
Submitted:
06 September 2023
Posted:
11 September 2023
You are already at the latest version
Abstract
Cyanobacteria are promising organisms for the sustainable production of various biotechnologi-cal interesting products. Due to their energy production via photosynthesis, the cultivation of cyanobacteria expands the CO2 cycle. Most cyanobacteria form biofilms on surfaces in their natu-ral environment by surrounding the cells with a self-produced matrix of extracellular polymeric substances (EPS) that hold the cells together. These special growth properties need special reac-tors for cultivation. By immobilizing cyanobacteria on carriers, systems currently established in industry could also be used for biofilm formers. Various artificial carriers for immobilized growth of cyanobacteria and microalgae have already been described in the literature. However, the use of waste materials or natural biodegradable carriers would be more sustainable and is, therefore, the focus of this study.
Dried Luffa cylindrica, zeolite, and corn stalks were investigated for their use as carriers for cyanobacteria. L. cylindrica was shown to be an excellent natural carrier for (i) Anabaena cylindrica (ii) Nostoc muscorum 1453-12a, and (III) Nostoc muscorum 1453-12b. Higher or at least similar growth rates were achieved when cyanobacteria were cultivated with L. cylindrica compared to submerged cultivation. Additionally, the production of EPS and C-phycocyanin was increased at least 1.4-fold in all strains by culturing on L. cylindrica. The improved growth could be explained on the one hand by the high surface area of L. cylindrica and its properties, and on the other hand by the release of growth-promoting nutrients from L. cylindrica to the medium.
Keywords:
biofilm characterization
; cyanobacteria
; phototrophic biofilms
; carrier
; loofah sponge
; Luffa cylindrica
1. Introduction
Cyanobacteria are among the oldest organisms on earth and are particularly known for their oxygenic photosynthesis [1]. These gram-negative bacteria with photosynthetic activity can convert CO2 and water into organic compounds under the influence of light and produce oxygen as a by-product. They occur in nature both aquatically in freshwater or marine ecosystems and terrestrially on soils or even on rocks and tree trunks. Cyanobacteria colonize many ecological niches due to their adaptability to extreme environmental conditions [2]. Cyanobacteria are widely used in various fields, such as the food industry, biofuel production due to their high lipid content, and medical applications due to their antibacterial secondary metabolites [3,4,5]. On an industrial scale, aquatic cyanobacteria, which achieve good productivity submersed, are mostly used. However, most cyanobacteria grow in their natural form as a biofilm surrounded by a self-produced matrix of extracellular polymeric substances (EPS) on surfaces and often show poorer growth rates in classical submerged systems such as open ponds or closed tubular systems [6]. To improve growth, special biofilm photobioreactors that mimic the natural habitat of different strains have been increasingly developed in recent years and are summarized by Ennaceri et al. [7]. Another approach is to use existing photobioreactor systems and provide the cyanobacteria with a surface in the form of different carriers.
Cyanobacteria offer promising biotechnological products [8] but grow very slowly compared to industrially used strains such as Escherichia coli or Bacillus subtilis. However, cyanobacteria offer the advantage of expanding the CO2 cycle due to their energy production via photosynthesis, thus counteracting the ongoing climate change. Furthermore, they do not require an additional sugar source, which means that no plate-tank problem arises. In addition, biofilm-forming cyanobacteria are very tolerant of fluctuating cultivation conditions [9], which means that close monitoring and control of the processes is not necessary in every case, which in turn can save energy and thus CO2. Overall, the cultivation of cyanobacteria is therefore sustainable. To maintain this aspect of sustainability, the carriers used for the cultivation of biofilm-forming cyanobacteria in submerged systems should also be biodegradable and thus sustainable. So far, various materials have been successfully used as carriers for immobilized cultivation of cyanobacteria such as fibers and fabrics [10,11], open-cell foams such as polyurethane (PUR) [12,13], or high-density polyethylene (HDPE) [14]. Plastics are always problematic if a product is to be produced sustainably. Furthermore, if the plastic is to be reused, recovery is costly and professional disposal is necessary. This study aims to find new biodegradable carriers for the cultivation of cyanobacteria and to overcome these problems. The advantages of biodegradable carriers are for instance the production of natural fertilizer in the form of biodegradable carriers overgrown with cyanobacteria. It would also be conceivable to use the overgrown carriers directly as animal feed. But also, the processing of biomass can be simplified by using natural materials as carriers. Thus, harvesting (in comparison to plastic carriers) is simplified and the potential products can be isolated directly from the carriers since no harmful substances are contained in them. Furthermore, the production of carriers should not be complicated like the production of fibers and fabrics. For this reason, the influence of different natural, biodegradable carriers on biofilm-forming cyanobacteria (terrestrial strains Nosotc muscorum 1453-12a and 1453-12b as well as aquatic strain Anabaena cylindrica) is investigated in this work, which would also be transferable to an industrial scale. The use of cyanobacteria on an industrial scale allows the development of new sustainable production organisms that simultaneously expand the CO2 cycle leach and thus counteract climate change.
2. Materials and Methods
2.1. Pre-culture
The strains (Anabaena cylindrica 1403-2, Nostoc muscorum 1453-12a and Nostoc muscorum 1453-12b) were ordered at the Culture Collection of Algae at the University of Göttingen, Germany (SAG). Pre-cultures were cultivated for 14 days in an illuminated shaking incubator (Multitron S-000115689, Infors HT, Bottmingen, Switzerland) at 120 rpm with 2.5 cm eccentricity using 250 ml Erlenmeyer flasks without baffles containing 50 ml of standard BG11 medium [15]. A constant temperature (30 °C) and illumination (140 µmolphotons m−2 s−1) were set. Every four weeks the biomass was centrifuged to change the medium and 50 mL of new BG11 medium was added. For harvesting of pre-cultures, cell suspension was transferred into a 50 ml plastic reaction vessel and centrifuged for 15 min at 8.000 g (centrifuge 383K, Hermle Labortechnik GmbH, Wehingen, Germany). The supernatant was discarded, and the biomass pellet was used for further experiments.
2.2. Cultivation of cyanobacteria on various carrier
100 mL Erlenmeyer flasks were filled to approximately 25 mL with the different carriers used (Corn stalk pieces of 1 cm3, dried Luffa cylindrica pieces of 1 cm3, and zeolite (CAS: 1318-02-1, Merck, Germany) with a particle size of 1–2.5 mm). This corresponded to approximately 1 g L. cylindrica, 6 g zeolite, and 10 g corn stalk. All carriers were sterilized by autoclaving. Following this, the Erlenmeyer flasks were filled with 50 mL BG11 and inoculated with approximately 0.2 g of Nostoc muscorum 1453-12a cell wet weight (CWW). Cultivation was carried out over a cultivation period of 14 days in a shaking incubator at 30 °C and 120 rpm. Illumination was continuous at a light intensity of 100 µmolphotons m 2 s−1. After 14 days, the supernatant and the overgrown surfaces were each transferred to a 50 mL plastic reaction tube, and the biomass growing at the flask edge was transferred to a 2 mL plastic reaction tube. The wet biomass was then lyophilized (LOC-1M Alpha 2-4, Christ, Osterode, Germany) for 24 h at -20 °C and 1 mbar to determine the cell dry weight (CDW). The overgrown carriers were dried at 80 °C in a drying oven (VT 5050 EKP, Haraeus Holding GmbH, Hanau, Germany). Thereafter, the fraction of cells that grew immobilized on the carrier, in the supernatant, and on the flask wall were determined gravimetrically. Since the salt of the BG11 medium also influences the masses of the individual components, three Erlenmeyer flasks were filled only with BG11 medium and the corresponding carrier (n = 3). After 14 days of cultivation, the carrier and the supernatant were treated the same way as the other Erlenmeyer flasks to determine the impact of salt on the weight of the carrier.
2.3. Determination of released growth-promoting substances by Luffa cylindrica
The aim was to investigate whether L. cylindrica releases growth-promoting substances into the medium. For this purpose, L. cylindrica was first ground using a coffee grinder (type 364 NIVONA Apperete GmbH Nuremberg, Germany). The resulting powdery material was weighed in a fine balance and transferred to a 50 ml plastic reaction tube. It was then made up with ultrapure water until a concentration of 20 respectively 40 g L−1 was reached in each plastic reaction tube. The plastic reaction tubes were then placed in an overhead shaker (Intellimixer, Neolab, Heidelberg, Germany) and incubated for 24 hours at room temperature. This was followed by centrifugation (Rotina 48 R, Hermle AG Hettich, Gosheim, Germany) at 4,000 rpm for 30 min. After decantation, the supernatants were used to prepare BG11 medium instead of using ultrapure water as usual.
2.4. Cultivation of N. muscorum 1453-12a in BG11 and BG11 with soaked L. cylindrica
N. muscorum 143-12a was cultivated with BG11 medium and with the BG11 medium to which the supernatant of the soaked L. cylindrica was added (BG11 medium + Luffa extract) to investigate the influence of potential growth-promoting substances from L. cylindrica on the growth of cyanobacteria. 50 mL of each medium were inoculated with 0.1 g of N. muscorum 1453-12a CWW. Cultivation was carried out in 250 mL Erlenmeyer flasks over a cultivation period of 14 days in a shaking incubator at 30 °C and 120 rpm. Illumination was continuous at a light intensity of 140 µmolphotons m 2 s−1.
2.5. Determination of CDW, EPS, Phycobiliproteins, and Pigments
A physical method modified according to Stiefelmaier and Strieth [16] was used to extract the EPS from the CWW. First, 5 ml of pre-heated (55 °C) deionized water was added to CWW in a 50 ml plastic reaction tube and incubated for 30 min at 55 °C in an overhead shaker (Intellimixer, Neolab, Heidelberg, Germany). The suspension was then treated in an ultrasonic bath (Sonorex Digiplus, Bandelin, Berlin, Germany) for 10 minutes at 20 °C and an ultrasound intensity of 100 W and was then centrifuged at 4,000 rpm for 15 minutes to isolate the supernatant, which contained the dissolved EPS. The supernatant was transferred to a new 50 ml plastic reaction tube. Both, the supernatant and the CWW pellet were lyophilized at 20 °C and 1 mbar for 48 hours in a freeze dryer. The mass of the obtained EPS and CDW were determined gravimetrically. The CDW was used for extraction of chlorophyll-a, carotenoids, and the phycobiliproteins C-phycocyanin, phycoerythrin, and allophycocyanin.
For extraction of phycobiliproteins, the lyophilized CDW was resuspended in 1mL 0.5 M potassium phosphate buffer (PPB), and 50 Ma-% glass beads were added (diameter: 0.7–0.9 mm). Cell disruption was performed using a vibrating mill (MM 301, Retsch Technology GmbH, Haan, Germany) for 10 min at a frequency of 30 s−1. Subsequently, the digested biomass was transferred to a 15 ml plastic reaction tube and 0.25 ml of the 0.5 M PPB was added per milligram of CDW. After incubation at 55 °C for 30 min in a drying oven, the suspension was stored at 4 °C for 24 h in the dark. Then, the suspension was centrifuged at 4,000 rpm for 10 min. The supernatant was transferred to a new 15 mL plastic reaction tube and the absorbance was measured at wavelengths 615, 652, and 562 nm (Cary 60 UV-Vis, Agilent Technologies, Santa Clara, USA). The concentrations of C-phycocyanin, allophycocyanin, and phycoerythrin were calculated according to Bennet and Bogorad [17] using the measured absorbance.
After extraction of phycobiliproteins, chlorophyll-a, and carotenoid were extracted from the remaining CDW. For this purpose, 0.5 ml of methanol saturated with CaCO3 was added per milligram of CDW. The suspension was then incubated for 24 h at 4 °C in the dark. After centrifugation at 4,000 rpm for 15 min, the supernatant was transferred to a new 15 mL plastic reaction tube. The concentration of chlorophyll a and carotenoid content were calculated by measuring the absorbances at 461, 665, 652, and 665nm according to Porra [18] and Chamovitz [19].
2.6. Analyzation of the cultivation medium
Using ion chromatography (model 930 Compact IC Flex from Deutsche METROHM GmbH & Co. KG), the supernatants were analyzed to determine the composition of anions and cations in the medium over the cultivation period. Additionally, components of the medium were quantified by compact ion exchange chromatography with an inline system for dialysis (930 Compact IC Flex, Metrohm, Filderstadt, Germany) with a conductivity detector over the cultivation period. Anions (chloride, nitrite, nitrate, phosphate, and sulfate) were measured with an anion column (Metrosep A Supp 5–250/4.0, Metrohm) using 1 mM NaHCO3 and 3.2 mM Na2CO3 as mobile phase at a flow rate of 0.7 mL min−1. Cations (ammonium, magnesium, sodium, potassium) were measured with a cation column (Metrosep C6-250/4.0, Metrohm) using 4 mM nitric acid and 0.7 mM dipicolin acid at a flow rate of 0.9 mL min−1. In both cases, the oven temperature was set to 35 °C. HPLC analysis of sugar monomers was carried out with an HPLC system (ESA Inc. 542 autosampler (Chelmsford, Massachusetts, USA), Azura pump P 6.1 L (Knauer GmbH, Berlin, Germany)) equipped with a refractive index detector, and a BioRad Aminex HPX- 87H column (300 × 7.8 mm) (Hercules, California, USA). A column temperature (80 °C), and 2.5 mM H2SO4 as mobile phase at a flow rate of 0.6 mL min−1 were used
3. Results
3.1. Screening for biodegradable carriers as cultivation surface for phototrophic biofilms
The production of biomass and biotechnological valuable products as well as the waste after the downstream process can be made more sustainable using biodegradable carriers as cultivation surfaces. Therefore, different carriers were tested including (i) corn stalk (waste), (ii) L. cylindrica, and (iii) zeolite. On the corn stalk, cyanobacteria grew exclusively on the flask wall and faded after a few days (see Table 1). No growth could be detected on the corn stalk. Since cyanobacteria do not grow axenically [20], the sugar contained in the growth bodies probably favors the growth of an unknown contaminant that overgrew the cyanobacteria. After 14 days of cultivation of N. muscorum 1453-12a with zeolite, 89.08 ± 7.00% of the cells were in the supernatant, while the rest grew immobilized on the flask wall (see Figure 1 and Table 2). In contrast, Filippidis et al. [21] demonstrated a decrease of cyanobacteria in seawater by 74.93% for filamentous and 50.94% for colonial strains by the addition of zeolite. They were also able to demonstrate the accumulation of cyanobacteria in the pores of the zeolite by scanning electron microscopy. Filippidis et al. used zeolite with a particle size of <0.5 mm, while in these experiments a larger particle size of 1–2.5 mm was used, which should provide a higher growth surface on the outer surface of the particles. During cultivation in the Erlenmeyer flask, the cells were probably transported to the top of the flask wall by the heavy zeolite particles during shaking, where they grew immobilized. In the experiments of Filippidis et al. [21], the zeolite was merely added to seawater without agitating the water.

Table 1.
Summarized results of cultivation of N. muscorum 1453-12a in Erlenmeyer flasks with various carriers. Cultivation parameters: 100 mL shaking flasks, 25 mL BG11-medium, 1 g L. cylindrica, 6 g zeolite, 6 g 3D-printed Luffa, 30 °C, 100 µmolPhotons m−2s−1, n = 3.
Table 1.
Summarized results of cultivation of N. muscorum 1453-12a in Erlenmeyer flasks with various carriers. Cultivation parameters: 100 mL shaking flasks, 25 mL BG11-medium, 1 g L. cylindrica, 6 g zeolite, 6 g 3D-printed Luffa, 30 °C, 100 µmolPhotons m−2s−1, n = 3.
| Carrier | Picture of Erlenmeyer Flask Filled with the Carrier before Cultivation | Picture of Erlenmeyer Flask Filled with the Carrier after Cultivation | Growth | Notes |
|---|---|---|---|---|
 | ||||
This probably makes it more possible for cells to diffuse into the particles. On L. cylindrica, 97 ± 4% of the cells grew immobilized (see Figure 1 and Table 1) which can probably be attributed to the structure of L. cylindrica. In-na et al. [22] used L. cylindrica as a 3D scaffold to form a natural biocomposite by immobilization of cyanobacteria for CO2 fixation. Other studies immobilized microalgae and fungi on Luffa plants in wastewater treatment due to good immobilization quotes caused by the natural high surface of L. cylindrica [23,24]. However, it must be said that none of these studies investigated the growth of cyanobacteria or the impact of L. cylindrica on the composition and product formation of cyanobacteria.
Figure 1.
Proportion of CDW on the surface, flask wall, and supernatant to total CWW of N. muscorum 1453-12a with Luffa (=Luffa cylindrica), zeolite, and 3D-printed Luffa. Cultivation parameters: 100 mL shaking flasks, 25 mL BG11-medium, 1 g L. cylindrica, 6 g zeolite,6 g 3D-printed Luffa, 30 °C, 120 rpm, 140 µmolPhotons m−2s−1, n = 3.
Figure 1.
Proportion of CDW on the surface, flask wall, and supernatant to total CWW of N. muscorum 1453-12a with Luffa (=Luffa cylindrica), zeolite, and 3D-printed Luffa. Cultivation parameters: 100 mL shaking flasks, 25 mL BG11-medium, 1 g L. cylindrica, 6 g zeolite,6 g 3D-printed Luffa, 30 °C, 120 rpm, 140 µmolPhotons m−2s−1, n = 3.
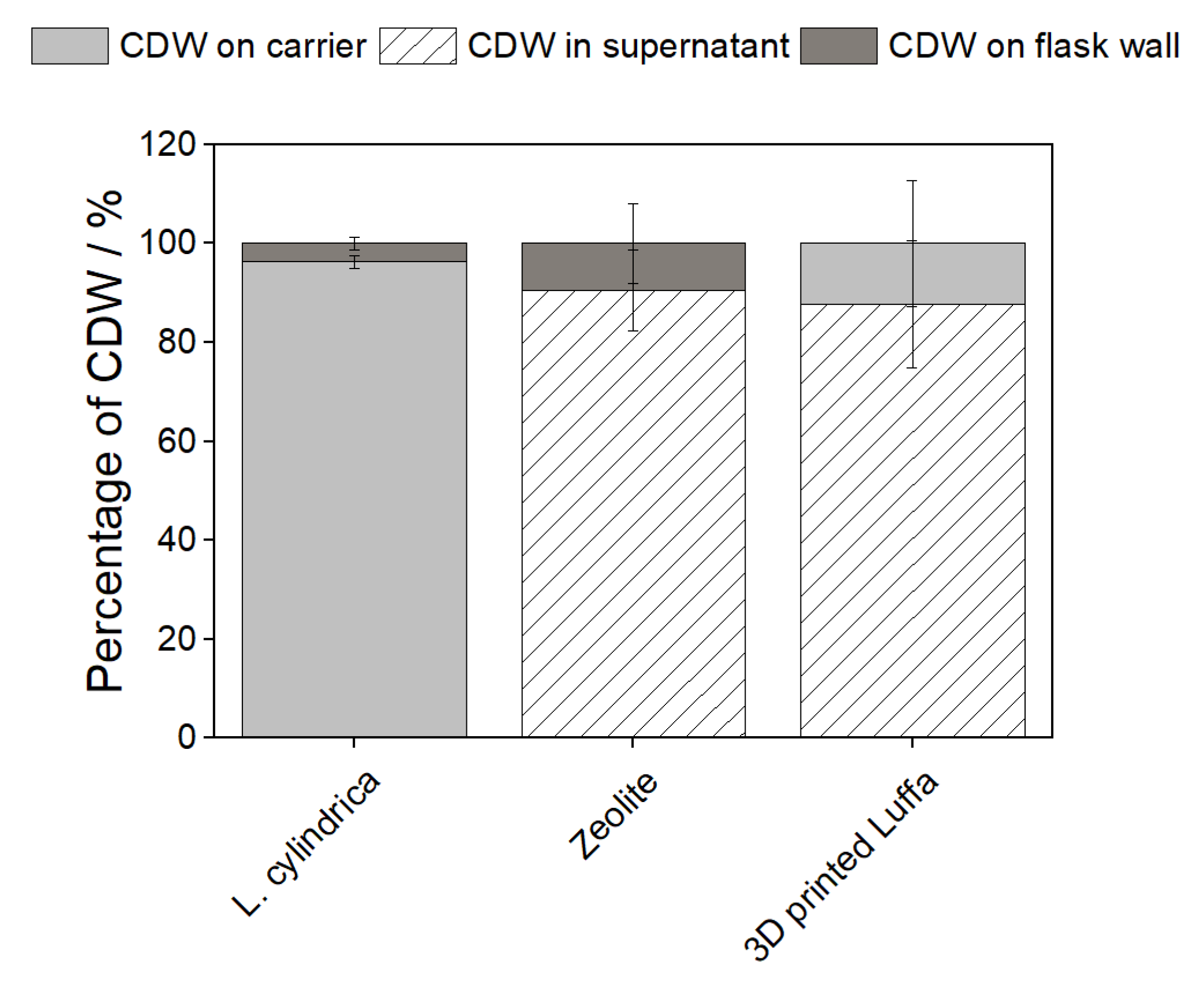
It is composed of rough fibers [207] that together form growth bodies with a high porosity of about 79–93% with a high specific pore volume of 21–29 cm3 g−1 [25]. These properties make the sponge act as a kind of sieve. By shaking the Erlenmeyer flasks, the cells probably became entangled in the pores of the L. cylindrica and stuck to it. To investigate exactly this influence, an artificial L. cylindrica with the same structure was 3D-printed from plastic, and the growth of N. muscorum 1453-12a was investigated as with the L. cylindrica carriers by using 5 pieces of 3D-printed Luffa (6 g). However, on the 3D-printed Luffa, 88 ± 13% of the cells were recovered in the supernatant (see Figure 1 and Table 1). This means that the cells were not only filtered out of the medium during cultivation on the natural L. cylindrica, but that active and natural immobilization took place. Cell adhesion of bacteria depends on hydrophobicity [26], which in turn depends on surface roughness. L. cylindrica has a moderately hydrophilic surface [27]. Hydrophilic properties were established for N. muscorum 1453-12a (corresponding to N. muscorum BB 90.3) after surface-associated cultivation in air and underwater [28], which explains the high immobilization rate on L. cylindrica. Additionally, a high surface roughness favors the adhesion of biofilms [29]. The lower surface roughness of the 3D-printed Luffa could be a reason for the poor attachment of cells to the surface, as it plays a crucial role, especially during the initial recruitment phase of biofilms. In future work, cultivations on wood-printed Luffa will take place to test the influence of the material on the immobilization of cyanobacteria.
In summary, it was shown that L. cylindrica is ideally suited for immobilized cultivation of N. muscorum 1453-12a.
3.1.1. Influence of Luffa cylindrica on the growth of various Cyanobacteria
Higher biomass productivity was observed for two strains, whereby A. cylindrica showed 1.23 times and N. muscorum 1453-12a 1.77 times higher growth rates compared to submerged cultivation. N. muscorum 1453-12b showed similar growth rates (see Figure 2A). This higher productivity compared to the cultivation without carrier could be attributed to the fact that 99% of bacteria grow in their natural form as biofilm [30] and thus like to attach to surfaces.
Figure 2.
Biomass production and EPS content of three different cyanobacteria cultivated for 14 days with and without Luffa cylindrica. CD = Cell Dry Weight. EPS = Extracellular Polymeric Substances. Cultivation parameters: 250 mL Erlenmeyer flasks, 50 mL BG11-medium, 20 g L. cylindrica, 30 °C, 120 rpm, 140 µmolPhotons m−2s−1, n = 3.
Figure 2.
Biomass production and EPS content of three different cyanobacteria cultivated for 14 days with and without Luffa cylindrica. CD = Cell Dry Weight. EPS = Extracellular Polymeric Substances. Cultivation parameters: 250 mL Erlenmeyer flasks, 50 mL BG11-medium, 20 g L. cylindrica, 30 °C, 120 rpm, 140 µmolPhotons m−2s−1, n = 3.
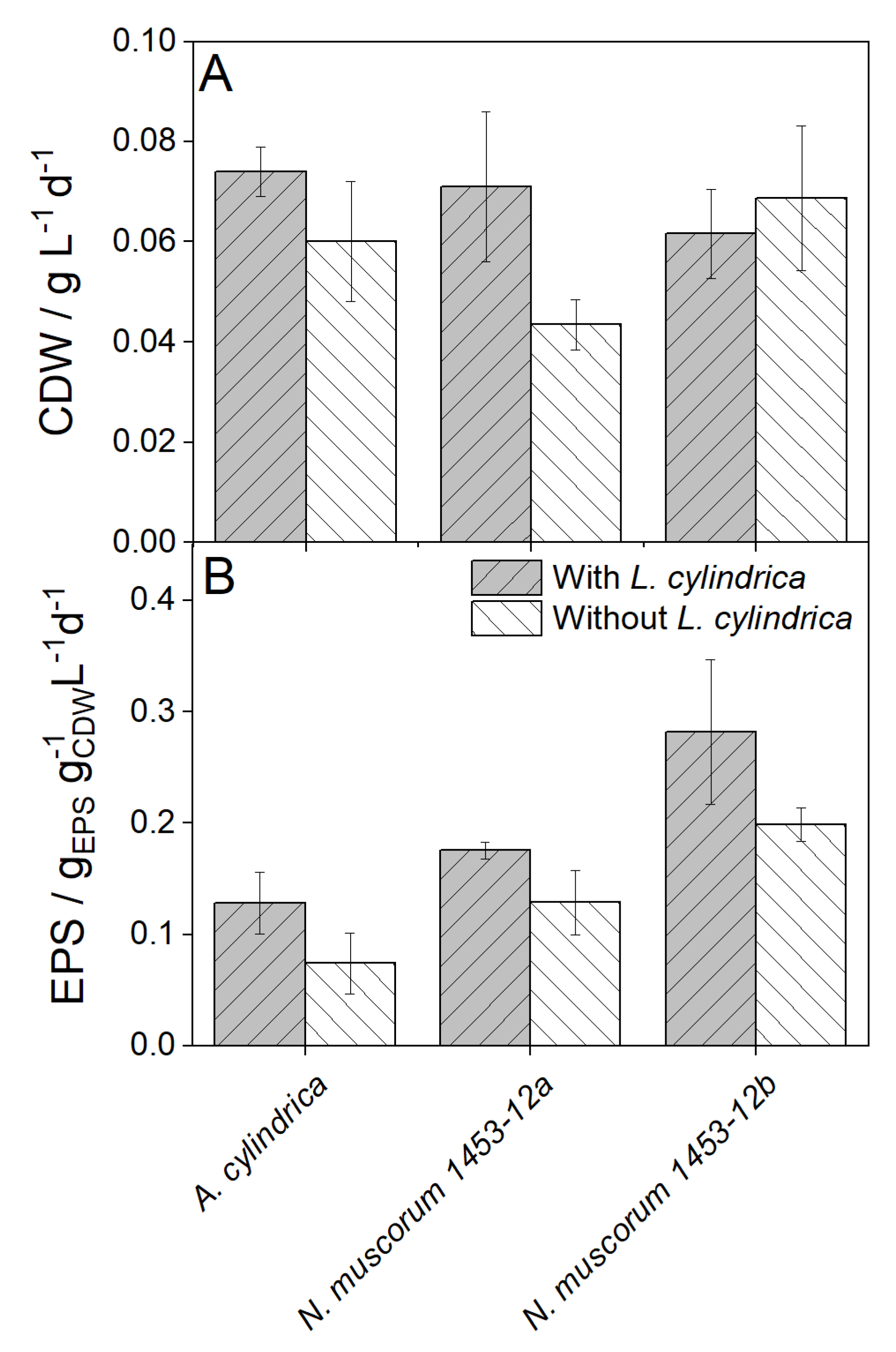
Also, many especially terrestrial cyanobacterial strains show enhanced growth when they are cultivated surface-associated compared to classical submerge cultivation in closed tubular systems for example [6]. Additionally, EPS contents were higher for all strains (A. cylindrica 1.75fold; N. muscorum 1453-12a 1.36fold and N. muscorum 1453-12b 1.4fold) when cyanobacteria were cultivated with L. cylindrica. This can have several reasons. One reason may be the immobilized growth. Here, Ekelhof and Melkonian [31] showed that Netrium digitu produced more EPS in the porous substrate reactor (i.e., immobilized) compared to submerged cultivation. Another reason could be increased nutrient availability due to substances released by L. cylindrica, which are then stored in the EPS [32]. Furthermore, It was shown for N. muscorum 1453-12b that 95 ± 5% of the cells were immobilized on L. cylindrica which is consistent with the results for N. muscorum 1453-12a. Although L. cylindrica causes relatively low light loss due to its high porosity of approximately 79-93%, this depends on the shape and volume of the L. cylindrica used for cultivation. Phycobiliproteins as well as pigment composition were investigated but no difference between both cultivation set-ups could be observed.
However, it could be shown, that biomass productivity can be improved when different cyanobacteria are cultivated with L. cylindrica. To find possible reasons for the better growth with loofah besides immobilization, potential growth-promoting substances from L. cylindrica were investigated in the next step.
3.2. Growth-promoting substances released by Luffa cylindrica
In the first step, the composition of the L. cylindrica used was determined according to the NREL method [33]. It consists of 14.89 ± 0.53 hemicellulose, 53.09 ± 1.75 cellulose, 9.70 ± 0.38 acid-insoluble lignin (AIL), 12.69 ± 0.42 acid-soluble lignin (ASL) and 0.35 ± 0.07 ash. These values match those described in the literature for L. cylindrica with hemicellulose (14–30%), cellulose (57–74%), lignin (1–20%) and ash (0.3–0.5%) [34,35]. Some cyanobacterial species can produce laccases that correlate with biomass formation. Laccase activities of up to 60 U ml−1 are described in the literature for Arthrospira platensis, but other species like for example Nostoc, Synechocystis, and Lyngbya strains show high productivities of laccase activity [36]. Future work should investigate whether the strains used in this work are also capable of producing laccases. If the strains used were able to produce laccases and thus degrade lignin, mixotrophic growth could explain the higher growth rates. In these experiments, the sugar concentration in the culture supernatant was determined and no sugars could be detected. However, released sugars might have been metabolized immediately by the cyanobacteria. To investigate whether the L. cylindrica releases growth-promoting substances into the medium, 20 g of ground L. cylindrica was soaked in distilled water for 24 hours, and then the supernatant was analysed for anions, cations, and sugars (see Table 2). It could be shown that no sugars were present in the supernatant. However, nutrients important for the growth of cyanobacteria such as nitrate, chloride, phosphate, and sulfate were present. N. muscorum 1453-12b showed enhanced growth in the growth phase when cultivated in the BG11 medium with Luffa extract compared to reference cultivation with BG11 (see Figure 3A). This could be attributed to the release of growth-promoting substances from L. cylindrica (see Table 2).
Table 2.
Ion concentrations of distilled water and 50 mL distilled water with 20 g soaked Luffa cylindrica measured with ion exchange chromatography and sugar concentration measured with HPLC.
Table 2.
Ion concentrations of distilled water and 50 mL distilled water with 20 g soaked Luffa cylindrica measured with ion exchange chromatography and sugar concentration measured with HPLC.
| Anions/Cations | The concentration of Ions in distilled water soaked with 20 g L. cylindrica [mg/L] | The concentration of Ions in distilled water [mg/L] |
|---|---|---|
| Nitrate | 4.36 ± 3.93 | 1.84 ± 1.60 |
| Nitrite | 0.91 ± 0.29 | 0.65 ± 0.02 |
| Chloride | 4.31 ± 1.23 | 0.61 ± 0.01 |
| Phosphate | 13.22 ± 9.92 | 0 |
| Sulfate | 10.51 ± 3.34 | 3.06 ± 0.02 |
| Sugar | 0 | 0 |
In this regard, the stationary phase was reached earlier when cultivated with Luffa extract than without, whereby the same biomass concentrations (~1.2 g L−1) were reached at the end (see Figure 3A). The power input in 250 mL Erlenmeyer flasks filled with 50 mL medium and a shaking frequency of 120 rpm (eccentricity 1.5 cm) is almost zero [37]. Accordingly, the gas exchange takes place only by diffusion at the surface. Oxygen limitations in cultivations of heterotrophic microorganisms in Erlenmeyer flasks have been described several times [38,39]. It is assumed that from a biomass concentration of 1.2 g L, the CO2 input into the system is no longer sufficient, resulting in the limitation of photosynthesis and, consequently, of growth. None of the specific nutrients were completely depleted at this time, which is also reflected in equal EPS contents in both cultivations (see Figure 3C). Only phosphate was completely depleted in the cultivation without Luffa extract after 14 days (see Figure 3D). was completely depleted. It can be assumed that higher differences in the cultivation with and without loofah extract with sufficient CO2 supply. supply can be determined.
Figure 3.
Impact of 40 g soaked Luffa cylindrica in 50 mL BG-11 on Nostoc muscorum 1453-12b compared to standard cultivation with BG-11. (A) = Cell dry weight (CDW). (B) = C-phycocyanin content. (C) = Extracellular polymeric substances (EPS). * Data are missing. (D) = Phosphate concentration over cultivation time. Cultivation parameter: 250 mL shaking flasks, 50 mL BG-11/50 mL BG11 soaked with 20 g L. cylindrica, 24 °C, cultivation time 14 days, continuous lighting at light intensity 140 µmolPhotons m−2s−1, No Day-Night-Rhythm, 120 rpm, n = 3.
Figure 3.
Impact of 40 g soaked Luffa cylindrica in 50 mL BG-11 on Nostoc muscorum 1453-12b compared to standard cultivation with BG-11. (A) = Cell dry weight (CDW). (B) = C-phycocyanin content. (C) = Extracellular polymeric substances (EPS). * Data are missing. (D) = Phosphate concentration over cultivation time. Cultivation parameter: 250 mL shaking flasks, 50 mL BG-11/50 mL BG11 soaked with 20 g L. cylindrica, 24 °C, cultivation time 14 days, continuous lighting at light intensity 140 µmolPhotons m−2s−1, No Day-Night-Rhythm, 120 rpm, n = 3.
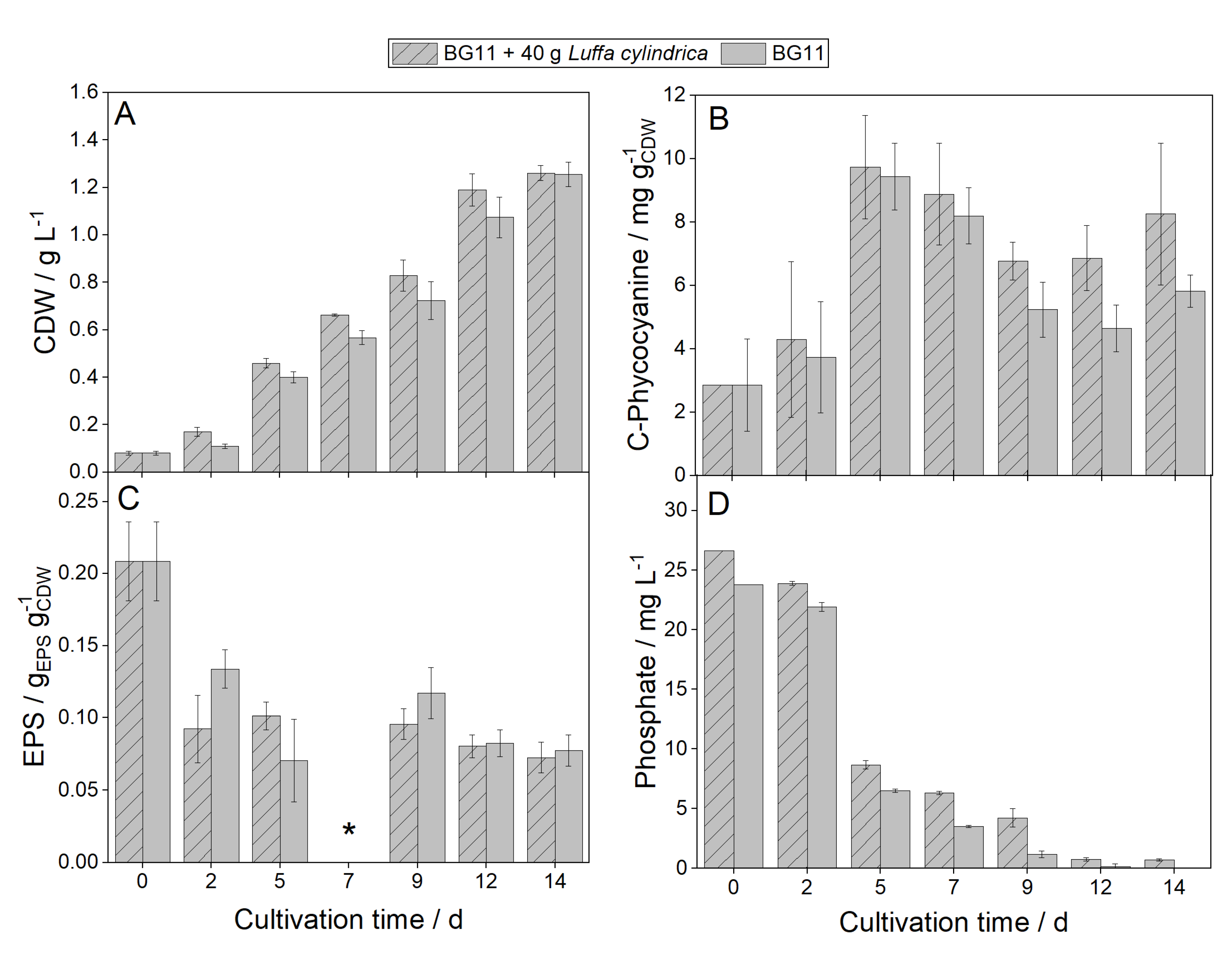
The concentration of phycoerythrin, and allophycocyanin as well as the pigments chlorophyll a, and carotenoids were identical in both cultivations. Only in the cultivation with Luffa extract higher C-phycocyanin concentrations were obtained (see Figure 3B). This is because C-phycocyanin functions, among other things, as a nitrogen storage [40]. Due to the higher availability of nitrate, more C-phycocyanin was formed, which in turn improved photosynthetic performance and thus led to higher growth rates. Therefore, nitrate concentration decreased more rapidly over the cultivation period when N. muscorum 1453-12b was cultivated with Luffa extract than without Luffa extract (data not shown). A further increase in growth by decreasing the L. cylindrica concentration from 40 to 20 g to produce the Luffa extract could not be detected.
It has been shown that the enhanced growth of cyanobacteria by the presence of L. cylindrica is due to the release of nutrients by L. cylindrica into the medium.
4. Conclusions
The goal was to find a way to use common reactor systems for the sustainable cultivation of biofilm-forming cyanobacteria. For this purpose, different sustainable and biodegradable carriers were tested. In summary, it was shown that Luffa cylindrica is ideally suited for immobilized cultivation of three different cyanobacteria (i) Anabaena cylindrica (ii) Nostoc muscorum 1453-12a, and (iii) Nostoc muscorum 1453-12b. It could be demonstrated, that the enhanced growth of cyanobacteria by the presence of L. cylindrica is, among other things, due to the release of nutrients by Luffa cylindrica into the medium. Two strains showed higher and one strain similar growth rates when cultivated on L. cylindrica compared to submerged cultivation. Additionally, the production of EPS and C-phycocyanin was increased at least 1.4-fold in all strains. In total, the natural structure (high pore volume) and properties (hydrophilic surface), as well as the growth-promoting substances released to the medium, make L. cylindrica a promising natural, biodegradable carrier for the sustainable cultivation of biofilm-forming cyanobacteria in industry. Thus, CO2-cycle can be expanded and biologically interesting products can be produced sustainably.
Author Contributions
Conceptualization, D.S. and J.S.; methodology, D.S. and J.K.; validation, D.S., J.K. and J.S; investigation, R.M. and J.K; resources, D.S.; writing—original draft preparation, D.S.; writing—review and editing, J.K. and J.S; visualization, D.S.; supervision, D.S and J.S..; project administration, D.S.; funding acquisition, D.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Carl-Zeiss Foundation and the German Research Foundation (DFG; Project number: STR 1650/1-1).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Whitton, B.A. Ecology of Cyanobacteria II: Their Diversity in Space and Time; Springer: Dordrecht, 2012; ISBN 9789400738553. [Google Scholar]
- Stanier, R.Y.; Cohen-Bazire, G. Phototrophic prokaryotes: the cyanobacteria. Annu. Rev. Microbiol. 1977, 31, 225–274. [Google Scholar] [CrossRef] [PubMed]
- Galetović, A.; Seura, F.; Gallardo, V.; Graves, R.; Cortés, J.; Valdivia, C.; Núñez, J.; Tapia, C.; Neira, I.; Sanzana, S.; et al. Use of Phycobiliproteins from Atacama Cyanobacteria as Food Colorants in a Dairy Beverage Prototype. Foods 2020, 9. [Google Scholar] [CrossRef]
- Vijayakumar, S.; Menakha, M. Pharmaceutical applications of cyanobacteria—A review. Journal of Acute Medicine 2015, 5, 15–23. [Google Scholar] [CrossRef]
- Nozzi, N.E.; Oliver, J.W.K.; Atsumi, S. Cyanobacteria as a Platform for Biofuel Production. Front. Bioeng. Biotechnol. 2013, 1, 7. [Google Scholar] [CrossRef] [PubMed]
- Lakatos, M.; Strieth, D. Terrestrial Microalgae: Novel Concepts for Biotechnology and Applications. In Progress in Botany Vol. 79; Cánovas, F.M., Lüttge, U., Matyssek, R., Eds.; Springer International Publishing; Imprint; Springer: Cham, 2018; pp. 269–312. ISBN 978-3-319-71412-7. [Google Scholar]
- Ennaceri, H.; Ishika, T.; Mkpuma, V.O.; Moheimani, N.R. Microalgal biofilms: Towards a sustainable biomass production. Algal Research 2023, 72, 103124. [Google Scholar] [CrossRef]
- Lau, N.-S.; Matsui, M.; Abdullah, A.A.-A. Cyanobacteria: Photoautotrophic Microbial Factories for the Sustainable Synthesis of Industrial Products. Biomed Res. Int. 2015, 2015, 754934. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.; Wang, J.; Chen, H.; Chen, D. Progress of marine biofouling and antifouling technologies. Chin. Sci. Bull. 2011, 56, 598–612. [Google Scholar] [CrossRef]
- Zhuang, L.-L.; Hu, H.-Y.; Wu, Y.-H.; Wang, T.; Zhang, T.-Y. A novel suspended-solid phase photobioreactor to improve biomass production and separation of microalgae. Bioresource Technology 2014, 153, 399–402. [Google Scholar] [CrossRef] [PubMed]
- Kaewprasit, C. Relationships Between a Cotton Fiber’s Specific Surface Area and Its Physical Properties. Textile Research Journal 2004, 74, 730–734. [Google Scholar] [CrossRef]
- Chetsumon, A.; Maeda, I.; Umeda, F.; Yagi, K.; Miura, Y.; Mizoguchi, T. Antibiotic production by the immobilized cyanobacterium,Scytonema sp. TISTR 8208, in a seaweed-type photobioreactor. J Appl Phycol 1994, 6, 539–543. [Google Scholar] [CrossRef]
- Chetsumon, A.; Fujieda, K.; Hirata, K.; Yagi, K.; Miura, Y. Optimization of antibiotic production by the cyanobacteriumScytonema sp. TISTR 8208 immobilized on polyurethane foam. J Appl Phycol 1993, 5, 615–622. [Google Scholar] [CrossRef]
- Walther, J.; Erdmann, N.; Stoffel, M.; Wastian, K.; Schwarz, A.; Strieth, D.; Muffler, K.; Ulber, R. Passively immobilized cyanobacteria Nostoc species BB 92.2 in a moving bed photobioreactor (MBPBR): design, cultivation and characterization. Biotechnol. Bioeng. 2022. [Google Scholar] [CrossRef]
- Rippka, R.; Herdman, M.; Waterbury, J.B. Generic Assignments, Strain Histories and Properties of Pure Cultures of Cyanobacteria. Microbiology 1979, 111, 1–61. [Google Scholar] [CrossRef]
- Strieth, D.; Stiefelmaier, J.; Wrabl, B.; Schwing, J.; Schmeckebier, A.; Di Nonno, S.; Muffler, K.; Ulber, R. A new strategy for a combined isolation of EPS and pigments from cyanobacteria. J. Appl. Phycol. 2020, 58, 419. [Google Scholar] [CrossRef]
- Bennett, A.; Bogorad, L. Complementary chromatic adaptation in a filamentous blue-green alga. J. Cell Biol. 1973, 58, 419–435. [Google Scholar] [CrossRef] [PubMed]
- Porra, R.J.; Thompson, W.A.; Kriedemann, P.E. Determination of accurate extinction coefficients and simultaneous equations for assaying chlorophylls a and b extracted with four different solvents: verification of the concentration of chlorophyll standards by atomic absorption spectroscopy. Biochimica et Biophysica Acta (BBA)—Bioenergetics 1989, 975, 384–394. [Google Scholar] [CrossRef]
- Chamovitz, D.; Sandmann, G.; Hirschberg, J. Molecular and biochemical characterization of herbicide-resistant mutants of cyanobacteria reveals that phytoene desaturation is a rate-limiting step in carotenoid biosynthesis. J. Biol. Chem. 1993, 268, 17348–17353. [Google Scholar] [CrossRef]
- Temraleeva, A.D.; Dronova, S.A.; Moskalenko, S.V.; Didovich, S.V. Modern methods for isolation, purification, and cultivation of soil cyanobacteria. Microbiology 2016, 85, 389–399. [Google Scholar] [CrossRef]
- Filippidis, A.; Moustaka-Gouni, M.; Papastergios, G.; Katsiapi, M.; Kantiranis, N. Cyanobacteria removal by Hellenic Natural Zeolite. Small Decentralized Water &.
- In-na, P.; Umar, A.A.; Wallace, A.D.; Flickinger, M.C.; Caldwell, G.S.; Lee, J.G. Loofah-based microalgae and cyanobacteria biocomposites for intensifying carbon dioxide capture. Journal of CO2 Utilization 2020, 42, 101348. [Google Scholar] [CrossRef]
- Akhtar, N.; Iqbal, J.; Iqbal, M. Removal and recovery of nickel(II) from aqueous solution by loofa sponge-immobilized biomass of Chlorella sorokiniana: characterization studies. J. Hazard. Mater. 2004, 108, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, M.; Edyvean, R.G.J. Loofa sponge immobilized fungal biosorbent: a robust system for cadmium and other dissolved metal removal from aqueous solution. Chemosphere 2005, 61, 510–518. [Google Scholar] [CrossRef] [PubMed]
- Ogbonna, J.C.; Liu, Y.-C.; Liu, Y.-K.; Tanaka, H. Loofa (Luffa cylindrica) sponge as a carrier for microbial cell immobilization. Journal of Fermentation and Bioengineering 1994, 78, 437–442. [Google Scholar] [CrossRef]
- Pringle, J.H.; Fletcher, M. Influence of substratum wettability on attachment of freshwater bacteria to solid surfaces. Appl. Environ. Microbiol. 1983, 45, 811–817. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Su, N.; Zhang, K.; Zhu, S.; Zhu, Z.; Qin, W.; Yang, Y.; Shi, Y.; Fan, S.; Wang, Z.; et al. Effect of fiber surface treatment on structure, moisture absorption and mechanical properties of luffa sponge fiber bundles. Industrial Crops and Products 2018, 123, 341–352. [Google Scholar] [CrossRef]
- Strieth, D. Produktive phototrophe Biofilme in Aerosolreaktoren, 1. Auflage; Verlag Dr. Hut: München, 2019; ISBN 978-3843939515. [Google Scholar]
- Characklis, W.G.; McFeters, G.A.; Marshall, K.C. Physiological ecology in biofilm systems; Wiley and sons: New York, 1990. [Google Scholar]
- Allaf, M.M.; Peerhossaini, H. Cyanobacteria: Model Microorganisms and Beyond. Microorganisms 2022, 10. [Google Scholar] [CrossRef]
- Ekelhof, A.; Melkonian, M. Microalgal cultivation in porous substrate bioreactor for extracellular polysaccharide production. J Appl Phycol 2017, 29, 1115–1122. [Google Scholar] [CrossRef]
- Schooling, S.R.; Beveridge, T.J. Membrane vesicles: an overlooked component of the matrices of biofilms. J. Bacteriol. 2006, 188, 5945–5957. [Google Scholar] [CrossRef] [PubMed]
- Sluiter, A.; Hames, B.; Hyman, D.; Payne, C.; Ruiz, R.; Scarlata, C.; Sluiter, J.; Templeton, D.; Wolfe, J. Determination of Total Solids in Biomass and Total Dissolved Solids in Liquid Process Samples: Laboratory Analytical Procedure (LAP). Technical Report NREL/TP-510-42621; National Renewable Energy Laboratory: Golden, CO, USA.
- Alhijazi, M.; Safaei, B.; Zeeshan, Q.; Asmael, M.; Eyvazian, A.; Qin, Z. Recent Developments in Luffa Natural Fiber Composites: Review. Sustainability 2020, 12, 7683. [Google Scholar] [CrossRef]
- Adeyanju, C.A.; Ogunniyi, S.; Ighalo, J.O.; Adeniyi, A.G.; Abdulkareem, S.A. A review on Luffa fibres and their polymer composites. J Mater Sci 2021, 56, 2797–2813. [Google Scholar] [CrossRef]
- Afreen, S.; Bano, F.; Ahmad, N.; Fatma, T. Screening and optimization of laccase from cyanobacteria with its potential in decolorization of anthraquinonic dye Remazol Brilliant Blue R. Biocatalysis and Agricultural Biotechnology 2017, 10, 403–410. [Google Scholar] [CrossRef]
- Büchs, J.; Zeols, B. Evaluation of Maximum to Specific Power Consumption Ratio in Shaking Bioreactors. J. Chem. Eng. Japan/JCEJ 2001, 34, 647–653. [Google Scholar] [CrossRef]
- Losen, M.; Frölich, B.; Pohl, M.; Büchs, J. Effect of oxygen limitation and medium composition on Escherichia coli fermentation in shake-flask cultures. Biotechnol Progress 2004, 20, 1062–1068. [Google Scholar] [CrossRef]
- Anderlei, T.; Büchs, J. Device for sterile online measurement of the oxygen transfer rate in shaking flasks. Biochemical Engineering Journal 2001, 7, 157–162. [Google Scholar] [CrossRef] [PubMed]
- Eriksen, N.T. Production of phycocyanin—a pigment with applications in biology, biotechnology, foods and medicine. Appl Microbiol Biotechnol 2008, 80, 1–14. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



