
Submitted:
13 September 2023
Posted:
15 September 2023
You are already at the latest version
Abstract
Our findings indicate that all five phytocannabinoids reduce HG-HL-induced -cell loss likely through reducing apoptosis and pyroptosis. The protective effects of CBD, THCV, CBC, and CBN were seen in the GSIS impairment by HG-HL. Although all five phytocannabinoids tested in this research demonstrated the capability to inhibit β-cell dedifferentiation induced by HG-HL, CBD seems to be more effective compared to the other phytocannabinoids, as indicated by the specific biomarker responses of β-cells and progenitor cells to CBD.
Keywords:
β-cell loss
; β-cell dedifferentiation
; diabetes
; GSIS
; THC
; CBD
; THCV
; CBC
; CBN
1. Introduction
Diabetes is characterized by abnormally high blood glucose levels and disruptions in the metabolism of carbohydrates, proteins, and fats. Moreover, diabetes is associated with the development of comorbidities such as atherosclerosis, retinopathy, neuropathy, and nephropathy [1]. Based on the projections made by the International Diabetes Federation (IDF), around 537 million individuals globally were diagnosed with diabetes in 2021. This is anticipated to increase to approximately 643 million by the year 2030 and further rise to about 783 million by 2045.
Type II diabetes mellitus (T2DM) is the predominant form of diabetes, representing the majority of cases worldwide. Although typically seen in adults, there has been a notable increase in the diagnosis of T2DM among children in recent years. Several key factors contribute to its development, including obesity, lack of physical activity, and the adoption of a Westernized diet [1].
The initial stage of T2DM involves the development of insulin resistance. Insulin resistance is a term used to describe a condition where cells, such as hepatocytes, adipocytes, and muscle cells, are unable to properly detect and respond to insulin. As a consequence, this leads to disruptions in the regulation of glucose and lipid levels within the body [2]. Insulin resistance often develops several years before the onset of T2DM and is characterized by high levels of insulin in the blood due to increased function and mass of β-cells. This prolonged strain on β-cells results in excessive insulin production and secretion, which can lead to glucolipotoxicity, inflammation, and ultimately β-cell dysfunction and loss, and thereby the development of T2DM [1].
Elevated levels of free fatty acids (FFAs) in the bloodstream caused by obesity and the consequent insulin resistance in adipose tissue have detrimental effects on the function and viability of β-cells through multiple mechanisms. These mechanisms include endoplasmic reticulum (ER) stress, proinflammatory responses, and impaired insulin signaling. As a result, these factors contribute to a decline in glucose stimulated insulin secretion (GSIS), trigger apoptosis, and ultimately lead to the loss of β-cells [3]. Consistently, the occurrence of persistent ER stress within β-cells, caused by an increased demand for insulin production during insulin resistance stage, can exacerbate the situation and contribute to the malfunction and decline of β-cells [4].
Recent findings indicate that the primary cause of diabetes is the malfunction of β-cells [5]. Consistently, the failure of β-cells in diabetes is primarily caused by an increase in β-cell dedifferentiation, rather than a decrease in β-cell proliferation or an increase in β-cell death. Studies have observed characteristics of β-cell dedifferentiation in mouse pancreatic insulinoma β-cells exposed to high levels of glucose for an extended period, without significant cell death occurring [6]. A recent investigation has revealed that individuals with T2DM exhibit a loss of identity of β-cells and their mass, which may further manifest as the acquisition of novel characteristics resembling other cell types within the islets. This study suggests that the dysfunction of β-cells, resulting in the onset of T2DM, can be attributed to the phenomenon of islet remodeling and dedifferentiation [7].
During β-cell dedifferentiation, there is a downregulation of genes that are typically expressed in β-cells, such as transcription factors, insulin, and genes involved in glucose metabolism. At the same time, there is an upregulation of genes that are normally suppressed in healthy β-cells, as well as genes associated with islet progenitor cells and other mature islet cell types. This process involves a shift in gene expression patterns away from the β-cell phenotype towards a more progenitor-like state [8]. The identity of pancreatic β-cells is regulated by the precise control of several key transcription factors, including pancreatic and duodenal homeobox 1 (PDX1), v-maf musculoaponeurotic fibrosarcoma oncogene homolog A (MafA), neurogenic differentiation 1 (NEUROD1), NK6 homeobox 1 (Nkx6.1), and forkhead box protein O1 (FOXO1) [9]. Under certain pathological conditions such as hyperglycemia and hyperlipidemia, β-cell dedifferentiation can occur as a result of increased oxidative stress, endoplasmic reticulum (ER) stress, and the presence of inflammatory cytokines [10]. Subsequent studies have further demonstrated the occurrence of β-cell dedifferentiation or trans-differentiation in both animal models and individuals with T2DM [11,12].
The endocannabinoid system is distributed extensively across the body and serves diverse regulatory functions in various organs [13]. By exerting regulatory influence over appetite, food intake, and the production of fats (lipogenesis), the activation of the endocannabinoid system, specifically through CB1R receptors in both central and peripheral regions, has been associated with metabolic disorders such as obesity, diabetes, and their related complications [4]. The existence of the endocannabinoid system has been documented in both rodent and human pancreatic islets [14].
Cannabis sativa contains a variety of phytocannabinoids that can modulate the endocannabinoid system through both receptor-dependent and receptor-independent mechanisms. Delta-9-tetrahydrocannabinol (THC) and cannabidiol (CBD) are the two major cannabinoids that have been extensively studied for their therapeutic potential. In addition to these major cannabinoids, minor phytocannabinoids such as tetrahydrocannabivarin (THCV), cannabichromene (CBC), and cannabinol (CBN) have also been shown to have potential therapeutic effects on various inflammatory and metabolic disorders, including T2DM, by targeting different components of the endocannabinoid system [4]. Although some publications have explored the potential antidiabetic properties of certain phytocannabinoids, a more extensive analysis is necessary to thoroughly examine the influence of major and minor phytocannabinoids on different stages involved in the onset of T2DM. These stages include the loss of β-cells, impaired GSIS, and β-cell dedifferentiation. In our present research, we aimed to elucidate the potential impacts of five major and minor phytocannabinoids on GSIS and β-cell viability and dedifferentiation through an in vitro study.
2. Materials and Methods
2.1. Chemicals and reagents
THC (6465-30-1), CBD (13956-29-1), THCV (T-094-1ML), CBC (20675-51-8), CBN (521-35-7), L-Glutamine (TMS-002-C), sodium pyruvate (S8636), HEPES (TMS-003-C), β-mercaptoethanol (ES-007), INS-1 832/13 Rat Insulinoma Cell (SCC207) were purchased from EMD Millipore Corporation (Temecula, CA, USA). Roswell Park Memorial Institute Medium (RPMI-1640) (350-060-CL) and PBS, 1X(311-010-CL), were acquired from Wisent Inc. (Saint-Jean-Baptiste, QC, Canada).
2.2. Cell culture and treatments
INS-1 832/13 Rat Insulinoma Cells were cultured in RPMI 1640 medium supplemented with 2 mM L-Glutamine, 1 mM sodium pyruvate, 10 mM HEPES, 0.05 mM β-mercaptoethanol, and 10% fetal bovine serum (FBS) ((10082147) acquired from Fisher Scientific Company, Ottawa, ON, Canada), along with 2.5 mM glucose. Cells within passages 5-10 were used for all experiments. After being cultured in a medium containing 2.5 mM glucose for two days, cells were exposed to 5 μM of individual phytocannabinoid and/or an equal quantity of the vehicle (methanol) for a duration of 2 hours. Subsequently, High Glucose (HG) (25 mM glucose) and High Lipid (HL) (400 μM palmitic acid) conditions were imposed for the next 48 hours. Following the experimental treatments, RNA and protein were extracted from the treated cells to perform quantitative real-time polymerase chain reaction (qRT-PCR) and western blot analysis, respectively.
2.3. MTT assay
The impact of five phytocannabinoids on β-cell viability under HG-HL conditions was evaluated using an MTT assay. Cells were seeded at a density of 5 × 104 cells/well in 100 μl of culture medium containing 5 μM of each phytocannabinoid or an equal volume of vehicle (methanol) for 2 hours. The cells were then incubated with medium containing phytocannabinoids, vehicle, and/or HG (25 mM glucose)-HL (400 μM palmitic acid) for 48 hours at +37°C and 5% CO2. After incubation, 10 μl of 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) labeling reagent (11465007001) purchased from MilliporeSigma Canada Ltd. Oakville, ON, Canada) was added to each well and the microplate was incubated for 4 hours in a humidified atmosphere (+37°C, 5% CO2). Next, 100 μl of solubilization solution was added to each well and the plate was left to stand overnight in the humidified incubator. The absorbance at 595 nm was measured using a plate reader (FLUOstar Omega, BMG LABTECH, Offenburg, Germany).
2.4. Western blotting
The cells were rinsed twice with cold PBS and then lysed using RIPA buffer. After centrifugation at 13,000 rpm for 15 minutes, the supernatant was collected and transferred to a new microtube. The protein content was determined using the Bradford assay and the lysates were used for western blotting. Proteins were separated on polyacrylamide gels of varying concentrations (8%, 10%, and 12%) and transferred to polyvinylidene difluoride (PVDF) Amersham Hybond® P membranes (RPN2020F) acquired from GE Healthcare, Oakville, ON, Canada. The membranes were blocked with a PBS containing 1% tween 20 (PBST) solution and 5% milk before being incubated with primary antibodies overnight at 4°C. After three washes with PBST, the membranes were incubated with secondary antibodies for two hours at room temperature and washed again with PBST. Immunoreactivity was identified using peroxidase-conjugated antibodies, and the ECL Plus Western Blotting Detection System (GE Healthcare, Oakville, ON, Canada) was utilized to render the reaction visible. The band intensities were quantified and normalized relative to β-Actin’s intensity through analysis with ImageJ. Primary antibodies used: Caspase 3 (9662s), Caspase 7 (12827), Caspase 1 (89332), Cleaved-PARP (9545s), Bax (2772s), PDX-1 (D59H3), FOXO1 (2880S), and TXNIP (14716S) were sourced from Cell Signal Technology, Whitby, ON, Canada. IL-1β (sc-12742) and secondary antibodies were obtained from Santa Cruz Biotechnology, Dallas, TX, U.S.A. Actin antibody (ab8227) was purchased from Abcam, Toronto, ON, Canada.
2.5. qRT-PCR
Total RNA was isolated using TRIzolTM (15596018, Invitrogen, Life Technologies Inc., Burlington, ON, Canada), and its concentration was subsequently determined using a nanodrop instrument (NanaDrop 2000c, ThermoFisher Scientific, Waltham, MA, USA). A portion of the total RNA was used for cDNA synthesis, utilizing 1 μg of total RNA and a cDNA synthesis kit, iScript™ Reverse Transcription Supermix (1708897, BioRad Laboratories, Saint-Laurent, QC, Canada). The resulting cDNA was then used as a template for q-PCR, with 1 μl of cDNA being used per reaction. The qRT-PCR reactions were performed using a SsAdvancedTM Universal Inhibitor-Tolerant SYBR Green Supermix (1725017, Bio-Rad Laboratories, Saint-Laurent, QC, Canada). We procured the primers necessary for our experiments from Eurofins (Ottawa, ON, Canada) (Supplementary Table S1).
2.6. Glucose stimulated insulin secretion (GSIS)
A GSIS assay was conducted to examine the effects of THC, CBD, THCV, CBC and CBN on the response of β-cells exposed to HG-HL conditions. Cells were plated at a density of 0.5x106/well in a 24-well plate and incubated for 2 days. Subsequently, cells were pretreated with media containing the phytocannabinoids at a concentration of 5 µM or vehicle control for 2 hours before being subjected to HG-HL conditions for 48 hours. The GSIS assay was performed using HEPES Balanced Salt Solution (HBSS) containing specific concentrations of various salts and bovine serum albumin at pH 7.2. Cells were washed twice with HBSS containing 2.5 mM glucose, with the second wash lasting for 1 hour. Each treatment was assigned two wells. One well was treated with HBSS containing the respective phytocannabinoid at a concentration of 5 µM and 2.5 mM glucose (as normal glucose), while the other well was treated with HBSS containing the respective phytocannabinoid at a concentration of 5 µM and 16.5 mM glucose (as high glucose). Both wells were incubated for 2 hours before the solution was removed for insulin ELISA analysis.
2.7. Enzyme-Linked Immunosorbent (ELISA)
In order to measure GSIS, ELISA assay was performed using Rat/Mouse Insulin ELISA kit (EZRMI-13K), purchased from Sigma Aldrich (Oakville, ON, Canada). The strips were placed in an empty plate holder and each well was washed thrice with 300 μL of diluted Wash Buffer. 10 μL of each sample was added to the wells, followed by the addition of 80 μL of Detection Antibody. The plate was sealed and incubated for 2 hours at room temperature on an orbital microtiter plate shaker rotating at a moderate speed of 400-500 rpm. The plate sealer was removed, and the solutions were decanted. The wells were washed thrice with diluted Wash Buffer and tapped on a paper towel to remove residual buffer. 100 μL of Enzyme Solution was added to each well and the plate was sealed and incubated with moderate shaking for 30 minutes. The wells were then washed six times with diluted Wash Buffer, followed by adding 100 μL of Substrate Solution to each well. The plate was sealed and shaken for 5-20 minutes. After removing the sealer, 100 μL of Stop Solution was added and the plate was briefly shaken to ensure complete mixing. The absorbance was measured at 450 nm and 590 nm using a plate reader (SpectraMax i3x Multi-Mode Microplate Reader, Molecular Devices, San Jose, USA).
2.8. Statistics
The collected data were subjected to statistical analysis using one-way analysis of variance (ANOVA) followed by Dunnett's and Turkey’s tests to compare the mean values, performed using GraphPad Prism 6 software.
3. Results
In all experiments, β-cells were cultivated in RPMI 1640 medium supplemented with 2.5 mM glucose over a span of two days. Subsequently, the cells were subjected to a pre-treatment phase involving exposure to 5 μM concentrations of individual phytocannabinoids as well as an equivalent volume of a vehicle substance for a duration of two hours. In the next 48 hours, the cells were subjected to HG-HL conditions, while simultaneously ensuring the presence of the phytocannabinoids and/or vehicle in the culture medium. In this study, our goal was to investigate the impact of five phytocannabinoids, including two major and three minor compounds, on β-cell loss and dedifferentiation. We selected an optimal dose of 5 μM based on our optimization trials and prior research [15,16]
3.1. Pre-treatment with 5 μM THC, CBD, THCV, CBC, and CBN Increased the Viability of β-cells under HG-HL Conditions.
We first looked at the effect of THC, CBD, THCV, CBC, and CBN on HG-HL-induced β-cell loss. The MTT assay showed that treating β-cells with 5 μM of each phytocannabinoid resulted in a significant increase in cell viability under HG-HL conditions (Figure 1). It is important to note that the initial cell viability in the 96-well plate was consistent, indicating that an equal number of cells were seeded in each well on day 0.
3.2. CBD, THCV, CBC, and CBN Improved GSIS in HG-HL-Treated β-cells
In order to investigate the impact of individual phytocannabinoids on impaired GSIS in HG-HL-challenged β-cells, we performed GSIS assay. The cells were pretreated with 5 μM of each phytocannabinoid or an equivalent volume of the vehicle (methanol) for a period of 2 hours. Following this, the experimental groups were subjected to High Glucose (HG) conditions (25 mM glucose) and High Lipid (HL) conditions (400 μM palmitic acid) for the subsequent 48 hours. Subsequently, the cells were washed with HBSS containing 2.5 mM glucose, and GSIS was assessed using HBSS containing both 2.5 mM and 16.5 mM glucose for each treatment. It is important to note that the phytocannabinoids were present in the HBSS during the GSIS assay.
Based on our findings, THC did not demonstrate any favorable effects on impaired GSIS in HG-HL-induced β-cells. However, THCV, CBD, CBC, and CBN exhibited notable improvements in this response compared to the vehicle (Figure 2).
3.3. The Phytocannabinoids Mitigated HG-HL-Induced β-cells Loss Through Reduction of Apoptosis and Pyroptosis
To elucidate the underlying mechanisms by which the phytocannabinoids mitigate β-cell loss induced by HG-HL conditions, we performed western blot analysis of Caspase-7, Caspase-3, Caspase-1, Cleaved PARP, and Pro-IL-1β, the key molecules involved in apoptosis and pyroptosis. The results demonstrated that THC, THCV, and CBD reduced the levels of Pro-Caspase-7 in HGHL-challenged β-cells, while CBC and CBN were not effective (Figure 3a). However, all five phytocannabinoids showed inhibitory effects on the levels of Cleaved-Caspase-7 (C-Caspase-7) (Figure 3b). In addition to Caspase-7, Caspase-3 is a pivotal biomarker of apoptosis. Notably, all five phytocannabinoids investigated in this study exhibited suppressive effects on the levels of Cleaved-Caspase-3 (C-Caspase-3) in HG-HL-induced β-cells (Figure 3c). These results suggest that five phytocannabinoids inhibit the HG-HL-triggered apoptosis in β-cells via downregulation of C-Caspase-3 and C-Caspase-7.
Pyroptosis, another form of cellular death, is mediated by the activation of the Nlrp3 inflammasome, cleavage of Pro-Caspase-1, and subsequent production of the mature form of IL-1β and Gasdermin N. This process leads to the release of proinflammatory cytokines, pore formation in the cell membrane, cell swelling, and eventual bursting, resulting in the recruitment of immune cells and initiation of inflammatory responses, and eventually cell loss [17]. Our findings indicated that THC, CBD, and CBC downregulated the levels of activated C-Caspase-1 in HG-HL-challenged β-cells (Figure 3d). Furthermore, all five phytocannabinoids demonstrated a mitigative effect on the immature form of IL-1β in HG-HL-challenged β-cells compared to the HG-HL-only group (Figure 3e).
PARP, an enzyme involved in DNA repair, is a well-established marker of apoptosis. Cleavage of PARP by activated Caspases, such as Caspase-3 and Caspase-7, serves as a characteristic biochemical indicator of apoptotic cell death. In our study, all five phytocannabinoids reduced the PARP cleavage and, consequently, the generation of the 89 kDa N-terminal fragment (Figure 3f). This reduction in PARP cleavage is likely mediated through the mitigation of Caspase-3 and Caspase-7 activity.
3.4. Treatment with THC Restores the Decreased Level of PDX-1 Protein and NEUROD1 and Slc2A2 Transcripts in HG-HL-Induced β-cells
PDX1, also known as the pancreas/duodenum homeobox protein 1 or insulin promoter factor 1, is a transcription factor that plays a crucial role in the early development of the pancreas during embryogenesis, as well as in the differentiation of pancreatic cell lineages and the maintenance of β-cell function [18]. Based on the results obtained from western blot analysis, all three concentrations of THC were able to replenish the levels of PDX-1 in HG-HL-stimulated β-cells (Figure 4-a1). However, this effect is likely unrelated to the influence of THC on the expression of the PDX-1 gene, as evidenced by qRT-PCR results for PDX-1 (Figure 4b). Additionally, THC had a positive effect on the response of FOXO1 protein in HG-HL-induced β-cells (Figure 4-a3). Our western blot results also indicated that treatment with THC reduce the induced level of THXNIP in HG-HL-treated β-cells (Figure 4-a2).
To further explore the effects of THC on β-cell dedifferentiation, the expression of several genes involved in this process was examined.
Rats and mice have two insulin genes, Ins1 and Ins2, which are structurally similar but not identical. Ins1 is a duplication of the ancestral Ins2 gene. The two preproinsulins, which are coded by these two genes, exhibit closely related polypeptide sequences, differing only in a few amino acids. Although both genes are functional, their relative expression in rodent β-cells is not well understood [19]. Some studies suggest that the two insulins are present in roughly equal amounts [20], while others indicate that either Ins1 or Ins2 is the predominant gene in mice [21,22]. According to our results, the treatment with 5 μM THC did not significantly affect the transcription of the Ins1 and Ins2 genes in HG-HL-challenged β-cells (Figure 4b).
NEUROD1 is a transcription factor that was first discovered as a transactivator of the insulin gene [23]. Further research has shown that it plays a broader role in regulating β-cell function by directly activating genes involved in β-cell maturation and function. Studies on mice have shown that NEUROD1 is required for the development of functional α- and β-cells in a cell type- and age-dependent manner. During pancreas development in mice, NEUROD1 is expressed by differentiating endocrine cells and later becomes restricted to insulin-producing β-cells and a subset of glucagon-expressing α-cells [24]. Inactivating mutations in the NEUROD1 gene can cause different types of diabetes [25]. Accordingly, homozygous mutations can result in permanent neonatal diabetes mellitus, while heterozygous mutations can lead to maturity-onset diabetes [26]. In our experiments, 5 μM THC restored the level of NEUROD1 gene in HG-HL-challenged β-cells (Figure 4b).
The Slc2A2 gene is responsible for producing the GLUT2 glucose transporter, which is part of a family of transporters that help glucose enter cells through the plasma membrane. GLUT2 is mainly found in liver cells, pancreatic β-cells, and in the intestines and kidneys. It plays a crucial role in the process of GSIS and is necessary for proper β-cell function in rodents. However, human β-cells also have GLUT1 and GLUT3 transporters and do not depend solely on GLUT2 for this process [27]. Like in the case of NEUROD1, THC upregulated Slc2A2 in HG-HL-induced β-cells (Figure 4b).
The MafA insulin gene transcription factor, which is found in mature β-cells, does not play a role in endocrine specification during embryonic development [28,29]. However, the ectopic expression of MafA, PDX1, NeuroD1 or Ngn3 can transform adult pancreatic acinar cells or liver cells into insulin-producing cells [30,31]. Reduced MafA expression is also an early sign of β-cell dysfunction in human T2DM [32]. The results of the current study showed that administering 5μM THC somewhat restored the expression of MafA in HG-HL-challenged β-cells, although the difference was not significant (p=0.08) (Figure 4b).
The biomarkers related to progenitor cells such as Pou5f1, Nanog, and L-MYC transcription factors are upregulated in T2DM [33]. In the current study, we looked at the response of two of these transcription factors, Pou5f1, Nanog, to all five phytocannabinoids. Our findings indicate that THC has the ability to downregulate the expression of both Pou5f1 and Nanog transcription factors in HG-HL-challenged β-cells (Figure 4b).
3.5. CBD Treatment Modulates the Response of Most Genes and Proteins Associated with β-Cell Dedifferentiation
Our results demonstrate that CBD has a stimulatory effect on the response of the PDX-1 protein in HG-HL-challenged β-cells, with all three doses of CBD eliciting a similar response (Figure 5-a1). Interestingly, CBD was found to increase the level of TXNIP in HG-HL-induced β-cells (Figure 5-a2). Treatment with 5μM CBD upregulated the expression of Ins1, PDX-1, Slc2A2, NEUROD1, MafA, and FOXO1 genes in HG-HL-induced β-cells, while not affecting the response of Ins2 (Figure 5b). Like THC, CBD was able to downregulate Pou5f1 and Nanog progenitor cell-related biomarkers in HG-HL-induced β-cells. However, unlike THC, it also downregulated these two transcription factors in the control groups (Figure 5b).
3.6. THCV Treatment Restores the Levels of PDX-1 protein and Ins-1, NEUROD1 and Slc2A2 Transcripts in HG-HL-Treated β-cells
Our findings indicate that the administration of 5 μM THCV elicits a stimulating impact on the expression of the PDX-1 protein within β-cells exposed to HG-HL, as illustrated in Figure 6-a1. Unlike CBD, THCV was observed to decrease the level of TXNIP in HG-HL-induced β cells (Figure 6-a2). Treatment with 5 μM THCV upregulated the expression of Ins1, NEUROD1, MafA, and FOXO1 genes in HG-HL-challenged β-cells, while having no effect on the response of Ins2 and PDX-1 transcripts (Figure 6b). THCV was capable of downregulating Pou5f1 and Nanog biomarkers associated with progenitor cells in HG-HL-challenged β-cells. Furthermore, 5 μM THCV exhibited an inhibitory effect on the expression of Slc2A2.
3.7. Treatment with CBC Modulates Elevated TXNIP Protein Levels and Reduced Slc2A2 Transcript Levels in HG-HL-Induced β-cells
Our findings suggest that the introduction of 5 μM CBC did not elicit any discernible changes in the level of the PDX-1 protein within β-cells subjected to high levels of glucose and lipids, as depicted in Figure 7-a1. In a similar fashion to THCV, CBC reduced the expression of TXNIP in HG-HL-induced β-cells (Figure 7-a2). While the treatment of HG-HL-challenged β-cells with 5 μM CBC resulted in an upregulation of Slc2A2 and FOXO1 gene expression, it did not display any positive influence on the response of Ins1, Ins2, PDX-1, NEUROD1, and MafA (Figure 7b). Conversely, CBC was capable of downregulating Pou5f1 and Nanog biomarkers associated with progenitor cells in HG-HL-challenged β-cells.
3.8. CBN Treatment Reduces Elevated TXNIP Levels and Increases the PDX-1 Protein and Slc2A Transcript Levels in HG-HL-Induced β-cells
5 μM CBN positively impacted the PDX-1 protein levels in HG-HL-induced β-cells (Figure 8-a1). Like THCV and CBC, CBN reduced the expression of TXNIP in HG-HL-challenged β-cells (Figure 8-a2). While 5 μM CBN treatment in HG-HL-challenged β-cells increased Slc2A2 and FOXO1 gene expression, it did not have any positive effects on the response of Ins1, Ins2, PDX-1, NEUROD1, and MafA (Figure 8b). Additionally, CBN was able to reduce the expression of Pou5f1 and Nanog biomarkers associated with progenitor cells in HG-HL-challenged β-cells (Figure 8b).
4. Discussion
4.1. The effect of THC on HG-HL-challenged β-cells
THC, the primary psychoactive phytocannabinoid synthesized by Cannabis sativa, is widely recognized. It acts as an agonist of cannabinoid receptor 1 (CB1R) and a partial agonist of cannabinoid receptor 2 (CB2R). Notably, the psychotropic and appetite-enhancing effects attributed to THC stem from its activation of CB1R, mimicking the behavioral patterns observed with anandamide (AEA ) [34]. As an agonist of CB1R, THC may not be considered a potential treatment for prediabetes, diabetes, or its complications. Reports on the effectiveness of THC in mitigating insulin resistance and the progression of diabetes and its complications are inconsistent [35,36,37]. However, some studies have suggested that THC may exert therapeutic effects on diabetes and its complications through its partial agonistic effects on CB2R [38], potent antioxidant activities, and/or agonistic effects on PPARs, resulting in anti-inflammatory properties [39].
Our results indicate that 5 µM THC can mitigate β-cell loss under HG-HL conditions, likely through the inhibition of apoptosis and/or pyroptosis. THC was found to downregulate several key apoptotic biomarkers, including C-Caspase-3, pro- and C-Caspase-7, and C-PARP. Pyroptosis is a type of cell death mediated by the activation of the Nlrp3 inflammasome and subsequent cleavage of Pro-Caspase-1 into Caspase-1. In our work, THC was found to have an inhibitory effect on C-Caspase-1, suggesting another mechanism by which THC may mitigate HG-HL-induced β-cell loss. Dysregulation of IL-1β in β-cells plays an important role in pyroptosis and the initiation of β-cell loss in T2DM [4]. THC has been shown to have an inhibitory effect on the response of Pro-IL-1β, providing further evidence for its mitigative effects on pyroptosis. Thioredoxin-interacting protein (TXNIP), also known as thioredoxin-binding protein-2, is a member of the α-arrestin protein family and is regulated by various cellular stress factors. It acts as an endogenous inhibitor of thioredoxin (Trx) by reducing its capacity, leading to increased cellular oxidative stress. In addition to its role in regulating redox homeostasis, TXNIP is also involved in the regulation of multiple signaling pathways [40]. TXNIP can induce the activation of the Nlrp3 inflammasome and subsequent pyroptosis. Our western blot results showed that THC can decrease the level of this protein in β-cells exposed to HG-HL conditions, suggesting an additional mechanism by which THC mitigates pyroptosis. It is important to note that THC may partially inhibit HG-HL-induced apoptosis by reducing TXNIP levels, due to the crucial role of TXNIP in triggering ROS and subsequent apoptosis in HG-HL-challenged β-cells.
Despite the fact that THC upregulated the impaired transcription of the Slc2A2 gene in HG-HL-challenged β-cells, it did not improve GSIS in HG-HL-stimulated β-cells. This may be due to the agonistic effects of THC on CB1R, leading to a reduction in intracellular cAMP and Ca2+, activation of tyrosine kinases, and interaction between CB1R and insulin receptor [4].
THC did not exhibit any restoring effects on HG-HL-triggered downregulation of the Ins1 and Ins2 genes. This may be attributed to the association between CB1R activation and a decrease in intracellular cAMP levels, leading to the inhibition of CREBP phosphorylation. Given the presence of a CREB-responsive element in the promoter region of the insulin-encoding gene, this may provide an explanation for the lack of significant effects of THC on the expression of these genes [41].
Additionally, 5 µM THC increased the level of NEUROD1 transcript, as well as PDX-1 protein, and decreased the levels of Nanog and Pou5f1 mRNA in β-cells exposed to HG-HL conditions. This suggests that THC may mitigate HG-HL-induced β-cell dedifferentiation (Figure 9).
4.2. The effect of CBD on HG-HL-challenged β-cells
CBD is a prominent non-psychoactive phytocannabinoid extracted from the Cannabis sativa plant. Recent reports have demonstrated that CBD exhibits a wide range of noteworthy pharmacological properties. Due to its potent anti-inflammatory properties, CBD has the potential to serve as a treatment for insulin resistance, diabetes, and associated complications [4]. Similar to THC, CBD exerted a suppressive effect on HG-HL-induced β-cell loss by inhibiting apoptosis and/or pyroptosis. In line with this, CBD demonstrated a downregulatory effect on crucial apoptotic biomarkers, including C-Caspase-3, pro- and C-Caspase-7, and C-PARP.
At a concentration of 5μM, CBD downregulated two significant biomarkers of pyroptosis, including C-Caspase-1 and pro-IL-1β. In line with these findings, it has been demonstrated that both CBD and THC can reduce the release of IL-1β by inhibiting the first and second phases of Nlrp3 inflammasome activation [15]. Interestingly, CBD was found to upregulate the protein TXNIP in HG-HL-challenged β-cells.
Unlike THC, CBD improved impaired GSIS response in HG-HL-stimulated β-cells, likely mediated through the upregulation of MafA and subsequent upregulation of Slc2A2. Treatment of HG-HL-challenged β-cells with 5 μM CBD increased the level of Ins-1, but not Ins-2. This may be related to the antagonistic effects of CBD on CB1R and subsequent increase in intracellular cAMP. It is noteworthy that other transcription factors studied in this research could also affect the transcription of the gene encoding insulin. In this regard, PDX-1 exerts direct regulatory control over insulin transcription via the formation of a complex with transcriptional coactivators at the proximal insulin promoter [42]. Furthermore, MafA has been identified as a key regulator of insulin gene transcription [43]. It is noteworthy that NEUROD1 has also been shown to play a crucial role in inducing insulin gene transcription in pancreatic β-cells [44]. The increase in PDX-1, MafA, and NEUROD1 mRNA levels with 5 μM CBD provides potential evidence for the anti-dedifferentiation activity of CBD on β-cells (Figure 9). Interestingly, CBD reduced the expression of two key progenitor-related transcription factors, Nanog and Pou5f1, in HG-HL-challenged β-cells, further supporting the inhibitory effects of CBD on HG-HL-challenged β-cell dedifferentiation.
Peroxisome proliferator-activated receptor gamma (PPARγ) is a nuclear transcription factor that has been demonstrated to regulate the expression of key β-cell genes, including PDX-1, Slc2A2, and glucokinase [37,45]. CBD has exhibited agonistic effects on PPARγ activity, suggesting that CBD may exert its protective effects on HG-HL-challenged β-cell survival, at least in part, through the activation of PPARγ. Furthermore, a correlation between PPARγ activation and the downregulation of proinflammatory cytokines, including IL-1β, has been well-established [46]. This suggests that the inhibitory effects of CBD on pyroptosis may be mediated, at least in part, through the activation of PPARγ. It is noteworthy that due to the role of HG-HL-induced CB1R activation in triggering apoptosis and activating Nlrp3 inflammasome [4], CBD may exert some of its inhibitory effects on HG-HL-induced apoptosis and pyroptosis in β-cells through suppressing CB1R.
4.3. The effect of THCV on HG-HL-challenged β-cells
THCV is a natural compound that is structurally similar to THC, but with a propyl group. It is commonly present in dried cannabis plants, albeit in small quantities [39]. THCV has been characterized as neutral antagonist of CB1R, meaning it inhibits the signaling of this receptor. Given its ability to modulate CB1R signaling, THCV has been suggested as a potential therapeutic agent for addressing conditions associated with excessive activation of peripheral CB1R, such as obesity, insulin resistance, diabetes, and their related complications [47].
In this work, we have demonstrated that THCV possesses inhibitory effects on HG-HL-induced β-cells loss, likely achieved by reducing apoptosis and/or pyroptosis. This conclusion is further supported by observing the effects of THCV on Pro- and Cleaved forms of Caspase-7, Caspase-3, PARP, Caspase-1, and Pro-IL-1β.
Since the activation of CB1R by HG-HL promotes apoptosis and pyroptosis in β-cells, the inhibitory effects of THCV on these processes may occur through CB1R suppression. Interestingly, THCV has shown inhibitory effects on TXNIP response in HG-HL-induced β-cells, providing an additional target for THCV to downregulate apoptosis and pyroptosis. Notably, our unpublished reports suggest that THCV upregulates PPARγ in macrophages, presenting another potential target for THCV to downregulate both apoptosis and pyroptosis. THCV has also shown improvement in the HG-HL-induced impaired GSIS in β-cells. The inhibitory effects of THCV on CB1R and subsequent elevation of intracellular Ca2+ partially explain its protective effects on HG-HL-induced impaired GSIS in β-cells (Figure 9). Furthermore, THCV has been found to upregulate MafA, suggesting another target through which it exerts its protective effects on HG-HL-induced impaired GSIS. THCV selectively upregulates the expression of Ins1, not Ins2, potentially mediated by increased levels of PDX-1, and elevated NEUROD1 and MafA mRNA. Apart from its stimulatory impact on NEUROD1, MafA, and PDX-1, THCV downregulates Nanog and Pou5f1 in HG-HL-challenged β-cells, further supporting its protective properties against β-cell dedifferentiation caused by HG-HL (Figure 9).
4.4. The effect of CBC on HG-HL-challenged β-cells
CBC, or cannabichromene, was discovered as the second phytocannabinoid isolated from the cannabis plant following THC. The interaction between CBC and the cannabinoid receptors CB1R and CB2R is characterized by a weak affinity, suggesting that CBC may exert its effects through mechanisms independent of these receptors. In addition to CB1R and CB2R, CBC has been found to target TRPA1 receptors and adenosine receptors [48].
CBC appears to affect cellular endocannabinoid reuptake and suppress the function of monoacylglycerol lipase (MAGL). These additional modes of action contribute to the overall effects of CBC.
Importantly, CBC has demonstrated effectiveness in reducing inflammatory responses in macrophages, which implies its potential therapeutic benefits in diabetes. It has been observed that CBC exerts its anti-inflammatory effects by activating TRPA1 receptors, leading to a decrease in intracellular levels of nitric oxide (NO), interferon-gamma (INFγ), and interleukin-10 (IL-10). Nevertheless, it appears that CB1R-mediated signaling can modulate CBC's activity [49].
Similar to THC, CBD, and THCV, CBC demonstrated the ability to alleviate β-cell loss induced by HG-HL conditions, most likely through the mitigation of apoptosis and/or pyroptosis. This was supported by the response of key biomarkers associated with apoptosis/pyroptosis, such as Pro-Caspase-7, C-Caspase-7, C-Caspase-3, C-PARP, C-Caspase-1, and Pro-IL-1β, to CBC treatment in HG-HL-challenged β-cells (Figure 9).
Similar to THCV, CBC also downregulated the response of TXNIP in HG-HL-challenged β-cells, indicating a potential target for CBC to exert its inhibitory effects on HG-HL-induced apoptosis and pyroptosis.
Furthermore, CBC was found to improve impaired GSIS caused by HG-HL conditions, partially attributed to the upregulation of Slc2A2. However, CBC did not have positive effects on the response of Ins1 and Ins2, possibly due to the negative impacts on MafA, NEUROD1, and PDX-1. In contrast, it downregulated the response of Nanog and Pou5f1, which are genes associated with progenitor cells.
Regulation of TRPA1 may be an additional potential mechanism by which CBC may exert its protective effects on impaired GSIS induced by HG-HL conditions. TRPA1 is a member of the transient receptor potential (TRP) protein superfamily, which consists of cation channels found in various cell types and involved in different signaling pathways. Unlike other TRP channels, TRPA1 is an ion channel located in the cell membrane and exhibits unique functionality. Experimental evidence has confirmed the functional expression of TRPA1 in pancreatic INS-1 β-cells [50]. TRPA1 possesses the capacity to facilitate the transportation of cation ions, particularly calcium ions, into cells and thereby the stimulation of insulin secretion [51].
4.5. The effect of CBN on HG-HL-challenged β-cells
Similar to other phytocannabinoids analyzed in this study, CBN demonstrated the ability to mitigate β-cell loss induced by HG-HL conditions. However, CBN exhibited weaker inhibitory effects on apoptotic and pyroptotic biomarkers, including C-Caspase-3, C-Caspase-7, C-PARP, C-Caspase-1, and Pro-IL-1β, compared to other phytocannabinoids (Figure 9).
CBN, particularly at a concentration of 5 μM, could downregulate the response of TXNIP in HG-HL-induced β-cells, suggesting an additional target through which CBN exerted its inhibitory effects on HG-HL-induced apoptosis and pyroptosis in β-cells. Furthermore, CBN demonstrated a protective impact on impaired GSIS in β-cells exposed to HG-HL conditions, partly attributed to the positive effects of CBN on the expression of Slc2A2.
While CBN increased the PDX-1 protein level, it did not restore the decreased PDX-1 transcript level in HG-HL-induced β-cells. This suggests that CBN may influence PDX-1 protein levels by targeting post-transcriptional mechanisms. CBN treatment showed an inhibitory effects on the expression of NEUROD1 and MafA, which may explain why CBN downregulated Ins1 and Ins2. Similar to CBD and CBC, CBN downregulated the expression of Nanog and Pou5f1 in both HG-HL-challenged β-cells and control groups, suggesting potential protective properties of CBN against HG-HL-induced β-cell dedifferentiation (Figure 9).
5. Conclusions
In our study, we examined the effects of five phytocannabinoids' on HG-HL-induced β-cell loss and dedifferentiation. All compounds showed potential in reducing β-cell loss, likely by inhibiting apoptosis and pyroptosis processes. Four phytocannabinoids, excluding THC, improved impaired GSIS in β-cells. CBD upregulated β-cell-specific biomarkers and downregulated progenitor cell markers the most. THCV enhanced cell viability and GSIS, CBC improved both loss and GSIS, and 10 μM CBC significantly reduced TXNIP levels. While CBN also mitigated β-cell loss and GSIS impairment, it was less effective in reducing apoptotic and pyroptotic markers compared to other phytocannabinoids (Figure 9).
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1: Primers used in the experiments.
Author Contributions
Conceptualization, I.K. and E.G.G.; Methodology, E.G.G., B.W. and D.-P.L.; Validation, I.K., B.W. and O.K; Formal analysis, E.G.G., B.W. and I.K.; Resources, I.K. and O.K.; Data curation, E.G.G. and I.K.; Writing—original draft preparation, E.G.G.; Writing—review and editing, E.G.G., I.K. and O.K.; Visualization, E.G.G.; Supervision, I.K. and O.K.; Project administration, I.K. and O.K.; Funding acquisition, I.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by MITACS, grant number IT28033.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Raw Western blot images were submitted to Cells.
Acknowledgments
We acknowledge the technical support of Rommy Rodriguez-Juarez and Rocio Rodriguez-Juarez.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Holt, R. I.; Cockram, C.; Flyvbjerg, A.; Goldstein, B. J. Textbook of diabetes. John Wiley & Sons: 2017.
- Nagappan, A.; Shin, J.; Jung, M.H. Role of Cannabinoid Receptor Type 1 in Insulin Resistance and Its Biological Implications. Int. J. Mol. Sci. 2019, 20, 2109. [Google Scholar] [CrossRef] [PubMed]
- Back, S.H.; Kaufman, R.J. Endoplasmic Reticulum Stress and Type 2 Diabetes. Annu. Rev. Biochem. 2012, 81, 767–793. [Google Scholar] [CrossRef] [PubMed]
- Ghasemi-Gojani, E.; Kovalchuk, I.; Kovalchuk, O. Cannabinoids and terpenes for diabetes mellitus and its complications: from mechanisms to new therapies. Trends Endocrinol. Metab. 2022, 33, 828–849. [Google Scholar] [CrossRef] [PubMed]
- DeFronzo, R.A.; Eldor, R.; Abdul-Ghani, M. Pathophysiologic Approach to Therapy in Patients With Newly Diagnosed Type 2 Diabetes. Diabetes Care 2013, 36, S127–S138. [Google Scholar] [CrossRef]
- John, A.N.; Morahan, G.; Jiang, F.-X. Incomplete Re-Expression of Neuroendocrine Progenitor/Stem Cell Markers is a Key Feature of β-Cell Dedifferentiation. J. Neuroendocr. 2017, 29. [Google Scholar] [CrossRef]
- Amo-Shiinoki, K.; Tanabe, K.; Hoshii, Y.; Matsui, H.; Harano, R.; Fukuda, T.; Takeuchi, T.; Bouchi, R.; Takagi, T.; Hatanaka, M.; et al. Islet cell dedifferentiation is a pathologic mechanism of long-standing progression of type 2 diabetes. J. Clin. Investig. 2021, 6. [Google Scholar] [CrossRef]
- Khin, P.-P.; Lee, J.-H.; Jun, H.-S. A Brief Review of the Mechanisms of β-Cell Dedifferentiation in Type 2 Diabetes. Nutrients 2021, 13, 1593. [Google Scholar] [CrossRef]
- Moin, A.S.M.; Butler, A.E. Alterations in Beta Cell Identity in Type 1 and Type 2 Diabetes. Curr. Diabetes Rep. 2019, 19, 1–12. [Google Scholar] [CrossRef]
- Efrat, S. Beta-cell dedifferentiation in type 2 diabetes: concise review. Stem Cells 2019, 37, 1267–1272. [Google Scholar] [CrossRef]
- Butler, A. E.; Dhawan, S.; Hoang, J.; Cory, M.; Zeng, K.; Fritsch, H.; Meier, J. J.; Rizza, R. A.; Butler, P. C. β-cell deficit in obese type 2 diabetes, a minor role of β-cell dedifferentiation and degranulation. The Journal of Clinical Endocrinology & Metabolism 2016, 101, 523–532. [Google Scholar]
- Spijker, H.S.; Song, H.; Ellenbroek, J.H.; Roefs, M.M.; Engelse, M.A.; Bos, E.; Koster, A.J.; Rabelink, T.J.; Hansen, B.C.; Clark, A.; et al. Loss of β-Cell Identity Occurs in Type 2 Diabetes and Is Associated with Islet Amyloid Deposits. Diabetes 2015, 64, 2928–2938. [Google Scholar] [CrossRef] [PubMed]
- Pacher, P.; Kogan, N. M.; Mechoulam, R. Beyond THC and endocannabinoids. Annual review of pharmacology and toxicology 2020, 60, 637–659. [Google Scholar] [CrossRef] [PubMed]
- Shin, H.; Han, J.H.; Yoon, J.; Sim, H.J.; Park, T.J.; Yang, S.; Lee, E.K.; Kulkarni, R.N.; Egan, J.M.; Kim, W. Blockade of cannabinoid 1 receptor improves glucose responsiveness in pancreatic beta cells. J. Cell. Mol. Med. 2018, 22, 2337–2345. [Google Scholar] [CrossRef] [PubMed]
- Suryavanshi, S.V.; Zaiachuk, M.; Pryimak, N.; Kovalchuk, I.; Kovalchuk, O. Cannabinoids Alleviate the LPS-Induced Cytokine Storm via Attenuating NLRP3 Inflammasome Signaling and TYK2-Mediated STAT3 Signaling Pathways In Vitro. Cells 2022, 11, 1391. [Google Scholar] [CrossRef]
- Kowalczuk, A.; Marycz, K.; Kornicka, J.; Groborz, S.; Meissner, J.; Mularczyk, M. Tetrahydrocannabivarin (THCV) Protects Adipose-Derived Mesenchymal Stem Cells (ASC) against Endoplasmic Reticulum Stress Development and Reduces Inflammation during Adipogenesis. Int. J. Mol. Sci. 2023, 24, 7120. [Google Scholar] [CrossRef]
- Bergsbaken, T.; Fink, S.L.; Cookson, B.T. Pyroptosis: host cell death and inflammation. Nat. Rev. Genet. 2009, 7, 99–109. [Google Scholar] [CrossRef]
- Zhu, Y.; Liu, Q.; Zhou, Z.; Ikeda, Y. PDX1, Neurogenin-3, and MAFA: critical transcription regulators for beta cell development and regeneration. Stem Cell Res. Ther. 2017, 8, 1–7. [Google Scholar] [CrossRef]
- Roderigo-Milne, H.; Hauge-Evans, A.C.; Persaud, S.J.; Jones, P.M. Differential expression of insulin genes 1 and 2 in MIN6 cells and pseudoislets. Biochem. Biophys. Res. Commun. 2002, 296, 589–595. [Google Scholar] [CrossRef]
- Koranyi, L.; Permutt, M.A.; Chirgwin, J.M.; Giddings, S.J. Proinsulin I and II gene expression in inbred mouse strains. Mol. Endocrinol. 1989, 3. [Google Scholar] [CrossRef]
- Wentworth, B.M.; Rhodes, C.; Schnetzler, B.; Gross, D.J.; Halban, P.A.; Villa-Komaroff, L. The ratio of mouse insulin I:insulin II does not reflect that of the corresponding preproinsulin mRNAs. Mol. Cell. Endocrinol. 1992, 86, 177–186. [Google Scholar] [CrossRef]
- Leroux, L.; Desbois, P.; Lamotte, L.; Duvillié, B.; Cordonnier, N.; Jackerott, M.; Jami, J.; Bucchini, D.; Joshi, R. L. Compensatory responses in mice carrying a null mutation for Ins1 or Ins2. Diabetes 2001, 50 (Suppl. S1), S150. [Google Scholar] [CrossRef] [PubMed]
- Naya, F.J.; Stellrecht, C.M.; Tsai, M.J. Tissue-specific regulation of the insulin gene by a novel basic helix-loop-helix transcription factor. Minerva Anestesiol. 1995, 9, 1009–1019. [Google Scholar] [CrossRef] [PubMed]
- Romer, A.I.; Singer, R.A.; Sui, L.; Egli, D.; Sussel, L. Murine Perinatal β-Cell Proliferation and the Differentiation of Human Stem Cell–Derived Insulin-Expressing Cells Require NEUROD1. Diabetes 2019, 68, 2259–2271. [Google Scholar] [CrossRef] [PubMed]
- Rubio-Cabezas, O.; Minton, J.A.; Kantor, I.; Williams, D.; Ellard, S.; Hattersley, A.T. Homozygous Mutations in NEUROD1 Are Responsible for a Novel Syndrome of Permanent Neonatal Diabetes and Neurological Abnormalities. Diabetes 2010, 59, 2326–2331. [Google Scholar] [CrossRef]
- Malecki, M.T.; Jhala, U.S.; Antonellis, A.; Fields, L.; Doria, A.; Orban, T.; Saad, M.; Warram, J.H.; Montminy, M.; Krolewski, A.S. Mutations in NEUROD1 are associated with the development of type 2 diabetes mellitus. Nat. Genet. 1999, 23, 323–328. [Google Scholar] [CrossRef]
- Ono, Y.; Kataoka, K. MafA, NeuroD1, and HNF1β synergistically activate the Slc2a2 (Glut2) gene in β-cells. Journal of Molecular Endocrinology 2021, 67, 71–82. [Google Scholar] [CrossRef]
- Nishimura, W.; Kondo, T.; Salameh, T.; El Khattabi, I.; Dodge, R.; Bonner-Weir, S.; Sharma, A. A switch from MafB to MafA expression accompanies differentiation to pancreatic β-cells. Dev. Biol. 2006, 293, 526–539. [Google Scholar] [CrossRef]
- Nishimura, W.; Bonner-Weir, S.; Sharma, A. Expression of MafA in pancreatic progenitors is detrimental for pancreatic development. Dev. Biol. 2009, 333, 108–120. [Google Scholar] [CrossRef]
- Kaneto, H.; Matsuoka, T.-A.; Nakatani, Y.; Miyatsuka, T.; Matsuhisa, M.; Hori, M.; Yamasaki, Y. A Crucial Role of MafA as a Novel Therapeutic Target for Diabetes. J. Biol. Chem. 2005, 280, 15047–15052. [Google Scholar] [CrossRef]
- Zhou, Q.; Brown, J.; Kanarek, A.; Rajagopal, J.; Melton, D.A. In vivo reprogramming of adult pancreatic exocrine cells to β-cells. Nature 2008, 455, 627–632. [Google Scholar] [CrossRef]
- Butler, A.E.; Robertson, R.P.; Hernandez, R.; Matveyenko, A.V.; Gurlo, T.; Butler, P.C. Beta cell nuclear musculoaponeurotic fibrosarcoma oncogene family A (MafA) is deficient in type 2 diabetes. Diabetologia 2012, 55, 2985–2988. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Dai, C.; Guo, M.; Taylor, B.; Harmon, J.S.; Sander, M.; Robertson, R.P.; Powers, A.C.; Stein, R. Inactivation of specific β cell transcription factors in type 2 diabetes. J. Clin. Investig. 2013, 123, 3305–3316. [Google Scholar] [CrossRef] [PubMed]
- Kovalchuk, I.; Pellino, M.; Rigault, P.; Van Velzen, R.; Ebersbach, J.; Ashnest, J.; Mau, M.; Schranz, M.; Alcorn, J.; Laprairie, R. The genomics of Cannabis and its close relatives. Annual Review of Plant Biology 2020, 71, 713–739. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; E Kaminski, N.; Fischer, L.J. Examination of the immunosuppressive effect of Δ9-tetrahydrocannabinol in streptozotocin-induced autoimmune diabetes. Int. Immunopharmacol. 2001, 1, 699–712. [Google Scholar] [CrossRef]
- Coskun, Z.M.; Bolkent, S. Evaluation of Δ(9)-tetrahydrocannabinol metabolites and oxidative stress in type 2 diabetic rats. Iran J Basic Med Sci 2016, 19, 154–8. [Google Scholar]
- Altınok, A.; Coşkun, Z.; Karaoğlu, K.; Bolkent, S.; Akkan, A.; Özyazgan, S. Δ9-tetrahydrocannabinol treatment improved endothelium-dependent relaxation on streptozotocin/nicotinamide-induced diabetic rat aorta. Acta Physiol. Hung. 2015, 102, 51–59. [Google Scholar] [CrossRef]
- Pertwee, R. The diverse CB1 and CB2 receptor pharmacology of three plant cannabinoids: Δ9-tetrahydrocannabinol, cannabidiol and Δ9-tetrahydrocannabivarin. British journal of pharmacology 2008, 153, (2), 199–215. [Google Scholar] [CrossRef]
- Russo, E.B. Taming THC: potential cannabis synergy and phytocannabinoid-terpenoid entourage effects. Br. J. Pharmacol. 2011, 163, 1344–1364. [Google Scholar] [CrossRef]
- Jiang, N.; Liu, J.; Guan, C.; Ma, C.; An, J.; Tang, X. Thioredoxin-interacting protein: A new therapeutic target in bone metabolism disorders? Frontiers in Immunology 2022, 13, 955128. [Google Scholar] [CrossRef]
- Fu, Z.; R Gilbert, E.; Liu, D. Regulation of insulin synthesis and secretion and pancreatic Beta-cell dysfunction in diabetes. Current diabetes reviews 2013, 9, (1), 25–53. [Google Scholar] [CrossRef]
- Iype, T.; Francis, J.; Garmey, J. C.; Schisler, J. C.; Nesher, R.; Weir, G. C.; Becker, T. C.; Newgard, C. B.; Griffen, S. C.; Mirmira, R. G. Mechanism of insulin gene regulation by the pancreatic transcription factor Pdx-1: application of pre-mRNA analysis and chromatin immunoprecipitation to assess formation of functional transcriptional complexes. Journal of Biological Chemistry 2005, 280, (17), 16798–16807. [Google Scholar] [CrossRef]
- Zhao, L.; Guo, M.; Matsuoka, T.-A.; Hagman, D.K.; Parazzoli, S.D.; Poitout, V.; Stein, R. The Islet β Cell-enriched MafA Activator Is a Key Regulator of Insulin Gene Transcription. J. Biol. Chem. 2005, 280, 11887–11894. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Moore, M.; Marcora, E.; Lee, J. E.; Qiu, Y.; Samaras, S.; Stein, R. The NeuroD1/BETA2 sequences essential for insulin gene transcription colocalize with those necessary for neurogenesis and p300/CREB binding protein binding. Molecular and cellular biology 1999, 19, (1), 704–713. [Google Scholar] [CrossRef]
- Welters, H.; Meyer, J.; Smith, D. M.; Kulkarni, R. N. In Role of PPAR gamma in Pancreatic beta-Cell Proliferation, DIABETES, 2009; AMER DIABETES ASSOC 1701 N BEAUREGARD ST, ALEXANDRIA, VA 22311-1717 USA: pp A412-A412.
- Olefsky, J. M.; Glass, C. K. Macrophages, inflammation, and insulin resistance. Annual review of physiology 2010, 72, 219–246. [Google Scholar] [CrossRef]
- Rock, E. M.; Sticht, M. A.; Duncan, M.; Stott, C.; Parker, L. A. Evaluation of the potential of the phytocannabinoids, cannabidivarin (CBDV) and Δ9-tetrahydrocannabivarin (THCV), to produce CB1 receptor inverse agonism symptoms of nausea in rats. British journal of pharmacology 2013, 170, (3), 671–678. [Google Scholar] [CrossRef]
- nbsp, *!!! REPLACE !!!*; Pollastro, F.; Minassi, A.; Fresu, L.G. Cannabis Phenolics and their Bioactivities. Curr. Med. Chem. 2018, 25, 1160–1185. [Google Scholar] [CrossRef]
- Romano, B.; Borrelli, F.; Fasolino, I.; Capasso, R.; Piscitelli, F.; Cascio, M.; Pertwee, R.; Coppola, D.; Vassallo, L.; Orlando, P.; et al. The cannabinoid TRPA1 agonist cannabichromene inhibits nitric oxide production in macrophages and ameliorates murine colitis. Br. J. Pharmacol. 2013, 169, 213–229. [Google Scholar] [CrossRef] [PubMed]
- Cao, D.-S.; Zhong, L.; Hsieh, T.-H.; Abooj, M.; Bishnoi, M.; Hughes, L.; Premkumar, L.S. Expression of Transient Receptor Potential Ankyrin 1 (TRPA1) and Its Role in Insulin Release from Rat Pancreatic Beta Cells. PLOS ONE 2012, 7, e38005. [Google Scholar] [CrossRef]
- Wei, Y.; Cai, J.; Zhu, R.; Xu, K.; Li, H.; Li, J. Function and therapeutic potential of transient receptor potential ankyrin 1 in fibrosis. Front. Pharmacol. 2022, 13, 1014041. [Google Scholar] [CrossRef]
Figure 1.
The protective effect of THC, THCV, CBD, CBC, and CBN on the HG-HL-induced β-cell loss. All five phytocannabinoids demonstrated an increase in cell viability of HG-HL-induced β-cells. The provided data represents the mean value with SD, n=3. The abbreviations used in the figure are as follows: Ct+Meth (Control+Methanol) and HGHL (High Glucose+High Lipid). The asterisks show significant difference, four – p<0.0001; ns – non-significant.
Figure 1.
The protective effect of THC, THCV, CBD, CBC, and CBN on the HG-HL-induced β-cell loss. All five phytocannabinoids demonstrated an increase in cell viability of HG-HL-induced β-cells. The provided data represents the mean value with SD, n=3. The abbreviations used in the figure are as follows: Ct+Meth (Control+Methanol) and HGHL (High Glucose+High Lipid). The asterisks show significant difference, four – p<0.0001; ns – non-significant.
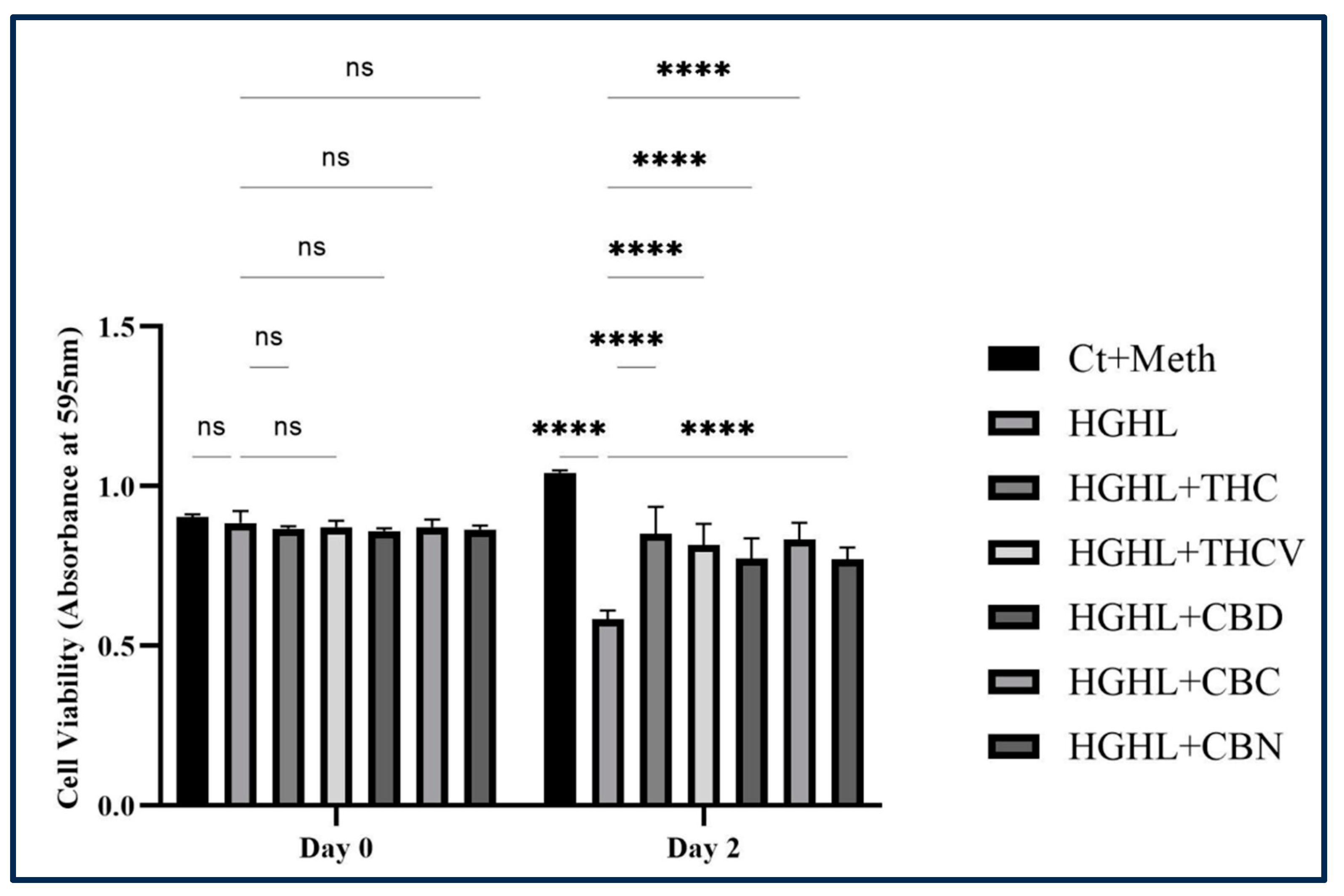
Figure 2.
The impact of THC, THCV, CBD, CBC, and CBN on GSIS in HG-HL-induced β-cells. Cells were pre-treated with 5 μM phytocannabinoids or vehicle for 2 h, followed by incubation with 25 mM glucose and 400 μM palmitic acid for 48 hours. Glucose-stimulated insulin secretion (GSIS) was then evaluated using phytocannabinoids in the HBSS solution. The provided data represents the mean value with SD, n=3 measurements. The abbreviations used in the figure are as follows: Ct+Meth (Control+Methanol) and HGHL (High Glucose+High Lipid). The asterisks show significant difference, where two – p<0.01, three – p<0.001, four – p<0.0001.
Figure 2.
The impact of THC, THCV, CBD, CBC, and CBN on GSIS in HG-HL-induced β-cells. Cells were pre-treated with 5 μM phytocannabinoids or vehicle for 2 h, followed by incubation with 25 mM glucose and 400 μM palmitic acid for 48 hours. Glucose-stimulated insulin secretion (GSIS) was then evaluated using phytocannabinoids in the HBSS solution. The provided data represents the mean value with SD, n=3 measurements. The abbreviations used in the figure are as follows: Ct+Meth (Control+Methanol) and HGHL (High Glucose+High Lipid). The asterisks show significant difference, where two – p<0.01, three – p<0.001, four – p<0.0001.
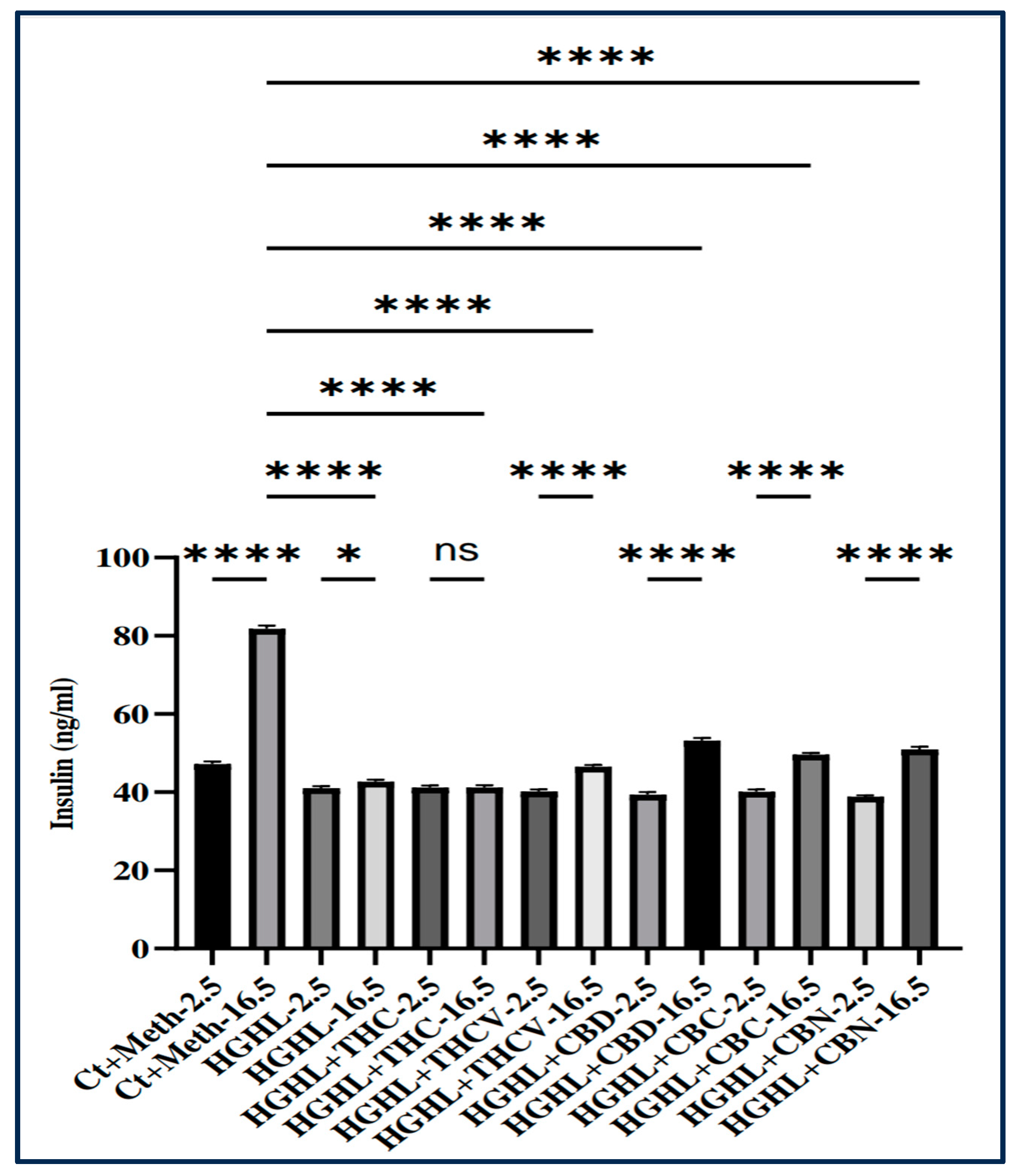
Figure 3.
Analysis of the level of proteins involved in apoptosis and pyroptosis in HG-HL-challenged β-cells in response to all five phytocannabinoids. a) The response of Pro-Caspase-7 to THC, THCV, CBD, CBC, and CBN in HG-HL-challenged β-cells. b) The response of C-Caspase-7 to THC, THCV, CBD, CBC, and CBN in HG-HL-challenged β-cells. c) The impact of 5 μM of each phytocannabinoid on the level of C-Caspase 3 (17 KDa) in HG-HL-stimulated β-cells. d) Western blot analysis of C-Caspase-1, with actin used for normalization, in response to all five phytocannabinoids in HG-HL-challenged β-cells. e) The response of pro-IL-1β to the phytocannabinoids in HG-HL-challenged β-cells. f) The level of C-PARP in HG-HL-challenged β-cells treated with the phytocannabinoids. Panels under graphics show the representative images of Western blots. The provided data represents the mean value with SD, n=3 measurements. The abbreviations used in the figure are as follows: Ct+Meth (Control+Methanol) and HG-HL (High Glucose+High Lipid). The asterisks show significant difference, where one – p<0.05, two – p<0.01, three – p<0.001, four - p<0.0001, ns – non-significant.
Figure 3.
Analysis of the level of proteins involved in apoptosis and pyroptosis in HG-HL-challenged β-cells in response to all five phytocannabinoids. a) The response of Pro-Caspase-7 to THC, THCV, CBD, CBC, and CBN in HG-HL-challenged β-cells. b) The response of C-Caspase-7 to THC, THCV, CBD, CBC, and CBN in HG-HL-challenged β-cells. c) The impact of 5 μM of each phytocannabinoid on the level of C-Caspase 3 (17 KDa) in HG-HL-stimulated β-cells. d) Western blot analysis of C-Caspase-1, with actin used for normalization, in response to all five phytocannabinoids in HG-HL-challenged β-cells. e) The response of pro-IL-1β to the phytocannabinoids in HG-HL-challenged β-cells. f) The level of C-PARP in HG-HL-challenged β-cells treated with the phytocannabinoids. Panels under graphics show the representative images of Western blots. The provided data represents the mean value with SD, n=3 measurements. The abbreviations used in the figure are as follows: Ct+Meth (Control+Methanol) and HG-HL (High Glucose+High Lipid). The asterisks show significant difference, where one – p<0.05, two – p<0.01, three – p<0.001, four - p<0.0001, ns – non-significant.
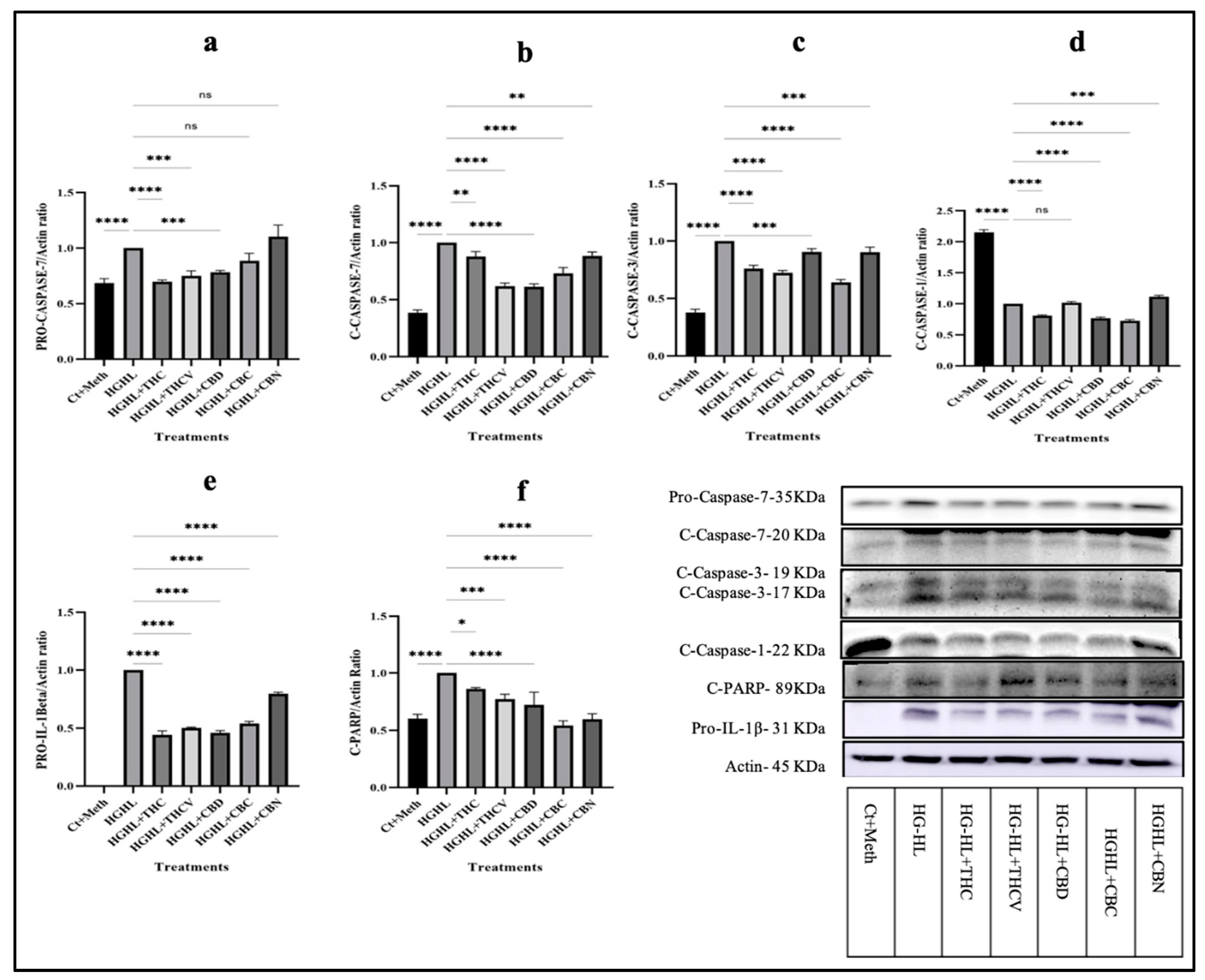
Figure 4.
qPCR and western blot analyses of key genes/proteins involved in β-cells dedifferentiation in response to THC in HG-HL-induced β-cells. a) Western blot analysis of TXNIP (a1), PDX-1 (a2), and forkhead box protein O1 (FOXO1) (a3), their expression levels were normalized to actin level in HG-HL-challenged β-cells. Panels under graphics show the representative images of Western blots. b) The normalized mRNA expression levels of Ins1, Ins2, PDX-1, FOXO1, NEUROD1, MafA, Slc2A2, Nanog, and Pou5f1 in HG-HL-challenged β-cells treated with 5 µM THC. The provided data represents the mean value with SD, n=3 measurements/replications. The abbreviations used in the figure are as follows: Ct+Meth (Control+Methanol) and HGHL (High Glucose+High Lipid). The asterisks show significant difference, where one – p<0.05, two – p<0.01, three – p<0.001, four - p<0.0001, ns – non-significant.
Figure 4.
qPCR and western blot analyses of key genes/proteins involved in β-cells dedifferentiation in response to THC in HG-HL-induced β-cells. a) Western blot analysis of TXNIP (a1), PDX-1 (a2), and forkhead box protein O1 (FOXO1) (a3), their expression levels were normalized to actin level in HG-HL-challenged β-cells. Panels under graphics show the representative images of Western blots. b) The normalized mRNA expression levels of Ins1, Ins2, PDX-1, FOXO1, NEUROD1, MafA, Slc2A2, Nanog, and Pou5f1 in HG-HL-challenged β-cells treated with 5 µM THC. The provided data represents the mean value with SD, n=3 measurements/replications. The abbreviations used in the figure are as follows: Ct+Meth (Control+Methanol) and HGHL (High Glucose+High Lipid). The asterisks show significant difference, where one – p<0.05, two – p<0.01, three – p<0.001, four - p<0.0001, ns – non-significant.
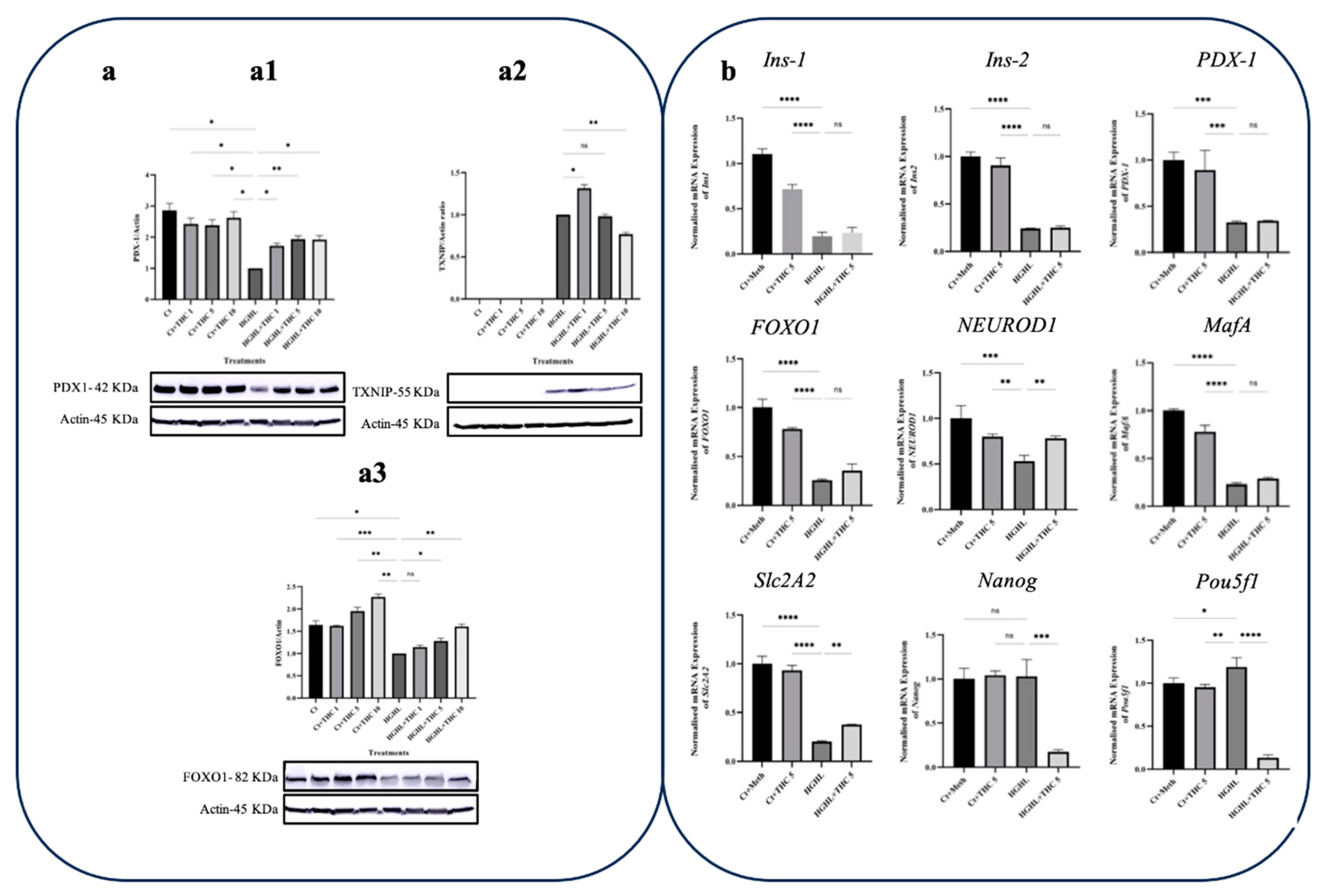
Figure 5.
The effect of CBD on the levels of key genes and proteins involved in β-cell dedifferentiation in HG-HL challenged β-cells. a) Western blot analysis of TXNIP (a1), PDX-1 (a2), and FOXO1 (a3), their expression levels were normalized to actin level in HG-HL-challenged β-cells. Panels under graphics show the representative images of Western blots. b) The normalized mRNA expression levels of Ins1, Ins2, PDX-1, FOXO1, NEUROD1, MafA, Slc2A2, Nanog, and Pou5f1 in HG-HL-challenged β-cells treated with 5 µM CBD. The provided data represents the mean value with SD, n=3 measurements/replications. The abbreviations used in the figure are as follows: Ct+Meth (Control+Methanol) and HGHL (High Glucose+High Lipid). The asterisks show significant difference, where one – p<0.05, two – p<0.01, three – p<0.001, four - p<0.0001, ns – non-significant.
Figure 5.
The effect of CBD on the levels of key genes and proteins involved in β-cell dedifferentiation in HG-HL challenged β-cells. a) Western blot analysis of TXNIP (a1), PDX-1 (a2), and FOXO1 (a3), their expression levels were normalized to actin level in HG-HL-challenged β-cells. Panels under graphics show the representative images of Western blots. b) The normalized mRNA expression levels of Ins1, Ins2, PDX-1, FOXO1, NEUROD1, MafA, Slc2A2, Nanog, and Pou5f1 in HG-HL-challenged β-cells treated with 5 µM CBD. The provided data represents the mean value with SD, n=3 measurements/replications. The abbreviations used in the figure are as follows: Ct+Meth (Control+Methanol) and HGHL (High Glucose+High Lipid). The asterisks show significant difference, where one – p<0.05, two – p<0.01, three – p<0.001, four - p<0.0001, ns – non-significant.
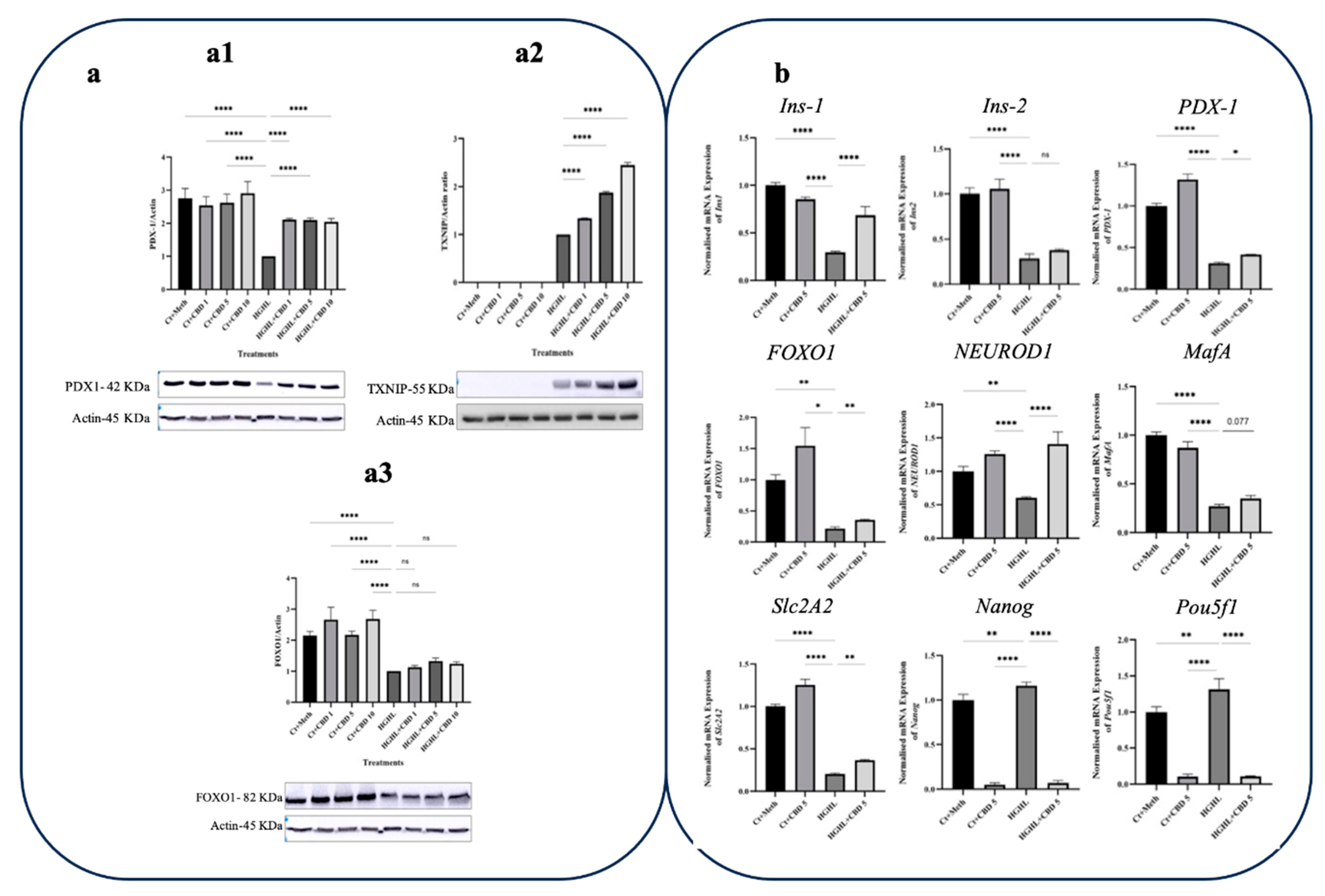
Figure 6.
qPCR and western blot analyses of transcripts/proteins associated with β-cells dedifferentiation in response to THCV. a) Western blot analysis of TXNIP (a1), PDX-1 (a2) and FOXO1 (a3), their expression levels were normalized to actin level in HG-HL-challenged β-cells. Panels under graphics show the representative images of Western blots. b) The normalized mRNA expression levels of Ins1, Ins2, PDX-1, FOXO1, NEUROD1, MafA, Slc2A2, Nanog, and Pou5f1 in HG-HL-challenged β-cells treated with 5 µM THCV. The provided data represents the mean value with SD, n=3 measurements/replications. The abbreviations used in the figure are as follows: Ct+Meth (Control+Methanol) and HGHL (High Glucose+High Lipid). The asterisks show significant difference, where one – p<0.05, two – p<0.01, three – p<0.001, four - p<0.0001, ns – non-significant.
Figure 6.
qPCR and western blot analyses of transcripts/proteins associated with β-cells dedifferentiation in response to THCV. a) Western blot analysis of TXNIP (a1), PDX-1 (a2) and FOXO1 (a3), their expression levels were normalized to actin level in HG-HL-challenged β-cells. Panels under graphics show the representative images of Western blots. b) The normalized mRNA expression levels of Ins1, Ins2, PDX-1, FOXO1, NEUROD1, MafA, Slc2A2, Nanog, and Pou5f1 in HG-HL-challenged β-cells treated with 5 µM THCV. The provided data represents the mean value with SD, n=3 measurements/replications. The abbreviations used in the figure are as follows: Ct+Meth (Control+Methanol) and HGHL (High Glucose+High Lipid). The asterisks show significant difference, where one – p<0.05, two – p<0.01, three – p<0.001, four - p<0.0001, ns – non-significant.
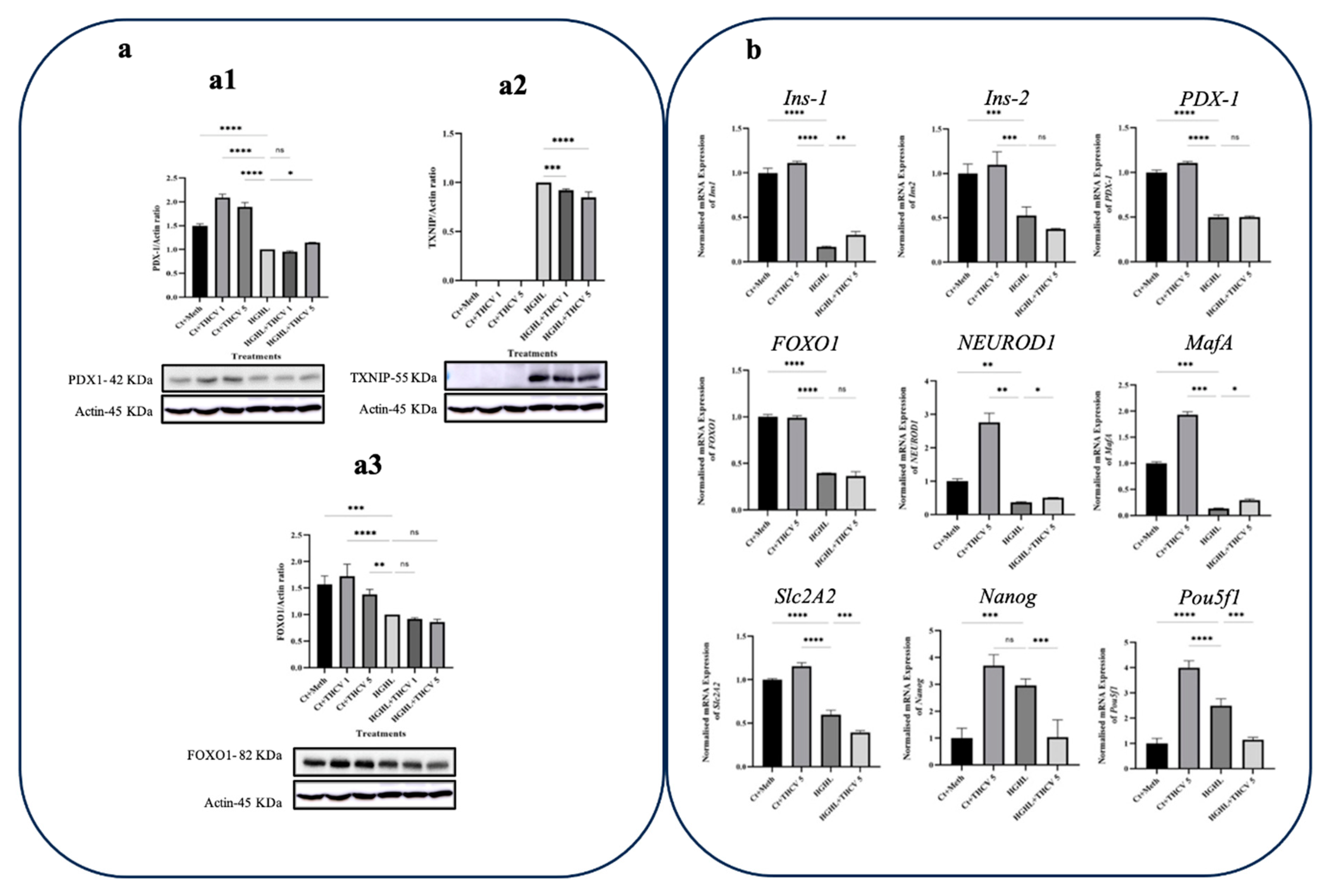
Figure 7.
The influence of CBC on the expression of critical transcripts and Proteins in β-Cell dedifferentiation under HG-HL conditions. a) Western blot analysis of TXNIP (a1), PDX-1 (a2), and FOXO1 (a3); their expression levels were normalized to actin level in HG-HL-challenged β-cells. Panels under graphics show the representative images of Western blots. b) The normalized mRNA expression levels of Ins1, Ins2, PDX-1, FOXO1, NEUROD1, MafA, Slc2A2, Nanog, and Pou5f1 in HG-HL-challenged β-cells treated with 5 µM CBC. Data represents the mean value with SD, n=3 measurements/replications. The abbreviations used in the figure are as follows: Ct+Meth (Control+Methanol) and HGHL (High Glucose+High Lipid). The asterisks show significant difference, where one – p<0.05, two – p<0.01, three – p<0.001, four - p<0.0001, ns – non-significant.
Figure 7.
The influence of CBC on the expression of critical transcripts and Proteins in β-Cell dedifferentiation under HG-HL conditions. a) Western blot analysis of TXNIP (a1), PDX-1 (a2), and FOXO1 (a3); their expression levels were normalized to actin level in HG-HL-challenged β-cells. Panels under graphics show the representative images of Western blots. b) The normalized mRNA expression levels of Ins1, Ins2, PDX-1, FOXO1, NEUROD1, MafA, Slc2A2, Nanog, and Pou5f1 in HG-HL-challenged β-cells treated with 5 µM CBC. Data represents the mean value with SD, n=3 measurements/replications. The abbreviations used in the figure are as follows: Ct+Meth (Control+Methanol) and HGHL (High Glucose+High Lipid). The asterisks show significant difference, where one – p<0.05, two – p<0.01, three – p<0.001, four - p<0.0001, ns – non-significant.
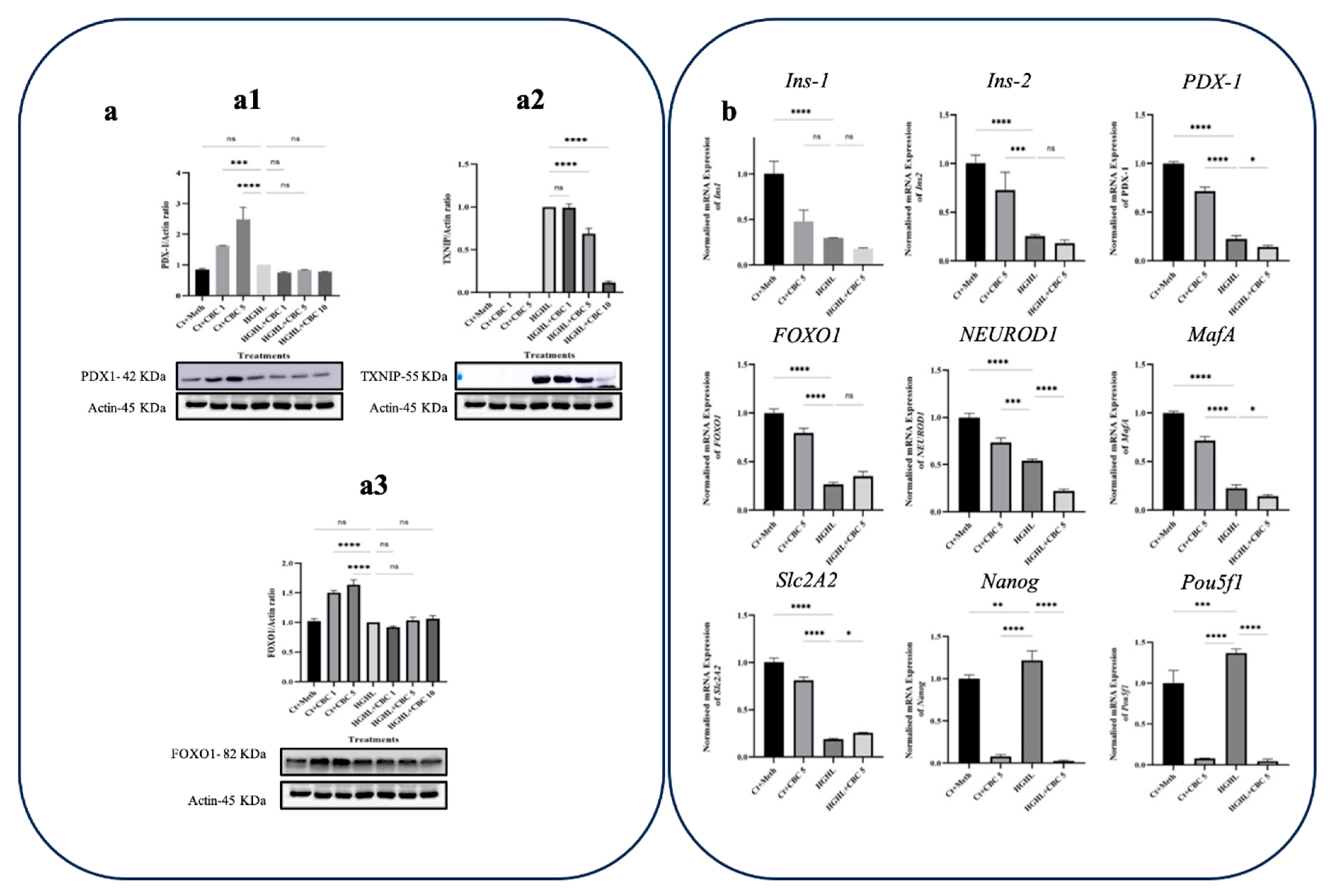
Figure 8.
qPCR and western blot analyses of transcripts and proteins associated with β-cell dedifferentiation in response to CBN. a) Western blot analysis of TXNIP (a1), PDX-1 (a2), and FOXO1 (a3), their expression levels were normalized to actin level in HG-HL-challenged β-cells. Panels under graphics show the representative images of Western blots. b) The normalized mRNA expression levels of Ins1, Ins2, PDX-1, FOXO1, NEUROD1, MafA, Slc2A2, Nanog, and Pou5f1 in HG-HL-challenged β-cells treated with 5 µM CBN. The provided data represents the mean value with SD, n=3 measurements/replications. The abbreviations used in the figure are as follows: Ct+Meth (Control+Methanol) and HGHL (High Glucose+High Lipid). The asterisks show significant difference, where one – p<0.05, two – p<0.01, three – p<0.001, four - p<0.0001, ns – non-significant.
Figure 8.
qPCR and western blot analyses of transcripts and proteins associated with β-cell dedifferentiation in response to CBN. a) Western blot analysis of TXNIP (a1), PDX-1 (a2), and FOXO1 (a3), their expression levels were normalized to actin level in HG-HL-challenged β-cells. Panels under graphics show the representative images of Western blots. b) The normalized mRNA expression levels of Ins1, Ins2, PDX-1, FOXO1, NEUROD1, MafA, Slc2A2, Nanog, and Pou5f1 in HG-HL-challenged β-cells treated with 5 µM CBN. The provided data represents the mean value with SD, n=3 measurements/replications. The abbreviations used in the figure are as follows: Ct+Meth (Control+Methanol) and HGHL (High Glucose+High Lipid). The asterisks show significant difference, where one – p<0.05, two – p<0.01, three – p<0.001, four - p<0.0001, ns – non-significant.
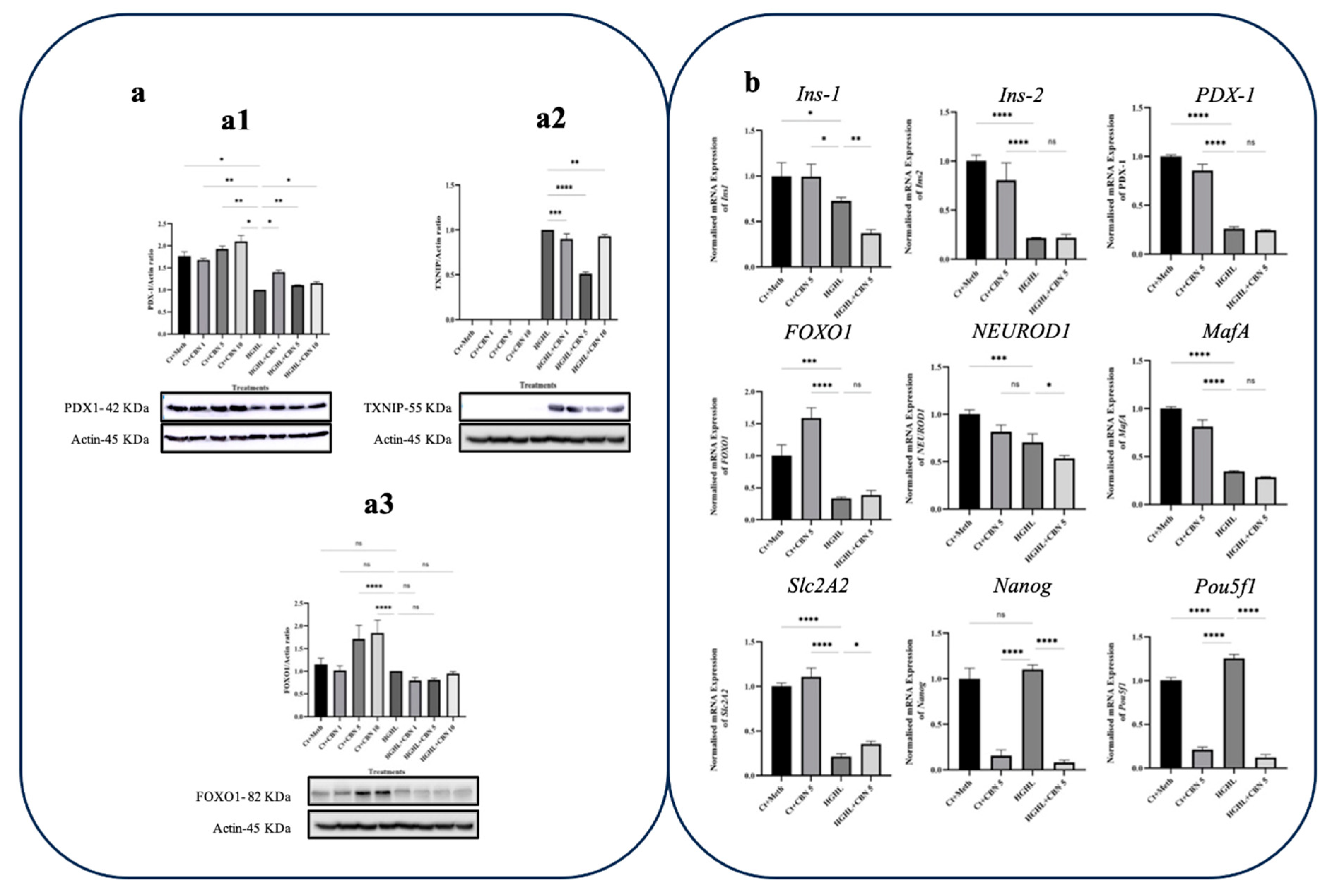
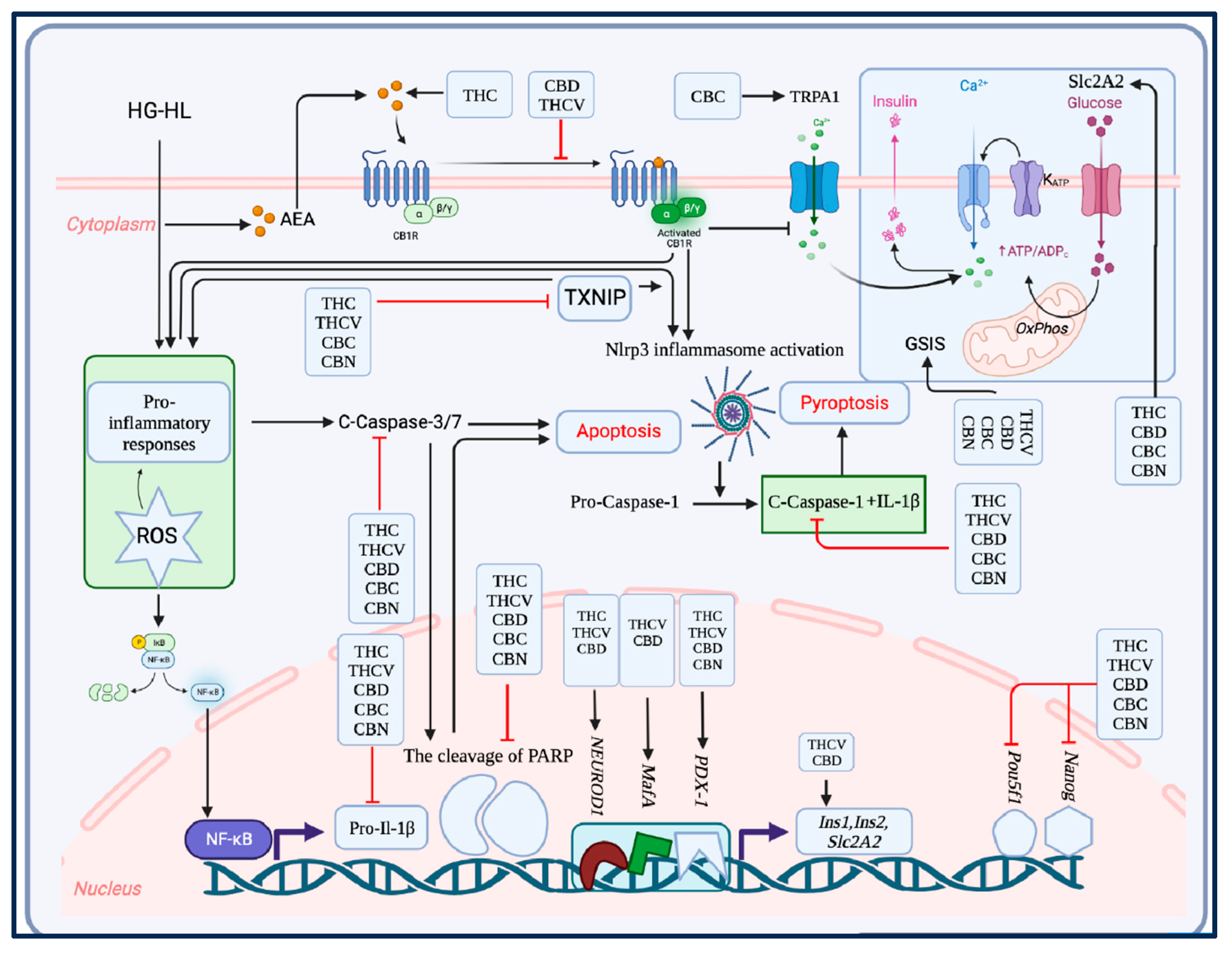
Figure 9.
The key findings of the study. In this study, all five phytocannabinoids examined exhibited the capacity to protect β-cells from the detrimental effects of HG-HL conditions, preventing their loss and dedifferentiation. All of the phytocannabinoids demonstrated a potential in mitigating the loss of β-cells induced by HG-HL, potentially by reducing apoptosis and pyroptosis processes. Like THCV, CBC and CBN, CBD showed a pronounced improvement in impaired GSIS in β-cells under HG-HL conditions, which may be attributed to the upregulation of MafA and Slc2A2. Furthermore, CBD upregulated the expression of most β-cell-specific biomarkers while downregulating progenitor cell biomarkers, implying its potential in the mitigation of HG-HL-induced β-cell dedifferentiation. THCV not only enhanced the viability of β-cells under HG-HL conditions but also improved impaired GSIS, similar to CBD, by upregulating MafA and Slc2A2. Likewise, CBC rescued both β-cell loss and impaired GSIS induced by HG-HL. Notably, THC, THCV, CBC, and CBN significantly reduced TXNIP levels in HG-HL-challenged β-cells, indicating an additional potential target through which these phytocannabinoids could regulate HG-HL-induced reactive oxygen species (ROS), apoptosis, and pyroptosis. Although CBN also mitigated β-cell loss induced by HG-HL and impaired GSIS, it was comparatively less effective than other phytocannabinoids in reducing levels of apoptotic and pyroptotic biomarkers, as demonstrated by our results.
Figure 9.
The key findings of the study. In this study, all five phytocannabinoids examined exhibited the capacity to protect β-cells from the detrimental effects of HG-HL conditions, preventing their loss and dedifferentiation. All of the phytocannabinoids demonstrated a potential in mitigating the loss of β-cells induced by HG-HL, potentially by reducing apoptosis and pyroptosis processes. Like THCV, CBC and CBN, CBD showed a pronounced improvement in impaired GSIS in β-cells under HG-HL conditions, which may be attributed to the upregulation of MafA and Slc2A2. Furthermore, CBD upregulated the expression of most β-cell-specific biomarkers while downregulating progenitor cell biomarkers, implying its potential in the mitigation of HG-HL-induced β-cell dedifferentiation. THCV not only enhanced the viability of β-cells under HG-HL conditions but also improved impaired GSIS, similar to CBD, by upregulating MafA and Slc2A2. Likewise, CBC rescued both β-cell loss and impaired GSIS induced by HG-HL. Notably, THC, THCV, CBC, and CBN significantly reduced TXNIP levels in HG-HL-challenged β-cells, indicating an additional potential target through which these phytocannabinoids could regulate HG-HL-induced reactive oxygen species (ROS), apoptosis, and pyroptosis. Although CBN also mitigated β-cell loss induced by HG-HL and impaired GSIS, it was comparatively less effective than other phytocannabinoids in reducing levels of apoptotic and pyroptotic biomarkers, as demonstrated by our results.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



