
Submitted:
08 September 2023
Posted:
11 September 2023
You are already at the latest version
Abstract
This study compared growth performance, serum biochemical indicators, rumen fermentation parameters, rumen bacterial structure, and fecal bacterial structure of cattle and yaks, fed for two months, given a feed containing concentrate to roughage ratio of 7: 3 on dry matter basis. Compared with cattle, yak showed better growth performance. The serum biochemical results showed that the albumin: globulin ratio in yaks serum was significantly higher than that in cattle. Aspartate aminotransferase, indirect bilirubin, creatine kinase, lactate dehydrogenase and total cholesterol were significantly lower in yaks than that in cattle. Rumen pH, acetate to propionate ratio, and acetate were lower in yaks than in cattle while the lactate in yaks was higher than in cattle. There were significant differences in the structure of ruminal as well as fecal bacteria between cattle and yaks. The prediction of rumen bacterial function showed that there was metabolic difference between cattle and yaks. In general, the metabolic pathway of cattle was mainly riched in de novo synthesis of nucleotides, while that of yaks was mainly riched in the metabolic utilization of nutrients. This study provides a basis for understanding the rumen ecology under the condition of high concentrate diet.
Keywords:
yaks
; cattle
; blood metabolism
; rumen bacterial
; fecal bacterial
1. Introduction
The grass-withering period is long in the Qinghai-Tibetan plateau due to its harsh environment. Yaks mainly live on grazing at the Qinghai-Tibetan plateau. A large number of studies on the adaptability of cattle and yaks show that yaks are better able to adapt to the pasture conditions of the Qinghai-Tibet Plateau [1,2], and yaks have a higher lignocellulose degradation rate than cattle [3,4]. This is related to the long-term grazing life of yaks, which also leads to a greater abundance of total cellulose-degrading bacteria in the rumen [5].
However, under the condition of long-term low nutrition levels, it is unfavorable for the production performance of yaks, in the cold season, the yaks can lose up to 25% of their body weight [6]. With the increase in the demand for milk and meat products and the gradual transformation of yak breeding mode from the initial grazing life to grazing plus supplementary feeding and captive breeding, it is particularly important to study the physiological response of yaks under high concentrated feeding conditions.
There are a lot of research reports that feeding large amounts of a high-concentrate diet can improve production performance, but cause nutritional metabolic diseases such as rumen acidosis, which change the structure and composition of the rumen microbial community [7], furthur leads to rapid fermentation in the rumen and a lower pH due to elevated levels of volatile fatty acids and lactate [8]. In addition, studies report that a large amount of fermentable carbohydrates flowing into the hindgut will cause hindgut acidosis, and even affect liver function through the increase of lipopolysaccharide, biogenic a,mine and other substances [9,10,11].
However, there is less information on the metabolic capacity, rumen microbiota, and health status of yaks comparing cattle given high-concentrate diets. Therefore, in the current study, we hypothesized that there is a difference in the metabolic ability of cattle and yaks to high-concentrate diets, including the differences in growth performance, blood biochemistry, rumen fermentation, and ruminal and fecal bacterial structure between yaks and cattle. To achieve this goal, rumen fluid, serum and fecal samples from yak and cattle after two months of feeding period to investigate differences in relevant indicators.
2. Materials and Methods
This experiment was conducted at Fengju breeding professional cooperative of Datong County, Qinghai Province, China (altitude approximately 2900 m; 37°40′N latitude and 101°52′E longitude) in 2021 from July to September. The site has a plateau continental climate. The protocol and all experimental procedures on the animals were approved by the Committee of Animal Use, Academy of Science and Veterinary Medicine, Qinghai University (Protocol QMKY202002).
2.1. Feeding and management
Datong yaks and native cattle, ten males each of the same age (approximate age 36-months) were selected for the study. Before the experiment, the cowshed was thoroughly cleaned and disinfected, and all animals were housed individually in single pens (1.5m×3m). The pre-feeding adaptation period was two weeks, and the data collection period was of two months. All the animals were provided a total mixed ration (Table 4) twice daily at 8:00 and 17:00. At the same time, the feeding trough, sink and cowshed were cleaned, and the experimental animals were free access to feed and water round the clock. Feed and water intake was recorded daily. Animals were weighed before and after the experiment to calculate the average daily gain and feed to gain ratio.
2.2. Sample collection
Before morning feeding every week in the formal trial period, the collected dietary samples were mixed evenly and then sampled by quartering method. The samples were baked to constant weight at 65 °C, crushed and stored at - 20 °C for testing. Basal diet samples were analysed for DM, ether extract (EE), and CP according to AOAC [12] using the standard procedures: 934.01, 942.05, and 920.39, respectively. The NDF and ADF contents were assessed according to Van Soest [13]. The samples of serum and rumen fluid of all experimental individuals were collected on day 60 before morning feeding. The blood samples were collected via the jugular vein and then were centrifuged at 3,200 g for 20 min. The serum samples were stored at -80 °C with 1.5 mL sterile microtubes for the later analysis. Ruminal fluid samples were collected using a flexible oral stomach tube with a metal strainer. The oral stomach tube was washed with clean water and the first 200 mL of ruminal fluid was discarded during the collection to avoid contamination between individuals. The collected rumen fluid samples were frozen immediately in liquid nitrogen for later analysis for bacterial community.
2.3. Biochemical indexes determination for blood and fermentation parameters for rumen fluid
Total protein, albumin, globulin, glucose, total cholesterol, triglyceride, aspartate aminotransferase, alanine aminotransferase, alkaline phosphatase, total bilirubin, direct bilirubin, indirect bilirubin, creatine kinase, lactate dehydrogenase, calcium, phosphorus in serum were measured by an automatic biochemical analyzer (BS-280, Mindray Bio-Medical Electronics Co., Ltd., Shenzhen, Guangdong, China). The pH value of rumen fluid was measured immediately after collection with pH meter (Ecoscan pH 5, Singapore). Lactate was determined with the Lactate Assay Kit (Nanjing Jiancheng Bioengineering Institute, Nanjing, China). For the analyses of the volatile fatty acids, 1 ml of rumen supernatant was added into 1.5 ml centrifuge tube, and then added 200 μl deproteinized metaphosphate solution containing crotonic acid. The mixture was filtered by filter (size 0.22 µm, SCAA-102; ANPEL, Shanghai, China) after centrifugation at 12,000 g for 10 min. Then the volatile fatty acids were analyzed using GC (Agilent 7890B, Agilent, California, United States), keeping column temperature of 135 ◦C, an injection temperature of 200 ◦C, FID detector temperature of 200 ◦C, and a carrier gas (N2) pressure of 0.06 MPa [14].
2.4. DNA extraction, 16S rRNA gene amplification, and high-throughput sequencing for rumen
The E.Z.N.A ® kit (Omega Bio-tek, Norcross, GA, USA) was used to extract the total DNA of rumen bacteria. The concentration and the purity of the extracted DNA were determined using a NanoDrop 2000 UV-vis Spectrophotometer (Thermo Scientific, Wilmington, DE, USA). The qualified genomic DNA samples and corresponding fusion primers were used to configure the PCR reaction system for amplification. The primers were 515-F (5”-GTGCCAGCMGCCGCGGTAA-3”) and 806-R (5”-GGACTACCVGGGTATCTAAT-3”). The PCR amplification products were purified using Agincourt AMPure XP magnetic beads and dissolved in the elution buffer, labeled, and the library was completed. The fragment range and concentration of the library were detected using Agilent 2100 Bioanalyzer. The qualified libraries were sequenced on the HiSeq platform according to the size of the inserted fragments. The reads that matched the primers were intercepted with the software cutadapt v2.6 to remove the contamination of primers and linkers, and the fragments of the target region were obtained. The final clean data were obtained by removing the low-complexity reads. The sequence was spliced using the software FLASH (Fast Length Adjustment of Short reads, v1.2.11) to obtain the tags of highly variable region. The sequences with 99% similarity were then assigned to the same operational taxonomic units (OTUs) using the Vsearch plug-in in QIIME2. The QIIME 2 software was used to analyze the α andβ-diversity. The principal coordinate analysis (PCoA) based on the unweighted UniFrac distance was used to compare the bacteria communities between yaks and cattle. Based on the LDA value with a threshold of 3, the linear discriminant analysis (LDA) effect size (LEfSe) analysis between groups from 16S rRNA sequencing was analyzed. PICRUSt software was used for predicting the function of the rumen bacteria with the KEGG database [15], and then used STAMP to analyze the differences between groups. The raw sequence data were subjected to the Sequence Read Archive (SRA) of the NCBI under accession number PRJNA931567.
2.5. Statistical analysis
The analysis of blood biochemical indexes was conducted by independent samples T-test. The P-value < 0.05 was considered significant, and the results were presented in the form of mean ± standard error of the mean (SEM) (SPSS 21 version, IBM Corp., Chicago, IL, USA). The drawing of different indicators was conducted using GraphPad Prism 7.0 (GraphPad software, San Diego, CA). The analysis of rumen and fecal bacteria were plotted using the online website (https://www.omicstudio.cn; https://www.bioincloud.tech)
3. Results
3.1. Growth performance, blood biochemical indices and rumen fermentation parameters
At the same age, yaks had lower dry matter intake, daily water intake, average daily gain, and feed to gain ratio than those of cattle (P < 0.050; Table 1). The albumin: globulin ratio, urea, and uric acid were significantly higher in yaks than in cattle (P < 0.050; Table 2). Aspartate aminotransferase (P = 0.012), indirect bilirubin (P = 0.021), creatine kinase (P = 0.047), lactate dehydrogenase (P < 0.001) and total cholesterol (P = 0.001) were significantly lower in yaks than that in cattle. No differences were found in the serum concentrations of alanine aminotransferase, total protein, albumin, globulin, alkaline phosphatase, total bilirubin, direct bilirubin, glucose, calcium, phosphorus, and triglyceride (P > 0.05) of yaks and cattle (Table 2). The acetate, acetate to propionate ratio and pH of rumen fluid were significantly higher in cattle than in yaks, while the lactate content was significantly higher in yaks than in cattle (P < 0.05). No significant differences were found in other rumen fermentation parameters (P > 0.05; Table 3).
3.2. Ruminal and fecal bacteria structure
Alpha diversity was measured by evenness, observed features, faith-pd, and Shannon’s diversity index. Alpha-diversity metrics showed no significant difference between cattle and yaks in fluid samples (P > 0.05; Figure 1). Based on the unweighted UniFrac distance, the principal coordinate analysis (PCoA) showed that there was an obvious difference in the rumen bacterial community between yaks and cattle (Figure 2A). The Venn diagram shows that in rumen fluid samples, the common OTUs are 1,250 between cattle and yaks, and the unique OTUs are 790 and 1,122, respectively (Figure 2B). Differential bacteria were identified from the kingdom to genus levels using the LEfSe analysis by setting the LDA value to 3. The results showed that the rumen bacterial structure of cattle and yaks was significantly different (Figure 2C). At the phylum level, the main bacteria of cattle and yaks are Firmicutes and Bacteroidetes. The relative abundance of Spirochaeae in rumen fluid of cattle is significantly higher than that in yak (P < 0.05, Figure 3A). At the genus level, the bacteria with differences were Prevotella 1(P = 0.024), Quinella (P = 0.026), Unclassified Bacteroides BS11 gut group (P = 0.049), Villonellaceae UCG-001 (P = 0.008), Bacteroides BS11 gut group (P= 0.001), Treponema 2 (P = 0.038), Ruminococcus 1 (P = 0.008), Uncultured-Bacteroidales BS11 gut group (P = 0.001;Figure 3B, 6B). In addition, the function prediction results of ruminal bacteria show that the pathway of S−adenosyl−L−methionine cycle I, adenosine deoxyribonucleotides denovo biosynthesis II, guanosine deoxyribonucleotides denovo biosynthesis II and NAD biosynthesis I (fromaspartate) were significantly rich in cattle (P < 0.05), while the pathway of 4−aminobutanoate degradation V, sulfate reduction I (assimilatory), super pathway of menaquinol−8 biosynthesis II and sulfate assimilation and cysteine biosynthesis were significantly rich in yak (P < 0.05) (Figure 7). Based on Spearman’s correlation analysis, the different genera of rumen bacteria and the phenotypic indicators correlation significantly (| r | > 0.6, P < 0.05; Figure 8).
After binning reads at 99% similarity, a total of 6,533 OTUs were detected by 16S rRNA gene sequencing, the sequence numbers in fecal samples of cattle and yaks were 56,785±891 and 58,054±433, respectively. Alpha-diversity metrics showed no significant difference between cattle and yaks in fecal samples (P > 0.05; Figure 4). The principal coordinate analysis (PCoA) showed that there was little difference in the feces bacterial community between yaks and cattle (Figure 5A). The Venn diagram shows that in fecal samples, the common OTUs are 1336 between cattle and yaks in fecal samples, and the unique OTUs are 732 and 622, respectively (Figure 5B). The LEfSe analysis by setting the LDA value to 3 showed that the fecal bacterial structure of cattle and yaks was significantly different (Figure 5C). At the phylum level, the main bacteria of cattle and yaks are Firmicutes and Bacteroidetes. The relative abundance of Spirochaeae in fecal of cattle is significantly lower than that in yak (P < 0.05, Figure 6A). At the genus level, Treponema 2 (P = 0.033), Phascolactobacterium (P = 0.007) and Ruminococcaceae (P = 0.037) were significantly different in feces of yaks and cattle.
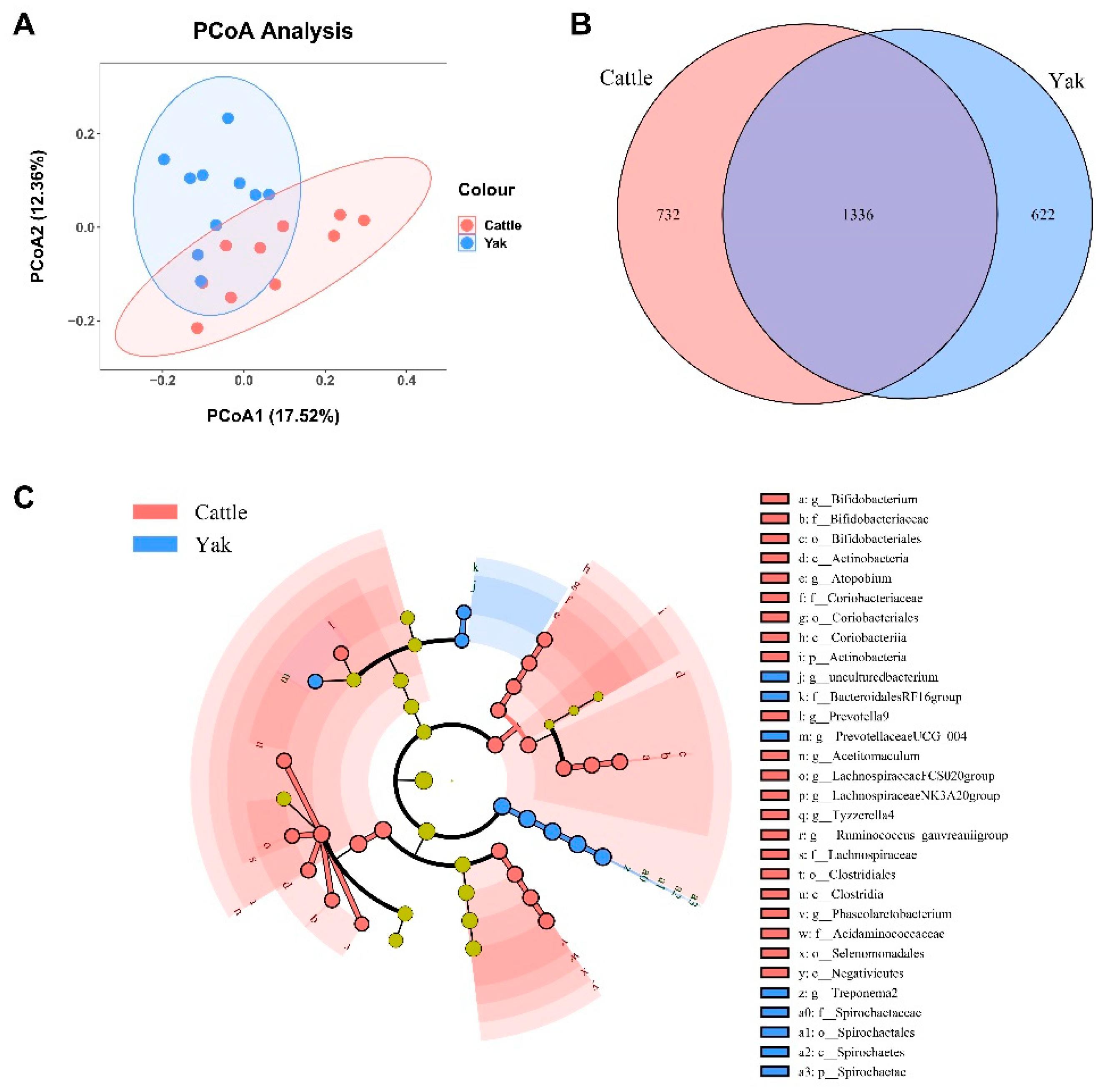
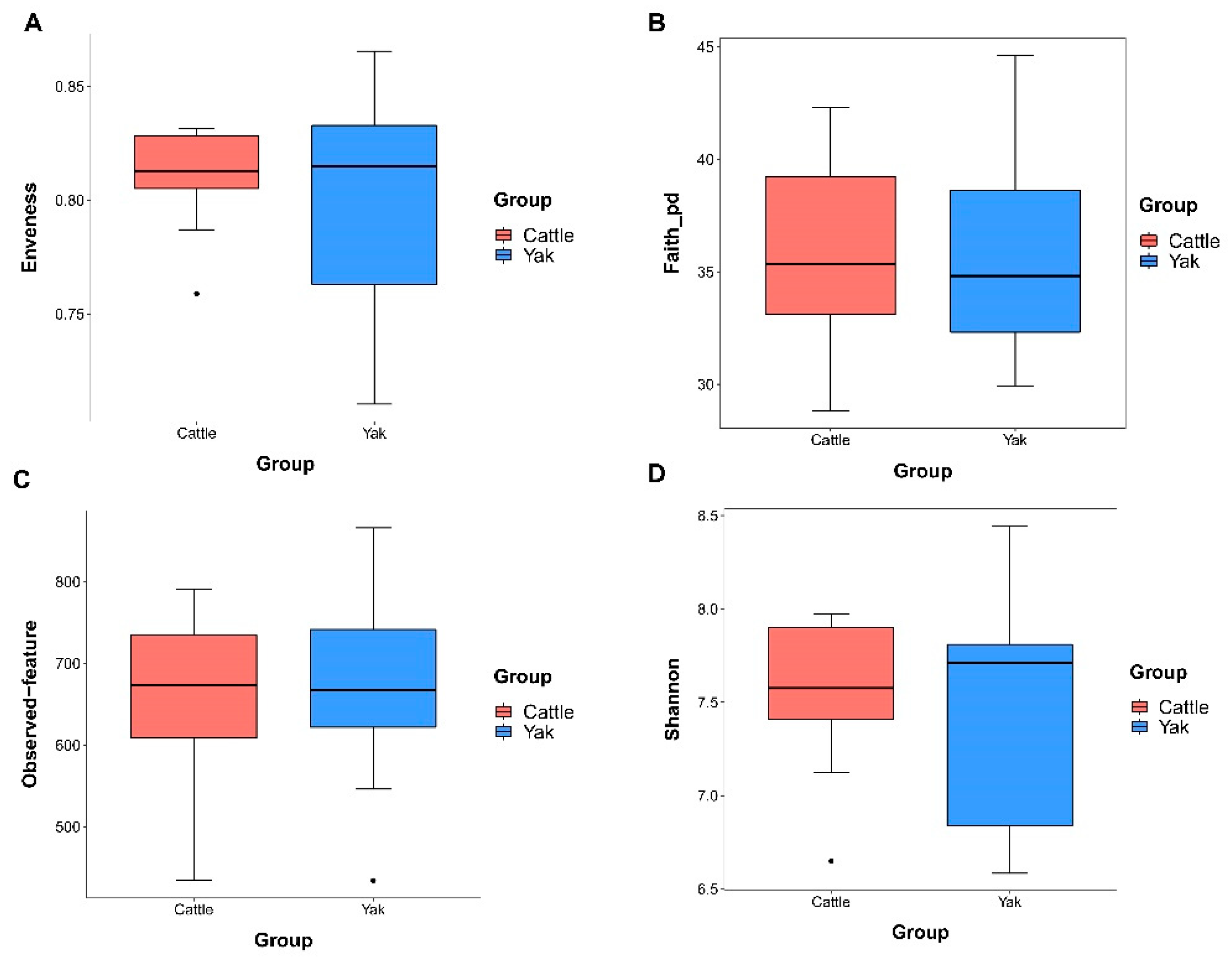
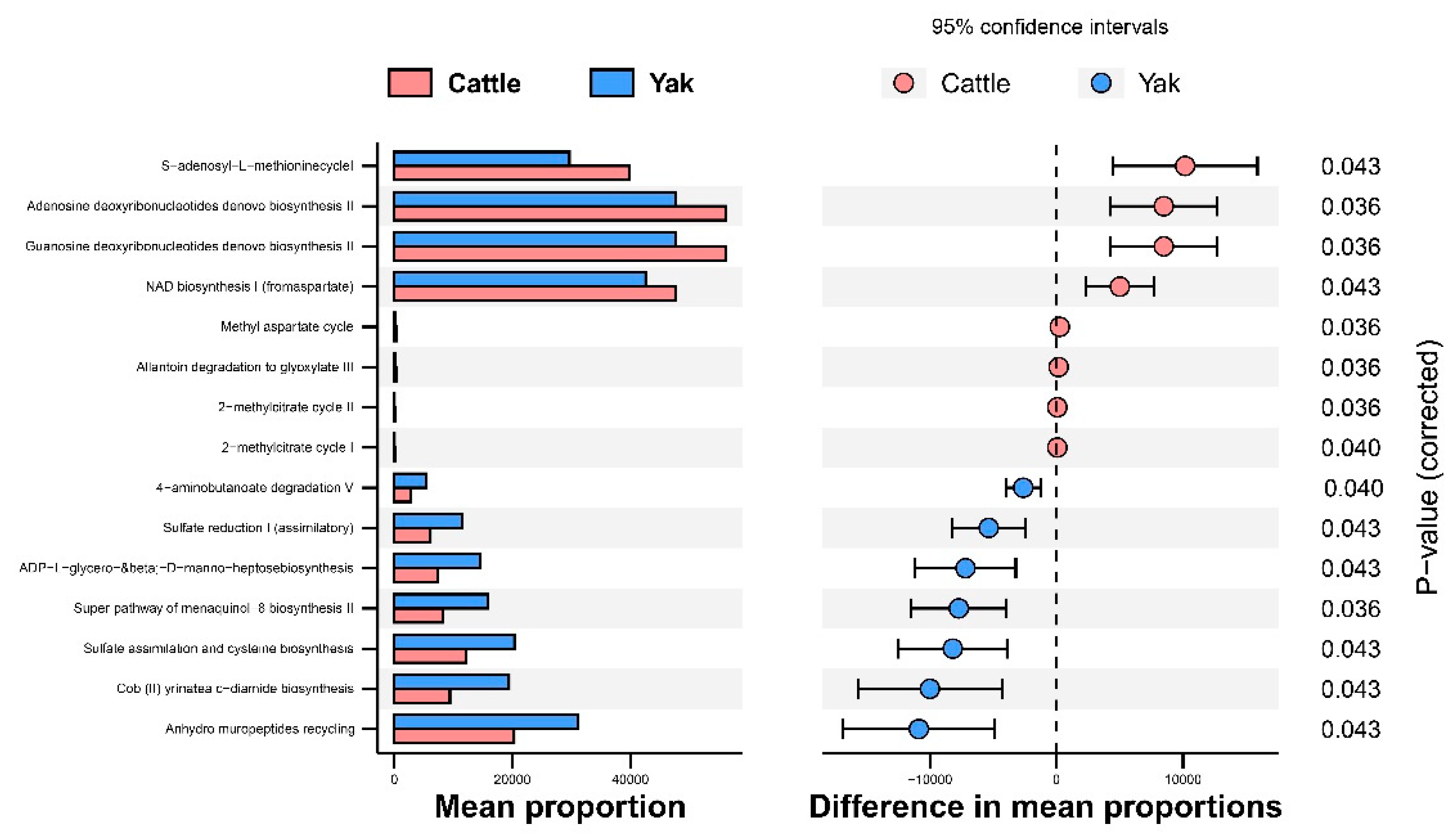
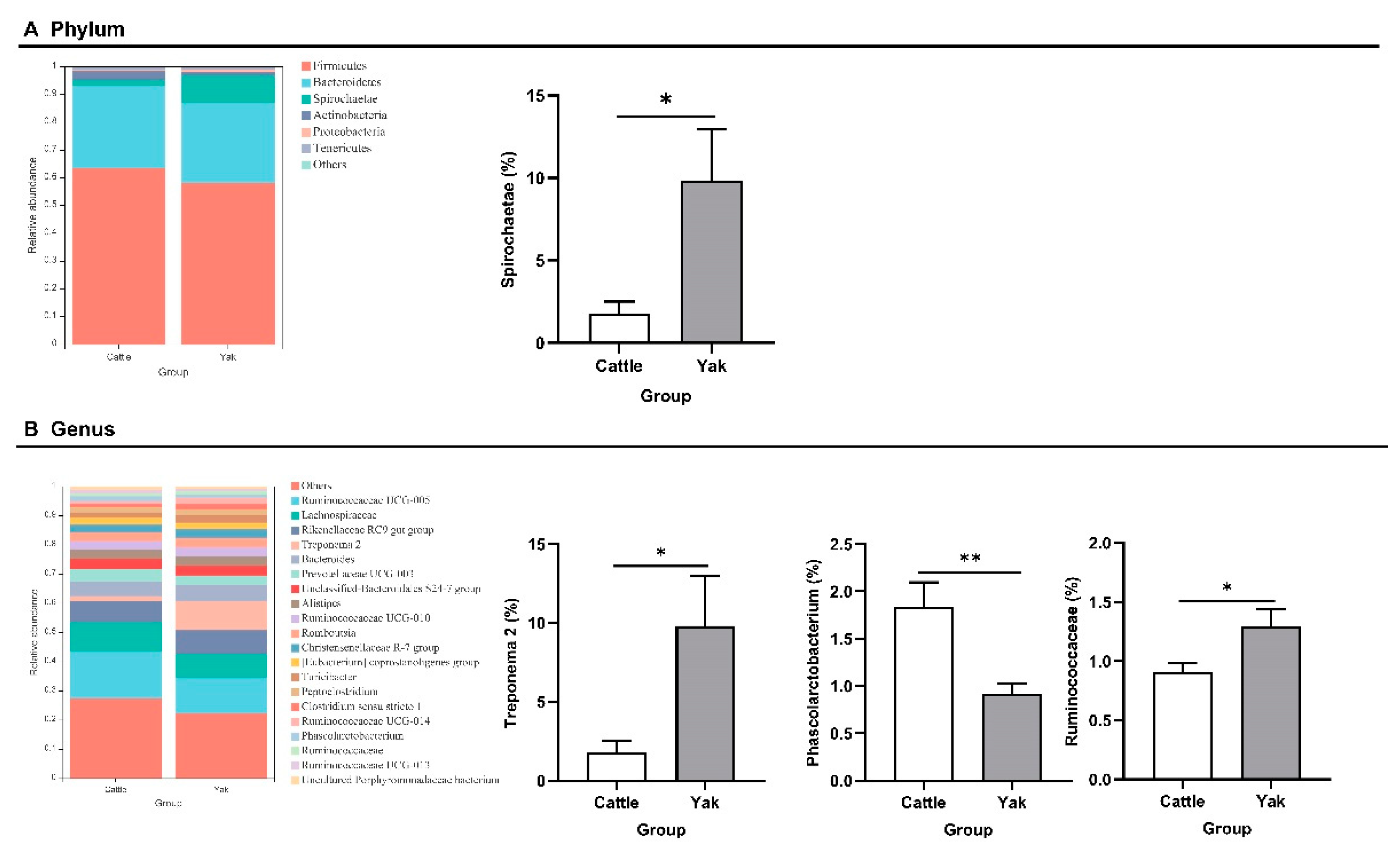
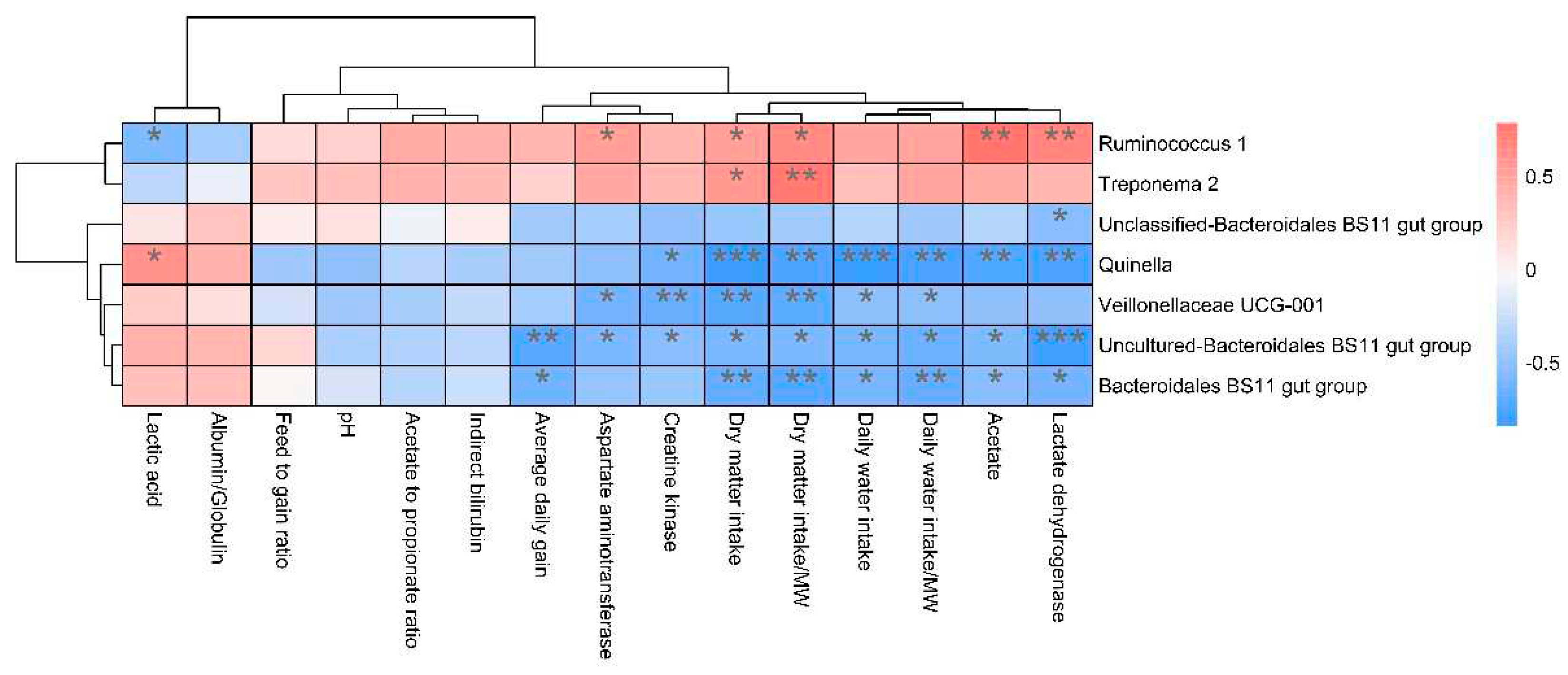
Figure 4.
α diversity and richness in fecal bacterial in yaks and cattle given high concentrate diets.
Figure 4.
α diversity and richness in fecal bacterial in yaks and cattle given high concentrate diets.
Figure 5.
(A) PCoA plots and (B) Venn diagrams of the fecal bacterial community. (C) Cladogram shows fecal bacteria from phylum level to genus level based on the linear discriminant analysis (LDA) effect size (LEfSe) analysis in yaks and cattle given high concentrate diets.
Figure 5.
(A) PCoA plots and (B) Venn diagrams of the fecal bacterial community. (C) Cladogram shows fecal bacteria from phylum level to genus level based on the linear discriminant analysis (LDA) effect size (LEfSe) analysis in yaks and cattle given high concentrate diets.
Figure 6.
(A) Fecal bacterial composition at the phylum level (the relative abundance > 0.5%) and bacteria in cattle and yaks (B) Fecal bacterial composition at the genus level (top 20) and bacteria in cattle and yaks given high concentrate diets. A single asterisk (*) indicates a signifcant diference at P<0.05, while double asterisks (**) indicate a signifcant diference at P<0.01.
Figure 6.
(A) Fecal bacterial composition at the phylum level (the relative abundance > 0.5%) and bacteria in cattle and yaks (B) Fecal bacterial composition at the genus level (top 20) and bacteria in cattle and yaks given high concentrate diets. A single asterisk (*) indicates a signifcant diference at P<0.05, while double asterisks (**) indicate a signifcant diference at P<0.01.
Figure 7.
Stamp analysis in KEGG metabolic pathway of rumen bacteria in yaks and cattle given high concentrate diets.
Figure 7.
Stamp analysis in KEGG metabolic pathway of rumen bacteria in yaks and cattle given high concentrate diets.
Figure 8.
Correlation analysis between rumen differential bacteria and significantly different growth performance, serum biochemical indexes and rumen fermentation parameters in cattle and yaks given high concentrate diets. A single asterisk (*) indicates a signifcant diference at P<0.05, double asterisks (**) indicate a signifcant diference at P<0.01, and triple asterisks (***) indicate a signifcant diference at P<0.001.
Figure 8.
Correlation analysis between rumen differential bacteria and significantly different growth performance, serum biochemical indexes and rumen fermentation parameters in cattle and yaks given high concentrate diets. A single asterisk (*) indicates a signifcant diference at P<0.05, double asterisks (**) indicate a signifcant diference at P<0.01, and triple asterisks (***) indicate a signifcant diference at P<0.001.
4. Discussion
4.1. Production efficiency, serum biochemistry and rumen fermentation parameters
Under the condition of high-concentrate diet, cattle showed a higher average daily gain, which may be due to the fact that yaks mainly graze, while cattle mainly feed in barns providing high-concentrate diet. Accordingly, also buffalo show lower daiy weight gain than bovines [16]. However, compared with cattle, yaks had significantly lower feed to gain ratio, indicating that, given high concentrate diet, yaks have better feed utilization capacity than cattle. These results are consistent with those studies [17,18,19] in which yak has been reported to energy-saving utilization mode. High concentrate diet can cause changes in nutrient metabolism of the body, and hence affect the immune response and health of the body [20], which can be reflected through blood biochemical indicators. Alkaline phosphatase is an important physiological index that can reflect liver function and protein metabolism [21], it increases in the serum with diseases. In this study, the alkaline phosphatase content of cattle and yaks is between 17.5 to 387 U/L, which is within the normal range [22]. However, the content of alkaline phosphatase in cattle is significantly higher than that of yaks. Whether it has pathological significance should be specifically discussed in combination with gene expression and tissue slices. The albumin/globulin ratio represents the synthesis of blood protein, which is widely used in clinical biochemistry to identify abnormal protein synthesis. When it is below the normal range, it may represent inflammation, immune deficiency, or liver and kidney diseases [23]. In a study, albumin/globulin range is 0.45-1.31 in dairy cows [24]. In this study, the value in the serum of cattle and yak is within the normal range, indicating that there is no obvious metabolic disease in both of them under the condition of high concentrate diet. Urea is the final product of protein, which was negatively correlated with nitrogen use efficiency in ruminants [25]. In this study, the urea content in the serum of cattle was significantly lower than that of yaks, indicating that the higher nitrogen position and protein synthesis of cattle under the condition of high concentrate diet led to the higher average daily gain of cattle. There are also research reports that the serum urea level in yak was also higher than that found in buffalo [26].The reference range of creatine kinase in buffalo serum is 14.66- 309.80 (U/L) [27]. Creatine kinase of yak is within the normal range, but the content in cattle is significantly higher, and also higher than the normal range. The reason may be that the differences between breeds cause the differences in the normal range. However, it is undeniable that the abnormal rise of creatine kinase in serum may be due to the cell damage caused by metabolic disorders, which makes its contents released into the serum [28]. As a marker of cell damage and inflammatory mediators, lactate dehydrogenase is often used as an indicator for clinical diagnosis [29]. Reference range of lactate dehydrogenase in buffalo serum is 186.72- 917.43 (U/L) [27]. In the present study, the level of lactate dehydrogenase in cattle and yaks serum is beyond the normal range and it is significantly higher in cattle than in yak. This may be due to the inflammatory reaction of cells, which leads to the abnormal increase of lactate dehydrogenase content. Total cholesterol level in buffalo was 0.90- 1.99 mmol/L [27]. In addition, the level of total cholesterol were significantly lower in yaks than that in cattle, which also found the same level in buffalo [26]. Cholesterol is actively metabolized to produce bile acid [30], and bile acid has a strong function of decomposing adipose tissue and protein, enabling the small intestine to quickly decompose these nutrients and absorb them. Thus, it plays a very important role in the metabolism and absorption of body nutrients [31]. Bile acid can also participate in a variety of signal pathways in the body, including fibrosis and immune homeostasis, to manage the immune and inflammatory responses of the body [32,33]. In this study, the total cholesterol level of cattle is significantly higher than that of yaks and exceeds the normal range, which may have been caused by its cholesterol metabolism disorder.
The pH and the concentrations of VFAs are the main indicators of the status of rumen fermentation [34]. In the present study, the ratio of acetate to propionate ranged between 3.10 and 2.67. A ratio greater than 2.2 was reported to have a positive effect on the rumen and productive performance of cattle [35], which would indicate that all the yaks and cattle had a well-functioning rumen. The acetate was significantly higher in cattle than in yaks in the study, which is linked closely to fiber degradation [36]. This shows that cattle are more effective in utilizing fiber in high-concentrate diets compared to yaks. In addition, the lactate content was significantly higher in yaks than in cattle, this may also indicate that yaks can more effectively utilize starch in high-concentrate diets, which further reduces the pH value in the rumen of yaks. It may be an important reason why the rumen pH of yaks is lower than that of cattle. To sum up, under the condition of high-concentrate diet, cattle have higher average daily gain than yaks, but yaks have a healthier metabolism and lower feed to gain ratio. These characteristics may be related to the long-term survival conditions and diet level of cattle and yaks.
4.2. Rumen and fecal bacterial structure in cattle and yaks
Rumen microbial structure was affected by the proportion of concentrate in diet [37]. Microbial structure influences the health status of animals. Higher the microbial diversity, better is the health of the animals [38]. The α-diversity index of yaks tends to be higher than that of cattle, which may be the result of feeding forage with high fiber content around the year, and it contains many rare and precious types of grass, which may help increase the diversity of rumen bacteria in yak and improve immunity. At the genus level, Prevotella_1 was the predominant genus, which has a wide metabolic niche due to genetic relatedness or high genetic variability that enables it to occupy different ecological niches within the rumen [39]. This genus utilizes starch and proteins to produce succinate and acetate [40]. Therefore, the difference between acetate and lactate may be partly attributed to Prevotella_1. In the present study, the relative abundance of Prevotella in the rumen and level of aspartate aminotransferase in serum of cattle were higher than in yaks. In a previous study [41], Prevotella has been shown to have a role in mediating inflammatory reaction, which results in degeneration of liver adipose tissue and increased inflammation, consequently increasing the serum aspartate aminotransferase. This may be the important reason why the content of aspartate aminotransferase in cattle serum is significantly higher than that in yaks. However, the specific mechanism of Prevotella mediated inflammatory response needs further study. It has been reported that Quinella is significantly higher in grazing yaks than in cattle [42], which are reportedly that lactate and propionate formation are major fermentation pathways of Quinella. Analysis of Quinella genome shows that there is a potential gene for hydrogen uptake, but no metabolic pathway for hydrogen production [43]. In addition, in the correlation analysis results of this study, Quinella and lactate content are significantly positively correlated, which may be an important reason for the high lactate content in the rumen of yaks. Bacteroidales BS11 has been observed to be higher relative abundance in moose given diet mainly composed of hemicellulose and lignin [44]. Genome-enabled metabolic analyses results showed that these bacteria can ferment hemicellulose monomeric sugars to short-chain fatty acids. The relative abundance of this genus in the rumen of the yak was significantly higher than that of cattle. Veillonellaceae belongs to Firmicutes, which can degrade and utilize cellulose to provide the hydrogen and energy. High relative abundance of Bacteroidales BS11 and Veillonellaceae in the rumen of yak may be the result of adaptation to forage with high hemicellulose and lignin content for a long time, further causing differences among breeds. Even under the condition of high concentrate diet, yaks still had higher number of Bacteroidales BS11 and Veillonellaceae than in cattle. It has been reported that Treponema is a common potential pathogen, which plays a very important role in mediating inflammatory reaction [45,46]. However, there are few reports on its pathogenicity, which may be because its metabolites can promote the expression of inflammatory factors and lead to the generation of body diseases. Therefore, higher number of Treponema in the rumen of cattle may aggravate the body’s inflammatory response, indicating how it interacts with the host during the feeding of high concentrate diets and play an important role in mitigating the damage caused by high concentrate diets.
In addition, the prediction results of rumen bacterial function showed that there was metabolic difference potential between cattle and yaks. In general, the metabolic pathway of cattle was mainly oriented towards the de novo synthesis of nucleotides, while that of yaks was mainly oriented towards the metabolic utilization of nutrients. The 4-aminobutanoate is a major endogenous neurotransmitter in the mammalian brain, which can generate succinic semialdehyde in the rumen under the action of aminobutyric acid transaminase and convert it into succinate, entering the tricarboxylic acid cycle. The prediction result of rumen bacterial function also shows that yaks have more significant enrichment of 4-aminobutyrate degradation pathway than cattle, which means that yaks have more energy substances to produce [47]. Sulfate in the diet is biochemically reduced to sulfide by sulfate reducing bacteria in the rumen, which can compete with the methane generation pathway to reduce methane production [48]. It is reported that compared with the high-fiber diet, the high-starch diet significantly enhances the rumen fermentation ability, reduces methane production by inhibiting the proliferation of methane bacteria, and promote the proliferation of hydrogen trophic bacteria with respiratory function. In this study, the prediction results of rumen bacterial function show that yaks had stronger sulfate reduction capacity than cattle, so it can be assumed that under high concentrate diet, yaks have stronger ability to inhibit methane generation than cattle [49]. This is consistent with the comparative differences of rumen bacterial such as Quinella in yaks and cattle. Cysteine is the end product of the sulfate assimilation pathway which widely exists in organisms [50]. It is the starting material for the synthesis of methionine, and also participates in the formation of biotin, thiamine, etc. Menaquinol- 8 is considered as a vitamin, which plays a very important role in promoting the absorption of proteins and lipids [51]. Therefore, it is speculated that the higher levels of menaquinol-8 biosynthesis and sulfate assimilation and cysteine biosynthesis may have resulted in more nutrients metabolization and absorption under the condition of high concentrate diet in yaks than in cattle.
The results of fecal bacterial diversity analysis showed that Ruminococcaceae play important role in the degradation of cellulose and fermentation of plant fibers [52] and are associated with the feed efficiency of dairy cattle [53]. Treponema 2 is a fibrolytic bacteria belonging to Spirochaetae, which can digest hemicellulose. These bacteria have been shown to increase in relative abundance in the rumen of sheep fed high-fiber diet [54]. The results of this study show that the number of Spirochaetae and Treponema 2 were significantly higher in yaks than in cattle. Therefore, it is speculated that the fermentation ability of yaks’ hindgut may be better than that of cattle. However, the specific comparison of the fermentation capacity of the hindgut of yak and cattle should be discussed in combination with the specific research on the disappearance rate of nutrients in the hindgut and fermentation parameters. In the present study, whether α-diversity or β- diversity, the differences between the rumen bacterial structure of cattle and yaks are more obvious than that of their feces. The possible reason is that the rumen is the main place for ruminants to digest nutrients. Most nutrients in the diet are digested in the rumen, and only a small amount of undigested nutrients enter the hindgut. The diet is the most important factor affecting intestinal microorganisms, therefore, the difference of rumen microorganisms between cattle and yak is greater than fecal microorganisms. However, at the phylum level, Spirochaetaes between cattle and yaks shows differences not only in rumen, but also in feces, which play an important role in degrading plant polymers materials such as xylan, pectin, and arabinogalactan [55]. But interestingly, the relative abundance of Spirochaetae in cattle is higher than that of yaks in the rumen, but it is the opposite in the feces, which may be due to the difference of available substrates. Most of the plant polymers materials in cattle are fermented in the rumen, while the unused polymers materials in yaks flow into the hindgut for fermentation. Therefore, it is speculated that the digestive capacity of the hindgut of yaks may be stronger than that of cattle, however, further discussion is required in combination with specific research. It has been reported that the characteristics of the rumen and fecal microbiome of cattle on alfalfa diet, differ slightly between the fecal microbiota of bloated versus non-bloated steers compared with rumen microbiota [56]. It is possible that in the same host, the physiological functions of microorganisms colonized in different parts are different, which is consistent with the research of de Oliveira [57].
5. Conclusions
In general, yaks and cattle have been mainly grazed and captive breeding for a long time, respectively. When feeding the high-concentrate diet, they showed a difference in growth performance, blood biochemical indices, rumen fermentation parameters, and rumen microbiota. In addition, the fecal bacterial structures of cattle and yaks are also different, suggesting that yaks may have stronger digestive capacity in the hindgut. In the future, the functions of microbiota and their response to high concentrated diet in the gastrointestinal tract of yaks, especially in the hindgut, are needed in-depth study using metagenomics and transcriptomics. In conclusion, under the conditions of high concentrated diet, the study is of fundamental significance to understanding the growth performance, blood biochemistry, rumen fermentation, ruminal and fecal bacterial structure between yaks and cattle, which provides a theoretical basis for the fattening method and precision feeding of yaks, so as to maximize the feed efficiency on the basis of ensuring the health of yaks, then it can alleviate the pressure of grassland ecosystem and reduce environmental pollution.
Author Contributions
YZ, YJ, NJ, BB, SW and MS completed the experiment. LX and WD completed the data analysis. CY, MY and HL conceived and designed the paper. LX, CY and HL wrote and revised the article.
Funding
This work was supported by The National Key Research and Development Program of China (2022YFD1302103、2022YFD1602311、2022YFD1602301), Top Talent project of “Kunlun Talents–High-level Innovation and Entrepreneurship Talents” in Qinghai Province (2020), Key Laboratory of Plateau Grazing Animal Nutrition and Feed Science of Qinghai Province (2022-ZJ-Y17).
Institutional Review Board Statement
The study was approved and supervised by Committee of Animal Use, Academy of Science and Veterinary Medicine, Qinghai University.
Informed Consent Statement
Not applicable.
Data Availability Statement
The raw sequence data were subjected to the Sequence Read Archive (SRA) of the NCBI under accession number PRJNA931567.
Acknowledgments
We thank Fengju breeding professional cooperative of Datong County, Qinghai Province, China for providing the site for animal experiments and all the people who participated in the sampling.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Qiu, Q.; Zhang, G.; Ma, T.; Qian, W.; Wang, J.; Ye, Z.; Cao, C.; Hu, Q.; Kim, J.; Larkin, D.M.; et al. The yak genome and adaptation to life at high altitude. Nat Genet 2012, 44, 946–949. [Google Scholar] [CrossRef] [PubMed]
- Mipam, T.D.; Wen, Y.; Fu, C.; Li, S.; Zhao, H.; Ai, Y.; Li, L.; Zhang, L.; Zou, D. Maternal phylogeny of a newly-found yak population in china. Int J Mol Sci 2012, 13, 11455–11470. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Zhu, Y.; Luo, Y.; Song, L.; Liu, D.; Liu, L.; Chen, F.; Wang, M.; Li, J.; Zeng, X.; et al. Metagenomic insights into the fibrolytic microbiome in yak rumen. PLoS One 2012, 7, e40430. [Google Scholar] [CrossRef]
- Wei, Y.Q.; Long, R.J.; Yang, H.; Yang, H.J.; Shen, X.H.; Shi, R.F.; Wang, Z.Y.; Du, J.G.; Qi, X.J.; Ye, Q.H. Fiber degradation potential of natural co-cultures of Neocallimastix frontalis and Methanobrevibacter ruminantium isolated from yaks (Bos grunniens) grazing on the Qinghai Tibetan Plateau. Anaerobe 2016, 39, 158–164. [Google Scholar] [CrossRef] [PubMed]
- An, D.; Dong, X.; Dong, Z. Prokaryote diversity in the rumen of yak (Bos grunniens) and Jinnan cattle (Bos taurus) estimated by 16S rDNA homology analyses. Anaerobe 2005, 11, 207–215. [Google Scholar] [CrossRef]
- Long, R.J.; Dong, S.K.; Wei, X.H.; Pu, X.P.J.L.P.S. The effect of supplementary feeds on the bodyweight of yaks in cold season. 2005, 93, 197–204. [Google Scholar] [CrossRef]
- Khafipour, E.; Li, S.; Plaizier, J.C.; Krause, D.O. Rumen microbiome composition determined using two nutritional models of subacute ruminal acidosis. Appl Environ Microbiol 2009, 75, 7115–7124. [Google Scholar] [CrossRef]
- Motoi, Y.; Oohashi, T.; Hirose, H.; Hiramatsu, M.; Miyazaki, S.; Nagasawa, S.; Takahashi, J. Turbidimetric-kinetic assay of endotoxin in rumen fluid or serum of cattle fed rations containing various levels of rolled barley. J Vet Med Sci 1993, 55, 19–25. [Google Scholar] [CrossRef]
- Ohtaki, T.; Ogata, K.; Kajikawa, H.; Sumiyoshi, T.; Asano, S.; Tsumagari, S.; Horikita, T. Effect of high-concentrate corn grain diet-induced elevated ruminal lipopolysaccharide levels on dairy cow liver function. J Vet Med Sci 2020, 82, 971–977. [Google Scholar] [CrossRef]
- Khafipour, E.; Krause, D.O.; Plaizier, J.C. A grain-based subacute ruminal acidosis challenge causes translocation of lipopolysaccharide and triggers inflammation. J Dairy Sci 2009, 92, 1060–1070. [Google Scholar] [CrossRef]
- Wang, D.S.; Zhang, R.Y.; Zhu, W.Y.; Mao, S.Y.J.L.S. Effects of subacute ruminal acidosis challenges on fermentation and biogenic amines in the rumen of dairy cows. 2013, 155, 262–272. [Google Scholar] [CrossRef]
- Helrich, K.; Scott, P.M. Official Methods of Analysis of the Association of Official Analytical Chemist. 2005. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J Dairy Sci 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Jin, W.; Cheng, Y.; Zhu, W. The community structure of Methanomassiliicoccales in the rumen of Chinese goats and its response to a high-grain diet. J Anim Sci Biotechnol 2017, 8, 47. [Google Scholar] [CrossRef] [PubMed]
- Langille, M.G.; Zaneveld, J.; Caporaso, J.G.; McDonald, D.; Knights, D.; Reyes, J.A.; Clemente, J.C.; Burkepile, D.E.; Vega Thurber, R.L.; Knight, R.; et al. Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nat Biotechnol 2013, 31, 814–821. [Google Scholar] [CrossRef]
- Infascelli, F.; Gigli, S.; Campanile, G. Buffalo meat production: Performance infra vitam and quality of meat. Vet.Res. Commun. 2004, 28, 143–148. [Google Scholar] [CrossRef]
- Mi, J.; Zhou, J.; Huang, X.; Long, R. Lower Methane Emissions from Yak Compared with Cattle in Rusitec Fermenters. PLoS One 2017, 12, e0170044. [Google Scholar] [CrossRef]
- Zhang, Z.; Xu, D.; Wang, L.; Hao, J.; Wang, J.; Zhou, X.; Wang, W.; Qiu, Q.; Huang, X.; Zhou, J.; et al. Convergent Evolution of Rumen Microbiomes in High-Altitude Mammals. Curr Biol 2016, 26, 1873–1879. [Google Scholar] [CrossRef]
- A, Y.Z.; A, J.W.Z.; C, X.S.G.B.; A, G.X.C.; C, L.M.D.B.; A, H.C.W.; D, L.W.L.; B, R.J.L.A.J.L.S. Influences of dietary nitrogen and non-fiber carbohydrate levels on apparent digestibility, rumen fermentation and nitrogen utilization in growing yaks fed low quality forage based-diet. 2012, 147, 139–147. [Google Scholar]
- Ma, N.; Abaker, J.A.; Wei, G.; Chen, H.; Shen, X.; Chang, G. A high-concentrate diet induces an inflammatory response and oxidative stress and depresses milk fat synthesis in the mammary gland of dairy cows. J Dairy Sci 2022, 105, 5493–5505. [Google Scholar] [CrossRef]
- Cozzi, G.; Ravarotto, L.; Gottardo, F.; Stefani, A.L.; Contiero, B.; Moro, L.; Brscic, M.; Dalvit, P. Short communication: reference values for blood parameters in Holstein dairy cows: effects of parity, stage of lactation, and season of production. J Dairy Sci 2011, 94, 3895–3901. [Google Scholar] [CrossRef]
- Hatate, K.; Shinya, K.; Matsuo-Sato, A.; Sasaki, S.; Devkota, B.; Takahashi, M.; Hirata, T.; Yamagishi, N. Changes in the plasma levels of several bone markers in newborn calves during the first two days of life. J Vet Med Sci 2016, 78, 337–340. [Google Scholar] [CrossRef] [PubMed]
- Eckersall, P.D.J.C.B.o.D.A. Proteins, Proteomics, and the Dysproteinemias. 2008, 117–155. [Google Scholar]
- Alberghina, D.; Giannetto, C.; Vazzana, I.; Ferrantelli, V.; Piccione, G. Reference intervals for total protein concentration, serum protein fractions, and albumin/globulin ratios in clinically healthy dairy cows. J Vet Diagn Invest 2011, 23, 111–114. [Google Scholar] [CrossRef] [PubMed]
- Lewis, D.J.T.J.o.A.S. Blood-Urea concentration in Relation to Protein Utilization in. 1957, 48, 438–446. [Google Scholar] [CrossRef]
- Lombardi, P.; Musco, N.; Cutrignelli, M.I.; Mollica, M.P.; Infascelli, F. The association of aloe and β-carotene supplementation improves oxidative stress and inflammatory state in pregnant buffalo cows. Buffalo Bulletin. 2017, 36(3), 497–503. [Google Scholar]
- Abd Ellah, M.R.; Hamed, M.I.; Ibrahim, D.R.; Rateb, H.Z. Serum biochemical and haematological reference intervals for water buffalo Bubalus bubalis heifers. J S Afr Vet Assoc 2014, 85, e1–e7. [Google Scholar] [CrossRef]
- Andretta, I.; Kipper, M.; Lehnen, C.R.; Lovatto, P.A. Meta-analysis of the relationship of mycotoxins with biochemical and hematological parameters in broilers. Poult Sci 2012, 91, 376–382. [Google Scholar] [CrossRef]
- Tzouvelekis, A.; Kouliatsis, G.; Anevlavis, S.; Bouros, D. Serum biomarkers in interstitial lung diseases. Respir Res 2005, 6, 78. [Google Scholar] [CrossRef]
- Bilotta, M.T.; Petillo, S.; Santoni, A.; Cippitelli, M. Liver X Receptors: Regulators of Cholesterol Metabolism, Inflammation, Autoimmunity, and Cancer. Front Immunol 2020, 11, 584303. [Google Scholar] [CrossRef]
- Li, T.; Chiang, J.Y. Bile acid signaling in metabolic disease and drug therapy. Pharmacol Rev 2014, 66, 948–983. [Google Scholar] [CrossRef] [PubMed]
- Joyce, S.A.; Gahan, C.G. Bile Acid Modifications at the Microbe-Host Interface: Potential for Nutraceutical and Pharmaceutical Interventions in Host Health. Annu Rev Food Sci Technol 2016, 7, 313–333. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.L.; Takeda, K.; Sundrud, M.S. Emerging roles of bile acids in mucosal immunity and inflammation. Mucosal Immunol 2019, 12, 851–861. [Google Scholar] [CrossRef] [PubMed]
- Liang, P.; Liu, S.; Xu, F.; Jiang, S.; Yan, J.; He, Q. Corrigendum: powdery mildews are characterized by contracted carbohydrate metabolism and diverse effectors to adapt to obligate biotrophic lifestyle. Front Microbiol 2019, 10, 1. [Google Scholar] [CrossRef] [PubMed]
- Russell, J.B. The importance of pH in the regulation of ruminal acetate to propionate ratio and methane production in vitro. J Dairy Sci 1998, 81(12), 3222–3230. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Cao, P.; Wang, L.; Zhao, Z.; Chen, Y.; Yang, Y. Bacterial community diversity associated with different levels of dietary nutrition in the rumen of sheep. Appl Microbiol Biotechnol 2017, 101(9), 3717–3728. [Google Scholar] [CrossRef]
- Brown, M.S.; Ponce, C.H.; Pulikanti, R. Adaptation of beef cattle to high-concentrate diets: performance and ruminal metabolism. J Anim Sci 2006, 84 Suppl, E25–33. [Google Scholar] [CrossRef]
- Marchesi, J.R.; Adams, D.H.; Fava, F.; Hermes, G.D.; Hirschfield, G.M.; Hold, G.; Quraishi, M.N.; Kinross, J.; Smidt, H.; Tuohy, K.M.; et al. The gut microbiota and host health: a new clinical frontier. Gut 2016, 65, 330–339. [Google Scholar] [CrossRef]
- Liu, H.; Xu, T.; Xu, S.; Ma, L.; Han, X.; Wang, X.; Zhang, X.; Hu, L.; Zhao, N.; Chen, Y.; et al. Effect of dietary concentrate to forage ratio on growth performance, rumen fermentation and bacterial diversity of Tibetan sheep under barn feeding on the Qinghai-Tibetan plateau. PeerJ 2019, 7, e7462. [Google Scholar] [CrossRef]
- Stevenson, D.M.; Weimer, P.J. Dominance of Prevotella and low abundance of classical ruminal bacterial species in the bovine rumen revealed by relative quantification real-time PCR. Appl Microbiol Biotechnol 2007, 75, 165–174. [Google Scholar] [CrossRef]
- Henao-Mejia, J.; Elinav, E.; Jin, C.; Hao, L.; Mehal, W.Z.; Strowig, T.; Thaiss, C.A.; Kau, A.L.; Eisenbarth, S.C.; Jurczak, M.J.; et al. Inflammasome-mediated dysbiosis regulates progression of NAFLD and obesity. Nature 2012, 482, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Ren, Q.; Si, H.; Yan, X.; Liu, C.; Ding, L.; Long, R.; Li, Z.; Qiu, Q. Bacterial communities in the solid, liquid, dorsal, and ventral epithelium fractions of yak (Bos grunniens) rumen. Microbiologyopen 2020, 9, e963. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Altermann, E.; Leahy, S.C.; Jauregui, R.; Jonker, A.; Henderson, G.; Kittelmann, S.; Attwood, G.T.; Kamke, J.; Waters, S.M.; et al. Genomic insights into the physiology of Quinella, an iconic uncultured rumen bacterium. Nat Commun 2022, 13, 6240. [Google Scholar] [CrossRef]
- Solden, L.M.; Hoyt, D.W.; Collins, W.B.; Plank, J.E.; Daly, R.A.; Hildebrand, E.; Beavers, T.J.; Wolfe, R.; Nicora, C.D.; Purvine, S.O.; et al. New roles in hemicellulosic sugar fermentation for the uncultivated Bacteroidetes family BS11. Isme j 2017, 11, 691–703. [Google Scholar] [CrossRef]
- Wu, Y.; Yao, Y.; Dong, M.; Xia, T.; Li, D.; Xie, M.; Wu, J.; Wen, A.; Wang, Q.; Zhu, G.; et al. Characterisation of the gut microbial community of rhesus macaques in high-altitude environments. BMC Microbiol 2020, 20, 68. [Google Scholar] [CrossRef]
- Nordhoff, M.; Moter, A.; Schrank, K.; Wieler, L.H. High prevalence of treponemes in bovine digital dermatitis-a molecular epidemiology. Vet Microbiol 2008, 131, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Bessman, S.P.; Rossen, J.; Layne, E.C. Gamma-Aminobutyric acid-glutamic acid transamination in brain. J Biol Chem 1953, 201, 385–391. [Google Scholar] [CrossRef]
- van Zijderveld, S.M.; Gerrits, W.J.; Apajalahti, J.A.; Newbold, J.R.; Dijkstra, J.; Leng, R.A.; Perdok, H.B. Nitrate and sulfate: Effective alternative hydrogen sinks for mitigation of ruminal methane production in sheep. J Dairy Sci 2010, 93, 5856–5866. [Google Scholar] [CrossRef]
- Li, Q.S.; Wang, R.; Ma, Z.Y.; Zhang, X.M.; Jiao, J.Z.; Zhang, Z.G.; Ungerfeld, E.M.; Yi, K.L.; Zhang, B.Z.; Long, L.; et al. Dietary selection of metabolically distinct microorganisms drives hydrogen metabolism in ruminants. Isme j 2022, 16, 2535–2546. [Google Scholar] [CrossRef]
- Harada, E.; Yamaguchi, Y.; Koizumi, N.; Hiroshi, S. Cadmium stress induces production of thiol compounds and transcripts for enzymes involved in sulfur assimilation pathways in Arabidopsis. Journal of Plant Physiology 2002, 159, 445–448. [Google Scholar] [CrossRef]
- Dowd, P.; Ham, S.W.; Naganathan, S.; Hershline, R. The mechanism of action of vitamin K. Annu Rev Nutr 1995, 15, 419–440. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, W.H. The cellulosome and cellulose degradation by anaerobic bacteria. Appl Microbiol Biotechnol 2001, 56, 634–649. [Google Scholar] [CrossRef] [PubMed]
- Myer, P.R.; Wells, J.E.; Smith, T.P.; Kuehn, L.A.; Freetly, H.C. Cecum microbial communities from steers differing in feed efficiency. J Anim Sci 2015, 93, 5327–5340. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Yang, C.; Guan, L.L.; Wang, J.; Xue, M.; Liu, J.X. Persistence of Cellulolytic Bacteria Fibrobacter and Treponema After Short-Term Corn Stover-Based Dietary Intervention Reveals the Potential to Improve Rumen Fibrolytic Function. Front Microbiol 2018, 9, 1363. [Google Scholar] [CrossRef] [PubMed]
- Paster, B.J.; Canale-Parola, E. Physiological diversity of rumen spirochetes. Appl Environ Microbiol 1982, 43, 686–693. [Google Scholar] [CrossRef]
- Azad, E.; Derakhshani, H.; Forster, R.J.; Gruninger, R.J.; Acharya, S.; McAllister, T.A.; Khafipour, E. Characterization of the rumen and fecal microbiome in bloated and non-bloated cattle grazing alfalfa pastures and subjected to bloat prevention strategies. Sci Rep 2019, 9, 4272. [Google Scholar] [CrossRef]
- de Oliveira, M.N.; Jewell, K.A.; Freitas, F.S.; Benjamin, L.A.; Tótola, M.R.; Borges, A.C.; Moraes, C.A.; Suen, G. Characterizing the microbiota across the gastrointestinal tract of a Brazilian Nelore steer. Vet Microbiol 2013, 164, 307–314. [Google Scholar] [CrossRef]
Figure 1.
α diversity and richness in ruminal bacterial in yaks and cattle given high concentrate diets.
Figure 1.
α diversity and richness in ruminal bacterial in yaks and cattle given high concentrate diets.
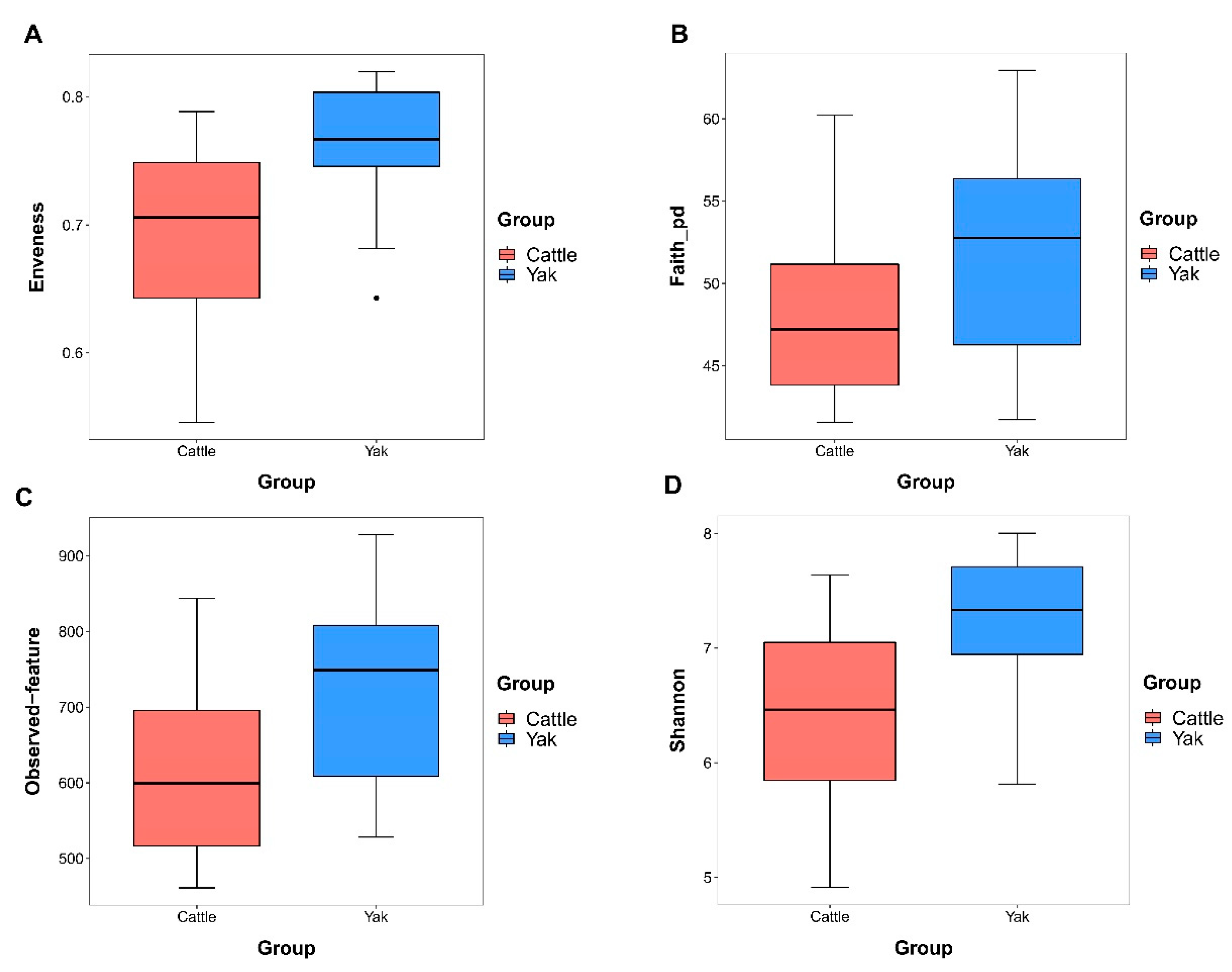
Figure 2.
(A) PCoA plots and (B) Venn diagrams of the rumen bacterial community. (C) Cladogram shows rumen bacteria from phylum level to genus level based on the linear discriminant analysis (LDA) effect size (LEfSe) analysis in yaks and cattle given high concentrate diets.
Figure 2.
(A) PCoA plots and (B) Venn diagrams of the rumen bacterial community. (C) Cladogram shows rumen bacteria from phylum level to genus level based on the linear discriminant analysis (LDA) effect size (LEfSe) analysis in yaks and cattle given high concentrate diets.
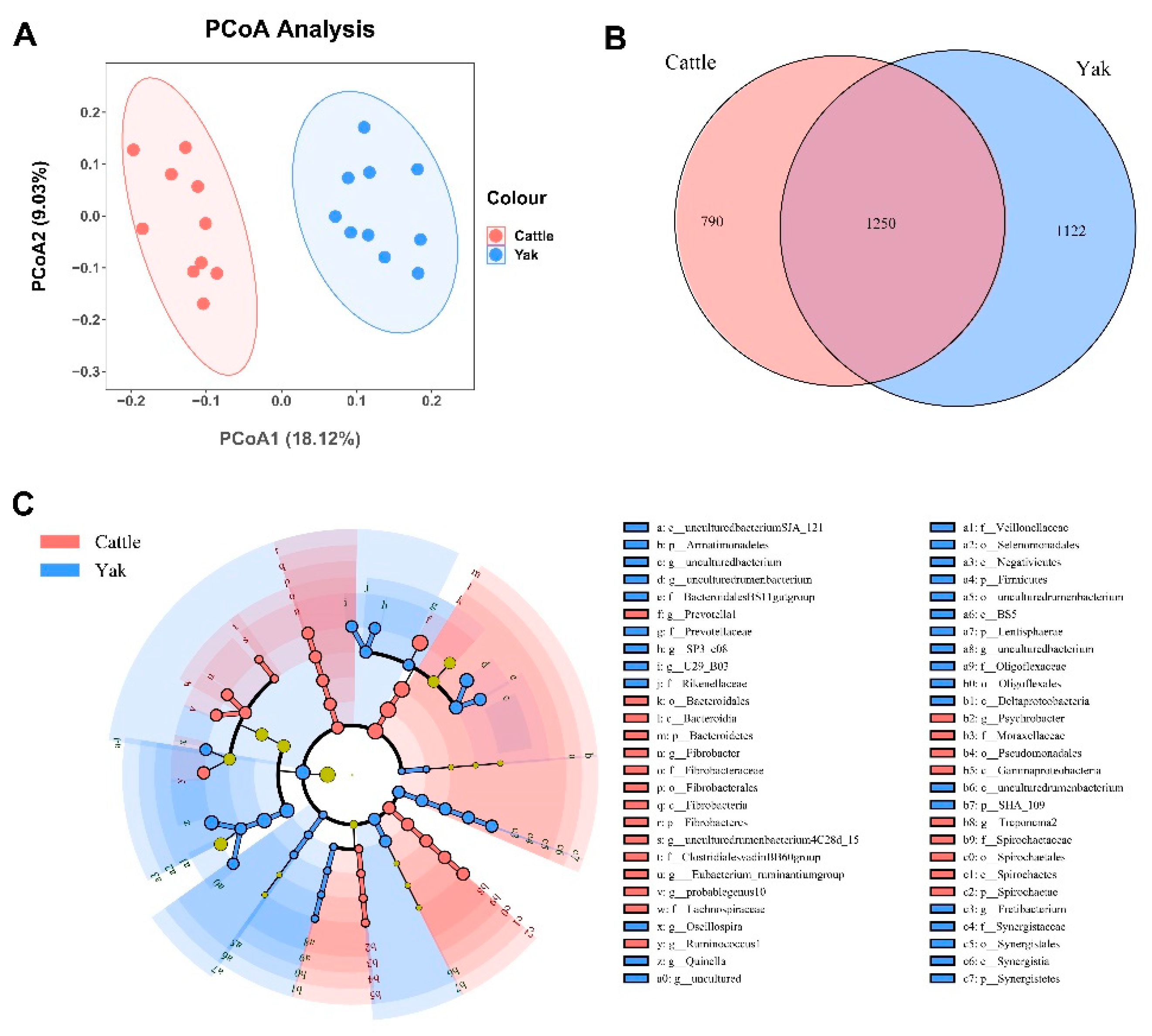
Figure 3.
(A) The rumen bacterial composition at the phylum level (the relative abundance > 0.5%) and bacteria in cattle and yaks. (B) The rumen bacterial composition at the genus level (top 20) and bacteria cattle and yaks given high concentrate diets. A single asterisk (*) indicates a signifcant diference at P<0.05, while double asterisks (**) indicate a signifcant diference at P<0.01.
Figure 3.
(A) The rumen bacterial composition at the phylum level (the relative abundance > 0.5%) and bacteria in cattle and yaks. (B) The rumen bacterial composition at the genus level (top 20) and bacteria cattle and yaks given high concentrate diets. A single asterisk (*) indicates a signifcant diference at P<0.05, while double asterisks (**) indicate a signifcant diference at P<0.01.
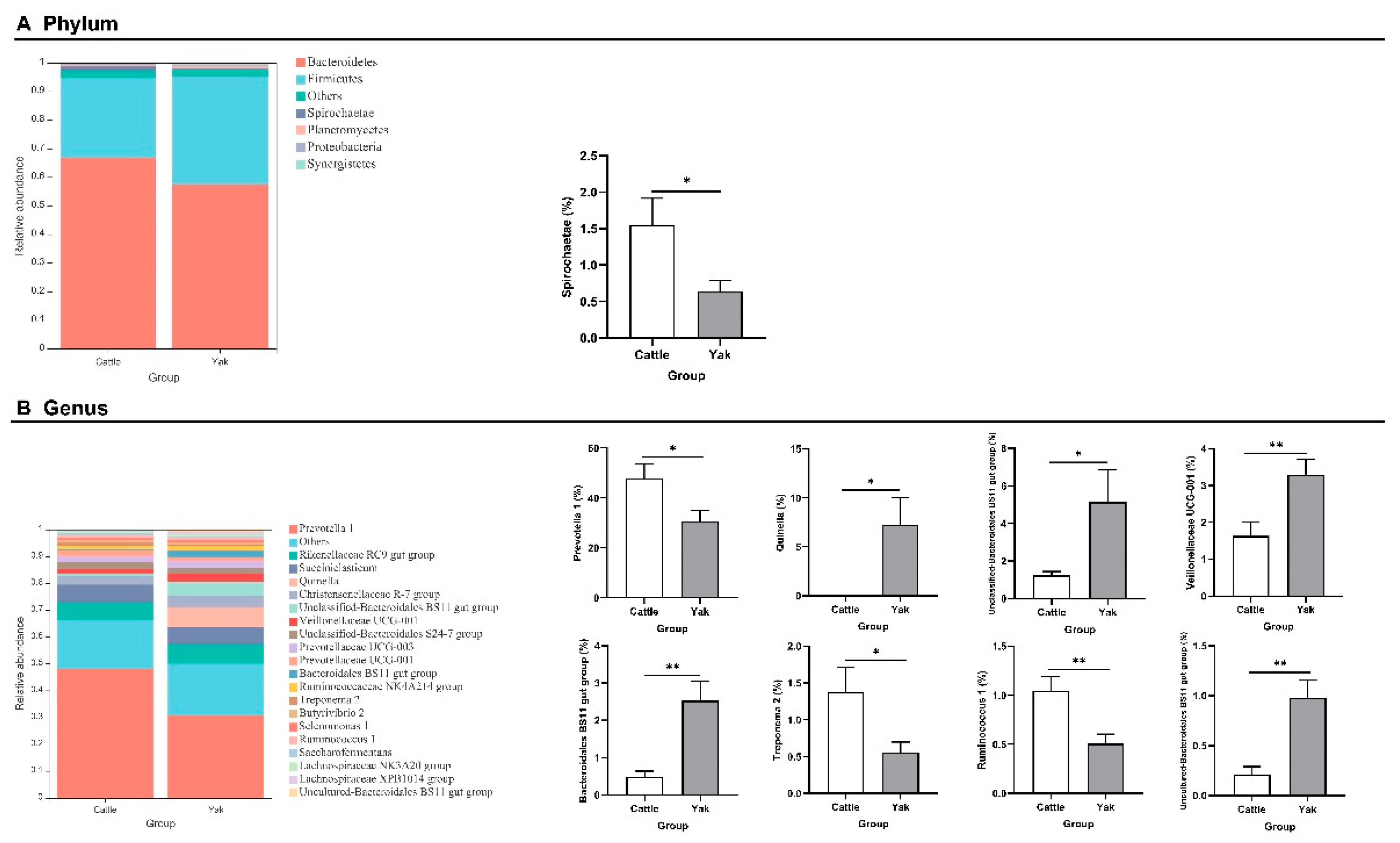

Table 4.
Feed ingredients and chemical composition of diets (DM basis).
| Ingredients, % | Nutrient levels, % | ||
|---|---|---|---|
| Oat hay | 10 | DM | 60.89 |
| Corn silage | 20 | Calcium | 0.49 |
| Corn kernels | 40 | Phosphorus | 0.58 |
| Bran | 17 | Ether extract | 2.77 |
| Soybean meal | 5 | Crude protein | 16.19 |
| Rapeseed cake | 5 | 2ME, MJ/kg | 11.96 |
| Mineral-vitamin premix1 | 2 | NDF | 29.44 |
| Baking soda | 1 | ADF | 16.54 |
1 The premix provided the following per kilogram of diet: vitamin A, 150 kIU/kg; vitamin D3, 30 kIU/kg; vitamin E, 600 IU/kg; vitamin K3, 10 mg/kg; vitamin B1, 10 mg/kg; vitamin B2, 28 mg/kg; vitamin B12, 100 μg/kg; vitamin B9, 4 mg/kg; vitamin B3, 200 mg/kg; vitamin B5, 50 mg/kg; vitamin B7, 2 mg/kg; NaCl, 225 g/kg; I, 16.5 mg/kg; Se, 6 mg/kg; Co, 4 mg/kg; Cu, 0.2 g/kg; Fe, 2.25 g/kg; P, 21 g/kg; Ca, 130 g/kg; Zn, 1.1 g/kg; Mg, 15.5 g/kg; and Mn, 1.6 g/kg. 2 ME was calculated value (Liu, 2018), while the other values were measured.
Table 1.
Dry matter intake, water intake, average daily gain and feed to gain ratio in yaks and cattle given high concentrate diets.
Table 1.
Dry matter intake, water intake, average daily gain and feed to gain ratio in yaks and cattle given high concentrate diets.
| Items | Cattle | Yak | P-value |
|---|---|---|---|
| Initial body weight (kg) | 256.9±8.6 | 194.3±13.7 | 0.001 |
| Dry matter intake (kg/d) | 9.63±0.51 | 5.49±0.32 | <0.001 |
| Daily water intake (kg/d) | 35.59±1.07 | 20.38±0.96 | <0.001 |
| Dry matter intake/MW(g/kgW0.75) | 128.11±5.52 | 92.24±3.76 | <0.001 |
| Daily water intake/MW(g/kgW0.75) | 475.97±19.13 | 343.96±14.50 | <0.001 |
| Average daily gain (kg/d) | 1.09±0.08 | 0.87±0.05 | 0.039 |
| Feed to gain ratio | 9.09±0.60 | 6.41±0.49 | 0.004 |
Table 2.
Serum biochemical indexes in cattle and yaks given high concentrate diets.
| Items | Cattle | Yak | P-value |
|---|---|---|---|
| Aspartate aminotransferase (U/L) | 91.80±7.62 | 64.40±3.94 | 0.005 |
| Alanine aminotransferase (U/L) | 27.20±1.97 | 30.50±2.07 | 0.263 |
| Total protein (g/L) | 76.55±2.48 | 73.71±3.17 | 0.491 |
| Albumin (g/L) | 35.50±0.48 | 37.24±0.91 | 0.113 |
| Globulin (g/L) | 41.07±2.70 | 36.48±2.67 | 0.242 |
| Albumin/Globulin | 0.89±0.06 | 1.07±0.06 | 0.048 |
| Alkaline phosphatase (U/L) | 218.50±22.93 | 165.60±13.54 | 0.066 |
| Total bilirubin (μmol/L) | 1.98±0.23 | 1.23±0.33 | 0.082 |
| Direct bilirubin (μmol/L) | 0.53±0.09 | 0.50±0.22 | 0.904 |
| Indirect bilirubin (μmol/L) | 1.45±0.15 | 0.81±0.20 | 0.021 |
| Creatine kinase (U/L) | 477.80±170.74 | 113.00±12.53 | 0.047 |
| Lactate dehydrogenase (IU/L) | 1431.60±55.05 | 940.00±53.21 | <0.001 |
| Glucose (mmol/L) | 4.43±0.12 | 4.58±0.24 | 0.578 |
| Urea (mmol/L) | 2.95±0.33 | 4.29±0.35 | 0.012 |
| Uric acid (μmol/L) | 16.40±1.83 | 34.40±3.06 | <0.001 |
| Calcium (mmol/L) | 2.71±0.06 | 2.53±0.10 | 0.144 |
| Phosphorus (mmol/L) | 2.01±0.13 | 2.24±0.12 | 0.219 |
| Total cholesterol (mmol/L) | 2.81±0.16 | 1.90±0.18 | 0.001 |
| Triglyceride (mmol/L) | 0.26±0.02 | 0.34±0.04 | 0.101 |
Table 3.
Ruminal parameters in cattle and yaks given high concentrate diets.
| Items | Cattle | Yak | P-value |
|---|---|---|---|
| pH | 6.13±0.03 | 6.00±0.03 | 0.006 |
| Lactate (mmol/L) | 0.25±0.04 | 0.38±0.04 | 0.028 |
| TVFA (mmol/L) | 76.02±7.70 | 85.11±6.88 | 0.390 |
| Acetate (%) | 59.34±0.81 | 56.04±0.50 | 0.003 |
| Propionate (%) | 19.48±0.79 | 21.13±0.65 | 0.125 |
| Butyrate (%) | 15.81±0.87 | 17.14±0.65 | 0.232 |
| Isobutyrate (%) | 1.56±0.20 | 1.51±0.19 | 0.861 |
| Isovalerate (%) | 2.02±0.26 | 2.13±0.26 | 0.773 |
| Valerate (%) | 1.80±0.21 | 2.06±0.24 | 0.428 |
| Acetate:Propionate | 3.10±0.15 | 2.67±0.07 | 0.022 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



