
Submitted:
12 September 2023
Posted:
13 September 2023
You are already at the latest version
Abstract
Previous studies on the mechanisms underlying willed actions reported that the premotor cortex may be involved in the construction of motor awareness. However, its exact role is still under investigation. Here we investigated the role of the dorsal premotor cortex (PMd) in motor awareness by modulating its activity applying inhibitory rTMS to PMd, before a specific motor awareness task (under three conditions: without stimulation, after rTMS and after sham stimulation). During the task, subjects had to trace straight lines to a given target, receiving a visual feedback of the line trajectories on a computer screen. Crucially, in most trials the trajectories on the screen were deviated and to produce straight lines, subjects had to correct their movements towards the opposite direction. After each trial, participants were asked to judge whether the line seen on the computer screen corresponded to the line actually drawn. Results show that participants in the no stimulation condition did not recognize the perturbation until 14 degrees of deviation. Importantly, active, but not sham, rTMS significantly modulated motor awareness, decreasing the amplitude of the angle at which participants became aware of the trajectory correction. These results suggest that PMd plays a crucial role in action self- monitoring.
Keywords:
Self-monitoring
; Motor Awareness
; Premotor Cortex
; TMS
; low-frequency
; Awareness
1. Introduction
Although many of the processes underlying motor programming and execution are not accessible to consciousness, we are aware that we are moving (motor awareness) and of our desire to act (motor intention). Blakemore et al. [1] proposed that, for intentional movements, motor commands are selected and sent to the muscles to perform the action, while at the same time a prediction is made about the sensory consequences of the movement. This prediction (indicated as forward model) is based on an efference copy of the planned motor act and is compared, by a comparator system, with the actual feedback of the executed movement. According to this proposal the forward model to be compared with the sensory feedbacks is the neural signal upon which motor awareness is constructed [2,3]. Therefore, motor awareness seems to precede, rather than follow, the actual execution of an intentional action being, within certain limits, dissociated from it. In a seminal experiment, Libet demonstrated that subjects become aware of a hand movement before the actual onset of muscle contraction [4], whereas Haggard and Magno [5] found that interfering, through single-pulse TMS, with the activity of the left primary motor cortex (M1) resulted in significant delays of right-hand movements but had little effect on the time the subjects experienced the movement, (assessed by asking participants to indicate the position of a rotating clock hand). Conversely, single-pulse TMS of the anterior frontal areas (with the coil placed at the standard FCz site) resulted in smaller delays in actual RTs but larger delays in the assessment of the timing of manual response, an ability related to motor awareness. This shows that motor awareness does not co-vary with the experimentally induced delay in motor response. In other words, once the intention to perform an action has been formed, the motor response may be delayed but the motor awareness, already triggered by the intentional stance, is not affected. Motor awareness can be reported even in absence of any willed movement. Indeed, brain damaged patients with anosognosia for hemiplegia (that is patients who deny their paralysis) subjectively report the feeling of having performed an action with the paralysed limb [6,7]. This phenomenal experience has its measurable counterpart in the fact that the pretended action with the paralysed hand actually affects the spatiotemporal parameters of the healthy hand movements [8,9]. Interestingly, on the bases of lesional data in anosognosic patients, Berti and colleagues [6,10] have proposed that the right premotor cortex (PM, especially area 6) is part of a neural circuit for motor monitoring, therefore contributing to the operation of one of the comparator systems described by Blakemore et al.’s [1] (see also Haggard, 2005 [11]). In particular, previous studies suggested the involvement of premotor [6,12,13] and insular cortices [6,12,14] for the process of conscious motor monitoring, basal ganglia, insulo-frontal, temporal and parietal structures for explicit and implicit motor awareness [15], and mesial-frontal [5,16,17] and posterior-parietal areas [18] for the intentional component of the motor act. More recently, non-invasive brain stimulation evidence [19] have shown that cathodal transcranial Direct Current Stimulation (tDCS) of the PM, but not of the PPC, affects subjects' self-confidence about their contralateral (left) hand motor performance, consistent with the idea of a role of right PM in the conscious monitoring of voluntary motor acts. On the contrary, tDCS over PM does not interfere with monitoring of involuntary muscle contractions induced by TMS over the hand motor area [20].
Taken together previous findings suggest a role of right PM in the conscious control of voluntary action, but they do not provide direct evidence of its causal involvement. With the present study, we aimed at exploring the role of the right PM in motor monitoring by means of repetitive Transcranial Magnetic Stimulation (rTMS), which allows to draw causal links between the stimulated brain regions and the observed behaviours [21,22]. In particular, subjects were requested to draw straight line either with the left hand (i.e. the hand contralateral to the stimulated side) or with the right (ipsilateral) hand. During the execution of the requested movements, visual feedback of subjects’ actual motor performance was, in most of the trials, experimentally deviated from the real trajectory to create a mismatch between the movement they performed and the movements they viewed on a computer screen. This mismatch would lead the subjects to correct their trajectory in the opposite direction in order to draw a straight line. Previous studies (e.g. [23,24,25,26]) had shown that, within certain limits of deviation, subjects did not become aware of the modified trajectory they performed. In other words, in the manipulated trial, until certain degrees of deviation, subjects still believed to trace a straight line, as requested by the task. Therefore, in this experimental setting, subjects made (erroneous) movements they were not aware of. In Fourneret and Jeannerod’s interpretation (1998), this finding demonstrated that subjects became aware of the movement they intended to perform (a straight movement) and not of the movement they actually performed (a deviated movement). This is again consistent with the idea that motor awareness is mainly constructed on a predictive code and not exclusively on the actual sensory feedback. In our experiment, in order to interfere with motor awareness we applied, before the execution of the task, 1 Hz rTMS over the right PMd cortex [27]. The task was also performed after sham rTMS stimulation of the same area.
Our first prediction was that if the PM has a pivotal role in motor awareness, then interfering with its activity by means of rTMS, when the subjects perform their ‘deviated’ trajectories, would further affect their action monitoring. However, it is worth noting that while an inhibitory TMS usually worsens subjects’ responses, by decreasing the activity of the targeted areas, [28,29,30,31,32], a few studies have unexpectedly shown that inhibitory rTMS of the right PMd could enhance subjects’ performance [33,34,35]. Consequently, if rTMS has an inhibitory effect on motor monitoring, we should expect an increase in the angle at which subjects become aware of the deviated trajectory (i.e. a decrease in motor awareness), whereas if rTMS has an enhancing effect on motor monitoring, we should expect a decrease in the angle at which subjects become aware of the deviated trajectory (i.e. an increase in motor awareness). As for the side of the body where right rTMS may have an effect, if the right PMd has a control only over the contralateral hand we would expect a modulation of motor monitoring only for the left hand action. However, if the right PM controls awareness of both hand movements then we should expect to find a modulation of motor monitoring for both hands. Finally, we expect to observe modulation of motor awareness in the active but not in the sham rTMS condition.
Crucially, and independently from the outcome of the stimulation, a modulation of rTMS on subjects’ capability of detecting action deviation would be another fundamental step for demonstrating the key role of the PMd in the construction of motor awareness.
2. Materials and Methods
2.1. Participants
Fourteen 14 healthy right-handed healthy volunteers (11 women, range age: 21-57 years, mean age 25.8; SE=2.5) participated in the study. Participants were selected according to the TMS exclusion criteria [36] and they gave their informed consent to participate in the study, previously approved by the Bioethical Committee of the University of Turin (Italy). The study was performed in compliance with the Declaration of Helsinki for human participants. The Edinburgh Handedness Inventory-revised [37] test was administered to ensure that all subjects were actually right-handed.
2.2. Set up
The experimental setup consisted of a 30x40 cm graphic tablet placed inside a wooden box on a desk and connected to the computer. The computer was also connected to a LCD screen placed on top of the wooden box, 30 cm above the graphic tablet. A hole in the wooden box enabled the participant to insert one of the hand inside the box, thus excluding it from the subjects' view (See Figure 1). The subject was seated on a comfortable fixed chair in front of the desk (both the graphic tablet and the screen were aligned to the subjects' trunk midline) and he/she could only see the screen below the chin. The chair was fixed so that the distance between the subject and the desk was kept equal across conditions (i.e. 60 cm). The hand inside the box held a pen stylus, while the other hand simply rested on the participants’ leg.
2.3. Procedures
The task was to trace a straight line from the starting position to the target position. During the task execution, while tracing the vertical line on the tablet, the subjects could only see the line appearing on the computer screen. In the Artificially Deviated (AD) trials, the output of the graphic tablet was processed by the computer using a simple algorithm for adding a constant linear directional bias to the right or left of varying amplitude, so that the trajectory drawn by the subject could appear displaced to the left or right according to an angle defined by the bias (i.e. Left Deviation-LD from -1° to -25° and Right Deviation-RD, from +1° to + 25°). The trajectory on the screen was the only visual feedback of the actual movement available to the subject and, after the target was reached, they were asked to report whether the trajectory they saw on the screen corresponded to the movement they actually performed or not, by saying “Yes” or “No” (see Figure 1). Subjects performed 51 trials in each experimental condition (i.e. No Stimulation, rTMS and Sham). One trial with 0° deviation was also included, indicating a perfect coherence between the visual feedback and the actual movement. Participants performed the task in three different conditions: No Stimulation (NS), after real 1 Hz rTMS (rTMS), delivered to the right PMd (900 pulses, at 90% of resting Motor Threshold, rMT), after sham stimulation (SHAM), with the coil held perpendicularly to the right PMd. For each condition, participants performed the task with the right (RH) and the left (LH) hand. NS, active and sham stimulation measurements were collected in 3 different days with an interval of about one month between sessions, in order to prevent potential learning. The order of the 3 conditions, as well as the order of the hand, was randomized across participants. Within participants, the order of the hand used for the task was maintained across sessions.
2.4. Transcranial Magnetic Stimulation
In the rTMS and Sham conditions, the subject performed the task soon after 15 minutes of real repetitive TMS or sham stimulation, respectively. In the sham stimulation, the coil was positioned over the same area of the real stimulation, but it was held in a position perpendicular to the subjects' scalp. RTMS was performed with a Magstim Rapid2 system with a focal coil (70-mm figure-of-eight). Participants’ resting Motor Threshold (rMT) was defined as the lowest pulse intensity able to elicit a visible twitch in the abductor pollicis brevis muscle of the right hand in at least five of ten consecutive stimulations of the motor hotspot [38]. The average resting Motor Threshold was 53.4 (SD= 5.38) of maximum machine output. Then, fifteen minutes of low-frequency rTMS (900 pulses, 1 Hz at 90% of rMT) was delivered over the right dorsal premotor cortex (PMd), defined as 2 cm anterior and 1 cm medial to the previously defined M1 hotspot [39]. Soon after the end of the stimulation, participants were asked to look at the screen in order to start the experimental task.
2.5. Data Analysis
Statistical analyses were conducted using Statistica 6.0. Only data from trials with manipulated angles were used for the analysis, so that we excluded the trials without deviation (i.e. the 0). In order to establish at which degree each subject became fully aware of the deviation, the angle at which the subjects recognized the existence of a mismatch between what they saw on the screen and what they had actually traced was recorded and analyzed. Therefore, the dependent variable for data analysis was the angle of deviation (toward the left and/or the right side) at which the subject started to consistently answer “No” (e.g., at the 14 degree), for at least two consecutive degrees. On the degree chose using the above criteria, a 3X2X2 ANOVA with three within-subject factors, COND (No Stimulation-NS, rTMS, and Sham), SIDE (Right and Left DEVIATION), and HAND (Left and Right Hand) was used to directly test the differential effects of NS vs. Real and vs. Sham rTMS.
3. Results
Analyses reveal that the factor COND was significant [F (2, 50) = 7.3; p= .001; partial η 2 = 0.228]. Crucially, post-hoc analyses (Duncan’s test) showed that participants became aware of their deviation at significantly smaller angle soon after the rTMS (p=0.0006, mean= 11.09, SE= 0.9) with respect the BS (mean= 14.34, SE= 1.1) and the Sham condition (p= 0.01, mean= 13.3, SE= 1.1), independently from the hand used to perform the task and from the direction of the deviation (see Figure 2). This result shows that rTMS facilitated awareness compared to the other conditions.
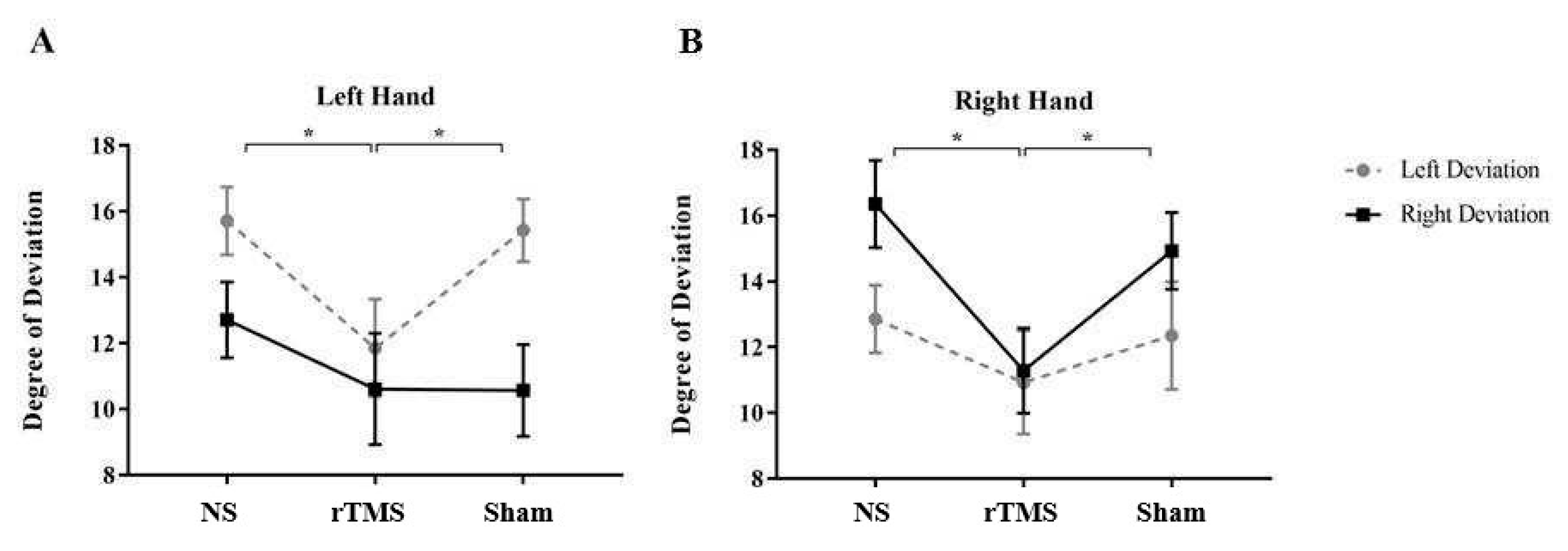
Results also showed a significant three-way interaction between COND X SIDE X HAND [F (2, 50) = 3.79; p= .02 partial η 2 = 0.228]. Post hoc analysis (Duncan’s test) revealed that when the task is performed with the Left Hand, subjects were significantly more aware about their own performance soon after rTMS (p= 0.003, mean= 11.53, SE= 1.6) than BS (mean= 16, SE= 1) and Sham (p= 0.008, mean= 15.46, SE= 1.41) in the Left Deviated trials. Conversely, when subjects performed the task with the Right Hand they were significantly more aware about the deviation in the rTMS condition (mean= 11.28, SE= 1.46) compared to the BS (p=0.0009, mean= 16.35, SE= 1.19) and the Sham (p= 0.01, mean= 14.92, SE= 1.3) in the Right Deviated trials (see Figure 3).
Taken together, these results suggest that rTMS of PMd significantly affected participants conscious self-monitoring, and that the deviation direction, as well as the hand used to perform the task, influenced subjects’ action self-monitoring.
4. Discussion
In the present study, we examined the role of the right PMd in action self-monitoring in healthy volunteers by evaluating motor awareness modulation through the application of inhibitory rTMS. To obtain a behavioural measure of motor awareness, we referred to the well-known Fourneret and Jeannerod paradigm where healthy volunteers are requested to judge their action in a task in which self-generated movements were experimentally deviated [23,24,25].
More specifically, we investigated whether the application of low frequency rTMS over the right PMd may affect the monitoring of trajectory deviations. Consistent with our expectations, we found a modulation of subjects’ motor awareness when movement perturbation was at 11 degrees of deviation. Interestingly, although the results reported in previous studies usually show worsening performance when inhibitory rTMS is applied [32,36] we found that inhibitory rTMS over the right PMd improved the participants’ motor awareness (i.e. it decreased the angle at which they became aware of the deviation). This is in line with some results reported in previous studies showing facilitatory effects of the low frequency rTMS protocol [33,34,35].
One possible explanation for the observed facilitation following 1Hz rTMS protocol is that it may be due to the phenomenon known as paradoxical facilitation (PF), for which behavioral facilitation may result from disruption or inhibition of brain activity [40,41,42]. PF was first described in brain damaged patients who performed better than normal subjects on specific tasks (e.g. [43]), for a review see [44]. Recently, it has been reported that PF can be induced by low frequency rTMS in healthy participants. For example, Buetefisch and co-workers [45] have shown that participants’ accuracy on a task that required ahigher level of precision for both hands increased after low frequency rTMS applied to the left M1. In a previous study, Avanzino et al. [46] demonstrated an improvement of ipsilateral motor accuracy following 1 Hz rTMS over M1 that outlasted the period of stimulation up to 30 min. Similarly, Schwarzkopf et al. [47] have demonstrated that low intensity TMS over the visual cortex facilitated detection of weak motion signals, whereas higher intensities led to impaired detection of stronger motion signals (see also Pascual-Leone et al., (2012) for a review [48]. Therefore, in healthy subjects the effects of low-frequency rTMS on the brain can either worsen [32,49], or improve subjects’ performance [33,34,35]. However, the specific mechanisms of how non-invasive brain stimulation induces PF in healthy individuals are not completely understood yet. One of the recently proposed explanations can be referred to the stochastic resonance model, which postulates that introducing small amounts of noise into a system may promote low-level signals, which, in turn, boost functions within that system. Whatever the explanation, the crucial finding of our study is the modulation of conscious experience obtained by delivering rTMS over the PMd. This demonstrates the causal relationship between premotor cortex and motor awareness, confirming that the premotor cortex can be considered an important hub of the circuit related to the construction of the conscious experience of actions.
As for the body side controlled by the right PM, our results show that the stimulation of the right PMd affects motor awareness for both, contralateral (left) and ipsilateral (right) hands. These findings showing a right hemispheric control on both hands would suggest a right hemispheric specialization for motor monitoring mechanisms. It is worth noting, however, that studies coming from different experimental paradigms have suggested that also the left pre-motor cortex seems to be involved in motor action monitoring. For instance, there are a few cases of anosognosia for the right hemiplegia (that is anosognosia following left, instead of right, brain damage [50] while Fornia et al., [51] in an awake surgery experiment found that Direct Electrical Stimulation (DES) on PMC dramatically altered the patients' motor awareness, making them unconscious of the motor arrest induced by the same stimulation. Given all these results, we might suggest that while the right hemisphere may have a control for motor action executed by both hands, the left hemisphere may have a control only on the right hand. This proposal is reminiscent of one of the theories put forward to explain the deployment of attention in space and the data on neglect [32,52]. Further investigation should consider this possibility.
Finally, we have also found an unexpected result: an effect on the participants’ awareness of the deviation direction related to the hand used to perform the task. Indeed, in the no- stimulation condition, participants were less aware of deviating from a straight trajectory in the Left Deviated trials when the task was performed with the left hand, and in the Right Deviated trials when the task was performed with the right hand. We can speculate on this result by referring to the well-known Simon effect, an attentional effect described as a stimulus-induced bias in response selection [53], in which manual responses to a visual stimulus are facilitated when there is congruence between the stimulus side and the responding hand (stimulus-response compatibility effect [54]. In our experimental setting a condition of hand-space compatibility was realised when the subject had to perform the task with the right (or left) hand and the line that was projected on the screen deviated towards the right (or left) space. The hand-space contingency created by the AD would facilitate and enhance attention to that space by interfering with the generation of awareness of the movements toward the opposite space that the subjects had to perform in order to correct the trajectories (see Freud et al., 2015 for the presence of the Simon effect in motor trajectory task [55]).
5. Limitations of the study
The present study has two main limitations. First, we did not perform stimulation of the left PMd, which prevents any definite conclusion about the role of the right PMd on motor awareness. Considering that, the left pre-motor cortex also seems to be involved in action control [51], it will be crucial to investigate the effect of the stimulation of the left PMd on motor awareness. Second, we did not investigate the role of the right Parietal Cortex in our task, which, according to some authors is involved in motor intention [18]. Therefore, further studies targeting the Parietal Cortex are needed to clarify the role of different brain areas in action monitoring.
6. Conclusions
Our results, showing a modulation on motor awareness by the application of rTMS on right PMd demonstrated that this region plays a crucial role in action self-monitoring. Although the interference with its activity improved the subjects motor awareness, our study suggests that one of the comparator mechanisms proposed by the Blakemore et al. [1] model, responsible forthe conscious monitoring of motor acts, is located in the right PM [6]. Given the functional enhancement effect that we found with the delivery of rTMS to PMd, it is worth considering this procedure as a possible treatment of motor awareness disorders. As already pointed out a limitation of our study is that we did not test the effect of rTMS over the left PMC. This would be crucial to draw firm conclusions about the different involvement of the two hemispheres in action self-monitoring. Therefore, further investigations, also targeting different areas, both in the right and left hemisphere are needed to clarify the different components of the motor monitoring circuit and their specific role in generating the conscious experience of action.
Author Contributions
Conceptualization, A.S., R.R. A.P., F.G., and A.B.; methodology, A.S., A.P., F.G., R.R. and A.B.; software, A.P.; validation, A.S. and A.P.; formal analysis, A.S., P.S. and A.P.; investigation, A.S. and P.S.; data curation, A.S. and P.S.; writing—original draft preparation, A.S., and P.S.; writing—review and editing, A.S., P.S., A.P., F.G., R.R., A.B.; supervision, R.R. and A.B.; project administration, A.S. and A.B.; funding acquisition, A.B. All authors have read and agreed to the published version of the manuscript.
Funding
The study has been funded by a San Paolo Foundation (EU accelerating grant 2012) grants to A.B.
Institutional Review Board Statement
The study was conducted according to the guidelines of the Declaration of Helsinki, and approved by the Ethics Committee of University of Turin (protocol code 24001, 20/07/2015).
Informed Consent Statement
Prior to the experiment, all participants were given a detailed expla-nation of the experimental procedures and provided written informed consent to participate.
Acknowledgments
We would like to thank Ms Federica Morello and Ms Sabrina Montanaro for the support during data collection.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Blakemore, S.-J.; Wolpert, D.M.; Frith, C.D. Abnormalities in the awareness of action. Trends Cogn. Sci. 2002, 6, 237–242. [Google Scholar] [CrossRef]
- The American Association for Research into Nervous and Mental Diseases; Heilman, K.M.; Barrett, A.M.; Adair, J.C. Possible mechanisms of anosognosia: a defect in self–awareness. Philos. Trans. R. Soc. B: Biol. Sci. 1998, 353, 1903–1909. [Google Scholar] [CrossRef]
- Frith, C.D.; Blakemore, S.-J.; Wolpert, D.M. Abnormalities in the awareness and control of action. Philos. Trans. R. Soc. B: Biol. Sci. 2000, 355, 1771–1788. [Google Scholar] [CrossRef] [PubMed]
- Libet, B. Unconscious cerebral initiative and the role of conscious will in voluntary action. Behavioral and Brain Sciences 1985, 8, 529–539. [Google Scholar] [CrossRef]
- Haggard, P.; Magno, E. Localising awareness of action with transcranial magnetic stimulation. Exp. Brain Res. 1999, 127, 102–107. [Google Scholar] [CrossRef]
- Berti, A.; Bottini, G.; Gandola, M.; Pia, L.; Smania, N.; Stracciari, A.; Castiglioni, I.; Vallar, G.; Paulesu, E. Shared Cortical Anatomy for Motor Awareness and Motor Control. Science 2005, 309, 488–491. [Google Scholar] [CrossRef]
- Berti, A.; Pia, L. Understanding Motor Awareness Through Normal and Pathological Behavior. Curr. Dir. Psychol. Sci. 2006, 15, 245–250. [Google Scholar] [CrossRef]
- Garbarini, F.; Rabuffetti, M.; Piedimonte, A.; Pia, L.; Ferrarin, M.; Frassinetti, F.; Gindri, P.; Cantagallo, A.; Driver, J.; Berti, A. ‘Moving’ a paralysed hand: bimanual coupling effect in patients with anosognosia for hemiplegia. Brain 2012, 135, 1486–1497. [Google Scholar] [CrossRef]
- Pia, L.; Spinazzola, L.; Rabuffetti, M.; Ferrarin, M.; Garbarini, F.; Piedimonte, A.; Driver, J.; Berti, A. Temporal coupling due to illusory movements in bimanual actions: Evidence from anosognosia for hemiplegia. Cortex 2013, 49, 1694–1703. [Google Scholar] [CrossRef]
- Pia, L.; Neppi-Modona, M.; Ricci, R.; Berti, A. The anatomy of anosognosia for hemiplegia: a metanalysis. Cortex 2004, 40, 367–377. [Google Scholar] [CrossRef]
- Haggard, P. Conscious intention and motor cognition. Trends Cogn. Sci. 2005, 9, 290–295. [Google Scholar] [CrossRef] [PubMed]
- Besharati, S.; Forkel, S.J.; Kopelman, M.; Solms, M.; Jenkinson, P.M.; Fotopoulou, A. The affective modulation of motor awareness in anosognosia for hemiplegia: Behavioural and lesion evidence. Cortex 2014, 61, 127–140. [Google Scholar] [CrossRef] [PubMed]
- Garbarini, F.; Cecchetti, L.; Bruno, V.; Mastropasqua, A.; Fossataro, C.; Massazza, G.; Sacco, K.; Valentini, M.C.; Ricciardi, E.; Berti, A. To Move or Not to Move? Functional Role of Ventral Premotor Cortex in Motor Monitoring During Limb Immobilization. Cereb. Cortex 2018, 29, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Karnath, O. Awareness of the functioning of one's own limbs mediated by the insular cortex? J Neurosci. 2005, 25, 7134–8 7. [Google Scholar] [CrossRef] [PubMed]
- Moro, V.; Pernigo, S.; Zapparoli, P.; Cordioli, Z.; Aglioti, S.M. Phenomenology and neural correlates of implicit and emergent motor awareness in patients with anosognosia for hemiplegia. Behav. Brain Res. 2011, 225, 259–269. [Google Scholar] [CrossRef]
- Lau, H.C.; Rogers, R.D.; Haggard, P.; Passingham, R.E. Attention to intention. Science 2004, 303, 1208–10. [Google Scholar] [CrossRef]
- Fried, I.; Mukamel, R.; Kreiman, G. Internally Generated Preactivation of Single Neurons in Human Medial Frontal Cortex Predicts Volition. Neuron 2011, 69, 548–562. [Google Scholar] [CrossRef]
- Desmurget, M.; Reilly, K.T.; Richard, N.; Szathmari, A.; Mottolese, C.; Sirigu, A. Movement Intention After Parietal Cortex Stimulation in Humans. Science 2009, 324, 811–813. [Google Scholar] [CrossRef]
- Bolognini, N.; Zigiotto, L.; Carneiro, M.I.S.; Vallar, G. “How Did I Make It?”: Uncertainty about Own Motor Performance after Inhibition of the Premotor Cortex. J. Cogn. Neurosci. 2016, 28, 1052–1061. [Google Scholar] [CrossRef]
- Bruno, V.; Fossataro, C.; Bolognini, N.; Zigiotto, L.; Vallar, G.; Berti, A.; Garbarini, F. The role of premotor and parietal cortex during monitoring of involuntary movement: A combined TMS and tDCS study. Cortex 2017, 96, 83–94. [Google Scholar] [CrossRef]
- Pascual-Leone, A.; Walsh, V.; Rothwell, J. Transcranial magnetic stimulation in cognitive neuroscience – virtual lesion, chronometry, and functional connectivity. Current Opinion in Neurobiology 2000, 10, 232–237. [Google Scholar] [CrossRef] [PubMed]
- Sliwinska, M.W.; Vitello, S.; Devlin, J.T. Transcranial Magnetic Stimulation for Investigating Causal Brain-behavioral Relationships and their Time Course. J. Vis. Exp. 2014. [Google Scholar] [CrossRef]
- Fourneret, P.; Jeannerod, M. Limited conscious monitoring of motor performance in normal subjects. Neuropsychologia 1998, 36, 1133–1140. [Google Scholar] [CrossRef]
- Slachevsky, A.; Pillon, B.; Fourneret, P.; Pradat-Diehl, P.; Jeannerod, M.; Dubois, B. Preserved Adjustment but Impaired Awareness in a Sensory-Motor Conflict following Prefrontal Lesions. J. Cogn. Neurosci. 2001, 13, 332–340. [Google Scholar] [CrossRef] [PubMed]
- Slachevsky, A.; Pillon, B.; Fourneret, P.; Renié, L.; Levy, R.; Jeannerod, M.; Dubois, B. The prefrontal cortex and conscious monitoring of action: An experimental study. Neuropsychologia 2003, 41, 655–665. [Google Scholar] [CrossRef]
- Nielsen, T.I. VOLITION: A NEW EXPERIMENTAL APPROACH. Scand. J. Psychol. 1963, 4, 225–230. [Google Scholar] [CrossRef]
- Christensen, M.S.; Lundbye-Jensen, J.; Grey, M.J.; Vejlby, A.D.; Belhage, B.; Nielsen, J.B. Illusory Sensation of Movement Induced by Repetitive Transcranial Magnetic Stimulation. PLOS ONE 2010, 5, e13301. [Google Scholar] [CrossRef]
- Ando, A.; Salatino, A.; Giromini, L.; Ricci, R.; Pignolo, C.; Cristofanelli, S.; Ferro, L.; Viglione, D.J.; Zennaro, A. Embodied simulation and ambiguous stimuli: The role of the mirror neuron system. Brain Res. 2015, 1629, 135–142. [Google Scholar] [CrossRef]
- Ricci, R.; Salatino, A.; Siebner, H.R.; Mazzeo, G.; Nobili, M. Normalizing Biased Spatial Attention With Parietal rTMS in a Patient With Focal Hand Dystonia. Brain Stimul. 2014, 7, 912–914. [Google Scholar] [CrossRef]
- Salatino, A.; Berra, E.; Troni, W.; Sacco, K.; Cauda, F.; D’agata, F.; Geminiani, G.; Duca, S.; Dimanico, U.; Ricci, R. Behavioral and neuroplastic effects of low-frequency rTMS of the unaffected hemisphere in a chronic stroke patient: A concomitant TMS and fMRI study. Neurocase 2013, 20, 615–626. [Google Scholar] [CrossRef]
- Salatino, A.; Momo, E.; Nobili, M.; Berti, A.; Ricci, R. Awareness of Symptoms Amelioration Following Low-frequency Repetitive Transcranial Magnetic Stimulation in a Patient With Tourette Syndrome and Comorbid Obsessive-compulsive Disorder. Brain Stimul. 2014, 7, 341–343. [Google Scholar] [CrossRef]
- Salatino, A.; Chillemi, G.; Gontero, F.; Poncini, M.; Pyasik, M.; Berti, A.; Ricci, R. Transcranial Magnetic Stimulation of Posterior Parietal Cortex Modulates Line-Length Estimation but Not Illusory Depth Perception. Front. Psychol. 2019, 10, 1169. [Google Scholar] [CrossRef]
- Chambers, C.D.; Bellgrove, M.A.; Gould, I.C.; English, T.; Garavan, H.; McNaught, E.; Kamke, M.; Mattingley, J.B. Dissociable Mechanisms of Cognitive Control in Prefrontal and Premotor Cortex. J. Neurophysiol. 2007, 98, 3638–3647. [Google Scholar] [CrossRef] [PubMed]
- Boroojerdi, B.; Phipps, M.; Kopylev, L.; Wharton, C.; Cohen, L.; Grafman, J. Enhancing analogic reasoning with rTMS over the left prefrontal cortex. Neurology 2001, 56, 526–528. [Google Scholar] [CrossRef]
- Dräger, B.; Breitenstein, C.; Helmke, U.; Kamping, S.; Knecht, S. Specific and nonspecific effects of transcranial magnetic stimulation on picture–word verification. Eur. J. Neurosci. 2004, 20, 1681–1687. [Google Scholar] [CrossRef]
- Rossi, S.; Antal, A.; Bestmann, S.; Bikson, M.; Brewer, C.; Brockmöller, J.; Carpenter, L.L.; Cincotta, M.; Chen, R.; Daskalakis, J.D.; et al. Safety and recommendations for TMS use in healthy subjects and patient populations, with updates on training, ethical and regulatory issues: Expert Guidelines. Clin. Neurophysiol. 2020, 132, 269–306. [Google Scholar] [CrossRef] [PubMed]
- Oldfield, R.C. The assessment and analysis of handedness: The Edinburgh inventory. Neuropsychologia 1971, 9, 97–113. [Google Scholar] [CrossRef] [PubMed]
- Ricci, R.; Salatino, A.; Li, X.; Funk, A.P.; Logan, S.L.; Mu, Q.; Johnson, K.A.; Bohning, D.E.; George, M.S. Imaging the neural mechanisms of TMS neglect-like bias in healthy volunteers with the interleaved TMS/fMRI technique: preliminary evidence. Front. Hum. Neurosci. 2012, 6, 326. [Google Scholar] [CrossRef] [PubMed]
- Kimberley, T.J.; Borich, M.R.; Arora, S.; Siebner, H.R. Multiple sessions of low-frequency repetitive transcranial magnetic stimulation in focal hand dystonia: clinical and physiological effects. Restor. Neurol. Neurosci. 2013, 31, 533–542. [Google Scholar] [CrossRef]
- Miniussi, C.; Rossini, P.M. Transcranial magnetic stimulation in cognitive rehabilitation. Neuropsychol. Rehabilitation 2011, 21, 579–601. [Google Scholar] [CrossRef]
- Levasseur-Moreau, J.; Brunelin, J.; Fecteau, S. Non-invasive brain stimulation can induce paradoxical facilitation. Are these neuroenhancements transferable and meaningful to security services? Front. Hum. Neurosci. 2013, 7, 449. [Google Scholar] [CrossRef]
- Sarasso, P.; Ninghetto, M.; Salatino, A.; Ronga, I.; Bongiardina, A.; Iarrobino, I.; Neppi-Modona, M.; Ricci, R. Everything is (still) illuminated: Dual right cathodal-left anodal tDCS of PPC prevents fatigue on a visual detection task. Brain Stimul. 2018, 12, 187–189. [Google Scholar] [CrossRef]
- Làdavas, E.; Petronio, A.; Umiltà, C. The Deployment of Visual Attention in the Intact Field of Hemineglect Patients. Cortex 1990, 26, 307–317. [Google Scholar] [CrossRef]
- Kapur, N. Paradoxical functional facilitation in brain-behaviour research. Brain 1996, 119, 1775–1790. [Google Scholar] [CrossRef] [PubMed]
- Buetefisch, C.M.; Hines, B.; Shuster, L.; Pergami, P.; Mathes, A.; Wischnewski, M.; Kowalski, G.M.; Rink, F.; Belagaje, S.R.; Haut, M.W.; et al. Motor demand-dependent improvement in accuracy following low-frequency transcranial magnetic stimulation of left motor cortex. J. Neurophysiol. 2011, 106, 1614–1621. [Google Scholar] [CrossRef] [PubMed]
- Avanzino, L.; Bove, M.; Trompetto, C.; Tacchino, A.; Ogliastro, C.; Abbruzzese, G. 1-Hz repetitive TMS over ipsilateral motor cortex influences the performance of sequential finger movements of different complexity. Eur. J. Neurosci. 2008, 27, 1285–1291. [Google Scholar] [CrossRef] [PubMed]
- Schwarzkopf, D.S.; Silvanto, J.; Rees, G. Stochastic Resonance Effects Reveal the Neural Mechanisms of Transcranial Magnetic Stimulation. J. Neurosci. 2011, 31, 3143–3147. [Google Scholar] [CrossRef]
- Pascual-Leone, A. , Horvath, J.C., Robertson, E.M. “Enhancement of normal cognitive abilities through noninvasive brain stimulation” in Cortical Connectivity: Brain Stimulation for Assessing and Modulating Cortical Connectivity and Function, eds R. Chen, and J.C. Rothwell (Berlin; Heidelberg: Springer), 2012. [Google Scholar]
- Ianì, F.; Burin, D.; Salatino, A.; Pia, L.; Ricci, R.; Bucciarelli, M. The beneficial effect of a speaker’s gestures on the listener’s memory for action phrases: The pivotal role of the listener’s premotor cortex. Brain Lang. 2018, 180-182, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Baier, B.; Vucurevic, G.; Müller-Forell, W.; Glassl, O.; Geber, C.; Dieterich, M.; Karnath, H.-O. Anosognosia for hemiparesis after left-sided stroke. Cortex 2014, 61, 120–126. [Google Scholar] [CrossRef]
- Fornia, L.; Puglisi, G.; Leonetti, A.; Bello, L.; Berti, A.; Cerri, G.; Garbarini, F. Direct electrical stimulation of the premotor cortex shuts down awareness of voluntary actions. Nat. Commun. 2020, 11, 1–11. [Google Scholar] [CrossRef]
- Heilman, K.M. , Valenstein, E., Watson, R.T. Neglect and related disorders. Semin Neurol. 2000, 20, 463–470. [Google Scholar] [CrossRef] [PubMed]
- Simon, J. R. The effects of an irrelevant directional cue on human information processing. In R. W. Proctor & T. G. Reeve (Eds.), Stimulus-response compatibility (pp. 31-86). 1990, Amsterdam: North-Holland.
- Umiltá, C. , Nicoletti, R. Spatial Stimulus-Response Compatibility. Advances in Psychology 1990, 65, 89–116. [Google Scholar]
- Freud, E.; Aisenberg, D.; Salzer, Y.; Henik, A.; Ganel, T. Simon in action: the effect of spatial congruency on grasping trajectories. Psychol. Res. 2013, 79, 134–142. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Experimental set up and procedure. The experimental setup consisted of a 30x40 cm graphic tablet placed inside a wooden box on a desk and connected to the computer. An LCD screen was placed on top of the wooden box. A hole in the wooden box enabled the participants to insert one of the hands inside the box, thus excluding it from their view. The subjects were seated on a comfortable fixed chair in front of the desk (both the graphic tablet and the screen were aligned to the subjects' trunk midline) and they could only see the screen below the chin (Panel A). The subjects were instructed to reach, with the pen tip, a yellow target (4x4 mm) located on the sagittal axis at 22 cm from the starting point, by drawing a continuous line as fast as possible. After the target was reached, they were asked to report whether the trajectory they saw on the screen corresponded to the movement they actually performed or not, by saying “Yes” or “No”. Subjects performed 51 trials in each experimental condition (i.e. No Stimulation, rTMS and Sham). For each trial, the software applied, in a random order, the trajectory deviation, which could range from 25° to the left (LD, i.e. -25° from the 0, with negative values indicating a leftward perturbation) to 25° on the right (RD, i.e. + 25° from the 0), with a trial for each degree of deviation. One 0° deviation trial was also included, indicating a perfect coherence between the visual feedback and the actual movement (Panel B).
Figure 1.
Experimental set up and procedure. The experimental setup consisted of a 30x40 cm graphic tablet placed inside a wooden box on a desk and connected to the computer. An LCD screen was placed on top of the wooden box. A hole in the wooden box enabled the participants to insert one of the hands inside the box, thus excluding it from their view. The subjects were seated on a comfortable fixed chair in front of the desk (both the graphic tablet and the screen were aligned to the subjects' trunk midline) and they could only see the screen below the chin (Panel A). The subjects were instructed to reach, with the pen tip, a yellow target (4x4 mm) located on the sagittal axis at 22 cm from the starting point, by drawing a continuous line as fast as possible. After the target was reached, they were asked to report whether the trajectory they saw on the screen corresponded to the movement they actually performed or not, by saying “Yes” or “No”. Subjects performed 51 trials in each experimental condition (i.e. No Stimulation, rTMS and Sham). For each trial, the software applied, in a random order, the trajectory deviation, which could range from 25° to the left (LD, i.e. -25° from the 0, with negative values indicating a leftward perturbation) to 25° on the right (RD, i.e. + 25° from the 0), with a trial for each degree of deviation. One 0° deviation trial was also included, indicating a perfect coherence between the visual feedback and the actual movement (Panel B).
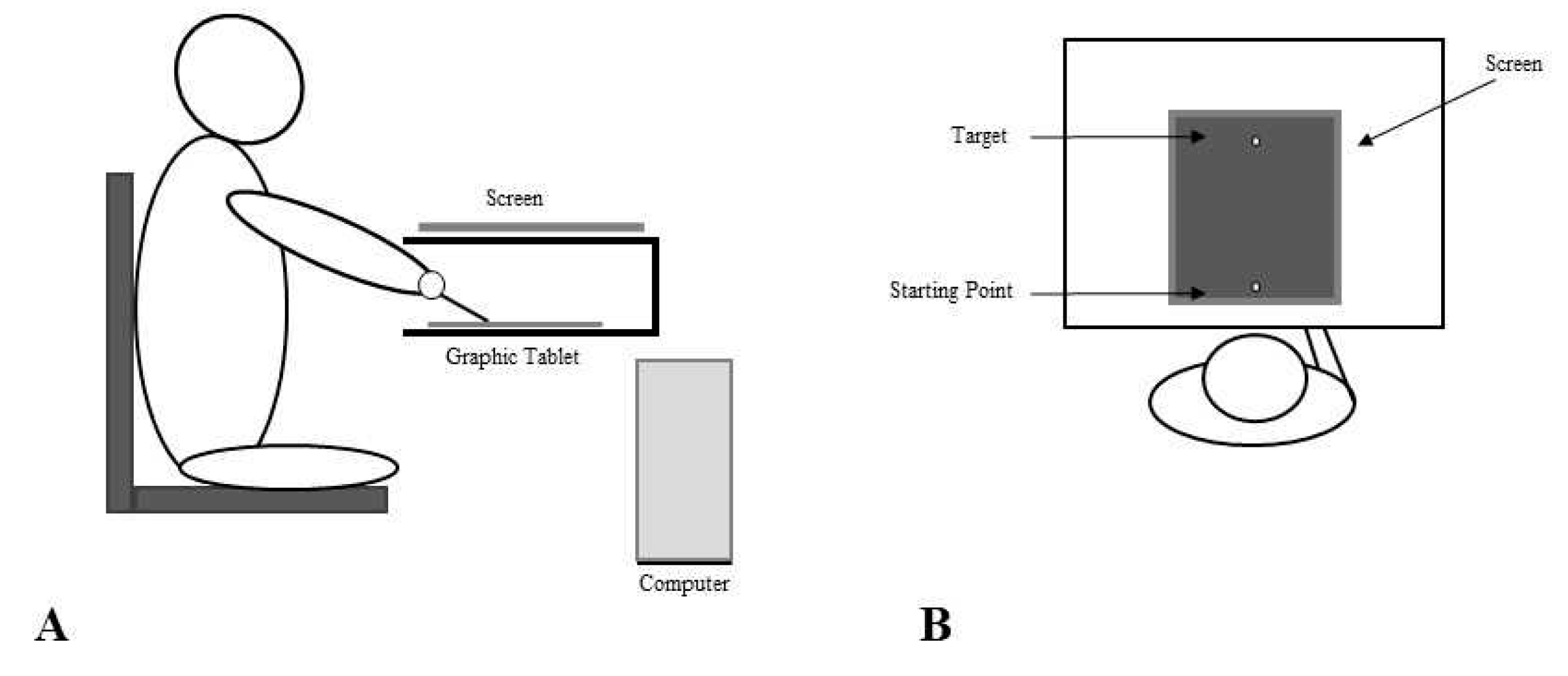
Figure 2.
Mean degree at which subjects became aware of the artificial deviation in the three conditions. Mean degree at which subjects became aware of the artificial deviation in the BS (mean= 14.34, SE= 1.1), soon after the rTMS (mean= 11.09, SE= 0.9) and in the Sham condition (mean= 13.3, SE= 1.1). Error bars represent standard error of means; ∗, significant. NS= No Stimulation, rTMS= repetitive Transcranial Magnetic Stimulation, Sham= Sham stimulation.
Figure 2.
Mean degree at which subjects became aware of the artificial deviation in the three conditions. Mean degree at which subjects became aware of the artificial deviation in the BS (mean= 14.34, SE= 1.1), soon after the rTMS (mean= 11.09, SE= 0.9) and in the Sham condition (mean= 13.3, SE= 1.1). Error bars represent standard error of means; ∗, significant. NS= No Stimulation, rTMS= repetitive Transcranial Magnetic Stimulation, Sham= Sham stimulation.
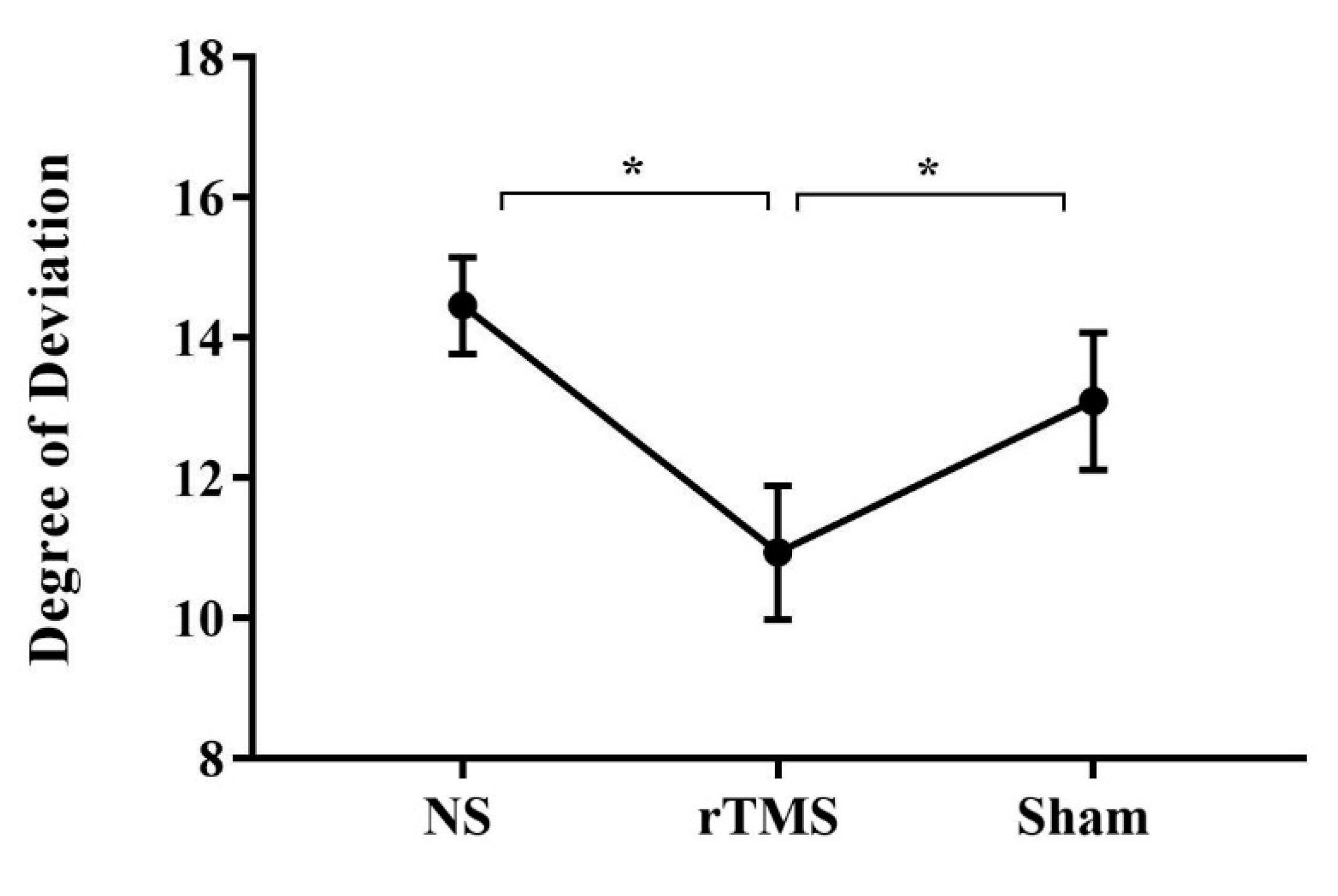
Figure 3.
Mean degree at which subjects became aware of the artificial deviation in the three conditions and with the two deviations. Mean degree at which subjects became aware of the artificial deviation in the three conditions and with the two deviations when the task was performed with the left hand (Panel A) and with the right hand (Panel B). Error bars represent standard error of means; ∗, significant. NS= No Stimulation, rTMS= repetitive Transcranial Magnetic Stimulation, Sham= Sham stimulation. Taken together, these results indicate that rTMS of PMd significantly affected participants conscious self-monitoring, and that the deviation direction, as well as the hand used to perform the task, influenced subjects’ action self-monitoring.
Figure 3.
Mean degree at which subjects became aware of the artificial deviation in the three conditions and with the two deviations. Mean degree at which subjects became aware of the artificial deviation in the three conditions and with the two deviations when the task was performed with the left hand (Panel A) and with the right hand (Panel B). Error bars represent standard error of means; ∗, significant. NS= No Stimulation, rTMS= repetitive Transcranial Magnetic Stimulation, Sham= Sham stimulation. Taken together, these results indicate that rTMS of PMd significantly affected participants conscious self-monitoring, and that the deviation direction, as well as the hand used to perform the task, influenced subjects’ action self-monitoring.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



