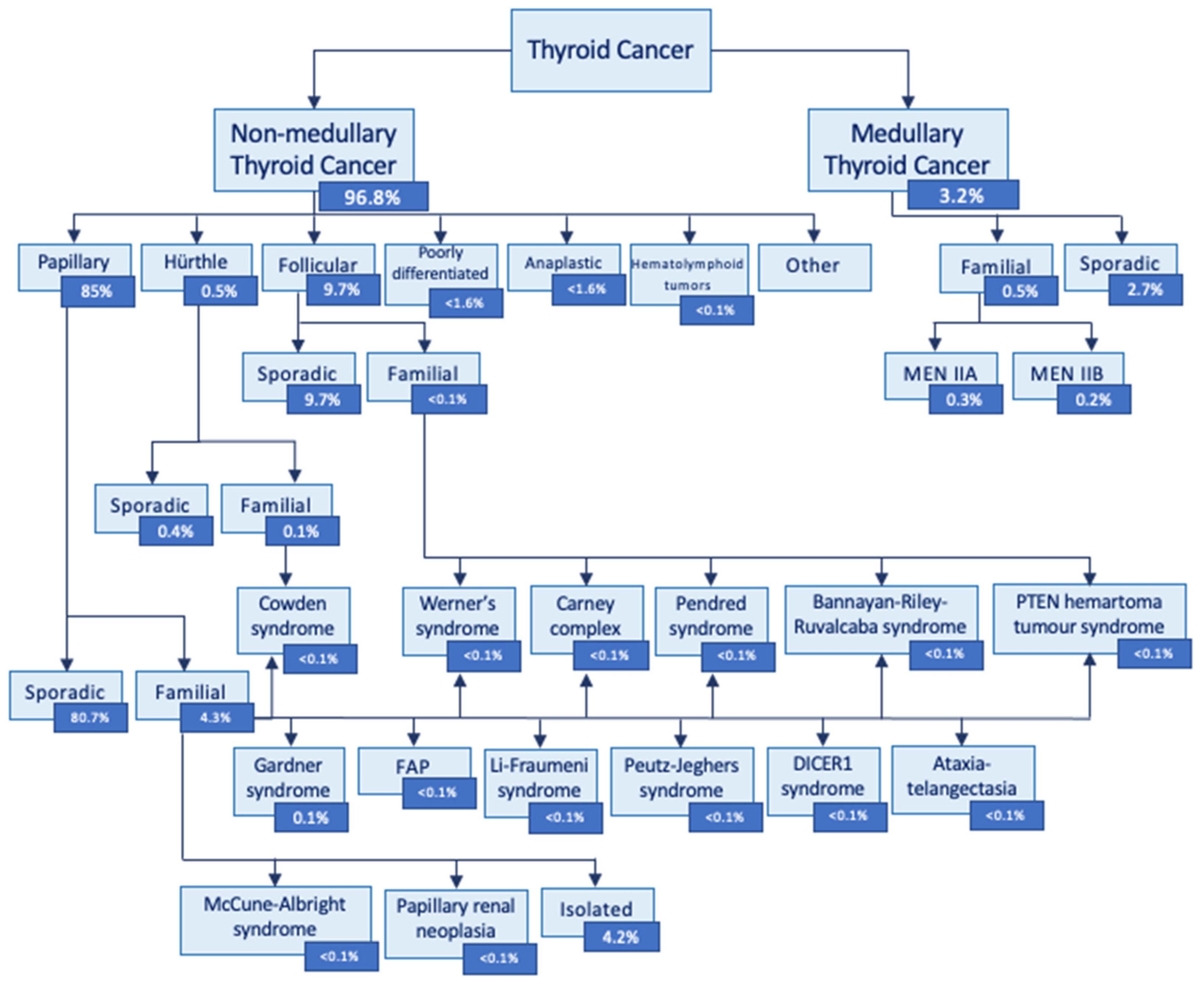
Figure 2.
Workflow for assessing the prevalence of germline mutation predisposing to the differentiated thyroid cancer in a cohort of Polish individuals using a genome-wide association study.
Figure 2.
Workflow for assessing the prevalence of germline mutation predisposing to the differentiated thyroid cancer in a cohort of Polish individuals using a genome-wide association study.
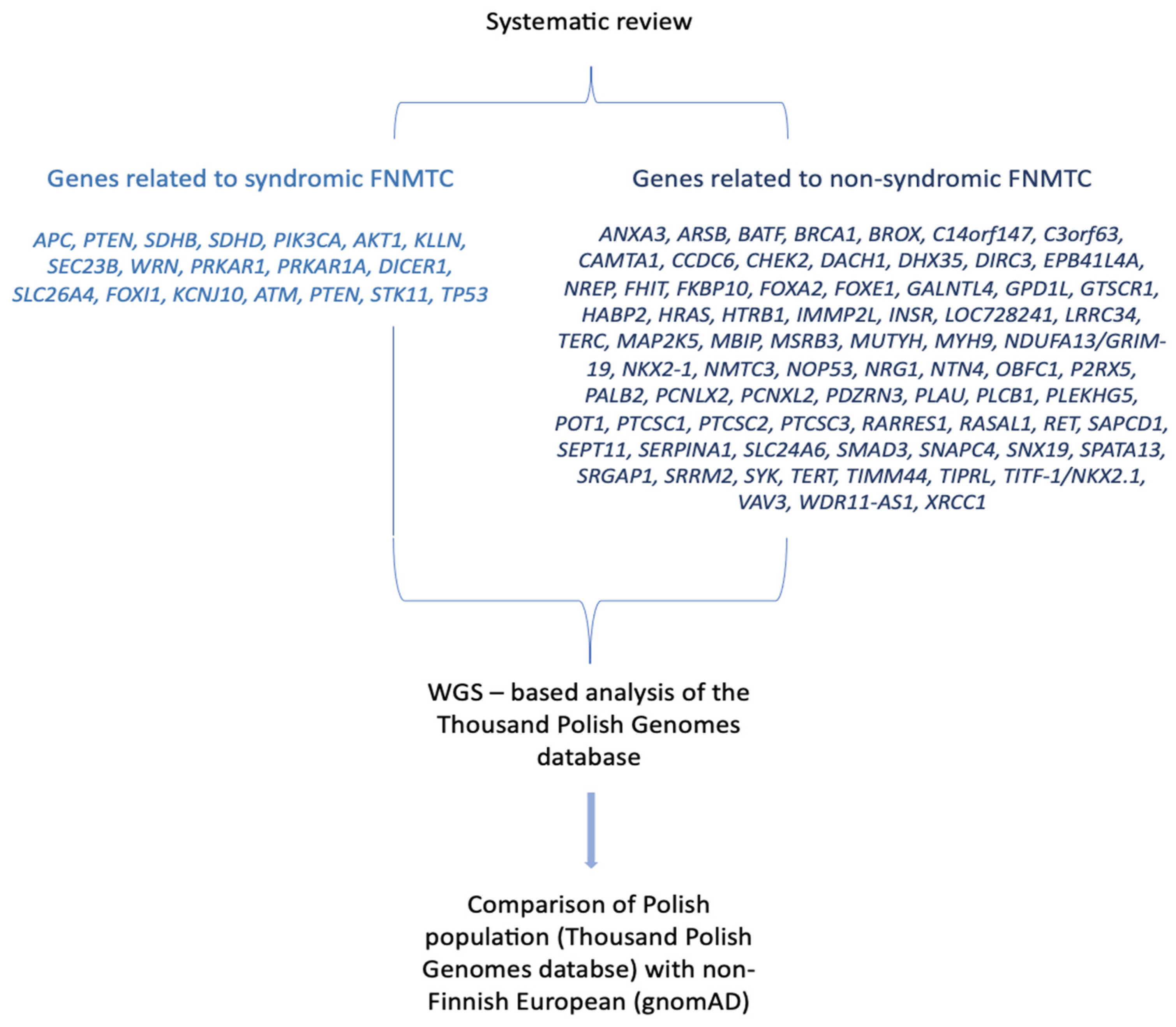
Figure 3.
Allele frequencies of ClinVar variants in 23 DTC associated genes. All displayed variants are statistically significant (q-value < 0.05) in difference in frequency between POL and gnomAD NFE. Most of those genetic variants are more frequent among Polish population, except from DICER1 and KCNJ10.
Figure 3.
Allele frequencies of ClinVar variants in 23 DTC associated genes. All displayed variants are statistically significant (q-value < 0.05) in difference in frequency between POL and gnomAD NFE. Most of those genetic variants are more frequent among Polish population, except from DICER1 and KCNJ10.
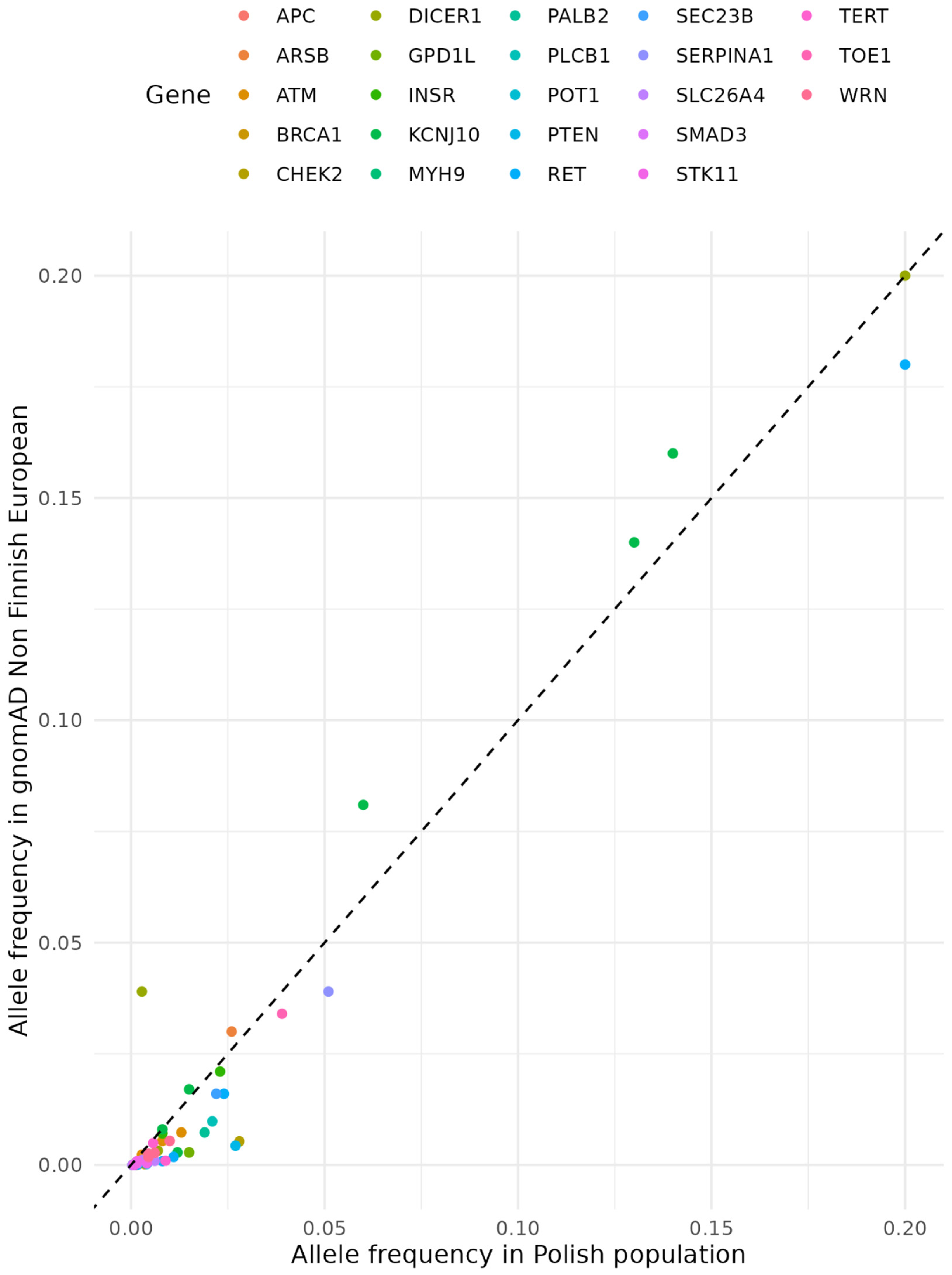

Table 1.
Genetic syndromes and genes related to syndromic familial non-medullary thyroid cancer; adapted from Kamani et al. [12].
Table 1.
Genetic syndromes and genes related to syndromic familial non-medullary thyroid cancer; adapted from Kamani et al. [12].
| Name | Gene | Mode of Inheritance | Thyroid cancer histological subtype* | Phenotypes other than thyroid cancer |
|---|---|---|---|---|
| FAP and Gardner’s syndrome | APC | Autosomal dominant | PTC | Colorectal carcinoma, ampullary carcinoma, hepatoblastoma, medulloblastoma |
| Cowden syndrome |
PTEN, SDHB-D, PIK3CA, AKT1, KLLN, SEC23B |
Autosomal dominant | cPTC, fvPTC, FTC |
Multiple hamartomas, follicular thyroid carcinoma, benign thyroid nodules, breast cancer, endometrial cancer |
| Werner syndrome | WRN | Autosomal recessive | PTC, FTC, ATC |
Premature ageing, scleroderma-like skin changes, cataracts, subcutaneous calcifications, muscular atrophy, diabetes |
| Carney complex | PRKAR1 | Autosomal dominant | PTC, FTC | Spotty skin pigmentation, cardiac myxomas, endocrine tumors |
| DICER1 syndrome | DICER1 | Autosomal dominant | PTC, DTC | Endocrine tumors (thyroid, parathyroid, pituitary, pineal gland, endocrine pancreas, paragangliomas, medullary, adrenocortical, ovarian, and testicular tumors |
| Pendred syndrome | SLC26A4, FOXI1, KCNJ10 | Autosomal recessive | PTC, FTC, ATC | Sensorineural deafness/hearing impairment, goiter, and an abnormal organification of iodide with or without hypothyroidism |
| Ataxia-telangiectasia | ATM | Autosomal recessive | PTC | Cerebellar degeneration, telangiectasia, immunodeficiency, recurrent sinopulmonary infections, radiation sensitivity, premature aging, lymphoid cancer, poor growth, gonadal atrophy, insulin resistant diabetes |
| Bannayan- Riley- Ruvalcaba syndrome | PTEN | Autosomal dominant | PTC, FTC | Macrocephaly, hamartomatous tissue overgrowth, lipomas, pigmented macules on the penis, developmental delay, large birth weight, joint hyperextensibility, endometrial cancer, renal cell carcinoma, Lhermitte–Duclos disease |
| Peutz-Jeghers syndrome | STK11 | Autosomal dominant | PTC, DTC | Gastrointestinal polyposis, mucocutaneous pigmented macules, breast cancer, uterine cancer, cervical cancer, lung cancer, ovarian cancer, testicular cancers |
| PTEN hamartoma tumor syndrome | PTEN | Autosomal dominant | FTC, PTC, fvPTC |
Breast cancer, endometrial cancer, gastrointestinal hamartomas, Lhermitte-Duclos disease, macrocephaly, macular pigmentation of the glans penis, multiple mucocutaneous lesions, autism spectrum disorder, colon cancer, esophageal glycogenic acanthosis, lipomas, mental retardation, renal cell carcinoma, testicular lipomatosis, thyroid adenoma, multinodular goiter |
| Li-Fraumeni syndrome | TP53 | Autosomal dominant | cPTC, fvPTC | Adrenocortical carcinomas, breast cancer, central nervous system tumors, osteosarcomas, soft-tissue sarcomas, leukemia, lymphoma, gastrointestinal cancers, cancers of head and neck, kidney, larynx, lung, skin, ovary, pancreas, prostate, and testis |
*PTC (papillary thyroid cancer), FTC (follicular thyroid cancer), ATC (anaplastic thyroid cancer), cPTC – classical variant of PTC, fvPTC (follicular variant of PTC), DTC – differentiated thyroid cancer.
Table 2.
Genes with significant (q-value <0.05) differences in pathogenic variant burden among Polish population (Thousand Polish Genomes database) compared with non-Finnish European (gnomAD); as indicated in the Table 1 in supplement, most of the genetic variants analyzed are more frequent among Polish population, however two exceptions are indicated in red below.
Table 2.
Genes with significant (q-value <0.05) differences in pathogenic variant burden among Polish population (Thousand Polish Genomes database) compared with non-Finnish European (gnomAD); as indicated in the Table 1 in supplement, most of the genetic variants analyzed are more frequent among Polish population, however two exceptions are indicated in red below.
| Gene | Type of thyroid cancer* | Hereditary syndromes | Other cancers | Reference |
|---|---|---|---|---|
| APC | PTC with cribriform pattern | FAP and Gardner’s syndrome | Colorectal cancer, ampullary carcinoma, hepatoblastoma, medulloblastoma | Kamani et al. 2022 [34]; Cetta et al. 2015 [34]2023-10-07 12:25:00 PM |
| ARSB | DTC | N/A | N/A | Figlioli et al. 2014 [35] |
| ATM | PTC | Ataxia-telangiectasia | Cerebellar degeneration, telangiectasia, immunodeficiency, recurrent sinopulmonary infections, radiation sensitivity, premature ageing, lymphoid cancer, poor growth, gonadal atrophy, insulin resistant diabetes | Kamani et al. 2022 [12] |
| PTC, DTC | N/A | N/A | Dombernowsky et al. 2008 [36]; Akulevich et al. 2009 [37]; Gu et al. 2014 [38]; Wójcicka 2014 [39] |
|
| BRCA1 | PTC | N/A | N/A | Wójcicka et al. 2014 [39] |
| CHEK2 | Non-syndromic DTC | N/A | Breast cancer, prostate cancer | Wang et al. 2019 [40] |
| PTC | N/A | N/A | Wójcicka et al. 2014 [41]; Siołek et al. 2015 [42]; Zhou et al. 2021[41] |
|
| DICER1 | PTC, DTC | DICER1 syndrome | Endocrine tumors (parathyroid, pituitary, pineal gland, endocrine pancreas, paragangliomas, medullary, adrenocortical, ovarian, and testicular tumors | Rutter et al. 2016 [43] |
| Nephroblastoma, NMTC pleuropulmonary blastoma, cystic nephroma, multinodular goiter, thyroid adenoma, sex cord tumor | Zhou et al.2021 [41] | |||
| GPD1L | DTC | N/A | N/A | Figloli et al. 2014 [35] |
| INSR | PTC | N/A | N/A | Son et al. 2017 [44] |
| FTC | N/A | N/A | Lai et al. 2017 [45] | |
| KCNJ10 | ATC | Pendred | Sensorineural deafness/hearing impairment, goiter, and an abnormal organification of iodide with or without hypothyroidism | Liu et al. 2016 [46]; Yang et al. 2023 0/0/00 0:00:00 AM |
| MYH9 | PTC, FTC | N/A | N/A | Wang et al. 2017 [47] |
| PALB2 | PTC | N/A | N/A | Kamihara et al. 2022 [48] |
| PLCB1 | PTC | N/A | N/A | Bakhsh et al. 2018 [49] |
| PLEKHG5 | PTC | N/A | N/A | Sarquis et al. 2020 [50] |
| PTEN | cPTC, fvPTC, FTC | Cowden syndrome | Multiple hamartomas, follicular thyroid carcinoma, benign thyroid nodules, breast cancer, endometrial cancer | Bevan et al. 2001 [51]; Pilarski et al. 2013 [52]; Hendricks et al. 2021 [53]; Bubien et al. 2013 [54]; Jonker et al. 2020 [55]; Ngeow et al. 2020 0/0/00 0:00:00 AM |
| PTC, FTC | Bannayan-Riley-Ruvalcaba syndrome | Macrocephaly, hamartomatous tissue overgrowth, lipomas, pigmented macules on the penis, developmental delay, large birth weight, joint hyperextensibility, endometrial cancer, renal cell carcinoma, Lhermitte–Duclos disease | ||
| FTC, PTC, fvPTC |
PTEN hamartoma tumor syndrome | Breast cancer, Endometrial cancer, FTC, Gastrointestinal hamartomas, Lhermitte-Duclos disease, Macrocephaly, Macu- lar pigmentation of the glans penis, Multiple mucocutaneous lesions, Autism spectrum disorder, Colon cancer, Esophageal glycogenic acanthosis, Lipomas, Mental retardation, Renal cell carcinoma, Testicular lipomatosis, PTC, fvPTC, thyroid adenoma, MNG | ||
| RET | PTC | FAP | Colorectal carcinoma, ampullary carcinoma, hepatoblastoma, medulloblastoma | Cetta et al. 2015 [34] |
| SEC23B | cPTC, fvPTC, FTC | Cowden syndrome | Multiple hamartomas, follicular thyroid carcinoma, benign thyroid nodules, breast cancer, endometrial cancer | Yehia et al. 2015 [56] |
| SERPINA1 | PTC | N/A | N/A | Vierlinger et al. 2011 [57] |
| SLC26A4 | FTC, DTC | Pendred syndrome | Sensorineural deafness/hearing impairment, goiter, and an abnormal organification of iodide with or without hypothyroidism | Makhlouf et al. 2016 [58] |
| SMAD3 | DTC | N/A | N/A | Gudmundsson et al. 2017 [59] |
| PTC | N/A | N/A | Zhang et al. 2014 [60]; | |
| STK11 | PTC, DTC | Peutz-Jeghers syndrome | Gastrointestinal (GI) polyposis, mucocutaneous pigmented macules, breast cancer, uterine cancer, cervical cancer, lung cancer, ovarian cancer, testicular cancers | Buryk et al. 2015 [61]; Wei et al. 2016 [62] |
| TERT | DTC | N/A | N/A | Gudmundsson et al. 2017 [59] |
| PTC | N/A | N/A | Kim et al. 2022 [63]; Alzahrani et al. 2022 0/0/00 0:00:00 AM |
|
| ATC | N/A | N/A | Abe et al. 2021 [64] | |
| TOE1 | PTC | FAP | Colorectal carcinoma, ampullary carcinoma, hepatoblastoma, medulloblastoma | ClinVar [30] |
| WRN | PTC, FTC, ATC |
Werner syndrome | Premature aging, scleroderma-like skin changes, cataracts, subcutaneous calcifications, muscular atrophy, diabetes | Lauper et al. 2013 [65] |
* PTC (papillary thyroid cancer), FTC (follicular thyroid cancer), ATC (anaplastic thyroid cancer), cPTC – classical variant of PTC, fvPTC (follicular variant of PTC), DTC – differentiated thyroid cancer.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



