
Submitted:
07 September 2023
Posted:
11 September 2023
You are already at the latest version
Abstract
In their natural environment, bacteria are exposed to various stresses. The stress-response sigma factor SigB of gram-positive Bacillus subtilis is the best-characterized example. Unlike Bacillus subtilis, the gram-positive bacterium Streptomyces coelicolor A3(2) contains nine SigB homologues (SigBFGHIKLMN) with a major role in differentiation and response to osmotic stress. We previously constructed a two-plasmid system to identify promoters recognized by these sigma factors. Interestingly, almost all identified promoters were recognized by two or more SigB homologues. However, no specific sequences characteristic for these recognition groups were found. To examine this cross-recognition in vivo in S. coelicolor A3 (2), one of these promoters was cho-sen, which drives the expression of the sporulation-specific gene ssgB. The ssgBp promoter was inserted into a luciferase reporter plasmid and conjugated to S. coelicolor M145 and nine mutant strains containing deleted individual sigB homologous genes. Luciferase reporter activity indi-cated differential activity of this promoter in these mutant strains, suggesting overlapping pro-moter recognition by these SigB homologues. To determine which nucleotides in the ̶ 10 re-gion are responsible for the selection of a specific SigB homologue, several mutant promoters with altered last three nucleotides in this region were prepared and tested in the two-plasmid system. Some mutant promoters were specifically recognized by some SigB homologues. Mutant promoters were inserted into a luciferase reporter plasmid and conjugated to S. coelicolor A3(2) and these nine mutant strains. Luciferase reporter activity indicated differential activity of these ssgBp mutant promoters, indicating overlapping promoter recognition by these SigB homologues in S. coelicolor A3(2).
Keywords:
differentiation
; promoter
; regulation
; sigma factor
; Streptomyces
; stress response
1. Introduction
Gram-positive soil-dwelling bacteria of the genus Streptomyces are among the best producers of biologically active secondary metabolites, including many antibiotics and antitumor compounds. The production of these secondary metabolites is temporally and genetically coordinated with the process of morphological differentiation of streptomycetes. This process is characterized by several cell types and usually takes place in surface-grown Streptomyces strains on a solid agar media. The spores initially germinate and form branched hyphae characterised by apical growth (the so-called vegetative substrate mycelium). At this stage, cell division does not lead to cell separation, but longer multi-nucleoid compartments are formed, which are separated by occasional cross-walls. In response to various signals (nutrition, quorum sensing, environmental stresses, etc.), white hyphae break surface tension and emerge from the substrate mycelium (forming so-called aerial mycelium) and grow similarly by tip extension. After hyphal growth ceases, sporulation is initiated and the tubulin-like GTPase FtsZ polymerizes into long filaments at regular sites of aerial hyphae, forming a ladder of Z rings that recruit additional cell division proteins to coordinate peptidoglycan synthesis at these regularly-spaced nascent division septa. Finally, cytokinesis coordinated with DNA segregation leads to long chains of unigenomic exospores with a characteristic colour [1,2].
Unlike unicellular bacteria, streptomycetes lack the canonical Min and Noc systems or nucleoid-occlusion proteins, but sporulation septation is controlled by a family of SsgA-like proteins (SALPs) unique to differentiating actinomycetes. The archetypal membrane associated SALP, SsgB, in cooperation with its paralogue SsgA, recruits FtsZ to septal sites. The correct localization of SsgB is regulated by the transmembrane protein SepG, and two dynamin-like proteins DynA and DynB and membrane-anchoring protein SepF stabilize the Z-ring [3,4].
In addition to these cell division proteins, streptomycete differentiation is regulated by two separate classes of regulators. Regulators encoded by bld (bald) genes control the transition from substrate mycelium to aerial mycelium. Mutants in the bld genes are unable to erect aerial hyphae and appear bald, lacking the characteristic fuzzy morphology. Regulators encoded by whi (white) genes control the differentiation of aerial hyphae into spore chains. Mutants in the whi genes form aerial hyphae blocked at specific stages of sporogenesis and appear white [1]. In addition to these regulators, c-di-GMP is a key signal molecule controlling the transitions between these stages. In the first stage (transition from substrate to aerial mycelium), c-di-GMP positively affects the binding of the key repressor BldD to the large regulon of genes, thereby repressing their transcription and this transition. After the initiation of differentiation, the level of c-di-GMP decreases, causing dissociation of BldD and derepression of its regulon. In the second stage (sporogenesis of aerial hyphae), c-di-GMP controls the activity of the sporulation-specific sigma factor WhiG. It mediates a complex with its specific anti-sigma factor RsiG, thereby preventing the activation of the WhiG regulon of sporulation-specific genes. A decrease in c-di-GMP during sporogenesis causes the release of WhiG from this complex to activate these genes [5,6].
In their natural habitats, streptomycetes are exposed to various environmental and nutritional stresses, and the regulation of differentiation is associated with stress-related signals [7,8]. The ability to modulate gene expression in response to the stress conditions is mediated in many bacteria by stress-response sigma factors of RNA polymerase that control the expression of genes encoding proteins required to overcome these adverse conditions. The stress-response sigma factor SigB of gram-positive Bacillus subtilis is the best-characterized example. Its activity is regulated by the RsbW/RsbV partner-switching mechanism. Under non-stress conditions, SigB is sequestered by the anti-sigma factor RsbW. The release of the sigma factor from this complex is accomplished by the anti-anti-sigma factor RsbV, which is dephosphorylated by specific PP2C-type phosphatases RsbU/RsbP under stress conditions and sequesters RsbW [9,10].
In contrast to B. subtilis, the genome of the best-characterized strain Streptomyces coelicolor A3(2) contains up to 65 sigma factor genes, including genes encoding nine close SigB homologues (SigBFGHIKLMN), 45 RsbW homologues, 17 RsbV homologues, and 44 RsbU/RsbP homologues, suggesting relatively complex regulation of these SigB homologues compared to B. subtilis [10,11,12]. The primary function of these nine SigB homologues is the control of morphological differentiation and response to osmotic stress. Phenotypic analysis of mutants for several sigB homologues revealed a specific role in controlling various stages of morphological differentiation. SigF (SCO4035) controls the late stages of morphological differentiation, spore maturation and pigmentation [13]. SigN (SCO4034) plays a role in the early stage of morphological differentiation, in the subapical stem region of aerial hyphae [14]. SigK (SCO6529) plays a negative role in morphological differentiation and secondary metabolism [15]. SigL (SCO7278) is thought to play a role in the sporulation of aerial hyphae and SigM (SCO7324) in efficient sporulation [16]. On the other hand, SigI (SCO3068) is probably involved in the response to osmotic stress [17]. No specific role for SigG (SCO7341) was originally proposed [18], however, the sigG gene was expressed during spore germination, suggesting its role in controlling this process [19]. Interestingly, phenotypic analysis of the genes for SigH (SCO5243) and SigB (SCO0600) suggested their dual role in controlling morphological differentiation and response to osmotic stress [16, 20, 21, 22. 23].
Regulation of SigB homologues in S. coelicolor A3(2) is more complex than that of SigB regulation in B. subtilis due to the large number of proposed regulators identified in this strain [10,11,12]. Some of them have been experimentally verified to activate SigB homologues by a partner-switching phosphorylation mechanism in S. coelicolor A3(2). The anti-sigma factor RsbA (SCO0599) and the anti-anti-sigma factor RsbV (SCO7325) are involved in the osmotic stress activation of SigB [24]. The anti-sigma factor PrsI (SCO3036) and the anti-anti-sigma factor ArsI (SCO3067) are involved in the activation of SigI [17]. The anti-sigma factor RsmA (SCO7313) specifically interacts with SigM, but no specific anti-anti-sigma factor has been identified in this regulation [25]. The anti-sigma factor UshX/PrsH (SCO5244) specifically interacts and negatively regulates SigH [22,26,27] and the anti-anti-sigma factor BldG (SCO3549) is involved in the osmotic stress activation of this sigma factor by sequestering UshX [28]. This regulation is more complex because BldG specifically interacts with and is phosphorylated by several anti-sigma factors, including RsfA (SCO4677) and SCO7328, which interact with four other sigma factors SigF, SigG, SigK, SigM [29]. All these data suggest a linked regulation of stress response and differentiation in S. coelicolor A3(2).
Using a heterologous two-plasmid E. coli system, we previously identified the promoters recognized by these SigB homologues. Interestingly, almost all identified promoters were recognized by two or more SigB homologues, suggesting their cross-recognition by these sigma factors. Although analysis of these promoters indicated some distinct groups of promoters recognized by these sigma factors, their sequence analysis did not reveal any specific sequences characteristic for these recognition groups. All promoters showed high similarity in the ̶ 35 and ̶ 10 regions, which is consistent with the high similarity of SigB homologues in regions 2.4 and 4.2, which are involved in the recognition of the ̶ 10 and ̶ 35 promoter regions [30].
In this study, we attempted to further characterize this cross-recognition in vivo in S. coelicolor A3(2). We selected a representative promoter ssgBp that controls the expression of the sporulation-specific essential gene ssgB and analyzed its expressing in vivo using a luciferase reporter system in the wild-type strain S. coelicolor A3(2) and nine mutant strains of S. coelicolor containing deleted individual sigma factor genes. To see which nucleotides in the -10 region are responsible for the selection of a specific SigB homologue, we additionally prepared mutant ssgBpA-G promoters in this region and tested them in the E. coli two-plasmid system and in vivo with the luciferase reporter system in S. coelicolor. The results showed differential activity of this promoter in these mutant strains, indicating overlapping promoter recognition by these nine SigB homologues in vivo in S. coelicolor A3(2), and some mutant promoters were specifically recognized by some of SigB homologues.
2. Results and Discussion
2.1. In vivo Cross-Recognition of the highly conserved ssgBp promoter by Nine SigB Homologues
Using a heterologous two-plasmid E. coli system, we previously identified 24 promoters dependent on nine S. coelicolor A3(2) SigB homologues, but many of them were recognized by two or more SigB homologues [30]. One of the identified promoters, SCO1541p directs the expression of the key sporulation-specific cell division gene ssgB. This ssgBp promoter (Figure 1a) was only recognized by SigF in the two-plasmid system [30]. However, its in vivo verification by S1-nuclease mapping revealed no significant dependence on SigF, but partial dependence upon SigH [31].
The ssgB gene encodes the key sporulation-specific protein SsgB in S. coelicolor A3(2) [32,33]. During the initial stages of sporulation of aerial hyphae, SsgB recruits GTPase FtsZ to the septum site in sporulating aerial hyphae and promotes the formation of Z-rings [4]. The sequence of the SsgB protein is extremely conserved within the genus Streptomyces. Of the 78 strains analyzed, it showed only two amino acids (aa) variations at the C-terminus of its 137 aa sequence [34]. Therefore, we performed a similar analysis of the ssgBp promoter with genome sequences in databases. BlastN analysis revealed similar very high promoter sequence conservation only in Streptomyces genomes with 545 hits. It was highly conserved with 100% identity in the regions -35, -10, up to the ATG initiation codon of the ssgB gene (Figure 1b). This is unprecedented conservation. A similar analysis of the hrdBp promoter, which directs the expression of the major vegetative sigma factor in Streptomyces [35] showed a standard similarity within the Streptomyces genus (from 78 to 98% identity in the 50 nt sequence up to TSS), similar to the comparison of the entire hrdB gene (from 86.9 to 99.7% identity in the 1536 nt sequence).
To examine the dependence of the ssgBp promoter on all nine SigB homologues in vivo in S. coelicolor A3(2), we inserted the ssgBp promoter as a 750-bp BamHI-XhoI DNA fragment from pF81 [30,31], in the luciferase reporter plasmid pLux-kasOp (Figure 2a). This new PhiC31 phage-based integration plasmid contains a synthetic luxCDABE operon from plasmid pMU1s* [37], a kanamycin resistance (KanR) gene compatible for selection with apramycin resistance (AprR) of S. coelicolor mutants in sigB homologous genes, and is able to integrate as a single copy into the Streptomyces chromosome. The resulting recombinant plasmid pLux-ssgB (Figure 2a) was conjugated to wild type (WT) S. coelicolor M145 and nine mutant strains containing deleted individual sigB homologous genes. The luminescence of eight independent KanR clones was determined during differentiation on the solid SFM medium.
In accordance with the previous results [31], in WT S. coelicolor M145 strain, the ssgBp promoter was not active in the substrate mycelium and was activated after appearance on the aerial mycelium (Figure 2b). The absence of ssgBp activity in the substrate mycelium can be explained by repression of the promoter with the transcriptional regulator BldD, which acts as a repressor of several key developmental genes during vegetative growth. Indeed, its binding site was identified in the ssgBp promoter (Figure 1a) [36]. The ssgBp promoter activity increased continuously until a maximum activity at 72 h, averaging 48,181 relative luminescence units (RLU), where the aerial mycelium was almost completely formed and some spore chains were visible. Then, during sporulation, its activity dropped dramatically (Figure 2b). In accordance with the work [31], the promoter was substantially dependent on the sigma factor SigH. However, it also was partially dependent on three other SigB homologues, SigI, SigN, and SigG. It may explain the weak activity of the ssgBp promoter in the S. coelicolor ΔsigH mutant [31]. The time course of in vivo expression of the ssgBp promoter is in accordance with a role for SsgB in the early stages of aerial mycelium septation, where SsgB localizes to septal Z-ring sites in the early stages of aerial mycelium septation [4].
In accordance with the previous results [31], the ssgBp promoter was not dependent on the sigma factor SigF. It is in contrast to our results in the heterologous two-plasmid E. coli system, where the ssgBp promoter was recognized by SigF [30]. SigF has a key role in late stages of morphological differentiation, spore maturation and pigmentation [13]. Its sigF gene is expressed only during sporulation and is spatially located in the prespore compartment of sporulating aerial hyphae [38,39]. Based on the in vivo activity of the ssgBp promoter, it was not active during this late developmental stage (Figure 2b). Therefore, its dependence in the heterologous two-plasmid system is artificial. The recognition of the ssgBp promoter by SigF in the late stages of development of S. coelicolor A3(2) may be blocked by a putative repressor activated at this stage. This could explain such a dramatic decrease in ssgBp activity at this stage. Indeed, several regulatory proteins have been found to bind to the ssgBp promoter [40,41], which may be candidates for this regulation.
In accordance with the previous transcriptional analysis of the ssgBp promoter by S1-nuclease mapping [31], the ssgBp promoter was substantially dependent on SigH. Its in vivo activity in the S. coelicolor ΔsigH mutant peaked at 92 h, averaging only 12,008 RLU. Then, like WT strain, it dramatically dropped during sporulation (Figure 2b). The shift in its maximal activity in this newly prepared S. coelicolor ΔsigH mutant strain compared to the WT strain may be due to a shift in the time course of differentiation. Unlike the previously prepared strain, S. coelicolor M145, sigH::tsr, which partially affected sporulation [22], this new S. coelicolor ΔsigH mutant strain had a similar sporulation phenotype to WT S. coelicolor M145 strain, in agreement with the results in [23]. Transcription of sigH is developmentally regulated and one of its promoters is strongly activated during aerial mycelium formation, similar to the ssgBp promoter. Interestingly, this promoter was similarly directly repressed by BldD [20,21]. Similarly, the SigH protein is also activated at this developmental stage [27,30]. These results are consistent with the expression of the ssgBp promoter and activation of SigH occurs at a similar developmental stage to its recognized ssgBp promoter.
The ssgBp promoter was also quite strongly dependent on SigI. Its in vivo activity in the S. coelicolor sigI mutant peaked at 88 h, averaging only 20,422 RLU, and decreased dramatically during sporulation (Figure 2b). However, the S. coelicolor sigI mutant was not affected in differentiation and sigI expression was constitutive during all developmental stages. In addition, SigI is regulated by a partner-switching phosphorylation mechanism with the anti-sigma factor PrsI and the anti-anti-sigma factor ArsI [17]. This shift in ssgBp activity compared to WT strain may be explained by some aberrant activation of other SigB homologues that recognize the promoter in the absence of SigI.
The ssgBp promoter was also partially dependent on SigN. Its in vivo activity in the S. coelicolor sigN mutant peaked similarly as in WT strain at 72 h, averaging 36,145 RLU, and decreased dramatically during sporulation (Figure 2b). SigN plays a role in morphological differentiation in the subapical stem region of aerial hyphae, and sigN expression is upregulated during aerial mycelium formation [14]. This time course is consistent with the ssgB expression. Therefore, it could partially participate in its regulation.
The ssgBp promoter may be partially dependent on SigG. Its in vivo activity in the S. coelicolor sigN mutant peaked at 88 h, averaging 39,817 RLU, and decreased dramatically during sporulation (Figure 2b). However, no specific role could be assigned to this sigma factor and no sigG expression was detected during differentiation [18]. However, its gene was expressed during spore germination, suggesting its role in this process [19]. This could also explain the shift in its activity compared to WT strain in the S. coelicolor sigG mutant.
As a control, the pLux-kasOp plasmid containing the strong kasOp* promoter [42] was similarly conjugated to these strains and the luciferase reporter activity was similarly measured in eight independent KanR clones. The promoter was active in the early stages of growth in substrate mycelium and its activity in WT S. coelicolor M145 strain increased to the maximum level at 40 h (on average 100,555 RLU) and then decreased continuously during differentiation. In contrast to the ssgBp promoter, the activity of the kasOp* promoter was similar in all mutant strains (Figure 2c).
2.2. Mutagenesis of the ssgBp promoter to identify specific nucleotides for Nine SigB Homologues
Almost all of the 24 promoters identified by the E. coli two-plasmid system as dependent on nine S. coelicolor A3(2) SigB homologues were recognized by two or more SigB homologues. Although analysis of these promoters suggested some distinct groups, no specific sequences characteristic for these recognition groups were identified. All promoters showed high similarity in regions ̶ 35 and ̶ 10, which were similar to the consensus sequence of promoters recognized by B. subtilis SigB (GTTTAA ̶ N12-14 ̶ GGGA/TAA/T) [30]. These results are consistent with the analysis of S. coelicolor A3(2) SigB homologues, where all were highly similar to B. subtilis SigB, mainly in regions 2.4 and 4.2, which are involved in the recognition of the ̶ 10 and ̶ 35 promoter regions [10]. Although there was high similarity in both promoter regions, the ̶ 10 region contains almost invariant first three nucleotides (GGG) but quite variable three following nucleotides [30].
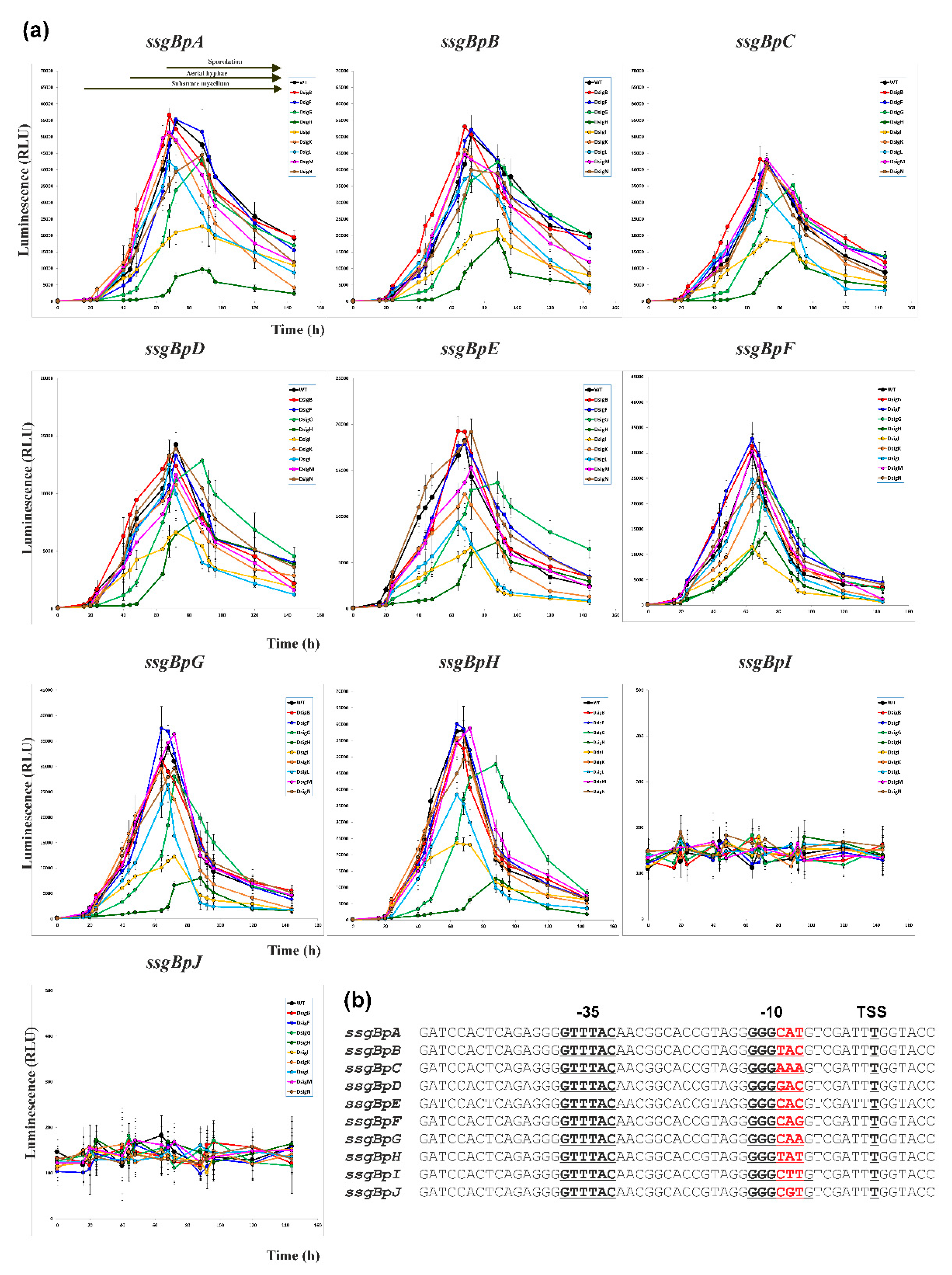
To investigate the role of these nucleotides in the ̶ 10 region of the ssgBp promoter, we prepared several mutants in this region (Figure 3a) and analyzed the mutated ssgBp promoters in the E. coli two-plasmid system. All promoters were constructed using a mixture of complementary primers covering the 55 nt core of the ssgBp promoter and the resulting DNA fragments were inserted in pSB40N [43], resulting in pSB40N-ssgBpA, pSB40N-ssgBpB, pSB40N-ssgBpC, pSB40N-ssgBpD, pSB40N-ssgBpE, pSB40N-ssgBpF, pSB40N-ssgBpG, pSB40N-ssgBpH, pSB40N-ssgBpI, and pSB40N-ssgBpJ. The original WT ssgBpA ̶ 10 region (TGGCAT) (Figure 1a) was changed to the more conservative GGGCAT (Figure 3a). Subsequently, all plasmids containing WT ssgBpA and mutated (ssgBpB-J) promoters were transformed into E. coli containing a compatible expression pAC plasmid with a particular sigma factor gene (pAC-sigB, pAC-sigF1, pAC-sigF2, pAC-sigG, pAC-sigH1, pACsigH2, pAC-sigI, pAC-sigK, pAC-sigL, pAC-sigM, pAC-sigN) and promoter dependence on sigma factors screened on MacConkey agar plates as previously described [30]. The results showed that six mutant promoters (ssgBpD, E, F, G, I, J) were not recognized by any sigma factor in the E. coli two-plasmid system (Figure 3b; Supplementary Figure S1).
The WT ssgBpA promoter (with the ̶ 10 GGGCAT region) was recognized by full-length sigma factor SigF (pAC-sigF1) and N-terminally truncated SigF (pAC-sigF2), similarly to plasmid pF81 with a 750-bp DNA fragment covering the entire ssgBp promoter region [30]. These results confirmed the previous hypothesis that this short N-terminal repeat region in SigF has no function for its activity. However, the ssgBpA promoter was also recognized by two other sigma factors, SigH1 and SigL, confirming the dependence of the promoter on SigH (as described above). This could be explained by the higher activity of the promoter in the E. coli two-plasmid system due to the presence of the conserved GGG in the ̶ 10 region. Furthermore, these results are consistent with previous data that only the short form of SigH (pAC-sigH1) is active and the larger form of SigH (pAC-sigH2) is inactive, suggesting some inhibitory effect of this N-terminal region on sigma factor function [30]. Therefore, like many other promoters dependent on SigB homologues of S. coelicolor A3(2) [30], ssgBpA is cross-recognized by several SigB homologues (SigF, SigH, SigL).
Interestingly, the ssgBpB mutant promoter (with the ̶ 10 GGGTAC region) was recognized by the sigma factors SigB, SigF, SigL and SigM. Therefore, changing the last three nucleotides of the ̶ 10 region from GGGCAT to GGGTAC dramatically altered promoter recognition. This change caused the ssgBpB promoter to be recognized not by SigH but by two other SigB homologues, SigB and SigM. However, this change did not alter the recognition by SigF and SigL.
The ssgBpC mutant promoter (with the ̶ 10 GGGAAA region) was recognized by the sigma factors SigB, SigF, and SigL. Therefore, changing the last three nucleotides of the ̶ 10 region from GGGCAT to GGGAAA similarly altered promoter recognition. This change caused the ssgBpC promoter to be recognized not by SigH but by SigB. Like the previous mutant promoter, this change did not alter the recognition by SigF and SigL.
The ssgBpH mutant promoter (with the ̶ 10 GGGTAT region) was recognized by the sigma factors SigB, SigF, SigH, and SigL. Interestingly, unlike the previous mutant promoters, changing the last three nucleotides of the ̶ 10 region from GGGCAT to GGGTAT did not affect recognition by SigF, SigH, and SigL, but the ssgBpH promoter was also recognized by SigB.
These results suggest a critical role for the last three nucleotides for promoter recognition by SigB homologues of S. coelicolor A3(2). Because the A nucleotide at the fifth position is highly conserved and essential for the promoter activity, as described below, the two variable nucleotides at the fourth and sixth positions in the ̶ 10 region are likely responsible for the partial selectivity of promoter recognition by several SigB homologues. SigH prefers GGGCAT but also recognizes GGGTAT. SigB recognizes GGGTAC, GGGAAA, and GGGTAT, but the presence of C at the fourth position in the ̶ 10 region prevents its recognition. SigM only recognizes GGGTAC and the presence of any other nucleotide at the sixth position in the ̶ 10 region prevents its recognition. Interestingly, SigL and SigF recognize all four regions analyzed, GGGCAT, GGGTAC, GGGAAA, and GGGTAT, and are therefore less selective in recognizing the ̶ 10 region.
Considering the cross-recognition table for 24 promoters dependent on SigB homologues [30], these conclusions are partially applicable to these SigB homologues. However, the ̶ 35 region also has a certain specific role for this selectivity, although analysis of this region in promoters did not reveal any specific sequence for SigB homologues. This region has the consensus sequence GTTTNN, where the last two nucleotides are highly variable [30]. Therefore, both regions are likely to play a role in this selectivity. Indeed, the representative B. subtilis SigB-dependent promoter Pctc (GTTTAA ̶ N14 ̶ GGGTAT) was active with all nine S. coelicolor A3(2) SigB homologues in the E. coli two-plasmid system [30].
2.3. In vivo Cross-Recognition of mutant ssgBp promoters by Nine SigB Homologues
To examine the dependence of mutated ssgBp promoters on SigB homologues in vivo in S. coelicolor A3(2), we inserted the ssgBpA-J promoters (Figure 3a) as 55-bp BamHI-KpnI DNA fragments into the luciferase reporter plasmid pLux-kasOp (Figure 2a) digested with the same restriction enzymes. The resulting recombinant plasmids (pLux-ssgBpA, pLux-ssgBpB, pLux-ssgBpC, pLux-ssgBpD, pLux-ssgBpE, pLux-ssgBpF, pLux-ssgBpG, pLux-ssgBpH, pLux-ssgBpI, pLux-ssgBpJ) were conjugated to WT S. coelicolor M145 and nine mutant strains containing deleted individual sigB homologous genes. The luminescence of eight independent KanR clones was determined during differentiation on the solid SFM medium (Figure 4).
Similar to the pLux-ssgBp plasmid containing a 750-bp DNA fragment covering the entire ssgBp promoter region (Figure 2), the ssgBpA promoter was also not active in the substrate mycelium and was activated after the appearance of the aerial mycelium in WT S. coelicolor M145 strain. Its activity similarly increased to a maximum activity at 72 h, averaging 54,636 RLU. This activity was higher than that of the pLux-ssgBp plasmid, indicating a higher activity of this core promoter with an altered, more conserved GGG in the ̶ 10 region. Interestingly, unlike pLux-ssgBp, the decrease of the ssgBpA promoter activity during sporulation was not as dramatic (Figure 4). Similar to the pLux-ssgBp plasmid, the ssgBpA promoter was most dependent on SigH (maximum at 88 h, on average 9,722 RLU) and SigI (maximum at 88 h, on average 22,836 RLU), and partially on SigN (maximum at 88 h, on average 44,494 RLU) and SigG (maximum at 88 h, on average 43,242 RLU). However, it also was partially dependent on SigL (maximum at 68 h, on average 42,446 RLU). Similar to pLux-ssgBp plasmid, the ssgBpA promoter was not dependent on SigF, although it was recognized by this sigma factor in the E. coli two-plasmid system.
Two mutant promoters, ssgBpI (with the ̶ 10 GGGCTT region) and ssgBpJ (with the ̶ 10 GGGCGT region), were not active in any strain used. Their activity was at the level of background activity (approx. 150 RLU) (Figure 4). Both promoters were also not recognized by any sigma factor in the E. coli two-plasmid system. These results indicated an essential nucleotide A in the ̶ 10 region for promoter recognition by all nine SigB homologues. This nucleotide A was indeed highly conserved in the identified 24 promoters dependent on nine SigB homologues in S. coelicolor A3(2) and also in the consensus sequence of promoters recognized by B. subtilis SigB (GTTTAA ̶ N12-14 ̶ GGGA/TAA/T) [30].
The ssgBpB mutant promoter (with the ̶ 10 GGGTAC region), which was dependent on four sigma factors (SigB, SigF, SigL, SigM) in the E. coli two-plasmid system, was also not active in the substrate mycelium and was activated after appearance of the aerial mycelium in WT S. coelicolor M145 strain. Similar to the ssgBpA promoter, its activity increased to a maximum activity at 72 h, averaging 50,462 RLU. This activity was similar as ssgBpA (54,636 RLU), and similarly decreased during sporulation (Figure 4). Interestingly, consistent with the results in the E. coli two-plasmid system, the ssgBpB promoter was less dependent on SigH than ssgBpA (maximum at 88 h, on average 18,979 RLU) but similarly dependent on SigI (maximum at 88 h, on average 21,879 RLU). Similarly, it was partially dependent on SigN (maximum at 72 h, on average 39,917 RLU), SigG (maximum at 88 h, on average 42,338 RLU) and SigL (maximum at 72 h, on average 38,610 RLU). Consistent with the results in the E. coli two-plasmid system, the promoter was partially dependent on SigM (maximum at 68 h, on average 44,557 RLU). However, in contrast to these results, it was not dependent on SigB and SigF. In the case of SigF, this is probably an artificial dependence, as in the case of the WT ssgBp promoter (as described above). In the case of SigB, it is probably also an artificial dependence, since SigB is not active during sporulation on solid media and is only activated by osmotic stress [30].
The ssgBpC mutant promoter (with the ̶ 10 GGGAAA region), which was dependent on three sigma factors (SigB, SigF, SigL) in the E. coli two-plasmid system, was also not active in the substrate mycelium and was activated after the appearance of the aerial mycelium in WT S. coelicolor M145 strain. Similar to the ssgBpA promoter, its activity increased to a maximum activity at 72 h, averaging 42,461 RLU. This activity was lower than ssgBpA (54,636 RLU) but similarly decreased during sporulation (Figure 4). Consistent with the results in the E. coli two-plasmid system, the ssgBpB promoter was less dependent on SigH than ssgBpA (maximum at 88 h, averaging 15,550 RLU), but similarly dependent on SigI (maximum at 72 h, averaging 18,739 RLU). Consistent with the results in the E. coli two-plasmid system, the promoter was partially dependent on SigL (maximum at 68 h, on average 33,638 RLU). Like the ssgBpA promoter, also the ssgBpC promoter was partially dependent on SigG (maximum at 88 h, on average 35,333 RLU). Unlike ssgBpA, however, the ssgBpC promoter was not dependent on SigN (maximum at 72 h, on average 42,028 RLU). Similar to the ssgBpB promoter, in contrast to the results of the E. coli two-plasmid system, the promoter was also not dependent on SigB and SigF, and this dependence is artificial as described above.
The ssgBpH mutant promoter (with the ̶ 10 GGGTAT region), which was dependent on four sigma factors (SigB, SigF, SigH, SigL) in the E. coli two-plasmid system, was also not active in the substrate mycelium and was activated after appearance of the aerial mycelium in WT S. coelicolor M145 strain. Similar to the ssgBpA promoter, its activity increased to a maximum activity at 68 h, averaging 58,093 RLU. This activity was higher than ssgBpA (54,636 RLU) but similarly decreased during sporulation (Figure 4). This higher activity of the ssgBpH promoter can be explained by higher similarity of the ̶ 10 region with the consensus sequence of promoters recognized by B. subtilis SigB (GTTTAA ̶ N12-14 ̶ GGGA/TAA/T) [30]. Consistent with the results in the E. coli two-plasmid system, the ssgBpH promoter was more dependent on SigH (maximum at 88 h, on average 12,621 RLU) and partially dependent on SigL (maximum at 64 h, on average 38,326 RLU). Like ssgBpA, it was similarly more dependent on SigI (maximum at 64 h, on average 23,533 RLU) and partially on SigN (maximum at 68 h, on average 49,073 RLU) and SigG (maximum at 88 h, on average 47,752 RLU). Similar to the ssgBpB promoter, in contrast to the results of the E. coli two-plasmid system, the promoter was also not dependent on SigB and SigF, and this dependence is artificial, as describe above.
The latter mutant promoters (ssgBpD,E,F,G) were not dependent on any sigma factor in the E. coli two-plasmid system. They were not active in the substrate mycelium and activated after the appearance of the aerial mycelium in all strains. The activity of the ssgBpD promoter (with the ̶ 10 GGGGAC region) increased to a maximum activity at 72 h, averaging 14,255 RLU. This activity was substantially lower than ssgBpA (54,636 RLU) but similarly decreased during sporulation (Figure 4). Unlike the previously analyzed promoters, it was only partially dependent on SigH (maximum at 88 h, on average 8,203 RLU) and SigI (maximum at 72 h, on average 6,632 RLU). The activity of the ssgBpE promoter (with the ̶ 10 GGGCAC region) increased to a maximum activity at 68 h, averaging 18,242 RLU. Again, this activity was substantially lower than ssgBpA (54,636 RLU) but similarly decreased during sporulation (Figure 4). The promoter was partially dependent on SigH (maximum at 88 h, on average 7,247 RLU), SigI (maximum at 72 h, on average 6,661 RLU), SigL (maximum at 64 h, on average 9,372 RLU), SigG (maximum at 88 h, on average 13,685 RLU) and SigK (maximum at 68 h, on average 12,415 RLU). The activity of the ssgBpF promoter (with the ̶ 10 GGGCAG region) increased to a maximum activity at 64 h, averaging 29,781 RLU. This activity was lower than ssgBpA (54,636 RLU) but similarly decreased during sporulation (Figure 4). In contrast to the previously analyzed promoters, it was only partially dependent on SigH (maximum at 72 h, on average 14,091 RLU) and SigI (maximum at 64 h, on average 11,465 RLU). The activity of the ssgBpG promoter (with the ̶ 10 GGGCAA region) increased to a maximum activity at 68 h, averaging 33,683 RLU. This activity was lower than ssgBpA (54,636 RLU) but similarly decreased during sporulation (Figure 4). The promoter was substantially dependent on SigH (maximum at 88 h, on average 7,980 RLU) and SigI (maximum at 72 h, on average 12,216 RLU) and partially dependent on SigL (maximum at 68 h, on average 26,381 RLU).
These results partially confirmed the dependence of these promoters on SigB homologues in the E. coli two-plasmid system. Indeed, both the ssgBpA and ssgBpH promoters were most dependent on SigH in vivo in S. coelicolor A3(2). However, all other mutant promoters (except inactive ssgBpI and ssgBpJ) were still partially dependent on this sigma factor. Therefore, as discussed above, both promoter regions play a role in this selectivity, and some nucleotides in the ̶ 35 region allow partial recognition of these mutant promoters. All four promoters that were dependent on SigL in the E. coli two-plasmid system (ssgBpA,B,C,H), were partially dependent on this sigma factor in vivo in S. coelicolor A3(2). In addition, the ssgBpB promoter was partially dependent on SigM, consistent with the results in the E. coli two-plasmid system. In contrast to the results of the E. coli two-plasmid system, the mutant promoters were not dependent on SigB and SigF in vivo in S. coelicolor A3(2) and this dependence is artificial, as describe above.
Similar to the pLux-ssgBp plasmid containing a 450-bp DNA fragment covering the entire ssgBp promoter region (Figure 2), all ssgBpA-G mutant promoters were also quite strongly dependent on SigI in vivo in S. coelicolor A3(2) (Figure 4). It was unexpected because this sigma factor was only active with one of the 24 SigB homologue-dependent promoters analyzed in the E. coli two-plasmid system, and this SCO2026p promoter was additionally cross-regulated by four other SigB homologues, SigB, SigF, SigH, SigL, although it contains ̶ 10 GGGCAT region [30]. As mentioned above, this region was preferred by SigH in the E. coli two-plasmid system with ssgBp mutant promoters. As noted above, sigI expression was constitutive during all developmental stages, and SigI is regulated with the anti-sigma factor PrsI and the anti-anti-sigma factor ArsI [17]. These results indicate that SigI is less selective for the ̶ 10 region, but its strong dependence lies in another region, probably the ̶ 35 region of the ssgBp promoter. Similarly, some of the mutant promoters were partially dependent on SigG (ssgBpA,B,C,E,F,G,H) and SigN (ssgBpA,B,F,H) in vivo in S. coelicolor A3(2), as noted above for ssgBp promoter in the pLux-ssgBp plasmid. Both sigma factors recognize several promoters of the 24 SigB homologue-dependent promoters analyzed in the E. coli two-plasmid system, which were additionally cross-regulated by several SigB homologues, and the promoters contain variable ̶ 10 regions [30]. Therefore, these results similarly suggest that both sigma factors are less selective for the ̶ 10 region and this partial dependence may lie in another region, probably the ̶ 35 region of the ssgBp promoter.
In vivo activity of the mutant promoters suggests a role for particular nucleotides in the ̶ 10 region. As noted above, nucleotide A at the fifth position is highly conserved and essential for the promoter activity. The two variable nucleotides at the fourth and sixth positions in the ̶ 10 region, which are likely responsible for the partial promoter selectivity, also affected promoter activity. The ssgBpH promoter with the ̶ 10 GGGTAT region, which was identical to the consensus sequence of the B. subtilis SigB-dependent promoter, had the highest activity (Figure 4). Promoters containing a pyrimidine nucleotide T or C in these positions (GGGCAT, GGGTAC) had slightly lower activity. However, the ssgBpE promoter with two Cs in these positions (GGGCAC) had dramatically reduced activity by approximately 70% (Figure 4). Therefore, the presence of a single T (more similar to the consensus sequence GGGTAT) in these positions had a substantially positive effect on promoter activity. Replacing both Ts with As (GGGAAA) in the ssgBpC promoter reduced activity by approximately 26% (Figure 4). However, substitution of A for C in this region (GGGCAA) in the ssgBpG promoter further reduced activity by approximately 44% (Figure 4), indicating that A is more active than C. The presence of C and G in these positions had a significant negative effect on promoter activity. The ssgBpF promoter (GGGCAG) had reduced activity by approximately 50% and the ssgBpD promoter (GGGGAC) by approximately 76% (Figure 4).
In conclusion, these results indicate a critical role of the last three nucleotides for promoter recognition and selectivity by SigB homologues and overlapping promoter recognition by these SigB homologues in vivo in S. coelicolor A3(2). Nucleotide A at position 5 is highly conserved and essential for promoter activity, and two variable nucleotides at positions 4 and 6 in the ̶ 10 region are likely responsible for the partial selectivity of promoter recognition by SigB homologues.
3. Materials and Methods
3.1. Bacterial strains, culture conditions, and plasmids
All strains and plasmids used in this study are listed in Supplementary Table S1. Conditions for growth and transformation of E. coli strain were described in [44]. Luria-Bertani (LB) medium was used for the growth. When necessary, media were supplemented with 50 μg/ml kanamycin (Kan) (SERVA, Heidelberg, Germany), 40 μg/ml chloramphenicol (Clm) (Sigma-Aldrich, Darmstadt, Germany), 100 μg/ml ampicillin (Amp) (Sigma-Aldrich, Darmstadt, Germany), 50 μg/ml apramycin (Apr) (Sigma-Aldrich, Darmstadt, Germany), and 20 μg/ml nalidixic acid (NA) (Sigma-Aldrich, Darmstadt, Germany). The strain E. coli XL1-Blue was used to detect dependence of promoters on sigma factors by the two-plasmid system. The E. coli colonies containing sigma factor-dependent promoters were screened on MacConkey agar plates (BIOMARK Laboratories, India) with Amp, Clm, and 1 mM isopropyl-β-D-thiogalactopyranoside (IPTG) [30]. E. coli DH5α strain (ThermoFisher Scientific, Waltham, MA, USA) was used for standard cloning experiments. Non-methylating E. coli ET12567/pUZ8002 was used to transfer plasmids by conjugation from E. coli into Streptomyces strains [45]. Growth and sporulation of S. coelicolor A3(2) were performed as described in [45]. Rich solid medium SFM [45] was used for growth of Streptomyces strains to various developmental stages; 24 h – vegetative substrate mycelium, 40 h – beginning of aerial mycelium formation, 64 h – septation of aerial hyphae, 88 h – spore maturation.
3.2. Recombinant DNA techniques
Standard DNA manipulation methods were performed as described in [44]. To construct recombinants plasmids containing mutated ssgBpA-J promoters, two complementary primers for a specific promoter, ssgBpA-JDir and ssgBpA-Jrev (Supplementary Table S2) were annealed (mixed 20 pmol of each primer in water, incubate for 2 min at 100 °C and allow to cool to room temperature for 15 min), mixed with 100 ng of pSB40N digested with BamHI and XhoI and ligated by the standard method as described in [44]. The mixtures were transformed into E. coli DH5a with AmpR selection, resulting in pSB40N-ssgBpA, pSB40N-ssgBpB, pSB40N-ssgBpC, pSB40N-ssgBpD, pSB40N-ssgBpE, pSB40N-ssgBpF, pSB40N-ssgBpG, pSB40N-ssgBpH, pSB40N-ssgBpI, and pSB40N-ssgBpJ. The nucleotide sequences of all constructs were verified by sequencing using the -47 primer (Supplementary Table S2).
3.3. Detection of E. coli clones containing ssgBp promoters and cross-recognition by nine SigB homologous sigma factors
The procedure is described in [30]. To examine the cross-recognition of WT or mutant ssgBp promoters by nine SigB homologous sigma factors (SigB, SigF, SigG, SigH, SigI, SigK, SigL, SigM, SigN) from S. coelicolor A3(2), a specific plasmid containing the promoter DNA fragment (inserted in pBS40N) was transformed into E. coli strain XL1-Blue containing a compatible expression plasmid with the sigma factor gene under the control of the trc promoter (pAC-sigB, pAC-sigF1, pAC-sigF2, pAC-sigG, pAC-sigH1, pAC-sigH2, pAC-sigI, pAC-sigK, pAC-sigL, pAC-sigM, pAC-sigN) or pAC5mut2 expression vector as a negative control. Transformation mixtures were plated on MacConkey plates with Amp, Cm, and 1 mM IPTG a grown for 72 h at 37 °C.
3.4. Construction of deletion mutants in S. coelicolor A3(2)
Construction of mutants with the deleted sigH, sigK, sigL, sigM, sigN genes in S. coelicolor M145 was carried out using a PCR targeting REDIRECT procedure [46]. A disruption cassette containing oriT and AprR gene was prepared by PCR with primers SigHD1Dir and SigHD1Rev for sigH, SigKD1Dir and SigKD1Rev for sigK, SigLD1Dir and SigLD1Rev for sigL, SigMD1Dir and SigMD1Rev for sigM, SigND1Dir and SigND1Rev for sigN (Supplementary Table S2) from the upstream and downstream regions of a specific gene using a gel-purified 1384-bp EcoRI-HindIII fragment from pIJ773 as a template. 1 μg of the amplified DNA fragment purified on a Wizard column (Promega, USA) was electroporated into E. coli BW25113 containing plasmid pIJ790 and a specific SuperCos1-derived cosmid for each sigma factor gene, 2St7G11 for sigH, St5C7 for sigK, St5H1 for sigL, St55F8 for sigM and 2St5D60 for sigN [47]. The resulting verified mutant cosmid was transformed into E. coli ET12567/pUZ8002 and the deleted allele was moved to S. coelicolor M145 by conjugation. Colonies were screened for AprR and sensitivity to Kan, indicating a double crossover. Three or four such colonies with a similar phenotype were selected and confirmed by Southern blot hybridization analysis (data not shown). One such confirmed colony for each sigma factor gene was selected for further studies, resulting in S. coelicolor ΔsigH, S. coelicolor ΔsigK, S. coelicolor ΔsigL, S. coelicolor ΔsigM, and S. coelicolor ΔsigN. The phenotype of the mutants grown on solid SFM medium was similar to that of WT S. coelicolor M145 (Supplementary Figure S2)
3.5. Construction of a New luxCDABE-based luciferase reporter plasmid and Bioluminescence Measurement
A new PhiC31 phage-based integration plasmid was constructed containing a synthetic luxCDABE operon from plasmid pMU1s* [37] and KanR gene compatible for selection with AprR of S. coelicolor mutants in sigB homologous genes. A 6,3-kb BamHI-EcoRI fragment from pMU1s-kasOp1 was cloned into pSB40BR5-phiC31int digested with BamHI and MfeI, resulting in pLux-kasOp (Figure 2a). The ssgBp promoter was inserted as a 750-bp BamHI-XhoI DNA fragment from pF81 into pBluescript II SK digested with the same enzymes, resulting in pSsgBp1A. Subsequently, the ssgBp DNA fragment was cloned from this plasmid as a 450-bp BamHI-KpnI DNA fragment into pLux-kasOp digested with the same enzymes, resulting in pLux-ssgBp (Figure 2a).
The ssgBpA-J mutant promoters were inserted as 55-bp BamHI-KpnI DNA fragments from pSB40N-ssgBpA, pSB40N-ssgBpB, pSB40N-ssgBpC, pSB40N-ssgBpD, pSB40N-ssgBpE, pSB40N-ssgBpF, pSB40N-ssgBpG, pSB40N-ssgBpH, pSB40N-ssgBpI and pSB40N-ssgBpJ into pLux-kasOp digested with the same enzymes, resulting in pLux-ssgBpA-J. The nucleotide sequence of all the recombinant plasmids was checked by sequencing using the luxCrev primer (Supplementary Table S2). The resulting recombinant plasmids were conjugated to WT S. coelicolor M145 and nine mutant strains containing deleted individual sigB homologous genes with KanR selection. Eight independent KanR clones from each construct were picked and sporulated twice on solid SFM medium with Kan and NA and finally on solid SFM medium with Kan. Spores from each clone, approximately 105 CFU, were inoculated into each well of white 96-well plates (Sigma-Aldrich, Darmstadt, Germany) containing 250 μl of solid SFM medium and grown into a confluent lawn. Plates were incubated at 28 °C and luminescence in RLU was measured in a Synergy HT microplate reader (Bio-Tek Instruments, Winooski, VT, USA). The background luminescence of the well inoculated with S. coelicolor M145 strain alone was approximately 150 RLU.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: Bacterial strains and plasmids used in this study; Table S2: Oligonucleotides used in this study; Figure S1: Cross-recognition of the ssgBpA-J promoters with all nine SigB homologous sigma factors from S. coelicolor A3(2) in the E. coli two-plasmid system; Figure S2: Phenotypic analysis of S. coelicolor sigma factor mutant strains.
Author Contributions
JK conceived and designed research. BS, RN, FO, RJ, DC and LF conducted experiments. BC, RN and JK analyzed data. BS and JK wrote the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by 2/0026/20 grant from the Slovak Academy of Sciences.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are available on request.
Acknowledgments
We are grateful to Dr. Bertold Gust (John Innes Centre, Norwich, UK) for kindly providing all the plasmids and strains used in PCR-targeting REDIRECT system; the system has been supplied by Plant Bioscience Ltd. (Norwich, UK). We thank Helen Kieser (John Innes Centre, Norwich, UK) for the cosmids 2St7G11, St5C7, St5H1, St55F8 and 2St5D60 and and to Justin Nodwell (McMaster University, Ontario, Canada) for the plasmid pMU1s*.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| aa | amino acid(s) |
| Amp | ampicillin |
| Apr | apramycin |
| AprR | Apr resistance |
| B | Bacillus |
| CFU | colony forming units |
| Clm | chloramphenicol |
| E | Escherichia |
| h | hour(s) |
| IPTG | isopropyl-β-D-thiogalactopyranoside |
| Kan | kanamycin |
| KanR | kanamycin resistance |
| kb | kilobase |
| LB | Luria-Bertani (medium) |
| NA | nalidixic acid |
| nt | nucleotide(s) |
| PCR | polymerase chain reaction |
| RLU | relative luminescence unit(s) |
| S | Streptomyces |
| TSS | transcription start site |
| WT | wild type |
References
- Chater, K.F. Differentiation in Streptomyces: the properties and programming of diverse cell types. In Streptomyces: molecular biology and biotechnology; Dyson, P.J., Ed.; Caister Academic Press: Norfolk, UK, 2011; pp. 43–86. [Google Scholar]
- McCormick, J.R.; Flardh, K. Signals and regulators that govern Streptomyces development. FEMS Microbiol. Rev. 2012, 36, 206–231. [Google Scholar] [CrossRef] [PubMed]
- Schlimpert, S.; Wasserstrom, S.; Chandra, G.; Bibb, M.J.; Findlay, K.C.; Flärd, K.; Buttner, M.J. Two dynamin-like proteins stabilize FtsZ rings during Streptomyces sporulation. Proc. Natl. Acad. Sci. U.S.A. 2017, 114, E6176–E6183. [Google Scholar] [CrossRef] [PubMed]
- Willemse, J.; Borst, J.W.; de Waal, E.; Bisseling, T.; van Wezel, G.P. Positive control of cell division: FtsZ is recruited by SsgB during sporulation of Streptomyces. Genes Dev. 2011, 25, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Bush, M.J.; Tschowri, N.; Schlimpert, S.; Flard, K.; Buttner, M.J. c-di-GMP signaling and the regulation of developmental transitions in streptomycetes. Nat. Rev. Microbiol. 2015, 13, 749–760. [Google Scholar] [CrossRef]
- Gallagher, K.A.; Schumacher, M.A.; Bush, M.J.; Bibb, M.J.; Chandra, G.; Holmes, N.A.; Zeng, W.; Henderson, M.; Zhang, H.; Findlay, K.C.; Brennan, R.G.; Buttner, M.J. c-di-GMP arms an anti-σ to control progression of multicellular differentiation in Streptomyces. Mol Cell 2020, 77, 586–599. [Google Scholar] [CrossRef]
- van der Heul, H.U.; Bilyk, B.L.; McDowall, K.J.; Seipke, R.F.; van Wezel, G.P. ; Regulation of antibiotic production in Actinobacteria: new perspectives from the post-genomic era. Nat. Prod. Rep. 2018, 35, 575–604. [Google Scholar] [CrossRef]
- Vohradsky, J.; Li, X.M.; Dale, G.; Folcher, M.; Nguyen, L. , Viollier, P.H.; Thompson, C.J. Developmental control of stress stimulons in Streptomyces coelicolor revealed by statistical analysis of global gene expression patters. J. Bacteriol. 2000, 182, 4979–4986. [Google Scholar] [CrossRef]
- Ayala, F.R.; Bartolini, M.; Grau, R. The stress-responsive alternative sigma factor SigB of Bacillus subtilis and its relatives: an old friend with new functions. Front. Microbiol. 2020, 11, 1761. [Google Scholar]
- Kormanec, J.; Sevcikova, B.; Novakova, R.; Homerova, D.; Rezuchova, B.; Mingyar, E. The complex regulatory network in the regulation of stress-response sigma factors in Streptomyces coelicolor A3(2). In Stress and Environmental Regulation of Gene Expression and Adaptation in Bacteria; de Bruijn, F.J., Ed.; John Wiley & Sons, Inc: NJ, 2016; pp. 321–336. [Google Scholar]
- Bentley, S.D.; Chater, K.F.; Cerdeno-Tarraga, A.M.; Challis, G.L.; Thomson, N.R.; James, K.D.; Harris, D.E.; Quail, M.A.; Kieser, H.; Harper, D.; Bateman, A.; Brown, S.; Chandra, G.; Chen, C.; Collins, M.; Cronin, A.; Fraser, A.; Goble, A.; Hidalgo, J.; Hornsby, T.; Howarth, S.; Huang, C.H.; Kieser, T.; Larke, L.; Murphy, L.; Oliver, K.; O’Neil, S.; Rabbinowitsch, E.; Rajandream, M.A.; Rutherford, K.; Rutter, S.; Seeger, K.; Saunders, D.; Sharp, S.; Squares, R.; Squares, S.; Taylor, K.; Warren, T.; Wietzorrek, A.; Woodward, J.; Barrell, B.G.; Parkhill, J.; Hopwood, D.A. Complete genome sequence of the model actinomycete Streptomyces coelicolor A3(2). Nature 2002, 417, 141–147. [Google Scholar] [CrossRef]
- Mittenhuber, G. A phylogenetic study of the general stress response sigma factor σB of Bacillus subtilis and its regulatory proteins. J. Mol. Microbiol. 2002, 4, 427–452. [Google Scholar]
- Potuckova, L.; Kelemen, G.H.; Findlay, K.C.; Lonetto, M.A.; Buttner, M.J.; Kormanec, J. A new RNA polymerase sigma factor, σF, is required for the late stages of morphological differentiation in Streptomyces spp. Mol. Microbiol. 1995, 17, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Dalton, K.A.; Thibessard, A.; Hunter, J.I.; Kelemen, G.H. A novel compartment, the ‘subapical stem’of the aerial hyphae, is the location of a sigN-dependent, developmentally distinct transcription in Streptomyces coelicolor. Mol. Microbiol. 2007, 64, 719–737. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.-M.; Zhou, Z.; Hou, X.-P.; Guan, W.-J.; Li, Y.-Q. Reciprocal regulation between sigK and differentiation programs in Streptomyces coelicolor. J. Bacteriol. 2009, 91, 6473–6481. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.J.; Karoonuthaisiri, N.; Kim, H.S.; Park, J.H.; Cha, C.J.; Kao, C.M.; Roe, J.H. A master regulator σB governs osmotic and oxidative response as well as differentiation via a network of sigma factors in Streptomyces coelicolor. Mol. Microbiol. 2005, 57, 1252–1264. [Google Scholar] [CrossRef]
- Homerova, D.; Sevcikova, B.; Rezuchova, B.; Kormanec, J. Regulation of an alternative sigma factor σI by a partner switching mechanism with an anti-sigma factor PrsI and an anti-anti-sigma factor ArsI in Streptomyces coelicolor A3(2). Gene 2012, 492, 71–80. [Google Scholar] [CrossRef]
- Kormanec, J.; Homerova, D.; Barak, I.; Sevcikova, B. A new gene, sigG, encoding a putative alternative sigma factor of Streptomyces coelicolor A3(2). FEMS Microbiol. Lett. 1999, 172, 153–158. [Google Scholar] [CrossRef]
- Strakova, E.; Zikova, A.; Vohradsky, J. Interference of sigma factor controlled networks by using numerical modeling applied to microarray time series data of the germinating prokaryote. Nucleic Acids Res. 2013, 42, 748–763. [Google Scholar] [CrossRef]
- Kelemen, G.H.; Viollier, P.; Tenor, J.L.; Marri, L.; Buttner, M.J.; Thompson, C.J. A connection between stress and development in the multicellular prokaryote Streptomyces coelicolor A3(2). Mol. Microbiol. 2001, 40, 804–814. [Google Scholar] [CrossRef]
- Kormanec, J.; Sevcikova, B.; Halgasova, N.; Knirschova, R.; Rezuchova, B. Identification and transcriptional characterization of the gene encoding the stress- response sigma factor σH in Streptomyces coelicolor A3(2). FEMS Microbiol. Lett. 2000, 189, 31–38. [Google Scholar]
- Sevcikova, B.; Benada, O.; Kofronova, O.; Kormanec, J. Stress-response sigma factor σH is essential for morphological differentiation of Streptomyces coelicolor A3(2). Arch. Microbiol. 2001, 177, 98–106. [Google Scholar] [CrossRef]
- Viollier, P.H.; Kelemen, G.H.; Dale, G.E.; Nguyen, K.T.; Buttner, M.J.; Thompson, C.J. Specialized osmotic stress response systems involve multiple SigB-like sigma factors in Streptomyces coelicolor. Mol. Microbiol. 2003. 47, 699–714. [CrossRef]
- Lee, E.J.; Cho, Y.H.; Kim, H.S.; Ahn, B.E.; Roe, J.H. Regulation of σB by an anti- and anti-anti-sigma factor in Streptomyces coelicolor in response to osmotic stress. J. Bacteriol. J. Bacteriol. 2004, 186, 8490–8498. [Google Scholar] [CrossRef] [PubMed]
- Gaskell, A.A.; Crack, J.C.; Kelemen, G.H.; Hutchings, M.I.; Le Brun, N.E. RmsA is an anti-sigma factor that modulates its activity through [2Fe-2S] cluster cofactor. J. Biol. Chem. 2007, 282, 31812–31820. [Google Scholar] [CrossRef] [PubMed]
- Sevcikova, B.; Kormanec, J. Activity of the Streptomyces coelicolor stress-response sigma factor σH is regulated an anti-sigma factor. FEMS Microbiol. Lett. 2002, 209, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Viollier, P.H.; Weihofen, A.; Folcher, M.; Thompson, C.J. Post-translational regulation of the Streptomyces coelicolor stress responsive sigma factor, SigH, involves translational control, proteolytic processing, and an anti-sigma factor homolog. J. Mol. Biol. 2003, 325, 637–649. [Google Scholar] [CrossRef] [PubMed]
- Sevcikova, B.; Rezuchova, B.; Homerova, D.; Kormanec, J. The anti-anti sigma factor BldG is involved in activation of stress-response sigma factor σH in Streptomyces coelicolor A3(2). J. Bacteriol. 2010, 192, 5674–5681. [Google Scholar] [CrossRef]
- Sevcikova, B.; Rezuchova, B.; Mingyar, E.; Homerova, D.; Novakova, R.; Feckova, L.; Kormanec, J. Pleiotropic anti-anti-sigma factor BldG is phosphorylated by several anti-sigma factor kinases in the process of activating multiple sigma factors in Streptomyces coelicolor A3(2). Gene 2020, 755, 144883. [Google Scholar] [CrossRef]
- Sevcikova, B.; Rezuchova, D.; Mazurakova, V.; Homerova, D.; Novakova, R.; Feckova, L.; Kormanec, J. Cross-recognition of promoters by the nine SigB homologues present in Streptomyces coelicolor A3(2). Int. J. Mol. Sci. 2021, 22, 7849. [Google Scholar] [CrossRef]
- Kormanec, J.; Sevcikova, B. The stress-response sigma factor σH controls the expression of ssgB, a homologue of the sporulation-specific cell division gene ssgA in Streptomyces coelicolor A3(2). Mol. Gen. Genet. 2002, 267, 536–543. [Google Scholar] [CrossRef]
- Keijser, B.J.F.; Noens, E.E.E.; Kraal, B.; Koerten, H.K.; van Wezel, G.P. The Streptomyces coelicolor ssgB gene is required for early stages of sporulation. FEMS Microbiol. Lett. 2003, 225, 59–67. [Google Scholar] [CrossRef]
- Sevcikova, B.; Kormanec, J. The ssgB gene, encoding a member of the regulon of stress-response sigma factor σH, is essential for aerial mycelium formation in Streptomyces coelicolor A3(2). Arch. Microbiol. 2003, 180, 380–384. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.; Willemse, J.; Voskamp, P.; Li, X.; Prota, A.E.; Lamers, M.; Pannu, N.; Abrahams, J.P.; van Wezel, G.P. Ectopic positioning of the cell division plane is associated with single amino acid substitutions in the FtsZ-recruiting SsgB in Streptomyces. Open Biol. 2021, 11, 200409. [Google Scholar] [CrossRef] [PubMed]
- Buttner, M.J.; Chater, K.F.; Bibb, M.J. C Cloning, Disruption, and Transcriptional Analysis of Three RNA Polymerase Sigma Factor Genes of Streptomyces coelicolor A3(2). J. Bacteriol. 1990, 172, 3367–3378. [Google Scholar] [CrossRef] [PubMed]
- den Hengst, C.D.; Tran, N.T.; Bibb, M.J.; Chandra, G.; Leskiw, B.K.; Buttner, M.J. Genes essential for morphological development and antibiotic production in Streptomyces coelicolor are targets of BldD during vegetative growth. Mol. Microbiol. 2010, 78, 361–379. [Google Scholar] [CrossRef] [PubMed]
- Craney, A.; Hohenauer, T.; Xu, Y.; Navani, N.K.; Li, Y.; Nodwell, J. A synthetic luxCDABE gene cluster optimized for expression in high-GC bacteria. Nucleic Acids Res. 2007, 35, e46. [Google Scholar] [CrossRef]
- Kelemen, G.H.; Brown, G.L.; Kormanec, J.; Potuckova, L.; Chater, K.F.; Buttner, M.J. The position of the sigma-factor genes, whiG and sigF, in the hierarchy controlling the development of spore chains in the aerial hyphae of Streptomyces coelicolor A3(2). Mol. Microbiol. 1996, 21, 593–603. [Google Scholar] [CrossRef]
- Sun, J.; Kelemen, G.H.; Fernandez-Abalos, J.M.; Bibb, M.J. Green fluorescent protein as a reporter for spatial and temporal gene expression in Streptomyces coelicolor A3(2). Microbiology-SGM 1999, 145, 2221–2227. [Google Scholar] [CrossRef]
- Bush, M.J.; Chandra, G.; Al-Bassam, M.M.; Findlay, K.C.; Buttner, M.J. BldC delays entry into development to produce a sustained period of vegetative growth in Streptomyces venezuelae. mBio 2019, 10, e02812–18. [Google Scholar] [CrossRef]
- Kim, S.H.; Traag, B.A.; Hasan, A.H.; McDowall, K.J.; Kim, B-G. ; van Wezel, G.P. Transcriptional analysis of the cell division-related ssg genes in Streptomyces coelicolor reveals direct control of ssgR by AtrA. Antonie van Leeuwenhoek 2015, 108, 201–213. [Google Scholar] [CrossRef]
- Wang, W.; Li, X.; Wang, J.; Xiang, S.; Feng, X.; Yang, K. An engineered strong promoter for streptomycetes. Appl. Environ. Microbiol. 2013, 79, 4484–4492. [Google Scholar] [CrossRef]
- Kormanec, J.; Sevcikova, B. Identification and transcriptional analysis of a cold shock inducible gene, cspA, in Streptomyces coelicolor A3(2). Mol. Gen. Genet. 2000, 264, 251–256. [Google Scholar] [CrossRef] [PubMed]
- Ausubel, F.M.; Brent, R.; Kingstone, R.E.; Moore, D.O.; Seidman, J.S.; Smith, J.A.; Struh, K. Current Protocols in Molecular Biology; Wiley: New York, 1995. [Google Scholar]
- Kieser, T.; Bibb, M.J.; Buttner, M.J.; Chater, K.F.; Hopwood, D.A. Practical Streptomyces genetics; The John Innes Foundation: Norwich UK,, 2000. [Google Scholar]
- Gust, B.; Challis, G.L.; Fowler, K.; Kieser, T.; Chater, K.F. PCR-targeted Streptomyces gene replacement identifies a protein domain needed for biosynthesis of the sesquiterpene soil odor geosmin. Proc. Natl. Acad. Sci. USA 2003, 100, 1541–1546. [Google Scholar] [CrossRef] [PubMed]
- Redenbach, M.; Kieser, H.M.; Denapaite, D.; Eichner, A.; Cullum, J.; Kinashi, H.; Hopwood, D.A. A set of ordered cosmids and a detailed genetic and physical map for the 8 Mb Streptomyces coelicolor A3 (2) chromosome. Mol. Microbiol. 1996, 21, 77–96. [Google Scholar] [CrossRef] [PubMed]
- Kelemen, G.H.; Brian, P.; Flard, K.; Chamberlin, L.; Chater, K.F.; Buttner, M.J. Developmental regulation of transcription of whiE, a locus specifying the polyketide spore pigment in Streptomyces coelicolor A3(2). J. Bacteriol. 1998, 180, 2515–2521. [Google Scholar] [CrossRef]
- Novakova, R.; Homerova, D.; Csolleiova, D.; Rezuchova, D.; Sevcikova, B.; Javorova, R.; Feckova, L.; Kormanec, J. A stable vector for efficient production of heterologous proteins and secondary metabolites in streptomycetes. Appl. Microbiol. Biotechnol. 2022, 106, 7285–7299. [Google Scholar] [CrossRef]
- Novakova, R.; Sevcikova, B.; Kormanec, J. A method for the identification of promoters recognized by RNA polymerase containing a particular sigma factor: Cloning of a developmentally regulated promoter and corresponding gene directed by the Streptomyces aureofaciens sigma factor RpoZ. Gene 1998, 208, 43–50. [Google Scholar] [CrossRef]
- Sevcikova, B.; Mazurakova, V.; Kormanec, J. Characterization of the alternative sigma factor σG in Streptomyces coelicolor A3(2). Folia Microbiol. 2005, 50, 47–58. [Google Scholar] [CrossRef]
- Kormanec, J.; Sevcikova, B. Stress-response sigma factor σH directs expression of the gltB gene encoding glutamate synthase in Streptomyces coelicolor A3(2). Biochim. Biophys. Acta 2002, 1577, 149–154. [Google Scholar] [CrossRef]
Figure 1.
(a) Nucleotide sequence of the S. coelicolor A3(2) ssgBp promoter region. The deduced protein products are listed in the single-letter amino acid code in the second position of each codon. The ̶ 10 and ̶ 35 boxes and the transcription start site (TSS) of the ssgBp promoter are marked in bold and underlined. Highlighted in green is the predicted binding site of BldD [36]. Details of individual genes and their products are given in Genbank Acc. No. AL939109. (b) A comparison of the ssgBp promoter from S. coelicolor A3(2) with several representative homologous promoters in Streptomyces spp. The ssgBp promoter regions are from Streptomyces genome sequences with the GenBank Acc. No. in parenthesis: S. lividans TK24 (CP009124), S. glaucescens GLA.O (CP009438), S. coelicolor A3(2) (AL939109), S. ambofaciens ATCC 23877 (CP012382), S. chartreusis ATCC 14922 (CP023689), S. nodosus ATCC 14899 (CP009313), S. alboniger ATCC 12461 (CP023695), S. avermetilis NBRC 14893 (BA000030), S. cyanogenus S136 (CP071839), S. clavuligerus ATCC 27064 (CP027858), S. venezuelae ATCC 10712 (FR845719), S. rimosus ATCC 10970 (CP023688), S. noursei ATCC 11455 (CP011533), S. griseus subsp. griseus NBRC 13350 (NC_010572), S. lavendulae subsp. lavendulae CCM 3239 (CP024985). The positions of the ̶ 10, ̶ 35 boxes, TSS and ATG codon are indicated above the sequences. The numbers correspond to the positions of the analyzed genome sequences.
Figure 1.
(a) Nucleotide sequence of the S. coelicolor A3(2) ssgBp promoter region. The deduced protein products are listed in the single-letter amino acid code in the second position of each codon. The ̶ 10 and ̶ 35 boxes and the transcription start site (TSS) of the ssgBp promoter are marked in bold and underlined. Highlighted in green is the predicted binding site of BldD [36]. Details of individual genes and their products are given in Genbank Acc. No. AL939109. (b) A comparison of the ssgBp promoter from S. coelicolor A3(2) with several representative homologous promoters in Streptomyces spp. The ssgBp promoter regions are from Streptomyces genome sequences with the GenBank Acc. No. in parenthesis: S. lividans TK24 (CP009124), S. glaucescens GLA.O (CP009438), S. coelicolor A3(2) (AL939109), S. ambofaciens ATCC 23877 (CP012382), S. chartreusis ATCC 14922 (CP023689), S. nodosus ATCC 14899 (CP009313), S. alboniger ATCC 12461 (CP023695), S. avermetilis NBRC 14893 (BA000030), S. cyanogenus S136 (CP071839), S. clavuligerus ATCC 27064 (CP027858), S. venezuelae ATCC 10712 (FR845719), S. rimosus ATCC 10970 (CP023688), S. noursei ATCC 11455 (CP011533), S. griseus subsp. griseus NBRC 13350 (NC_010572), S. lavendulae subsp. lavendulae CCM 3239 (CP024985). The positions of the ̶ 10, ̶ 35 boxes, TSS and ATG codon are indicated above the sequences. The numbers correspond to the positions of the analyzed genome sequences.
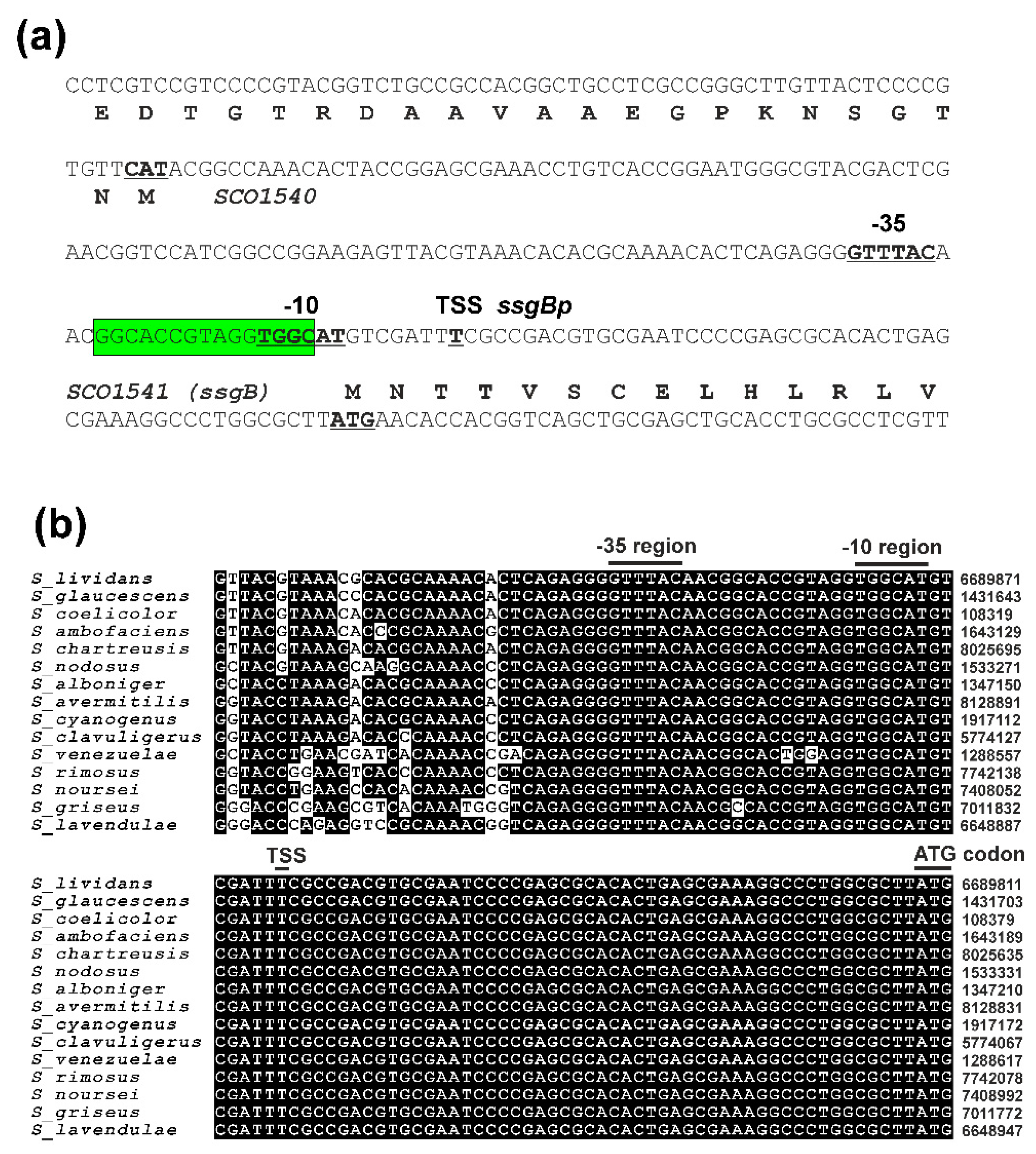
Figure 2.
(a) Schemes of the luxCDABE reporter PhiC31 phage integration plasmids pLux-ssgBp and pLux-kasOp containing a synthetic luxCDABE operon from plasmid pMU1s* [37] and the ssgBp or kasOp* promoter. Arrows indicate the corresponding genes and bent arrows promoters. The plasmid backbone contains the KanR neo gene (blue arrow), the AmpR bla gene (red arrow), the oriT origin of transfer (red bar), the phiC31int integrase gene (green arrow) together with the attP site (green bar), the fdT terminator (dark grey box), and the E. coli origin of replication ColE1 (light grey box). The relevant restriction sites are indicated. (b, c) The luciferase activity of the luxCDABE operon after fusion with the corresponding promoters. The plasmids were conjugated to WT S. coelicolor M145 and nine mutant strains containing deleted individual sigB homologous genes (DsigB,F,G,H,I,K,L,M,N). The luminescence of eight independent KanR clones was measured in a Synergy HT microplate reader in RLU after growth and differentiation on solid SFM medium in 96-well plates at the indicated time points. Each time point represents the mean of eight independent measurements. The error bar indicates the standard deviation from the mean. The arrows above the graph indicate the developmental stages. The presence of vegetative substrate mycelium, aerial mycelium and spores was assessed visually and by microscopic examinations at relevant time points. .
Figure 2.
(a) Schemes of the luxCDABE reporter PhiC31 phage integration plasmids pLux-ssgBp and pLux-kasOp containing a synthetic luxCDABE operon from plasmid pMU1s* [37] and the ssgBp or kasOp* promoter. Arrows indicate the corresponding genes and bent arrows promoters. The plasmid backbone contains the KanR neo gene (blue arrow), the AmpR bla gene (red arrow), the oriT origin of transfer (red bar), the phiC31int integrase gene (green arrow) together with the attP site (green bar), the fdT terminator (dark grey box), and the E. coli origin of replication ColE1 (light grey box). The relevant restriction sites are indicated. (b, c) The luciferase activity of the luxCDABE operon after fusion with the corresponding promoters. The plasmids were conjugated to WT S. coelicolor M145 and nine mutant strains containing deleted individual sigB homologous genes (DsigB,F,G,H,I,K,L,M,N). The luminescence of eight independent KanR clones was measured in a Synergy HT microplate reader in RLU after growth and differentiation on solid SFM medium in 96-well plates at the indicated time points. Each time point represents the mean of eight independent measurements. The error bar indicates the standard deviation from the mean. The arrows above the graph indicate the developmental stages. The presence of vegetative substrate mycelium, aerial mycelium and spores was assessed visually and by microscopic examinations at relevant time points. .
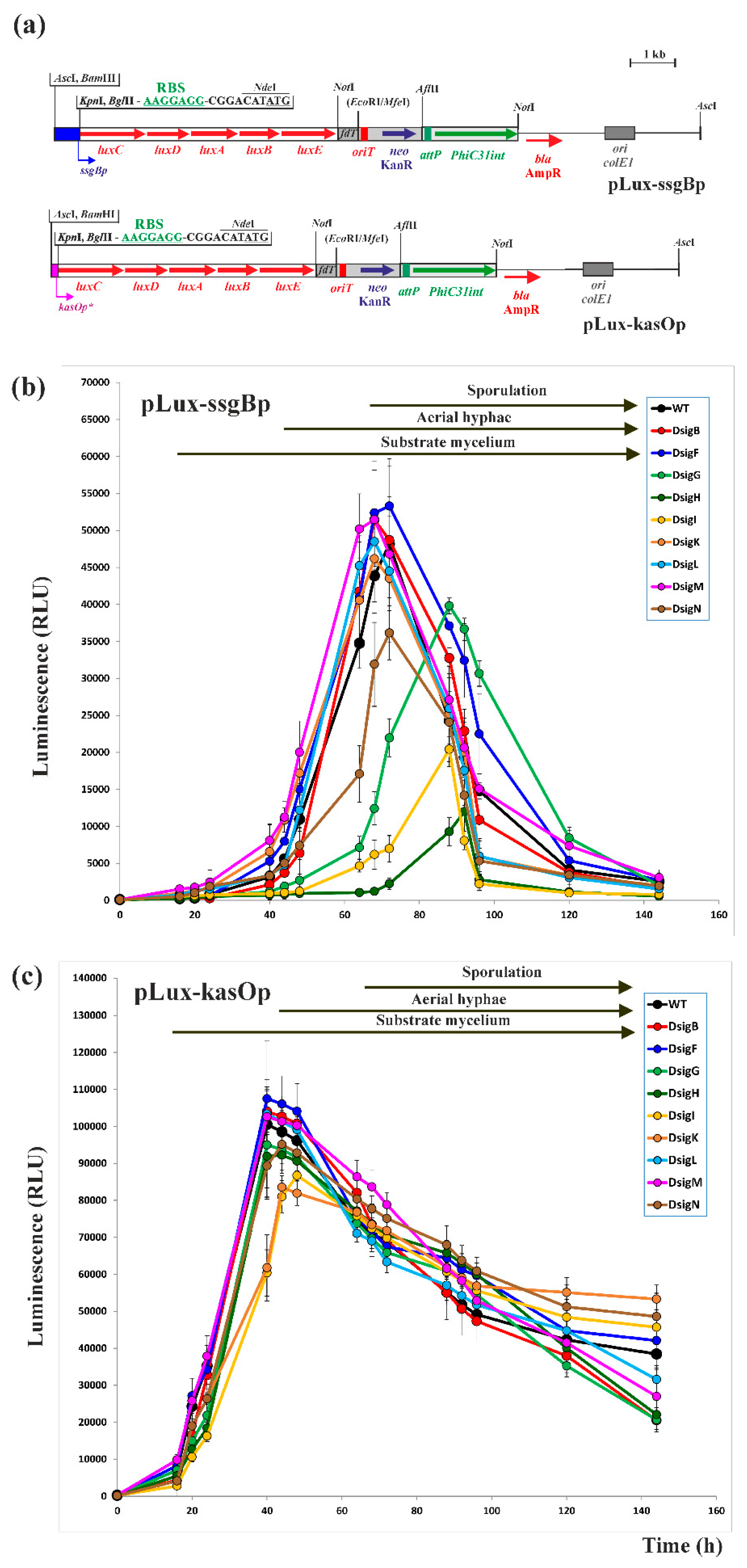
Figure 3.
(a) Nucleotide sequences of the WT ssgBpA promoter and mutant ssgBpB-J promoters with the altered ̶ 10 region. The ̶ 10 and ̶ 35 boxes and TSS of the promoters are indicated in bold and underlined. (b) Cross-recognition of the ssgBpA-J promoters with all nine SigB homologous sigma factors from S. coelicolor A3(2) in the E. coli two-plasmid system with a red box corresponding to a positive result and a white box to a negative result. MacConkey agar plates (with Amp, Clm, IPTG) with transformation mixtures of all plasmids containing the promoters with combinations of expression plasmid pAC5mut2 and the vector with cloned sigma factor genes under the control of the trc promoter (pAC-sigB, pAC-sigF1, pAC-sigF2, pAC-sigG, pAC-sigH1, pACsigH2, pAC-sigI, pAC-sigK, pAC-sigL, pAC-sigM, pAC-sigN) are shown in Supplementary Figure S1.
Figure 3.
(a) Nucleotide sequences of the WT ssgBpA promoter and mutant ssgBpB-J promoters with the altered ̶ 10 region. The ̶ 10 and ̶ 35 boxes and TSS of the promoters are indicated in bold and underlined. (b) Cross-recognition of the ssgBpA-J promoters with all nine SigB homologous sigma factors from S. coelicolor A3(2) in the E. coli two-plasmid system with a red box corresponding to a positive result and a white box to a negative result. MacConkey agar plates (with Amp, Clm, IPTG) with transformation mixtures of all plasmids containing the promoters with combinations of expression plasmid pAC5mut2 and the vector with cloned sigma factor genes under the control of the trc promoter (pAC-sigB, pAC-sigF1, pAC-sigF2, pAC-sigG, pAC-sigH1, pACsigH2, pAC-sigI, pAC-sigK, pAC-sigL, pAC-sigM, pAC-sigN) are shown in Supplementary Figure S1.

Figure 4.
(a) The luciferase activity of the luxCDABE operon after fusion with the ssgBpA-J promoters. Plasmids pLux-ssgBpA, pLux-ssgBpB, pLux-ssgBpC, pLux-ssgBpD, pLux-ssgBpE, pLux-ssgBpF, pLux-ssgBpG, pLux-ssgBpH, pLux-ssgBpI, pLux-ssgBpJ were conjugated to WT S. coelicolor M145 (WT) and nine mutant strains containing deleted individual sigB homologous genes (DsigB,F,G,H,I,K,L,M,N). The luminescence of eight independent KanR clones was measured in a Synergy HT microplate reader in RLU after growth and differentiation on solid SFM medium in 96-well plates at the indicated time points. Each time point represents the mean of eight independent measurements. The error bar indicates the standard deviation from the mean. The arrows above the graph indicate the developmental stages. The presence of vegetative substrate mycelium, aerial mycelium and spores was assessed visually and by microscopic examinations at relevant time points. (b) Nucleotide sequences of the WT ssgBpA promoter and mutant ssgBpB-J promoters with the altered ̶ 10 region. The ̶ 10 and ̶ 35 boxes and TSS of the promoters are indicated in bold and underlined and changed nucleotides in red.
Figure 4.
(a) The luciferase activity of the luxCDABE operon after fusion with the ssgBpA-J promoters. Plasmids pLux-ssgBpA, pLux-ssgBpB, pLux-ssgBpC, pLux-ssgBpD, pLux-ssgBpE, pLux-ssgBpF, pLux-ssgBpG, pLux-ssgBpH, pLux-ssgBpI, pLux-ssgBpJ were conjugated to WT S. coelicolor M145 (WT) and nine mutant strains containing deleted individual sigB homologous genes (DsigB,F,G,H,I,K,L,M,N). The luminescence of eight independent KanR clones was measured in a Synergy HT microplate reader in RLU after growth and differentiation on solid SFM medium in 96-well plates at the indicated time points. Each time point represents the mean of eight independent measurements. The error bar indicates the standard deviation from the mean. The arrows above the graph indicate the developmental stages. The presence of vegetative substrate mycelium, aerial mycelium and spores was assessed visually and by microscopic examinations at relevant time points. (b) Nucleotide sequences of the WT ssgBpA promoter and mutant ssgBpB-J promoters with the altered ̶ 10 region. The ̶ 10 and ̶ 35 boxes and TSS of the promoters are indicated in bold and underlined and changed nucleotides in red.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



