
Submitted:
08 September 2023
Posted:
11 September 2023
You are already at the latest version
Abstract
Abstract: Microflora play an important role in the fermentation of blueberry wine, influencing the flavor and nutrient formation. Commercial yeasts give blueberry wines an average flavor profile that does not highlight the specific aroma and origin of the blueberry. In the present study, ITS1-ITS2 region sequencing analysis was performed using Illumina MiSeq high-throughput technology to sequence fermented blueberry wine samples of three Vaccinium ashei varieties, Gardenblue, Powderblue, and Britewell, from the Majiang appellation in Guizhou province to analyze the trends of fungal communities and the diversity of compositional structures in different periods of blueberry wine fermentation. The study's results revealed that 114 genera from 7 phyla were detected in 9 samples from different fermentation periods of blueberry wine. The main fungal phyla were Ascomycota, Basidiomycota, Kickxellomycota, Chytridiomycota, and Olpidiomycota. The main fungal genera were Hanseniaspora, Saccharomyces, unidentified, Aureobasidium, Penicillium, Mortierella, Colletotrichum, etc. Hanseniaspora was dominant in the pre-fermentation stage of blueberry wine, accounting for more than 82%; Saccharomyces was the dominant genera in the middle and late fermentation stages of blueberry wine, with Saccharomyces accounting for more than 72% in the middle of fermentation and 93% in the late fermentation stage. This study screened indigenous flora for the natural fermentation of blueberry wine in the Majiang production area of Guizhou, improved the flavor substances of the blueberry wine, highlighted the characteristics of the production area, and made the blueberry wine have the characteristic flavor of the production area.
Keywords:
Majiang region
; dry red blueberry wine
; fungal community
; fungal dynamic analysis
; Illumina MiSeq high-throughput technology
1. Introduction
Microorganisms are the richest resource of biodiversity on Earth, and exploring microbial diversity is important for the study of microorganisms and human diseases[1]. The blueberry wine fermentation system consists of different species, quantities, and qualities of yeasts, bacteria, and molds, and is a complex and dynamic microbial ecosystem that is constantly evolving and changing[2]. Among them, the growth, metabolism, and apoptosis activities of yeast and lactic acid bacteria convert the sugars in blueberry into alcohol, so it is of great theoretical and practical significance to explore the fungal community characteristics and succession law in the fermentation process of blueberry wine for the optimization of the wine fermentation process. The color, taste, and aroma of blueberry wine are inseparable from the interaction of microorganisms in the parameters of fermentation. Blueberry wine currently has two types of fermentation: natural fermentation and commercial yeast-inoculated fermentation. Commercial yeast inoculation fermentation is used to control the fermentation by adding commercial yeast that has been maturely used in the winemaking industry. This method allows for stable brewing during the fermentation of blueberry wine, but it also produces a single flavor of blueberry wine[3]. The natural fermentation method of blueberry wine can have the characteristics and style of the original production area because the natural fermentation process retains the microflora of the blueberry origin[4].
In recent years, high-throughput sequencing technology has been widely used in food research due to its advantages[5]. The existence of complex and variable micro-ecosystems during the fermentation of blueberry mash, which reflects the fermentation status of blueberry mash in real time, plays a crucial role in the flavor and quality of blueberry wine. Most of the studies were mainly on the composition structure of bacterial colonies and fungal colonies on the peel of different blueberry varieties, but there were few studies on the trend and structural variability of fungal colonies in different fermentation stages of different blueberry varieties, and few types of research have been reported for the fermentation process of different blueberry varieties in Majiang appellation in Guizhou.
Therefore, in this study, we used ITS-ITS2 high-throughput sequencing technology to dynamically analyze the fungal communities of different blueberry varieties at different fermentation periods to assess the composition and abundance of fungal communities and provide a basis for further research on the structural changes of blueberry wine flora and its role in the fermentation process in the Majiang region of Guizhou.
2. Materials and Methods
2.1. Experimental materials
The Vaccinium ashei used in this study came from the ecological blueberry industrial park in Ukapeng of Majiang County Guanghua Agriculture Limited. Ukapeng Ecological Blueberry Industrial Park is centered on the Blue Dream Valley Scenic Area in Republican Village, Longshan Town. The base was built in 1998 and mostly planted with Britewell, Baldwin-T-117, Powderblue, Bluerain, O’Neal, Gardenblue, and other blueberry varieties. The blueberry base is based on blueberry cultivation land and relies on different contractors to cultivate blueberry plants. The area of Uka village is about 9.73 square kilometers, with 818 mu of arable land, except for a few flat lands for rice, corn, and chrysanthemum planting, and a large area of blueberry is neatly planted on the terraces on the hills and mountains with the slope ranging from 6 ° to 20 °, and the horizontal terraces are more than 2 m wide. The peak growth of Ukapeng eco-blueberries is usually in May and June[6]. The heavy metal content in the soil profile of the Majiang production area in Guizhou exceeded the standard, but mainly at the bottom of the contour, the growth of blueberries was not significantly affected[7]. Pest control in new blueberry growing areas is mainly applied at safe intervals during the rest period[8].
The blueberries were randomly harvested at the ripening stage from three varieties of blueberries, namely Gardenblue, Powderblue, and Britewell, hand-harvested from the plants using sterile gloves, the blueberries were preserved in sterile vacuum bags and transferred to the laboratory under refrigeration for blueberry winemaking.
2.2. Experimental methods
2.2.1. The fermentation process of blueberry wine
Hu et al. studied the fermentation process of blueberry wine[9]. Fresh blueberry berries were fermented with sterile gloves and hand crushed and pressed. Fermentation of the three blueberry varieties was monitored daily using a densitometer, and 50 mL fermentation broth samples were collected at three different fermentation stages (pre-fermentation, mid-fermentation and, post-fermentation): pre-fermentation (day 2), mid-fermentation (day 4) and post-fermentation (day 6). Thirty mL of blueberry ferment broth samples were drawn into sterile centrifuge tubes under aseptic conditions and stored in an ultra-low temperature refrigerator (-80°C) for sequencing of the fungi. Another 1 mL of blueberry fermentation broth samples were aspirated into 99 mL of sterile saline under aseptic conditions and homogenized for 30 min for media experiments[10].
2.2.2. Extraction of genetic DNA from blueberry wine samples
Extraction of genetic DNA is performed using the MOBIO PowerSoil® DNA Isolation Kit in the following steps. The main steps include Placing 0.25 ml of blueberry wine sample in a 750 μl tube, mixing gently with good offices, adding 60 μl of Reagent 1, mixing upside down, and placing the tube on a vortexer adapter at 3200 RMP for 10 min. Transfer the supernatant to a clean 2 ml collection tube, add reagent ② and reagent ③, and continue to vortex and incubate. Add reagent ④ to allow the DNA to adsorb onto the membrane, and then add reagent ⑤ to elute. Finally, the extracted plasmid DNA was spiked onto a 1% agarose gel, and nucleic acid electrophoresis was performed after sampling to obtain the extracted DNA[11].
2.2.3. PCR amplification and MiSeq sequencing of DNA extracted from blueberry wine fermentation broth samples
Primer sequences for fungal ITS1-ITS2 region: (5’-CTTGGTCATTTAGAGGAAGTAAA-3’) and (3’-TGCGTTCTTCATCGATGC-5’, 5 μmol/L) 1 μL each, BSA (2 ng/μL) 3 μL, 2×Taq Plus Master Mix 12.5 μL, and ddH2O 7.5 μL. The amplification process is as follows: 94 ° C preproduction for 5 minutes, decomposed for 94 seconds at 94 ° C., annealed at 55 ° C for 30 seconds, extended at 72 ° C for 1 minute, 28 cycles, and finally extended at 72 ° C for 10 minutes, and each sample was repeated three times. Detection of PCR amplification products by nucleic acid electrophoresis using 1% agarose gel for amplified target band size, and purified by Agencourt AMPure XP Nucleic Acid Purification Kit was used for purification. Subsequent library construction and sequencing were entrusted to the Nanjing branch of Beijing Ovison Biotechnology Co.
2.3. Data processing
Miseq sequencing is obtained from Pair-End (PE) double-end sequence data, using Trimmomatic and Pear to quality control the measured Fastq data to get Fasta sequence; then using Flash and Pear to splice (merge) the two-end sequence according to the overlapping relationship of PE, and finally to get raw-tags; remove the chimeras of Fasta sequences by chime method according to the known database, and remove them by self-comparison (denovo) method for the unknown database, and remove the short sequences that do not meet the requirements to get high-quality sequences clean-tags[12].
The clean tags were clustered to generate OTUs[13]. The RDP Classifier algorithm was used to compare and analyze the OTU representative sequences and annotate the species information of their communities at the phylum, genus, and species level (phylum, genus, species) to obtain the species taxonomic information corresponding to each OTU. Sequence alignment was performed using mafft, fast tree for tree building, and species (genus) evolutionary relationships were visualized and displayed according to the Python language. PCA statistical analysis and graphing were performed using R language.
3. Results
3.1. Sequence data and OTUs analysis
After sequencing samples from different fermentation periods of Gardenblue, Powderblue, and Britewell blueberry wines, the quality sequence distribution statistics obtained are shown in Table 1. A total of 482852 valid sequences were collected from 9 blueberry fermentation broth samples, among which the sequence lengths in the range of 420-440 accounted for the majority of the valid sequences. This experimental result indicates that the detection results are excellent and can be applied to the later experimental analysis. The dilatability curves allow the sequencing depth of the samples to be derived[14]. From the dilution curves of the fungal ITS1-ITS2 regions of different varieties of blueberries before, during, and after fermentation under the 97% similarity condition shown in Figure 1, it can be seen that when the number of fungal OTUs reached about 70, the dilution curves of the samples of pre-fermentation of Gardenblue and late fermentation of Britewell had begun to level off, and the sampling depth was sufficient, and the sequencing number could cover all taxa. The remaining 7 samples also had sufficient sequencing depth when the number of fungal OTUs reached about 100, and the taxa were covered completely.
The OTU-Venn diagram (Figure 2) labels the number of OTUs in the three varieties of blueberry wine samples, with the main objects being those that are conventional in blueberry wine samples and those that are specific to blueberry wine samples. As can be seen in Figure 2, the samples of the three blueberry varieties have 128 OTUs in common, of which only Gardenblue has 14, only Powderblue has 24, and 38 are specific to Britewell. This shows that the fungal communities of the three blueberry varieties have overlapping parts, and also have their own varieties of unique fungal communities. To better represent the OTU distribution of blueberry wine samples, a petal chart of OTU distribution (Figure 3) shows the number of endemic and overlapping OTUs in the full sample of blueberry wine. As can be seen in Figure 3, the total number of OTUs for all blueberry wine samples was 32, indicating the presence of 32 fungal communities that survived the entire fermentation process of blueberry wine.
From Figure 3, the total number of OTUs in the nine samples of Gardenblue, Powderblue, and Britewell before, during, and after fermentation was 32. 133, 159, and 177 for Gardenblue, 79, 132, and 141 for Powderblue, and 135, 186, and 109 for Britewell before, during, and after fermentation, respectively. This shows that there are both similarities and differences in the bacterial flora of different varieties of blueberries in the same appellation during fermentation.
3.2. Analysis and discussion of alpha diversity data of fungal community zones during the fermentation of blueberry wine
Alpha diversity index table for fungal detection[15], The alpha diversity index table contains six main indices, namely: chao1 index, coverage index, observed_species, PD_whole_tree, Shannon and Simpson indices, by which the experimental results can reflect the richness and diversity of samples of three different blueberry wines at different fermentation stages in the Majiang appellation of Guizhou. chao1 and observed_species indices are mainly to estimate the number of species contained in the fungal community in blueberry wine samples. Also, these two indices can represent the OUT estimate and the actual value of the sample community respectively, which can reflect the abundance of fungal communities at each stage of blueberry wine fermentation. The coverage is a test of microbial coverage in blueberry wine, and the higher the value, the lower the probability that new species are not detected in the samples of different fermentation stages of blueberry wine. The coverage index of all blueberry wine samples was 1.00, indicating that the coverage of blueberry wine samples was 100%, indicating the actual situation of true actual value bacteria in samples of different varieties of blueberry wine at different fermentation stages and reflecting the reliability of the experimental data. As can be seen from Table 2, Chao1 reflects the richness and diversity of fungal species in the blueberry wine samples, which is the number of species in the sample, and the more species in the sample, the richer the sample. Species richness does not consider the relative abundance of each species, it gives the same weight to species with low relative abundance as those with high relative abundance. As shown in Table 2: The largest fungal richness index Chao1 for different varieties of blueberries before, during, and after fermentation was for Britewell in the middle of fermentation, and the smallest Chao1 index was for Gardenblue in the pre-fermentation period, indicating that Britewell in the middle of fermentation was the most abundant species among these nine samples, and the abundance of Gardenblue reached the minimum value in the pre-fermentation period. Among them, the PD_whole_tree index responded to the changes of fungal communities in the samples during the fermentation of blueberry wine and the diversity, reflecting the trend and variation of fungi in blueberry wine samples. Uniformity reflects the relative abundance of different species. Diversity increases with increasing species richness and evenness. The diversity indices, Shannon and Simpson, are a composite indicator of species richness and evenness in a sample higher Shannon and Simpson values indicate higher diversity of community species[16-18]. From Table 2, it can be seen that Powderblue had the highest fungal community diversity among different varieties in the same fermentation stage, followed by Britewell and the lowest Gardenblue. Among different fermentation stages of the same variety, all had the highest fungal community diversity in the pre-fermentation stage, followed by the middle stage, and the lowest in the late fermentation stage.
3.3. Alpha diversity analysis of fungal flora during wine fermentation
Based on the differentiation of ITS1-ITS2 regions for fungal species, representative sequences of OTUs corresponding to the largest abundance were selected for tree building by genus using mafft and fast tree methods. From Figure 4, it can be seen that there are mainly 114 genera belonging to 7 phyla in the dry red blueberry wine-making habitat, namely: Ascomycota, Kickxellomycota, Chytridiomycota, Olpidiomycota, Basidiomycota, Mortierellomycota, and Glomeromycota, of which 83 genera belong to Ascomycetes, 24 to Basidiomycota, 2 to Glomeromycota, 2 to Chytridiomycota, 1 to Kickxellomycota, 1 to Olpidiomycota, and One genus belongs to the Mortierellomycota. This indicates that the fungal diversity in the dry red blueberry wine-making habitat is very rich, and the dry red blueberry wine-making is the result of the joint action of many microorganisms, while the Ascomycetes is the dominant phylum.
3.4. Analysis of fungal species and abundance
Comparison of fungal communities in pre, mid, and late- fermentation fungal samples from different varieties of blueberries using visual bar graphs[19] revealed the presence of major phyla and genera in all samples, but with differences in abundance (Figure 5). From the figure, it can be seen that at the level of the fungus phylum, Ascomycetes are present throughout the fermentation process of blueberry wine, with the relative abundance pairs of Ascomycetes in the pre-fermentation fungal communities of Gardenblue, Powderblue, and Britewell being 92.35 %, 93.67 % and 92.92 %, respectively, and still increasing as fermentation progressed, and at the late fermentation stages of Gardenblue, Powderblue, and Britewell being 98.85 %, 99.06 %, 99.36 %, the percentage of Ascomycetes in the blueberry wine samples of all three blueberry varieties showed an increasing trend as fermentation proceeded; followed by the unidentified phylum; followed by the Basidiomycota, the relative abundance of the Basidiomycota in the fungal communities of Gardenblue, Powderblue and Britewell at the pre-fermentation stage were 0.28 %, 0.28 % and 0.69 %, respectively, and gradually decreased as fermentation proceeded, with the abundance of at the late stage of fermentation, only 0.22 %, 0.25 % and 0.11 % were found in the fungal communities of Gardenblue, Powderblue and Britewell, respectively.
At the genus level, the fungi in the pre-and mid-fermentation samples of Gardenblue, Powderblue, and Britewell were distributed in seven genera: Hanseniaspora, Saccharomyces, unidentified, Aureobasidium, Penicillium, Mortierella, and Colletotrichum. The relative abundance of Hanseniaspora was the highest in the pre-fermentation samples of Gardenblue, Powderblue, and Britewell, with 84.72 %, 80.26 %, and 81.50 %, respectively; followed by Saccharomyces with 10.23 %, 4.97 %, and 3.43 %, respectively; the abundance of the remaining genera was very small. The relative abundance of Saccharomyces was the highest in the mid-fermentation Gardenblue, Powderblue, and Britewell samples, with 90.40 %, 63.84 %, and 62.92 %, respectively, followed by Hanseniaspora with 2.62 %, 33.70 %, and 33.87 %, respectively, while the abundance of the remaining genera was very low. The relative abundance of the Saccharomyces was the highest in the late fermentation samples of Gardenblue, Powderblue, and Britewell with 96.18 %, 92.40 %, and 90.62 %, respectively, followed by Hanseniaspora with 1.22 %, 3.55 %, and 7.87 %, respectively, while the abundance of the remaining genera was minimal. From the relative abundance of Saccharomyces on the fourth day, it is clear that the fermentation rate of Gardenblue was faster than that of Powderblue and Britewell, and the Saccharomyces were dominant by the fourth day.
On the whole, the genus Saccharomyces began to exist at the beginning of fermentation, increased rapidly in number in the middle of fermentation, and became absolutely dominant in the middle and late stages of fermentation, while the genus Hanseniaspora was present in large numbers at the beginning of fermentation and became absolutely dominant, and its relative abundance decreased rapidly in the middle stage as fermentation progressed and was lower in the late stage. Thus, it can be seen that in the fermentation process of Majiang appellation Gardenblue, Powderblue, and Britewell blueberry wines, the Hanseniaspora genus was the first to develop in the early stage and gradually gave way to Saccharomyces in the middle and late stages.
Nine samples from different fermentation periods of three varieties of blueberry wine were analyzed using the spearman test, and the 20 fungal genera with the highest absolute abundance in the blueberry wine samples were extracted for correlation association analysis, while the analysis results were plotted against the fungal phylum as a legend. The data with p-value > 0.05 or correlation value |R|< 0.4 were plotted to obtain Figure 6. It can be seen from the figure that the association status between Hanseniaspora and Saccharomyces during the fermentation of blueberry wine was negatively correlated. The correlation between Aureobasidium and Hanseniaspora was positive during the fermentation of blueberry wine.
3.5. Dynamic changes of fungal flora
The top 20 genera in terms of relative abundance values were selected and clustered according to their abundance information in samples at different fermentation stages, and the fungal thermogram was used to visually analyze the distribution of fungal communities of the three blueberry varieties Gardenblue, Powderblue, and Britewell at different fermentation stages and different blueberry varieties at the same fermentation stage during the fermentation of blueberry wine. The relative abundance of Saccharomyces, Hanseniaspora, etc, the dominant fungal community was present in all fermentation stages of the dry red blueberry wine, and its fungal flora showed similar trends in the samples, as can be seen in Figure 7. For example, the abundance of Hanseniaspora, Aureobasidium, and Penicillium in the pre-fermentation samples, Gardenblue was similar to Britewell and higher than in the mid and late-fermentation samples.
From Table 3, it can be seen that the three groups of bacteria involved in the fermentation process of blueberry wine such as H. uvarum, S. cerevisiae, A. pullulans, M. elongata, etc. Saccharomyces cerevisiae was the most commonly used organism species in fermentation and dominated in the pre-fermentation stage in this experiment. The main product of wild-type S. cerevisiae is ethanol, and traditional S. cerevisiae strains have good fermentation performance, are well tolerated under anaerobic conditions, and can ferment sugars to produce high ethanol yields and production rates[20].S. cerevisiae has a short growth cycle, efficient sugar conversion capacity, competes for limited nutrient capacity, is easily S. cerevisiae, as a dominant strain in fermentation, is rich in nutrients such as proteins, amino acids, vitamins, and bioactive substances, and promotes the production of volatile aroma components such as esters, alcohols, terpenoids, and volatile thiol substances using its metabolic activities[21, 22]. S. cerevisiae has synergistic effects with other yeasts to improve wine quality and shape-rich aromas together with other yeasts[23]. As can be seen from Table 3, the fungal community of blueberries in the process of fermentation into blueberry wine is constantly changing, with S.cerevisiae becoming the dominant flora, accounting for more than 90%.
3.6. Analysis of fungal OTUs of different blueberry varieties before, during and after fermentation
The 32 shared OTUs were analyzed for abundance, resulting in the top 20 OTUs in terms of abundance, as shown in Table 3, namely: Saccharomyces cerevisiae, Hanseniaspora uvarum, Hanseniaspora osmophila, Hanseniaspora vineae, Pseudaleuria sp, Aureobasidium pullulans, Mortierella alpina, Mortierella elongata, Stachybotrys microspora, and Guehomyces pullulans.
Saccharomyces cerevisiae is a traditional ethanol-producing strain with good fermentation performance, good growth capacity under anaerobic conditions, the ability to ferment glucose to produce high ethanol yields and production rates, and high tolerance to ethanol and other inhibitors[24-26]. S. cerevisiae, as the dominant flora in mid and late fermentation, also promotes the production of volatile aroma substances such as alcohols, esters, terpenoids, and volatile thiols through its own metabolic activities and the release of non-aromatic precursors[27]. Hanseniaspora uvarum is the most abundant non-enological yeast commonly found in vineyards and winemaking environments[28]. The glycosidases secreted by H. uvarum mainly promote the hydrolysis of desmethyl isoprenoids and some terpene glycosides, and less so for C6 compound glycosides, thereby increasing the temperate fruit and floral characteristics of the wine[29]. Hanseniaspora osmophila and others can ferment higher levels of acetate compounds such as phenyl acetate and isopentyl acetate[30]. Zhang et al[31] found that Hanseniaspora vineae and Saccharomyces cerevisiae mixed fermentation had a synergistic effect on the production of ethyl 2-phenylacetate. ethyl 2-phenylacetate has a peachy aroma and is present in apples, grapes, poplar berries, tea, beer, whiskey, and brandy, and is a desired aroma compound in wine. In this experiment, Hanseniaspora had a high relative abundance (around 80%) in the pre-fermentation period; Saccharomyces dominated in the middle and late stages, with relative abundance up to 96%, and they acted synergistically to play an important role in improving blueberry wine quality and aroma shaping, etc.
Aureobasidium pullulans, a class of yeast-like fungi[32], is the predominant pectinolytic species (pectinolytic strain) in the grape skin flora and is present in the must and pre-fermentation. Although low-temperature maceration for wine production enhances the aroma of the wine, the color and phenolics of the wine are affected. Marin et al[33] applied A. pullulans to cold maceration and low-temperature fermentation before fermentation and found a significant increase in total anthocyanin content, total polyphenol index and color intensity (CI), and a higher content of polymer pigments at 9 months of storage in the bottle. This indicates that A. pullulans affects the color and stability of the wine and plays an important role in the quality of the wine. In this experiment, the relative abundance of A. pullulans was detected at 1.11 %, 0.24 %, and 0.19 % before and during the middle of the fermentation of Gardenblue, 0.25 %, 0.01 %, and 0.001 % during the three fermentation periods of Powderblue, and 1.30 %, 0.15 %, and 0.07 % during the three fermentation periods of Britewell, respectively. The relative abundance of A. pullulans decreased gradually during the fermentation of different blueberry wine varieties and was the lowest in the Powderblue samples.
Microbial cells usually contain only 2 % to 3 % of oil, but under specific culture conditions, certain microbial dry bodies can contain up to 80-90% oil[34]. Some molds have a strong lipid production capacity, and the lipids produced also contain varying degrees of γ-linoleic acid (GLA), arachidonic acid, and other beneficial components for humans, so scholars at home and abroad have conducted in-depth studies on them. Mortierella alpina has a strong lipid synthesis capacity, and its lipid accumulation can reach more than 50 % of its dry weight, where the production of Arachidonic acid can exceed 40% of the total fatty acid content[35], which has been applied to commercial production and has good application value and development prospects. strains such as Mortierella elongata and Mortierella alpina contain γ-linoleic acid (GLA), and the process of GLA production by Peridiomycetes is mature and can produce annually several hundred tons per year[36]. It was found that M. elongata, M. alpina, and other mycobacteria also contain EPA and DHA[37]. Although these two species were detected in this experiment, their relative abundance was small. Among the three species, M. alpina was the most abundant in Britewell, with abundances of 0.01 %, 0.04 %, and 0.02 % before, during, and after fermentation, respectively. The abundance of M. elongata in Gardenblue was 0.03 %, 0.09 %, and 0.07 % before, during, and after fermentation, respectively; the relative abundance in Powderblue was 0.10 %, 0.14 %, and 0.03 % during the three fermentation periods, respectively; and 0.17 %, 0.04 % and 0.09 % during the three fermentation periods in Britewell, respectively. The relative abundance of M. elongata was greatest during the pre-fermentation period of Britewell and decreased significantly in the middle and late stages, while it increased in the middle of the mash of Powderblue blueberries, and the specific factors influencing this need to be studied in depth. It is worth acknowledging that both bacteria have a role in enhancing the nutritional value of blueberry wine.
Stachybotrys microspora is a filamentous fungus with cellulolytic activity. The basic process of cellulose hydrolysis is the production of cellobiose and some cellooligosaccharides by the synergistic action of endoglucanase and exoglucanase, which in turn are hydrolyzed by β-glucosidase to produce glucose[38]. By studying the wild-type strain N1 of S. microspora and its mutant A19, Abdeljalil et al[39]found that Stachybotrys produced much higher yields of glucanase, β-glucanase, and protease than the current commercial lyase "zymolyase", in addition to endoglucanase and exoglucanase. In this experiment, the abundance of S. microspora was small, accounting for 0.0027 %, 0.027 %, and 0.045 % in the three stages of fermentation for Gardenblue, 0.03 %, 0.05 %, and 0.0053 % in the three stages of fermentation for Powderblue, and 0.0053 %, 0.04 %, and 0.01 % in the three stages of fermentation for Britewell. As a whole, the abundance of this bacterium increased in the middle of fermentation. From previous studies, this bacterium has some roles in blueberry wine such as hydrolysis of cellulose to produce glucose and promotion of blueberry juice maceration to stabilize the color.
Guehomyces pullulans were found to produce pectinase even at low temperatures (8°C)[40]. Pectinase has a very important role in juice yield, clarification, and promotion of nutrient leaching in grape juice[41-43]. Zhang et al[44] found G. pullulans to be the dominant bacterium in traditional Korean rice wine distillers. This bacterium was also detected in nine samples of this experiment, but the relative abundance was low and the relative abundance did not change significantly at different fermentation stages, accounting for only about 0.01 %.
3.7. Beta diversity analysis of fungal flora during blueberry wine fermentation
As shown in Figure 8, PC1 and PC2 explained 88.94% and 5.52% of the information about the fungal communities of the samples of different species before and during the middle of the fermentation period, respectively, and the sum of both was greater than 80%, and the results were reliable. There were some distances between the nine samples, indicating that the fungal communities of the nine samples had some differences; in comparison, the three samples of Gardenblue were closer and the differences were not significant, indicating that the fungal communities of Gardenblue samples in different fermentation periods were highly similar; the distance between Powderblue samples in different fermentation periods was larger and the differences were significant, indicating that the fermentation the distance between Gardenblue samples of different fermentation periods was large and the difference was significant, indicating that the fungal communities of Gardenblue samples of different fermentation periods were very different from those of Powderblue samples of pre-middle and late fermentation. Overall, the similarity of fungal communities between Gardenblue samples in different fermentation periods was high, and the difference of fungal communities between Powderblue and Britewell samples was high.
The species clustering tree diagram of the different sample fungal communities is represented by the taxonomic levels from phylum to genus by circles radiating from the inside to the outside. The circles in Figure 9 represent a classification at different levels, and the circle size increases as the relative abundance of species under that classification increases. The circle colors represent the differences between species at different levels, yellow are species with no significant differences, red indicates a class of microbial taxa that can be clearly distinguished in their group, while green indicates microbial taxa that can play an important role in the corresponding group, and the other circle colors have the same meaning [45]. As shown in Figure 9, the color of the fungal community species clustering tree of different samples was mostly yellow, indicating that the fungal species in the fermentation process of Gardenblue, Powderblue, and Britewell were mostly the same two fungal communities different from Powderblue and Britewell were present in Gardenblue. The results of Biomarker follow-group coloring showed that the main fermentation process was stable in Gardenblue, Powderblue, and Britewell samples, and no other different species dominated or disrupted the fermentation process with yeast as the dominant species.
4. Conclusions
The fungal community structure of the fermentation process of three blueberry varieties in Majiang appellation was studied by high throughput sequencing technology, which revealed that Hanseniaspora was the dominant group in the pre-fermentation mash and Saccharomyces was predominant in the middle and late stages of fermentation. Saccharomyces cerevisiae, Hanseniaspora uvarum, Hanseniaspora osmophila, Hanseniaspora vineae, Pseudaleuria sp, Aureobasidium pullulans. Mortierella alpina, Mortierella elongata, Stachybotrys microspora, Guehomyces pullulans relative to Hanseniaspora and Saccharomyces, the relative abundance of several other bacteria was small, accounting for only about 0.001 % to 1.300 %, respectively; The fungal composition of different fermentation stages of the same species differed significantly, except for the Gardenblue samples.
Among the three blueberry wines fermented under this colony structure, Powderblue was better than Gardenblue and Britewell in terms of overall sensory quality, especially in terms of persistence and fullness on the palate, good structure and complexity, and strong tannin, which is related to its own varietal characteristics, but also to the richness of the fungal community during fermentation and the significant differences in the community composition at different stages of fermentation. The Gardenblue dry red blueberry wine has good fullness and pleasant fruit aroma, which is significant during the fermentation process, but it is a little bit bland in terms of taste, whether it is related to the small difference in fungal composition in different fermentation stages needs to be further verified. In addition to the high sugar content of the fruits, Saccharomyces cerevisiae, which was dominant in the middle and late samples, also played an important role. In conclusion, the results of this experiment can provide a theoretical basis for future research on community structure and its influence on blueberry wine style shaping in Majiang region, Guizhou.
Author Contributions
Conceptualization, J.S., and B.H.; methodology, J.S.; software, J.S.; validation, J.S., and M.Z.; formal analysis, S.X.; investigation, J.S.; resources, M.Z.; data curation, S.X.; writing—original draft preparation, J.S.; writing—review and editing, S.X. and B.H.; visualization, J.S.; supervision, M.Z.; project administration, B.H.; funding acquisition, B.H. All authors have read and agreed to the published version of the manuscript.
Funding
“Qian southeast science and cooperation support [2023] No. 13”.
Data Availability Statement
The data sets generated during and/or analyzed during the current study are available with the corresponding author on reasonable request.
Acknowledgments
We acknowledge financial support by the National Natural Science Foundation of China. (Project No. 31271857).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Eldarov, M.A.; Kishkovskaia, S.A.; Tanaschuk, T.N.; Mardanov, A.V. Genomics and biochemistry of Saccharomyces cerevisiae wine yeast strains. Biochemistry 2016, 81, 1650–1668. [Google Scholar] [CrossRef] [PubMed]
- Caridi, A.; Sidari, R.; Pulvirenti, A.; Blaiotta, G. Genetic Improvement of wine yeasts for opposite adsorption activity of phenolics and ochratoxin A during red winemaking. Food Biotechnol. 2020, 34, 352–370. [Google Scholar] [CrossRef]
- A, M. A.; B, A. P.; B, S. J.; C, G. B.; A, M. B.; A, J. C. , Influence of winemaking techniques on the resveratrol content, total phenolic content and antioxidant potential of red wines. Food Chemistry 2012, 131(2), 513–518. [Google Scholar]
- Bagheri, B.; Bauer, F.F.; Setati, M.E. The Impact of Saccharomyces cerevisiae on a Wine Yeast Consortium in Natural and Inoculated Fermentations. Front. Microbiol. 2017, 8, 1988. [Google Scholar] [CrossRef] [PubMed]
- Reuter, J.; Spacek, D. V.; Snyder, M. , High-Throughput Sequencing Technologies. Molecular Cell 2015, 58(4), 586–597. [Google Scholar] [CrossRef] [PubMed]
- Xu Hong, Planning and design study of special blueberry industry tourism park in Majiang County, Guizhou. China Agri-cultural Resources and Zoning 2016, 37, 204–207.
- Li, Q.; Li, C.; Wang, H.; Wei, X.; Liu, Y.; Yang, R.; Wen, X. Geochemical Characteristics of Heavy Metals in Soil and Blueberries of the Core Majiang Blueberry Production Area. Bull. Environ. Contam. Toxicol. 2021, 106, 57–64. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, F.; Wang, B.; Qiu, D. Identification, Characterization and Expression Analysis of Anthocyanin Biosynthesis-related bHLH Genes in Blueberry (Vaccinium corymbosum L.). 2021.
- Hu, B.; Gao, J.; Xu, S.; Zhu, J.; Fan, X.; Zhou, X. Quality evaluation of different varieties of dry red wine based on nuclear magnetic resonance metabolomics. Appl. Biol. Chem. 2020, 63, 1–8. [Google Scholar] [CrossRef]
- Hu, B.; Zhou, M.; Su, J.; Lin, L.; Xu, S. Study of Fungal Communities in Dry Red Wine Fermentation in Linfen Appellation, Shanxi. Fermentation 2022, 8, 475. [Google Scholar] [CrossRef]
- Walch, G.; Knapp, M.; Rainer, G.; Peintner, U. Colony-PCR Is a Rapid Method for DNA Amplification of Hyphomycetes. J. Fungi 2016, 2, 12. [Google Scholar] [CrossRef]
- Ghosh, S.; Namin, S.M.; Jung, C. Differential Bacterial Community of Bee Bread and Bee Pollen Revealed by 16s rRNA High-Throughput Sequencing. Insects 2022, 13, 863. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Li, Y.; Wang, Y.; Luo, X.; Du, F.; Liu, W.; Xie, L.; Chen, J.; Ren, Z.; Hou, S.; et al. The microbial diversity in industrial effluents makes high-throughput sequencing-based source tracking of the effluents possible. Environ. Res. 2022, 212, 113640. [Google Scholar] [CrossRef] [PubMed]
- Amato, K.R.; Yeoman, C.J.; Kent, A.; Righini, N.; Carbonero, F.; Estrada, A.; Gaskins, H.R.; Stumpf, R.M.; Yildirim, S.; Torralba, M.; et al. Habitat degradation impacts black howler monkey (Alouatta pigra) gastrointestinal microbiomes. ISME J. 2013, 7, 1344–1353. [Google Scholar] [CrossRef] [PubMed]
- Gail, M. H.; Yunhu, W.; Jianxin, S. , Power of Microbiome Beta-Diversity Analyses Based on Standard Reference Samples. American Journal of Epidemiology.
- Martínez-Salgado, S.; Romero-Arenas, O.; Morales-Mora, L. A.; Luna-Cruz, A.; Hoyos, P. A. , First Report of Macrophomina phaseolina Causing Charcoal Rot of Peanut (Arachis hypogaea L.) in Mexico. Plant Disease 2021.
- Zhao, Y.; Sun, Y.; Chen, W.; Zhao, Y.; Bai, Y. , remote sensing The Potential of Mapping Grassland Plant Diversity with the Links among Spectral Diversity, Functional Trait Diversity, and Species Diversity. Remote Sensing 2021, 13. [Google Scholar]
- Ken; Kelley; Francis; Bilson; Darku; Bhargab; Chattopadhyay, Sequential accuracy in parameter estimation for population correlation coefficients. Psychological Methods 2019.
- Oberauner, L.; Zachow, C.; Lackner, S.; Högenauer, C.; Smolle, K.-H.; Berg, G. The ignored diversity: complex bacterial communities in intensive care units revealed by 16S pyrosequencing. Sci. Rep. 2013, 3, srep01413. [Google Scholar] [CrossRef] [PubMed]
- Ameni, G.; Kebede, A.M.; Zewude, A.; Abdulla, M.G.; Asfaw, R.; Gobena, M.M.; Kyalo, M.; Stomeo, F.; Gumi, B.; Sori, T. Equine Histoplasmosis in Ethiopia: Phylogenetic Analysis by Sequencing of the Internal Transcribed Spacer Region of rRNA Genes. Front. Cell. Infect. Microbiol. 2022, 12, 789157. [Google Scholar] [CrossRef] [PubMed]
- Vidyasagar-E-mail, C.; Kumar–E-mail, P. S.; Vijayakumar–E-mail, P.; Alekya–E-mail, S.; Umamahesh, K.; Reddy, O. V. S. , Comparative structural and functional analysis of the PGU1 protein from Saccharomyces bayanus with other Saccharomyces species. Bioinformation 2022, 18(5), 464–469. [Google Scholar] [CrossRef] [PubMed]
- Manuel, S. L. J.; Sylwia, J.; Aslihan, E. K.; Jungho, L.; Roger, S.; Vanbogaert, I. N. A. , The oleaginous yeast Starmerella bombicola reveals limitations of Saccharomyces cerevisiae as a model for fatty acid transport studies. FEMS Yeast Research (1), 1.
- Santiago-Urbina, J.A.; Arias-García, J.A.; Ruiz-Terán, F. Yeast species associated with spontaneous fermentation of taberna, a traditional palm wine from the southeast of Mexico. Ann. Microbiol. 2015, 65, 287–296. [Google Scholar] [CrossRef]
- Hu, B.; Cao, Y.; Zhu, J.; Xu, W.; Wu, W. Analysis of metabolites in chardonnay dry white wine with various inactive yeasts by 1H NMR spectroscopy combined with pattern recognition analysis. AMB Express 2019, 9, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Oh, E.J.; Jin, Y.-S. Engineering of Saccharomyces cerevisiae for efficient fermentation of cellulose. FEMS Yeast Res. 2020, 20. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Zhu, J.; Zhao, Q.; Gao, J.; Zhang, H.; Hu, B. Quality evaluation of Cabernet Sauvignon wines in different vintages by 1H nuclear magnetic resonance-based metabolomics. Open Chem. 2021, 19, 385–399. [Google Scholar] [CrossRef]
- Wendland, G. J. , The Whiff of Wine Yeast Innovation: Strategies for Enhancing Aroma Production by Yeast during Wine Fermentation. Journal of Agricultural and Food Chemistry 2019, 67. [Google Scholar]
- Quintero-Blanco, J.; Delodi, E.; Garzón, A.; Jimenez, J. Sexually-Driven Combinatorial Diversity in Native Saccharomyces Wine Yeasts. Fermentation 2022, 8, 569. [Google Scholar] [CrossRef]
- Wang, Z.; Chen, K.; Liu, C.; Ma, L.; Li, J. Effects of glycosidase on glycoside-bound aroma compounds in grape and cherry juice. J. Food Sci. Technol. 2023, 60, 761–771. [Google Scholar] [CrossRef] [PubMed]
- Viana, F.; Gil, J.V.; Vallés, S.; Manzanares, P. Increasing the levels of 2-phenylethyl acetate in wine through the use of a mixed culture of Hanseniaspora osmophila and Saccharomyces cerevisiae. Int. J. Food Microbiol. 2009, 135, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Bza, B.; Dxa, B.; Cda, B.; Gya, B. , Synergistic effect enhances 2-phenylethyl acetate production in the mixed fermentation of Hanseniaspora vineae and Saccharomyces cerevisiae. Process Biochemistry 2020, 90, 44–49. [Google Scholar]
- Li, T.; Yang, W.; Wu, S.; Selosse, M.-A.; Gao, J. Progress and Prospects of Mycorrhizal Fungal Diversity in Orchids. Front. Plant Sci. 2021, 12, 646325. [Google Scholar] [CrossRef]
- Merin, G.; Morata, V. I. Application of a grape surface majority pectinolytic species, Aureobasidium pullulans, to low-temperature red winemaking: development and stability of wine colour. Journal of Wine Research 2020, 31. [Google Scholar] [CrossRef]
- Yan, Q.; Pfleger, B.F. Revisiting metabolic engineering strategies for microbial synthesis of oleochemicals. Metab. Eng. 2019, 58, 35–46. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, C.; Feng, J.; Liu, Y.; Yang, Q.; Chen, H.; Gu, Z.; Zhang, H.; Chen, W.; Chen, Y.Q. Role of dihydrofolate reductase in tetrahydrobiopterin biosynthesis and lipid metabolism in the oleaginous fungus Mortierella alpina. Microbiology 2016, 162, 1544–1553. [Google Scholar] [CrossRef]
- Zhao, H.; Lv, M.; Liu, Z.; Zhang, M.; Wang, Y.; Ju, X.; Song, Z.; Ren, L.; Jia, B.; Qiao, M.; et al. High-yield oleaginous fungi and high-value microbial lipid resources from Mucoromycota. BioEnergy Res. 2020, 14, 1196–1206. [Google Scholar] [CrossRef]
- Shah, A.M.; Yang, W.; Mohamed, H.; Zhang, Y.; Song, Y. Microbes: A Hidden Treasure of Polyunsaturated Fatty Acids. Front. Nutr. 2022, 9, 827837. [Google Scholar] [CrossRef] [PubMed]
- Lane, S.; Dong, J.; Jin, Y.-S. Value-added biotransformation of cellulosic sugars by engineered Saccharomyces cerevisiae. Bioresource Technology: Biomass, Bioenergy, Biowastes, Conversion Technologies, Biotransformations. Bioresour. Technol. 2018, 260, 380–394. [Google Scholar] [CrossRef] [PubMed]
- Abdeljalil, S.; Ben Hmad, I.; Saibi, W.; Amouri, B.; Maalej, W.; Kaaniche, M.; Koubaa, A.; Gargouri, A. , Investigations on Hydrolytic Activities fromStachybotrys microsporaand Their Use as an Alternative in Yeast DNA Extraction. Applied Biochemistry & Biotechnology 2014, 172, 1599–1611. [Google Scholar]
- Cavello; Ivana; Cavalitto; Sebastian; Garmedia; Gabriela; Vero; Silvana; Albanesi; Agustin, Pectinolytic yeasts from cold environments: novel findings of Guehomyces pullulans, Cystofilobasidium infirmominiatum and Cryptococcus adeliensis producing pectinases. Extremophiles: Life under extreme conditions 2017, 21, 319–329.
- Liu, G. L. , The contribution of indigenous non-Saccharomyces wine yeast to improved aromatic quality of Cabernet Sauvignon wines by spontaneous fermentation. LWT-Food Science & Technology 2016, 71 (Null).
- Renault, P.; Coulon, J.; de Revel, G.; Barbe, J.-C.; Bely, M. Increase of fruity aroma during mixed T. delbrueckii/S. cerevisiae wine fermentation is linked to specific esters enhancement. Int. J. Food Microbiol. 2015, 207, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Miller, K. V.; Noguera, R.; Beaver, J.; Medina-Plaza, C.; Oberholster, A.; Block, D. E. , A Mechanistic Model for the Extraction of Phenolics from Grapes During Red Wine Fermentation. Molecules (Basel, Switzerland) 2019, 24. [Google Scholar] [CrossRef]
- Zhang, S.; Chen, X.; Zhong, Q.; Zhuang, X.; Bai, Z. Microbial Community Analyses Associated with Nine Varieties of Wine Grape Carposphere Based on High-Throughput Sequencing. Microorganisms 2019, 7, 668. [Google Scholar] [CrossRef] [PubMed]
- Kioroglou, D.; Mas, A.; Portillo, M.C. High-Throughput Sequencing Approach to Analyze the Effect of Aging Time and Barrel Usage on the Microbial Community Composition of Red Wines. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef]
Figure 1.
Dilution curve of different samples.CGE1 is the pre-fermentation Gardenblue blueberry wine sample, CGM2 is the mid-fermentation Gardenblue blueberry wine sample, CGL3 is the late fermentation Gardenblue blueberry wine sample, CSE1 is the pre-fermentation Powderblue blueberry wine sample, CSM2 is the mid-fermentation Powderblue blueberry wine sample, CSL3 is the late fermentation Powderblue blueberry wine sample, ME1 is the pre-fermentation Britewell blueberry wine sample, MM2 is the mid-fermentation Britewell blueberry wine sample, ML3 is the late fermentation Britewell blueberry wine sample.
Figure 1.
Dilution curve of different samples.CGE1 is the pre-fermentation Gardenblue blueberry wine sample, CGM2 is the mid-fermentation Gardenblue blueberry wine sample, CGL3 is the late fermentation Gardenblue blueberry wine sample, CSE1 is the pre-fermentation Powderblue blueberry wine sample, CSM2 is the mid-fermentation Powderblue blueberry wine sample, CSL3 is the late fermentation Powderblue blueberry wine sample, ME1 is the pre-fermentation Britewell blueberry wine sample, MM2 is the mid-fermentation Britewell blueberry wine sample, ML3 is the late fermentation Britewell blueberry wine sample.
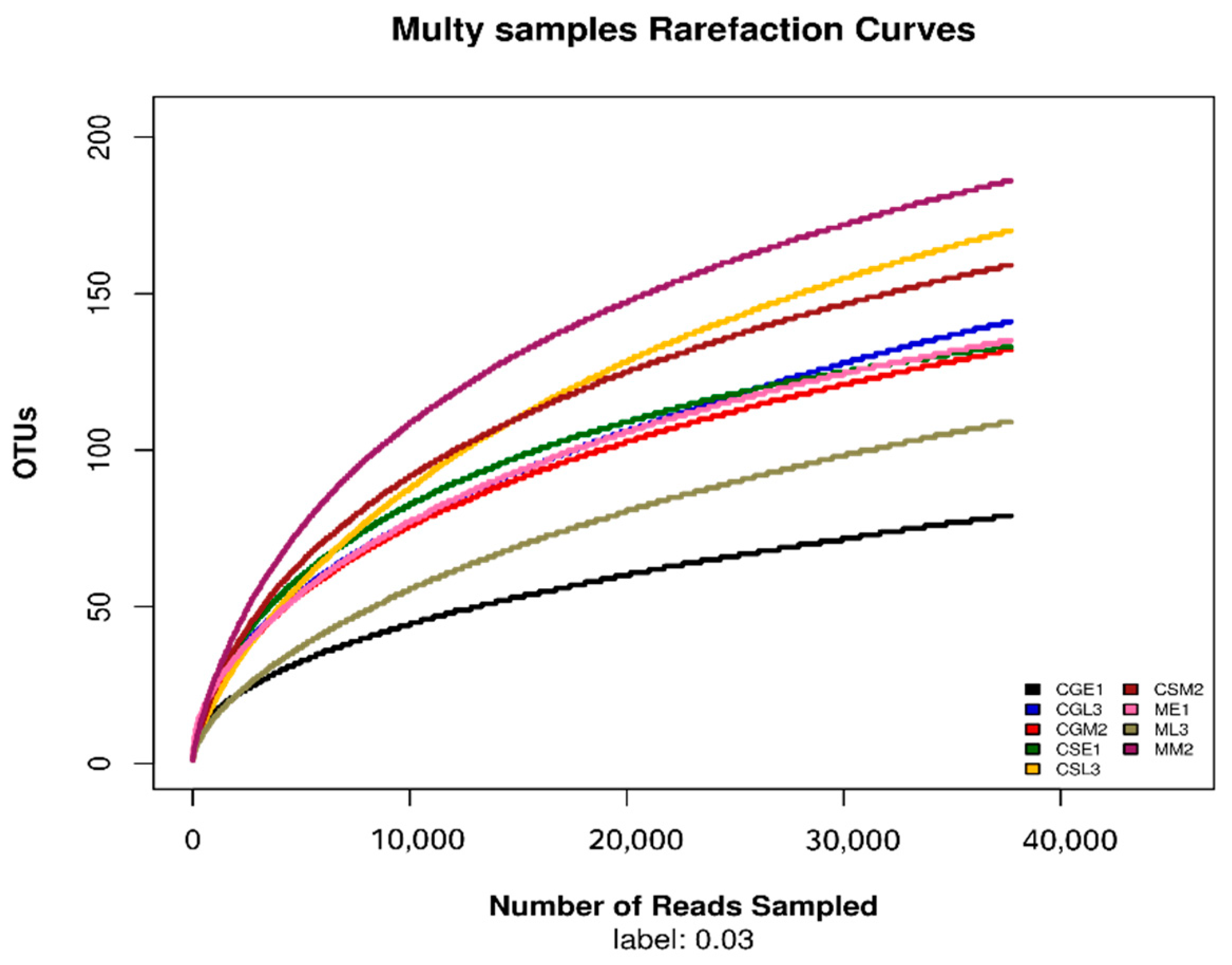
Figure 2.
Venn diagram of OTU distribution. S is a sample of Powderblue, G is a sample of Gardenblue, and M: is a sample of Britewell.
Figure 2.
Venn diagram of OTU distribution. S is a sample of Powderblue, G is a sample of Gardenblue, and M: is a sample of Britewell.
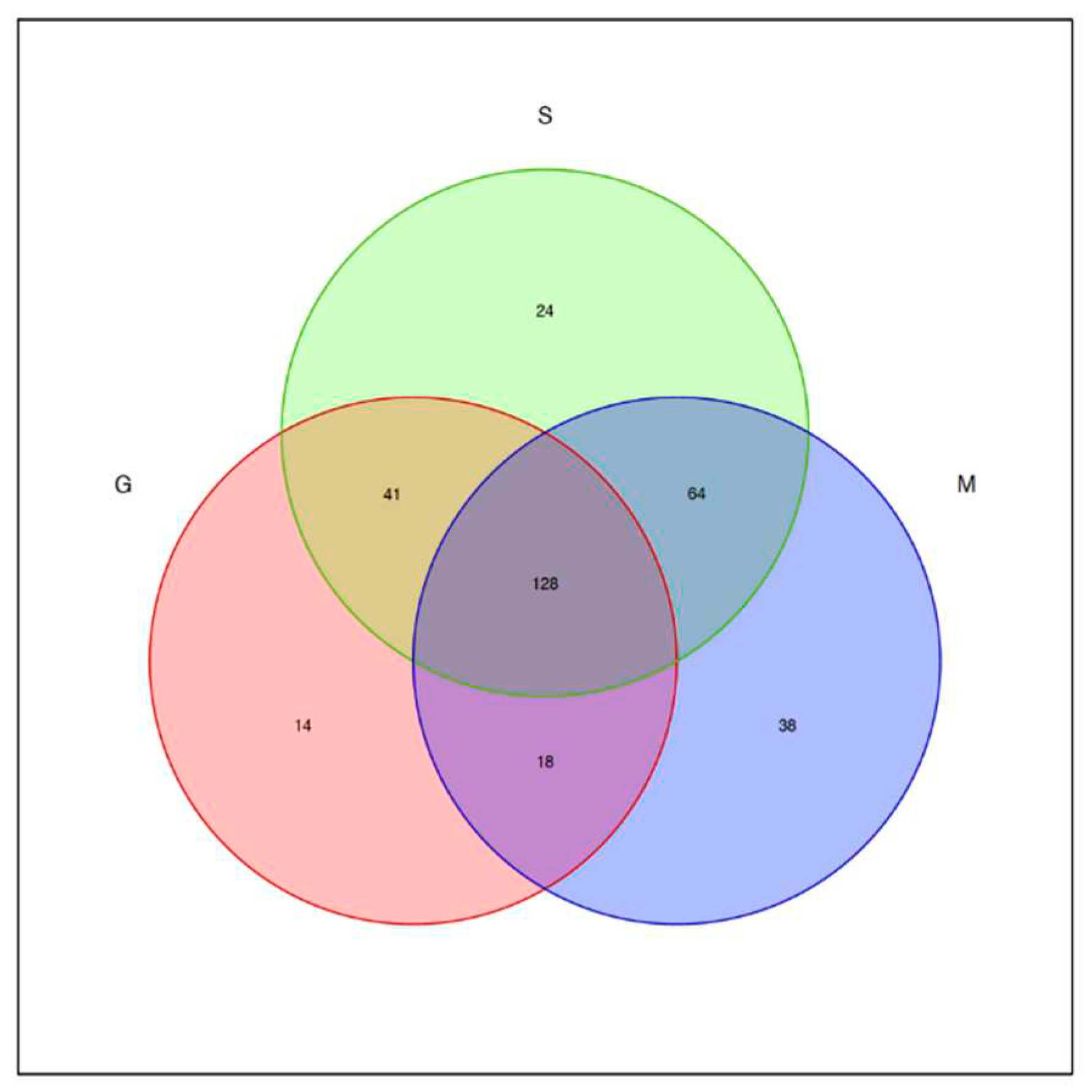
Figure 3.
Flower plot. CGE1 is the pre-fermentation Gardenblue blueberry wine sample, CGM2 is the mid-fermentation Gardenblue blueberry wine sample, CGL3 is the late fermentation Gardenblue blueberry wine sample, CSE1 is the pre-fermentation Powderblue blueberry wine sample, CSM2 is the mid-fermentation Powderblue blueberry wine sample, CSL3 is the late fermentation Powderblue blueberry wine sample, ME1 is the pre-fermentation Britewell blueberry wine sample, MM2 is the mid-fermentation Britewell blueberry wine sample, ML3 is the late fermentation Britewell blueberry wine sample. The core numbers inside the central circle indicate the number of otu present in all nine blueberry wine samples, and the numbers on the outer petals indicate the number of otu specific to that sample at different times for different blueberry varieties.
Figure 3.
Flower plot. CGE1 is the pre-fermentation Gardenblue blueberry wine sample, CGM2 is the mid-fermentation Gardenblue blueberry wine sample, CGL3 is the late fermentation Gardenblue blueberry wine sample, CSE1 is the pre-fermentation Powderblue blueberry wine sample, CSM2 is the mid-fermentation Powderblue blueberry wine sample, CSL3 is the late fermentation Powderblue blueberry wine sample, ME1 is the pre-fermentation Britewell blueberry wine sample, MM2 is the mid-fermentation Britewell blueberry wine sample, ML3 is the late fermentation Britewell blueberry wine sample. The core numbers inside the central circle indicate the number of otu present in all nine blueberry wine samples, and the numbers on the outer petals indicate the number of otu specific to that sample at different times for different blueberry varieties.
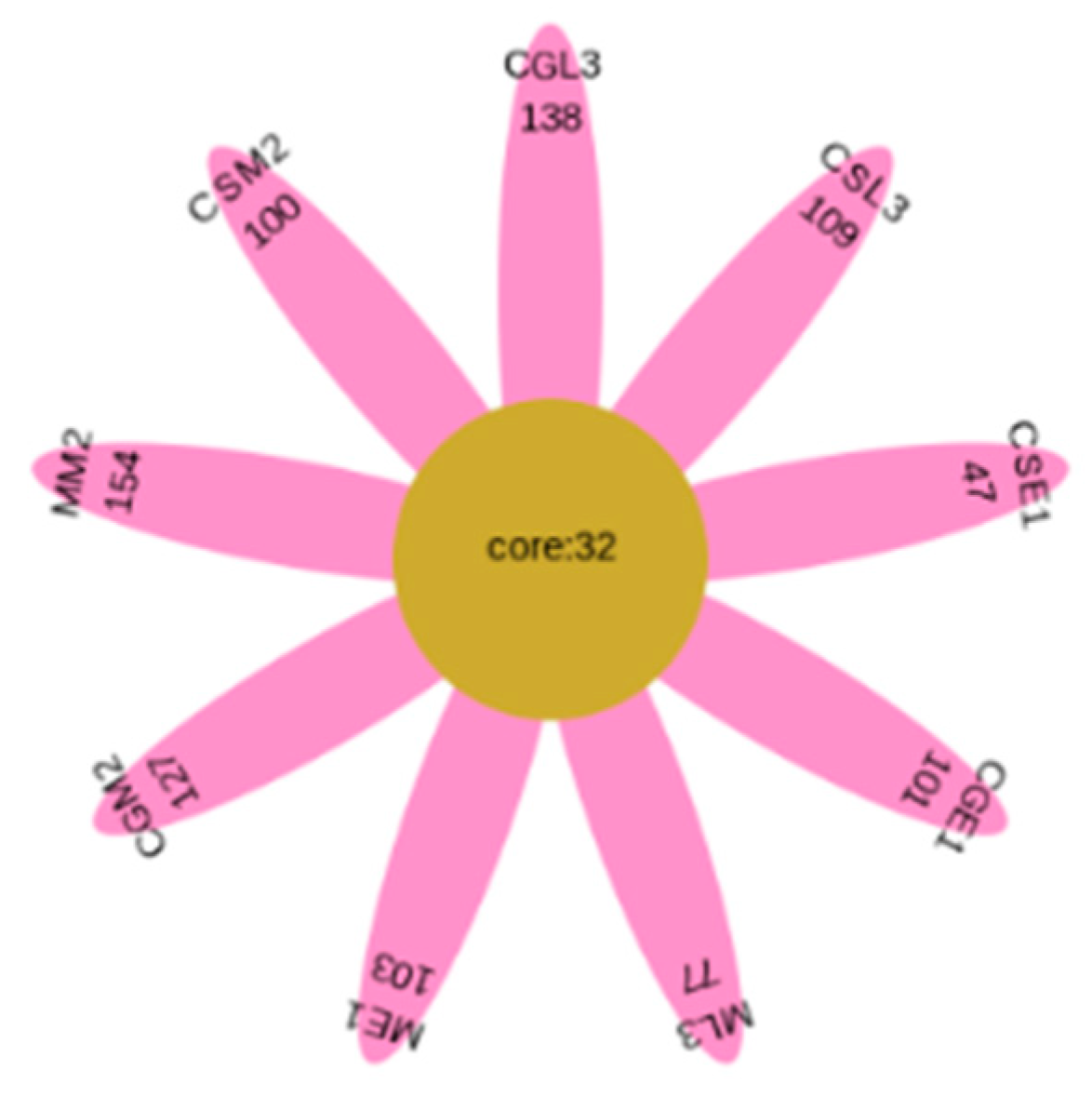
Figure 4.
Phylogenetic tree of fungi in wine-making habitat in Majiang, Guizhou province. In this figure, the first column starting from the left shows the blueberry wine sample information, including the phylum and genus of the fungus. The second column shows the fungal phylum classification corresponding to the fungal genus classification in the blueberry wine sample. In this case, the outermost circle of the phylogenetic tree shows the relative abundance of each fungal number in the blueberry wine sample, with the length of the different blocks of each color representing the corresponding level of relative abundance.
Figure 4.
Phylogenetic tree of fungi in wine-making habitat in Majiang, Guizhou province. In this figure, the first column starting from the left shows the blueberry wine sample information, including the phylum and genus of the fungus. The second column shows the fungal phylum classification corresponding to the fungal genus classification in the blueberry wine sample. In this case, the outermost circle of the phylogenetic tree shows the relative abundance of each fungal number in the blueberry wine sample, with the length of the different blocks of each color representing the corresponding level of relative abundance.
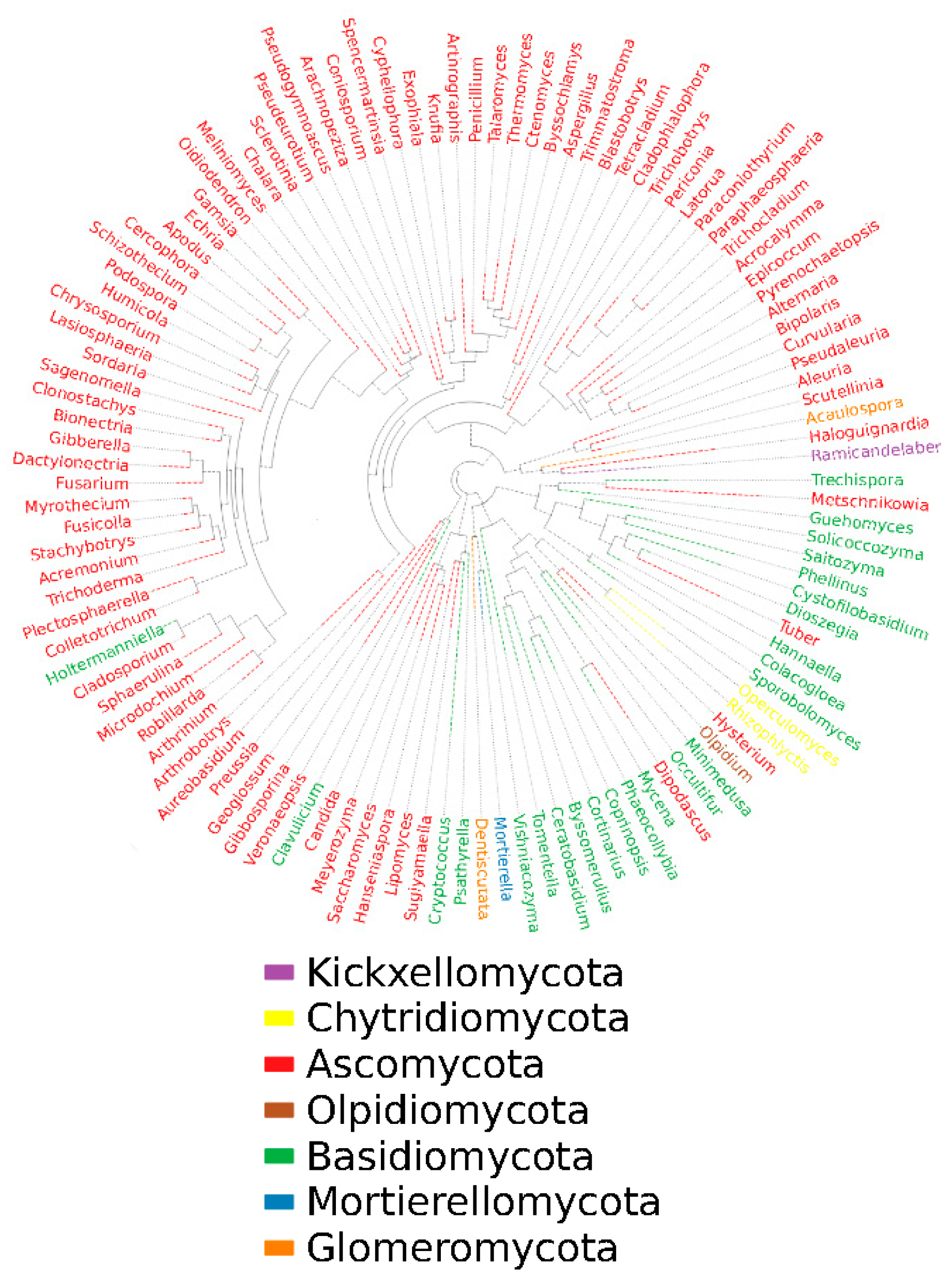
Figure 5.
Differences in fungal communities of different blueberry varieties before, during and after fermentation at the level of phylum(A) and genus(B). * CGE1 is the pre-fermentation Gardenblue blueberry wine sample, CGM2 is the mid-fermentation Gardenblue blueberry wine sample, CGL3 is the late fermentation Gardenblue blueberry wine sample, CSE1 is the pre-fermentation Powderblue blueberry wine sample, CSM2 is the mid-fermentation Powderblue blueberry wine sample, CSL3 is the late fermentation Powderblue blueberry wine sample, ME1 is the pre-fermentation Britewell blueberry wine sample, MM2 is the mid-fermentation Britewell blueberry wine sample, ML3 is the late fermentation Britewell blueberry wine sample.
Figure 5.
Differences in fungal communities of different blueberry varieties before, during and after fermentation at the level of phylum(A) and genus(B). * CGE1 is the pre-fermentation Gardenblue blueberry wine sample, CGM2 is the mid-fermentation Gardenblue blueberry wine sample, CGL3 is the late fermentation Gardenblue blueberry wine sample, CSE1 is the pre-fermentation Powderblue blueberry wine sample, CSM2 is the mid-fermentation Powderblue blueberry wine sample, CSL3 is the late fermentation Powderblue blueberry wine sample, ME1 is the pre-fermentation Britewell blueberry wine sample, MM2 is the mid-fermentation Britewell blueberry wine sample, ML3 is the late fermentation Britewell blueberry wine sample.
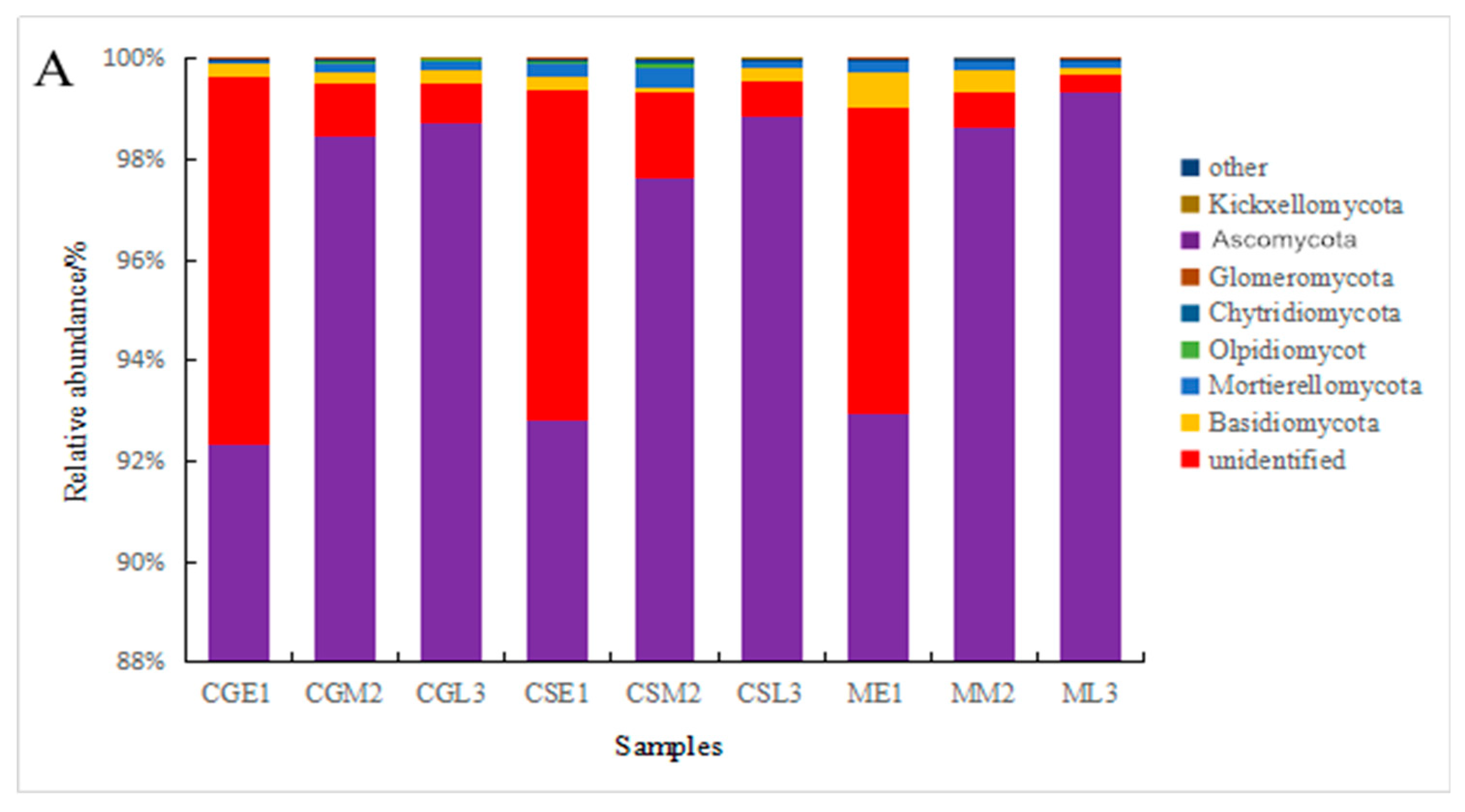
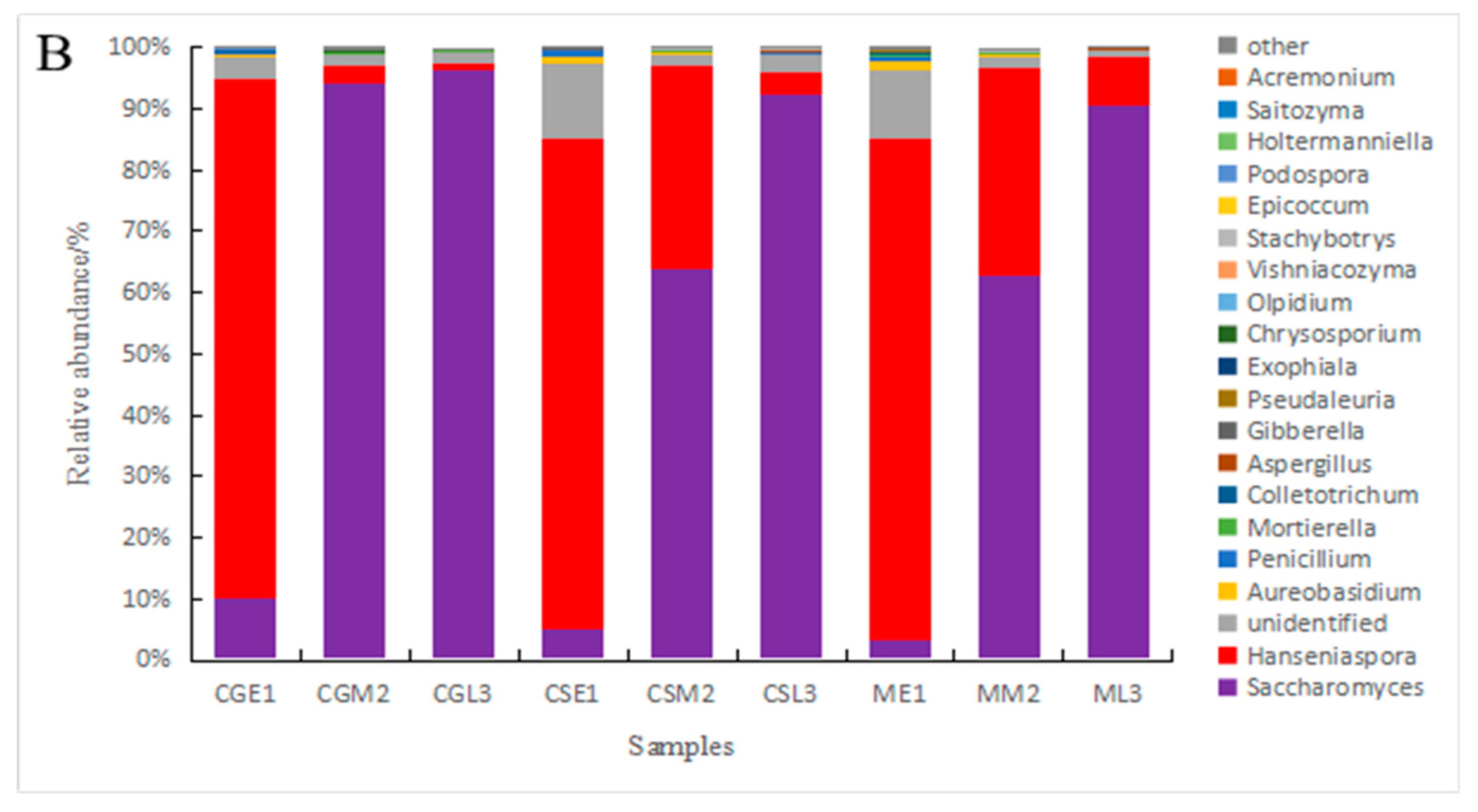
Figure 6.
Analysis of species assemblages at the fungal genus level in blueberry wine samples.
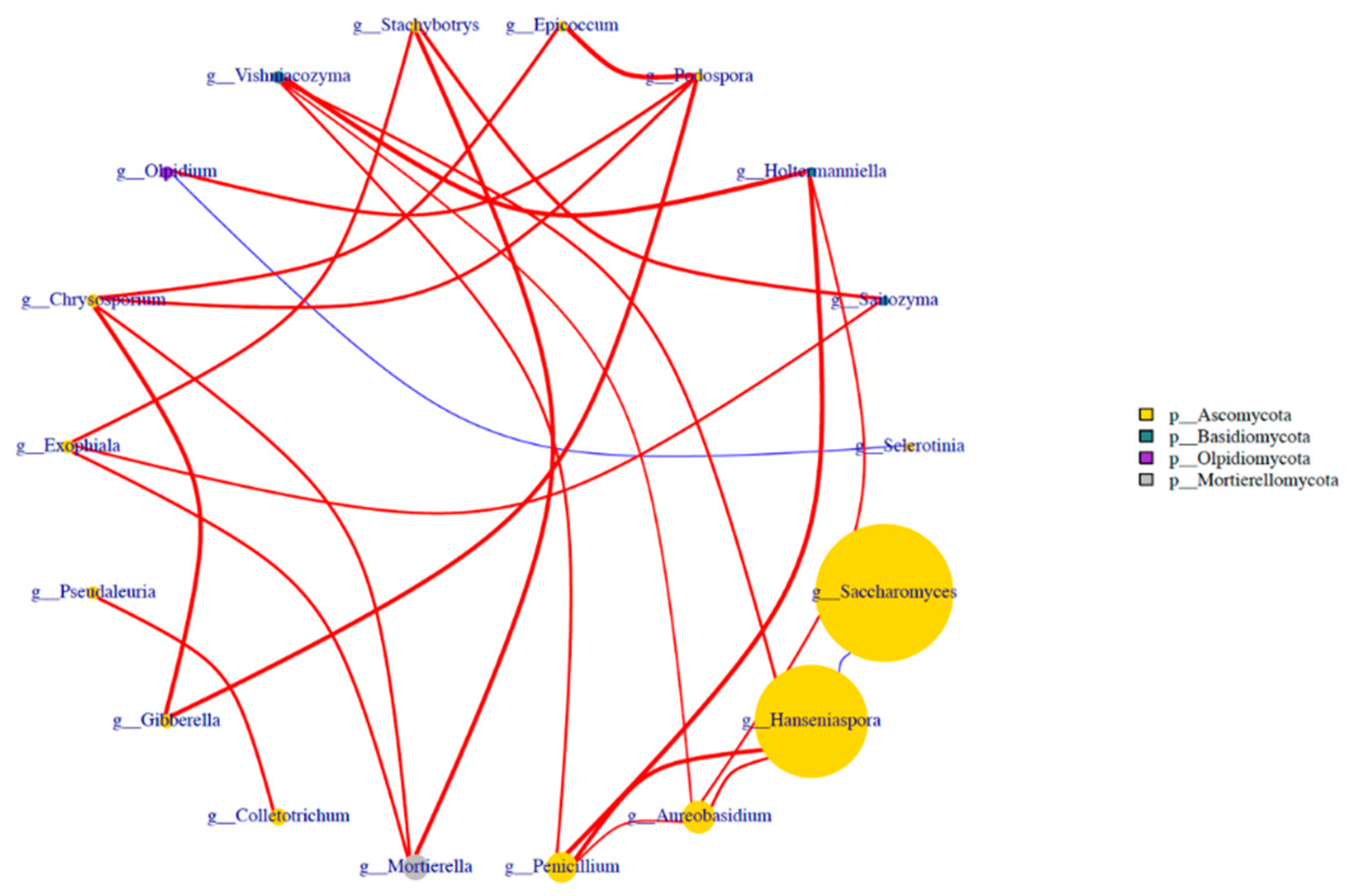
Figure 7.
OUT and texonomic level Heatmap. CGE1 is the pre-fermentation Gardenblue blueberry wine sample, CGM2 is the mid-fermentation Gardenblue blueberry wine sample, CGL3 is the late fermentation Gardenblue blueberry wine sample, CSE1 is the pre-fermentation Powderblue blueberry wine sample, CSM2 is the mid-fermentation Powderblue blueberry wine sample, CSL3 is the late fermentation Powderblue blueberry wine sample, ME1 is the pre-fermentation Britewell blueberry wine sample, MM2 is the mid-fermentation Britewell blueberry wine sample, ML3 is the late fermentation Britewell blueberry wine sample. The left side of the graph shows the cluster analysis of the affiliation of the blueberry wine samples, and the bottom of the graph indicates the abundance of the species, with darker colors representing the higher abundance of the species and lighter colors representing the lower abundance of the species.
Figure 7.
OUT and texonomic level Heatmap. CGE1 is the pre-fermentation Gardenblue blueberry wine sample, CGM2 is the mid-fermentation Gardenblue blueberry wine sample, CGL3 is the late fermentation Gardenblue blueberry wine sample, CSE1 is the pre-fermentation Powderblue blueberry wine sample, CSM2 is the mid-fermentation Powderblue blueberry wine sample, CSL3 is the late fermentation Powderblue blueberry wine sample, ME1 is the pre-fermentation Britewell blueberry wine sample, MM2 is the mid-fermentation Britewell blueberry wine sample, ML3 is the late fermentation Britewell blueberry wine sample. The left side of the graph shows the cluster analysis of the affiliation of the blueberry wine samples, and the bottom of the graph indicates the abundance of the species, with darker colors representing the higher abundance of the species and lighter colors representing the lower abundance of the species.
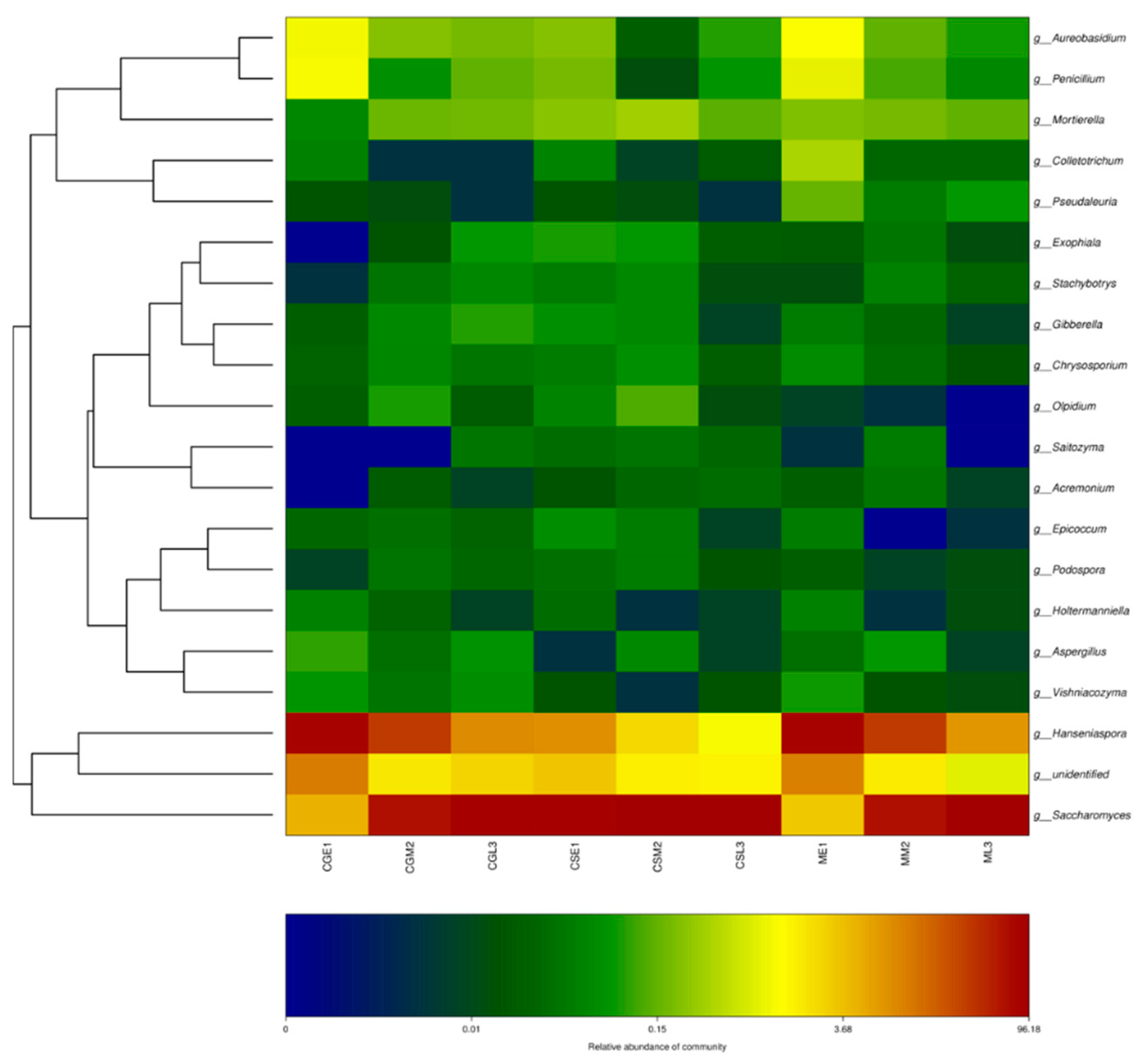
Figure 8.
Principal component analysis diagram of fungal communities in different samples. S is Powderblue blueberry wine sample, M is Britewell blueberry wine sample, G is Gardenblue blueberry wine sample. CGE1 is the pre-fermentation Gardenblue blueberry wine sample, CGM2 is the mid-fermentation Gardenblue blueberry wine sample, CGL3 is the late fermentation Gardenblue blueberry wine sample, CSE1 is the pre-fermentation Powderblue blueberry wine sample, CSM2 is the mid-fermentation Powderblue blueberry wine sample, CSL3 is the late fermentation Powderblue blueberry wine sample, ME1 is the pre-fermentation Britewell blueberry wine sample, MM2 is the mid-fermentation Britewell blueberry wine sample, ML3 is the late fermentation Britewell blueberry wine sample.
Figure 8.
Principal component analysis diagram of fungal communities in different samples. S is Powderblue blueberry wine sample, M is Britewell blueberry wine sample, G is Gardenblue blueberry wine sample. CGE1 is the pre-fermentation Gardenblue blueberry wine sample, CGM2 is the mid-fermentation Gardenblue blueberry wine sample, CGL3 is the late fermentation Gardenblue blueberry wine sample, CSE1 is the pre-fermentation Powderblue blueberry wine sample, CSM2 is the mid-fermentation Powderblue blueberry wine sample, CSL3 is the late fermentation Powderblue blueberry wine sample, ME1 is the pre-fermentation Britewell blueberry wine sample, MM2 is the mid-fermentation Britewell blueberry wine sample, ML3 is the late fermentation Britewell blueberry wine sample.
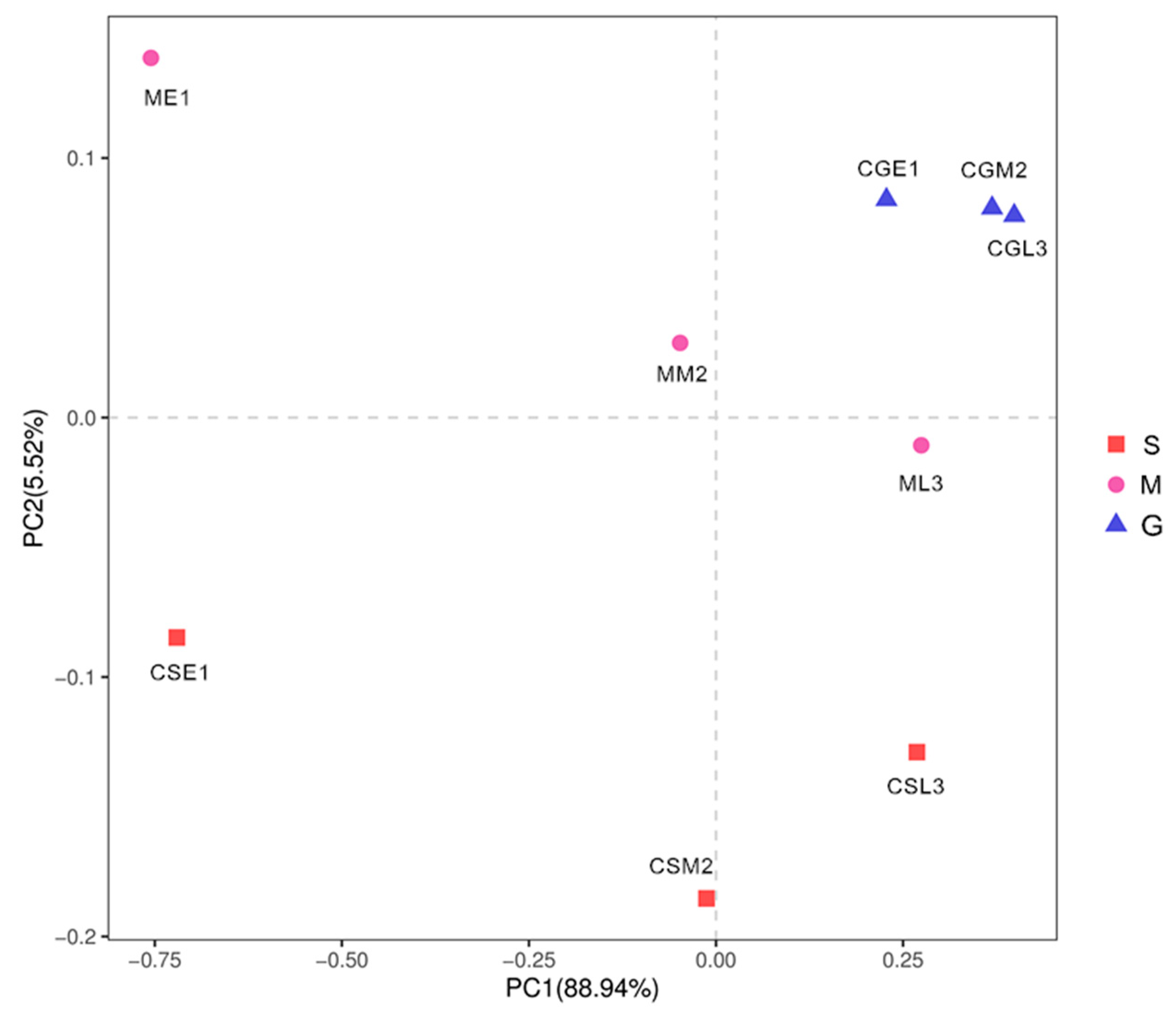
Figure 9.
Clustering tree diagram of fungal community species in different samples. CGE1 is the pre-fermentation Gardenblue blueberry wine sample, CGM2 is the mid-fermentation Gardenblue blueberry wine sample, CGL3 is the late fermentation Gardenblue blueberry wine sample, CSE1 is the pre-fermentation Powderblue blueberry wine sample, CSM2 is the mid-fermentation Powderblue blueberry wine sample, CSL3 is the late fermentation Powderblue blueberry wine sample, ME1 is the pre-fermentation Britewell blueberry wine sample, MM2 is the mid-fermentation Britewell blueberry wine sample, ML3 is the late fermentation Britewell blueberry wine sample.
Figure 9.
Clustering tree diagram of fungal community species in different samples. CGE1 is the pre-fermentation Gardenblue blueberry wine sample, CGM2 is the mid-fermentation Gardenblue blueberry wine sample, CGL3 is the late fermentation Gardenblue blueberry wine sample, CSE1 is the pre-fermentation Powderblue blueberry wine sample, CSM2 is the mid-fermentation Powderblue blueberry wine sample, CSL3 is the late fermentation Powderblue blueberry wine sample, ME1 is the pre-fermentation Britewell blueberry wine sample, MM2 is the mid-fermentation Britewell blueberry wine sample, ML3 is the late fermentation Britewell blueberry wine sample.
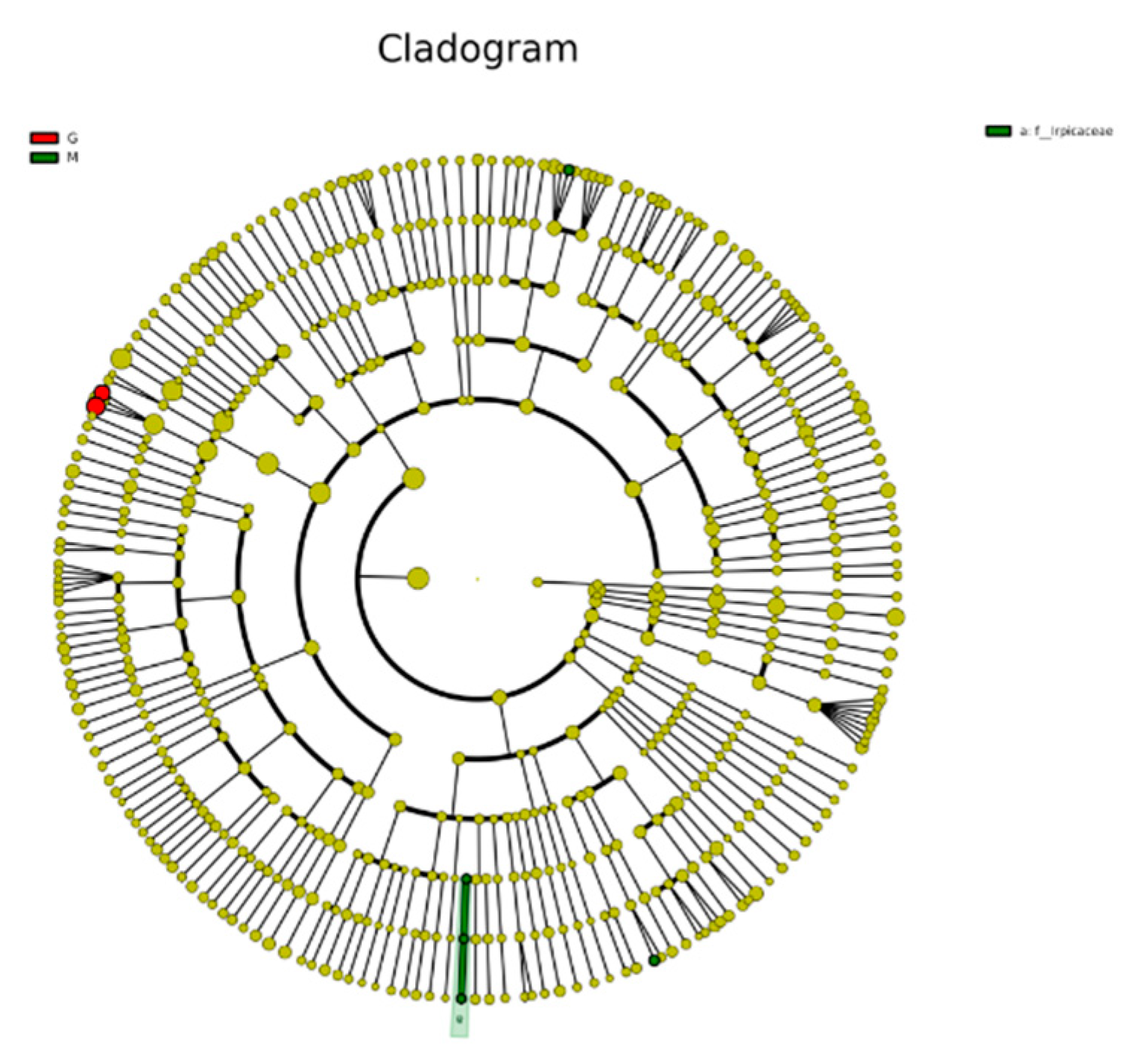

Table 1.
Sample sequence length distribution.
| Sequence length gradient | Number of Sequencs |
|---|---|
| 0-200 | 1109 |
| 200-260 | 28770 |
| 260-320 | 1521 |
| 320-360 | 17800 |
| 360-380 | 136776 |
| 380-400 | 11 |
| 400-420 | 11 |
| 420-440 | 296628 |
| 440-460 | 197 |
| 460-480 | 10 |
| 480-500 | 10 |
| 500-520 | 9 |
| 520-540 | 0 |
| 540-560 | 0 |
| 560-600 | 0 |
Table 2.
Statistical table of Alpha Diversity Index of samples.
| Name of Samples | chao1 | coverage | observed_species | PD_whole_tree | shannon | simpson |
|---|---|---|---|---|---|---|
| CGE1 | 130.00 | 1.00 | 79.00 | 19.65 | 1.69 | 0.50 |
| CGM2 | 203.55 | 1.00 | 132.00 | 30.62 | 1.60 | 0.53 |
| CGL3 | 217.25 | 1.00 | 141.00 | 32.62 | 1.00 | 0.25 |
| CSE1 | 150.03 | 1.00 | 133.00 | 30.64 | 0.95 | 0.25 |
| CSM2 | 220.60 | 1.00 | 159.00 | 37.49 | 0.55 | 0.11 |
| CSL3 | 233.17 | 1.00 | 170.00 | 37.29 | 0.42 | 0.07 |
| ME1 | 176.93 | 1.00 | 135.00 | 29.58 | 1.43 | 0.36 |
| MM2 | 239.82 | 1.00 | 186.00 | 42.59 | 1.39 | 0.50 |
| ML3 | 156.00 | 1.00 | 109.00 | 26.59 | 0.65 | 0.17 |
* CGE1 is the pre-fermentation Gardenblue blueberry wine sample, CGM2 is the mid-fermentation Gardenblue blueberry wine sample, CGL3 is the late fermentation Gardenblue blueberry wine sample, CSE1 is the pre-fermentation Powderblue blueberry wine sample, CSM2 is the mid-fermentation Powderblue blueberry wine sample, CSL3 is the late fermentation Powderblue blueberry wine sample, ME1 is the pre-fermentation Britewell blueberry wine sample, MM2 is the mid-fermentation Britewell blueberry wine sample, ML3 is the late fermentation Britewell blueberry wine sample.
Table 3.
Proportion distribution of representative strains in different samples.
| Name of strains | CGE1 | CGM2 | CGL3 | CSE1 | CSM2 | CSL3 | ME1 | MM2 | ML3 |
|---|---|---|---|---|---|---|---|---|---|
| S.cerevisiae | 4.9720% | 63.8434% | 86.3965% | 86.2348% | 94.4048% | 96.1841% | 3.4287% | 62.9180% | 90.6234% |
| H.uvarum | 68.9374% | 22.6035% | 3.4897% | 8.6447% | 2.5589% | 1.1376% | 79.5656% | 32.4918% | 6.4225% |
| A.pullulans | 1.1111% | 0.2413% | 0.1962% | 0.2493% | 0.0159% | 0.0955% | 1.2967% | 0.1511% | 0.0796% |
| M.elongata | 0.0345% | 0.0902% | 0.0796% | 0.1034% | 0.1405% | 0.0345% | 0.1671% | 0.0398% | 0.0902% |
| O.brassicae | 0.0159% | 0.0849% | 0.0133% | 0.0451% | 0.1246% | 0.0080% | 0.0053% | 0.0027% | - |
| S.microspora | 0.0027% | 0.0292% | 0.0477% | 0.0345% | 0.0530% | 0.0080% | 0.0080% | 0.0424% | 0.0186% |
| M.alpina | 0.0027% | 0.0265% | 0.0080% | 0.0265% | 0.0212% | 0.0504% | 0.0133% | 0.0796% | 0.0292% |
| H.takashimae | 0.0424% | 0.0186% | 0.0053% | 0.0239% | 0.0027% | 0.0053% | 0.0424% | 0.0027% | 0.0080% |
| P.sp | - | - | - | 0.0186% | 0.0159% | - | - | - | - |
| M.sp | 0.0106% | - | 0.0133% | 0.0133% | 0.0053% | - | 0.0053% | 0.0027% | - |
| D.erythropus | - | 0.0027% | - | 0.0106% | 0.0106% | 0.0027% | 0.0053% | - | 0.0053% |
| G.pullulans | 0.0053% | 0.0106% | 0.0186% | 0.0080% | 0.0106% | 0.0080% | 0.0106% | 0.0133% | 0.0133% |
* CGE1 is the pre-fermentation Gardenblue blueberry wine sample, CGM2 is the mid-fermentation Gardenblue blueberry wine sample, CGL3 is the late fermentation Gardenblue blueberry wine sample, CSE1 is the pre-fermentation Powderblue blueberry wine sample, CSM2 is the mid-fermentation Powderblue blueberry wine sample, CSL3 is the late fermentation Powderblue blueberry wine sample, ME1 is the pre-fermentation Britewell blueberry wine sample, MM2 is the mid-fermentation Britewell blueberry wine sample, ML3 is the late fermentation Britewell blueberry wine sample. - indicates relative abundance below 0.0001% or not detected.
Table 4.
Different samples share OUT.
| OTUs | level | taxonomy |
|---|---|---|
| OTU-1 | species | Saccharomyces cerevisiae |
| OTU-2 | species | Hanseniaspora uvarum |
| OTU-4 | species | Hanseniaspora osmophila |
| OTU-5 | species | Hanseniaspora vineae |
| OTU-48 | species | Pseudaleuria sp |
| OTU-102 | species | Aureobasidium pullulans |
| OTU-119 | species | Fungi sp |
| OTU-122 | species | Mortierella alpina |
| OTU-131 | species | Lasiosphaeriaceae sp |
| OTU-135 | species | Colletotrichum salsolae |
| OTU-136 | species | Chrysosporium synchronum |
| OTU-138 | species | Bipolaris drechsleri |
| OTU-148 | species | Fungi sp |
| OTU-192 | species | Stachybotrys microspora |
| OTU-196 | species | Podospora sp |
| OTU-217 | species | Holtermanniella takashimae |
| OTU-221 | species | Penicillium sp |
| OTU-222 | species | Guehomyces pullulans |
| OTU-241 | species | Gibberella baccata |
| OTU-262 | species | Mortierella elongata |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



