
Submitted:
12 September 2023
Posted:
13 September 2023
You are already at the latest version
Abstract
The study investigated the effect of a multi-strain probiotic compound containing Bacillus coagulans, Enterococcus faecalis, Clostridium butyricum and Bacillus mesentericus as in-feed zinc oxide (ZnO) alterna-tive on growth performance, diarrhea incidence, antioxidant status, lipid profile, stress and immunity in weaned piglets. A total of 72 piglets were randomly divided into 3 groups with four replicates of six piglets each. The details of the groups were as follows; (i) weaned control group (WC) received basal diet, (ii) weaned probiotic group (WB) received basal diet and probiotics and (iii) positive control (PC) group received basal diet with 2500 mg/kg ZnO. The experiment lasted for 28 days. Probiotic supple-mentation improved growth performance and reduced diarrhea rate. Probiotics supplementation im-proved lipid profile; significantly lower levels of total cholesterol and low-density lipoprotein cholesterol and higher level of high-density lipoprotein cholesterol in WB group as compared to those of the control group (WC) were recorded. Probiotic supplementation stimulated antioxidant defense system by in-creasing total antioxidant capacity and decreasing lipid peroxidation. Probiotic supplementation down-regulated the stress biomarkers like serum cortisol and serum heat shock proteins. WB group showed higher serum levels of IgG and IgM throughout the study period and higher IgA at day 28 as compared to WC. These data suggest that supplementation of the probiotic minimizes the weaning stress, thereby improves the growth performance, lipid profile, antioxidant status and systemic as well as mucosal immunity. Therefore, the multi-strain probiotic compound may be used to replace ZnO in weaned piglets.
Keywords:
Probiotics
; pigs
; growth performance
; lipid profile
; oxidative stress
; cytokines
; immunity
1. Introduction
In modern intensive farming system, weaning is practiced to enhance the breeding efficiency of sows and economic profit of the farm [1]. In modern farms, it is general practice to wean piglets at three to four weeks of age [2]. Weaning, though a standard management practice, is a stressful and traumatic event for piglets due to sudden dietary, social and environmental changes [3]. Such periods of multiple stressors are linked to severe enteric infection, diarrhea, reduced feed conversion efficiency, loss of weight and death in extreme cases [4,5] leading to enormous economic loss to swine industry. At weaning, as the digestive capacity of a piglet is poor, opportunistic pathogens (mainly Escherichia coli and Salmonella) residing at the gastrointestinal tract, ferment undigested feed materials and generate toxic metabolites which damage the intestinal mucosa and ultimately results in diarrhea and poor performance of the piglet [3,6]. Moreover, stressors associated with weaning disrupt or weaken antioxidant defense system of the piglets, making them more prone to stress and infection [7]. Sometimes in commercial farming, piglets are weaned at only one or two weeks of age which amplifies the detrimental effects of weaning [8]. Therefore, reduction of weaning stress is extremely important and key to profitable pig farming.
In-feed administration of antibiotics and zinc oxide (ZnO) have been widely used to combat post-weaning diarrhea and growth improvement in piglets [9,10]. However, use of some antibiotics as growth promoters has been banned in the European Union, China, Japan and Korea due to increasing occurrence of antibiotic resistance in animals as well as in human, the ultimate consumer of the animal produce [11,12,13]. In recent years, concern over use of ZnO has been raised as extensive use of ZnO is linked to environmental heavy metal contamination [10]. Moreover, development of antibiotic-resistant microorganisms is a common consequence of application of high doses of dietary ZnO [14,15]. Considering the negative consequences of in-feed ZnO administration on environment and public health, the European Union has recommended to phase out medicinal application of ZnO in pig production by 2022 [16].
Consequently, in the recent past, much emphasis has been paid to find out a safe and practical alternative to ZnO to alleviate weaning stress and to maintain swine health and performance. Among the strategies that have been proposed, probiotics supplementation has proven to be an effective alternative to in-feed ZnO use, because of its potential to stimulate the intestinal immune system, antioxidant status, nutrient digestibility and to increase the production of antimicrobial peptides and cytokines in the intestinal tract [16,17,18]. Lactic acid bacteria (LAB) which include a variety of bacterial genera like, Lactobacillus, Bacillus, Bifidobacterium, Streptococcus, Enterococcus, and some other microbes, are the most frequently used microorganism as probiotic agents [19]. Among these LAB, spore-forming Bacillus spp. have been considered the most promising as their spores can endure hostile environments and allow for extensive storage at room temperature [20].
The effectiveness of a probiotic is highly strain specific; some strains provide more benefits to the host than others [21]. B. coagulans alone [22] or in combination with E. faecalis and C. butyricum [23] was found beneficial to weaning piglets. However, the combined effect of B. coagulans, E. faecalis, C. butyricum and B. mesentericus on weaning piglets has not been investigated extensively. The study was designed with the hypothesis that the multi-strain probiotic compound could be an alternative to ZnO administration in alleviation of weaning stress in piglets. Therefore, the objective of the current study was to investigate the combined effect of multi-strain probiotic supplementation including B. mesentericus TO-A, B. coagulans SNZ1969, C. butyricum TO-A and E. faecalis T-110on growth performance, lipid metabolites, antioxidant defense system and serum cytokine profiles of piglets at weaning in an island ecosystem.
2. Materials and Methods
2.1. Experimental area
The present study was conducted at the ICAR-CIARI institute pig farm, Port Blair, South Andaman, a district of Andaman and Nicobar Islands (ANI). The ANI, an archipelago made up of 572 islands and islets, is situated (Lat. 6° to 14° North and Lon. 92° to 94° East) in the meeting point of the Andaman Sea (East Side) and the Bay of Bengal (West Side). It has a total surface area of 8249 sq. km. and a 1,962 km. coastline.
2.2. Experimental Period
The current work was conducted during January to February, 2022. The highest and lowest air temperatures during the study period were 33.5 °C and 21.8 °C, respectively. The average relative humidity ranged from 64.0 % to 79.5 %. Temperature Humidity Index (THI) varied from 75.6 to 81.5 during the study.
2.3. Experimental Animals
The experiment was conducted on indigenous Andaman local pig (ALP). ALP is generally reared by the tribal farmers of these islands on semi-intensive system of management. Under intensive condition, they perform extremely well and attains market weight of 65-70 kg at the age of 9 months. They provide livelihood and nutritional security to the farmers particularly the tribal farmers of ANI.
2.4. Source of the Probiotic
The probiotic (BIFILAC) used in the present study was purchased from a commercial company (Tablets India Limited, Chennai, India). It is a multi-strain probiotic and contains four microorganisms; the details are presented in Table 1. The probiotic formulation was reported to safe and offers no side effect [24].
2.5. Study design
The study included 72 clinically healthy piglets (ALP, weaned at 27 ± 1 day) with initial average body weight of 8.77 ± 0.15 (mean ± SD, kg). The piglets were randomly divided into 3 groups according to sex and body weight with 24 animals in each group (6 piglets per pen; 4 pens/replicates per treatment). Each pen (1.5 x 1.3 m2) with six animals (3 males and 3 females) was considered as an experimental unit. Throughout the trial, each pen had access to ad libitum feed and drinking water. the pens have concrete floors and height of 0.7 meter which allow adequate natural ventilation. The piglets were grouped as follows; (a) weaned negative control group (WC) received basal diet without probiotics, (b) positive control (PC) group received basal diet with 2500 mg/kg ZnO as reported previously [25] and (c) weaned probiotic group (WB) received basal diet with probiotics (0.1 % with feed). The dose of the probiotics was standardized using a pilot study in which the above-mentioned dose was found most effective in promotion of growth and in alleviation of weaning stress. The composition of the basal diet is provided in Table 2. Probiotics and ZnO were mixed with experimental diets using a feed miller. The experiment was for 28 days.
2.6. Estimation of Diarrhea Rate of Piglets
For estimation of diarrhea incidence, feces of piglets were examined visually every morning and afternoon. Scoring of the feces and severity of diarrhea were done as per procedure recommended by Walsh et al. [27] as following; 1 = hard feces; 2 = slightly soft feces; 3 = soft, partially formed feces; 4 = loose, semi-liquid feces; and 5 = watery, mucous-like feces. Fecal consistency score of grade 4 to 5 for 2 consecutive days was considered as diarrhea. Diarrhea rate was calculated as previously described [28].
2.7. Production Parameters
All the piglets in each group were weighed individually on day 0, 7, 14, 21 and 28 with an electronic balance. Daily feed consumption was recorded throughout the study. Average daily feed intake (ADFI), average daily gain (ADG), and feed to gain ratio (F:G) were calculated using standard methodologies.
2.8. Blood Sampling
Each piglet of every group was bled (∼ 10 ml) on 0 day, and weekly interval thereafter up to forth week from cranial vena cava into a vacutainer containing clot activator (Hebei Xinle Sci &Tech Co., Ltd. Hebei Province, China), following standard aseptic condition. Serum was separated by keeping the tubes at room temperature for 30 minutes followed by centrifuged at 1200 × g for 10 minutes at 4 °C. Serum was stored at -80 °C till further use.
2.9. Lipid Profile Analysis
Serum lipid profile including concentration of total cholesterol (TC), triglycerides (TG) and high-density lipoprotein cholesterol (HDLc) were determined by enzymatic methods using commercially available kits (Jeev Diagnostics Pvt. Ltd., Chennai, India; Spinreact, S.A., Spain and Pathozyme Diagnostics, Kholapur, India, respectively). The concentration of LDLc was determined as reported previously as: LDLc = TC – HDLc – (TG/5) [29]. Additionally, cardiac risk factor (CRF = TC/HDLc) and atherogenic index (AI= (TC-HDLc)/HDLc were also evaluated as previously described [30,31]. Lipid profile was measured on day 0, 7, 14 and 28; CRF and AI were calculated for day 14 and 28.
Total antioxidant activity (T-AOC) of serum samples were assessed by commercial kit brought from HiMedia laboratories company (Mumbai, India). Serum T-AOC was detected with reduction of Cu (II)-chromogen complex to Cu (I) complex and absorbance was measured at 460 nm.
Serum activities of glutathione -S-transferase (GSH), superoxide dismutase (SOD), catalase were estimated by commercially available kits brought from Cayman chemicals company (Ann Arbor, Michigan, USA).
Serum malonyldialdehyde (MDA) levels were measured to determine the degree of lipid peroxidation as previously described [30]. The MDA concentration of serum samples were estimated with 2-thiobarbituric acid and the variations in absorbance were read at 534 nm.
T-AOC and MDA were measured at day 0 and weekly interval thereafter throughout the study whereas SOD, catalase and GSH levels were evaluated on day 0, 7, 14 and 28.
2.10. Measurement of Stress Biomarkers
Total nitric oxide (TNO) and heat shock proteins (HSPs) were observed at day 0 and at weekly interval till the end of experiment, whereas serum cortisol concentration was measured on day 0, 7, 14 and 28.
2.11. Antioxidant and Oxidative Profile
2.11.1. Nitric Oxide Assay
TNO level was analyzed by commercially available NO Estimation kit (HiMedia laboratories, Mumbai, India).
2.11.2. Serum Cortisol Assay
Level of cortisol in serum was measured by commercial cortisol detection kit by Arsh Biotech company (Life Technologies, Delhi, India) using biotin double antibody sandwich technology.
2.11.3. Determination of serum HSPs
Serum heat shock proteins (HSP90, HSP70, HSP40, and HSP20) were determined by commercial double antibody sandwich ELISA kits by Arsh Biotech company (Life Technologies, Delhi, India).
2.12. Immune Parameters
Serum immunoglobulin concentrations including IgM, IgG and IgA were determined by commercial kits brought from Arsh Biotech company (Delhi, India).
Serum interleukin concentrations including IL-1β, IL-2, IL-4, IL-6, IFN-γ and IL-12 were determined by porcine ELISA based kits brought from Arsh Biotech company (Life Technologies, Delhi, India). All the parameters were evaluated on day 0, 7, 14 and 28.
2.13. Statistical Analysis
Before analysis, Shapiro-Wilk statistics were performed to check the data for normality. Data had a homoscedastic distribution and a normal shape. One-way repeated measures analysis of variance (ANOVA) or a within-subjects ANOVA was applied to determine the significant differences be-tween groups at a particular time point in GraphPad Prism software (http://www.graphpad.com). Pen was served as an experimental unit in analysis of diarrhea rate, F:G and ADFI while, each pig was treated as the experimental unit for other experimental parameters of this study. The analyzed data of each parameter was presented as mean (M) ± standard deviation (SD). The statistical significance was defined as the mean values with a significance level of p < 0.05.
3. Results
3.1. Production Parameters and Diarrhea Incidence
Effect of currently used probiotic formulation on production parameters and diarrhea incidences are presented in Table 3. No differences in body weights among three groups at day 0 and day 7 were observed. Thereafter, significantly higher body weights in WB and PC groups as compared to WC group were recorded on day 14, day 21 and day 28. But, it did not show any changes between WB and PC group. WB and PC groups showed greater average daily weight gain on 0-14 day and 14-28 days as compared to those of WC group. Moreover, the overall ADG (0-28 D) of WC group was significantly lower than WB and PC group. Significantly ADFI in PC as compared to WC was recorded for 0-14 D whereas it did not vary between WB and PC group. Overall ADFI (0-28 D) did not differ significantly among the groups. WB and PC groups showed significantly lower feed to gain ration (F:G) as compared to the control group (WC) for 0-14 D, 14-28 D and overall (0-28 D). The diarrhea rates in piglets supplemented with the probiotic compound (WB) or in-feed ZnO (PC) were lower than that of the negative control group (WC).
3.2. Lipid Panel Analysis
TC did not vary significantly among the groups up to day 14 (Figure 1a). At day 28, TC was significantly greater in WC group than those of other two groups (WB and PC), whereas WB and PC did not record any difference.
Down-regulated HDLc concentration (Figure 1b) was observed in WC group as compared to WB and PC groups on day 14 and 28 whereas the concentration was higher in WB group than PC group on day 28.
At day 7 and day 14, TG concentration in WC group was significantly greater as compared to other two group (WB and PC), whereas, it did not vary between WB and PC group (Figure 1c). Day 28 recorded no significant difference in TG concentration among the groups.
Regarding LDLc concentration (Figure 1d), significantly higher value on Day 28 was observed in WC as compared to the other two groups, whereas, WB and PC was unchanged. On the other time points, there was no significant change among the groups.
3.3. Antioxidant Profiles and Oxidative Stress Indicators
Significantly lower T-AOC (Figure 2a) in WC group than that of other two groups (WB, PC) was observed throughout the study period, whereas no significant difference between WB and PC was detected.
Activities of SOD (Figure 2b), catalase (Figure 2c) and concentrations of GSH (Figure 2d) in PC and WB were significantly greater than the WC group on both day 14 and 28, whereas it did not vary among the two groups (WB and PC) except for SOD and catalase at day 7, in which both were found higher in PC group than those of WB group.
At every week under consideration, the WC group showed significantly greater MDA concentration than WB and PC group. No significant difference in MDA concentration between WB and PC was observed throughout the study period except at day 7 in which WB group showed significantly higher MDA concentration than PC group (Figure 2e).
3.4. Evaluation of Stress Parameters
3.4.1. Total Serum Nitric Oxide Concentration
WC group showed significantly higher serum TNO concentration (Figure 3a) on day 7 and 14 in comparison to WC and PC group whereas, there was no significant difference among the three groups on day 21 and 28.
3.4.2. Serum Cortisol
Significantly higher cortisol concentrations (Figure 3b) in WC and WB as compared that of PC were recorded on day 7. At day 14 and 28, significantly reduced concentrations of cortisol in WB and PC groups as compared to WC group was observed while, there was no significant difference between WB and PC on those days.
3.4.3. Serum Heat Shock Proteins (HSPs)
Serum concentrations of four HSPs (HSP20, HSP70, HSP40 and HSP90) were evaluated (Figure 4) in this study. Concentrations of all four HSP isoforms in WC group were found up-regulated in all the time points as compared to those of WB and PC groups except for HSP20 and HSP70 on day 7 and HSP90 on day 28. HSP20 and HSP70 levels didn't vary significantly between WC and WB at day 7. Concentrations of all four HSPs between WB and PC groups were found insignificant in all the time points.
3.5. Immune Parameters of Serum
WB group recorded significantly greater levels of IgM and IgG on day 14 and 28 and IgA on day 28 than that of the other two groups, whereas the values in the WC group were found significantly lower in all the time points for IgM and IgG and day 7 for IgA than those of other two groups (Figure 5a-c).
On day 14 and day 28, IL-1β, IL-2, IL-6, IFN-γ and IL-12 concentrations were lower in WB and PC group than the WC group, while it did not vary among WB and PC group. WC showed reduced serum IL-4 concentrations at day 7 and day 14 as compared to the other two groups. At day 28, in WB group showed significantly greater concentration of IL-4 than the other two groups (WC and PC) (Figure 5d-i).
4. Discussion
Weaning is the most stressful event a piglet [3]. Weaned piglets must adapt to the stressful condition rapidly to increase their growth and performance [4]. Probiotics can help the weaning piglets adapt to this stressful condition by modulating the intestinal microbial population and stimulating the immune system of the host, which can then reduce diarrhea incidence, enhance gut health and growth performance [21,22,32]. A number of bacterial species have been used as probiotics. Among them lactic acid bacteria and butyric acid bacteria are the most commonly used in swine production [33]. In the present study the multi-strain probiotic compound containing B. mesentericus TO-A, B. coagulans SNZ1969, C. butyricum TO-A and E. faecalis T-110showed beneficial effect on growth parameters, lipid profile, antioxidant defense system and immune parameters in weaning piglets; thus, may be a good alternative to in-feed ZnO supplementation.
Body weight, ADFI, F:G and ADG are vital parameters of animal performance in pig industry. Here, the weaned piglets supplemented with probiotics (WB) demonstrated higher body weight, fortnightly ADG and overall ADG than those of the weaned negative control group suggesting beneficial effects of currently used multi-strain probiotic formulation. Moreover, as compared to the negative control group, the probiotic group had significantly lower diarrhea rate as well as F:G . Probiotics supplemented group and positive control group did not show any significant difference in growth and incidence of diarrhea. Similar results were reported by Cai et al. [34] who observed improved growth performance in weaned piglets supplemented with Bacillus based probiotics. Our findings were also supported by a former study in which weaned pigs received a complex probiotic formulation (L. paracasei, E. faecium and B. subtilis) had greater body weight, improved ADG and lower F:G than negative control [35]. Probiotics produce various enzymes, including arabinose, α-amylase, maltase, cellulase, levansucrase, dextranase, alkaline protease, β-glucanas, and neutral protease which enhance the nutrient digestibility in the gut [36,37]. However, the results of probiotic supplementation are inconsistent. Previous studies of Giang et al. [38] and Méndez-Palacios et al. [39] reported that the inclusion of Bacillus-based or Lactobacillus-based feed additives were failed to enhance the growth and production parameters of newly weaned piglets. The outcome of these studies may have been altered due to several other factors including diet composition, feed form and their interaction with probiotics, stains, probiotics doses, age of the pigs, surrounding environment and strategies of probiotics supplementation [40].
At weaning, piglets experience fasting due to abrupt change in feed; this stimulates fat mobilization from its reservoirs to support energy deficit [41]. High cholesterol concentration especially high concentration of LDL, and high level of TG in the bloodstream are closely related with atherosclerotic cardiovascular disease [42,43]. In the present study, current probiotics formulation improved the lipid profile of WB group which had significantly greater HDLc and lower TC and LDLc concentration as compared to the WC group. Our observations were in accordance with previous studies. Yu et al. [44] found that selenium in combination with complex probiotics (L. acidophilus, L. pentose, and B. subtilis) tended to reduce TC, VLDL, and TG and increase HDLc concentrations. Kim al. [45] found that cholesterol concentration was significantly decreased in the pigs fed with Lactobacillus-based probiotic. From this, it may be inferred that probiotic supplementation in weaning piglets improves the lipid profile by decreasing the TC and LDL concentration in serum and by increasing the HDLc concentration.
Weaning stress often induces oxidative stress [10,46]. In normal cellular metabolism, NO synthase (NOS) and NAD(P)H oxidase isoforms generate reactive oxygen species (ROS) and reactive nitrogen species (RNS) respectively [47]. At low/moderate concentrations, ROS and RNS are involved in a variety of physiological roles including cell signaling pathways and mitogenic response [48]. But when ROS and RNS concentration exceeds the normal cellular level, they cause potential damage to essential biomolecules (DNA, proteins, and lipids), starting a free radical chain reaction [49]. Therefore, oxidative stress and subsequent overproduction of ROS and RNS reduce immune response, increase susceptibility to pathogenic microorganisms, induce enterocyte apoptosis with cell cycle arrest in the gastrointestinal tract and eventually decrease production performance [10,50]. The current study observed that the serum TNO concentration between WB and PC did not vary throughout the experimental period, whereas the concentration was significantly increased on day 7 and 14 in WC than the other two groups. Furthermore, we determined MDA concentration to investigate whether weaning stress led to oxidative damage. Cell membranes or plasma membranes are rich in polyunsaturated fatty acids that make susceptible to free radical assault because of their multiple double bonds [51]. As a result of this oxidation of lipid molecules, MDA is produced, which interacts with biomolecules and exerts cytotoxic and genotoxic effects [52]. So, MDA is used as a biomarker of oxidative damage [53]. In this study, WB group recorded lower serum MDA concentration than WC group. These results suggest that the probiotics supplementation can reduce lipid peroxidation and oxidative damage. Similar findings were reported in previous reports [54,55].
Decrease in total NO and MDA concentration in probiotics supplemented piglets over un-supplemented piglets indicated improvement in the antioxidant system. The improved serum T-AOC offered additional support for an improvement in the antioxidant defense system for probiotic treated piglets. The antioxidant defense system in the body comprises of several antioxidant enzymes such as superoxide dismutase (SOD), glutathione peroxidase (GSH-Px), and catalase [56]. SOD converts superoxide radicals to less toxic H2O2 and then it is further processed to non-toxic water either by GSH-Px or catalase [57]. Probiotics can enhance the T-AOC by increasing the production of these antioxidant enzymes [58]. In our study, WB group showed elevated T-AOC, SOD, catalase and GSH levels on day 14 and 28 than WC group. Increased T-AOC and antioxidant enzyme levels in the probiotic supplemented group further supported the idea that currently used probiotic formulation may contribute to improve the antioxidant profiles of weaned piglets. Similar observations were reported earlier [54,55]. LAB can degrade free radicals by producing intracellular enzymatic (SOD and catalase) and non-enzymatic antioxidants (glutathione and thioredoxin) [59,60]. This explains how the current probiotics improved the antioxidant status of weaned piglets.
Stresses associated with weaning especially oxidative stress enhance the production of heat shock proteins (HSPs) [61]. HSPs are highly conserved intracellular proteins that are conserved across the species, but cell death or tissue injury caused by physiological stress may increase their release into serum [62]. HSPs are involved in a variety of physiological functions including, protein synthesis and homeostasis, improve antioxidant defense system, protect gut epithelium from oxidative stress and inflammation and inhibit apoptotic pathways [63]. In our study, the piglets with probiotic supplementation (WB) showed significantly decreased serum levels of HSPs (HSP70, HSP40, HSP90 and HSP20) as compared to the un-supplemented group (WC). This indicates downregulating effect of probiotics supplementation on serum HSPs levels in weaning piglets. Weaning increases formation of free radicals which induce the production of HSPs [61,64]. So, the down regulating effect of probiotic on serum HSPs might be due to decreased formation of free radicals and increase in total antioxidant capacity. Similar results were also reported Gan et al. [65] in which selenium-enriched probiotic supplementation reduced mRNA expression of HSPs in heat stressed piglets.
Cortisol is an important biomarker of stress. Weaning is a stressful event for a piglet and most frequently cause a marked rise in cortisol level [66]. Cortisol is assumed to be produced by the hypothalamic-pituitary-adrenal (HPA) axis [67]. Under stress like weaning stress, HPA axis activates and increases the production of cortisol hormone which is a healthy adaptive response of body to cope up with the situation [68]. Cortisol, up to a certain level, is beneficial to the host, but at high level and chronic persistence may have deleterious effects on productivity [69]. In our study, serum cortisol concentration significantly decreased in probiotic group on day 14 and 28 as compared to the negative control group, whereas there was no difference with positive control group. These findings clearly indicate that weaning stress enhances the production of cortisol hormone, which can be down regulated by probiotic supplementation. These findings provide another evidence that dietary probiotic supplementation can alleviate the weaning stress. Our findings were in accordance with previous studies; Burdick Sanchez et al. [70] found that serum cortisol concentration was reduced in pigs fed with Lactobacillus acidophilus fermentation product. Similarly, Wang et al. [71] observed that dietary supplementation of L. fermentum I5007 in weaned piglets could decrease the diquat induced plasma cortisol level.
Immune system modulation is one of the crucial roles of probiotics. It has been reported that probiotics interact with gut microbiota, epithelial cells and immune cells which in turn stimulate the immune function and antibody production [72,73]. IgAs are the predominant isotype expressed in all the mucosal tissues and aids in mucosal immunity. The most prevalent antibody, IgG, is present in blood and extracellular fluid and plays important roles in systemic immune response. IgM is the main antibody produced at the initial stage of antibody mediated immune response and the major component of natural antibody [74]. In the present study, the probiotic group displayed higher levels of serum IgG and IgM at all the time points and higher IgA at day 28 as compared to the negative control group. When compared with the positive control group, higher serum levels of all the Igs were detected in the probiotic group at the end of the experiment. These findings were in accordance with previous studies. Dong et al. [75] demonstrated that the serum IgM and IgA levels were improved in weaned piglets treated with complex probiotic containing L. plantarum and B. subtilis. Dlamini et al. [76] also observed increased IgG levels in weaned piglets upon supplementation of a combined probiotics. Our results suggested that direct-fed complex probiotics enhanced mucosal as well as humoral immunity of the weaned piglets. These significant increases may be due to the persistence of current probiotic bacteria in the intestinal tract and acting as an immune adjuvant to the humoral immune system and therefore stimulating antibody production. We next determined the concentrations of serum pro-inflammatory cytokines including IL-2, IL-1β, IFN-γ, IL-6, and IL-12 and anti-inflammatory cytokine IL-4 to assess the effects of weaning stress on intestinal or systemic inflammatory response. Cytokines are small and cell signaling molecules which plays major roles in the immune and inflammatory responses and overall homeostasis of the body [77]. Excess production of cytokines especially pro-inflammatory cytokines has negative influence on immune response and gut integrity which ultimately reduce the growth performance [78,79]. There is an intricate balance between pro and anti-inflammatory cytokines; anti-inflammatory cytokines suppress the production of pro-inflammatory cytokines and thus protect against intestinal inflammation and maintain the gut integrity [77]. Their harmony is therefore essential for the host immunological and inflammatory response. Abrupt changes in the dietary and environmental factors during weaning may lead to changes in the cytokine network and cause transient inflammation of the gut which may disrupt the barrier function [80]. In current study, as compared to un-supplemented group (WC), the concentrations of pro-inflammatory cytokines were lower in probiotics supplemented group. This was in agreement with a previous finding where Bacillus based compound probiotics (C. butyricum, B. subtilis, and B. licheniformis) modulated the inflammatory process in weaned piglets by decreasing serum pro-inflammatory cytokines (IL-1β, IL-6, and TNF-α) [81]. In case of IL-4, the anti-inflammatory cytokine, significantly increased levels on day 7, 14 and 28 in the probiotic group than the levels in negative control group were recorded. Similar results were published by Laskowska et al. [82] that showed the multi-microbial probiotic formulation "Bokashi" could raise the serum IL-4 level in pregnant sow. The experimental data of this study indicated that currently used multi-strain probiotic combination can reduce the transient inflammation and improve intestinal barrier function which eventually contribute to improved growth performance of weaned piglets.
5. Conclusions
It may be concluded that supplementation of the multi-strain Bacillus-based probiotic formulation containing Bacillus mesentericus, Bacillus coagulans, Enterococcus faecalis and Clostridium butyricum minimized the weaning stress, thereby improved feed intake, body weight, antioxidant activity, lipid profile, and systemic as well as mucosal immunity and overall growth performance of weaned piglets. Therefore, the multi-strain probiotic compound may be used to replace ZnO in weaned piglets.
Author Contributions
Conceptualization, A.K.D. and G.S.; methodology, G.S., S.S., S.M., P.P. and D.B.; software, A.K.D.; validation, D.B.; formal analysis, A.K.D.; investigation, G.S., P.P., S.S. and A.K.D.; resources, A.K.D. and D.B.; data curation, A.K.D.; writing—original draft preparation, A.K.D.; writing—review and editing, D.B.; visualization, S.M.; supervision, D.B.; project administration, A.K.D.; funding acquisition, A.K.D. The published version of the manuscript has been read and approved by all authors.
Funding
The current study was funded by All India Coordinated Research Project on Pig (AICRP on Pig), Indian Council of Agricultural Research, New Delhi, India with Grant number AICRP-Pig/ICAR-CIARI and National Bank for Agriculture and Rural Development (NABARD), Port Blair, Andaman and Nicobar Islands, India with Grant number NABARD/IDA in Piglets.
Institutional Review Board Statement
The animal study protocol was approved by the Institutional Ethics Committee of ICAR-Central Island Agricultural Research Institute (ICAR-CIARI), Port Blair, Andaman and Nicobar Islands, India (protocol code ICAR-CIARI/AS/23468 and date of approval 23.12.2021). Humane animal care was practiced throughout the study and every effort was made to minimize the suffering of the animals. All procedures were carried out in conformity with the relevant national regulations and guidelines.
Data Availability Statement
All data are available within the manuscript.
Conflicts of Interest
There is no competing interest amongst the authors. The study's design, data collection, analysis, or interpretation; the preparation of the manuscript; or the choice to publish the findings were all made independently of the funders.
References
- Wang, J.Q.; Yin, F.G.; Zhu, C. Evaluation of probiotic bacteria for their effects on the growth performance and intestinal microbiota of newly-weaned pigs fed fermented high-moisture maize. Livest. Sci. 2012, 145, 79–86. [Google Scholar] [CrossRef]
- Jensen, P. Observations on the maternal behaviour of free-ranging domestic pigs. Appl. Anim. Behav. Sci. 1986, 16, 131–142. [Google Scholar] [CrossRef]
- Tang, X.; Xiong, K.; Fang, R.; Li, M. Weaning Stress and Intestinal Health of Piglets: A Review. Front. Immunol. 2022, 13. [Google Scholar] [CrossRef] [PubMed]
- Prunier, A.; Heinonen, M.; Quesnel, H. High Physiological Demands in Intensively Raised Pigs: Impact on Health and Welfare*. Animal 2010, 4, 886–898. [Google Scholar] [CrossRef] [PubMed]
- Duthie, G.G.; Arthur, J.R.; Nicol, F.; Walker, M. Increased indices of lipid peroxidation in stress-susceptible pigs and effects of vitamin E. Res. Vet. Sci. 1989, 46, 226–230. [Google Scholar] [CrossRef] [PubMed]
- García, E.K.; deSouza, T.C.R.; Landín, G.M.; Barreyro, A.A.; Santos, M.G.B.; Soto, J.G.G. Microbial fermentation patterns, diarrhea incidence, and performance in weaned piglets fed a low protein diet supplemented with probiotics. Food Nutr. Sci. 2014, 5, 1776–1786. [Google Scholar] [CrossRef]
- Hao, Y.; Xing, M.; Gu, X. Research Progress on Oxidative Stress and Its Nutritional Regulation Strategies in Pigs. Animals 2021, 11. [Google Scholar] [CrossRef]
- Leibbrandt, V.D.; Ewan, R.C.; Speer, V.C.; Zimmerman, D.R. Effect of Weaning and Age at Weaning on Baby Pig Performance. J. Anim. Sci. 1975, 40, 1077–1080. [Google Scholar] [CrossRef]
- Faccin, J.E.G.; Laskoski, F.; Hernig, L.F.; Kummer, R.; Lima, G.F.R.; Orlando, U.A.D.; Goncalves, M.A.D.; Mellagi, A.P.G.; Ulguim, R.R.; Bortolozzo, F.P. Impact of Increasing Weaning Age on Pig Performance and Belly Nosing Prevalence in a Commercial Multisite Production System. J. Anim. Sci. 2020, 98. [Google Scholar] [CrossRef]
- Bonetti, A.; Tugnoli, B.; Piva, A.; Grilli, E. Towards Zero Zinc Oxide: Feeding Strategies to Manage Post-Weaning Diarrhea in Piglets. Animals 2021, 11, 642. [Google Scholar] [CrossRef]
- Casewell, M.; Friis, C.; Marco, E.; McMullin, P.; Phillips, I. The European Ban on Growth-Promoting Antibiotics and Emerging Consequences for Human and Animal Health. J. Antimicrob. Chemother. 2003, 52, 159–161. [Google Scholar] [CrossRef] [PubMed]
- Wen, R.; Li, C.; Zhao, M.; Wang, H.; Tang, Y. Withdrawal of Antibiotic Growth Promoters in China and Its Impact on the Foodborne Pathogen Campylobacter Coli of Swine Origin. Front. Microbiol. 2022, 13, 1004725. [Google Scholar] [CrossRef] [PubMed]
- Kyung-hyo, D.; Jae-won, B.; Wan-kyu, L. Antimicrobial Resistance Profiles of Escherichia Coli from Diarrheic Weaned Piglets after the Ban on Antibiotic Growth Promoters in Feed. Antibiotics 2020, 9, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Bednorz, C.; Oelgeschläger, K.; Kinnemann, B.; Hartmann, S.; Neumann, K.; Pieper, R.; Bethe, A.; Semmler, T.; Tedin, K.; Schierack, P.; Wieler, L.H.; Guenther, S. The broader context of antibiotic resistance: zinc feed supplementation of piglets increases the proportion of multi-resistant Escherichia coli in vivo. Int. J. Med. Microbiol. 2013, 303, 396–403. [Google Scholar] [CrossRef] [PubMed]
- Slifierz, M.J.; Friendship, R.; Weese, J.S. Zinc oxide therapy increases prevalence and persistence of methicillin-resistant Staphylococcus aureus in pigs: a randomized controlled trial. Zoonoses Public Health 2015, 62, 301–308. [Google Scholar] [CrossRef]
- Pejsak Z, Kaźmierczak P, Butkiewicz AF, Wojciechowski J, Woźniakowski G. Alternatives to Zinc Oxide in Pig Production. Pol. J. Vet. Sci. 2023, 26. [Google Scholar] [CrossRef]
- Delcenserie, V.; Martel, D.; Lamoureux, M.; Amiot, J.; Boutin, Y.; Roy, D. Immunomodulatory effects of probiotics in the intestinal tract. Curr. Issues Mol. Biol. 2008, 10, 37–54. [Google Scholar]
- Schlee, M.; Harder, J.; Köten, B.; Stange, E.F.; Wehkamp, J.; Fellermann, K. Probiotic lactobacilli and VSL#3 induce enterocyte beta-defensin 2. Clin. Exp. Immunol. 2008, 151, 528–535. [Google Scholar] [CrossRef]
- Fuller, R. Probiotics in man and animals. J. Appl. Bacteriol. 1989, 66, 365–378. [Google Scholar]
- Hong, H.A.; Le, H.D; Cutting, S.M. The use of bacterial spore formers as probiotics. FEMS Microbiol. Rev. 2005, 29, 813–835. [Google Scholar] [CrossRef]
- Weichselbaum, E. Probiotics and health: A review of the evidence. Nutr. Bull. 2009, 34, 340–373. [Google Scholar] [CrossRef]
- Sun, T.; Miao, H.; Zhang, C.; Wang, Y.; Liu, S.; Jiao, P.; Li, W.; Li, Y.; Huang, Z. Effect of Dietary Bacillus Coagulans on the Performance and Intestinal Microbiota of Weaned Piglets. Animal 2022, 16, 100561. [Google Scholar] [CrossRef]
- Lan, R.; Tran, H.; Kim, I. Effects of probiotic supplementation in different nutrient density diets on growth performance, nutrient digestibility, blood profiles, fecal microflora and noxious gas emission in weaning pig. J. Sci. Food Agric. 2017, 97, 1335–1341. [Google Scholar] [CrossRef]
- Narayanappa, D. Randomized double blinded controlled trial to evaluate the efficacy and safety of Bifilac in patients with acute viral diarrhea. Indian J. Pediatr. 2008, 75, 709–713. [Google Scholar] [CrossRef] [PubMed]
- Poulsen, H.D. Zinc oxide for weanling piglets. Acta Agriculturae Scandinavica A–Animal Sciences, 1995, 45, 159–167. [Google Scholar] [CrossRef]
- NRC (National Research Council). Nutrient Requirements of Swine, 11th ed.; National Academy Press: Washington, DC, USA, 2012. [Google Scholar]
- Walsh, A.M.; Sweeney, T.; O’Shea, C.J.; Doyle, D.N.; O’Doherty, J. V. Effect of Supplementing Different Ratios of Laminarin and Fucoidan in the Diet of the Weanling Piglet on Performance, Nutrient Digestibility, and Fecal Scoring. J. Anim. Sci. 2012, 90, 215–217. [Google Scholar] [CrossRef]
- Liu, H.; Hu, J.; Mahfuz, S.; Piao, X. Effects of Hydrolysable Tannins as Zinc Oxide Substitutes on Antioxidant Status, Immune Function, Intestinal Morphology, and Digestive Enzyme Activities in Weaned Piglets. Animals (Basel) 2020, 10, 757. [Google Scholar] [CrossRef]
- McNamara, J.R.; Cohn, J.S.; Wilson, P.W.F.; Schaefer, E.J. Calculated Values for Low-Density Lipoprotein Cholesterol in the Assessment of Lipid Abnormalities and Coronary Disease Risk. Clin. Chem. 1990, 36, 36–42. [Google Scholar] [CrossRef]
- Marbut, M.M.; Majeed, B.M.; Rahim, S.M.; Yuusif, M.N. Estimation of malondialdehyde as oxidative factor & glutathione as early detectors of hypertensive pregnant women. Tikrit. Med. J. 2009, 15, 63–69. [Google Scholar]
- Chakraborty, M.; Gouroju, S.; Singh, P.; Thatkar, P. V.; Bagchi, D. Tracking Lipid Profile and Atherogenic Indices in the Prediabetics of Andaman Nicobar Islands: A Retrospective Hospital-Based Study. J. Fam. Med. Prim. Care 2019, 8, 1117. [Google Scholar] [CrossRef]
- Liao, S.F.; Nyachoti, M. Using probiotics to improve swine gut health and nutrient utilization. Anim. Nutr. 2017, 3, 331–343. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Kuroki, Y.; Oka, K.; Takahashi, M.; Rao, S.; Sukegawa, S.; Fujimura, T. Effects of Dietary Supplementation with Enterococcus Faecium and Clostridium Butyricum, Either Alone or in Combination, on Growth and Fecal Microbiota Composition of Post-Weaning Pigs at a Commercial Farm. Front. Vet. Sci. 2019, 6, 432044. [Google Scholar] [CrossRef] [PubMed]
- Cai, L.; Indrakumar, S.; Kiarie, E.; Kim, I.H. Effects of a multi-strain Bacillus species–based direct-fed microbial on growth performance, nutrient digestibility, blood profile, and gut health in nursery pigs fed corn–soybean meal–based diets. J. Anim. Sci. 2015, 93, 4336–4342. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Zhang, M.; Zhao, L.; Ge, K.; Wang, Z.; Jun, L.; Ren, F. Growth performance and post-weaning diarrhea in piglets fed a diet supplemented with probiotic complexes. J. Microbiol. Biotechnol. 2018, 28, 1791–1799. [Google Scholar] [CrossRef]
- Bajagai et al. Probiotics in Animal Nutrition - Production, Impact and Regulation; 2016; Vol. 24; ISBN 2-9524354-0-5.
- Yi, R.; Pan, Y.; Long, X.; Tan, F.; Zhao, X. Enzyme Producing Activity of Probiotics and Preparation of Compound Enzyme. J. Chem. 2020, 2020. [Google Scholar] [CrossRef]
- Giang, H.H.; Viet, T.Q.; Ogle, B.; Lindberg, J.E. Effects of Supplementation of Probiotics on the Performance, Nutrient Digestibility and Faecal Microflora in Growing-Finishing Pigs. Asian-Australasian J. Anim. Sci. 2011, 24, 655–661. [Google Scholar] [CrossRef]
- Méndez-Palacios, N.; Méndez-Mendoza, M.; Vázquez-Flores, F.; Castro-Colombres, J.G.; Ramírez-Bribiesca, J.E. Productive and Economic Parameters of Pigs Supplemented from Weaning to Finishing with Prebiotic and Probiotic Feed Additives. Anim. Sci. J. 2018, 89, 994–1001. [Google Scholar] [CrossRef]
- Jørgensen, J.N.; Laguna, J.S.; Millán, C.; Casabuena, O.; Gracia, M.I. Effects of a Bacillus-based probiotic and dietary energy content on the performance and nutrient digestibility of wean to finish pigs. Anim. Feed Sci. Technol. 2016, 221, 54–61. [Google Scholar] [CrossRef]
- Dodson, M.V.; Hausman, G.J.; Guan, L.; Du, M.; Rasmussen, T.P.; Poulos, S.P.; Mir, P.; Bergen, W.G.; Fernyhough, M.E.; McFarland, D.C.; Rhoads, R.P.; Soret, B.; Reecy, J.M.; Velleman, S.G.; Jiang, Z. Lipid metabolism, adipocyte depot physiology and utilization of meat animals as experimental models for metabolic research. Int. J. Biol. Sci. 2010, 6, 691–699. [Google Scholar] [CrossRef]
- Austin, M.A. Plasma triglyceride and coronary heart disease. Arterioscler. Thromb. 1991, 11, 2–14. [Google Scholar] [CrossRef]
- Busnelli, M.; Manzini, S.; Froio, A.; Vargiolu, A.; Cerrito, M.G.; Smolenski, R.T.; Giunti, M.; Cinti, A.; Zannoni, A.; Leone, B.E.; et al. Diet Induced Mild Hypercholesterolemia in Pigs: Local and Systemic Inflammation, Effects on Vascular Injury – Rescue by High-Dose Statin Treatment. PLoS One 2013, 8, 80588. [Google Scholar] [CrossRef] [PubMed]
- Yu, I.T.; Ju, C.C.; Lin, J.; Wu, H.L.; Yen, H.T. Effects of probiotics and selenium combination on the immune and blood cholesterol concentration of pigs. J. Anim. Feed Sci. 2004, 13, 625–634. [Google Scholar] [CrossRef]
- Kim, D.; Min, Y.; Yang, J.; Heo, Y.; Kim, M.; Hur, C.G.; Lee, S.C.; Lee, H.K.; Song, K.D.; Heo, J.; et al. Multi-Probiotic Lactobacillus Supplementation Improves Liver Function and Reduces Cholesterol Levels in Jeju Native Pigs. Animals 2021, 11, 2309. [Google Scholar] [CrossRef] [PubMed]
- Novais, A.K.; Martel-Kennes, Y.; Roy, C.; Deschêne, K.; Beaulieu, S.; Bergeron, N.; Laforest, J.P.; Lessard, M.; Matte, J.J.; Lapointe, J. Tissue-Specific Profiling Reveals Modulation of Cellular and Mitochondrial Oxidative Stress in Normal- and Low-Birthweight Piglets throughout the Peri-Weaning Period. Animal 2020, 14, 1014–1024. [Google Scholar] [CrossRef] [PubMed]
- Kurutas, E.B. The Importance of Antioxidants Which Play the Role in Cellular Response against Oxidative/Nitrosative Stress: Current State. Nutr. J. 2016, 15. [Google Scholar] [CrossRef] [PubMed]
- Carvajal Carvajal, C. Reactive oxygen species: training, function and oxidative stress. Med. leg. Costa Rica. 2019, 36, 91–100. [Google Scholar]
- Griendling, K.K.; Touyz, R.M.; Zweier, J.L.; Dikalov, S.; Chilian, W.; Chen, Y.R.; Harrison, D.G.; Bhatnagar, A. Measurement of Reactive Oxygen Species, Reactive Nitrogen Species, and Redox-Dependent Signaling in the Cardiovascular System. Circ. Res. 2016, 119, e39–e75. [Google Scholar] [CrossRef]
- Zhu, L.; Cai, X.; Guo, Q.; Chen, X.; Zhu, S.; Xu, J. Effect of N-acetyl cysteine on enterocyte apoptosis and intracellular signalling pathways' response to oxidative stress in weaned piglets. Br. J. Nutr. 2013, 110, 1938–1947. [Google Scholar] [CrossRef]
- Yin, H.; Xu, L.; Porter, N.A. Free Radical Lipid Peroxidation: Mechanisms and Analysis. Chem. Rev. 2011, 111, 5944–5972. [Google Scholar] [CrossRef]
- Ayala, A.; Muñoz, M.F.; Argüelles, S. Lipid Peroxidation: Production, Metabolism, and Signaling Mechanisms of Malondialdehyde and 4-Hydroxy-2-Nonenal. Oxid. Med. Cell. Longev. 2014, 2014. [Google Scholar] [CrossRef]
- Del Rio, D.; Stewart, A.J.; Pellegrini, N. A review of recent studies on malondialdehyde as toxic molecule and biological marker of oxidative stress. Nutr. Metab. Cardiovasc. Dis. 2005, 15, 316–328. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhang, Q.; Zhuo, Y.; Fang, Z.; Che, L.; Xu, S.; Feng, B.; Lin, Y.; Jiang, X.; Zhao, X.; et al. Effects of Multi-Strain Probiotics and Perilla Frutescens Seed Extract Supplementation Alone or Combined on Growth Performance, Antioxidant Indices, and Intestinal Health of Weaned Piglets. Animals 2022, 12. [Google Scholar] [CrossRef]
- Zhang, S.; Wu, Z.; Heng, J.; Song, H.; Tian, M.; Chen, F.; Guan, W. Combined Yeast Culture and Organic Selenium Supplementation during Late Gestation and Lactation Improve Preweaning Piglet Performance by Enhancing the Antioxidant Capacity and Milk Content in Nutrient-Restricted Sows. Anim. Nutr. 2020, 6, 160. [Google Scholar] [CrossRef] [PubMed]
- Ighodaro, O.M.; Akinloye, O.A. First line defence antioxidants-superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPX): Their fundamental role in the entire antioxidant defence grid. Alexandria J. Med. 2018, 54, 287–293. [Google Scholar] [CrossRef]
- Fukai, T.; Ushio-Fukai, M. Superoxide Dismutases: Role in Redox Signaling, Vascular Function, and Diseases. Antioxid. Redox Signal. 2011, 15, 1583. [Google Scholar] [CrossRef]
- Wang, A.N.; Yi, X.W.; Yu, H.F.; Dong, B.; Qiao, S.Y. Free radical scavenging activity of Lactobacillus fermentum in vitro and its antioxidative effect on growing-finishing pigs. J. Appl. Microbiol. 2009, 107, 1140–1148. [Google Scholar] [CrossRef]
- Knauf, H.J.; Vogel, R.F.; Hammes, W.P. Cloning, sequence, and phenotypic expression of katA, which encodes the catalase of Lactobacillus sake LTH677. Appl. Environ. Microbiol. 1992, 58, 832–839. [Google Scholar] [CrossRef]
- Stecchini, M.L.; Del Torre, M.; Munari, M. Determination of peroxy radical-scavenging of lactic acid bacteria. Int. J. Food Microbiol. 2001, 64, 183–188. [Google Scholar] [CrossRef]
- Arvans, D.L.; Vavricka, S.R.; Ren, H.; Musch, M.W.; Kang, L.; Rocha, F.G.; Lucioni, A.; Turner, J.R.; Alverdy, J.; Chang, E.B. Luminal Bacterial Flora Determines Physiological Expression of Intestinal Epithelial Cytoprotective Heat Shock Proteins 25 and 72. Am. J. Physiol. Gastrointest. Liver Physiol. 2005, 288. [Google Scholar] [CrossRef]
- Sepponen, K.; Pösö, A.R. The inducible form of heat shock protein 70 in the serum, colon and small intestine of the pig: Comparison to conventional stress markers. Vet. J. 2006, 171, 519–524. [Google Scholar] [CrossRef]
- Hu, C.; Yang, J.; Qi, Z.; Wu, H.; Wang, B.; Zou, F.; Mei, H.; Liu, J.; Wang, W.; Liu, Q. Heat Shock Proteins: Biological Functions, Pathological Roles, and Therapeutic Opportunities. MedComm 2022, 3. [Google Scholar] [CrossRef] [PubMed]
- David, J.C.; Grongnet, J.F.; Lalles, J.P. Weaning Affects the Expression of Heat Shock Proteins in Different Regions of the Gastrointestinal Tract of Piglets. J. Nutr. 2002, 132, 2551–2561. [Google Scholar] [CrossRef] [PubMed]
- Gan, F.; Ren, F.; Chen, X.; Lv, C.; Pan, C.; Ye, G.; Shi, J.; Shi, X.; Zhou, H.; Shituleni, S.A.; Huang, K. Effects of selenium-enriched probiotics on heat shock protein mRNA levels in piglet under heat stress conditions. J. Agric. Food Chem. 2013, 61, 2385–2391. [Google Scholar] [CrossRef] [PubMed]
- Kick, A.R.; Tompkins, M.B.; Flowers, W.L.; Whisnant, C.S.; Almond, G.W. Effects of Stress Associated with Weaning on the Adaptive Immune System in Pigs. J. Anim. Sci. 2012, 90, 649–656. [Google Scholar] [CrossRef]
- Hinds, J.A.; Sanchez, E.R. The Role of the Hypothalamus–Pituitary–Adrenal (HPA) Axis in Test-Induced Anxiety: Assessments, Physiological Responses, and Molecular Details. Stress 2022, 2, 146–155. [Google Scholar] [CrossRef]
- Martínez-Miró, S.; Tecles, F.; Ramón, M.; Escribano, D.; Hernández, F.; Madrid, J.; Orengo, J.; Martínez-Subiela, S.; Manteca, X.; Cerón, J.J. Causes, Consequences and Biomarkers of Stress in Swine: An Update. BMC Vet. Res. 2016, 12. [Google Scholar] [CrossRef]
- Van Erp-Van Der Kooij, E.; Kuijpers, A.H.; Van Eerdenburg, F.J.C.M.; Dieleman, S.J.; Blankenstein, D.M.; Tielen, M.J.M. Individual behavioural characteristics in pigs - Influences of group composition but no differences in cortisol responses. Physiol. Behav. 2003, 78, 479–488. [Google Scholar] [CrossRef]
- Burdick Sanchez, N.C.; Carroll, J.A.; Broadway, P.R.; Bass, B.E.; Frank, J.W. Supplementation of a Lactobacillus Acidophilus Fermentation Product Can Attenuate the Acute Phase Response Following a Lipopolysaccharide Challenge in Weaned Pigs. Animal 2019, 13, 144–152. [Google Scholar] [CrossRef]
- Wang, A.N.; Cai, C.J.; Zeng, X.F.; Zhang, F.R.; Zhang, G.L.; Thacker, P.A.; Wang, J.J.; Qiao, S.Y. Dietary supplementation with Lactobacillus fermentum I5007 improves the anti-oxidative activity of weanling piglets challenged with diquat. J. Appl. Microbiol. 2013, 114, 1582–1591. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, P.; Zhang, X. Probiotics Regulate Gut Microbiota: An Effective Method to Improve Immunity. Molecules 2021, 26. [Google Scholar] [CrossRef]
- Tejada-Simon, M.V.; Lee, J.H.; Ustunol, Z.; Pestka, J.J. Ingestion of yogurt containing Lactobacillus acidophilus and Bifidobacterium to potentiate immunoglobulin A responses to cholera toxin in mice. J. Dairy Sci. 1999, 82, 649–660. [Google Scholar] [CrossRef] [PubMed]
- Charles A Janeway, J.; Travers, P.; Walport, M.; Shlomchik, M.J. The Distribution and Functions of Immunoglobulin Isotypes. 2001.
- Dong, X.; Zhang, N.; Zhou, M.; Tu, Y.; Deng, K.; Diao, Q. Effects of dietary probiotics on growth performance, faecal microbiota and serum profiles in weaned piglets. Anim. Prod. Sci. 2014, 54, 616–621. [Google Scholar] [CrossRef]
- Dlamini, Z.C.; Langa, R.L.S.; Aiyegoro, O.A.; Okoh, A.I. Effects of probiotics on growth performance, blood parameters, and antibody stimulation in piglets. South African J. Anim. Sci. 2017, 47, 766–775. [Google Scholar] [CrossRef]
- Hu, C.H.; Xiao, K.; Luan, Z.S.; Song, J. Early weaning increases intestinal permeability, alters expression of cytokine and tight junction proteins, and activates mitogen-activated protein kinases in pigs. J. Anim. Sci. 2013, 91, 1094–1101. [Google Scholar] [CrossRef]
- Johnson, R.W. Inhibition of growth by pro-inflammatory cytokines: an integrated view. J. Anim. Sci. 1997, 75, 1244–1255. [Google Scholar] [CrossRef] [PubMed]
- Groot, N. De; Fariñas, F.; Cabrera-Gómez, C.G.; Pallares, F.J.; Ramis, G. Weaning Causes a Prolonged but Transient Change in Immune Gene Expression in the Intestine of Piglets. J. Anim. Sci. 2021, 99, 1–12. [Google Scholar] [CrossRef]
- Bomba, L.; Minuti, A.; Moisá, S.J.; Trevisi, E.; Eufemi, E.; Lizier, M.; Chegdani, F.; Lucchini, F.; Rzepus, M.; Prandini, A.; et al. Gut Response Induced by Weaning in Piglet Features Marked Changes in Immune and Inflammatory Response. Funct. Integr. Genomics 2014, 14, 657–671. [Google Scholar] [CrossRef]
- Cao, G.; Tao, F.; Hu, Y.; Li, Z.; Zhang, Y.; Deng, B.; Zhan, X. Positive effects of a Clostridium butyricum-based compound probiotic on growth performance, immune responses, intestinal morphology, hypothalamic neurotransmitters, and colonic microbiota in weaned piglets. Food Funct. 2019, 10, 2926–2934. [Google Scholar] [CrossRef]
- Laskowska, E.; Jarosz, Ł.; Grądzki, Z. Effect of multi-microbial probiotic formulation bokashi on pro- and anti-inflammatory cytokines profile in the serum, colostrum and milk of sows, and in a culture of polymorphonuclear cells isolated from colostrum. Probiotics Antimicrob. Proteins. 2019, 11, 220–232. [Google Scholar] [CrossRef]
Figure 1.
Effect on serum lipid panel of weaned piglets. (a) Total cholesterol (TC) concentration; (b) High-density lipoprotein cholesterol (HDLc) concentration; (c) Triglycerides (TG) concentration; (d) Low-density lipoprotein cholesterol (LDLc) concentration; (e) Cardiac risk factor (CRF); (f) Atherogenic index (AI). Analyzed data were shown as M ± SD. A,B,CValues in a particular time point having different superscripts. WC: weaned negative control group, WB: weaned probiotic group and PC: positive control (ZnO) group.
Figure 1.
Effect on serum lipid panel of weaned piglets. (a) Total cholesterol (TC) concentration; (b) High-density lipoprotein cholesterol (HDLc) concentration; (c) Triglycerides (TG) concentration; (d) Low-density lipoprotein cholesterol (LDLc) concentration; (e) Cardiac risk factor (CRF); (f) Atherogenic index (AI). Analyzed data were shown as M ± SD. A,B,CValues in a particular time point having different superscripts. WC: weaned negative control group, WB: weaned probiotic group and PC: positive control (ZnO) group.
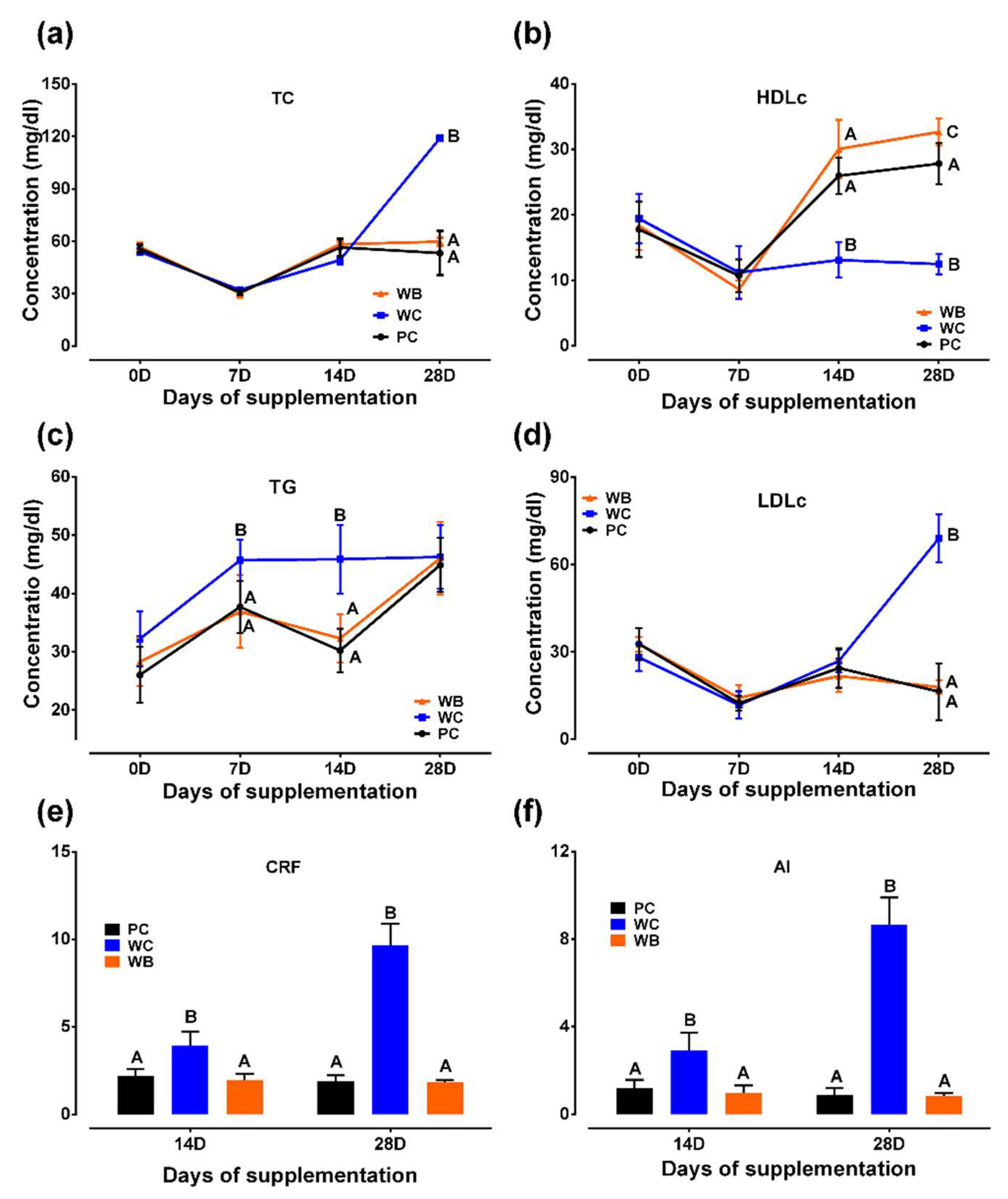
Figure 2.
Results of probiotic administration on antioxidant activity and oxidative stress markers of weaned piglets. (a) Total antioxidant capacity (T-AOC); (b) Superoxide dismutase (SOD) activity; (c) Catalase activity; (d) Glutathione-S-transferase (GSH) concentration; (e) Malonyldialdehyde (MDA) concentration. Analyzed data were shown as M ± SD. A,B,CValues in a particular time point having different superscripts. WC: weaned negative control group, WB: weaned probiotic group and PC: positive control (ZnO) group.
Figure 2.
Results of probiotic administration on antioxidant activity and oxidative stress markers of weaned piglets. (a) Total antioxidant capacity (T-AOC); (b) Superoxide dismutase (SOD) activity; (c) Catalase activity; (d) Glutathione-S-transferase (GSH) concentration; (e) Malonyldialdehyde (MDA) concentration. Analyzed data were shown as M ± SD. A,B,CValues in a particular time point having different superscripts. WC: weaned negative control group, WB: weaned probiotic group and PC: positive control (ZnO) group.
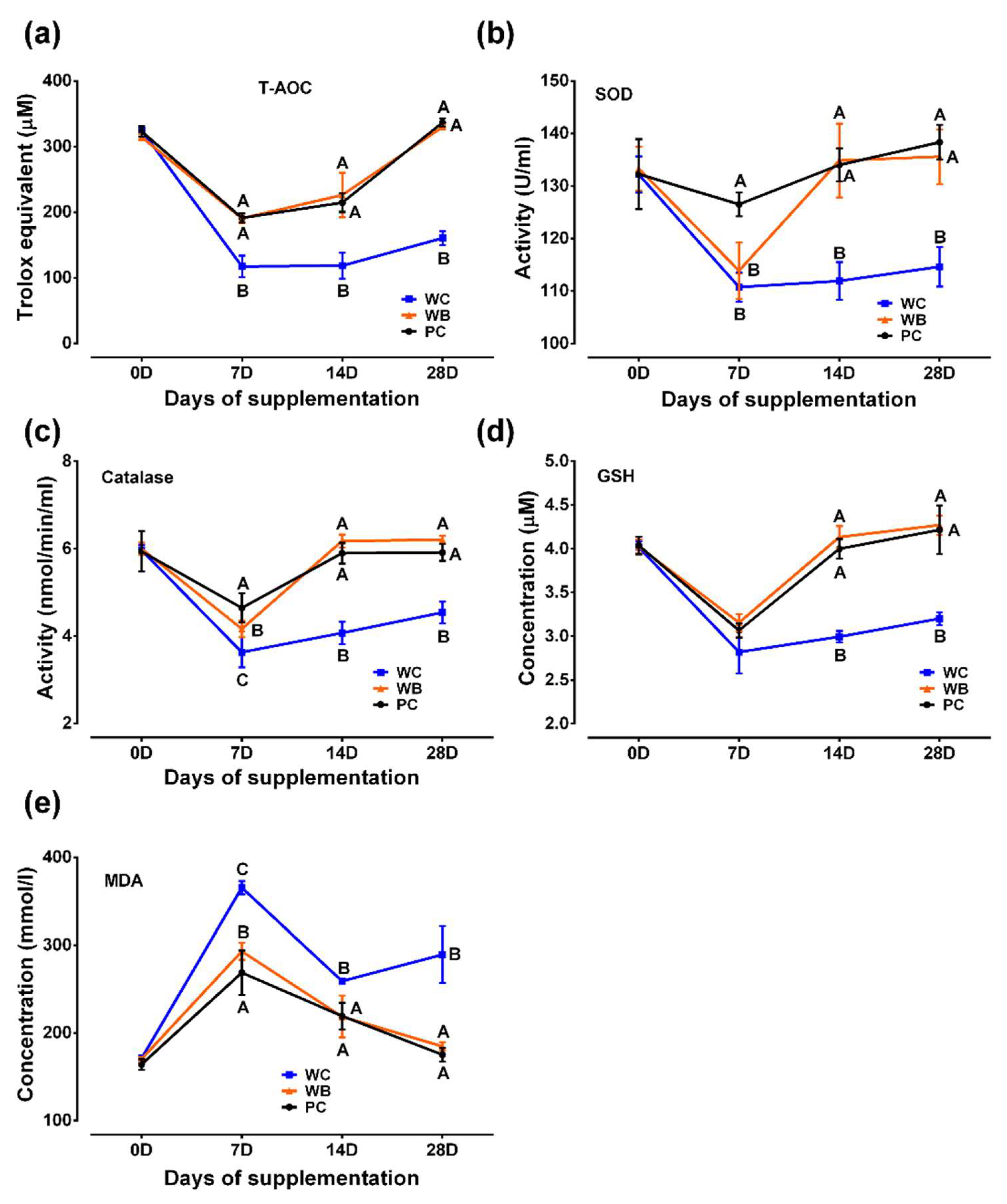
Figure 3.
Results of probiotic supplementation on total nitric oxide (TNO) and cortisol concentration in weaned piglets. (a) Serum TNO level, (b) Serum cortisol level. Analyzed data were shown as M ± SD. A,B,CValues in a particular time point having different superscripts. WC: weaned negative control group, WB: weaned probiotic group and PC: positive control (ZnO) group.
Figure 3.
Results of probiotic supplementation on total nitric oxide (TNO) and cortisol concentration in weaned piglets. (a) Serum TNO level, (b) Serum cortisol level. Analyzed data were shown as M ± SD. A,B,CValues in a particular time point having different superscripts. WC: weaned negative control group, WB: weaned probiotic group and PC: positive control (ZnO) group.
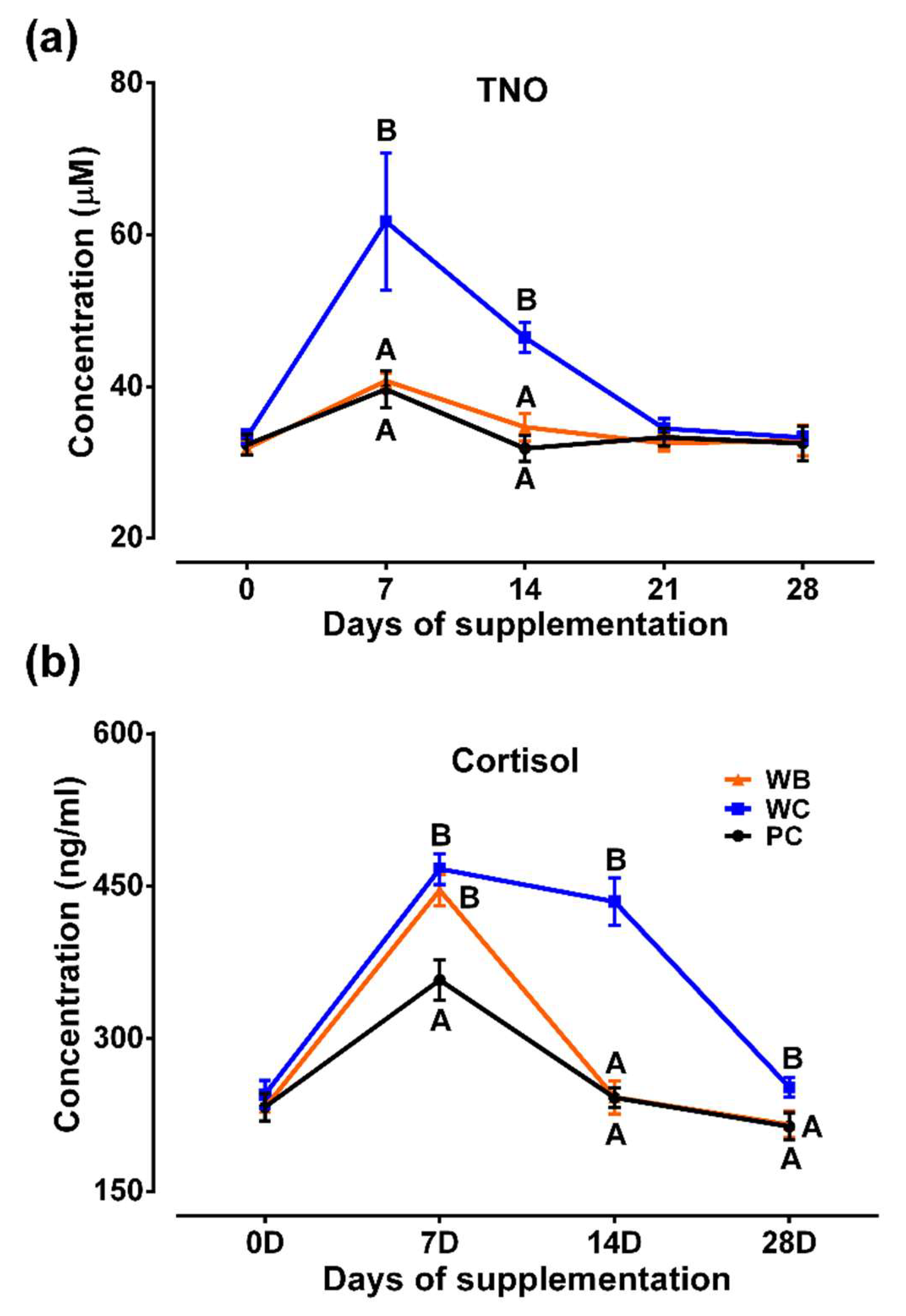
Figure 4.
Results of probiotic supplementation on heat shock proteins (HSPs) in weaned piglets. (a) HSP20; (b) HSP40; (c) HSP70; (d) HSP90. Analyzed data were shown as M ± SD. A,B,CValues in a particular time point having different superscripts. WC: weaned negative control group, WB: weaned probiotic group and PC: positive control (ZnO) group.
Figure 4.
Results of probiotic supplementation on heat shock proteins (HSPs) in weaned piglets. (a) HSP20; (b) HSP40; (c) HSP70; (d) HSP90. Analyzed data were shown as M ± SD. A,B,CValues in a particular time point having different superscripts. WC: weaned negative control group, WB: weaned probiotic group and PC: positive control (ZnO) group.
Figure 5.
Results of probiotic supplementation on serum immunoglobulins and cytokines in weaned piglets. (a) IgA; (b) IgG; (c) IgM; (d) IL-2; (e) IL-4; (f) IL-6; (g) IL-12; (h) IL-1β; (i) IFN-γ. Analyzed data were shown as M ± SD. A,B,CValues in a particular time point having different superscripts. WC: weaned negative control group, WB: weaned probiotic group and PC: positive control (ZnO) group.
Figure 5.
Results of probiotic supplementation on serum immunoglobulins and cytokines in weaned piglets. (a) IgA; (b) IgG; (c) IgM; (d) IL-2; (e) IL-4; (f) IL-6; (g) IL-12; (h) IL-1β; (i) IFN-γ. Analyzed data were shown as M ± SD. A,B,CValues in a particular time point having different superscripts. WC: weaned negative control group, WB: weaned probiotic group and PC: positive control (ZnO) group.
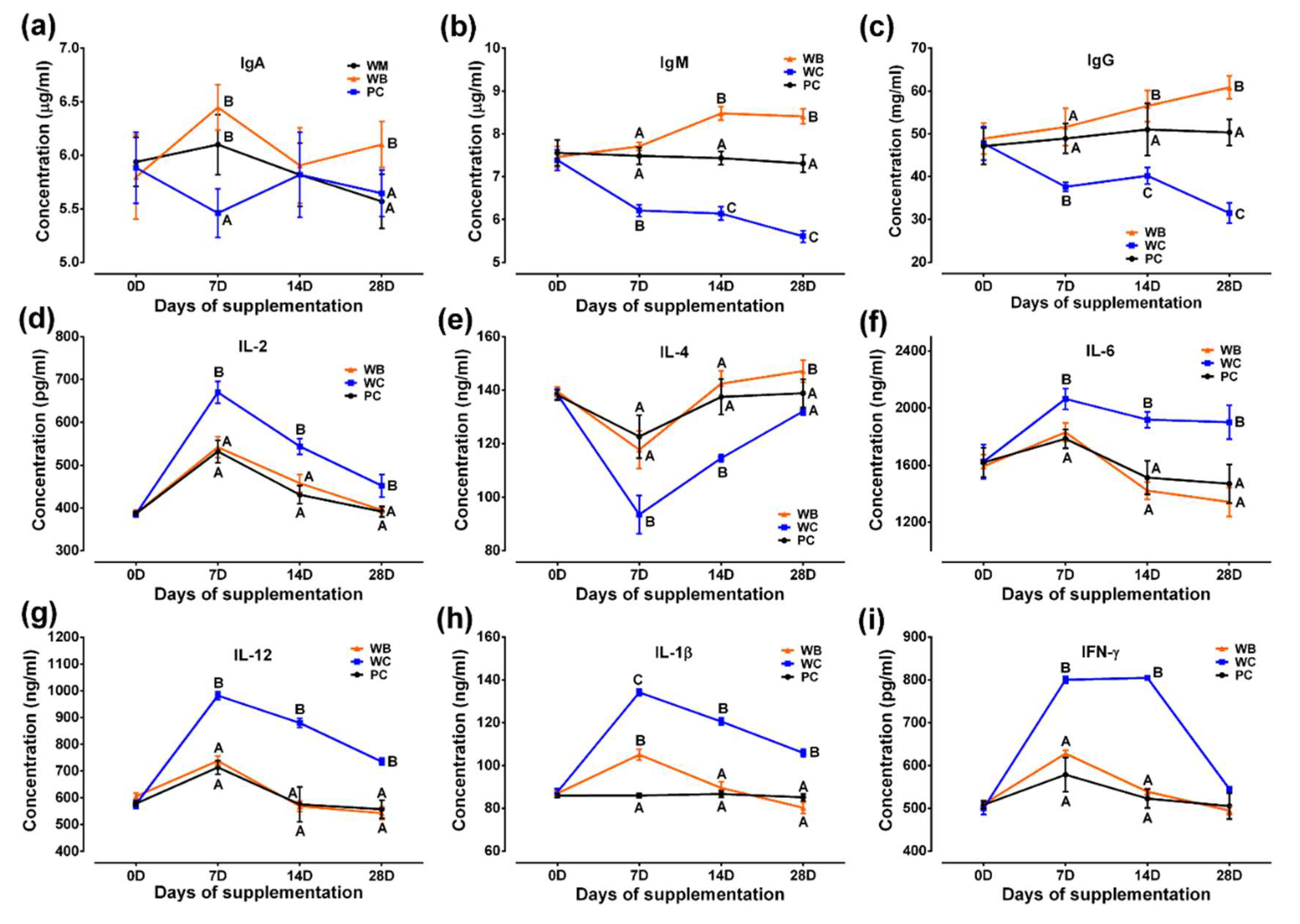

Table 1.
Composition of the probiotic.
| Microbial composition | Strain number | GenBank accession details | Deposition details | Content per gram of product |
|---|---|---|---|---|
| Enterococcus faecium | T-110 | AB687552: 16S DNA | 8936* | 3 x 107 CFU/g |
| CP006030: Complete genome | ||||
| CP006031: Complete plasmid | ||||
| Clostridium butyricum | TOA | AB687551: 16S DNA | 8935* | 2 x 106 CFU/g |
| CP014704: Chromosome 1 CP014705: Chromosome 2 | ||||
| CP014706: Plasmid | ||||
| Bacillus mesentericus | TOA | AB687550:16S DNA | 8934* | 1 x 106 CFU/g |
| CP005997: Complete genome | ||||
| Bacillus coagulans | SNZ 1969 | KC146407: 16S DNA | MTCC 5724 | 5 x 107 CFU/g |
*All the three strains were deposited with the international deposit agency in Japan.
Table 2.
Composition of basal diet and its nutritional levels.
| Ingredients | Percentage |
|---|---|
| Maize | 50.00 |
| Wheat bran | 15.00 |
| Soybean meal | 29.00 |
| Vitamin & trace min. mix. | 2.50 |
| MCP | 1.00 |
| Salt | 1.00 |
| CaCO3 | 1.28 |
| DL-Methionine | 0.22 |
| Chemical formula of basal diet | |
| Crude Protein (CP) | 20.72 |
| Calcium | 0.80 |
| Total phosphorus | 0.63 |
| Lysine | 0.80 |
| Methionine and Cystine | 0.70 |
| Metabolizable energy (ME) (kcal/kg) | 3382.31 |
The basal diet was prepared and given to the experimental piglets according to the nutritional guidelines of National Research Council [26]. Vitamin & trace mineral mix has (per kg feed): vitamin D3 (4000 IU); vitamin K (16 mg); vitamin E (80 IU); vitamin A (20,000 IU); Ca-pantothenate (50 mg); niacin (120 mg); riboflavin (20 mg); pyridoxine (6 mg); thiamine (4 mg); folic acid (2 mg); vitamin B12 (0.08 mg); biotin (0.08 mg); Mn (73 mg); Cu Zn (56 mg); (15 mg); Co (0.5 mg); Se (0.4 mg); I (0.3 mg).
Table 3.
Production Parameters and diarrhea incidence of piglets.
| Parameters | WC | WB | PC |
|---|---|---|---|
| Body Weight/kg | |||
| 0 D | 8.84 ± 0.12 | 8.79 ± 0.14 | 8.75 ± 0.12 |
| 7 D | 9.87 ± 0.33 | 10.11 ± 0.24 | 10.38 ± 0.18 |
| 14 D | 11.25 ± 0.13A | 11.79 ± 0.11B | 11.78 ± 0.14B |
| 21 D | 12.58 ± 0.17A | 13.63 ± 0.34B | 13.71 ± 0.38B |
| 28 D | 13.80 ± 0.22A | 14.88 ± 0.43B | 14.97 ± 0.45B |
| ADG/g | |||
| 0-14 D | 171.90 ± 10.70A | 214.17 ± 14.14B | 216.2 ± 15.98B |
| 14-28 D | 182.26 ± 17.69A | 220.48 ± 28.47B | 228.1± 31.70B |
| 0-28 D | 177.08 ± 7.21A | 217.32 ± 15.54B | 222.1 ± 16.76B |
| ADFI/g | |||
| 0-14 D | 417.73 ± 26.01A | 454.03 ± 29.98AB | 459.2 ± 33.94B |
| 14-28 D | 466.59 ± 45.30 | 485.05 ± 62.63 | 499.2 ± 69.32 |
| 0-28 D | 442.16 ± 18.45 | 469.54 ± 33.97 | 479.2 ± 36.78 |
| F:G | |||
| 0-14 D | 2.426 ± 0.03A | 2.11 ± 0.01B | 2.12 ± 0.01B |
| 14-28 D | 2.57 ± 0.02A | 2.20 ± 0.01B | 2.18 ± 0.01B |
| 0-28 D | 2.49 ± 0.01A | 2.16 ± 0.01B | 2.16 ± 0.01B |
| Diarrhea rate (%) | |||
| 0-28 D | 11.67 ± 1.31A | 4.67 ± 1.07B | 4.53± 0.78B |
Analyzed data were shown as M ± SD. A,BValues with different superscripts in a same row differ significantly. WC designates weaned negative control group, WB designates weaned probiotic group and PC designates positive control (ZnO) group.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



