Submitted:
13 May 2024
Posted:
14 May 2024
You are already at the latest version
Abstract
Phylogenomics has enriched our understanding that the Tree of Life can have network-like or reticulate structures among some taxa and genes. Non-vertical modes of evolution—such as hybridization/introgression and horizontal gene transfer—deviate from a strictly bifurcating tree model, causing non-treelike patterns. Here, we present a brief overview of a phylogenomic workflow for inferring organismal histories and compare methods for detecting reticulate evolution. We discuss how the timing of coalescent events can help disentangle introgression from incomplete lineage sorting and how horizontal gene transfer events can help determine the relative timing of speciation events. In doing so, we identify pitfalls of certain methods and discuss how to extend their utility across the Tree of Life. Workflows, methods, and future directions discussed herein underscore the need to embrace reticulate evolutionary patterns for understanding the timing and rates of evolutionary events, providing a clearer understanding of life’s history.
Keywords:
phylogenetics
; hybridization
; introgression
; horizontal gene transfer
; lateral gene transfer
; phylogenetic incongruence
; gene-tree-species-tree discordance
; divergence time estimation
Introduction
Phylogenomics—phylogenetic analysis using genome-scale data—has been used to infer the evolutionary history of diverse lineages across the Tree of Life, including animals, fungi, plants, bacteria, archaea, and viruses (Dunn et al. 2008; Misof et al. 2014; Wickett et al. 2014; Worobey et al. 2016; Simion et al. 2017; Parks et al. 2018; Shen et al. 2018; One Thousand Plant Transcriptomes Initiative 2019; Zhu et al. 2019; Li et al. 2021; Coleman et al. 2021; Galindo et al. 2021; Tahon et al. 2021). These studies have resolved numerous phylogenetic controversies, deepening our understanding of life's history (Capella-Gutiérrez et al. 2012; King and Rokas 2017; Williams et al. 2019; Pipes et al. 2021; Steenwyk et al. 2023a). Phylogenomics has also proven useful for delineating lineage relationships at taxonomic scales ranging from species to higher-order relationships (Muñoz-Gómez et al. 2017; Díaz-Tapia et al. 2017; Mateo-Estrada et al. 2019; Bringloe et al. 2021; Steenwyk et al. 2022b; Sierra-Patev et al. 2023). Species trees inferred using phylogenomics provide the framework for various comparative evolutionary genomic studies, such as determining gene duplication and loss events or studying phenotypic innovation (Zhang et al. 2014b; Steenwyk et al. 2019a; Fernández and Gabaldón 2020; Shen et al. 2020; Phillips et al. 2021; Li et al. 2022b; Opulente et al. 2023; Title et al. 2024).
Incongruence between the evolutionary histories of single loci and organisms (locus-tree-species-tree incongruence or discordance) can arise from various biological factors (Steenwyk et al. 2023a). These factors include reticulate evolutionary processes like hybridization/introgression—the interbreeding between distinct lineages—which can disrupt inferences of both the timing and pattern of historical divergences (Rieseberg et al. 2007; Racimo et al. 2015; Barley et al. 2018; Gonçalves et al. 2018; Gonçalves and Gonçalves 2019; Steenwyk et al. 2020a; Suvorov et al. 2022; Li et al. 2022a; Tiley et al. 2023). Hybridization/introgression has been documented in plants, algae, fungi, animals, and other lineages (Rieseberg et al. 2007; Neafsey et al. 2010; Stukenbrock 2016; Sousa et al. 2019; Edger et al. 2019; Edelman et al. 2019; Steenwyk et al. 2020a; Mixão and Gabaldón 2020; Bringloe et al. 2021; Wang et al. 2022). Among humans, loci originating from admixture events between early humans and Neanderthals have been associated with adaptation, phenotypic variation, and disease risk, including for severe COVID-19 (Sankararaman et al. 2016; Simonti et al. 2016; Dannemann and Kelso 2017; Dannemann et al. 2017; Zeberg and Pääbo 2020).
Hybridization can also result in allopolyploidy wherein the genome of the hybrid organism encodes (nearly) the entire genome of both parents. Allopolyploidy has been observed in numerous plants, fungi, and a few vertebrates (Ozkan et al. 2001; Session et al. 2016; Edger et al. 2019; Steenwyk et al. 2020a; Chen et al. 2022; Session and Rokhsar 2023). Genome evolution in allopolyploids can be rapid—marked by pronounced loss of genetic material (Ozkan et al. 2001)—or relatively stable, resulting in retention of both parental genomes (Steenwyk et al. 2020a, 2023b; Salojärvi et al. 2024). In either case, introgression/hybridization results in novel combinations of genes and genetic backgrounds that can, in turn, lead to distinct phenotypic profiles (Steenwyk et al. 2020a; Bautista et al. 2021).
Another mode of reticulate evolution, horizontal gene transfer (or lateral gene transfer) —the transfer of genetic material without sexual reproduction—also causes discordance between the locus-tree and the organismal history. Horizontal gene transfer has been documented in diverse organisms, especially among prokaryotes and archaea (Galtier 2007; Yue et al. 2012; Van Etten and Bhattacharya 2020; Arnold et al. 2022; Li et al. 2022a; Gophna and Altman-Price 2022; Gonçalves and Gonçalves 2022; Steenwyk et al. 2023b). Horizontal gene transfer can be advantageous, endowing recipient organisms with potentially novel functionality (Kominek et al. 2019; Gonçalves and Gonçalves 2019; Li et al. 2022a). In certain cases, complex patterns of horizontal gene transfer or lateral acquisition of entire gene clusters can occur, resulting in new metabolic capabilities such as alcohol fermentation and the biosynthesis of thiamine and siderophores in yeast (Gonçalves et al. 2018; Kominek et al. 2019; Gonçalves and Gonçalves 2019). Horizontally acquired genes can also facilitate adaptation to extreme environments. For example, ice-binding proteins originating from bacteria are thought to contribute to algal adaptation to Arctic environments (Dorrell et al. 2023), and mercuric reductase, an enzyme responsible for converting mercury to a less toxic form, was transferred from bacteria to extremophilic algae commonly isolated from environments with a high mercury concentration (Schönknecht et al. 2013). Among protists, approximately 1% of gene repertoires are estimated to have been horizontally acquired (Van Etten and Bhattacharya 2020). These observations emphasize the significance of horizontal gene transfer as a major evolutionary mode across the tree of life.
This review aims to pinpoint current challenges and identify future avenues for methodological advancement in detecting reticulate evolution in phylogenomics. To do so, we briefly outline notable steps for species tree inference—a common prerequisite for detecting reticulate evolution—and then compare methodologies for detecting and differentiating reticulate evolution from other biological factors contributing to incongruence between loci and organismal histories, such as incomplete lineage sorting. We also discuss how determining the relative timing of introgression/hybridization and horizontal gene transfer can inform the order of speciation events. For a more in-depth discussion of analytical sources of phylogenomic incongruence and methods to mitigate them, we refer the reader to previously published literature (e.g., (Philippe et al. 2017; Kapli et al. 2020; Steenwyk et al. 2023a)). The application and development of these methods hold promise for unraveling the confluence of evolutionary processes that shape the Tree of Life.
Overview of a Phylogenomic Workflow
The first step of phylogenomic tree inference involves acquiring high-quality genomic/transcriptomic data from the target taxa (Figure 1A) (Kapli et al. 2020; Cheon et al. 2020; Turnbull et al. 2023). We note that best practices for generating new sequence data involve depositing voucher specimens (preserved whole organisms and/or tissues) in an accredited biorepository for use by future researchers (Buckner et al. 2021). Thereafter, orthology inference is conducted among gene sequences (nucleotide or amino acid) encoded in the genomic/transcriptomic data. Relationships among orthologous genes can be described as one of three categories: one-to-one, one-to-many, and many-to-many (Fernández et al. 2019). Considering two haploid genomes, one-to-one orthologs are encoded in each genome once; one-to-many orthologs are encoded in one genome once and the other multiple times (implying gene duplication or loss); and many-to-many orthologs refer to a gene with multiple copies in each genome. Species tree inference often relies on one-to-one orthologs as phylogenomic markers because they (presumably) have not experienced duplication or loss (Li et al. 2017). (However, phylogenomic markers can also be noncoding sequences; thus, we broaden our terminology to be loci, instead of genes.) Moreover, these single-copy orthologs are often the substrate of many downstream molecular evolution analyses, such as selection measures, relative evolutionary rates, and gene-gene coevolution (Chikina et al. 2016; Kowalczyk et al. 2019; Steenwyk et al. 2021, 2022a; Álvarez-Carretero et al. 2023).
Once a curated set of phylogenomic markers has been obtained, the next step is multiple sequence alignment and trimming of each marker individually (Figure 1C). Multiple sequence alignment aims to determine the site-wise homology across a group of sequences, typically derived from different organisms (Katoh and Standley 2013; Sievers and Higgins 2018; Edgar 2022). Thereafter, alignments for each marker are commonly subjected to trimming, which involves the removal of specific sites or blocks of sites within the alignments (Talavera and Castresana 2007; Criscuolo and Gribaldo 2010; Tan et al. 2015; Steenwyk et al. 2020b). Next, an optimal model of sequence evolution is determined for each alignment for use in conducting phylogenetic inference (Kapli et al. 2020). The resulting single-locus phylogenies represent the inferred genealogical history for that locus among the sampled taxa.
Species tree inference often follows, which seeks to unite information from the genealogical histories among the sampled markers. Two commonly used approaches for species tree inference from genome-scale datasets are multiple sequence alignment concatenation (or simply concatenation) and coalescence (Figure 1D) (Rokas et al. 2003; Liu et al. 2009a; Steenwyk et al. 2023a). Each approach employs a different theoretical framework. Concatenation places the multiple sequence alignments from each marker end-to-end (concatenates them column-wise) to form a supermatrix, which then may be analyzed using a single model of sequence evolution or else partitioned with separate models for different markers or sites (e.g., third codon positions may evolve faster than first or second positions; (Kainer and Lanfear 2015)). Concatenation approaches assume all locus trees reflect the same species tree (Gatesy et al. 2017). In contrast, coalescence relies on the multi-species coalescent model, which accounts for discordance between locus trees stemming from processes like incomplete lineage sorting. There are two main coalescent-based approaches. In the one-step coalescent approach, single-locus phylogenies are estimated simultaneously with the species tree (Liu et al. 2008; Yang and Rannala 2010; Douglas et al. 2022). In two-step coalescent approaches, single-locus phylogenies are individually inferred and then used to construct a summary species-tree phylogeny (Liu et al. 2009b; Zhang et al. 2018).
Support for the resulting phylogeny can be assessed using, for example, bootstrapping, single-locus or -site support frequencies (also known as concordance factors), and phylogenomic subsampling (Figure 1E) (Edwards 2016; Zhang et al. 2018; Minh et al. 2020; Steenwyk et al. 2021, 2023a). Additional parameters to consider during phylogenomic inference, including ways to identify and ameliorate analytical sources of error, are reviewed elsewhere (Philippe et al. 2017; Kapli et al. 2020 p. 20; Steenwyk et al. 2023a). Furthermore, although we focused on multiple sequence alignment-based phylogenomics, we acknowledge the relevance of relatively new alternative data types in the phylogenomic era, such as synteny, retrotransposon insertion, and structure (Doronina et al. 2019; Parey et al. 2023; Schultz et al. 2023; Steenwyk and King 2023; Moi et al. 2023).
Reticulate Evolution: Identification and Relevance for Relative Divergences
Reticulate evolutionary processes, such as hybridization/introgression and horizontal gene transfer, result in loci that record different evolutionary histories than the whole organism (Dobzhansky 1982; Abbott et al. 2013; Steenwyk et al. 2023a). There is a spectrum of outcomes for hybridization ranging from adaptive changes due to ecological selection or compromised viability or fertility due to hybrid incompatibilities (Racimo et al. 2015; Moran et al. 2021). For example, sunflowers have adapted to novel environments and reabsorbed incipient species due to hybridization (Mallet 2005, 2008; Racimo et al. 2015; Buck et al. 2023). Hybrid progeny can have improved growth and reproductive success or be sterile (Zanewich et al. 2018; Qiao et al. 2019; Allen et al. 2020; Adavoudi and Pilot 2021). Similarly, horizontal gene transfer endows recipient organisms with novel genetic material and can be adaptive (Schönknecht et al. 2013; Gonçalves and Gonçalves 2019; Arnold et al. 2022; Li et al. 2022a; Gophna and Altman-Price 2022; Dorrell et al. 2023). For example, hybridization has been observed in microbial pathogens and thus may contribute to higher or lower organismal virulence (Lin et al. 2009; Depotter et al. 2016; Mixão and Gabaldón 2020).
Signatures of Hybridization/Introgression across the Genome, Gene Trees, and Sites
Comparative genomic and phylogenetic methods are available for identifying hybridization/introgression events (Scannell et al. 2006; Marcet-Houben and Gabaldón 2015; Ortiz-Merino et al. 2017; Steenwyk et al. 2020a, 2023b; Mixão and Gabaldón 2020). In the context of allopolyploid hybrids—where the genome of the hybrid organism contains (nearly) the complete genetic complement of both parental genomes and, therefore, two or more copies of most genes—ancient events can be identified by a burst of gene duplications and are supported by other lines of evidence such as synteny information (Chain et al. 2011; Marcet-Houben and Gabaldón 2015; Session et al. 2016). For example, the allopolyploid event leading to the radiation of Hawaiian mints was identified by signatures of ancient hybridization coupled with subgenome duplication (Tomlin et al. 2024).
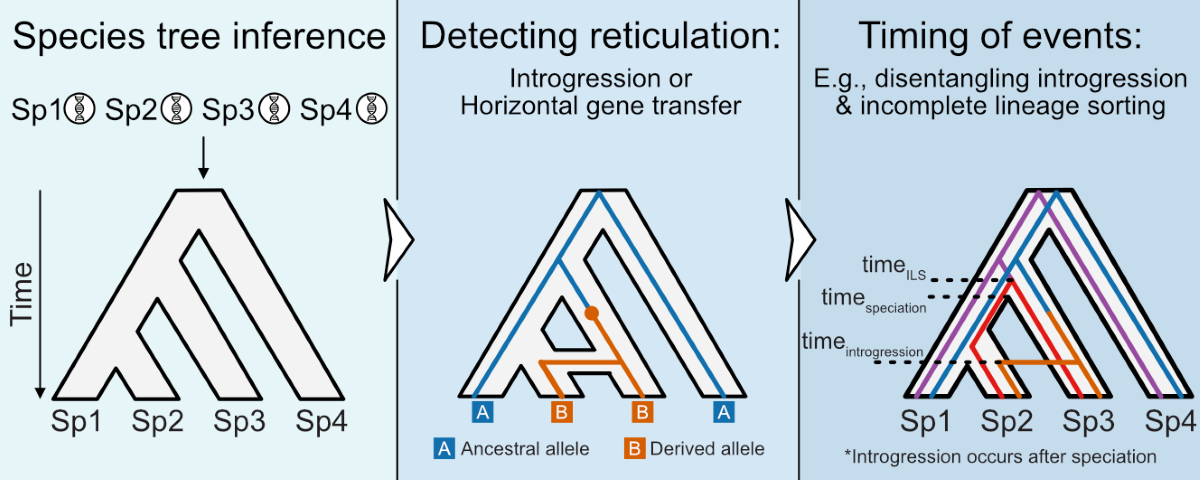
Among phylogenetic approaches, it is crucial to discriminate between incongruences among single-locus phylogenies stemming from hybridization between species versus incomplete lineage sorting—the random sorting of ancestral alleles that can, at times, result in single-loci with evolutionary histories distinct from the organismal history (Yu et al. 2013). Hybridization is favored when two nearly equally supported topologies (one of which is the species tree) are found among genome-wide single-locus phylogenies, which should especially be the case if hybridization was a recent event; incomplete lineage sorting is favored when three topologies are observed equally frequently for a given node, especially among cases of more recent divergences (Steenwyk et al. 2019b). The expected degree of incongruence stemming from incomplete lineage sorting can be modeled using the multispecies coalescent model. Deviations from that model, such as more incongruence than expected, may also be evidence of a past hybridization event (Degnan and Rosenberg 2009).
Hybridization events can also be detected using patterns of site patterns within a phylogenetic framework (Hibbins and Hahn 2022). For example, the D-statistic (or the ABBA-BABA test) is one pioneering approach in this area that leverages expectations about biallelic site patterns along a phylogeny (Figure 2A-E) (Green et al. 2010). Specifically, if the ABBA-BABA test detects asymmetric support between ABBA and BABA patterns at biallelic sites, then an introgression/hybridization event is suggested; in contrast, equal proportions of ABBA and BABA site patterns suggest the absence of introgression/hybridization and instead favor incomplete lineage sorting as the primary source of incongruence. Leveraging genome-scale data, the ABBA-BABA test can accurately quantify introgression across a wide parameter space (Zheng and Janke 2018). Variants of this test that leverage five taxa instead of four can further polarize the directionality of past introgression but are limited to symmetrical tree topologies (Pease and Hahn 2015; Eaton et al. 2015). Of note, detecting ancient hybridization/introgression events using these methods is challenged by analytical factors—such as the inherent difficulty of detecting site-wise orthology and saturation by multiple substitutions. Evaluating the limits of these methods to ancient events remains underexplored and is an avenue for future research.
Coalescent Times Differ between Incomplete Lineage Sorting and Introgression
The degree of incomplete lineage sorting can also differ depending on the timing between speciation events. When speciation occurs at a constant tempo, with sufficient time to accumulate mutations between cladogenic events, incongruence stemming from incomplete lineage sorting is expected to be low (Rokas and Carroll 2006). In contrast, when speciation events occur rapidly, such as during radiation events, the proportion of gene trees supporting all three possible topologies of a rooted triplet is expected to be roughly equal (Song et al. 2023). As a result, differentiating between the three topologies is challenging even with genome-scale data, prompting some to represent such divergences as a polytomy (Sayyari and Mirarab 2018). Several polytomies indicative of near-simultaneous radiation events have been identified in fungi and plants (One Thousand Plant Transcriptomes Initiative 2019; Li et al. 2021; Steenwyk et al. 2021), harkening back to what was earlier called a ‘star phylogeny’ with more limited data (Lara et al. 1996).
Analyses of coalescent times among single loci can help differentiate loci originating from introgression events compared to incomplete lineage sorting. In the case of incomplete lineage sorting, loci will coalesce before speciation, while in the case of hybridization, loci will coalesce after speciation (Song et al. 2023) (Figure 2F-I). This analysis relies on divergence-time analyses of single loci; however, statistical uncertainty can challenge these analyses due to a lack of information in an alignment, and differences in their underlying mutation rates (Koch and Carmona 2024). It is therefore strongly recommended to evaluate loci according to the rate of evolution and relative phylogenetic usefulness (Mongiardino Koch 2021). The influence of different clock model assumptions and time calibrations should also be systematically evaluated to parameterize the ‘chronospace’ of a given analysis (Smith et al. 2018; Mongiardino Koch 2021; Koch and Carmona 2024).
Horizontal Gene Transfer: High Throughput Screens and the Phylogenetic Gold Standard
The methods employed for detecting horizontally acquired loci vary in precision and accuracy. Early techniques relied on identifying deviations in gene sequence characteristics. In the case of very recent prokaryote-to-eukaryote horizontal gene transfer, detection could be achieved by observing genes that deviate in guanine-cytosine content, intron content, gene order, and codon usage across the host genome (Friedman and Ely 2012; Zhang et al. 2014a; Jaramillo et al. 2015; Gonçalves and Gonçalves 2022). In the phylogenomic era, these methods are often employed to support identifying horizontal gene transfer events rather than serving as primary detection tools.
Another approach is to conduct a high throughput screen by calculating the alien index—a score that compares the similarity between sequences within the target group and sequences from outgroup taxa (Gladyshev et al. 2008; Alexander et al. 2016)—of all genes in a host genome. Loci exhibiting alien indices indicative of potential horizontal gene transfer are then selected for further investigation through phylogenetic inference, the gold standard approach for horizontal gene transfer detection. Several software tools have been developed to calculate alien indices or similar metrics for assessing horizontal gene transfer. Examples include AvP, HGTector, and HGTphyloDetect (Zhu et al. 2014; Koutsovoulos et al. 2022; Yuan et al. 2023).
Phylogenetic trees that suggest horizontal gene transfer events are characterized by the confident placement of one or a few sequences within an unexpected taxonomic group (Figure 3A and B). For instance, in the case of prokaryote-to-eukaryote horizontal gene transfer, sequences in a eukaryotic genome may be nested deep within a prokaryotic lineage (Coelho et al. 2013; Husnik and McCutcheon 2018; Zhou et al. 2018; Gonçalves et al. 2018; Shen et al. 2018; Kominek et al. 2019; Gonçalves and Gonçalves 2019; Van Etten and Bhattacharya 2020; Irwin et al. 2021; Li et al. 2022a). The evidence for horizontal gene transfer can be strengthened using topology tests like the Kishino-Hasegawa and Shimodaira-Hasegawa tests (Kishino and Hasegawa 1989; Shimodaira and Hasegawa 1999). These tests compare the likelihood of a phylogeny constrained to reflect a vertical evolutionary scenario (the null hypothesis) with the observed topology, reflecting the occurrence of horizontal gene transfer (the alternative hypothesis) (Gonçalves et al. 2018; Shen et al. 2018).
Horizontal Gene Transfer Events Can Inform Relative Divergences
The timing of divergences plays an important part in understanding the evolutionary history on earth. For example, time-calibrated phylogeneies have been used to identify how plant and fungal symbioses led to radiations in both lineages (Lutzoni et al. 2018). One relatively underexplored method to determine relative divergences is to compare patterns of horizontal gene transfer events (Davín et al. 2018, 2022). Specifically, two competing hypotheses may suggest the origin of one clade preceded or came after the origin of another clade (Figure 3C concerning clades A and B compared to E and F, which are simplified as clade AB and clade EF, respectively). The robust identification of horizontal gene transfer events between lineages of interest may help support one hypothesis. For example, if transfer events are identified between the ancestor of clade AB into the ancestor of EF, this would support EF diversifying before clade AB (Figure 3D). In contrast, if a transfer event is identified between taxon A into the ancestor of clade EF, this would suggest that clade AB diversified before clade EF (Figure 3E). In other words, the expectation that donor clades are older than recipient clades means that careful determining of horizontal gene transfer events across a phylogeny can help determine relative divergence times.
Complex evolutionary histories among horizontally transferred loci may complicate the inference of relative divergences. For example, determining the precise origin of a horizontal gene transfer event in the recipient lineage can be challenging due to differential gene loss or pseudogenization. For example, horizontally acquired loci may be retained in some descendants in a recipient ancestor and lost in others. As a result, it will be difficult to determine at which branch in the organismal phylogeny a gene was acquired, introducing errors in relative divergence time estimates. Specifically, since donor lineages are older than recipient lineages, differential gene loss in a donor lineage may erroneously support a later divergence in the recipient clade. A similar issue can occur due to pseudogenization. Using the previous example, horizontal transfer into the ancestor of AB followed by the subsequent loss of the horizontally transferred gene in taxon B would erroneously support hypothesis two (Figure 3C-E).
This method also relies on detecting enough horizontal gene transfer events. Certain lineages are known for having higher rates of horizontal gene transfer events—like cyanobacteria, archaea, and fungi. Among these lineages, transfers have helped determine the relative divergences (Davín et al. 2018). However, other lineages, such as animals, tend to experience fewer horizontal gene transfer events (only from some viruses and transposable elements (Osmanski et al. 2023)), raising the question of how many horizontal gene transfers are needed to overcome detection limits from differential gene loss and other factors. This question raises the need for further methods development.
Nonetheless, relative divergence-time estimation using horizontal gene transfers may be particularly helpful for lineages lacking fossils, such as many soft-bodied fungi and microbes. Numerous examples exist of horizontal gene transfer of microbial loci to lineages with well-established fossils, like animals and plants (Yue et al. 2012; Li et al. 2022a). In this case, horizontal gene transfers between microbial lineages and those with well-established fossils can help constrain the evolution of microbial lineages in geologic time. Such analyses may help refine the timing of radiations between symbiotic lineages such as plants and fungi, helping to establish key evolutionary episodes, including how fungi may have helped plants colonize land (Lutzoni et al. 2018).
Time-Calibration of Inferred Phylogenetic Divergences
Divergence times among branches in a phylogenomic analysis can be estimated using fossils, mutation rates, horizontal gene transfers, or other temporal evidence to calibrate a molecular clock model (Ho and Phillips 2009; Dos Reis et al. 2016, 2018; Davín et al. 2018; Tiley et al. 2020). This procedure converts the relative divergences of molecular substitution rates to absolute time, often in units of thousands or millions of years ago. The resulting time-calibrated phylogenies, which may be referred to as ‘timetrees’ or ‘chronograms’, differ from uncalibrated phylogenies (‘phylograms’) in that the former is comparable to other evidence that is scaled to absolute time. Timetrees can be used to investigate causal eco-evolutionary dynamics relative to a broad array of independent evidence; for example, past changes in global temperature versus rates of lineage divergence (Oliveros et al. 2019; Schubert et al. 2019; S. Meseguer and Condamine 2020; Feijó et al. 2022), co-diversification among taxa (Sabrina Pankey et al. 2022; Nelsen et al. 2023), and rates of speciation among related clades (Harvey et al. 2020; Upham et al. 2021).
Approaches to estimating divergence times can be divided into node dating, tip dating, and fossil-free dating. Node dating places temporal constraints (i.e., calibrations) on a bifurcating internal node of a phylogeny. In contrast, tip dating places calibrations on terminal taxa that existed at some time in the past (Ho and Phillips 2009; Heath et al. 2014). The ages of serially sampled taxa—usually fossils or viruses and other microbes (Stadler and Yang 2013)—are the most reliable data for calibrating divergence times in phylogenomic datasets. Fossils and their associated ages can calibrate divergence times at either nodes or tips, typically using a probability distribution to incorporate age uncertainty (Ho and Phillips 2009; Stadler and Yang 2013). A fossil's phylogenetic position relative to living members of a given clade must be inferred or assumed based on other data for that fossil to serve as a time calibration (Parham et al. 2012). Viruses and other microbes evolve rapidly enough that samples collected in the last few decades offer valuable tip calibrations analogous to the role of fossils in longer-lived mammals or plants (Volz et al. 2013; Andréoletti et al. 2022). The resulting ‘phylodynamic’ analyses can help expose the population-dynamic processes that generate the phylogenetic patterns inferred from phylogenomic datasets (Stadler et al. 2021; Andréoletti et al. 2022).
In both node and tip dating, clock models are used to extrapolate species divergence times from temporal constraints. Strict clock models assume a fixed mutation rate in all branches, which is often violated when comparing more distant relatives (e.g., the 2%-per-million-years rate long used for bird mitochondrial genes; (Ho 2007)). Indeed, strict clocks may lack biological realism, so this assumption is often relaxed, such as in autocorrelated clock models where closely related branches have similar mutation rates or, in uncorrelated models where each branch is given an independent rate (Drummond et al. 2006; Lepage et al. 2007; Steenwyk and Rokas 2023). Relaxed clocks allow greater flexibility for handling the observed molecular-rate variation among lineages, and thus they are in wide use today for all types of time-calibration strategies. Multi-species coalescent dating approaches additionally leverage information about ancestral population sizes to estimate species divergence times (Dos Reis et al. 2016, 2018; Flouri et al. 2022). Such coalescent dating approaches can be quite accurate when mutation rates are known from pedigrees (Tiley et al. 2020), and appear to be robust to small amounts of introgression in phylogenomic datasets (Huang et al. 2020; Tiley et al. 2023).
What if no fossils or other serial samples are available for a particular taxon? Two main options exist to calibrate divergences: use a fixed, strict clock model to project estimates back from tips, or use secondary calibrations derived from previous analyses. Secondary calibrations typically apply the divergence times estimated at a larger phylogenetic scale (from primary fossil or rate calibrations) for a sister taxon or outgroup, which can be used to calibrate the root node for a clade of interest (Shaul and Graur 2002). However, caution is required to avoid specifying overly precise secondary calibrations, given the strong assumptions involved (Schenk 2016).
Choosing which software to use for divergence-time estimation involves a trade-off between available compute resources and the desired level of biological realism. At one extreme, the most realistic models (e.g., BPP and StarBEAST (Flouri et al. 2018; Douglas et al. 2022)) will perform Bayesian inference to estimate multi-species coalescent parameters across thousands of gene genealogies, considering multiple rate priors, and integrating across both phylogenetic and temporal uncertainty to yield a posterior distribution of time-scaled trees. However, these ‘full methods’ do not scale to large numbers of taxa or distant relatives (Tiley et al. 2020; Jiao et al. 2021).
At the other extreme, concatenated sequence data can be used step-wise to first estimate the phylogenetic tree topology in units of substitutions/site, which can then be calibrated in a second step of divergence-time estimation. Step-wise methods most commonly use maximum-likelihood (e.g., r8s, treePL, RelTime; (Sanderson 2003; Smith and O’Meara 2012; Tao et al. 2020)), but can also be implemented using Bayesian inference in programs like BEAST or MrBayes, which often requires fixing the tree topology. Midway between these extremes is the use of concatenated sequence data to perform simultaneous estimation of topology and divergence times, generally as implemented in a Bayesian framework (e.g., BEAST, MCMCtree, MrBayes, PhyloBayes, RevBayes). This latter approach has been implemented in large datasets (e.g., 800 taxa by 40,000 sites; (Upham et al. 2019)), and continues to be aided by GPU-based computing libraries (Ayres et al. 2019). Strategies for setting priors can strongly impact divergence-time estimation and are thus a further key consideration, particularly since such analyses generally assume the monophyly of all time-constrained nodes (Barba-Montoya et al. 2017).
During divergence time estimation, a range of plausible dates is typically returned under the model’s experimental conditions. Thus, divergence-time results are communicated using confidence intervals, often of the middle 95% (from 2.5% to 97.5% of the resultant distribution). Divergence times can also be inferred using a bootstrapping approach for intractably large datasets (Liu et al. 2023). However, any divergence times communicated without a confidence interval should be viewed with caution given the strong assumptions involved in choosing a point estimate. Overall, the choices of node, tip, or fossil-free dating and strict or relaxed clocks depend on the question of interest, available molecular and morphological data, and prevalence of locus-tree-species-tree incongruence.
Conclusion
This review explores how to infer a species tree and subsequently detect reticulate evolutionary processes within phylogenomic datasets. We expect that future research avenues will seek to improve upon these methods in three main ways.
First, for detecting horizontal gene transfer, improvements in high-throughput tree-based methods will reduce the number of phylogenetic trees that need to be (semi)manually inspected and pruned from collections of putatively horizontally transferred genes. Currently, the alien index is relied upon for high-throughput screening, but it is prone to false positives and thus does not scale well to phylogenomic data.
Second, for detecting introgression, site-based approaches like the ABBA-BABA test will continue to be valuable among recently diverged species or populations, but model-based approaches are needed to test for hybridization at more ancient nodes where substitution saturation is expected (Swofford et al. 2001; Hibbins and Hahn 2022). Building on earlier methods for single loci (Huson et al. 2005), the node-by-node frequency of topologies discordant with the species tree will be skewed to one topology in cases of ancient hybridization whereas incomplete lineage sorting will yield two equally represented discordant topologies. Several studies have generated their own pipelines for analyzing sliding genomic windows to find signatures of ancient hybridization in this way (e.g., in butterflies, fruit flies, and mammals (Edelman et al. 2019; Suvorov et al. 2022; Foley et al. 2023)). However, high-throughput and general-purpose tools for these tests are needed. Developing automated methods for accurate introgression detection from phylogenomic datasets containing hundreds to thousands of taxa will illuminate the general prevalence of introgression across the Tree of Life.
Third, for dating divergence events using single loci, improvements in the accuracy of dates will help differentiate incomplete lineage sorting from introgression, given the expectation that introgressed loci will coalesce more recently than the corresponding species-tree divergence. Doing so will require confident inferences of per-locus substitution rates across genomic windows of different sizes, which is particularly difficult among ancient divergences, again due to substitution saturation. One method to potentially refine molecular clock models is to leverage long-term experimental evolution data, where mutation rates are known to vary (Lenski 2017; Wei et al. 2022). In other taxa, per-species estimates of de novo mutation rates can be obtained by trio-based sequencing of genomes from wild-caught mother-father-offspring (Bergeron et al. 2023; Suárez-Menéndez et al. 2023), which could be leveraged to calibrate divergence times more accurately than with external fossils. Such insights may improve models of the complex interrelationships between mutation rate, population size, natural selection, and the divergence of lineages as manifested in locus-tree-species-tree dynamics.
Taken together, we have identified numerous challenges and opportunities for further research to understand how reticulate evolution can inform—and has shaped—our knowledge of the Tree of Life.
Funding
J.L.S. is a Howard Hughes Medical Institute Awardee of the Life Sciences Research Foundation. N.S.U. was supported by Arizona State University start-up funds. H.V. received support from the Australian Biological Resources Study (4-G046WSD) and the Australian Research Council (DP200101613).
Acknowledgments
We thank Drs. Xing-Xing Shen and Yuanning Li for helpful discussion over the years. N.S.U. thanks Dr. William Murphy for valuable debate about hybridization and divergence times. Chat-GPT was used for initial editing, but the authors extensively edited the text thereafter.
Competing Interests
J.L.S. is an advisor for ForensisGroup Inc.
References
- Abbott R., D. Albach, S. Ansell, J. W. Arntzen, S. J. E. Baird, et al., 2013 Hybridization and speciation. J. Evol. Biol. 26: 229–246. [CrossRef]
- Adavoudi R., and M. Pilot, 2021 Consequences of Hybridization in Mammals: A Systematic Review. Genes 13: 50. [CrossRef]
- Alexander W. G., J. H. Wisecaver, A. Rokas, and C. T. Hittinger, 2016 Horizontally acquired genes in early-diverging pathogenic fungi enable the use of host nucleosides and nucleotides. Proc. Natl. Acad. Sci. U.S.A. 113: 4116–4121. [CrossRef]
- Allen R., H. Ryan, B. W. Davis, C. King, L. Frantz, et al., 2020 A mitochondrial genetic divergence proxy predicts the reproductive compatibility of mammalian hybrids. Proc. R. Soc. B. 287: 20200690. [CrossRef]
- Álvarez-Carretero S., P. Kapli, and Z. Yang, 2023 Beginner’s Guide on the Use of PAML to Detect Positive Selection, (K. Crandall, Ed.). Molecular Biology and Evolution 40: msad041. [CrossRef]
- Andréoletti J., A. Zwaans, R. C. M. Warnock, G. Aguirre-Fernández, J. Barido-Sottani, et al., 2022 The Occurrence Birth–Death Process for Combined-Evidence Analysis in Macroevolution and Epidemiology, (S. Höhna, Ed.). Systematic Biology 71: 1440–1452. [CrossRef]
- Arnold B. J., I.-T. Huang, and W. P. Hanage, 2022 Horizontal gene transfer and adaptive evolution in bacteria. Nat Rev Microbiol 20: 206–218. [CrossRef]
- Ayres D. L., M. P. Cummings, G. Baele, A. E. Darling, P. O. Lewis, et al., 2019 BEAGLE 3: Improved Performance, Scaling, and Usability for a High-Performance Computing Library for Statistical Phylogenetics, (D. Posada, Ed.). Systematic Biology 68: 1052–1061. [CrossRef]
- Barba-Montoya J., M. Dos Reis, and Z. Yang, 2017 Comparison of different strategies for using fossil calibrations to generate the time prior in Bayesian molecular clock dating. Molecular Phylogenetics and Evolution 114: 386–400. [CrossRef]
- Barley A. J., J. M. Brown, and R. C. Thomson, 2018 Impact of Model Violations on the Inference of Species Boundaries Under the Multispecies Coalescent. Systematic Biology 67: 269–284. [CrossRef]
- Bautista C., S. Marsit, and C. R. Landry, 2021 Interspecific hybrids show a reduced adaptive potential under DNA damaging conditions. Evol Appl 14: 758–769. [CrossRef]
- Bergeron L. A., S. Besenbacher, J. Zheng, P. Li, M. F. Bertelsen, et al., 2023 Evolution of the germline mutation rate across vertebrates. Nature 615: 285–291. [CrossRef]
- Bringloe T. T., D. Zaparenkov, S. Starko, W. S. Grant, C. Vieira, et al., 2021 Whole-genome sequencing reveals forgotten lineages and recurrent hybridizations within the kelp genus Alaria (Phaeophyceae), (M. Coleman, Ed.). J. Phycol. 57: 1721–1738. [CrossRef]
- Buck R., D. Ortega-Del Vecchyo, C. Gehring, R. Michelson, D. Flores-Rentería, et al., 2023 Sequential hybridization may have facilitated ecological transitions in the Southwestern pinyon pine syngameon. New Phytologist 237: 2435–2449. [CrossRef]
- Buckner J. C., R. C. Sanders, B. C. Faircloth, and P. Chakrabarty, 2021 The critical importance of vouchers in genomics. eLife 10: e68264. [CrossRef]
- Capella-Gutiérrez S., M. Marcet-Houben, and T. Gabaldón, 2012 Phylogenomics supports microsporidia as the earliest diverging clade of sequenced fungi. BMC Biol 10: 47. [CrossRef]
- Chain F. J., J. Dushoff, and B. J. Evans, 2011 The odds of duplicate gene persistence after polyploidization. BMC Genomics 12: 599. [CrossRef]
- Chen L., J. Xu, X. Sun, and P. Xu, 2022 Research advances and future perspectives of genomics and genetic improvement in allotetraploid common carp. Reviews in Aquaculture 14: 957–978. [CrossRef]
- Cheon S., J. Zhang, and C. Park, 2020 Is Phylotranscriptomics as Reliable as Phylogenomics?, (E. Teeling, Ed.). Molecular Biology and Evolution 37: 3672–3683. [CrossRef]
- Chikina M., J. D. Robinson, and N. L. Clark, 2016 Hundreds of Genes Experienced Convergent Shifts in Selective Pressure in Marine Mammals. Mol Biol Evol 33: 2182–2192. [CrossRef]
- Coelho M. A., C. Gonçalves, J. P. Sampaio, and P. Gonçalves, 2013 Extensive Intra-Kingdom Horizontal Gene Transfer Converging on a Fungal Fructose Transporter Gene, (J. Heitman, Ed.). PLoS Genet 9: e1003587. [CrossRef]
- Coleman G. A., A. A. Davín, T. A. Mahendrarajah, L. L. Szánthó, A. Spang, et al., 2021 A rooted phylogeny resolves early bacterial evolution. Science 372: eabe0511. [CrossRef]
- Criscuolo A., and S. Gribaldo, 2010 BMGE (Block Mapping and Gathering with Entropy): a new software for selection of phylogenetic informative regions from multiple sequence alignments. BMC Evol Biol 10: 210. [CrossRef]
- Dannemann M., and J. Kelso, 2017 The Contribution of Neanderthals to Phenotypic Variation in Modern Humans. The American Journal of Human Genetics 101: 578–589. [CrossRef]
- Dannemann M., K. Prüfer, and J. Kelso, 2017 Functional implications of Neandertal introgression in modern humans. Genome Biol 18: 61. [CrossRef]
- Davín A. A., E. Tannier, T. A. Williams, B. Boussau, V. Daubin, et al., 2018 Gene transfers can date the tree of life. Nat Ecol Evol 2: 904–909. [CrossRef]
- Davín A. A., D. Schrempf, T. A. Williams, P. Hugenholtz, and G. J. Szöllősi, 2022 Relative Time Inference Using Lateral Gene Transfers, pp. 75–94 in Environmental Microbial Evolution, Methods in Molecular Biology. edited by Luo H. Springer US, New York, NY.
- Degnan J. H., and N. A. Rosenberg, 2009 Gene tree discordance, phylogenetic inference and the multispecies coalescent. Trends in Ecology & Evolution 24: 332–340. [CrossRef]
- Depotter J. R., M. F. Seidl, T. A. Wood, and B. P. Thomma, 2016 Interspecific hybridization impacts host range and pathogenicity of filamentous microbes. Current Opinion in Microbiology 32: 7–13. [CrossRef]
- Díaz-Tapia P., C. A. Maggs, J. A. West, and H. Verbruggen, 2017 Analysis of chloroplast genomes and a supermatrix inform reclassification of the Rhodomelaceae (Rhodophyta), (K. Müller, Ed.). J. Phycol. 53: 920–937. [CrossRef]
- Dobzhansky T., 1982 Genetics and the Origin of Species. Columbia university press.
- Doronina L., O. Reising, H. Clawson, D. A. Ray, and J. Schmitz, 2019 True Homoplasy of Retrotransposon Insertions in Primates, (E. Susko, Ed.). Systematic Biology 68: 482–493. [CrossRef]
- Dorrell R. G., A. Kuo, Z. Füssy, E. H. Richardson, A. Salamov, et al., 2023 Convergent evolution and horizontal gene transfer in Arctic Ocean microalgae. Life Sci. Alliance 6: e202201833. [CrossRef]
- Dos Reis M., P. C. J. Donoghue, and Z. Yang, 2016 Bayesian molecular clock dating of species divergences in the genomics era. Nat Rev Genet 17: 71–80. [CrossRef]
- Dos Reis M., G. F. Gunnell, J. Barba-Montoya, A. Wilkins, Z. Yang, et al., 2018 Using Phylogenomic Data to Explore the Effects of Relaxed Clocks and Calibration Strategies on Divergence Time Estimation: Primates as a Test Case, (S. Ho, Ed.). Systematic Biology 67: 594–615. [CrossRef]
- Douglas J., C. L. Jiménez-Silva, and R. Bouckaert, 2022 StarBeast3: Adaptive Parallelized Bayesian Inference under the Multispecies Coalescent, (R. Bell, Ed.). Systematic Biology 71: 901–916. [CrossRef]
- Drummond A. J., S. Y. W. Ho, M. J. Phillips, and A. Rambaut, 2006 Relaxed Phylogenetics and Dating with Confidence, (D. Penny, Ed.). PLoS Biol 4: e88. [CrossRef]
- Dunn C. W., A. Hejnol, D. Q. Matus, K. Pang, W. E. Browne, et al., 2008 Broad phylogenomic sampling improves resolution of the animal tree of life. Nature 452: 745–749. [CrossRef]
- Eaton D. A. R., A. L. Hipp, A. González-Rodríguez, and J. Cavender-Bares, 2015 Historical introgression among the American live oaks and the comparative nature of tests for introgression: INTROGRESSION IN THE AMERICAN LIVE OAKS. Evolution 69: 2587–2601. [CrossRef]
- Edelman N. B., P. B. Frandsen, M. Miyagi, B. Clavijo, J. Davey, et al., 2019 Genomic architecture and introgression shape a butterfly radiation. Science 366: 594–599. [CrossRef]
- Edgar R. C., 2022 Muscle5: High-accuracy alignment ensembles enable unbiased assessments of sequence homology and phylogeny. Nat Commun 13: 6968. [CrossRef]
- Edger P. P., T. J. Poorten, R. VanBuren, M. A. Hardigan, M. Colle, et al., 2019 Origin and evolution of the octoploid strawberry genome. Nat Genet 51: 541–547. [CrossRef]
- Edwards S. V., 2016 Phylogenomic subsampling: a brief review. Zool Scr 45: 63–74. [CrossRef]
- Feijó A., D. Ge, Z. Wen, J. Cheng, L. Xia, et al., 2022 Mammalian diversification bursts and biotic turnovers are synchronous with Cenozoic geoclimatic events in Asia. Proc. Natl. Acad. Sci. U.S.A. 119: e2207845119. [CrossRef]
- Fernández R., T. Gabaldón, and C. Dessimoz, 2019 Orthology: definitions, inference, and impact on species phylogeny inference. [CrossRef]
- Fernández R., and T. Gabaldón, 2020 Gene gain and loss across the metazoan tree of life. Nat Ecol Evol 4: 524–533.
- Flouri T., X. Jiao, B. Rannala, and Z. Yang, 2018 Species Tree Inference with BPP Using Genomic Sequences and the Multispecies Coalescent, (A. D. Yoder, Ed.). Molecular Biology and Evolution 35: 2585–2593. [CrossRef]
- Flouri T., J. Huang, X. Jiao, P. Kapli, B. Rannala, et al., 2022 Bayesian Phylogenetic Inference using Relaxed-clocks and the Multispecies Coalescent, (R. Nielsen, Ed.). Molecular Biology and Evolution 39: msac161. [CrossRef]
- Foley N. M., V. C. Mason, A. J. Harris, K. R. Bredemeyer, J. Damas, et al., 2023 A genomic timescale for placental mammal evolution. Science 380: eabl8189. [CrossRef]
- Friedman R., and B. Ely, 2012 Codon Usage Methods for Horizontal Gene Transfer Detection Generate an Abundance of False Positive and False Negative Results. Curr Microbiol 65: 639–642. [CrossRef]
- Galindo L. J., P. López-García, G. Torruella, S. Karpov, and D. Moreira, 2021 Phylogenomics of a new fungal phylum reveals multiple waves of reductive evolution across Holomycota. Nat Commun 12: 4973. [CrossRef]
- Galtier N., 2007 A Model of Horizontal Gene Transfer and the Bacterial Phylogeny Problem, (M. Steel, Ed.). Systematic Biology 56: 633–642. [CrossRef]
- Gatesy J., R. W. Meredith, J. E. Janecka, M. P. Simmons, W. J. Murphy, et al., 2017 Resolution of a concatenation/coalescence kerfuffle: partitioned coalescence support and a robust family-level tree for Mammalia. Cladistics 33: 295–332. [CrossRef]
- Gladyshev E. A., M. Meselson, and I. R. Arkhipova, 2008 Massive Horizontal Gene Transfer in Bdelloid Rotifers. Science 320: 1210–1213. [CrossRef]
- Gonçalves C., J. H. Wisecaver, J. Kominek, M. S. Oom, M. J. Leandro, et al., 2018 Evidence for loss and reacquisition of alcoholic fermentation in a fructophilic yeast lineage. eLife 7: e33034. [CrossRef]
- Gonçalves C., and P. Gonçalves, 2019 Multilayered horizontal operon transfers from bacteria reconstruct a thiamine salvage pathway in yeasts. Proc. Natl. Acad. Sci. U.S.A. 116: 22219–22228. [CrossRef]
- Gonçalves P., and C. Gonçalves, 2022 Horizontal gene transfer in yeasts. Current Opinion in Genetics & Development 76: 101950. [CrossRef]
- Gophna U., and N. Altman-Price, 2022 Horizontal Gene Transfer in Archaea—From Mechanisms to Genome Evolution. Annu. Rev. Microbiol. 76: 481–502. [CrossRef]
- Green R. E., J. Krause, A. W. Briggs, T. Maricic, U. Stenzel, et al., 2010 A draft sequence of the Neandertal genome. science 328: 710–722. [CrossRef]
- Harvey M. G., G. A. Bravo, S. Claramunt, A. M. Cuervo, G. E. Derryberry, et al., 2020 The evolution of a tropical biodiversity hotspot. Science 370: 1343–1348. [CrossRef]
- Heath T. A., J. P. Huelsenbeck, and T. Stadler, 2014 The fossilized birth–death process for coherent calibration of divergence-time estimates. Proc. Natl. Acad. Sci. U.S.A. 111. [CrossRef]
- Hibbins M. S., and M. W. Hahn, 2022 Phylogenomic approaches to detecting and characterizing introgression, (M. Turelli, Ed.). Genetics 220: iyab173. [CrossRef]
- Ho S. Y. M., 2007 Calibrating molecular estimates of substitution rates and divergence times in birds. J Avian Biology 38: 409–414. [CrossRef]
- Ho S. Y. W., and M. J. Phillips, 2009 Accounting for Calibration Uncertainty in Phylogenetic Estimation of Evolutionary Divergence Times. Systematic Biology 58: 367–380. [CrossRef]
- Huang J., T. Flouri, and Z. Yang, 2020 A Simulation Study to Examine the Information Content in Phylogenomic Data Sets under the Multispecies Coalescent Model, (B. Su, Ed.). Molecular Biology and Evolution 37: 3211–3224. [CrossRef]
- Husnik F., and J. P. McCutcheon, 2018 Functional horizontal gene transfer from bacteria to eukaryotes. Nature Reviews Microbiology 16: 67–79.
- Huson D. H., T. Huson D. H., T. Klöpper, P. J. Lockhart, and M. A. Steel, 2005 Reconstruction of Reticulate Networks from Gene Trees, pp. 233–249 in Research in Computational Molecular Biology, Lecture Notes in Computer Science. edited by Miyano S., Mesirov J., Kasif S., Istrail S., Pevzner P. A., et al. Springer Berlin Heidelberg, Berlin, Heidelberg.
- Irwin N. A. T., A. A. Pittis, T. A. Richards, and P. J. Keeling, 2021 Systematic evaluation of horizontal gene transfer between eukaryotes and viruses. Nat Microbiol 7: 327–336. [CrossRef]
- Jaramillo V. D. A., S. A. Sukno, and M. R. Thon, 2015 Identification of horizontally transferred genes in the genus Colletotrichum reveals a steady tempo of bacterial to fungal gene transfer. BMC Genomics 16: 2. [CrossRef]
- Jiao X., T. Flouri, and Z. Yang, 2021 Multispecies coalescent and its applications to infer species phylogenies and cross-species gene flow. National Science Review 8: nwab127.
- Kainer D., and R. Lanfear, 2015 The Effects of Partitioning on Phylogenetic Inference. Molecular Biology and Evolution 32: 1611–1627. [CrossRef]
- Kapli P., Z. Yang, and M. J. Telford, 2020 Phylogenetic tree building in the genomic age. Nat Rev Genet 21: 428–444. [CrossRef]
- Katoh K., and D. M. Standley, 2013 MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Molecular Biology and Evolution 30: 772–780. [CrossRef]
- King N., and A. Rokas, 2017 Embracing Uncertainty in Reconstructing Early Animal Evolution. Current Biology 27: R1081–R1088. [CrossRef]
- Kishino H., and M. Hasegawa, 1989 Evaluation of the maximum likelihood estimate of the evolutionary tree topologies from DNA sequence data, and the branching order in hominoidea. J Mol Evol 29: 170–179. [CrossRef]
- Koch N. M., and P. M. Carmona, 2024 Chronospaces: an R package for the statistical exploration of divergence times reveals extreme dependence on molecular clocks and gene choice.
- Kominek J., D. T. Doering, D. A. Opulente, X.-X. Shen, X. Zhou, et al., 2019 Eukaryotic Acquisition of a Bacterial Operon. Cell 176: 1356-1366.e10. [CrossRef]
- Koutsovoulos G. D., S. Granjeon Noriot, M. Bailly-Bechet, E. G. J. Danchin, and C. Rancurel, 2022 AvP: A software package for automatic phylogenetic detection of candidate horizontal gene transfers, (M. Ziemann, Ed.). PLoS Comput Biol 18: e1010686. [CrossRef]
- Kowalczyk A., W. K. Meyer, R. Partha, W. Mao, N. L. Clark, et al., 2019 RERconverge: an R package for associating evolutionary rates with convergent traits, (A. Valencia, Ed.). Bioinformatics 35: 4815–4817. [CrossRef]
- Lara M. C., J. L. Patton, and M. N. F. Da Silva, 1996 The Simultaneous Diversification of South American Echimyid Rodents (Hystricognathi) Based on Complete Cytochrome b Sequences. Molecular Phylogenetics and Evolution 5: 403–413. [CrossRef]
- Lenski R. E., 2017 Experimental evolution and the dynamics of adaptation and genome evolution in microbial populations. The ISME Journal 11: 2181–2194. [CrossRef]
- Lepage T., D. Bryant, H. Philippe, and N. Lartillot, 2007 A General Comparison of Relaxed Molecular Clock Models. Molecular Biology and Evolution 24: 2669–2680. [CrossRef]
- Li Z., A. R. De La Torre, L. Sterck, F. M. Cánovas, C. Avila, et al., 2017 Single-Copy Genes as Molecular Markers for Phylogenomic Studies in Seed Plants. Genome Biology and Evolution 9: 1130–1147. [CrossRef]
- Li Y., J. L. Steenwyk, Y. Chang, Y. Wang, T. Y. James, et al., 2021 A genome-scale phylogeny of the kingdom Fungi. Current Biology 31: 1653-1665.e5. [CrossRef]
- Li Y., Z. Liu, C. Liu, Z. Shi, L. Pang, et al., 2022a HGT is widespread in insects and contributes to male courtship in lepidopterans. Cell 185: 2975-2987.e10. [CrossRef]
- Li Y., H. Liu, J. L. Steenwyk, A. L. LaBella, M.-C. Harrison, et al., 2022b Contrasting modes of macro and microsynteny evolution in a eukaryotic subphylum. Current Biology S0960982222016700. [CrossRef]
- Lin X., S. Patel, A. P. Litvintseva, A. Floyd, T. G. Mitchell, et al., 2009 Diploids in the Cryptococcus neoformans Serotype A Population Homozygous for the α Mating Type Originate via Unisexual Mating, (A. Andrianopoulos, Ed.). PLoS Pathog 5: e1000283. [CrossRef]
- Liu L., D. K. Pearl, R. T. Brumfield, and S. V. Edwards, 2008 Estimating species trees using multiple-allele DNA sequence data. Evolution 62: 2080–2091.
- Liu L., L. Yu, L. Kubatko, D. K. Pearl, and S. V. Edwards, 2009a Coalescent methods for estimating phylogenetic trees. Molecular Phylogenetics and Evolution 53: 320–328. [CrossRef]
- Liu L., L. Yu, D. K. Pearl, and S. V. Edwards, 2009b Estimating Species Phylogenies Using Coalescence Times among Sequences. Systematic Biology 58: 468–477. [CrossRef]
- Liu H., J. L. Steenwyk, X. Zhou, D. T. Schultz, K. M. Kocot, et al., 2023 A genome-scale Opisthokonta tree of life: toward phylogenomic resolution of ancient divergences. Evolutionary Biology.
- Lutzoni F., M. D. Nowak, M. E. Alfaro, V. Reeb, J. Miadlikowska, et al., 2018 Contemporaneous radiations of fungi and plants linked to symbiosis. Nat Commun 9: 5451. [CrossRef]
- Mallet J., 2005 Hybridization as an invasion of the genome. Trends in Ecology & Evolution 20: 229–237. [CrossRef]
- Mallet J., 2008 Hybridization, ecological races and the nature of species: empirical evidence for the ease of speciation. Phil. Trans. R. Soc. B 363: 2971–2986. [CrossRef]
- Marcet-Houben M., and T. Gabaldón, 2015 Beyond the Whole-Genome Duplication: Phylogenetic Evidence for an Ancient Interspecies Hybridization in the Baker’s Yeast Lineage, (L. D. Hurst, Ed.). PLoS Biol 13: e1002220. [CrossRef]
- Mateo-Estrada V., L. Graña-Miraglia, G. López-Leal, and S. Castillo-Ramírez, 2019 Phylogenomics Reveals Clear Cases of Misclassification and Genus-Wide Phylogenetic Markers for Acinetobacter, (L. Delaye, Ed.). Genome Biology and Evolution 11: 2531–2541. [CrossRef]
- Minh B. Q., H. A. Schmidt, O. Chernomor, D. Schrempf, M. D. Woodhams, et al., 2020 IQ-TREE 2: New Models and Efficient Methods for Phylogenetic Inference in the Genomic Era, (E. Teeling, Ed.). Molecular Biology and Evolution 37: 1530–1534. [CrossRef]
- Misof B., S. Liu, K. Meusemann, R. S. Peters, A. Donath, et al., 2014 Phylogenomics resolves the timing and pattern of insect evolution. Science 346: 763–767. [CrossRef]
- Mixão V., and T. Gabaldón, 2020 Genomic evidence for a hybrid origin of the yeast opportunistic pathogen Candida albicans. BMC Biol 18: 48. [CrossRef]
- Moi D., C. Moi D., C. Bernard, M. Steinegger, Y. Nevers, M. Langleib, et al., 2023 Structural phylogenetics unravels the evolutionary diversification of communication systems in gram-positive bacteria and their viruses. Bioinformatics.
- Mongiardino Koch N., 2021 Phylogenomic Subsampling and the Search for Phylogenetically Reliable Loci, (Y. Satta, Ed.). Molecular Biology and Evolution 38: 4025–4038. [CrossRef]
- Moran B. M., C. Payne, Q. Langdon, D. L. Powell, Y. Brandvain, et al., 2021 The genomic consequences of hybridization. eLife 10: e69016. [CrossRef]
- Muñoz-Gómez S. A., F. G. Mejía-Franco, K. Durnin, M. Colp, C. J. Grisdale, et al., 2017 The New Red Algal Subphylum Proteorhodophytina Comprises the Largest and Most Divergent Plastid Genomes Known. Current Biology 27: 1677-1684.e4. [CrossRef]
- Neafsey D. E., B. M. Barker, T. J. Sharpton, J. E. Stajich, D. J. Park, et al., 2010 Population genomic sequencing of Coccidioides fungi reveals recent hybridization and transposon control. Genome Res. 20: 938–946. [CrossRef]
- Nelsen M. P., C. S. Moreau, C. Kevin Boyce, and R. H. Ree, 2023 Macroecological diversification of ants is linked to angiosperm evolution. Evolution Letters 7: 79–87. [CrossRef]
- Oliveros C. H., D. J. Field, D. T. Ksepka, F. K. Barker, A. Aleixo, et al., 2019 Earth history and the passerine superradiation. Proc. Natl. Acad. Sci. U.S.A. 116: 7916–7925. [CrossRef]
- One Thousand Plant Transcriptomes Initiative, 2019 One thousand plant transcriptomes and the phylogenomics of green plants. Nature 574: 679–685. [CrossRef]
- Opulente D. A., A. L. Opulente D. A., A. L. LaBella, M.-C. Harrison, J. F. Wolters, C. Liu, et al., 2023 Genomic and ecological factors shaping specialism and generalism across an entire subphylum. Evolutionary Biology.
- Ortiz-Merino R. A., N. Kuanyshev, S. Braun-Galleani, K. P. Byrne, D. Porro, et al., 2017 Evolutionary restoration of fertility in an interspecies hybrid yeast, by whole-genome duplication after a failed mating-type switch, (L. Hurst, Ed.). PLoS Biol 15: e2002128. [CrossRef]
- Osmanski A. B., N. S. Paulat, J. Korstian, J. R. Grimshaw, M. Halsey, et al., 2023 Insights into mammalian TE diversity through the curation of 248 genome assemblies. Science 380: eabn1430. [CrossRef]
- Ozkan H., A. A. Levy, and M. Feldman, 2001 Allopolyploidy-Induced Rapid Genome Evolution in the Wheat ( Aegilops – Triticum ) Group. Plant Cell 13: 1735–1747. [CrossRef]
- Parey E., A. Louis, J. Montfort, O. Bouchez, C. Roques, et al., 2023 Genome structures resolve the early diversification of teleost fishes. Science 379: 572–575. [CrossRef]
- Parham J. F., P. C. J. Donoghue, C. J. Bell, T. D. Calway, J. J. Head, et al., 2012 Best Practices for Justifying Fossil Calibrations. Systematic Biology 61: 346–359. [CrossRef]
- Parks D. H., M. Chuvochina, D. W. Waite, C. Rinke, A. Skarshewski, et al., 2018 A standardized bacterial taxonomy based on genome phylogeny substantially revises the tree of life. Nat Biotechnol 36: 996–1004. [CrossRef]
- Pease J. B., and M. W. Hahn, 2015 Detection and Polarization of Introgression in a Five-Taxon Phylogeny. Systematic Biology 64: 651–662. [CrossRef]
- Philippe H., D. M. D. Vienne, V. Ranwez, B. Roure, D. Baurain, et al., 2017 Pitfalls in supermatrix phylogenomics. EJT. [CrossRef]
- Phillips M. A., J. L. Steenwyk, X.-X. Shen, and A. Rokas, 2021 Examination of Gene Loss in the DNA Mismatch Repair Pathway and Its Mutational Consequences in a Fungal Phylum, (K. Wolfe, Ed.). Genome Biology and Evolution 13: evab219. [CrossRef]
- Pipes L., H. Wang, J. P. Huelsenbeck, and R. Nielsen, 2021 Assessing Uncertainty in the Rooting of the SARS-CoV-2 Phylogeny, (H. Malik, Ed.). Molecular Biology and Evolution 38: 1537–1543. [CrossRef]
- Qiao H., W. Liu, Y. Zhang, Y. Zhang, and Q. Q. Li, 2019 Genetic admixture accelerates invasion via provisioning rapid adaptive evolution. Mol Ecol 28: 4012–4027. [CrossRef]
- Racimo F., S. Sankararaman, R. Nielsen, and E. Huerta-Sánchez, 2015 Evidence for archaic adaptive introgression in humans. Nature Reviews Genetics 16: 359–371.
- Rieseberg L. H., S.-C. Kim, R. A. Randell, K. D. Whitney, B. L. Gross, et al., 2007 Hybridization and the colonization of novel habitats by annual sunflowers. Genetica 129: 149–165. [CrossRef]
- Rokas A., B. L. Williams, N. King, and S. B. Carroll, 2003 Genome-scale approaches to resolving incongruence in molecular phylogenies. Nature 425: 798–804. [CrossRef]
- Rokas A., and S. B. Carroll, 2006 Bushes in the Tree of Life. PLoS Biol 4: e352. [CrossRef]
- S. Meseguer A., and F. L. Condamine, 2020 Ancient tropical extinctions at high latitudes contributed to the latitudinal diversity gradient*. Evolution 74: 1966–1987. [CrossRef]
- Sabrina Pankey M., D. C. Plachetzki, K. J. Macartney, M. Gastaldi, M. Slattery, et al., 2022 Cophylogeny and convergence shape holobiont evolution in sponge–microbe symbioses. Nat Ecol Evol 6: 750–762. [CrossRef]
- Salojärvi J., A. Rambani, Z. Yu, R. Guyot, S. Strickler, et al., 2024 The genome and population genomics of allopolyploid Coffea arabica reveal the diversification history of modern coffee cultivars. Nat Genet 56: 721–731. [CrossRef]
- Sanderson M. J., 2003 r8s: inferring absolute rates of molecular evolution and divergence times in the absence of a molecular clock. Bioinformatics 19: 301–302. [CrossRef]
- Sankararaman S., S. Mallick, N. Patterson, and D. Reich, 2016 The combined landscape of Denisovan and Neanderthal ancestry in present-day humans. Current Biology 26: 1241–1247.
- Sayyari E., and S. Mirarab, 2018 Testing for Polytomies in Phylogenetic Species Trees Using Quartet Frequencies. Genes 9: 132. [CrossRef]
- Scannell D. R., K. P. Byrne, J. L. Gordon, S. Wong, and K. H. Wolfe, 2006 Multiple rounds of speciation associated with reciprocal gene loss in polyploid yeasts. Nature 440: 341–345. [CrossRef]
- Schenk J. J., 2016 Consequences of Secondary Calibrations on Divergence Time Estimates, (A. Janke, Ed.). PLoS ONE 11: e0148228. [CrossRef]
- Schönknecht G., W.-H. Chen, C. M. Ternes, G. G. Barbier, R. P. Shrestha, et al., 2013 Gene Transfer from Bacteria and Archaea Facilitated Evolution of an Extremophilic Eukaryote. Science 339: 1207–1210. [CrossRef]
- Schubert M., T. Marcussen, A. S. Meseguer, and S. Fjellheim, 2019 The grass subfamily Pooideae: Cretaceous–Palaeocene origin and climate-driven Cenozoic diversification, (G. Jordan, Ed.). Global Ecol Biogeogr geb.12923. [CrossRef]
- Schultz D. T., S. H. D. Haddock, J. V. Bredeson, R. E. Green, O. Simakov, et al., 2023 Ancient gene linkages support ctenophores as sister to other animals. Nature. [CrossRef]
- Session A. M., Y. Uno, T. Kwon, J. A. Chapman, A. Toyoda, et al., 2016 Genome evolution in the allotetraploid frog Xenopus laevis. Nature 538: 336–343. [CrossRef]
- Session A. M., and D. S. Rokhsar, 2023 Transposon signatures of allopolyploid genome evolution. Nat Commun 14: 3180. [CrossRef]
- Shaul S., and D. Graur, 2002 Playing chicken ( Gallus gallus ): methodological inconsistencies of molecular divergence date estimates due to secondary calibration points. Gene 300: 59–61. [CrossRef]
- Shen X.-X., D. A. Opulente, J. Kominek, X. Zhou, J. L. Steenwyk, et al., 2018 Tempo and Mode of Genome Evolution in the Budding Yeast Subphylum. Cell 175: 1533-1545.e20. [CrossRef]
- Shen X.-X., J. L. Steenwyk, A. L. LaBella, D. A. Opulente, X. Zhou, et al., 2020 Genome-scale phylogeny and contrasting modes of genome evolution in the fungal phylum Ascomycota. Sci. Adv. 6: eabd0079. [CrossRef]
- Shimodaira H., and M. Hasegawa, 1999 Multiple Comparisons of Log-Likelihoods with Applications to Phylogenetic Inference. Molecular Biology and Evolution 16: 1114–1116. [CrossRef]
- Sierra-Patev S., B. Min, M. Naranjo-Ortiz, B. Looney, Z. Konkel, et al., 2023 A global phylogenomic analysis of the shiitake genus Lentinula. Proc. Natl. Acad. Sci. U.S.A. 120: e2214076120. [CrossRef]
- Sievers F., and D. G. Higgins, 2018 Clustal Omega for making accurate alignments of many protein sequences: Clustal Omega for Many Protein Sequences. Protein Science 27: 135–145. [CrossRef]
- Simion P., H. Philippe, D. Baurain, M. Jager, D. J. Richter, et al., 2017 A Large and Consistent Phylogenomic Dataset Supports Sponges as the Sister Group to All Other Animals. Current Biology 27: 958–967. [CrossRef]
- Simonti C. N., B. Vernot, L. Bastarache, E. Bottinger, D. S. Carrell, et al., 2016 The phenotypic legacy of admixture between modern humans and Neandertals. Science 351: 737–741. [CrossRef]
- Smith S. A., and B. C. O’Meara, 2012 treePL: divergence time estimation using penalized likelihood for large phylogenies. Bioinformatics 28: 2689–2690. [CrossRef]
- Smith S. A., J. W. Brown, and J. F. Walker, 2018 So many genes, so little time: A practical approach to divergence-time estimation in the genomic era, (H. Escriva, Ed.). PLoS ONE 13: e0197433. [CrossRef]
- Song H., Y. Wang, H. Shao, Z. Li, P. Hu, et al., 2023 Scaphopoda is the sister taxon to Bivalvia: Evidence of ancient incomplete lineage sorting. Proc. Natl. Acad. Sci. U.S.A. 120: e2302361120. [CrossRef]
- Sousa F., J. Neiva, N. Martins, R. Jacinto, L. Anderson, et al., 2019 Increased evolutionary rates and conserved transcriptional response following allopolyploidization in brown algae: GENOME EVOLUTION IN ALLOPOLYPLOID ALGAE. Evolution 73: 59–72. [CrossRef]
- Stadler T., and Z. Yang, 2013 Dating Phylogenies with Sequentially Sampled Tips. Systematic Biology 62: 674–688. [CrossRef]
- Stadler T., O. G. Pybus, and M. P. H. Stumpf, 2021 Phylodynamics for cell biologists. Science 371: eaah6266. [CrossRef]
- Steenwyk J. L., D. A. Opulente, J. Kominek, X.-X. Shen, X. Zhou, et al., 2019a Extensive loss of cell-cycle and DNA repair genes in an ancient lineage of bipolar budding yeasts, (S. Kamoun, Ed.). PLoS Biol 17: e3000255. [CrossRef]
- Steenwyk J. L., X.-X. Shen, A. L. Lind, G. H. Goldman, and A. Rokas, 2019b A Robust Phylogenomic Time Tree for Biotechnologically and Medically Important Fungi in the Genera Aspergillus and Penicillium, (J. P. Boyle, Ed.). mBio 10: e00925-19. [CrossRef]
- Steenwyk J. L., A. L. Lind, L. N. A. Ries, T. F. dos Reis, L. P. Silva, et al., 2020a Pathogenic Allodiploid Hybrids of Aspergillus Fungi. Current Biology 30: 2495-2507.e7. [CrossRef]
- Steenwyk J. L., T. J. Buida, Y. Li, X.-X. Shen, and A. Rokas, 2020b ClipKIT: A multiple sequence alignment trimming software for accurate phylogenomic inference, (A. Hejnol, Ed.). PLoS Biol 18: e3001007. [CrossRef]
- Steenwyk J. L., T. J. Buida, A. L. Labella, Y. Li, X.-X. Shen, et al., 2021 PhyKIT: a broadly applicable UNIX shell toolkit for processing and analyzing phylogenomic data, (R. Schwartz, Ed.). Bioinformatics 37: 2325–2331. [CrossRef]
- Steenwyk J. L., M. A. Phillips, F. Yang, S. S. Date, T. R. Graham, et al., 2022a An orthologous gene coevolution network provides insight into eukaryotic cellular and genomic structure and function. Sci. Adv. 8: eabn0105. [CrossRef]
- Steenwyk J. L., C. Steenwyk J. L., C. Balamurugan, H. A. Raja, C. Gonçalves, N. Li, et al., 2022b Phylogenomics reveals extensive misidentification of fungal strains from the genus Aspergillus. Evolutionary Biology.
- Steenwyk J. L., and A. Rokas, 2023 The dawn of relaxed phylogenetics. PLoS Biol 21: e3001998. [CrossRef]
- Steenwyk J. L., Y. Li, X. Zhou, X.-X. Shen, and A. Rokas, 2023a Incongruence in the phylogenomics era. Nature Reviews Genetics.
- Steenwyk J. L., S. L. Steenwyk J. L., S. L. Knowles, R. Bastos, C. Balamurugan, D. Rinker, et al., 2023b Evolutionary origin, population diversity, and diagnostics for a cryptic hybrid pathogen. Evolutionary Biology.
- Steenwyk J., and N. King, 2023 From Genes to Genomes: Opportunities and Challenges for Synteny-based Phylogenies. Preprints. [CrossRef]
- Stukenbrock E. H., 2016 The Role of Hybridization in the Evolution and Emergence of New Fungal Plant Pathogens. Phytopathology® 106: 104–112. [CrossRef]
- Suárez-Menéndez M., M. Bérubé, F. Furni, V. E. Rivera-León, M.-P. Heide-Jørgensen, et al., 2023 Wild pedigrees inform mutation rates and historic abundance in baleen whales. Science 381: 990–995. [CrossRef]
- Suvorov A., B. Y. Kim, J. Wang, E. E. Armstrong, D. Peede, et al., 2022 Widespread introgression across a phylogeny of 155 Drosophila genomes. Current Biology 32: 111-123.e5. [CrossRef]
- Swofford D. L., P. J. Waddell, J. P. Huelsenbeck, P. G. Foster, P. O. Lewis, et al., 2001 Bias in Phylogenetic Estimation and Its Relevance to the Choice between Parsimony and Likelihood Methods. Systematic Biology 50: 525–539. [CrossRef]
- Tahon G., P. Geesink, and T. J. G. Ettema, 2021 Expanding Archaeal Diversity and Phylogeny: Past, Present, and Future. Annu. Rev. Microbiol. 75: 359–381. [CrossRef]
- Talavera G., and J. Castresana, 2007 Improvement of Phylogenies after Removing Divergent and Ambiguously Aligned Blocks from Protein Sequence Alignments, (K. Kjer, R. Page, and J. Sullivan, Eds.). Systematic Biology 56: 564–577. [CrossRef]
- Tan G., M. Muffato, C. Ledergerber, J. Herrero, N. Goldman, et al., 2015 Current Methods for Automated Filtering of Multiple Sequence Alignments Frequently Worsen Single-Gene Phylogenetic Inference. Syst Biol 64: 778–791. [CrossRef]
- Tao Q., K. Tao Q., K. Tamura, and S. Kumar, 2020 Efficient Methods for Dating Evolutionary Divergences, pp. 197–219 in The Molecular Evolutionary Clock, edited by Ho S. Y. W. Springer International Publishing, Cham.
- Tiley G. P., J. W. Poelstra, M. Dos Reis, Z. Yang, and A. D. Yoder, 2020 Molecular Clocks without Rocks: New Solutions for Old Problems. Trends in Genetics 36: 845–856. [CrossRef]
- Tiley G. P., T. Flouri, X. Jiao, J. W. Poelstra, B. Xu, et al., 2023 Estimation of species divergence times in presence of cross-species gene flow, (B. Carstens, Ed.). Systematic Biology 72: 820–836. [CrossRef]
- Title P. O., S. Singhal, M. C. Grundler, G. C. Costa, R. A. Pyron, et al., 2024 The macroevolutionary singularity of snakes. Science 383: 918–923. [CrossRef]
- Tomlin C. M., S. Rajaraman, J. T. Sebesta, A.-C. Scheen, M. Bendiksby, et al., 2024 Allopolyploid origin and diversification of the Hawaiian endemic mints. Nat Commun 15: 3109. [CrossRef]
- Turnbull R., J. L. Steenwyk, S. J. Mutch, P. Scholten, V. W. Salazar, et al., 2023 OrthoFlow: phylogenomic analysis and diagnostics with one command. In Review.
- Upham N. S., J. A. Esselstyn, and W. Jetz, 2019 Inferring the mammal tree: Species-level sets of phylogenies for questions in ecology, evolution, and conservation, (A. J. Tanentzap, Ed.). PLoS Biol 17: e3000494. [CrossRef]
- Upham N. S., J. A. Esselstyn, and W. Jetz, 2021 Molecules and fossils tell distinct yet complementary stories of mammal diversification. Current Biology 31: 4195-4206.e3. [CrossRef]
- Van Etten J., and D. Bhattacharya, 2020 Horizontal Gene Transfer in Eukaryotes: Not if, but How Much? Trends in Genetics 36: 915–925. [CrossRef]
- Volz E. M., K. Koelle, and T. Bedford, 2013 Viral Phylodynamics, (S. Wodak, Ed.). PLoS Comput Biol 9: e1002947. [CrossRef]
- Wang M.-S., G. G. R. Murray, D. Mann, P. Groves, A. O. Vershinina, et al., 2022 A polar bear paleogenome reveals extensive ancient gene flow from polar bears into brown bears. Nat Ecol Evol 6: 936–944. [CrossRef]
- Wei W., W.-C. Ho, M. G. Behringer, S. F. Miller, G. Bcharah, et al., 2022 Rapid evolution of mutation rate and spectrum in response to environmental and population-genetic challenges. Nat Commun 13: 4752. [CrossRef]
- Wickett N. J., S. Mirarab, N. Nguyen, T. Warnow, E. Carpenter, et al., 2014 Phylotranscriptomic analysis of the origin and early diversification of land plants. Proc. Natl. Acad. Sci. U.S.A. 111. [CrossRef]
- Williams T. A., C. J. Cox, P. G. Foster, G. J. Szöllősi, and T. M. Embley, 2019 Phylogenomics provides robust support for a two-domains tree of life. Nat Ecol Evol 4: 138–147. [CrossRef]
- Worobey M., T. D. Watts, R. A. McKay, M. A. Suchard, T. Granade, et al., 2016 1970s and ‘Patient 0’ HIV-1 genomes illuminate early HIV/AIDS history in North America. Nature 539: 98–101. [CrossRef]
- Yang Z., and B. Rannala, 2010 Bayesian species delimitation using multilocus sequence data. Proceedings of the National Academy of Sciences 107: 9264–9269.
- Yu Y., R. M. Barnett, and L. Nakhleh, 2013 Parsimonious Inference of Hybridization in the Presence of Incomplete Lineage Sorting. Systematic Biology 62: 738–751. [CrossRef]
- Yuan L., H. Lu, F. Li, J. Nielsen, and E. J. Kerkhoven, 2023 HGTphyloDetect: facilitating the identification and phylogenetic analysis of horizontal gene transfer. Briefings in Bioinformatics 24: bbad035. [CrossRef]
- Yue J., X. Hu, H. Sun, Y. Yang, and J. Huang, 2012 Widespread impact of horizontal gene transfer on plant colonization of land. Nat Commun 3: 1152. [CrossRef]
- Zanewich K. P., D. W. Pearce, and S. B. Rood, 2018 Heterosis in poplar involves phenotypic stability: cottonwood hybrids outperform their parental species at suboptimal temperatures. Tree Physiology 38: 789–800. [CrossRef]
- Zeberg H., and S. Pääbo, 2020 The major genetic risk factor for severe COVID-19 is inherited from Neanderthals. Nature 587: 610–612. [CrossRef]
- Zhang R., H.-Y. Ou, F. Gao, and H. Luo, 2014a Identification of Horizontally-transferred Genomic Islands and Genome Segmentation Points by Using the GC Profile Method. CG 15: 113–121. [CrossRef]
- Zhang G., C. Li, Q. Li, B. Li, D. M. Larkin, et al., 2014b Comparative genomics reveals insights into avian genome evolution and adaptation. Science 346: 1311–1320. [CrossRef]
- Zhang C., M. Rabiee, E. Sayyari, and S. Mirarab, 2018 ASTRAL-III: polynomial time species tree reconstruction from partially resolved gene trees. BMC Bioinformatics 19: 153. [CrossRef]
- Zheng Y., and A. Janke, 2018 Gene flow analysis method, the D-statistic, is robust in a wide parameter space. BMC bioinformatics 19: 1–19.
- Zhou X., X.-X. Shen, C. T. Hittinger, and A. Rokas, 2018 Evaluating Fast Maximum Likelihood-Based Phylogenetic Programs Using Empirical Phylogenomic Data Sets. Molecular Biology and Evolution 35: 486–503. [CrossRef]
- Zhu Q., M. Kosoy, and K. Dittmar, 2014 HGTector: an automated method facilitating genome-wide discovery of putative horizontal gene transfers. BMC Genomics 15: 717. [CrossRef]
- Zhu Q., U. Mai, W. Pfeiffer, S. Janssen, F. Asnicar, et al., 2019 Phylogenomics of 10,575 genomes reveals evolutionary proximity between domains Bacteria and Archaea. Nat Commun 10: 5477. [CrossRef]
Figure 1.
A workflow for phylogenomic inference. (A) The first step in a phylogenomic workflow is data acquisition and preparation. This often entails identifying gene boundaries in genome assemblies or assembling transcripts in transcriptomes. (B) The next step is to identify orthologs using (left) de novo approaches—for example, all-by-all sequence similarity calculations followed by graph-based clustering—or (right) from predetermined sets of orthologs. (C) Orthologous groups of genes suitable for phylogenomics (i.e., one-to-one orthologs and SNAP-OGs) are subsequently aligned and trimmed. (D) The resulting multiple sequence alignments can be (left) concatenated into a supermatrix or (right) collections of single-locus phylogenies can be used in a coalescence-based approach. (E) Support for the inferred phylogeny can be evaluated using multiple approaches, such as bootstrap statistics, gene support frequencies / concordance factors, and phylogenomic subsampling. (F) Divergence time estimates can be inferred using node dating, tip dating, or fossil-free analyses. Branch lengths represent substitutions per site in the phylogeny on the left and time on the right. Grey boxes in the timetree represent confidence intervals. Silhouette images were obtained from PhyloPic (https://www.phylopic.org/); credit goes to their respective contributors.
Figure 1.
A workflow for phylogenomic inference. (A) The first step in a phylogenomic workflow is data acquisition and preparation. This often entails identifying gene boundaries in genome assemblies or assembling transcripts in transcriptomes. (B) The next step is to identify orthologs using (left) de novo approaches—for example, all-by-all sequence similarity calculations followed by graph-based clustering—or (right) from predetermined sets of orthologs. (C) Orthologous groups of genes suitable for phylogenomics (i.e., one-to-one orthologs and SNAP-OGs) are subsequently aligned and trimmed. (D) The resulting multiple sequence alignments can be (left) concatenated into a supermatrix or (right) collections of single-locus phylogenies can be used in a coalescence-based approach. (E) Support for the inferred phylogeny can be evaluated using multiple approaches, such as bootstrap statistics, gene support frequencies / concordance factors, and phylogenomic subsampling. (F) Divergence time estimates can be inferred using node dating, tip dating, or fossil-free analyses. Branch lengths represent substitutions per site in the phylogeny on the left and time on the right. Grey boxes in the timetree represent confidence intervals. Silhouette images were obtained from PhyloPic (https://www.phylopic.org/); credit goes to their respective contributors.
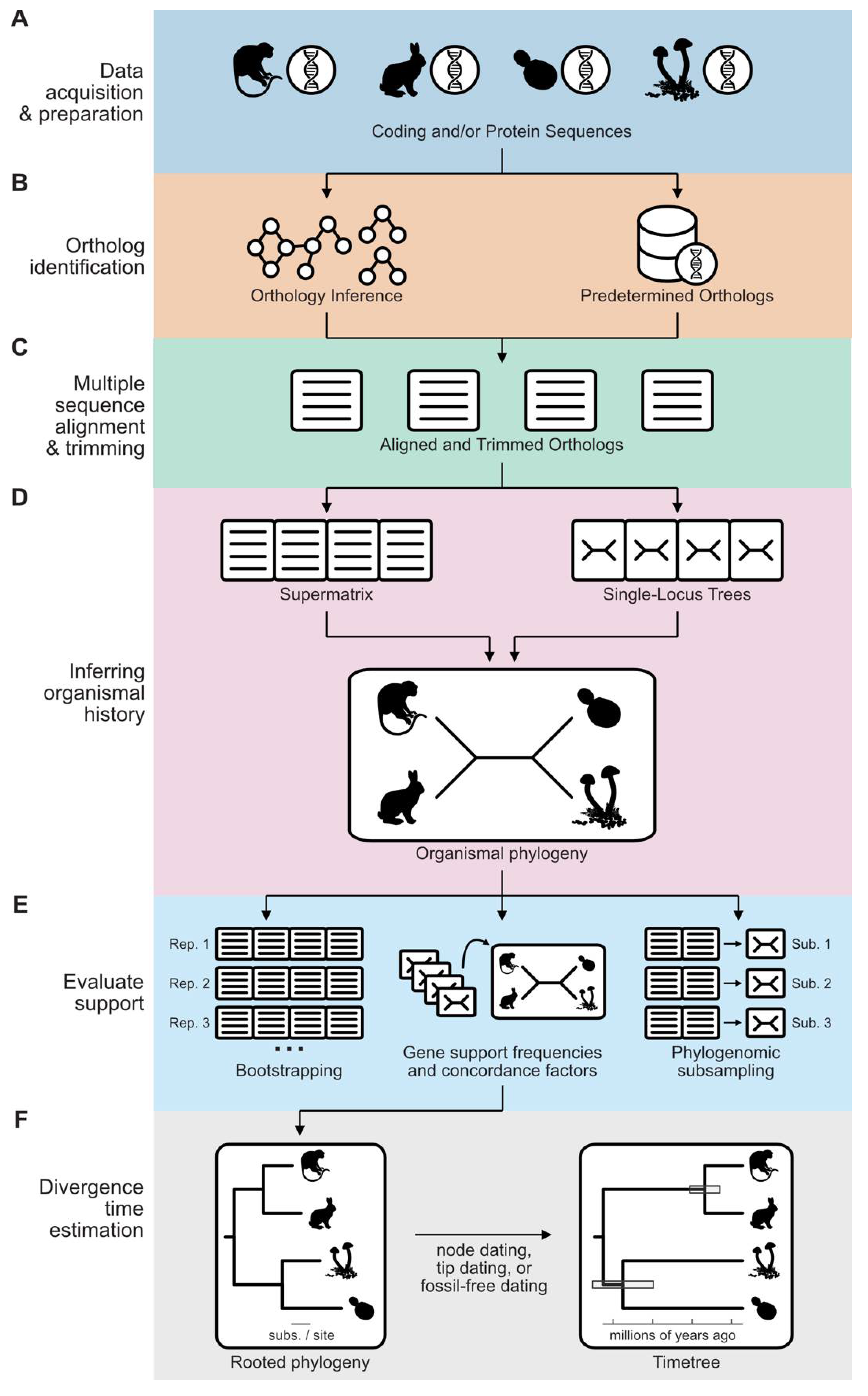
Figure 2.
Detection and differentiation of introgression/hybridization from incomplete lineage sorting. To detect introgression/hybridization in the (A) four-taxon case (represented as T1 through T4 where T4 is the outgroup), (B-E) the D-statistic or ABBA-BABA test can be used. (B) The orange dot represents a mutation from the ancestral allele ‘A’ (blue) to a derived allele ‘B’ (orange). The BBAA pattern, which is not directly accounted for in the ABBA-BABA test, is a biallelic site that follows the organismal phylogeny. Asymmetric patterns of ABBA and BABA biallelic site patterns suggest the occurrence of an introgression/hybridization event. The ABBA pattern can arise from (C) incomplete lineage sorting or (D) introgression/hybridization, whereas the (E) BABA pattern can only arise from incomplete lineage sorting; thus, unequal frequencies of ABBA and BABA patterns are suggestive of introgression/hybridization. (F-I) Coalescent times of loci subject to incomplete lineage sorting and introgression will differ. (F) Species tree depicting patterns of single-locus variation. Single locus phylogenies are also shown in panels G through I, wherein G and I depict incomplete lineage sort and H depicts the case of introgression. Of note, loci originating from introgression have a coalescent after speciation, whereas loci subject to incomplete lineage sorting coalescence before speciation.
Figure 2.
Detection and differentiation of introgression/hybridization from incomplete lineage sorting. To detect introgression/hybridization in the (A) four-taxon case (represented as T1 through T4 where T4 is the outgroup), (B-E) the D-statistic or ABBA-BABA test can be used. (B) The orange dot represents a mutation from the ancestral allele ‘A’ (blue) to a derived allele ‘B’ (orange). The BBAA pattern, which is not directly accounted for in the ABBA-BABA test, is a biallelic site that follows the organismal phylogeny. Asymmetric patterns of ABBA and BABA biallelic site patterns suggest the occurrence of an introgression/hybridization event. The ABBA pattern can arise from (C) incomplete lineage sorting or (D) introgression/hybridization, whereas the (E) BABA pattern can only arise from incomplete lineage sorting; thus, unequal frequencies of ABBA and BABA patterns are suggestive of introgression/hybridization. (F-I) Coalescent times of loci subject to incomplete lineage sorting and introgression will differ. (F) Species tree depicting patterns of single-locus variation. Single locus phylogenies are also shown in panels G through I, wherein G and I depict incomplete lineage sort and H depicts the case of introgression. Of note, loci originating from introgression have a coalescent after speciation, whereas loci subject to incomplete lineage sorting coalescence before speciation.

Figure 3.
Signatures of a horizontal gene transfer event and their applicability to relative divergence estimation. To detect horizontal gene transfer, (A) organismal histories are compared to (B) single-locus phylogenies. Horizontal gene transfer is suggested when sequences are placed within an unexpected taxonomic group in single-locus phylogenies. Here, an example of prokaryote-to-eukaryote transfer is depicted wherein an organism from the fungal kingdom (orange) is monophyletic with prokaryotic sequences (blue). The horizontal gene transfer event is depicted as a grey arrow. (C) Similar horizontal gene transfer events can be used to estimate relative divergences, especially in lineages where horizontal gene transfer occurs sufficiently frequently. Specifically, consider the scenario of two competing phylogenetic hypotheses wherein the clade containing taxa E and F diverged before or after the clade with A and B (hypotheses 1 and 2 or H1 and H2, respectively). (D) A horizontal gene transfer event that would support H1 would involve the ancestor of A and B transferring a locus to the ancestor of E and F, whereas (E) a horizontal gene transfer event that would support H2 would involve the transfer of a locus to E and F from a lineage along the stem branch leading to A. In panels D and E, the left panel depicts the horizontal gene transfer event and the rght panel depicts the phylogenetic tree of the horizontally transferred locus. HGT is an abbreviation for horizontal gene transfer. Silhouette images were obtained from PhyloPic (https://www.phylopic.org/); credit goes to their respective contributors.
Figure 3.
Signatures of a horizontal gene transfer event and their applicability to relative divergence estimation. To detect horizontal gene transfer, (A) organismal histories are compared to (B) single-locus phylogenies. Horizontal gene transfer is suggested when sequences are placed within an unexpected taxonomic group in single-locus phylogenies. Here, an example of prokaryote-to-eukaryote transfer is depicted wherein an organism from the fungal kingdom (orange) is monophyletic with prokaryotic sequences (blue). The horizontal gene transfer event is depicted as a grey arrow. (C) Similar horizontal gene transfer events can be used to estimate relative divergences, especially in lineages where horizontal gene transfer occurs sufficiently frequently. Specifically, consider the scenario of two competing phylogenetic hypotheses wherein the clade containing taxa E and F diverged before or after the clade with A and B (hypotheses 1 and 2 or H1 and H2, respectively). (D) A horizontal gene transfer event that would support H1 would involve the ancestor of A and B transferring a locus to the ancestor of E and F, whereas (E) a horizontal gene transfer event that would support H2 would involve the transfer of a locus to E and F from a lineage along the stem branch leading to A. In panels D and E, the left panel depicts the horizontal gene transfer event and the rght panel depicts the phylogenetic tree of the horizontally transferred locus. HGT is an abbreviation for horizontal gene transfer. Silhouette images were obtained from PhyloPic (https://www.phylopic.org/); credit goes to their respective contributors.
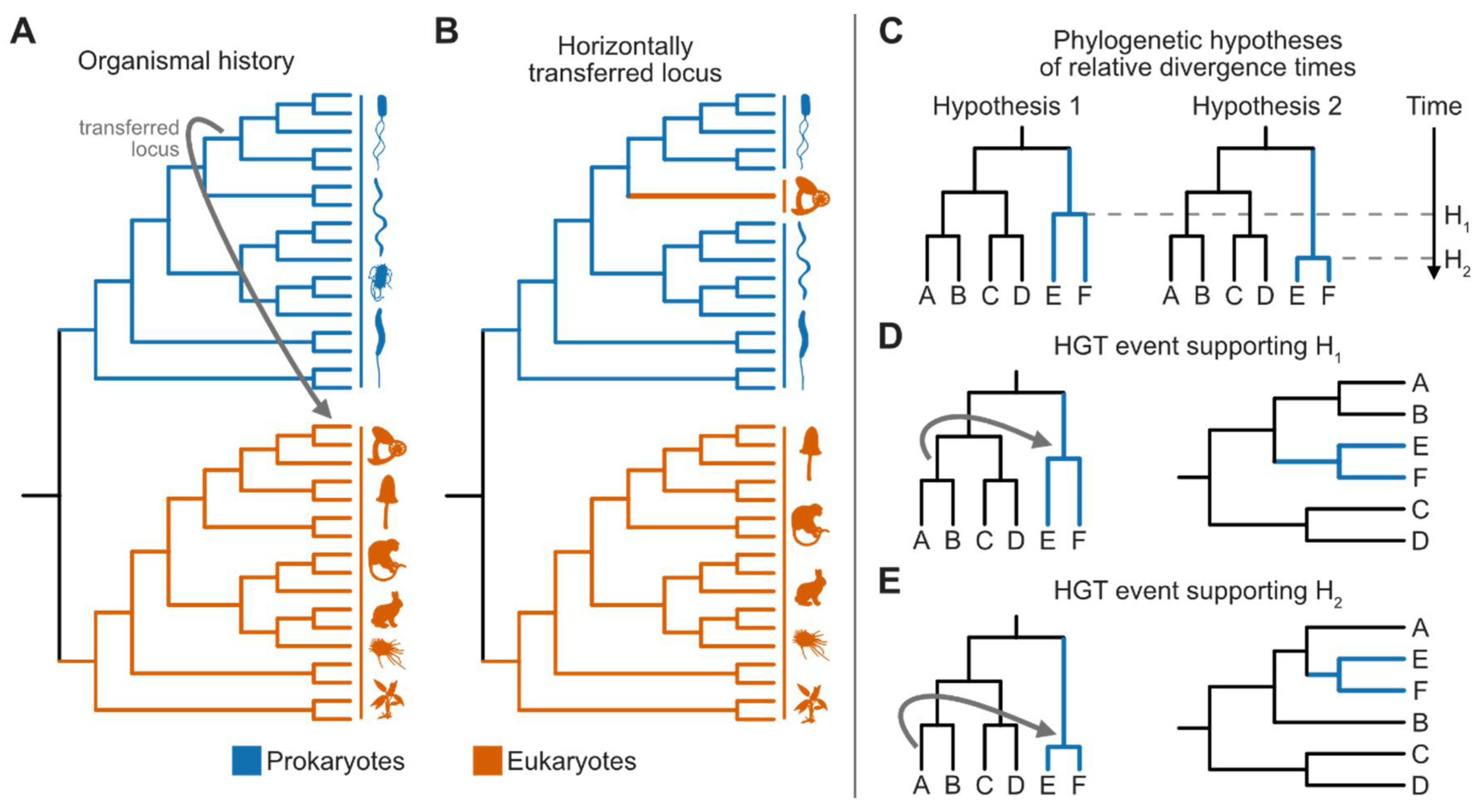
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



