
Submitted:
11 September 2023
Posted:
13 September 2023
You are already at the latest version
Abstract
Phospholipases are among the important elements involved in lipid-dependent cell signaling that lead to the induction of downstream pathways. In the current study, phospholipases D (PLDs) gene family was characterized and compared in two important oilseeds crops, Brassica napus and Camelina sativa. The results revealed that PLD has 33 members in Camelina sativa (CsPLD) and 41 members in Brassica napus (BnPLD). All studied PLDs showed a negative GRAVY value, indicating that PLDs are probably hydrophilic proteins. Phylogenetic analysis separated PLDs into five subfamilies; gamma, delta, beta, alpha, and zeta. According to evolution analysis, a different evolution process was observed between CsPLD and BnPLD. In addition, the results disclosed that the most of the PLD genes have been segmentally duplicated under purifying selection. Cis-regulatory elements related to ABA and auxin responsiveness were more found in upstream region of CsPLDs, while elements linked with MeJA responsiveness were more dis-tributed in promoter region of BnPLDs. Analysis of the expression data showed that PLD alpha genes have a wide expression and are expressed in most tissues. Quantitative expression analysis (qPCR) of CsPLD genes under salt stress, 200 mM of NaCl, was done in different time series. The results revealed that the CsPLD genes are involved in the response to salinity stress and their expression levels enhance with the increase of salinity stress time. The outcomes of this research will be useful for future molecular work related to lipid signaling in oilseed plants.
Keywords:
evolution analysis
; oilseeds
; regulatory system
; sequence analysis
; lipid signaling
1. Introduction
Plants exploit lipid signaling to regulate a variety of cellular processes in response to external stimuli [1]. In this regard, phospholipids which are the main components of biological membranes can play as intracellular signaling molecules [2,3]. As part of maintaining membrane stability and homeostasis as well as biotic and abiotic stress alleviation mechanisms, these molecules are released directly and transiently by the activity of phospholipases or lipid kinases [4,5]. Mediating different types of catalytic reactions, plant phospholipases are divided into A, C, and D classes. In both prokaryotes and eukaryotes, phospholipases D (PLDs) are the main enzymes that break the terminal phosphodiester bonds of phospholipids in plasmalemma and organellar membranes yielding phosphatidic acid (PA) and free soluble head groups (e.g., ethanolamine or choline) [6,7]. It has been shown that PLDs can modulate various process in plants at cell scale including programmed cell dead [8], interaction with cytoskeleton [9], pollen tube growth [10], vesicular transport [11], and stomata closure [12] or at whole plant level including seed germination [13], wounding [14], and hair root growth and patterning [15]. There are three PLDs subfamilies including SP-PLD, PX/PH-PLD and C2-PLD, in which SP-PLD subfamily is classified into α, β, γ, δ, ε, and ζ classes on the basis of their physicochemical catalytic, and structural features and sequence homology [2,6]. While each PLD appears to have a different physiological function, all classes have a phospholipid-binding region at their N-terminus, and two highly conserved C-terminus domains that are catalytically involved in lipase activity through the interactions of two catalytic HxKxxxxD (HKD) motifs [16,17].
PLD enzyme is found in both plant and animal cells, and its activity is regulated by various signaling pathways and cellular factors. Several PLD cDNAs have been cloned from higher plant species since the cloning of the first eukaryotic PLD cDNA from castor bean (Ricinus sativus L.) [18]. The members of the PLD gene family can now be studied and discussed at the genomic level in many plant species thanks to advances in genome sequencing technologies. Du et al. (2013) studied phospholipase D gene family in three Rosaceae species and identified three pairs of segmental PLD gene duplications in Malus × domestica and Prunus mume and four pairs in Fragaria vesca L. [19]. Out of 40 identified PLD genes, 33 genes represented conserved synteny and the authors suggested that PLD genes in Rosacea family could have a common ancestor and purifying selection could be the primary driving force behind PLD gene evolution. Using genome-wide analysis, 17 PLDs were discovered in rice across four chromosomes. In addition to the C2-PLDs and PXPH-PLDs, the SP-PLDs has also bene identified as a novel PLD subfamily in rice (which contains a signal peptide sequence rather than the PXPH or C2 domains) based on the protein domain structural analyses [13]. Liu et al. (2010) identified 18 and 11 PLD members in grape and poplar, respectively. PLD genes were clustered into φ, ζ, ε, δ, β/γ, and α subgroups where they have evolved from four preliminary ancestors as a results of gene duplications [17]. They also found that most of PLD gene family members in grape (10 out of 11) and poplar (13 out of 17) showed closer phylogenetically relationship together than with rice and Arabidopsis PLD gene members. Genome-wide in silico analysis of PLD genes from Corchorus olitorius and C. capsularis resulted in identification of 20 different motifs where β1 and γ1, and all members of δ1 classes in both Corchorus species represented the highest number of motifs [1]. Tissue-specific analysis of δ2, α2, and β1 subclasses revealed their roles involved in drought and salinity stresses. Genome-wide analysis of PLD gene family members in tree cotton (Gossypium arboretum) revealed 19 non-redundant genes with similar exon-intron architecture and motifs with highly conserved structures. There are 59 PLD gene members in alfalfa (Medicago sativa L.) grouped into ε, δ, γ, β, α, and ζ subclasses [3]. It has been demonstrated that cis-acting elements are different in PLD members within common subfamilies, but they represent similar physicochemical characteristics, sequence feature and domains. The PLD protein family has, however, not been bioinformatically analyzed in important crops within Brassicaceae family, including rapeseed (Barssica rapa L.) and camelina (Camelina sativa L.) and to date, no genome-wide comparative analysis of PLD gene family has been carried out in these oilseed crops. In the present study, we identified and compared PLD gene members from the whole genome of Brassica rapa and Camelina sativa. The available bioinformatics tools were applied to analyze various characteristics of PLD genes and their expression levels were analyzed in response to salinity stress. Our findings provide a better understanding of the evolution and structure of sequence and function of PLDs, which will be useful in future functional studies.
2. Materials and Methods
2.1. Identification and Characterization of PLD Family Members
In order to identify the members of the PLD gene family, the known sequence of these genes in Arabidopsis was run as queries against the genome of Brassica napus and Camelina sativa, using BLASTp tool of Ensembl Plants database [20]. The peptide sequences of the PLDs in Brassica napus (BnPLDs) and Camelina sativa (CsPLDs) were checked by CCD database [21] for confirmation. In addition, PLDs were identified in Glycine max and Oryza sativa similar to the previous method. In this study, the physicochemical properties, including number of amino acids, isoelectric point (pI), grand average of hydropathicity (GRAVY), molecular weight (MW), and instability index were enumerated for the candidate PLDs, using the ProtParam tools [22].
2.2. Phylogenetic Analysis of PLDs and Prediction of Phosphorylation Sites
To investigate the evolution of the PLD gene family, peptide sequences of CsPLDs and BnPLDs with the PLD characterized in Oryza sativa, Glycine max, and Arabidopsis thaliana were aligned using online tool, the Clustal Omega [23]. In the next step, the aligned sequences were introduced to IQ-TREE web server [24] to construct a phylogeny tree based on the Maximum likelihood (ML) method with 1000 bootstrap. Finally, the phylogeny tree of PLDs was visualized by Interactive Tree Of Life (iTOL) v6.3 [25]. In addition, the strong potential phosphorylation sites (potential value ≥ 0.90) of CsPLDs and BnPLDs were identified using the NetPhos 3.1 server [26].
2.3. Duplication Analysis of CsPLD and BnPLD Genes
In the present study, the coding sequence (CDS) of pairs of CsPLD and BnPLD genes were aligned and of pairs with an identity ≥0.85 were selected as a duplicated genes [27,28]. In addition, the synonymous (Ks) and non-synonymous (Ka) indexes were calculated for duplicated pairs using TBtools software [29]. Besides, the time of divergence for each duplicated pairs was estimated using the following equation, T = (Ks/2λ) × 10−6. (λ = 6.5 × 10−9).
2.4. Conserved Motifs and Promoter Analysis
To recognize the conserved protein motifs, CsPLDs and BnPLDs were analyzed using the MEME (version 5.0.5) [30], based on the default settings. Moreover, an upstream region (1500 bp) of the start codon in each CsPLD and BnPLD was screened, as a promoter site, to identify the putative cis- regulatory elements using the PlantCARE database [31].
2.5. Interaction Networks and Structure Analysis of CsPLD and BnPLD
In the current study, the interaction networks for CsPLDs and BnPLDs were constructed using the STRING database (v11.5) [32]. To better understand direct and indirect interactions, the first and second shell were adjusted to ≤ 20 and ≤ 5, respectively. Finally, the interaction networks were visualized using Cytoscape v3.8.2 [33]. In order to draw the three-dimensional structure, five CsPLDs and BnPLD proteins (one candidate from each subfamily) were selected and their structure of these proteins were predicted using the Phyre2 server [34].
2.6. Expression Analysis of CsPLD and BnPLD in Different Tissues
In the current study, the available transcriptome data (RNA-seq information) related to different stages of growth and development for B. napus and C. sativa were used to screen the expression profile of the BnPLD and CsPLD genes. The Camelina eFP Browser (https://bar.utoronto.ca/efp_camelina/cgi-bin/efpWeb.cgi) was used for CsPLD genes and the BrassicaEDB was used for BnPLD genes to extract expression data in terms of FPKM (fragments per kilobase of exon model per million mapped reads) value. Finally, data were illustrated in heatmaps based on the log2 transformed method using the TBtools software.
2.7. Plant Materials and Treatment
In this study, expression patterns of candidate CsPLD genes were evaluated in camelina seedlings under salt stress. In the first step, sterilized seeds were planted in pots containing peat moss and perlite (2:1) and were kept under a temperature of 24±2 °C and 16 hours of light and pots were irrigated every three days. Then, after five weeks, seedlings were irrigated with salt (200 mM of NaCl) and was repeated after 24 hours. Finally, leaves were harvested at different time points (6, 24, and 72 h after treatment). The experiment was carried out in three biological replicates.
2.8. Expression Analysis Using qPCR
The total RNA samples from leaves were extracted by an RNX kit (Sinaclon, Iran) according to manufacturer protocols and the cDNA was produced by a reverse transcriptase kit (Roche, Germany), based on manufacturer protocols. In this study, six CsPLD genes were selected for qPCR analysis based on phylogenetic analysis, one member from delta, beta, zeta, and gamma subfamily and two members from alpha subfamily. Besides, actin-2 gene (Csa15g026420) was selected as a housekeeping gene for normalizing the raw expression data. The Primer3 online tool (version 4.1.0) [35] was applied to design the specific primers of candidate CsPLDs (Table S1). Maxima SYBR Green/ROX qPCR Master Mix kit (Thermo Fisher, France) was used to analyze the expression levels of the CsPLDs by the ABI Step One, according to manufacturer protocols. Finally, the raw data were analyzed by the delta delta ct method [36].
3. Results
3.1. Physiochemical Properties of PLDs
In this study, 33 CsPLDs and 41 BnPLDs were recognized in genome of Camelina sativa and Brassica napus, respectively (Table S2). PLDs were compared in five plant species that diverse physiochemical properties were observed between PLDs (Table 1). For instance, 33% of CsPLDs were predicted as stable protein, while 61% of BnPLDs were stable. All studied PLDs showed a negative GRAVY value, indicating that PLDs are probably hydrophilic proteins. Based on the exon number, less exon number was recognized in alpha subfamily members.
3.2. Phylogenetic Analysis and Prediction of Post-Translational Modifications of PLD Gene Family
Phylogenetic analysis of PLD gene family in five plant species, including C. sativa, Arabidopsis, B. napus, O. sativa, and G. max could separate the five PLD subfamilies, including gamma, delta, beta, alpha, and zeta (Figure 1). According to phylogeny, subfamilies gamma, delta, and beta are more closely related. In addition, the most of PLDs were in alpha subfamily group. CsPLDs showed more close relationships to Arabidopsis PLDs than oilseed crops. The potential phosphorylation modifications, as an important post-translational modification, were predicted in PLD proteins (Figure 1). Alpha subfamily members showed less potential sites than other subfamilies. It seems that alpha proteins are less influenced by post-translational modifications.
3.3. Conserved Motifs in PLDs
In this study to investigate the structure of PLDs, the conserved motifs of the CsPLDs and BnPLDs were identified. A total of 12 motifs were detected that motifs 1, 6, 10, 5, 4, 3, and 2 were observed in almost of PLDs (Figure 2). However, the members of zeta subfamily were different from other PLD subfamilies based on the distribution of conserved motifs. For instance, motifs 8, 9, 11, and 12 were observed in subfamilies alpha, beta, delta, and gamma. Structural differences between PLDs reveal the functional specificity between different subfamilies. On the other hand, the PLD members that have the same pattern of motif distribution probably had a similar evolutionary process and have a common ancestor.
3.4. Duplication Events in PLD Gene Family
The CsPLDs and BnPLD genes were unevenly located on chromosomes of C. sativa and B. napus (Figure 3 and Table S3). For the genome of C. sativa, the highest number of CsPLDs were distributed on chromosomes 1, 8, 13, 15, and 19 (Figure 3a). BnPLDs were mainly distributed on chromosomes A03, A05, A06, C03, and C07 (Figure 3b). However, the chromosomal location for 9 genes from B. napus is unknown, presented with An and Cn in the Figure 3b. Duplication events between PLDs revealed that many duplication events have occurred in PLD gene family from C. sativa and B. napus (Figure 3 and Table S3). Besides, the results disclosed that all CsPLD genes have been segmentally duplicated, while in BnPLDs, three tandem duplications were observed. According to ka/ks index, selective pressure of all studied PLDs was purifying. Divergence time of duplication events estimated ranging 1.38 to 8.21 and 1.56 to 219.36 MYA in CsPLDs and BnPLDs, respectively.
3.5. Promoter Analysis
The upstream of CsPLDs and BnPLDs were screened to identify the putative cis-regulatory elements having roles in stress and defense responsiveness, hormone responsiveness, protein binding site, and plant growth and light responsiveness (Figure 4a,b; Table S4). Cis-regulatory elements related to hormones were more observed in promoter region of CsPLDs, while in BnPLDs, cis-regulatory elements (about 44%) had roles in stress responses. The promoter elements related to ABA and auxin responsiveness were more distributed in CsPLDs, while elements linked with MeJA responsiveness were more observed in promoter region of BnPLDs. This is suggestive of the participation of CsPLDs in response under abiotic stress and BnPLDs are associated with defense responses against biotic stresses by inducing the MeJA mechanism.
3.6. Interaction Network of CsPLDs and BnPLDS
In order to predict the interactions of PLDs with other proteins, an interaction network for PLDs in camelina and B. napus was constructed based on the available data (Figure 5a,b). As presented in Figure a, CsPLDs showed interactions with non-specific phospholipase C (nsPLC), guanine nucleotide-binding protein (GNBP), 1-acyl-sn-glycerol-3-phosphate acyltransferase (AGPAT), diacylglycerol kinase (DAGK), and phosphatase 2C. Besides, BnPLDs interacted with nsPLC, phosphatidic acid phosphohydrolase 2 (PAH2), AGPAT, lipid phosphate phosphatase 2 (LPP2), lysophosphatidyl acyltransferase 2 (LPAT2), monogalactosyldiacylglycerol synthase type C (MGD3), phosphatidic acid phosphatase (PAP2), alpha/beta-Hydrolases, and phosphatidyl-N-methylethanolamine (Figure 3b). PLDs strongly showed interactions with proteins that are involved in phosphonate and phosphinate metabolism, ether lipid metabolism, phosphatidic acid metabolic, and inositol phosphate metabolism. These results disclose that PLDs can be involved in various pathways in response to upstream signals. However, interaction network of CsPLDs and BnPLDs was somewhat different.
3.7. 3D Structure and Pocket Sites of Candidate PLD Subfamilies
Three dimensional structure of each subfamily of PLD gene family was predicted based on homology. Results revealed that PLDs in C. sativa and B. napus have similar 3D structure, while they showed different binding points according to the location and type of sequence in the pocket region (Figure 6). Based on the abundance of amino acids in the pocket region, PRO, TRP, LEU, and GLU were more present in zeta subfamily of BnPLD, while TRP and GLU more observed in CsPLD zeta. Moreover, LEU, and GLU in BnPLD alpha and GLY in CsPLD alpha were more present in pocket site. Interestingly, ALA was frequently distributed in pocket site of beta subfamily of both BnPLD and CsPLD. In candidate gamma subfamily proteins, LYS, GLN, and ALA in BnPLD and SER, and ASN in CsPLD were more observed. LYS and THR were highly distributed in BnPLD delta and CsPLD delta, respectively.
3.8. Expression Profile of PLDs in Different Tissues and Organs
Expression levels of PLDs were investigated in different tissues and organs of C. sativa and B. napus (Figure 7 and Figure 8). In C. sativa, three CsPLD genes from alpha subfamily including Csa15g020390, Csa19g022540, and Csa01g018370, showed high expression in all tissues and organs (Figure 7). Besides, two genes, Csa13g049040 and Csa02g016220, from gamma subfamily were more expressed in senescing leaf tissues. In B. napus, five genes from alpha subfamily, including BnaC01g35830D, BnaA01g28530D, BnaA01g28540D, BnaC05g37540D, and BnaA05g23740D, showed high expression in all studied tissues and organs, except for anther tissues (Figure 8). A delta subfamily gene, BnaA08g04540D, and two beta subfamily genes, BnaC05g00230D and BnaA10g00160D, were more expressed in anther. In bud tissues of B. napus, two genes from alpha subfamily, BnaC03g69220D and BnaA05g35700D, were more induced.
3.9. Expression Profile of CsPLD in Response to Salt Stress
To better understand the potential roles of the PLDs in camelina, the expression profile of six candidate CsPLD genes were investigated in response to 200 mM of NaCl, as salinity stress. The results indicate that CsPLD candidate genes are involved in the response of Camelina seedlings to salt stress, although they had different expression patterns (Figure 9). PLD beta1 (Csa06g048050) and PLD delta (Csa10g006270) showed a similar expression pattern, so that they their expression levels were upregulated after 24 hours of salt stress and showed the highest level of expression after 72 hours of salt stress. The expression pattern of one PLD gamma gene (Csa13g049040) was significantly down-regulated after 6 hours and its expression level was not significantly induced by salinity. Besides, the expression level of Csa01g006520 (which is a PLD zeta) was reduced early stages of salt stress (after 6 and 24 hours), while its expression was increased after 72 hours. Two members of the alpha subfamily presented a different expression pattern after 6 hours of salt stress, alpha 4 (Csa03g061750) increased expression and alpha 1 (Csa15g020390) decreased expression, while their expression levels of both genes were increased after 72 hours.
4. Discussion
Oilseed plants such as rapeseed play an important role in the production of healthy oils in the food basket. Camelina, as an almost new agricultural plant, also has a good percentage of healthy oil and also has high resistance to adverse environmental conditions. Besides this, phospholipases have a key role in regulating cellular signaling pathways in response to stresses. PLD, as a key element linked in lipid mediated signaling, which involved in various physiological and developmental processes [5,37]. In this study, two oilseed plants were compared based on the PLD gene family to gain a better understanding of their structures and functions. The results showed that PLD family has 41 members in B. napus and 33 members in C. sativa, which were more members compared to other plants such as rice, 17 PLDs, [13] and Arabidopsis, 12 PLDs [38], 13 in maize [39] and 19 in potato [37]. Ploidy level and genome size can have an effect on the number of gene family members in plant species [40]. Evolutionary analysis was able to separate the subfamilies (gamma, delta, beta, alpha, and zeta) well. These findings suggest that PLDs in the same group share conserved sequences, disclosing that they can have similar functions. However, less relationship was observed between CsPLDs and BnPLDs. These results indicate that the evolutionary process of PLD gene family has been somewhat different in C. sativa and B. napus. Moreover, it seems that the ancestral gene of PLD family were formed before C. sativa and B. napus, and after the separation of the species, a different evolutionary process has been passed in each species [41]. In addition, the Ka/Ks value of all duplicated CsPLD and BnPLD pairs was estimated to be less than 1.0, revealing that the CsPLD and BnPLD family members survived under strong selection pressure of purification [42,43]. However, it was stated that purifying selection can generate pseudogenes and/or genes with conserved functions [44,45]. Besides, it seems that the expansion in CsPLDs happened much earlier than BnPLDs, and the evolution process of PLD was different in CsPLD and BnPLD family.
Signaling of phospholipases activates pathways related to growth and development and response to stresses [12,46]. Direct and indirect interactions can be effective in expanding the effect of phospholipases. By examining the interaction network, it was determined that the members of CsPLD and BnPLD family interact more with other elements of lipid-dependent cell signaling such as non-specific phospholipase, diacylglycerol kinase, 1-acyl-sn-glycerol-3-phosphate acyltransferase, phosphatidic acid phosphohydrolase, lipid phosphate phosphatase, lysophosphatidyl acyltransferase, and phosphatase. However, the interaction network of BnPLDs was estimated to be different from CsPLDs. It was previously reported that the interaction of PLD alpha1 with GPA is effective in increasing resistance to osmotic stress by affecting the opening and closing of stomata through ABA signaling pathway [47]. Moreover, it was stated that PLDs can participate in the response to abiotic and biotic stresses by cooperating with abscisic acid and jasmonic acid-dependent pathways [13,14,48]. The presence of cis-regulatory elements in the upstream region of PLD genes can strengthen this hypothesis, although more additional studies are needed. Post-translational modifications such as phosphorylation are effective in activating or deactivating target proteins [51]. The evaluation of post-translational changes in BnPLDs and CsPLDs showed that they have different potentials and thus can contribute in different ways. For example, less phosphorylation site was predicted in PLD alpha proteins, suggesting that this subfamily is less affected by upstream signaling elements.
The results of the expression profile of CsPLD and BnPLD genes in different tissues indicated that the members of this gene family are expressed in all tissues and developmental stages. A comparison between subfamilies revealed that the alpha subfamily has a greater range of activity. Also, the analysis of the structure of CsPLD and BNPLD genes disclosed that the members of alpha subfamily have fewer introns/exons. It has been reported that genes with fewer exons/introns are induced faster [45,52]. It seems that the smaller number of introns has an effect on the duration of the mRNA splicing process before leaving the nucleus, and as a result, they are translated earlier [43,53]. The quantitative findings under salt stress revealed that candidate CsPLDs were significantly induced. It was reported that PLDs are affected by abiotic stresses such as drought, high and low temperature, salt stress [37,46,54,55]. CsPLD gamma1, CsPLD zeta2, CsPLD alpha1, CsPLD alpha4 are recognized as early elements in response to salt stress. With the persistence of salinity stress, all studied CsPLD genes, except for CsPLD gamma1 gene, showed upregulation, suggesting that CsPLDs are probably involved in both the initial responses to activate the downstream pathways and in the resistance response to reduce the damages of the salinity stress.
5. Conclusions
Phospholipases D (PLD) is an enzyme that plays a crucial role in cell signaling, lipid metabolism, and membrane dynamics. In the current study, PLD gene family is characterized in Brassica napus and Camelina sativa. CsPLD and BnPLD have had different evolutionary processes, although both of them have been under almost similar evolutionary pressures, which has caused their structure to be conserved. Among the PLD subfamilies, members of the alpha subfamily have more obvious differences from the others, which can be considered more in future functional works. According to the expression data, PLDs were identified as widely expressed genes that can be involved in most of the developmental stages of plants. Our findings revealed that CsPLDs are involved in the response to high salinity stress and are probably active in a multi-branched interaction network in increasing resistance and reducing stress damages.
Supplementary Materials
Table S1: List of CsPLD primers used in qPCR analysis; Table S2: List of the identified PLDs and their physiochemical properties in Camelina sativa, and Brassica napus. Table S3: List of the duplicated CsPLDs and BnPLDs along with Ka and Ks values; Table S4: List of putative cis-regulatory elements related to various developmental, hormones and stress responsive pathways in CsPLDs and BnPLDs.
Author Contributions
Conceptualization, P.H. and F.P.; methodology, P.H., M.A. and F.P.; software, Y.V., F.P., M.E. and H.A.; writing—original draft preparation, P.H., Y.V. and M.A.; writing—review and editing, P.H. and Y.V. All authors have read and agreed to the published version of the manuscript.
Data Availability Statement
The data that support the findings of this study are available from the corresponding author, Parviz Heidari, upon reasonable request.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Sadat, M.A.; Ullah, M.W.; Hossain, M.S.; Ahmed, B.; Bashar, K.K. Genome-wide in silico identification of phospholipase D (PLD) gene family from Corchorus capsularis and Corchorus olitorius: Reveals their responses to plant stress. J. Genet. Eng. Biotechnol. 2022, 20. [CrossRef]
- Takáč, T.; Novák, D.; Šamaj, J. Recent advances in the cellular and developmental biology of phospholipases in plants. Front. Plant Sci. 2019, 10, 432553. [CrossRef]
- Yuan, Y.; Yu, J.; Kong, L.; Zhang, W.; Hou, X.; Cui, G. Genome-wide investigation of the PLD gene family in alfalfa (Medicago sativa L.): Identification, analysis and expression. BMC Genomics 2022, 23. [CrossRef]
- Sagar, S.; Singh, A. Emerging role of phospholipase C mediated lipid signaling in abiotic stress tolerance and development in plants. Plant Cell Rep. 2021, 40, 2123–2133. [CrossRef]
- Hong, Y.; Zhao, J.; Guo, L.; Kim, S.C.; Deng, X.; Wang, G.; Zhang, G.; Li, M.; Wang, X. Plant phospholipases D and C and their diverse functions in stress responses. Prog. Lipid Res. 2016, 62, 55–74. [CrossRef]
- Tang, K.; Dong, C.; Liu, J. Genome-wide analysis and expression profiling of the phospholipase D gene family in Gossypium arboreum. Sci. China. Life Sci. 2016, 59, 130–141. [CrossRef]
- Deepika, D.; Singh, A. Plant phospholipase D: Novel structure, regulatory mechanism, and multifaceted functions with biotechnological application. Crit. Rev. Biotechnol. 2022, 42, 106–124. [CrossRef]
- Iakimova, E.T.; Michaeli, R.; Woltering, E.J. Involvement of phospholipase D-related signal transduction in chemical-induced programmed cell death in tomato cell cultures. Protoplasma 2013, 250, 1169–1183. [CrossRef]
- Pleskot, R.; Li, J.; Žárský, V.; Potocký, M.; Staiger, C.J. Regulation of cytoskeletal dynamics by phospholipase D and phosphatidic acid. Trends Plant Sci. 2013, 18, 496–504. [CrossRef]
- Pejchar, P.; Sekereš, J.; Novotný, O.; Žárský, V.; Potocký, M. Functional analysis of phospholipase Dδ family in tobacco pollen tubes. Plant J. 2020, 103, 212–226. [CrossRef]
- Mancuso, S.; Marras, A.M.; Mugnai, S.; Schlicht, M.; Žársky, V.; Li, G.; Song, L.; Xue, H.W.; Baluška, F. Phospholipase Dζ2 Drives Vesicular Secretion of Auxin for Its Polar Cell-Cell Transport in the Transition Zone of the Root Apex. Plant Signal. Behav. 2007, 2, 240. [CrossRef]
- Distéfano, A.M.; Scuffi, D.; García-Mata, C.; Lamattina, L.; Laxalt, A.M. Phospholipase Dδ is involved in nitric oxide-induced stomatal closure. Planta 2012, 236, 1899–1907. [CrossRef]
- Li, G.; Lin, F.; Xue, H.W. Genome-wide analysis of the phospholipase D family in Oryza sativa and functional characterization of PLDβ1 in seed germination. Cell Res. 2007 1710 2007, 17, 881–894. [CrossRef]
- Wang, C.; Zien, C.A.; Afitlhile, M.; Welti, R.; Hildebrand, D.F.; Wang, X. Involvement of Phospholipase D in Wound-Induced Accumulation of Jasmonic Acid in Arabidopsis. Plant Cell 2000, 12, 2237. [CrossRef]
- Wang, X. Regulatory Functions of Phospholipase D and Phosphatidic Acid in Plant Growth, Development, and Stress Responses. Plant Physiol. 2005, 139, 566–573. [CrossRef]
- Wang, X. Plant phospholipases. Annu. Rev. Plant Biol. 2001, 52, 211–231.
- Liu, Q.; Zhang, C.; Yang, Y.; Hu, X. Genome-wide and molecular evolution analyses of the phospholipase D gene family in Poplar and Grape. BMC Plant Biol. 2010, 10. [CrossRef]
- Wang, X.; Xu, L.; Zheng, L. Cloning and expression of phosphatidylcholine-hydrolyzing phospholipase D from Ricinus communis L. J. Biol. Chem. 1994, 269, 20312–20317. [CrossRef]
- Du, D.; Cheng, T.; Pan, H.; Yang, W.; Wang, J.; Zhang, Q. Genome-wide identification, molecular evolution and expression analyses of the phospholipase D gene family in three Rosaceae species. Sci. Hortic. (Amsterdam). 2013, 153, 13–21. [CrossRef]
- Bolser, D.; Staines, D.M.; Pritchard, E.; Kersey, P. Ensembl Plants: Integrating Tools for Visualizing, Mining, and Analyzing Plant Genomics Data. Methods Mol. Biol. 2016, 1374, 115–140. [CrossRef]
- Marchler-Bauer, A.; Derbyshire, M.K.; Gonzales, N.R.; Lu, S.; Chitsaz, F.; Geer, L.Y.; Geer, R.C.; He, J.; Gwadz, M.; Hurwitz, D.I.; et al. CDD: NCBI’s conserved domain database. Nucleic Acids Res. 2015, 43, D222–D226. [CrossRef]
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Duvaud, S.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. Protein identification and analysis tools on the ExPASy server. In The Proteomics Protocols Handbook; Humana Press: Totowa, NJ, 2005; pp. 571–607. [CrossRef]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [CrossRef]
- Nguyen, L.-T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating Maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, W256–W259. [CrossRef]
- Blom, N.; Sicheritz-Pontén, T.; Gupta, R.; Gammeltoft, S.; Brunak, S. Prediction of post-translational glycosylation and phosphorylation of proteins from the amino acid sequence. Proteomics 2004, 4, 1633–1649. [CrossRef]
- Musavizadeh, Z.; Najafi-Zarrini, H.; Kazemitabar, S.K.; Hashemi, S.H.; Faraji, S.; Barcaccia, G.; Heidari, P. Genome-Wide Analysis of Potassium Channel Genes in Rice: Expression of the OsAKT and OsKAT Genes under Salt Stress. Genes (Basel). 2021, 12, 784. [CrossRef]
- Puresmaeli, F.; Heidari, P.; Lawson, S. Insights into the Sulfate Transporter Gene Family and Its Expression Patterns in Durum Wheat Seedlings under Salinity. Genes (Basel). 2023, 14, 333. [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [CrossRef]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [CrossRef]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein–protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [CrossRef]
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J.E. The Phyre2 web portal for protein modeling, prediction and analysis. Nat. Protoc. 2015, 10, 845–858. [CrossRef]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3--new capabilities and interfaces. Nucleic Acids Res. 2012, 40. [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [CrossRef]
- Li, L.; Zhang, C.; Zhang, M.; Yang, C.; Bao, Y.; Wang, D.; Chen, Y.; Chen, Q. Genome-Wide Analysis and Expression Profiling of the Phospholipase D Gene Family in Solanum tuberosum. Biology (Basel). 2021, 10. [CrossRef]
- Qin, C.; Wang, X. The Arabidopsis Phospholipase D Family. Characterization of aCalcium-Independent and Phosphatidylcholine-Selective PLDζ1 withDistinct Regulatory Domains. Plant Physiol. 2002, 128, 1057. [CrossRef]
- Chen, L.; Cao, B.; Han, N.; Tao, Y.; Zhou, S.F.; Li, W.C.; Fu, F.L. Phospholipase D family and its expression in response to abiotic stress in maize. Plant Growth Regul. 2017, 81, 197–207. [CrossRef]
- Faraji, S.; Ahmadizadeh, M.; Heidari, P. Genome-wide comparative analysis of Mg transporter gene family between Triticum turgidum and Camelina sativa. BioMetals 2021, 4. [CrossRef]
- Gao, X.; Liu, J.; Yu, L.; Yuan, B.; … X.W. Genome-wide identification and expression analysis of APX gene family in Taraxacum kok-saghyz. Acta Bot.-Boreali-Occident. Sin. 2019, 39, 1935–1942.
- Juretic, N.; Hoen, D.R.; Huynh, M.L.; Harrison, P.M.; Bureau, T.E. The evolutionary fate of MULE-mediated duplications of host gene fragments in rice. Genome Res. 2005, 15, 1292–1297. [CrossRef]
- Faraji, S.; Filiz, E.; Kazemitabar, S.K.; Vannozzi, A.; Palumbo, F.; Barcaccia, G.; Heidari, P. The AP2/ERF Gene Family in Triticum durum: Genome-Wide Identification and Expression Analysis under Drought and Salinity Stresses. Genes (Basel). 2020, 11, 1464. [CrossRef]
- Wang, Y.; Tang, H.; DeBarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.; Jin, H.; Marler, B.; Guo, H. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49–e49. [CrossRef]
- Heidari, P.; Hasanzadeh, S.; Faraji, S.; Ercisli, S.; Mora-Poblete, F. Genome-Wide Characterization of the Sulfate Transporter Gene Family in Oilseed Crops: Camelina sativa and Brassica napus. Plants 2023, 12, 628. [CrossRef]
- Liu, B.; Yao, L.; Wang, W.; Gao, J.; Chen, F.; Wang, S.; Xu, Y.; Tang, L.; Jia, Y. Molecular cloning and characterization of phospholipase D from Jatropha curcas. Mol. Biol. Rep. 2010, 37, 939–946. [CrossRef]
- McDermott, M.I.; McDermott, M.; Wakelam, M.J.; Morris, A.J. Phospholipase d. cdnsciencepub.comM McDermott, MJO Wakelam, AJ MorrisBiochemistry Cell Biol. 2004•cdnsciencepub.com 2004, 82, 225–253.
- Ben Othman, A.; Ellouzi, H.; Planchais, S.; Vos, D. De; Faiyue, B. Phospholipases Df1 and Df2 have distinct roles in growth and antioxidant systems in Arabidopsis thaliana responding to salt stress. Planta 2017, 246, 721–735. [CrossRef]
- Guo L., Devaiah S.P., Narasimhan R., Pan X., Zhang Y., Zhang W., et al. Cytosolic Glyceraldehyde-3-Phosphate Dehydrogenases Interact with Phospholipase DΔ to Transduce Hydrogen Peroxide Signals in the Arabidopsis Response to. Plant Cell 2012, 24, 2200–2212. [CrossRef]
- Wang, P.; Shen, L.; Guo, J.; Jing, W.; Qu, Y.; Li, W.; … R.B.-T.P.; 2019, undefined Phosphatidic acid directly regulates PINOID-dependent phosphorylation and activation of the PIN-FORMED2 auxin efflux transporter in response to salt stress. Acad. Wang, L Shen, J Guo, W Jing, Y Qu, W Li, R Bi, W Xuan, Q Zhang, W ZhangThe Plant Cell, 2019•academic.oup.com. [CrossRef]
- Ahmadizadeh, M.; Rezaee, S.; Heidari, P. Genome-wide characterization and expression analysis of fatty acid desaturase gene family in Camelina sativa. Gene Reports 2020, 21, 100894. [CrossRef]
- Yaghobi, M.; Heidari, P. Genome-Wide Analysis of Aquaporin Gene Family in Triticum turgidum and Its Expression Profile in Response to Salt Stress. Genes (Basel). 2023, 14, 202. [CrossRef]
- Arab, M.; Najafi Zarrini, H.; Nematzadeh, G.; Heidari, P.; Hashemipetroudi, S.H.; Kuhlmann, M. Comprehensive Analysis of Calcium Sensor Families, CBL and CIPK, in Aeluropus littoralis and Their Expression Profile in Response to Salinity. Genes (Basel). 2023, 14, 753. [CrossRef]
- Darwish, E.; Testerink, C.; Khalil, M.; El-Shihy, O.; Munnik, T. Phospholipid Signaling Responses in Salt-Stressed Rice Leaves. Plant Cell Physiol. 2009, 50, 986–997. [CrossRef]
- Li, W.; Li, M.; Zhang, W.; Welti, R.; Wang, X. The plasma membrane-bound phospholipase Ddelta enhances freezing tolerance in Arabidopsis thaliana. Nat. Biotechnol. 2004, 22, 427–433. [CrossRef]
Figure 1.
Phylogenetic tree of PLD gene family in five plant species. The potential phosphorylation sites in PLD proteins. Subfamilies of PLD including gamma, delta, beta, alpha, and zeta are highlighted around phylogenetic tree. PLDs from C. sativa (started with Csa), Arabidopsis (started with AT), B. napus (started with Bna), O. sativa (started with Os), G. max (started with GLY) are studied.
Figure 1.
Phylogenetic tree of PLD gene family in five plant species. The potential phosphorylation sites in PLD proteins. Subfamilies of PLD including gamma, delta, beta, alpha, and zeta are highlighted around phylogenetic tree. PLDs from C. sativa (started with Csa), Arabidopsis (started with AT), B. napus (started with Bna), O. sativa (started with Os), G. max (started with GLY) are studied.
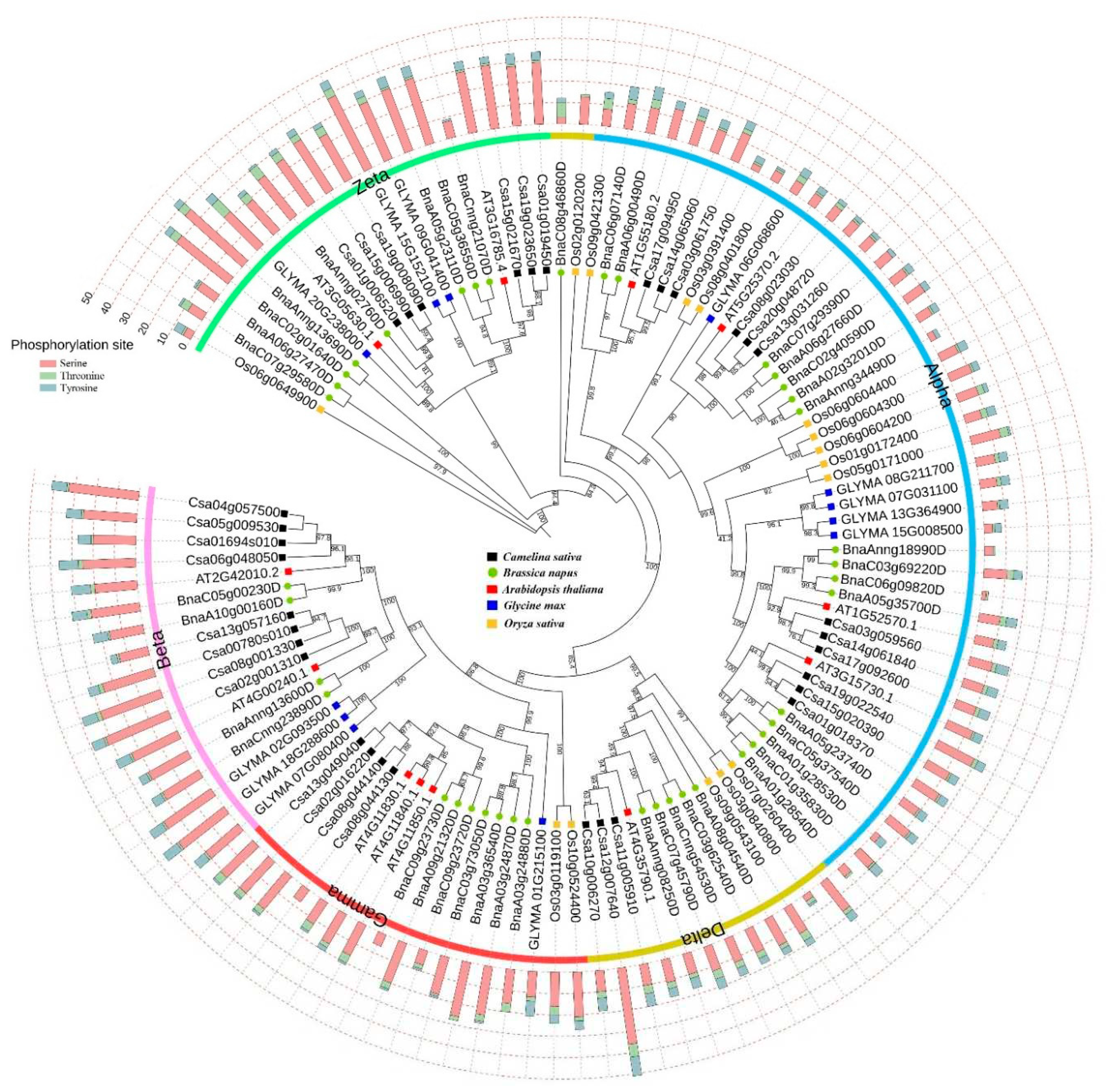
Figure 2.
Distribution the conserved motifs into protein sequence of PLD gene family. Subfamilies of PLD including gamma, delta, beta, alpha, and zeta are highlighted according to phylogeny analysis.
Figure 2.
Distribution the conserved motifs into protein sequence of PLD gene family. Subfamilies of PLD including gamma, delta, beta, alpha, and zeta are highlighted according to phylogeny analysis.

Figure 3.
Distribution of PLD family members in chromosomes of Camelina sativa (a), and Brassica napus (b). Gray lines show the duplicated genes. More details are presented in Table S3. .
Figure 3.
Distribution of PLD family members in chromosomes of Camelina sativa (a), and Brassica napus (b). Gray lines show the duplicated genes. More details are presented in Table S3. .
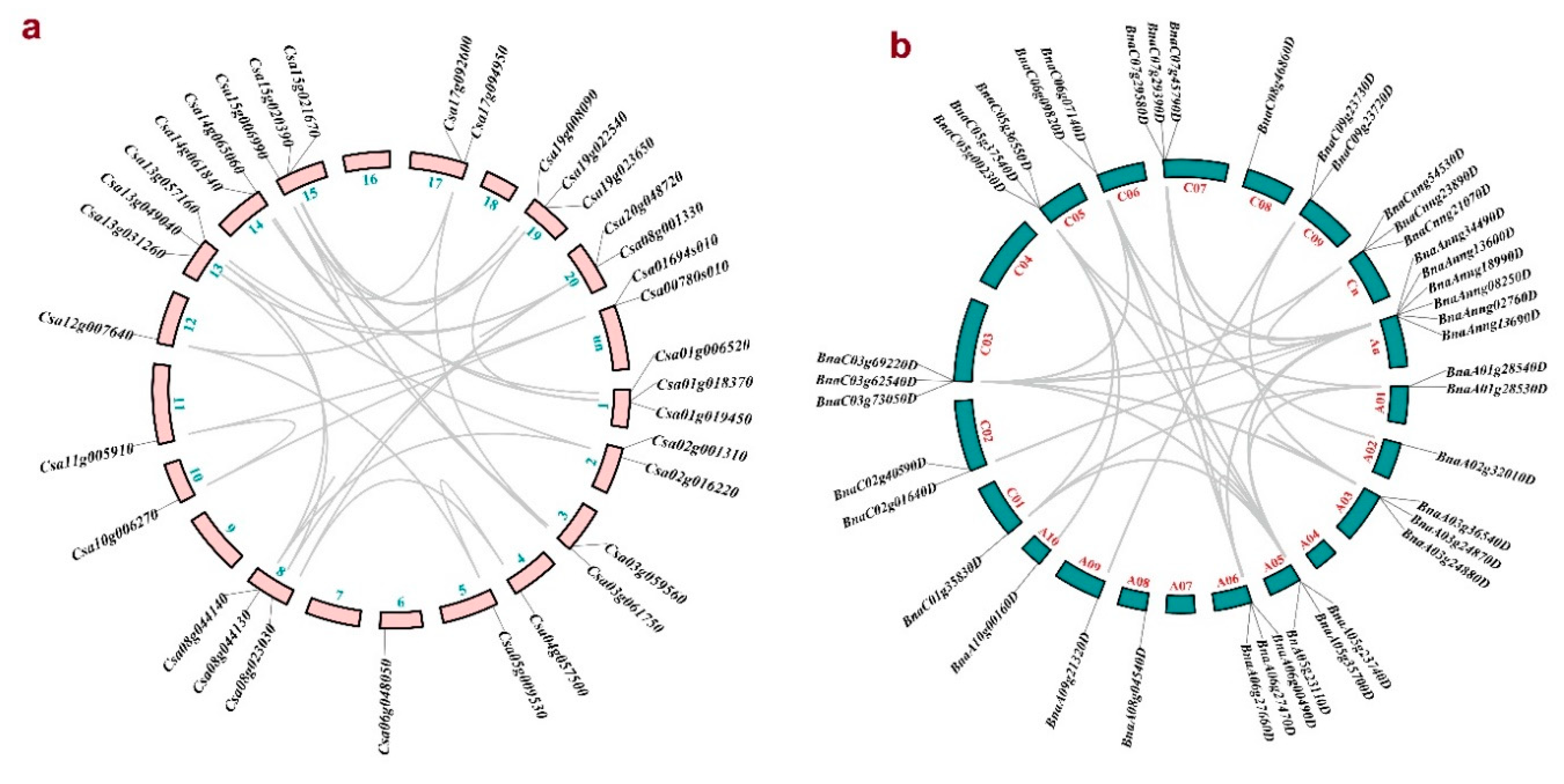
Figure 4.
Frequency of the putative cis-regulatory elements based their function in upstream region of CsPLDs (a) and BnPLDs (b). More details are presented in Table S4.
Figure 4.
Frequency of the putative cis-regulatory elements based their function in upstream region of CsPLDs (a) and BnPLDs (b). More details are presented in Table S4.
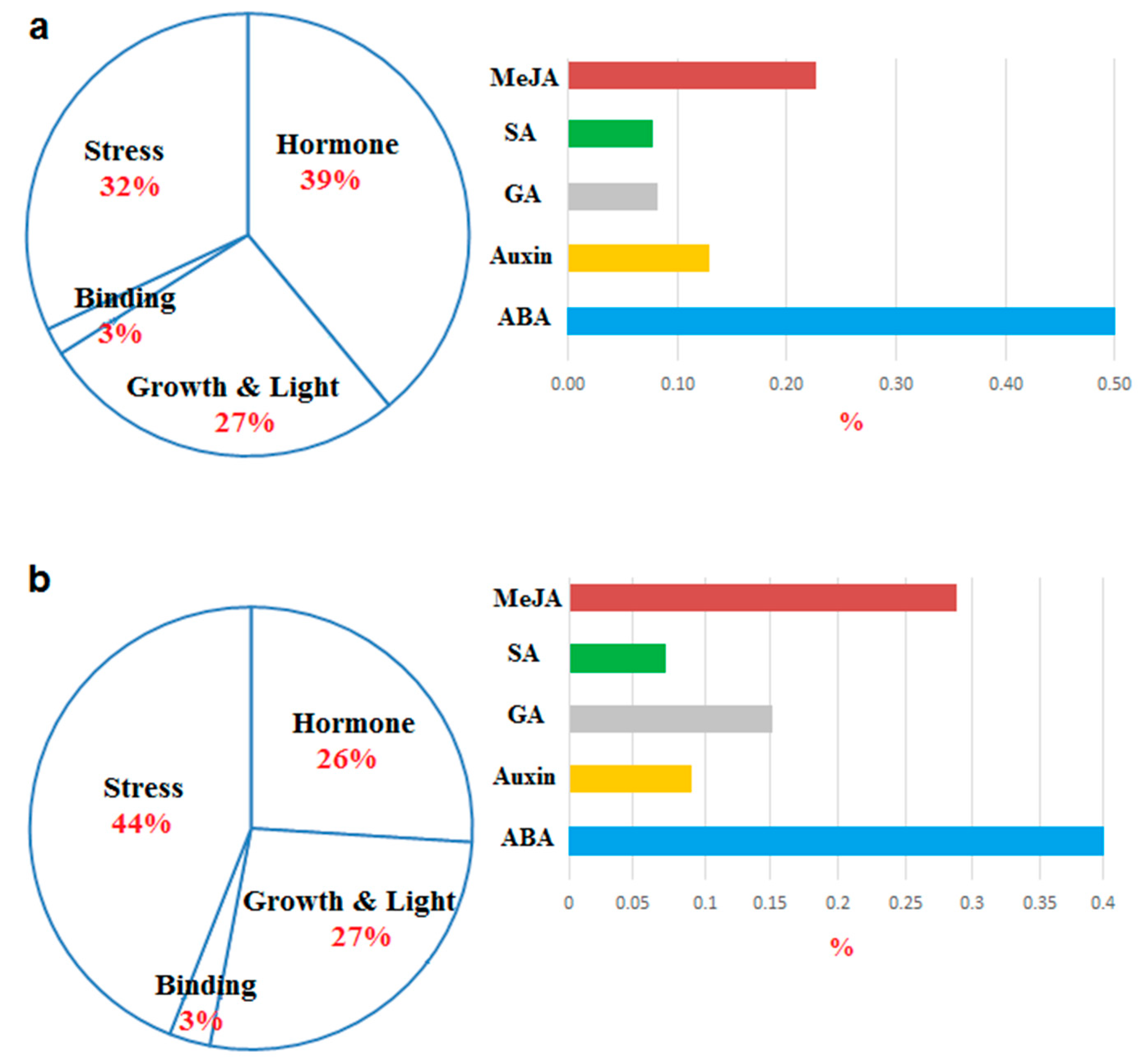
Figure 5.
Interaction network of CsPLDs (a), and BnPLDs (b). The yellow background is for PLD proteins. nsPLD: non-specific phospholipase; AGPAT: 1-acyl-sn-glycerol-3-phosphate acyltransferase; DAGK: diacylglycerol kinase; GNBP-a: guanine nucleotide-binding protein subunit alpha; GNBP-b: guanine nucleotide-binding protein subunit beta; PAH2: phosphatidic acid phosphohydrolase 2; LPP2: lipid phosphate phosphatase 2; LPAT2: lysophosphatidyl acyltransferase 2; MGD3: monogalactosyldiacylglycerol synthase type C; PAP2: phosphatidic acid phosphatase.
Figure 5.
Interaction network of CsPLDs (a), and BnPLDs (b). The yellow background is for PLD proteins. nsPLD: non-specific phospholipase; AGPAT: 1-acyl-sn-glycerol-3-phosphate acyltransferase; DAGK: diacylglycerol kinase; GNBP-a: guanine nucleotide-binding protein subunit alpha; GNBP-b: guanine nucleotide-binding protein subunit beta; PAH2: phosphatidic acid phosphohydrolase 2; LPP2: lipid phosphate phosphatase 2; LPAT2: lysophosphatidyl acyltransferase 2; MGD3: monogalactosyldiacylglycerol synthase type C; PAP2: phosphatidic acid phosphatase.
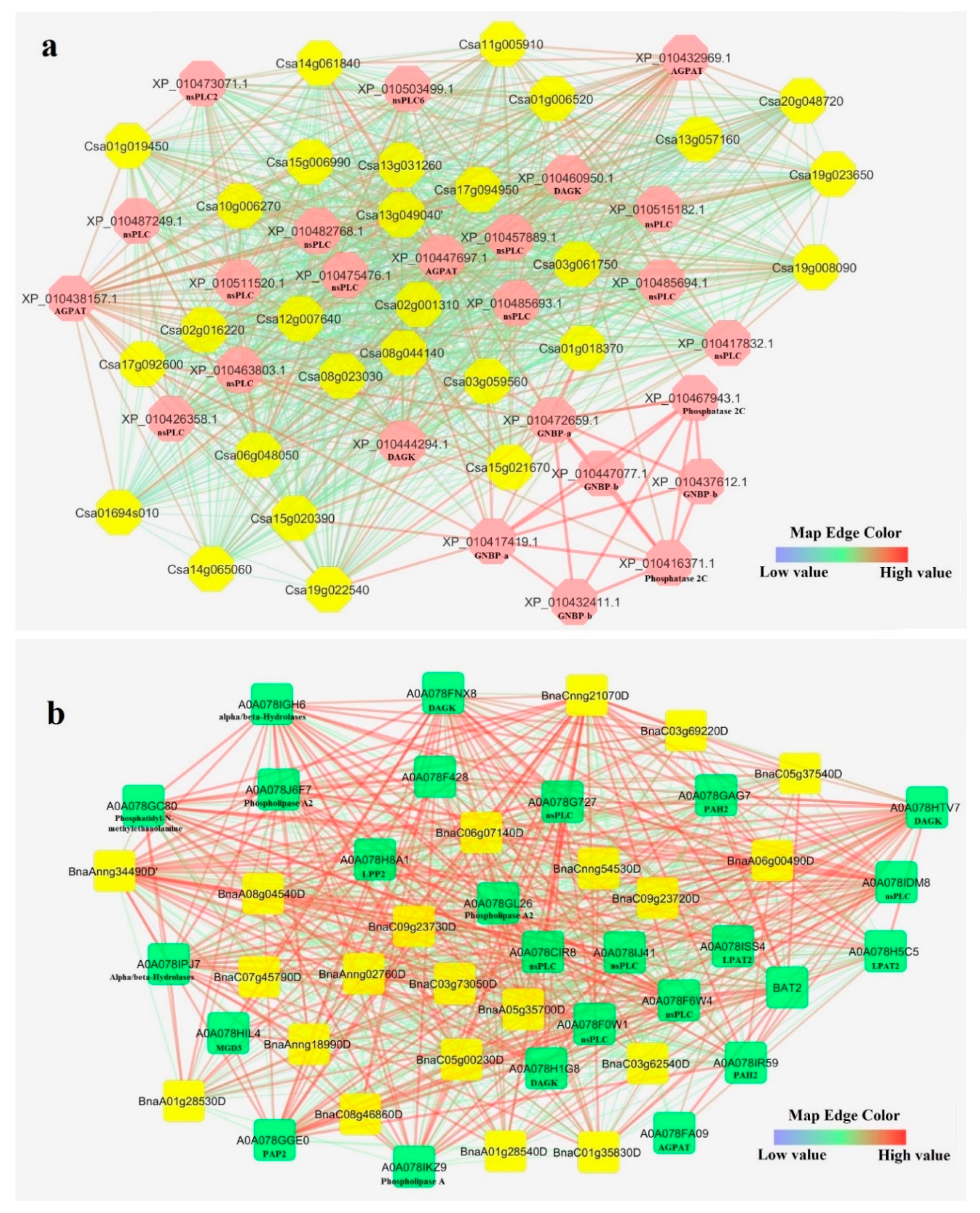
Figure 6.
3D structure and pocket sites of candidate PLD subfamilies. The binding residues are shown on protein structure.
Figure 6.
3D structure and pocket sites of candidate PLD subfamilies. The binding residues are shown on protein structure.

Figure 7.
Expression profile of CsPLDs in different tissues and developmental stages of C. sativa. Data were collected based on the available RNA-seq information.
Figure 7.
Expression profile of CsPLDs in different tissues and developmental stages of C. sativa. Data were collected based on the available RNA-seq information.
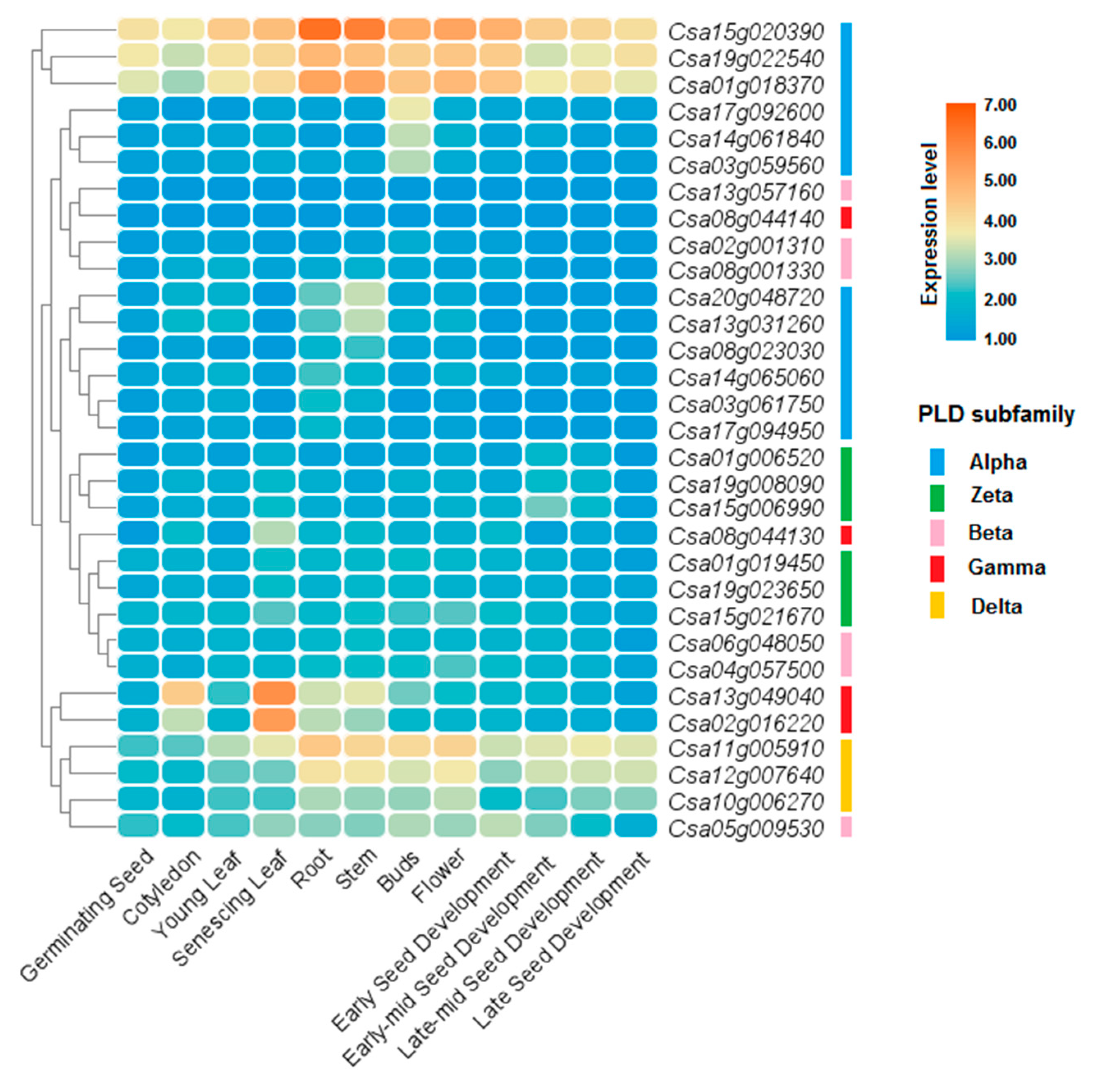
Figure 8.
Expression profile of BnPLDs in different tissues and developmental stages of B. napus. Data were collected based on the available RNA-seq information.
Figure 8.
Expression profile of BnPLDs in different tissues and developmental stages of B. napus. Data were collected based on the available RNA-seq information.
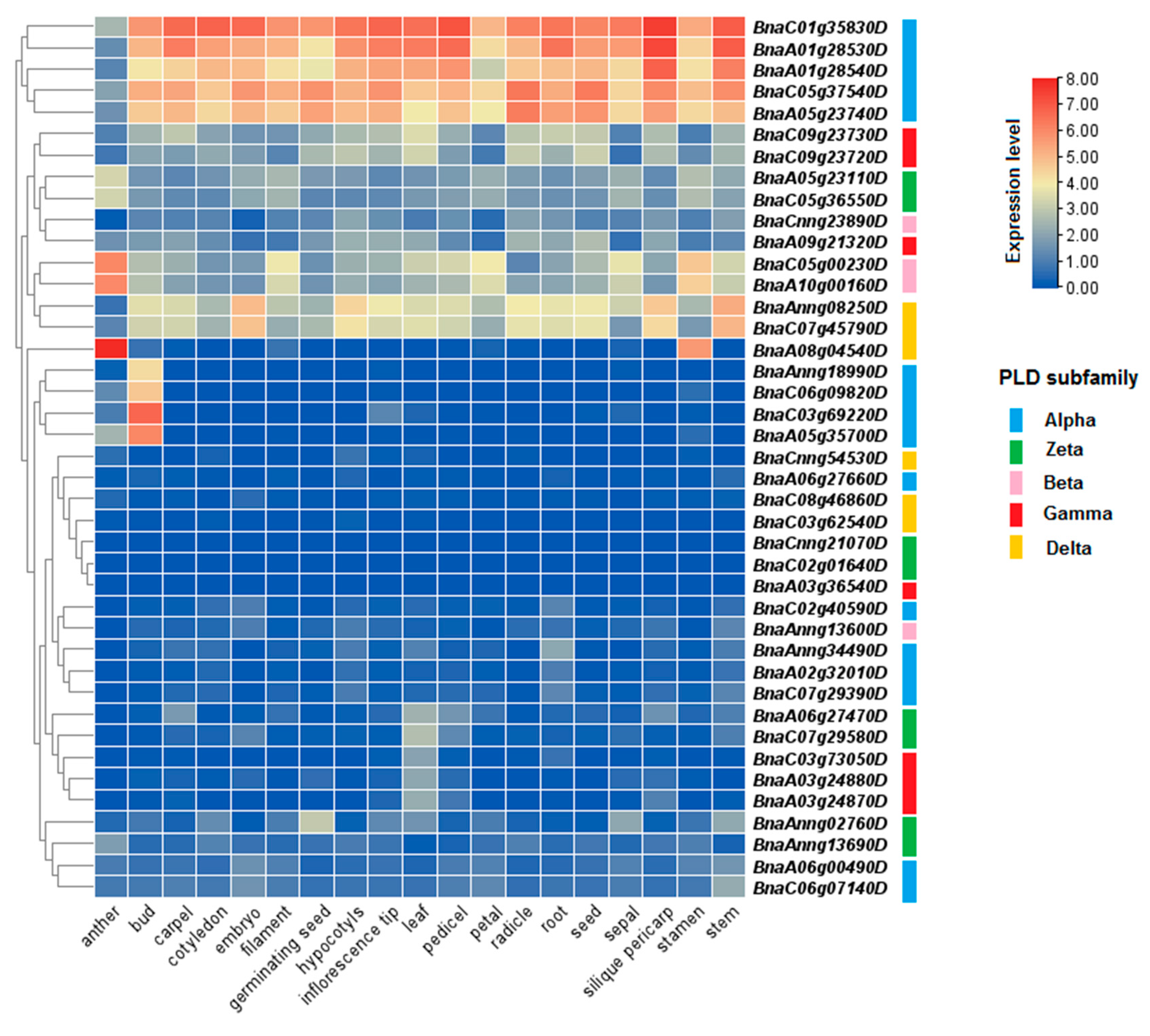
Figure 9.
Expression analysis of six CsPLDs in response to salinity stress. Asterisks, * and **, show significant differences P < 0.05 and P < 0.01, respectively, based on Student’s t-tests.
Figure 9.
Expression analysis of six CsPLDs in response to salinity stress. Asterisks, * and **, show significant differences P < 0.05 and P < 0.01, respectively, based on Student’s t-tests.
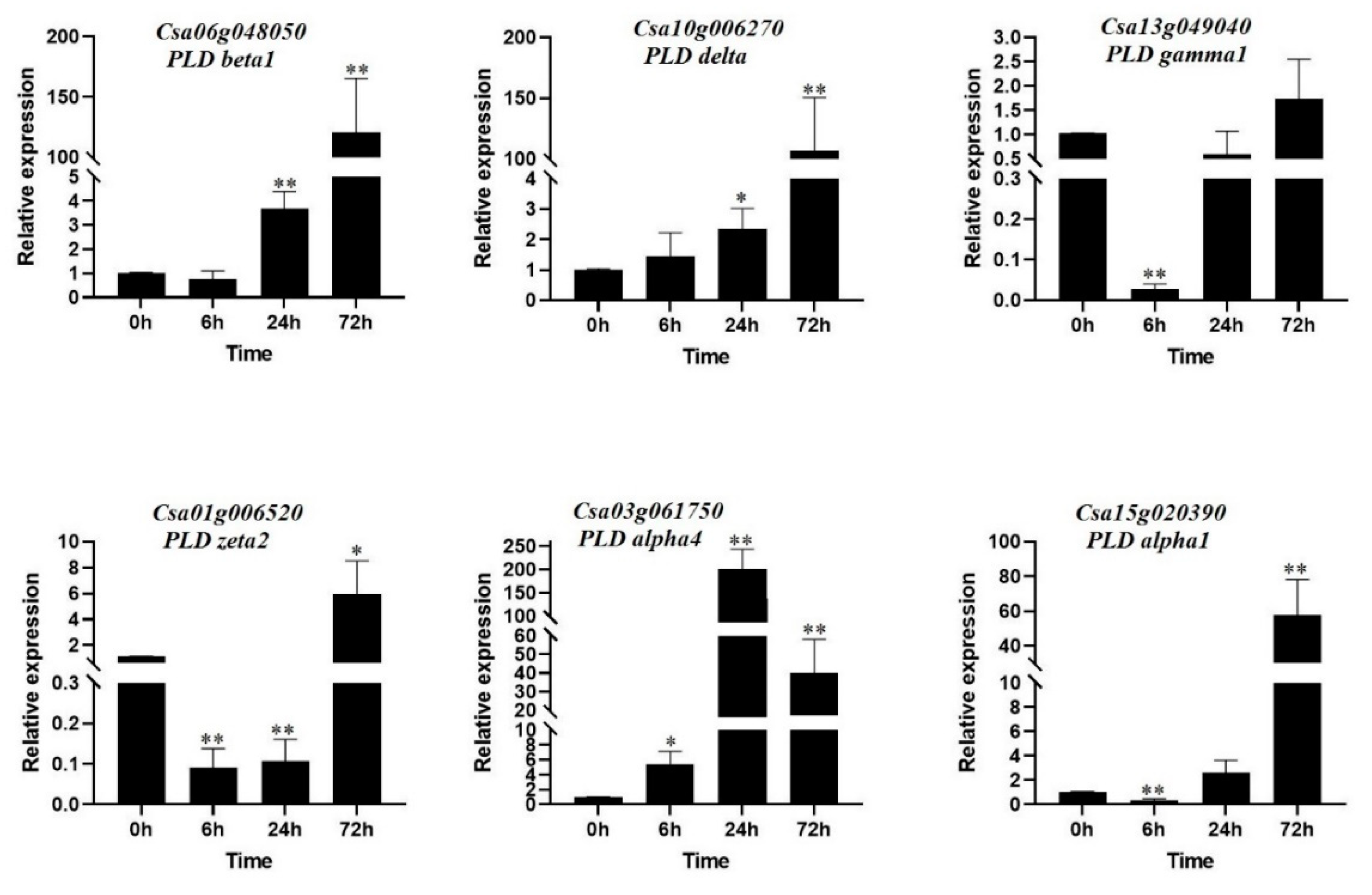

Table 1.
Summary of physiochemical properties of PLD family members in five plant species. Full details for CsPLDs and BnPLDs are presented in Tables S2.
Table 1.
Summary of physiochemical properties of PLD family members in five plant species. Full details for CsPLDs and BnPLDs are presented in Tables S2.
| Plant | Protein length | Exon number | MW (KDa) | pI | GRAVY | Stability |
|---|---|---|---|---|---|---|
| Camelina sativa | 350 - 1596 | 3 - 27 | 40.02 - 181.62 | 5.57 - 9.09 | -0.655 , -0.316 | 33% |
| Brassica napus | 213 - 1465 | 2 - 22 | 24.66 - 164.78 | 5.10 - 10.30 | -0.682 , -0.247 | 61% |
| Glycine max | 711 - 1126 | 3 - 22 | 81.22 - 127.63 | 5.48 - 7.65 | -0.532 , -0.374 | 25% |
| Arabidopsis thaliana | 762 - 1108 | 4 - 20 | 86.99 - 124.73 | 5.53 - 8.36 | -0.555 , -0.343 | 75% |
| Oryza sativa | 355 - 1046 | 1 - 12 | 39.31 - 116.93 | 5.56 - 11.23 | -0.622 , -0.147 | 67% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



