
Submitted:
11 September 2023
Posted:
12 September 2023
You are already at the latest version
Abstract
Alzheimer’s disease (AD) is the most common form of neurodegenerative disease worldwide. A large body of work implicates insulin resistance in the development and progression of AD such that AD may be a form of brain insulin resistance. Moreover, impairment in mitochondrial function, a common symptom of insulin resistance, now represents a fundamental aspect of AD pathobiology. Ceramides are a class of bioactive sphingolipids that have been hypothesized to drive insulin resistance. Here we describe preliminary work that tests the hypothesis that hyperinsulinemia pathologically alters cerebral mitochondrial function in AD mice via accrual of the ceramides. Homozygous male and female ApoE4 mice were given chronic injections of PBS (control), insulin, myriocin (an inhibitor of ceramide biosynthesis), or insulin and myriocin over four weeks. Cerebral ceramide content was assessed using liquid chromatography-mass spectrometry. Mitochondrial oxygen consumption rates were measured with high-resolution respirometry, and H2O2 emissions were quantified via biochemical assays on brain tissue from the cerebral cortex. Significant increases in brain ceramides and impairments in brain oxygen consumption were observed in the insulin-treated group. These hyperinsulinemia-induced impairments in mitochondrial function were reversed with the administration of myriocin. Altogether, these data demonstrate a causative role for insulin in promoting brain ceramide accrual and subsequent mitochondrial impairments that may be involved in AD expression and progression.
Keywords:
insulin resistance
; Alzheimer’s disease
; ApoE4
; dyslipidemia
; ceramides
; mitochondrial bioenergetics
; cerebral cortex
1. Introduction
Hyperinsulinemia, both the driving cause and clinical manifestation of insulin resistance [1], is the most common metabolic disorder worldwide [2,3]. Appropriately, much of the research on insulin resistance and hyperinsulinemia revolves around insulin’s cardiometabolic roles in the development of type 2 diabetes, hypertension [4], atherosclerosis [5], among others. However, once thought to be an insulin-independent organ [6,7], it is now widely accepted that the brain is also insulin-responsive, and therefore, is likely susceptible to fluctuations in peripheral insulin and insulin sensitivity.
Insulin is a hormone secreted by pancreatic β cells that dictates energy utilization within the body and regulates cellular glucose uptake. As a brain growth factor, insulin is necessary in development, but is also highly involved in other processes in adulthood [8]. The hormone is transported into the brain via saturable transporters in the endothelial cells of the blood-brain barrier. While the majority of glucose uptake in the brain occurs independent of insulin via insulin-insensitive glucose transporters GLUT1 and GLUT3 expressed in glia and neurons, respectively [9], regions involved in regulating whole-body energy homeostasis, cognition, and other functions (e.g., hypothalamus, hippocampus and cerebral cortex, olfactory bulb, cerebellum) express the insulin-sensitive GLUT transporter, GLUT4 [10,11]. Indeed, insulin signaling has been shown to play essential roles synapse density, regulating synaptogenesis and synaptic plasticity [12].
Alzheimer’s disease (AD) is the most common form of dementia and is characterized by neurodegeneration that progressively impairs cognition and behavior. The disease affects approximately 6.5 million Americans 65 years and older [13], which is projected to increase to 13.8 million by 2060. Interestingly, the global rise in AD parallels trends in insulin resistance and metabolic syndrome, hinting at an important relationship between peripheral insulin and brain function [14]. Indeed, this relationship has been explored by Kuusisto et al., who suggested the relevance of insulin and glucose metabolism in AD, and demonstrated that insulin resistance significantly correlates with AD [15]. It is now widely acknowledged that impairments in brain metabolism, insulin sensitivity, and mitochondrial function are core characteristics of the disease.
In AD, neurodegeneration follows a pattern such that brain regions involved in learning and memory, such as the hippocampus and entorhinal cortex, display neurodegeneration in early disease stages. However, as the disease progresses, neurodegeneration spreads to other cognitive areas of the brain, including areas of the cerebral cortex responsible for language, decision-making, sociality, and other more basic behaviors. Specifically, the connection between the hippocampus and prefrontal cortex is associated with cognitive dysfunction in AD and represents the link between cognition and emotion, which are both disrupted in AD progression [16].
Ceramides are a bioactive family of sphingolipids with structural and functional roles within the cell. Under normal conditions, ceramides found in cell membranes are typically associated with lipid rafts that provide structural support and also have roles in cell signaling that mediate cell growth, proliferation, senescence, and apoptosis, among other processes [17]. However, under obesogenic conditions, ceramides can take on maladaptive roles. Rapid increases in adiposity induce elevations in plasma free fatty acids (FFA). In peripheral organs, FFA can either be oxidized as energy substrates, ectopically stored in pools of triacylglycerols (TAG), or used to produce other lipid species [18]. Exceeding an organ’s TAG storage capacity can lead to the accumulation of toxic lipid metabolites (e.g., ceramides), which has been shown to activate inflammatory cytokines and promote both local and systemic metabolic dysfunction such as insulin resistance. Lipotoxic ceramides have been hypothesized to link peripheral adiposity and central insulin resistance, which have substantial implications for AD etiology and treatment [19].
Previous studies have demonstrated significant elevations in brain ceramide content in patients with AD and other neurodegenerative disorders [20] and have, as mediators of apoptosis, been suggested to drive neurodegeneration [21,22]. One study demonstrated that a high fat diet increases pro-ceramide gene expression in the liver but not brain of wildtype C57BL/6 mice [23]. The increase in hepatic ceramide synthesis machinery increased oxidative stress and markers of neurodegeneration in the temporal lobe, suggesting that brain insulin resistance may be mediated by hepatic production of ceramides that cross the blood-brain barrier and promote apoptosis. However, the effect of the HFD on the accumulation of ceramides in the brain was not directly assessed.
The accumulation of long-chain ceramides has been observed in post-mortem AD brains [24]. In this study, we explore whether hyperinsulinemia is sufficient to increase peripheral adiposity and increase ceramide accumulation within the brain of ApoE4 mice, a model of sporadic AD. We hypothesize that changes in brain ceramide content impairs mitochondrial function. The data presented here suggest ceramides may play a causal role in insulin-induced mitochondrial impairment in AD and represent a potential target for future research.
2. Results
2.1. Chronic Insulin Injections Increase Body Weight and Reduce Insulin Tolerance in Male and Female ApoE4 Mice
Over the four weeks of conditioning, insulin injections elicited a significant increase in body weight compared with all other groups at weeks three and four. Myriocin treatment reversed this effect (INS + MYR) (Figure 1a). The chronic insulin injections alone, consistent with our previous work in WT mice [25,26], elicited an elevation in fasting glucose that reverted to control levels with myriocin treatment (Figure 1b). Chronic insulin injections decreased insulin sensitivity, evidenced by a blunted reduction in plasma glucose concentrations following insulin injection in the ITT that was inhibited with MYR treatment (Figure 1c).
2.2. Insulin Increases Brain Ceramide Accrual in Male and Female ApoE4 Mice
To identify changes in brain ceramide concentrations following the induction of hyperinsulinemia, a broad spectrum of ceramide species were assessed. Ceramides with side chains C16:1, C20, C24, and C24:1 were significantly increased with insulin treatment and reverted to control levels when combined with MYR (Figure 2). Together, the increase in C16:1, C20, C24, and C24:1 ceramides yielded a robust increase in total ceramide levels with insulin injections that was rescued with myriocin administration (INS+MYR). Myriocin treatment alone also significantly reduced total ceramide content compared to control PBS-treated mice.
2.3. Insulin Disrupts Mitochondrial Function in Brain Tissue of Male and Female ApoE4 Mice
Mitochondrial oxygen consumption rates were measured to assess the effects of hyperinsulinemia on mitochondrial function in our APOE4 mice (Figure 3a). Insulin treatment significantly reduced respiration rates associated with oxidative phosphorylation (i.e. with the addition of ADP; GMP) and complex II-supported respiration (i.e. with the addition of succinate, GMSP) compared to PBS controls. A significant reduction was also observed in respiratory rate, an indicator of general mitochondrial fitness (Figure 3b), and complex II factor with insulin injections (Figure 3c). Myriocin treatment reverted RCR to control levels, but not CII factor.
2.4. Chronic Insulin Injections Direct Brain O2 Use Towards H2O2 Production in Male and Female ApoE4 Mice
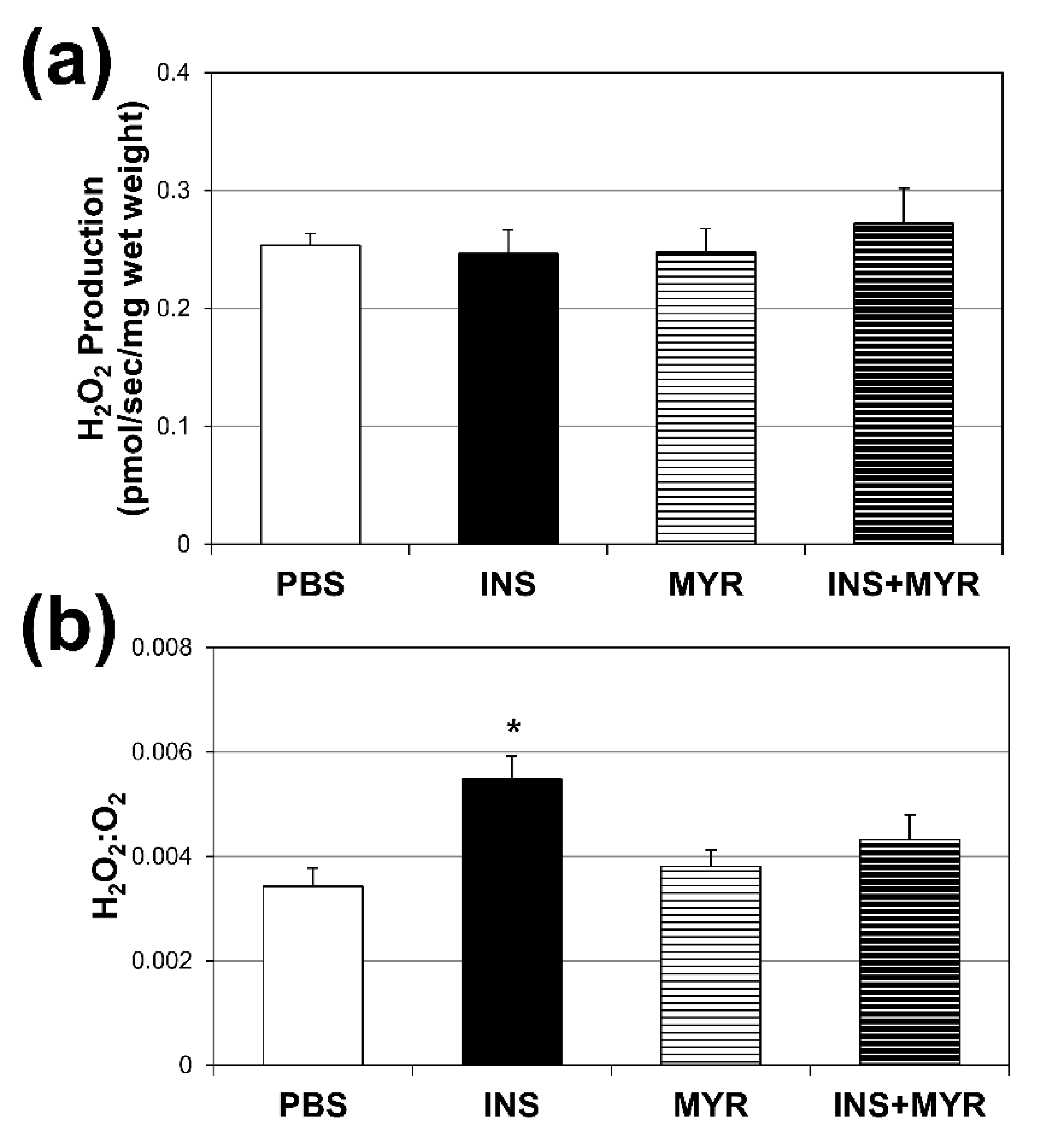
Absolute rates of H2O2 production did not vary across the treatment groups (Figure 4a). However, when compared with the amount of O2 consumed (Figure 4b), the INS group revealed a roughly 54% increase in H2O2 production per unit O2 consumed. This significant enhancement in H2O2 production per unit O2 consumption was once again rescued by inhibiting ceramide synthesis with MYR.
3. Discussion
The global rise in insulin resistance and metabolic dysfunction poses a substantial threat to cognitive health and increases the risk of developing dementias such as AD [27]. AD is a multifactorial disease involving the interplay of both genetics and the environment [28]. In this study we explored the intersection of an ApoE4 genetic background and hyperinsulinemia in mice. ApoE4, a variant of the lipoprotein ApoE, is the strongest genetic risk factor for developing AD [29], suggesting a link between disrupted lipid homeostasis and AD. Here we add support to this link by implicating ceramides, a sphingolipid, in AD mitochondrial dysfunction.
Although hyperinsulinemia and obesity alone are insufficient to cause AD [19], the data presented in this study demonstrate that hyperinsulinemia is sufficient to increase the accumulation of ceramides within the cortex of ApoE4 mice and significantly impair mitochondrial bioenergetics. These mitochondrial effects were reversed with the systemic administration of myriocin, a potent serine-palmitoyltransferase (SPT) inhibitor, which inhibits ceramide synthesis and demonstrates that hyperinsulinemia-induced mitochondrial impairments were mediated by ceramides. These results agree with previous reports that inhibiting ceramide synthesis improves virtually all metabolic disorders in rodents [30]. Although we did not assess the longitudinal effects of hyperinsulinemia on ceramide accrual and downstream mitochondrial dysfunction, these data demonstrate insulin resistance impairs mitochondrial function through ceramide accrual and suggest both insulin resistance and sphingolipids are relevant to AD.
We acknowledge that including only ApoE4 and not wildtype mice in this study limited our ability to draw conclusions about the detriment of insulin and the efficacy of myriocin. However, our observation that hyperinsulinemia elevated cortical ceramide content agreed with AD pathology, as previous studies have demonstrated significant elevations in brain ceramides in patients with AD and other neurodegenerative disorders [20]. These studies conclude that the normally tightly controlled regulation of ceramide synthesis is likely lost following neurodegeneration. They suggest that excessive ceramide accrual in the brain of AD patients is a consequence of disease mechanisms. Here we report that hyperinsulinemia induced elevations in cortex ceramide concentrations that impaired mitochondrial oxygen consumption (Figure 3A) and increased the rate of H2O2 production (Figure 4) in ApoE4 mice. These effects were reversed by inhibiting ceramide synthesis with myriocin, which demonstrates a causal role for ceramides and insulin in AD mitochondrial dysfunction and suggests that ceramide accrual may be more of an active driver than a mere consequence of the disease.
Chronic insulin exposure reduced peripheral insulin tolerance (Figure 1C), which supports previous findings that insulin alone, from endogenous or exogenous sources, promotes insulin resistance [25,31-34]. Myriocin treatment protected against deleterious changes in insulin sensitivity and other indicators of insulin resistance that were assessed.
In the current study, hyperinsulinemia increased cerebral cortex concentrations of C16:1, C20, C24, and C24:1 ceramide (Figure 2). We determined these elevations were due to an increase in de novo ceramide synthesis, as the insulin-induced increase in ceramides was reversed with myriocin treatment, an inhibitor of SPT, the rate-limiting step of de novo ceramide synthesis. From the data presented here, determining whether these ceramides were the result of an increase in central or peripheral de novo synthesis was not possible. However, the significant increase in body mass with insulin treatment (Figure 1A) was due to an increase in peripheral adipose mass. Therefore, the increase in brain ceramides was likely a result of ceramide synthesis in peripheral adipose stores and organ-specific triglyceride pools. This would further support the link between dysregulated peripheral lipid metabolism and Alzheimer’s disease but would require more explicit exploration in the future.
The de novo synthesis pathway is the most-studied and most-robust path to ceramide production [35]. The process begins in the endoplasmic reticulum, where the enzyme SPT condenses serine and palmitoyl-CoA to form 3-keto-sphinganine (3KSn), the 18-carbon backbone of other, more complex sphingolipids, which are extremely relevant to the brain and are heavily enriched in myelin sheaths surrounding CNS axons [36]. 3KSn is then reduced via 3KSn reductase to sphinganine, which is combined with a variable length fatty acyl-CoA by ceramide synthase (CerS) yielding dihydroceramide, which undergoes desaturation to produce ceramides. There are six CerS that catalyze the acylation of sphinganine and are limited in the fatty acyl-CoAs they can use as substrates. While we did not measure the expression of different synthases within the cortex, it would be interesting to know whether the increases in C16:1, C20, C24, and C24:1 ceramide were due to increases in CerS, which would give us further insight into the origins of these ceramides.
High plasma ceramide concentrations have been associated with hippocampal atrophy and cognitive impairment in Alzheimer’s disease [37-39]. More recent work has demonstrated that the plasma ratio of very long (C22-24) to long (C16-18) chain ceramides have more predictive relevance to Alzheimer’s disease than total ceramide concentrations [40]. They suggest that a lower plasma ratio of very long-chain to long-chain ceramides is associated with a higher risk of AD and may be a means of noninvasively assessing disease risk. In this study, we measured the concentrations of specific ceramide species in the cerebral cortex, but not plasma. Because we observed significant increases in total ceramide content in the cortex, it is likely that total ceramide content in the plasma also increased. It is possible that there was a lower ratio of very long-chain to long-chain ceramides in the plasma. However, this would require further study.
Together, these data add to the growing body of evidence suggesting AD is a metabolic disorder and can be characterized by impairments in brain energy homeostasis and mitochondrial function. More specifically, this work joins other research which suggest ceramides as drivers of AD and potential markers and drug targets of the disease [41-43]. While we did not study the onset of clinically-relevant (i.e., neurocognitive, and other behavioral) disease symptoms, we demonstrate a causative role for insulin in AD mitochondrial dysfunction via ceramides. We show that hyperinsulinemia is sufficient to increase cortical ceramide content, which has deleterious effects on mitochondrial bioenergetics, and that inhibiting ceramide synthesis is sufficient to reverse these insulin-induced changes. We interpreted these mitochondrial effects as detrimental due to the increase in H2O2 production rate and ceramide content, which are more closely associated with pathology than protection in the context of Alzheimer’s disease [20].
These findings are especially relevant due to the current widespread nature of metabolic dysfunction and excess adiposity. One study assessing US trends in obesity over recent decades have demonstrated that over 50% of young adults (ages 18-25) have overweight or obesity [44], which has substantial implications for neurocognitive health. The results presented here suggest that addressing hyperinsulinemia or ceramide synthesis through pharmacological or lifestyle intervention may be effective in alleviating the cognitive burden of obesity and insulin resistance and be protective against AD.
4. Materials and Methods
4.1. Animals
Adult male and female ApoE4 C57BL6 mice (average age of 4.5 months) were group-housed, maintained at 22 ± 1 °C, 60–70% humidity, with a 12-h light–dark cycle, and given ad libitum access to food (LabDiet 5001) and water. Mice were randomly assigned to one of four intraperitoneal injection treatments for 4 weeks: 1. vehicle PBS injections (daily); 2. insulin injections (INS; daily; 0.75 mg/kg); 3. myriocin injections in order to inhibit ceramide synthesis (MYR; thrice weekly; 3 mg/kg); and, 4. insulin and myriocin injections (INS+MYR; as indicated). At the conclusion of the study, brains were removed and cerebral cortex tissue was isolated and processed according to subsequent analysis (see below).
The following animal studies were conducted in accordance with the principles and procedures outlined in the National Institutes of Health Guide for the Care and Use of Laboratory Animals and were approved by the IACUC (Institutional Animal Care and Use Committee) at Brigham Young University. Additionally, experiments have been reported in compliance with the ARRIVE guidelines 2.0 on reporting animal experiments.
4.2. Insulin Tolerance Test
After four weeks conditioning, mice underwent insulin tolerance tests. Mice were fasted six hours then received an intraperitoneal injection of insulin (0.75 unit/kg of body weight). Blood glucose was determined at baseline (week 0), and weekly until sacrifice as indicated in the figures using the Bayer Contour glucose meter (New Jersey, USA).
4.3. Lipid Nalysis
Lipids were analyzed as described previously [45]. To isolate lipids, pellets were suspended in ice-cold chloroform-methanol (1:2), incubated for 15 min on ice, and then briefly vortexed. Aqueous and organic phases were separated by the addition of ice-cold water and chloroform. The organic phase was collected in a fresh vial and dried via vacuum centrifugation (Eppendorf Concentrator Plus). Lipids were then characterized and quantified using shotgun lipidomics on a Thermo ScientificLTQ Orbitrap XL mass spectrometer.
4.4. Mitochondrial Respirometry
High-resolution O2 consumption was determined at 37°C in permeabilized brain tissue using the Oroboros O2K Oxygraph (Innsbruck, Austria) with MiR05 respiration buffer as described previously [46,47]. Respiration was determined by all or parts of the following substrate-uncoupler inhibitor-titration (SUIT) protocol [48]: electron flow through complex I was supported by glutamate malate (10 and 2 mM, respectively) to determine O2 consumption from proton leak (GML). Following stabilization, ADP (2.5 mM) was added to determine oxidative phosphorylation capacity (GMP). Succinate was added (GMSP) for complex III electron flow into the Q-junction. Complex II-supported ETS was then measured by inhibiting complex I with rotenone (Rot; 0.5 M). Last, residual O2 consumption was measured by adding antimycin A (2.5 M) to block complex III action, effectively inhibiting electron flow. This value provides a rate of respiration that is used as a baseline.
4.5. H2O2 Emissions
H2O2 generation was measured using an Amplex Red Hydrogen Peroxide/Peroxidase Assay kit (Molecular Probes; A22188) as described previously [49]. A reaction mixture containing 50 μM Amplex Red and 0.1 unit/mL HRP in KRPG (Krebs-Ringer phosphate glucose) buffer was prepared (145 mM NaCl, 5.7 mM sodium phosphate, 4.86 mM KCl, 0.54 mM CaCl2, 1.22 mM MgSO4 and 5.5 mM glucose). The reaction mixture was pre-warmed in a 96-well plate with 100 μL of mixture per well. A 20 μL aliquot of tissue lysate suspended in KRPG buffer (~1.5 × 104 cells) were added to each well. Samples were incubated for 1 h. Fluorescence was measured with a microplate reader (Molecular Devices; San Jose, CA, USA).
4.6. Statistics
Values are reported as means ± the standard errors of the means. Statistical analyses were conducted by one-way analysis of variance (ANOVA) accounting for the four treatment groups. Student's t tests were used for comparisons between groups within each experiment, and differences were considered statistically significant at P < 0.05.
Author Contributions
Conceptualization, S.T.C. and B.T.B.; Methodology, S.T.C. and B.T.B.; Investigation, S.T.C. and B.T.B.; Formal Analysis, S.T.C. and B.T.B.; Writing—Original Draft Preparation, S.T.C., E.R.S., C.M.W., and B.T.B.; Writing—Review and Editing S.T.C., E.R.S., C.M.W., J.Y.S., C.M.H., C.E.W., A.M.T., and B.T.B.; Project Administration, B.T.B.; Supervision, B.T.B.; Validation, S.T.C. and B.T.B.; Visualization, S.T.C., E.R.S., and B.T.B.; and Funding Acquisition, B.T.B. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by internal university funds.
Institutional Review Board Statement
All animal experiments were performed in accordance with Institutional Animal Care and Use Committee (IACUC; 20-0203) protocols and followed National Institute of Health guidelines for the care and use of laboratory animals. IACUC protocols for all experiments were approved by the Institutional Animal Care and Use Committee of Brigham Young University.
Acknowledgments
We recognize the contributions of Brigham Young University, including the vivarium staff for supporting this research.
Conflicts of Interest
B.T.B. receives royalties from the sale of a book about insulin resistance. All other authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- Shanik, M.H.; Xu, Y.; Skrha, J.; Dankner, R.; Zick, Y.; Roth, J. Insulin resistance and hyperinsulinemia: is hyperinsulinemia the cart or the horse? Diabetes care 2008, 31, S262–S268. [Google Scholar] [CrossRef] [PubMed]
- Saklayen, M.G. The Global Epidemic of the Metabolic Syndrome. Curr Hypertens Rep 2018, 20, 12. [Google Scholar] [CrossRef] [PubMed]
- Roberts, C.K.; Hevener, A.L.; Barnard, R.J. Metabolic syndrome and insulin resistance: underlying causes and modification by exercise training. Compr Physiol 2013, 3, 1–58. [Google Scholar] [CrossRef] [PubMed]
- Katz, S.L.; MacLean, J.E.; Hoey, L.; Horwood, L.; Barrowman, N.; Foster, B.; Hadjiyannakis, S.; Legault, L.; Bendiak, G.N.; Kirk, V.G.; et al. Insulin Resistance and Hypertension in Obese Youth With Sleep-Disordered Breathing Treated With Positive Airway Pressure: A Prospective Multicenter Study. J Clin Sleep Med 2017, 13, 1039–1047. [Google Scholar] [CrossRef]
- Bornfeldt, K.E.; Tabas, I. Insulin resistance, hyperglycemia, and atherosclerosis. Cell Metab 2011, 14, 575–585. [Google Scholar] [CrossRef] [PubMed]
- Crone, C. Facilitated transfer of glucose from blood into brain tissue. The Journal of physiology 1965, 181, 103. [Google Scholar] [CrossRef] [PubMed]
- Hom, F.G.; Goodner, C.J.; Berrie, M.A. A (3H) 2-deoxyglucose method for comparing rates of glucose metabolism and insulin responses among rat tissues in vivo: validation of the model and the absence of an insulin effect on brain. Diabetes 1984, 33, 141–152. [Google Scholar] [CrossRef]
- Gerozissis, K. Brain insulin: regulation, mechanisms of action and functions. Cellular and molecular neurobiology 2003, 23, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Mueckler, M. Facilitative glucose transporters. European journal of biochemistry 1994, 219, 713–725. [Google Scholar] [CrossRef]
- Havrankova, J.; Roth, J.; BROWNSTEIN, M. Insulin receptors are widely distributed in the central nervous system of the rat. Nature 1978, 272, 827–829. [Google Scholar] [CrossRef]
- Hopkins, D.; Williams, G. Insulin receptors are widely distributed in human brain and bind human and porcine insulin with equal affinity. Diabetic Medicine 1997, 14, 1044–1050. [Google Scholar] [CrossRef]
- Chiu, S.-L.; Chen, C.-M.; Cline, H.T. Insulin receptor signaling regulates synapse number, dendritic plasticity, and circuit function in vivo. Neuron 2008, 58, 708–719. [Google Scholar] [CrossRef]
- 2022 Alzheimer's disease facts and figures. Alzheimer's & Dementia : the journal of the Alzheimer's Association 2022.
- Neth, B.J.; Craft, S. Insulin Resistance and Alzheimer's Disease: Bioenergetic Linkages. Front Aging Neurosci 2017, 9, 345. [Google Scholar] [CrossRef]
- Kuusisto, J.; Koivisto, K.; Mykkanen, L.; Helkala, E.L.; Vanhanen, M.; Hanninen, T.; Kervinen, K.; Kesaniemi, Y.A.; Riekkinen, P.J.; Laakso, M. Association between features of the insulin resistance syndrome and Alzheimer's disease independently of apolipoprotein E4 phenotype: cross sectional population based study. BMJ 1997, 315, 1045–1049. [Google Scholar] [CrossRef]
- Sampath, D.; Sathyanesan, M.; Newton, S.S. Cognitive dysfunction in major depression and Alzheimer's disease is associated with hippocampal-prefrontal cortex dysconnectivity. 2017. [CrossRef]
- Sokolowska, E.; Blachnio-Zabielska, A. The role of ceramides in insulin resistance. Frontiers in Endocrinology 2019, 10, 577. [Google Scholar] [CrossRef]
- Hammerschmidt, P.; Brüning, J.C. Contribution of specific ceramides to obesity-associated metabolic diseases. Cellular and Molecular Life Sciences 2022, 79, 395. [Google Scholar] [CrossRef] [PubMed]
- Suzanne, M. Insulin resistance and Alzheimer’s disease. BMB reports 2009, 42, 475. [Google Scholar]
- Filippov, V.; Song, M.A.; Zhang, K.; Vinters, H.V.; Tung, S.; Kirsch, W.M.; Yang, J.; Duerksen-Hughes, P.J. Increased ceramide in brains with Alzheimer's and other neurodegenerative diseases. J Alzheimers Dis 2012, 29, 537–547. [Google Scholar] [CrossRef] [PubMed]
- Jazvinšćak Jembrek, M.; Hof, P.R.; Šimić, G. Ceramides in Alzheimer’s disease: key mediators of neuronal apoptosis induced by oxidative stress and Aβ accumulation. Oxidative medicine and cellular longevity 2015, 2015. [Google Scholar] [CrossRef]
- Shubhra Chakrabarti, S.; Bir, A.; Poddar, J.; Sinha, M.; Ganguly, A.; Chakrabarti, S. Ceramide and sphingosine-1-phosphate in cell death pathways: relevance to the pathogenesis of Alzheimer's disease. Current Alzheimer Research 2016, 13, 1232–1248. [Google Scholar] [CrossRef] [PubMed]
- Lyn-Cook Jr, L.E.; Lawton, M.; Tong, M.; Silbermann, E.; Longato, L.; Jiao, P.; Mark, P.; Wands, J.R.; Xu, H.; de la Monte, S.M. Hepatic Ceramide May Mediate Brain Insulin Resistance and Neurodegeneration in Type 2 Diabetes and Non-alcoholic Steatohepatitis. Journal of Alzheimer's Disease 2009, 16, 715–729. [Google Scholar] [CrossRef] [PubMed]
- Cutler, R.G.; Kelly, J.; Storie, K.; Pedersen, W.A.; Tammara, A.; Hatanpaa, K.; Troncoso, J.C.; Mattson, M.P. Involvement of oxidative stress-induced abnormalities in ceramide and cholesterol metabolism in brain aging and Alzheimer's disease. Proceedings of the National Academy of Sciences 2004, 101, 2070–2075. [Google Scholar] [CrossRef] [PubMed]
- Hansen, M.E.; Tippetts, T.S.; Anderson, M.C.; Holub, Z.E.; Moulton, E.R.; Swensen, A.C.; Prince, J.T.; Bikman, B.T. Insulin increases ceramide synthesis in skeletal muscle. J Diabetes Res 2014, 2014, 765784. [Google Scholar] [CrossRef]
- Hodson, A.E.; Tippetts, T.S.; Bikman, B.T. Insulin treatment increases myocardial ceramide accumulation and disrupts cardiometabolic function. Cardiovasc Diabetol 2015, 14, 153. [Google Scholar] [CrossRef]
- Luchsinger, J.A.; Tang, M.-X.; Shea, S.; Mayeux, R. Hyperinsulinemia and risk of Alzheimer disease. Neurology 2004, 63, 1187–1192. [Google Scholar] [CrossRef] [PubMed]
- Gatz, M.; Reynolds, C.A.; Fratiglioni, L.; Johansson, B.; Mortimer, J.A.; Berg, S.; Fiske, A.; Pedersen, N.L. Role of genes and environments for explaining Alzheimer disease. Archives of general psychiatry 2006, 63, 168–174. [Google Scholar] [CrossRef]
- Sienski, G.A.-O.; Narayan, P.A.-O.; Bonner, J.A.-O.; Kory, N.A.-O.; Boland, S.A.-O.; Arczewska, A.A.; Ralvenius, W.A.-O.; Akay, L.A.-O.; Lockshin, E.A.-O.; He, L.; et al. APOE4 disrupts intracellular lipid homeostasis in human iPSC-derived glia. 2021. [CrossRef]
- Chaurasia, B.; Summers, S.A. Ceramides–lipotoxic inducers of metabolic disorders. Trends in Endocrinology & Metabolism 2015, 26, 538–550. [Google Scholar]
- Hodson, A.E.; Tippetts, T.S.; Bikman, B.T. Insulin treatment increases myocardial ceramide accumulation and disrupts cardiometabolic function. Cardiovascular Diabetology 2015, 14, 153. [Google Scholar] [CrossRef]
- Pontiroli, A.E.; Alberetto, M.; Pozza, G. Patients with insulinoma show insulin resistance in the absence of arterial hypertension. Diabetologia 1992, 35, 294–295. [Google Scholar] [CrossRef]
- Henry, R.R.; Gumbiner, B.; Ditzler, T.; Wallace, P.; Lyon, R.; Glauber, H.S. Intensive Conventional Insulin Therapy for Type II Diabetes: Metabolic effects during a 6-mo outpatient trial. Diabetes Care 1993, 16, 21. [Google Scholar] [CrossRef]
- Del Prato, S.; Leonetti, F.; Simonson, D.C.; Sheehan, P.; Matsuda, M.; DeFronzo, R.A. Effect of sustained physiologic hyperinsulinaemia and hyperglycaemia on insulin secretion and insulin sensitivity in man. Diabetologia 1994, 37, 1025–1035. [Google Scholar] [CrossRef] [PubMed]
- Bikman, B.T.; Summers, S.A. Ceramides as modulators of cellular and whole-body metabolism. The Journal of clinical investigation 2011, 121, 4222–4230. [Google Scholar] [CrossRef] [PubMed]
- Hussain, G.; Wang, J.; Rasul, A.; Anwar, H.; Imran, A.; Qasim, M.; Zafar, S.; Kamran, S.K.S.; Razzaq, A.; Aziz, N.; et al. Role of cholesterol and sphingolipids in brain development and neurological diseases. Lipids in Health and Disease 2019, 18, 26. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Nevado-Holgado, A.; Whiley, L.; Snowden, S.G.; Soininen, H.; Kloszewska, I.; Mecocci, P.; Tsolaki, M.; Vellas, B.; Thambisetty, M. Association between plasma ceramides and phosphatidylcholines and hippocampal brain volume in late onset Alzheimer’s disease. Journal of Alzheimer's Disease 2017, 60, 809–817. [Google Scholar] [CrossRef]
- Mielke, M.M.; Bandaru, V.V.R.; Haughey, N.J.; Rabins, P.V.; Lyketsos, C.G.; Carlson, M.C. Serum sphingomyelins and ceramides are early predictors of memory impairment. Neurobiology of aging 2010, 31, 17–24. [Google Scholar] [CrossRef]
- Mielke, M.M.; Haughey, N.J.; Bandaru, V.V.R.; Schech, S.; Carrick, R.; Carlson, M.C.; Mori, S.; Miller, M.I.; Ceritoglu, C.; Brown, T. Plasma ceramides are altered in mild cognitive impairment and predict cognitive decline and hippocampal volume loss. Alzheimer's & Dementia 2010, 6, 378–385. [Google Scholar]
- McGrath, E.R.; Himali, J.J.; Xanthakis, V.; Duncan, M.S.; Schaffer, J.E.; Ory, D.S.; Peterson, L.R.; DeCarli, C.; Pase, M.P.; Satizabal, C.L. Circulating ceramide ratios and risk of vascular brain aging and dementia. Annals of clinical and translational neurology 2020, 7, 160–168. [Google Scholar] [CrossRef]
- Czubowicz, K.; Jęśko, H.; Wencel, P.; Lukiw, W.J.; Strosznajder, R.P. The role of ceramide and sphingosine-1-phosphate in Alzheimer’s disease and other neurodegenerative disorders. Molecular neurobiology 2019, 56, 5436–5455. [Google Scholar] [CrossRef]
- Kurz, J.; Parnham, M.J.; Geisslinger, G.; Schiffmann, S. Ceramides as novel disease biomarkers. Trends in molecular medicine 2019, 25, 20–32. [Google Scholar] [CrossRef]
- Teitsdottir, U.D.; Halldorsson, S.; Rolfsson, O.; Lund, S.H.; Jonsdottir, M.K.; Snaedal, J.; Petersen, P.H. Cerebrospinal fluid C18 ceramide associates with markers of Alzheimer’s disease and inflammation at the pre-and early stages of dementia. Journal of Alzheimer's Disease 2021, 81, 231–244. [Google Scholar] [CrossRef]
- Ellison-Barnes, A.; Johnson, S.; Gudzune, K. Trends in obesity prevalence among adults aged 18 through 25 years, 1976-2018. JAMA 2021, 326, 2073–2074. [Google Scholar] [CrossRef] [PubMed]
- Hansen, M.; Tippetts, T.S.; Anderson, M.; Holub, Z.; Moulton, E.; Swensen, A.; Prince, J.; Bikman, B. Insulin increases ceramide synthesis in skeletal muscle. Journal of Diabetes Research 2014, 2014. [Google Scholar] [CrossRef]
- Pesta, D.; Gnaiger, E. High-Resolution Respirometry: OXPHOS Protocols for Human Cells and Permeabilized Fibers from Small Biopsies of Human Muscle. In Mitochondrial Bioenergetics: Methods and Protocols; Palmeira, C.M., Moreno, A.J., Eds.; Humana Press: Totowa, NJ, 2012; pp. 25–58. [Google Scholar]
- Smith, M.E.; Tippetts, T.S.; Brassfield, E.S.; Tucker, B.J.; Ockey, A.; Swensen, A.C.; Anthonymuthu, T.S.; Washburn, T.D.; Kane, D.A.; Prince, J.T.; et al. Mitochondrial fission mediates ceramide-induced metabolic disruption in skeletal muscle. Biochemical Journal 2013, 456, 427. [Google Scholar] [CrossRef] [PubMed]
- Jheng, H.-F.; Tsai, P.-J.; Guo, S.-M.; Kuo, L.-H.; Chang, C.-S.; Su, I.-J.; Chang, C.-R.; Tsai, Y.-S. Mitochondrial Fission Contributes to Mitochondrial Dysfunction and Insulin Resistance in Skeletal Muscle. Molecular and Cellular Biology 2012, 32, 309–319. [Google Scholar] [CrossRef] [PubMed]
- Napa, K.; Baeder, A.C.; Witt, J.E.; Rayburn, S.T.; Miller, M.G.; Dallon, B.W.; Gibbs, J.L.; Wilcox, S.H.; Winden, D.R.; Smith, J.H.; et al. LPS from P. gingivalis Negatively Alters Gingival Cell Mitochondrial Bioenergetics. Int J Dent 2017, 2017, 2697210. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Chronic Insulin Injections Increase Body Weight and Fasting Glucose Levels and Insulin Resistance. Male and female ApoE4 mice received injections of PBS (daily), insulin (INS; daily; 0.75 mg/kg), myriocin (MYR, thrice weekly; 3 mg/kg), or INS+MYR. Body weight (a) was tracked weekly. Fasting glucose levels (b) and insulin tolerance (c) were measured after 28 days of treatment. N=5. *P<0.05 and **P<0.01 for INS vs. other treatments. #P<0.05 for INS vs. INS+MYR.
Figure 1.
Chronic Insulin Injections Increase Body Weight and Fasting Glucose Levels and Insulin Resistance. Male and female ApoE4 mice received injections of PBS (daily), insulin (INS; daily; 0.75 mg/kg), myriocin (MYR, thrice weekly; 3 mg/kg), or INS+MYR. Body weight (a) was tracked weekly. Fasting glucose levels (b) and insulin tolerance (c) were measured after 28 days of treatment. N=5. *P<0.05 and **P<0.01 for INS vs. other treatments. #P<0.05 for INS vs. INS+MYR.
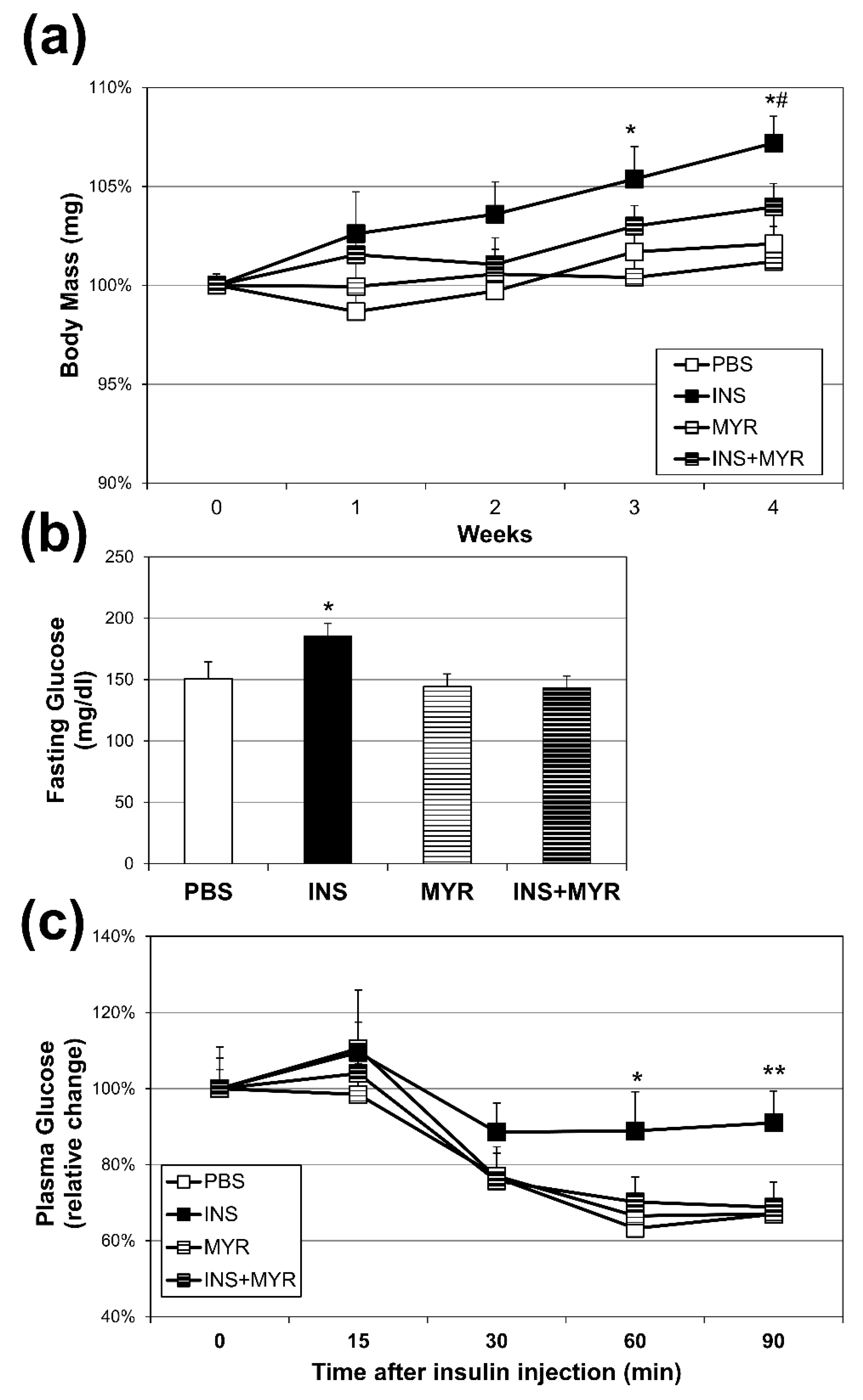
Figure 2.
Hyperinsulinemia Increases Ceramides in the ApoE4 Brain. Male and female ApoE4 mice received injections of PBS (daily), insulin (INS; daily; 0.75 mg/kg), myriocin (MYR, thrice weekly; 3 mg/kg), or INS+MYR. Following the 28-day treatment, lipids were isolated from the cerebral cortex for analysis of sphingolipids via liquid chromatography-mass spectrometry. N=5. *P<0.05 and **P<0.01 for treatment vs. PBS. #P<0.05 for treatment vs. INS.
Figure 2.
Hyperinsulinemia Increases Ceramides in the ApoE4 Brain. Male and female ApoE4 mice received injections of PBS (daily), insulin (INS; daily; 0.75 mg/kg), myriocin (MYR, thrice weekly; 3 mg/kg), or INS+MYR. Following the 28-day treatment, lipids were isolated from the cerebral cortex for analysis of sphingolipids via liquid chromatography-mass spectrometry. N=5. *P<0.05 and **P<0.01 for treatment vs. PBS. #P<0.05 for treatment vs. INS.
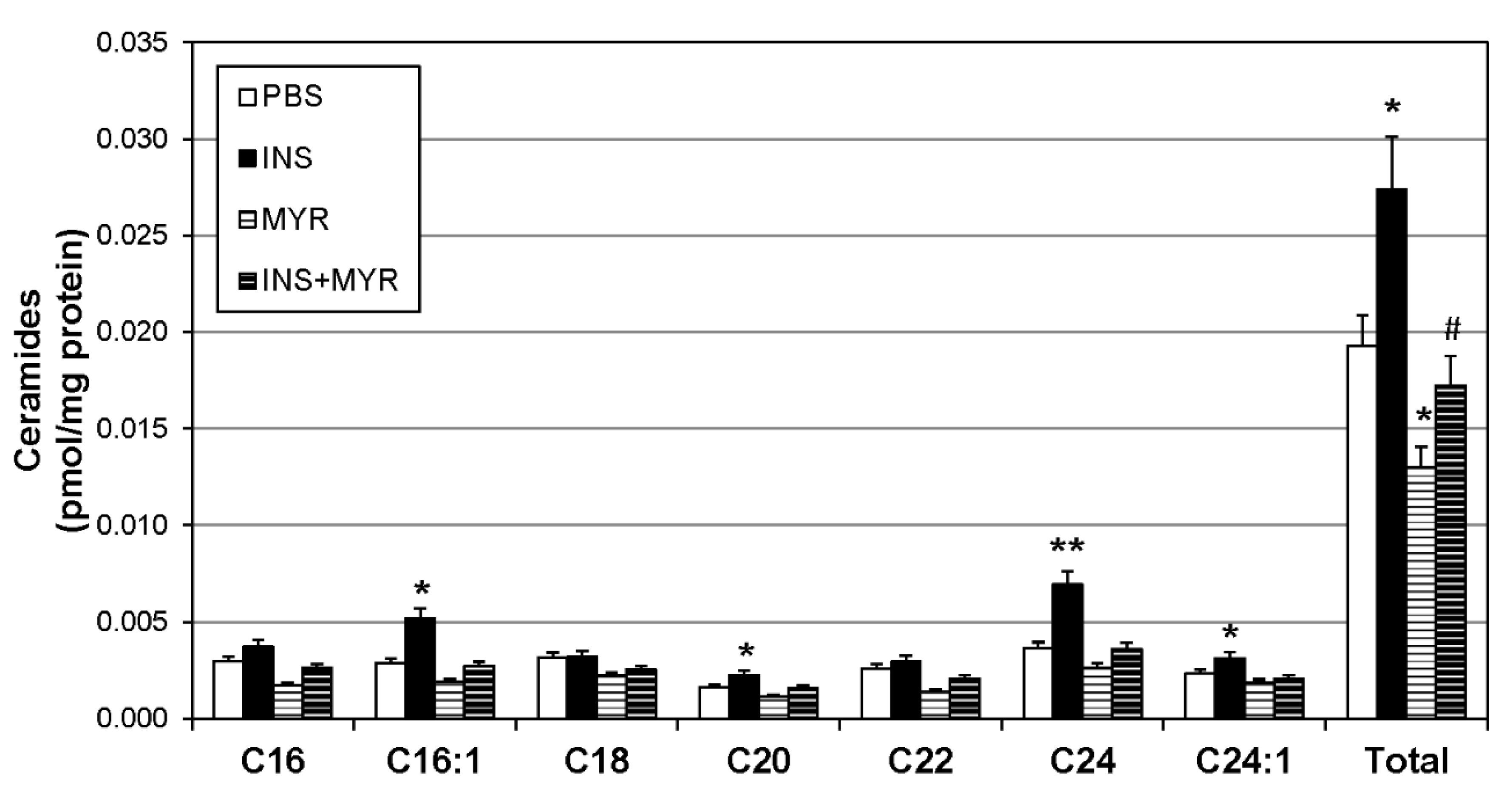
Figure 3.
Hyperinsulinemia Compromises Brain Mitochondrial Function. Male and female ApoE4 mice received injections of PBS (daily), insulin (INS; daily; 0.75 mg/kg), myriocin (MYR, thrice weekly; 3 mg/kg), or INS+MYR. High-resolution respirometry on cerebral cortex tissue was performed (a) using GML (glutamate (10 mM) + malate (2 mM)); GMp: (ADP (2.5 mM)); GMSp (succinate (10 mM)). Respiratory control ratio (RCR; b) and complex II factor (c) were determined, indicated in the Methods. N=5. *P<0.05 and **P<0.01 for treatment vs. PBS. #P<0.05 for treatment vs. INS.
Figure 3.
Hyperinsulinemia Compromises Brain Mitochondrial Function. Male and female ApoE4 mice received injections of PBS (daily), insulin (INS; daily; 0.75 mg/kg), myriocin (MYR, thrice weekly; 3 mg/kg), or INS+MYR. High-resolution respirometry on cerebral cortex tissue was performed (a) using GML (glutamate (10 mM) + malate (2 mM)); GMp: (ADP (2.5 mM)); GMSp (succinate (10 mM)). Respiratory control ratio (RCR; b) and complex II factor (c) were determined, indicated in the Methods. N=5. *P<0.05 and **P<0.01 for treatment vs. PBS. #P<0.05 for treatment vs. INS.
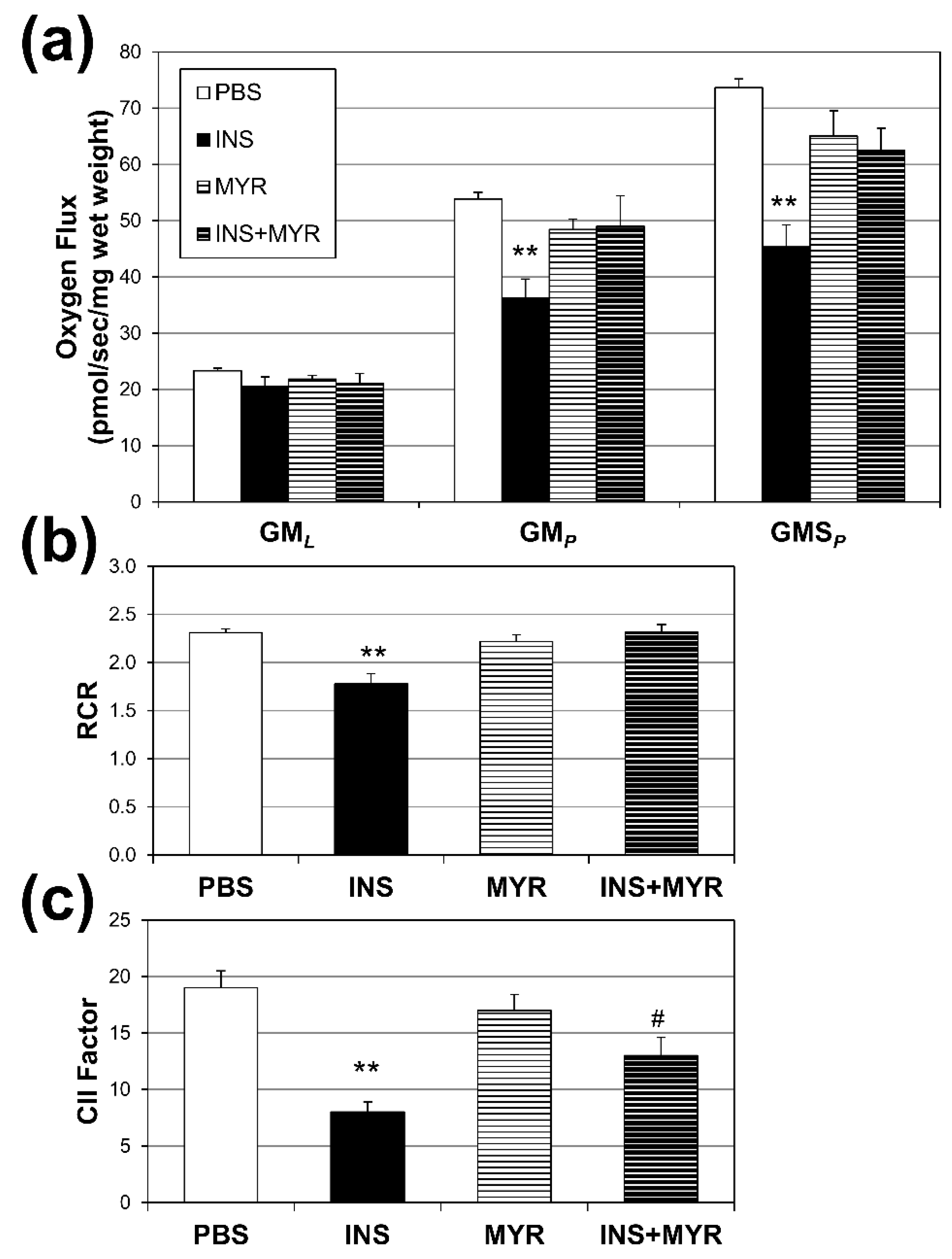
Figure 4.
Chronic Insulin Injections Increase Brain H2O2 production. Male and female ApoE4 mice received injections of PBS (daily), insulin (INS; daily; 0.75 mg/kg), myriocin (MYR, thrice weekly; 3 mg/kg), or INS+MYR for 28 days. Cerebral cortex lysate was used to determine fluorescence using Amplex Red (a) and the ratio of H2O2 to O2 consumed was determined (b). *P<0.05 and **P<0.01 for INS vs. other treatments. #P<0.05 for INS+MYR vs. PBS.
Figure 4.
Chronic Insulin Injections Increase Brain H2O2 production. Male and female ApoE4 mice received injections of PBS (daily), insulin (INS; daily; 0.75 mg/kg), myriocin (MYR, thrice weekly; 3 mg/kg), or INS+MYR for 28 days. Cerebral cortex lysate was used to determine fluorescence using Amplex Red (a) and the ratio of H2O2 to O2 consumed was determined (b). *P<0.05 and **P<0.01 for INS vs. other treatments. #P<0.05 for INS+MYR vs. PBS.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



