
Submitted:
14 September 2023
Posted:
18 September 2023
Read the latest preprint version here
Abstract
The objective of the this study was to reduce the benzo[a]pyrene-induced hepatotoxicity method in herbal medicine products and to give proof that neferine, daidzein, genistein possess antihepa-totoxic effects. B[a]P which classified as a group 1 carcinogen and metabolized to B[a]P-7,8-dihydrodiol-9,10-epoxide (BPDE) causes mutagenic DNA addition products. The reduc-tion of BPDE-DNA adduct formation by B[a]P-7,8-dihydrodiol-9,10-epoxide (BPDE) was derived by benzo[a]pyrene (B[a]P). In HepG2 cells, B[a]P exhibited toxicity and substance treatment of the cells with B[a]P with neferine in lotus and daidzein, genistein in soybean reduced the BPDE-DNA ad-ducts level. The level of B[a]P-metabolites in the substance treatment of the cells was presented that BPDE levels were reduced by neferine in lotus and daidzein, genisten in soybean. These results suggest that neferine in lotus and daidzein, genistein in soybean prevent B[a]P-induced hepato-toxicity for BPDE-DNA adduct formation.
Keywords:
herbal medicine products
; benzo[a]pyrene-induced hepatotoxicity
; B[a]P-7
; 8-dihydrodiol-9
; 10-epoxide (BPDE)
; benzo[a]pyrene (B[a]P)
; HepG2 cells
1. Introduction
Benzo[a]pyrene (B[a]P) was one of the kind of polycyclic aromatic hydrocarbon and have high molecular weight in a type of potent and ubiquitous atmospheric contaminant [1]. B[a]P was generated from incomplete combustion of the organic substances such as some foods, tobacco smoke, coal tar and automobile exhaust fumes [2]. Carcinogenic, mutagenic, cytotoxic properties of B[a]P was represented in recent years [3]. Inhalation of B[a]P causes various types of adverse health effects such as mutations and immunocompromised status. It damages to the cardiopulmonary and reproductive system [4]. Accordingly, the National Institute for Occupational Safety and Health (NIOSH) nominated guidelines for public safety that a worker be exposed to PAHs at a level of <100 μg/m3 in a workplace [5]. It was shown that the lipophilicity of PAHs keep them with low clearance and high residence in organs such as brain, lung and liver [6]. After inhalation, B[a]P was metabolized to B[a]P-7,8-dihydrodiol-9,10-epoxide (BPDE) [7]. The synthesis of reactive oxygen species (ROS) in the liver related with their hepatoxicity in terms of B[a]P-induced hepatotoxicity [8]. In this respect, neferine, daidzein and genistein were the substances which were analyzed to confirm the benzo[a]pyrene-induced hepatotoxicity in this study were constituents of natural herbal medicines such as lotus seeds (Nelumbo nucifera) and soybeans (Glycine max). Neferine is a bisbenzylisoquinoline alkaloid isolated from lotus seed sample and had a wide range of pharmacological activities [9]. Neferine which contained in the lotus seeds acted as a free radical scavenger which inhibited the formation of LPO due to the hydroxyl groups and enhanced the activity of superoxide dismutase present in the structure [10]. Lotus seeds were widely used in herbal medicines to treat skin diseases, diuretics, cancer, tissue inflammation, and antidote to poison [11]. Embryos of lotus seeds were used in the treatment of cardiovascular diseases, high fevers, insomnia, and nervous disorders [12]. Poornima et al reported that neferine was correlated with the possible pathway in HepG2 cells [13]. Daidzein and genistein which contained in the soybeans were the major isoflavones and were associated with beneficial properties on human health [14]. Soybeans were associated with a reduction of osteoporosis, cardiovascular disease, colon cancer, prostate cancer and breast cancer [15]. In this regard, daidzein and genistein which contained in the soybeans have antiosteoporosis, anticarcinogenic and antioxidant activities [16]. Sarao et al reported that isoflavones such as daidzein and genistein in soybeans correlated with the possible pathway in HepG2 cells [17]. However, the effects of neferine, daidzein, genistein on the reduction of benzo[a]pyrene-induced hepatotoxicity method in HepG2 cells not yet been represented. Previous studies represented the considerable efforts was made to relate the hepatotoxicity of PAHs [9]. It was analyzed through biochemical and histopathological approaches [18]. In addition, the intraperitoneal and oral administration of PAHs could affect the weight of liver [19]. Also, it cause the preneoplastic hepatocytes and synthesis of hepatic enzymes as well as liver congestion [20]. This was followed by other studies about the use of dietary agents such as polyphenols, terpenoids, flavonoids and natural herbal medicines in order to prevent the occurrence of various types of environment-induced diseases [21]. Therefore, the objective of this study was to conduct the antioxidative effects of natural herbal medicines in inhibiting the B[a]P-induced hepatotoxicity.
2. Materials and Methods
2.1. Chemicals and Materials
B[a]P, neferine, daidzein, genistein were acquired from Sigma-Aldrich Chemical (St. Louis, MO, USA). Trypsin-ethylenediaminetetraacetic acid (T-EDTA), penicillin/streptomycin, fetal bovine serum (FBS), sodium pyruvate and phosphate buffered saline (PBS), Minimum essential medium (MEM) were acquired from Gibco (Seoul, Korea). Dimethyl sulfoxide (DMSO) was acquired from VWR (Suwon, Korea). HPLC grade solvents (methanol and water) were purchased from J.T. Baker (USA). Trifluoroacetic acid (TFA) was purchased from DAEJUNG (Gyeonggi-do, Korea). Acetonitrile and ethyl acetate was obtained from Burdick & Jackson (Muskegon, MI, USA). EZ-CYTOX reagent was acquired from DOGEN (Daejeon, Korea). The lid (all plate type, 84.50 × 126.50 × 9.00) and immunoplate (96-well flat bottom cell culture plate) were acquired from SPL Life Sciences Co. Ltd. (Gyeonggi-do, Korea). 0.45 mm PTFE membrane filter paper (Chiyoda City, Japan) was acquired from Advantec Co., Ltd. A sterile 100 mm2 cell culture dish was acquired from SPL Life Sciences Co. Ltd. (Gyeonggi-do, Korea). QIAamp DNA Mini kit was acquired from Qiagen (Seoul, Korea). BPDE-DNA adduct enzyme-linked immunosorbent assay (ELISA) kit was acquired from Cell Biolabs (Seoul, Korea).
2.2. Sample preparations for isolation of neferine
Neferine was isolated from the Nelumbo nucifera GAERTN of lotus. Dried Nelumbo nucifera GAERTN of lotus was isolated in triplicate with 80% methanol by ultrasonication. The isolated solution was evaporated under 37°C to concentrate a volume of 0.5 L. The concentrated solution was dissolved in water. The water solution was divided in ethyl acetate, CHCl3, n-hexane and ethyl acetate fractions were eluted with water : methanol (1:4). The extracts were filtered by using 0.45 mm PTFE membrane filter paper (Chiyoda City, Japan) [22].
2.3. Sample preparations for isolation of daidzein, genistein
Daidzein and genistein were isolated from soybean seeds that grind into the powder. The soybean seeds were kept in polyethylene bag on frozen state until next use. 80% methanol solution was used as a solvent. The 10 mL of 80% methanol solution was added to a glass reactor and heated in the bath for 20 min. The 10 g of the soybean powder was added to the glass reactor. Daidzein and genistein were isolated for 60 min with centrifugation at 500 rpm. The 1.0 mL of extract were obtained in the reactor at 10 min and 1.0 mL of solvent was added into it. The extracts were filtered by using a 0.45 mm PTFE membrane filter paper (Chiyoda City, Japan). This filtrate was diluted and the powder of daidzein and genistein were prepared [23].
2.4. HepG2 cells culture and treatment
HepG2 cells were obtained from the Cell Bank of the Chinese Academy of Sciences (Shanghai, China). They were cultured with 10% FBS in MEM, 1 mM sodium pyruvate mixing 100 U/mL penicillin-streptomycin in a 100 mm2 cell culture dish with 5% CO2 at 37℃ in incubator. They were incubated in MEM at of B[a]P, neferine, daidzein, genistein for 48 h. They were prepared to analyzed the cell-free extracts.
2.5. Cell viability and proliferation assay
In order to assess the cytotoxicity of B[a]P, neferine, daidzein, genistein on HepG2 cells, we evaluated the cell viability. HepG2 cells were seeded on 1 × 104 cells/well in MEM in immunoplate (96-well flat bottom cell culture plate) with B[a]P, neferine, daidzein, genistein at 0, 1, 2, 5, 10, 20 µM concentrations for 48 h. After this procedure, EZ-CYTOX reagent 100 µL was added to each well in immunoplate (96-well flat bottom cell culture plate). The cells were incubated for 2 h. The absorbance at a wavelength of 450 nm by using a microplate reader (PerkinElmer Victor X4, Seoul, Korea) was subjected in these immunoplate (96-well flat bottom cell culture plate). The cell viability was compared between the plates treated with B[a]P, neferine, daidzein, genistein.
2.6. Cell isolation
HepG2 cells seeding 5×10⁴ cells in 100 mm2 dish and cultivated for 24 h for adherence. The medium then was replaced and treated with B[a]P, neferine, daidzein, and genistein for 48 h to achieve the performance characteristics. 1.0 mL of culture medium was isolated from a cell culture dish, translated into 1.5 mL EP tube and centrifuge at 15,000 rpm for 30 min at 4 °C. 1.0 mL of supernatant was put into the sample vials for HPLC-UV detection. Cell culture medium (CM) samples were stored at 4 °C. The medium was eliminated from the dishes, cleaned triplicate with PBS preheated at 37 °C to dispose the disturbance. To each dish 700 μL of KH2PO4 pre-cooled to 4 °C was added and the cells were scraped from the dish with cell scrapers. The scraped cells from each culture dish were collected into a 1.5 mL EP tube, freeze-thawed three times to break cells then centrifuged at 15,000 rpm for 30 min at 4 °C. 500 μL of supernatant was put into the sample vials for HPLC-UV detection.
2.7. HPLC-UV analysis for method validation
The stock solution was prepared at five different concentrations (3, 5, 10, 20, 40 ug/kg) through solving an adequate amount of neferine, daidzein, and genistein powder in 80 % methanol. The stock solution was kept in the refrigerator for examinations with five different concentrations (3, 5, 10, 20, 40 ug/kg) to be kept in the sample vials which then was injected into the system. The HPLC-UV analytical method was validated for linearity, recovery (%), limit of detection (LOD), limit of quantification (LOQ), and precision (%) in lotus and soybean samples. Dionex-C18 column (4.6 mm i.d. × 150 mm × 5 µm) was used in neferine. Hypersil-ODS C18 column (4.6 mm i.d. × 20 cm × 5 µm) was used in daidzein and genistein. A mixture of methanol with 0.1% Trifluoroacetic acid (TFA) in water, methanol and acetonitrile to select mobile phase. In mobile phase, neferine, daidzein, and genistein were injected to the system, and then according to the shape of the repeatability and peak, the appropriate results of the mobile phase were expressed. The flow rate was 1.0 mL/min. The UV wavelength was acquired from 205 nm in neferine and 262 nm in daidzein and genistein [22,23].
2.8. BPDE-DNA adduct formation analysis
HepG2 cells were seeded on immunoplate (96-well flat bottom cell culture plate) in MEM and treated with 10 µM B[a]P alone and 10 µM B[a]P with 20 µM neferine, daidzein, genistein for 48 h. QIAamp DNA Mini Kit (Qiagen, Stanford, CA, USA) isolated the DNA from the HepG2 cells. BPDE-DNA adduct ELISA Kit (Cell Biolabs, San Diego, CA, USA) formed the BPDE-DNA adduct. The absorbance for the BPDE-DNA adduct levels was measured at 450 nm wavelength by using a microplate reader (PerkinElmer Victor X4, Seoul, Korea).
2.9. The typical intracellular metabolites of B[a]P were measured by high performance liquid chromatography (HPLC)
HepG2 cells were treated with 10 µM B[a]P alone and 10 µM B[a]P with 20 µM neferine, daidzein, genistein in 100 mm2 cell culture dishes for 48 h and were mixed with ethyl acetate. The mixture was evaporated by using a vacuum rotary evaporator to add 50% acetonitrile and 0.1% acetic acid mixed solution. The 100 µL of the mixture was injected into an HPLC instrument (Dionex U3000 HPLC, Thermo Fisher Scientific, Sunnyvale, CA, USA) equipped with a Kinetex C18 Plus column (4.6 mm id × 250 mm × 5 µm; Phenomenex, Torrance, CA, USA). The flow rate was 1.2 mL/min at 30 ℃. The mobile phase was consisted of 0.1 % acetic acid in distilled water used as solvent A and 0.1 % acetic acid with 50% acetonitrile in distilled water used as solvent B. Chromatogram’s separation was conducted with the following gradient program: 50% B for 0-40 min and 25-30 min for 100% acetonitrile. The fragmentation and retention time were acquired from BPDE, B[a]P-7,8-dihydrodiol and B[a]P from the reference standards in the MRI Global Chemical Carcinogen Repository (Kansas City, MO, USA).
2.10. Statistical analysis
All experiments were performed in triplicate. All data was expressed as mean ± standard deviation (SD). Significances of differences between the treatment group (B[a]P with neferine, daidzein, genistein) and no-treatment group (B[a]P) were determined by using one-way ANOVA in SPSS. Statistical significance was expressed at p < 0.05.
3. Results
3.1. Reduction of B[a]P-induced hepatotoxicity by neferine, daidzein, genistein
Based on the regulations of the dose-dependent production of BPDE in B[a]P-induced cell deaths varied depending on BPDE-DNA adduct research in the treatment group (B[a]P with neferine, daidzein, genistein) and no-treatment group (B[a]P) were expressed. Accordingly, we studied the potential preventive effects of neferine, daidzein, and genistein against BPDE formation by reducing B[a]P-induced hepatotoxicity using well-characterized HepG2 cells. The toxicities of B[a]P (Figure 1A) and neferine (Figure 1B), daidzein (Figure 1C), genistein (Figure 1D) on HepG2 cells were evaluated by using cell viability assays. Because neferine, daidzein, genistein were non-toxic (20 µM for 48 h), B[a]P induced cell death in a dose-dependent manner in comparison with treatment in Figure 1A-D. The protective effects of neferine, daidzein, genistein against B[a]P-induced hepatotoxicity, B[a]P was applied to HepG2 cells together with various concentrations of neferine, daidzein, genistein. Neferine, daidzein, genistein reduced B[a]P-induced hepatotoxicity and restored cell viability up to 90% in a dose-dependent manner. These expressed that neferine, daidzein, genistein have a protective effect against B[a]P-induced hepatotoxicity. Therefore, the potential preventive effect of neferine, daidzein, genistein against B[a]P-induced hepatotoxicity was evaluated in Figure 2,3,4A. These formation was expressed by BPDE-DNA adduct enzyme-linked immunosorbent assay (ELISA) kit after treatment with B[a]P (10 µM), neferine, daidzein, genistein (20 µM), and B[a]P (10 µM) co-administered with neferine, daidzein, genistein (20 µM). It was resulted that B[a]P (10 µM) co-administered with neferine, daidzein, genistein (20 µM) treatment alone decreased the BPDE-DNA adduct level compared to the B[a]P treatment alone in Figure 2, Figure 3 and Figure 4B.
3.2. Reduction of intracellular B[a]P metabolites by neferine, daidzein, genistein
B[a]P was metabolized to B[a]P-7,8-dihydrodiol and BPDE according to the procedure. BPDE caused hepatotoxicity by increasing mutagenic adducts. The quantity of B[a]P, B[a]P-7,8-dihydrodiol and BPDE in the treated with HepG2 cells were expressed by using a high performance liquid chromatography (HPLC) system. The calculated amount of BPDE rapidly increased in the cells were treated with B[a]P alone. As a result, B[a]P-7,8-dihydrodiol levels increased. In contrast, BPDE levels decreased and B[a]P, B[a]P with neferine, daidzein, genistein compared to the levels with B[a]P treatment alone in Figure 5, Figure 6 and Figure 7.
3.3. Validation of analytical method for neferine, daidzein, genistein analysis
The HPLC chromatograms of neferine, daidzein and genistein standards, neferine, daidzein and genistein in a spiked sample were presented in Figure 8A−B, respectively.
Calibration curves were constructed for the neferine, daidzein and genistein at five concentrations (3, 5, 10, 20, and 40 μg/kg) by plotting concentration against the peak area. Regression analysis of each calibration curve yielded a slope of satisfactory linearity with values for the coefficient of determination (R2) > 0.99 over the tested concentration range. For the lotus and soybean matrices, the LOD was below 0.2 μg/kg (0.08−0.12 μg/kg) and the LOQ was below 0.6 μg/kg (0.24−0.36 μg/kg). The limit of detection (LOD), limit of quantification (LOQ) and linearity (R2) and calibration equations for validation of the neferine in lotus and daidzein and genistein in soybean matrices by using HPLC-UV analysis were presented in Table 1.
The recovery values of the neferine, daidzein and genistein (Table 2) were expressed through the peak areas of the neferine, daidzein and genistein standards. Recovery values of neferine in lotus, daidzein and genistein in soybean samples ranged from 82.95% to 111.46%. Precision was performed at five different concentrations (3, 5, 10, 20, and 40 μg/kg) and mean values of the five different concentrations (3, 5, 10, 20, and 40 μg/kg) in intra, interday of precision of the neferine in lotus, daidzein and genistein in soybean were expressed in Table 3. The results of the intraday analysis (performed three times a day) and the interday analysis (carried out simultaneously on 3 days) showed a precision (%CV) of <15%. More specifically, the precision of neferine in lotus, daidzein and genistein in soybean analysis were in the ranges of 0.24−6.82% (intraday), and 0.65−4.67% (interday), respectively. Collectively, these data show the effectiveness of the proposed methodology for lotus and soybean matrices. The authors observed no substantial differences in sample concentration and recoveries between the two methods over the concentration range studied (3.0−40.0 μg/kg).
3.4. Comparison of neferine, daidzein and genistein concentrations in lotus and soybean samples
The concentrations of the neferine, daidzein and genistein were determined. These data were presented for 70 samples of the concentrations of neferine, daidzein and genistein in lotus and soybean matrices using HPLC-UV analysis (Table 4). In the lotus and soybean samples, neferine was 0.78 μg/kg, daidzein was 1.40 μg/kg and genistein was 1.01 μg/kg. Among the lotus and soybean samples, sample 69 recorded the highest concentration of neferine (2.45 μg/kg), the highest concentration of daidzein (3.42 μg/kg) and the highest concentration of genistein (3.23 μg/kg).
4. Discussion
These results expressed that neferine, daidzein, genistein exerts an antigenotoxic effect by reducing the formation of BPDE-DNA adducts. B[a]P was known to regulate various physiological signaling pathways, leading to various side effects related to cellular toxicity. In this regard, neferine has been reported to regulate various signaling pathways related to B[a]P-induced cellular toxicity. Specifically, neferine was reported to inhibit the B[a]P-induced NF-κB pathway [24,25,26]. NF-κB was one of the inflammatory signaling pathways that can induce inflammatory responses related to B[a]P, and it was believed that neferine reduces cellular toxicity by inhibiting the NF-κB signaling pathway. Additionally, neferine was reported to inhibit the B[a]P-induced MAPK pathway [27,28,29]. MAPK was one of the various signaling pathways related to cell survival, growth, inflammation, stress responses, and others. B[a]P was known to regulate the MAPK pathway to induce cellular toxicity, and it was believed that neferine reduces cellular toxicity by inhibiting the MAPK signaling pathway. Furthermore, neferine was reported to inhibit the B[a]P-induced PI3K-Akt pathway [30]. PI3K-Akt was one of the signaling pathways related to cell survival and growth. B[a]P was known to regulate the PI3K-Akt signaling pathway to induce cellular toxicity, and it was believed that neferine reduces cellular toxicity by inhibiting the PI3K-Akt pathway.
Other similar studies express the well-characterized HepG2 cell to study the potential preventive effects of silymarin, quercetin and isorhamnetin on BPDE formation by reducing the cell toxicity induced by B[a]P. The toxicities of B[a]P and silymarin, quercetin and isorhamnetin on HepG2 cells were evaluated by using cell viability assays and the potential preventive effect of silymarin, quercetin and isorhamnetin against B[a]P-induced hepatotoxicity to confirm that silymarin, quercetin and isorhamnetin inhibits BPDE-DNA adduct formation compared to the effects of B[a]P alone and indicate that silymarin, quercetin and isorhamnetin attenuate B[a]P-induced hepatotoxicity by inhibiting BPDE-DNA adduct formation [31,32].
These results represented that neferine, daidzein, genistein reduced B[a]P-induced hepatotoxicity by the inhibition of BPDE-DNA adduct formation and excretion of BPDE. BPDE was one of the metabolites of B[a]P, which can form adducts with cellular DNA and molecules, leading to genetic mutations and cell damage. In particular, neferine has been found to be the most effective in reducing BPDE-DNA adduct levels which were believed to be due to its mechanisms such as antioxidant activity, modulation of metabolic enzymes, and enhancement of DNA repair. Neferine has been reported to have antioxidant activity, and this action was believed to inhibit the generation of BPDE-DNA adducts. Therefore, neferine has been reported to prevent DNA damage by reducing intracellular ROS (Radical Oxygen Species) and oxidative stress [33,34,35,36,37]. Additionally, B[a]P activates intracellular metabolic enzymes (CYP1A1 and CYP1B1) to produce BPDE, while neferine was reported to inhibit the expression of CYP1A1 and CYP1B1, thereby inhibiting the production of BPDE [28]. Furthermore, BPDE-DNA adducts cause DNA damage, and in order to repair it, cells need to activate DNA repair mechanisms. In this process, neferine was reported to prevent the formation of BPDE-DNA adducts and enhance DNA repair mechanisms, thus preventing DNA damage [38]. In this study, neferine was found to effectively inhibit the formation of intracellular B[a]P metabolites through the mechanisms mentioned above. However, further studies were needed to investigate the specific mechanisms by which neferine effectively inhibits the formation of intracellular B[a]P metabolites. In addition, studies have reported that daidzein and genistein also reduce intracellular B[a]P metabolites [39,40]. However, the results of this study showed that they were not as effective as neferine.
Other similar studies identify the mechanism of BPDE-DNA adduct formation by silymarin, quercetin and isorhamnetin and evaluated the amount of B[a]P metabolites produced. Silymarin, quercetin and isorhamnetin co-treatment with B[a]P resulted in greater production of B[a]P and B[a]P-7,8-dihydrodiol than B[a]P alone. These results represented that silymarin, quercetin and isorhamnetin reduced B[a]P-induced hepatotoxicity by the inhibition of BPDE-DNA adduct formation and excretion of BPDE [31,32]. The high linearity (R2 > 0.99) of our analytical method was similar to that reported for other spiked sample product matrices, including other lotus and soybean samples [12,22,23]. This indicates that the analytical method can accurately detect and quantify, and can be useful for detecting these substances in the medical industry. Generally, the lower the LOD and LOQ values, the higher the sensitivity of the analytical method, which allows for more sensitive detection. This helps to improve the accuracy and reliability of the analytical method. Therefore, it can be said that analytical results are more reliable when the LOD and LOQ values are lower. These same studies obtained values for LOD of 1.3−4.4 μg/mL and LOQ of 3.9−13.2 μg/mL, which were higher than the LOD and LOQ results of our validation tests [22,41]. It was noteworthy that the LOD and LOQ were similar between the lotus and soybean matrices.
The occurrence of neferine, daidzein and genistein in herbal medicines was typically a consequence of the ubiquitous presence of these compounds in the environment, unintentional production during cooking or a manufacturing process. To increase the recovery of these HPLC-UV methods, it was important to select an appropriate isolation solvent with similar polarity to the neferine, daidzein and genistein. Most of the relevant studies have reported that methanol was a suitable isolation solvent because it has adequate polarity and yields a good recovery for most compounds [42].
In this study, we established, validated, and applied an HPLC-UV method for the evaluation of neferine in lotus and daidzein and genistein in soybean matrices in Korea. In the context of these limits, all of the lotus and soybean samples analyzed in the current study were recognized as having safe levels of neferine, daidzein and genistein. The specific structures of the lotus and soybean samples analyzed in the current study were the same as those reported in a study of neferine, daidzein and genistein in various lotus and soybean samples [22,41].
Our study was consistent with those reported in other studies previously confirmed that each lotus and soybean samples contained safe neferine, daidzein and genistein levels. With few previously conducted studies on this topic, accurate quantitative analysis methods were required. Therefore, this study can provide basic scientific data for safety management.
5. Conclusions
In conclusion, our studies represented that B[a]P detoxification prevent the formation of BPDE-DNA adducts by reducing intracellular B[a]P metabolites. In HepG2 cells, neferine in lotus and daidzein, genistein in soybean reduced the BPDE-DNA adduct formation by B[a]P-7,8-dihydrodiol-9,10-epoxide (BPDE) derived by benzo[a]pyrene (B[a]P). These results suggest that neferine in lotus and daidzein, genistein in soybean prevent B[a]P-induced hepatotoxicity for BPDE-DNA adduct formation.
Author Contributions
Y.-Y.K.: Methodology, Visualization, Formal analysis, Writing—original draft preparation, reviewing and editing, H.-H.S.: Project administration, Writing—review and editing. Author was agreed to the published version of the manuscript.
Funding
This research was received no funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The datasets generated for this study are available on request to the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Phillips, D.H.; Venitt S. DNA and protein adducts in human tissues resulting from exposure to tobacco smoke. Int J Cancer. 2012, 131(12), 2733-2753. 12. [CrossRef]
- Baird, W.M.; Hooven. L.A.; Mahadevan. B. Carcinogenic polycyclic aromatic hydrocarbon-DNA adducts and mechanism of action. Environ Mol Mutagen. 2005, 45(2-3), 106-114. [CrossRef]
- Saunders. C.R.; Das, S.K.; Ramesh, A.; Shockley, D.C.; Mukherjee, S. Benzo[a]pyrene-induced acute neurotoxicity in the F-344 rat: role of oxidative stress. J Appl Toxicol. 2006, 26(5), 427-438. [CrossRef]
- Kim, K.H.; Jahan, S.A.; Kabir, E.; Brown, R.J. A review of airborne polycyclic aromatic hydrocarbons (PAHs) and their human health effects. Environ Int. 2013, 60, 71-80. [CrossRef]
- McCormick, S.; Snawder, J.E.; Chen, I.C.; Slone, J.; Calafat, A.M.; Wang, Y.; Meng, L.; Alexander-Scott M,; Breitenstein M.; Johnson B.; Meadows J.; Fairfield EC. Exposure assessment of polycyclic aromatic hydrocarbons in refined coal tar sealant applications. Int J Hyg Environ Health. 2022, 242, 113971. [CrossRef]
- Ramesh, A.; Inyang, F.; Hood, D.B.; Archibong, A.E.; Knuckles, M.E.; Nyanda, A.M. Metabolism, bioavailability, and toxicokinetics of benzo(alpha)pyrene in F-344 rats following oral administration. Exp Toxicol Pathol. 2001, 53(4), 275-290. 4.
- Li, F.; Xiang, B.; Jin, Y.; Li, C.; Ren, S.; Wu, Y.; Li, J.; Luo, Q. Hepatotoxic effects of inhalation exposure to polycyclic aromatic hydrocarbons on lipid metabolism of C57BL/6 mice. Environ Int. 2020, 134, 105000. [Google Scholar] [CrossRef]
- Jaeschke, H. Reactive oxygen and mechanisms of inflammatory liver injury. J Gastroenterol Hepatol. 2000, 15(7), 718–724. [Google Scholar] [CrossRef] [PubMed]
- Miller, K.P.; Ramos, K.S. Impact of cellular metabolism on the biological effects of benzo[a]pyrene and related hydrocarbons. Drug Metab Rev. 2001, 33(1), 1–35. [Google Scholar] [CrossRef]
- Zhao, L.; Wang, X.; Chang, Q.; Xu, J.; Huang, Y.; Guo, Q. Neferine, a bisbenzylisoquinline alkaloid attenuates bleomycin-induced pulmonary fibrosis. Eur. J. Pharmacol. 2010, 627, 304–312. [Google Scholar] [CrossRef] [PubMed]
- Mahbubur Rahman AHM. Traditional medicinal plants used in the treatment of different skin diseases of santals at abdullahpur village under akkelpur upazilla of joypurhat district, Bangladesh. Biomed. and Biotechnol. 2013, 1(2), 17-20.
- Chen, Y.; Fan, G.R.; Wu, H.L.; Wu, Y.T.; Mitchell, A. Separation, identification and rapid determination of liensine, isoliensinine and neferine from embryo of the seed of Nelumbo nucifera GAERTN by liquid chromatography coupled to diode array detector and tandem mass spectrometry. J. Pharm. Biomed. Anal. 2007, 43, 99–104. [Google Scholar] [CrossRef]
- Poornima, P.; Weng, C.F.; Padma, V.V. Neferine, an alkaloid from lotus seed embryo, inhibits human lung cancer cell growth by MAPK activation and cell cycle arrest. Biofactors. 2014, 40(1), 121–131. [Google Scholar] [CrossRef]
- Liggins, J.; Mulligan, A.; Runswick, S.; Bingham, S.A. Daidzein and genistein content of cereals. Eur. J. Clin Nutr. 2002, 56(10), 961–966. [Google Scholar] [CrossRef]
- Sirtori, C.R. Risks and benefits of soy phytoestrogens in cardiovascular diseases, cancer, climacteric symptoms and osteoporosis. Drug. Saf. 2001, 24(9), 665–682. [Google Scholar] [CrossRef]
- Adjakly, M.; Ngollo, M.; Boiteux, J.P.; Bignon, Y.J.; Guy, L.; Gallon, D.B. Genistein and daidzein: different molecular effects on prostate cancer. Anticancer Res. 2013, 33(1), 39–44. [Google Scholar]
- Sarao, L.; Kaur, S.; Malik, T.; Singh, A. Chapter 19 – Genistein and daidzein. Nutraceuticals and Health Care. 2022, 331–341. [Google Scholar]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the last 25 years. J Nat Prod. 2007, 70(3), 461–477. [Google Scholar] [CrossRef] [PubMed]
- Ji, X.; Li, Y.; He, J.; Shah, W.; Xue, X.; Feng, G.; Zhang, H.; Gao, M. Depletion of mitochondrial enzyme system in liver, lung, brain, stomach and kidney induced by benzo[a]pyrene. Environ Toxicol Pharmacol. 2016, 43, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Miao, W.; Lin, X.; Wu, T.; Shen, H.; Chen, S.; Li, Y.; Pan, Q.; Fu, Z. Sub-chronically exposing mice to a polycyclic aromatic hydrocarbon increases lipid accumulation in their livers. Environ Toxicol Pharmacol. 2014, 38(2), 353–363. [Google Scholar] [CrossRef] [PubMed]
- Mumtaz, M.M.; George, J.D.; Gold, K.W.; Cibulas, W.; DeRosa, C.T. ATSDR evaluation of health effects of chemicals. IV. Polycyclic aromatic hydrocarbons (PAHs): understanding a complex problem. Toxicol Ind Health. 1996, 12(6), 742-971. [CrossRef]
- Ryu, G.H.; Weon, J.B.; Yang, W.S.; Ma, C.J. Simultaneous determination of four compounds in a Nelumbo nucifera seed embryo by HPLC-DAD. J. Spectrosc. 2017, 1–6. [Google Scholar] [CrossRef]
- Arau´jo, J.M.A.; Silva, M.V.; Chaves, J.B.P. Supercritical fluid extraction of daidzein and genistein isoflavones from soybean hypocotyl after hydrolysis with endogenous ß-glucosidases. Food Chem. 2007, 105, 266–272. [Google Scholar] [CrossRef]
- Guan, G.; Han, H.; Yang, Y.; Jin, Y.; Wang, X.; Liu, X. Neferine prevented hyperglycemia-induced endothelial cell apoptosis through suppressing ROS/Akt/NF-κB signal. Endocrine. 2014, 47, 764–771. [Google Scholar] [CrossRef]
- Li, H.; Chen, W.; Chen, Y.; Zhou, Q.; Xiao, P.; Tang, R.; Xue, J. Neferine attenuates acute kidney injury by inhibiting NF-κB signaling and upregulating Klotho expression. Frontiers in Pharmacology. 2019, 10, 1197. [Google Scholar] [CrossRef]
- Wang, Y.K.; Li, W.Q.; Xia, S.; Guo, L.; Miao, Y.; Zhang, B.K. Metabolic activation of the toxic natural products from herbal and dietary supplements leading to toxicities. Frontiers in pharmacology. 2021, 12, 758468. [Google Scholar] [CrossRef]
- Eid, W.; Abdel-Rehim, W. Neferine enhances the antitumor effect of mitomycin-C in hela cells through the activation of p38-MAPK pathway. J. Cell. Biochem. 2017, 118(10), 3472–3479. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, S.; Wang, R.; Li, S.; Yuan, Y. Neferine exerts antioxidant and anti-inflammatory effects on carbon tetrachloride-induced liver fibrosis by inhibiting the MAPK and NF-κB/IκBα pathways. Evid. Based Complementary Altern. Med. 2021, 2021, 1–12. [Google Scholar] [CrossRef]
- Guolan, D.; Lingli, W.; Wenyi, H.; Wei, Z.; Baowei, C.; Sen, B. Anti-inflammatory effects of neferine on LPS-induced human endothelium via MAPK, and NF-κβ pathways. Die Pharmazie. 2018, 73(9), 541–544. [Google Scholar] [CrossRef] [PubMed]
- Poornima, P.; Weng, C.F.; Padma, V.V. Neferine from Nelumbo nucifera induces autophagy through the inhibition of PI3K/Akt/mTOR pathway and ROS hyper generation in A549 cells. Food chem. 2013, 141(4), 3598–3605. [Google Scholar] [CrossRef]
- Jee, S.C.; Kim, M.; Sung, J.S. Modulatory effects of silymarin on benzo[a]pyrene-induced hepatotoxicity. Int. J. Mol. Sci. 2020, 21, 2369–2384. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Jee, S.C.; Kim, K.S.; Kim, H.S.; Yu, K.N.; Sung, J.S. Quercetin and isorhamnetin attenuate benzo[a]pyrene-induced toxicity by modulating detoxification enzymes through the AhR and NRF2 signaling pathways. Antioxidants. 2021, 10, 787–801. [Google Scholar] [CrossRef] [PubMed]
- Jahan, N.; Chowdhury, A.; Li, T.; Xu, K.; Wei, F.; Wang, S. Neferine improves oxidative stress and apoptosis in benign prostate hyperplasia via Nrf2-ARE pathway. Redox Rep. 2021, 26(1), 1–9. [Google Scholar] [CrossRef]
- Jung, H.A.; Jin, S.E.; Choi, R.J.; Kim, D.H.; Kim, Y.S.; Ryu, J.H.; Choi, J.S. Anti-amnesic activity of neferine with antioxidant and anti-inflammatory capacities, as well as inhibition of ChEs and BACE1. Life Sciences. 2010, 87(13-14), 420-430. [CrossRef]
- Asokan, S.M.; Mariappan, R.; Muthusamy, S.; Velmurugan, B.K. Pharmacological benefits of neferine-A comprehensive review. Life Sciences. 2018, 199, 60–70. [Google Scholar] [CrossRef]
- Li, J.; Chou, H.; Li, L.; Li, H.; Cui, Z. Wound healing activity of neferine in experimental diabetic rats through the inhibition of inflammatory cytokines and nrf-2 pathway. Artif Cells Nanomed Biotehcnol. 2020, 48(1), 96–106. [Google Scholar] [CrossRef]
- Khan, A.; Bai, H.; Shu, M.; Chen, M.; Khan, A.; Bai, Z. Antioxidative and antiphotoaging activities of neferine upon UV-A irradiation in human dermal fibroblasts. Biosci. 2018, 38(6), BSR20181414. [Google Scholar]
- Somasundram, B.; Manogaran, P.; Vasudevan, M.; Viswanadha, V.P. Chemosensitizing effect of neferine on cisplatin-resistant colorectal cancer: Identification of potential candidate genes and pathways through whole transcriptome profiling. Phytomedicine Plus. 2022, 2(3), 100299. [Google Scholar] [CrossRef]
- Borradaile, N.M.; Dreu, L.E.; Wilcox, L.J.; Edwards, J.Y.; Huff, M.W. Soya phytoestrogens, genistein and daidzein, decrease apolipoprotein B secretion from HepG2 cells through multiple mechanisms. Biochem J. 2002, 366, 531–539. [Google Scholar] [CrossRef]
- Lu, R.; Zheng, Z.; Yin, Y.; Jiang, Z. Effect of genistein on cholesterol metabolism-related genes in HepG2 cell. J. Food Sci. 2019, 84(8), 2330–2336. [Google Scholar] [CrossRef] [PubMed]
- Nan, G.; Gao, Y.; Guo, L.; Meng, X.; Yang, G. Solid-liquid extraction of daidzein and genistein from soybean: Kinetic modeling of influential factors. Prep. Biochem. Biotechnol. 2018, 48(10), 946–953. [Google Scholar] [CrossRef] [PubMed]
- Maštovská, K.; Lehotay, S.J. Evaluation of common organic solvents for gas chromatographic analysis and stability of multiclass pesticide residues. J. Chromatogr. A. 2004, 1040, 259–272. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
HepG2 cell viability by evaluating cell viability assay treated with benzo[a]pyrene (B[a]P) (A) and neferine (B), daidzein (C), genistein (D) at 0, 1, 2, 5, 10, 20 µM concentrations for 48 h. All treatment group values were considerably different in comparison with the B[a]P (### p < 0.001, ## p < 0.01, # p < 0.05) and the controls (*** p < 0.001) in t-Test for Independent in SPSS.
Figure 1.
HepG2 cell viability by evaluating cell viability assay treated with benzo[a]pyrene (B[a]P) (A) and neferine (B), daidzein (C), genistein (D) at 0, 1, 2, 5, 10, 20 µM concentrations for 48 h. All treatment group values were considerably different in comparison with the B[a]P (### p < 0.001, ## p < 0.01, # p < 0.05) and the controls (*** p < 0.001) in t-Test for Independent in SPSS.
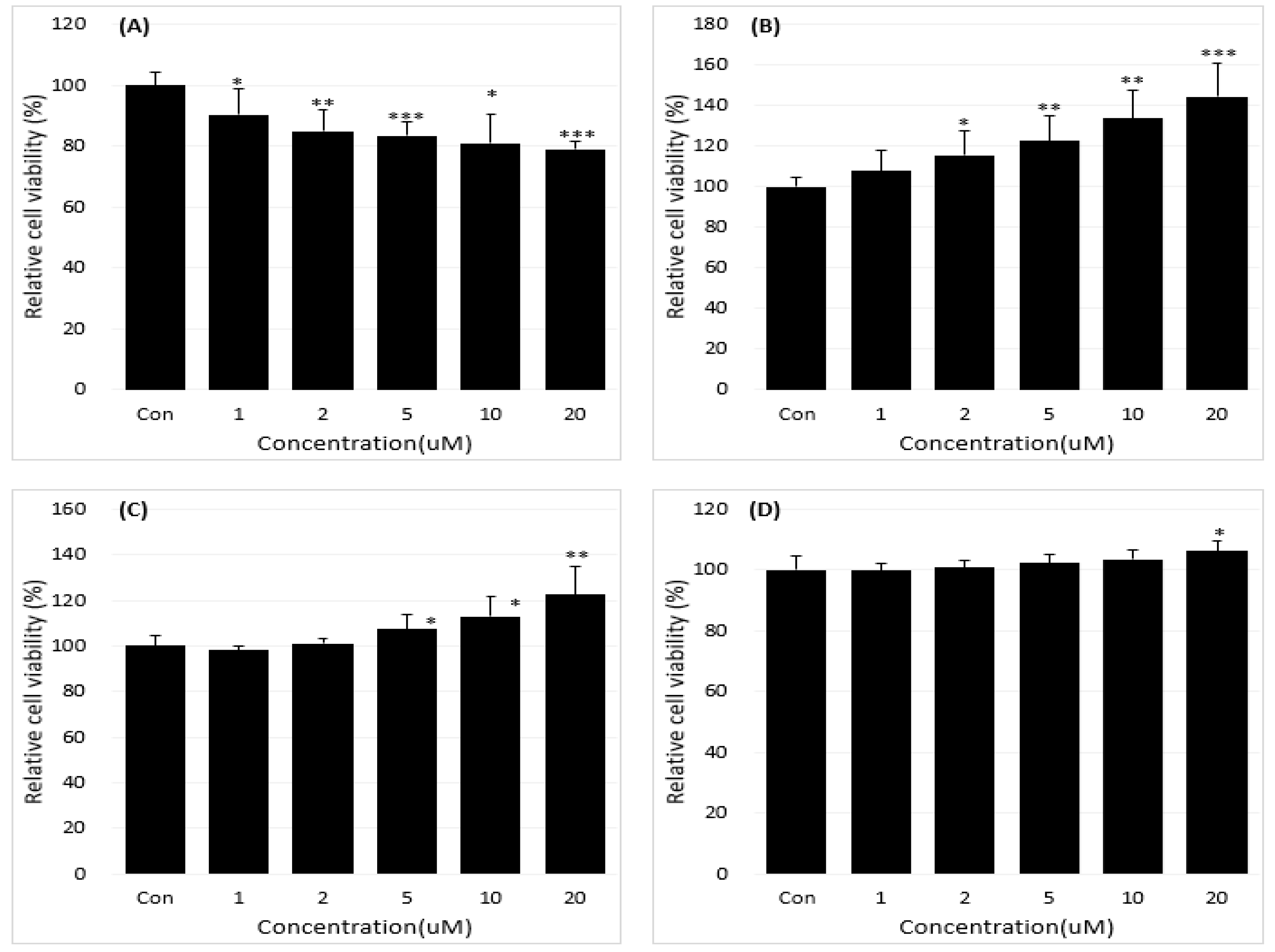
Figure 2.
Reduction of the B[a]P-induced hepatotoxicity in cells treated with 0, 1, 2, 5, 10, 20 µM concentrations of neferine for 48 h (A). The inhibitory effect of neferine on BPDE-DNA adduct formation by using enzyme-linked immunosorbent assay (ELISA) (B). All treatment group values were considerably different in comparison with the B[a]P (### p < 0.001, ## p < 0.01, # p < 0.05) and the controls (*** p < 0.001) in t-Test for Independent in SPSS.
Figure 2.
Reduction of the B[a]P-induced hepatotoxicity in cells treated with 0, 1, 2, 5, 10, 20 µM concentrations of neferine for 48 h (A). The inhibitory effect of neferine on BPDE-DNA adduct formation by using enzyme-linked immunosorbent assay (ELISA) (B). All treatment group values were considerably different in comparison with the B[a]P (### p < 0.001, ## p < 0.01, # p < 0.05) and the controls (*** p < 0.001) in t-Test for Independent in SPSS.
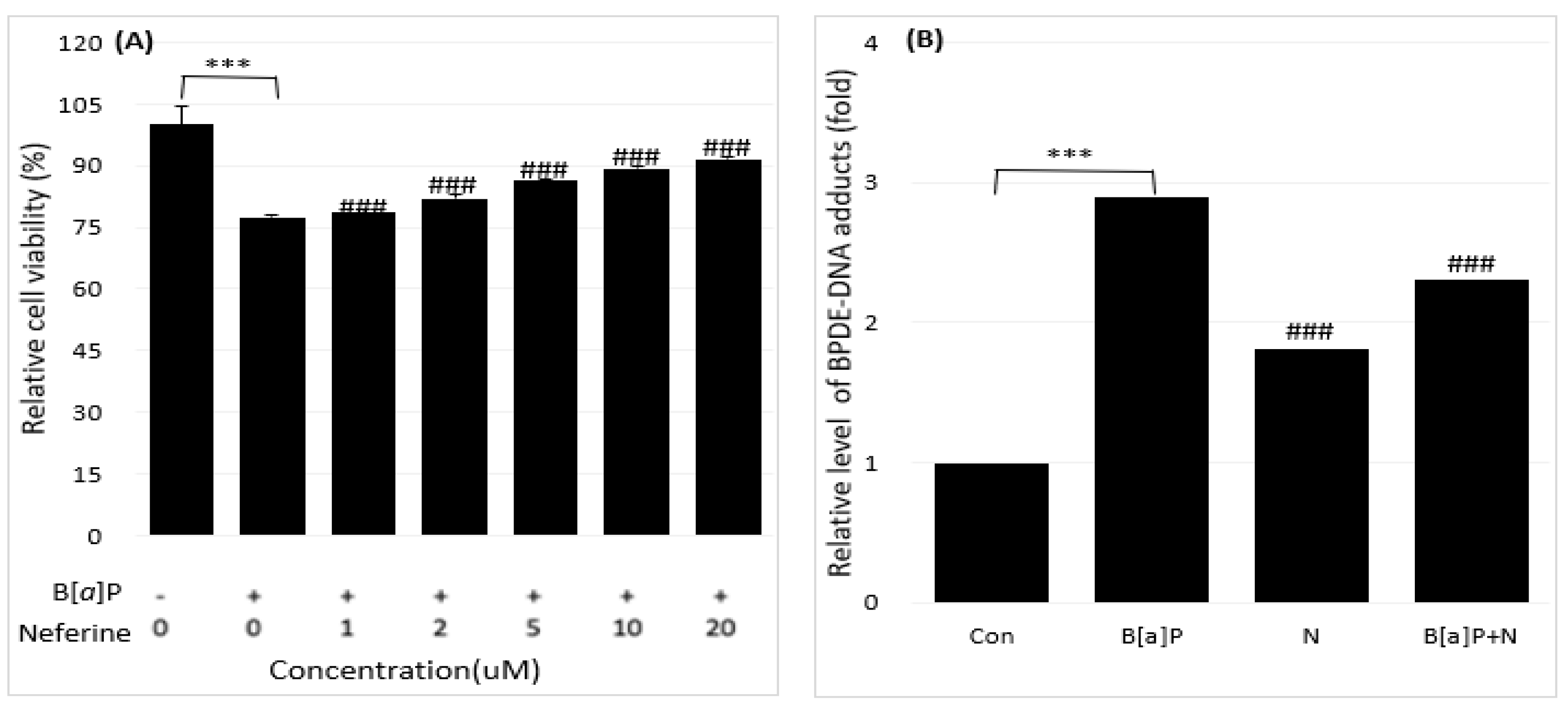
Figure 3.
Reduction of the B[a]P-induced hepatotoxicity in cells treated with 0, 1, 2, 5, 10, 20 µM concentrations of daidzein for 48 h (A). The inhibitory effect of daidzein on BPDE-DNA adduct formation by using enzyme-linked immunosorbent assay (ELISA) (B). All treatment group values were considerably different in comparison with the B[a]P (### p < 0.001, ## p < 0.01, # p < 0.05) and the controls (*** p < 0.001) in t-Test for Independent in SPSS.
Figure 3.
Reduction of the B[a]P-induced hepatotoxicity in cells treated with 0, 1, 2, 5, 10, 20 µM concentrations of daidzein for 48 h (A). The inhibitory effect of daidzein on BPDE-DNA adduct formation by using enzyme-linked immunosorbent assay (ELISA) (B). All treatment group values were considerably different in comparison with the B[a]P (### p < 0.001, ## p < 0.01, # p < 0.05) and the controls (*** p < 0.001) in t-Test for Independent in SPSS.
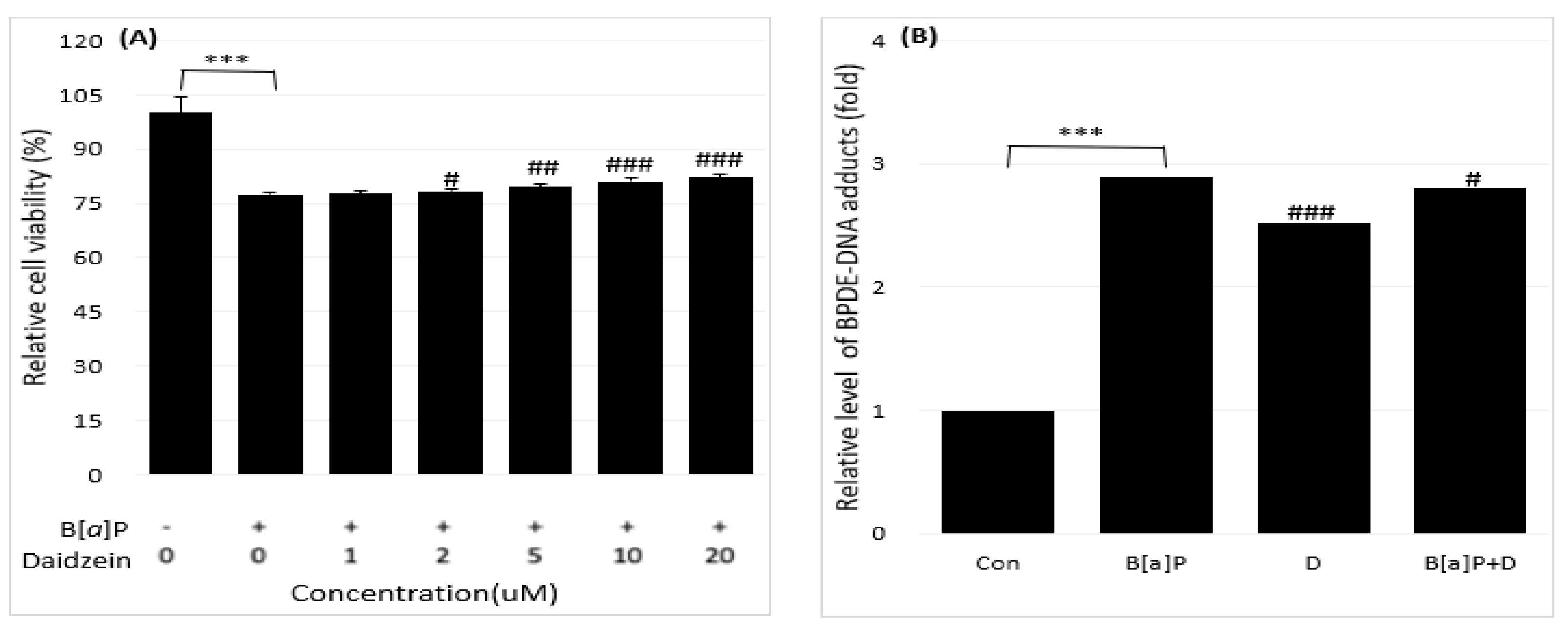
Figure 4.
Reduction of the B[a]P-induced hepatotoxicity in cells treated with 0, 1, 2, 5, 10, 20 µM concentrations of genistein for 48 h (A). The inhibitory effect of genistein on BPDE-DNA adduct formation by using enzyme-linked immunosorbent assay (ELISA) (B). All treatment group values were considerably different in comparison with the B[a]P (### p < 0.001, ## p < 0.01, # p < 0.05) and the controls (*** p < 0.001) in t-Test for Independent in SPSS.
Figure 4.
Reduction of the B[a]P-induced hepatotoxicity in cells treated with 0, 1, 2, 5, 10, 20 µM concentrations of genistein for 48 h (A). The inhibitory effect of genistein on BPDE-DNA adduct formation by using enzyme-linked immunosorbent assay (ELISA) (B). All treatment group values were considerably different in comparison with the B[a]P (### p < 0.001, ## p < 0.01, # p < 0.05) and the controls (*** p < 0.001) in t-Test for Independent in SPSS.
Figure 5.
The representative intracellular metabolites of B[a]P were measured by high performance liquid chromatography (HPLC). M3, BPDE; M2, B[a]P-7,8-dihydrodiol; M1, B[a]P that HepG2 cells were incubated with B[a]P (10 µM) and co-treated with neferine (20 µM) concentrations for 48 h.
Figure 5.
The representative intracellular metabolites of B[a]P were measured by high performance liquid chromatography (HPLC). M3, BPDE; M2, B[a]P-7,8-dihydrodiol; M1, B[a]P that HepG2 cells were incubated with B[a]P (10 µM) and co-treated with neferine (20 µM) concentrations for 48 h.
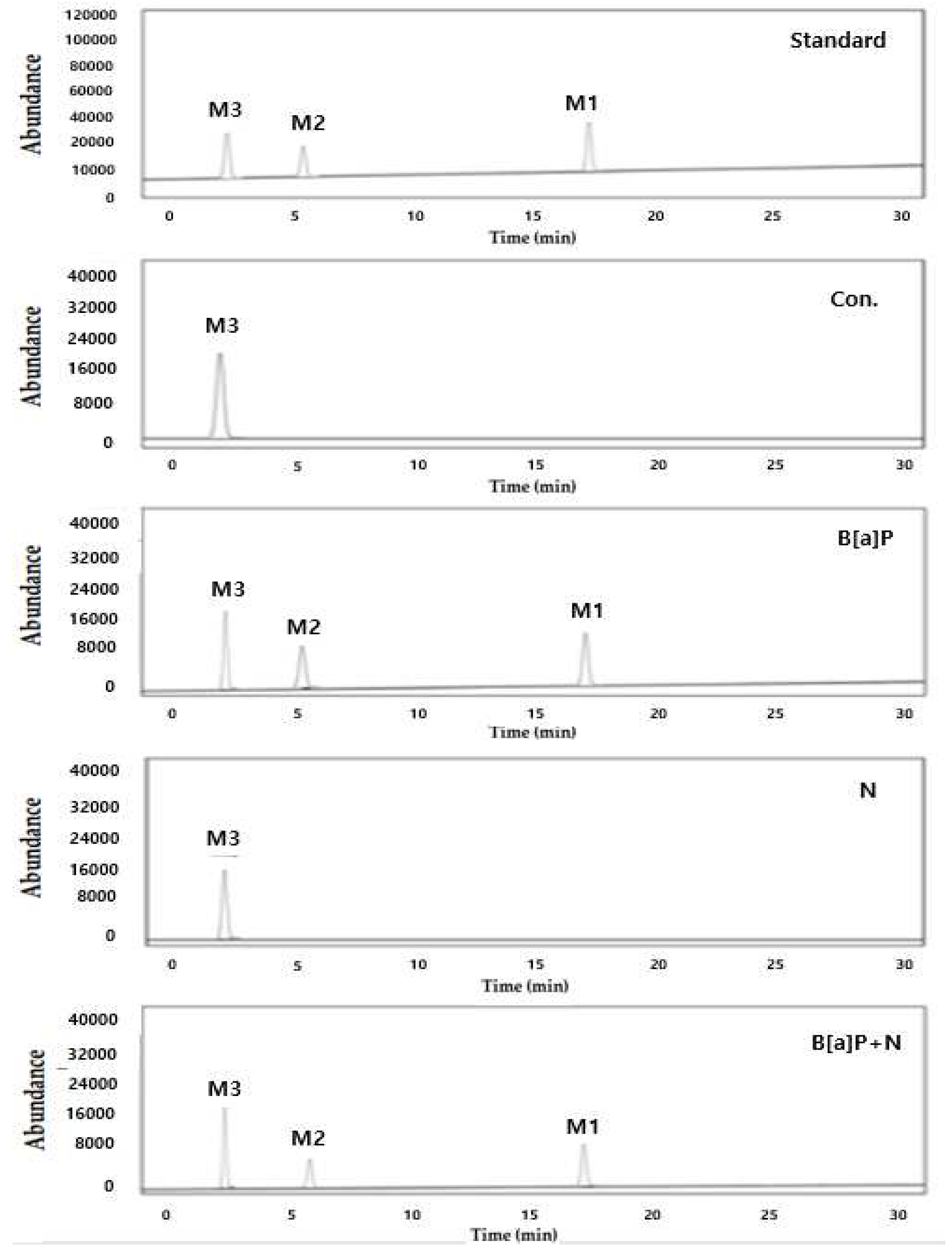
Figure 6.
The representative intracellular metabolites of B[a]P were measured by high performance liquid chromatography (HPLC). M3, BPDE; M2, B[a]P-7,8-dihydrodiol; M1, B[a]P that HepG2 cells were incubated with B[a]P (10 µM) and co-treated with daidzein (20 µM) concentrations for 48 h.
Figure 6.
The representative intracellular metabolites of B[a]P were measured by high performance liquid chromatography (HPLC). M3, BPDE; M2, B[a]P-7,8-dihydrodiol; M1, B[a]P that HepG2 cells were incubated with B[a]P (10 µM) and co-treated with daidzein (20 µM) concentrations for 48 h.
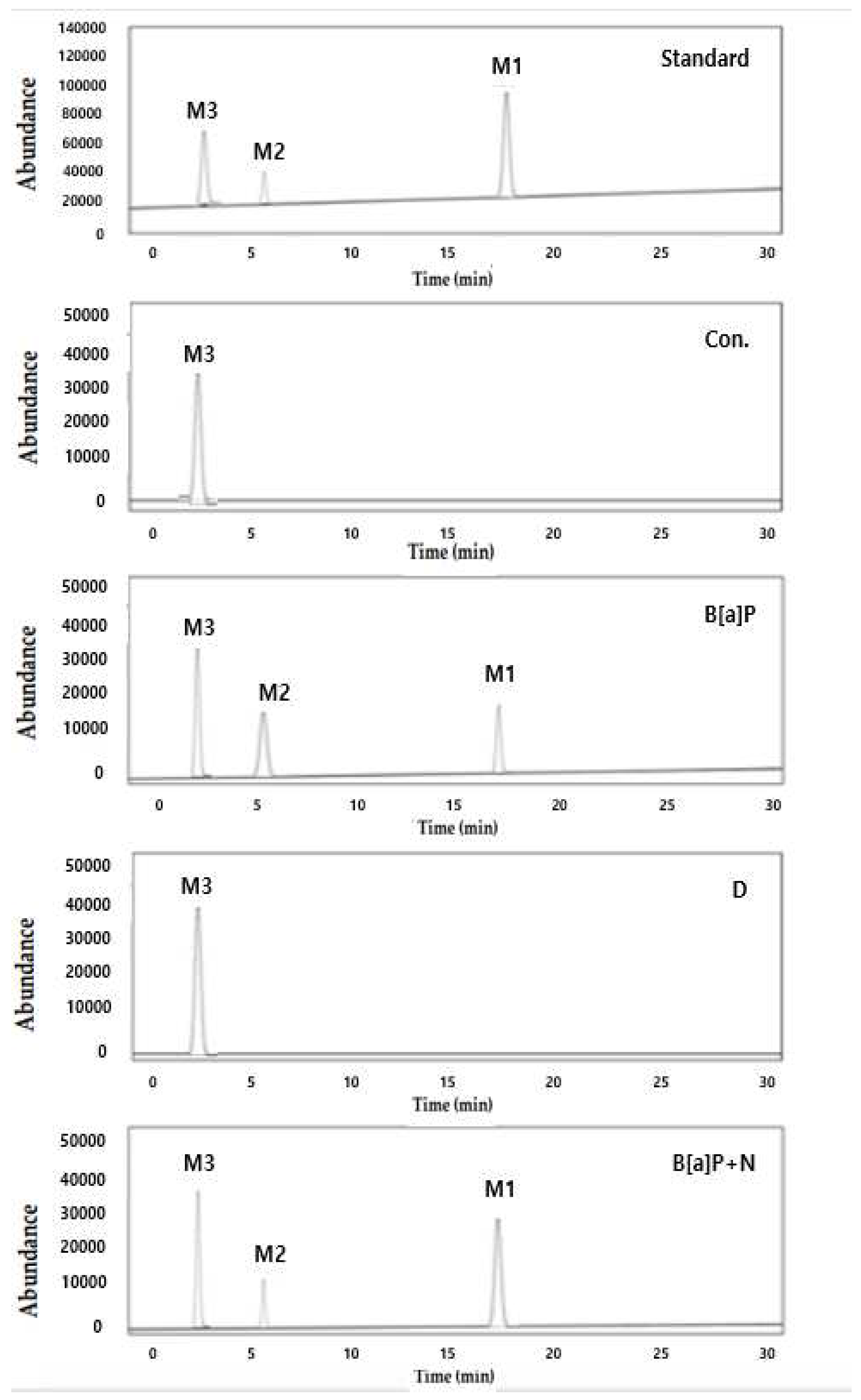
Figure 7.
The representative intracellular metabolites of B[a]P were measured by high performance liquid chromatography (HPLC). M3, BPDE; M2, B[a]P-7,8-dihydrodiol; M1, B[a]P that HepG2 cells were incubated with B[a]P (10 µM) and co-treated with genistein (20 µM) concentrations for 48 h.
Figure 7.
The representative intracellular metabolites of B[a]P were measured by high performance liquid chromatography (HPLC). M3, BPDE; M2, B[a]P-7,8-dihydrodiol; M1, B[a]P that HepG2 cells were incubated with B[a]P (10 µM) and co-treated with genistein (20 µM) concentrations for 48 h.
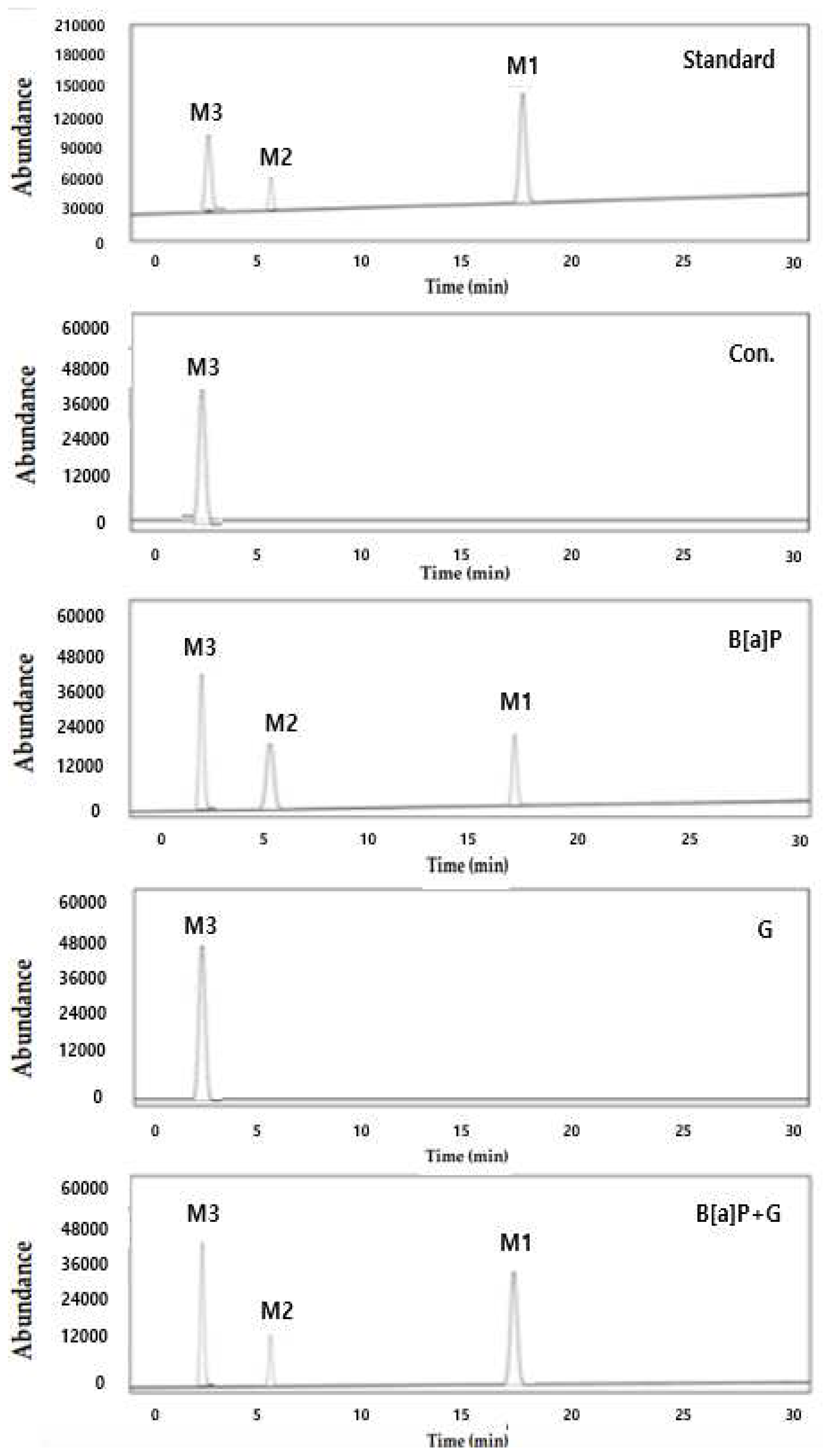
Figure 8.
The HPLC chromatograms of neferine, daidzein and genistein standards (A); neferine, daidzein and genistein in a spiked sample (B).
Figure 8.
The HPLC chromatograms of neferine, daidzein and genistein standards (A); neferine, daidzein and genistein in a spiked sample (B).
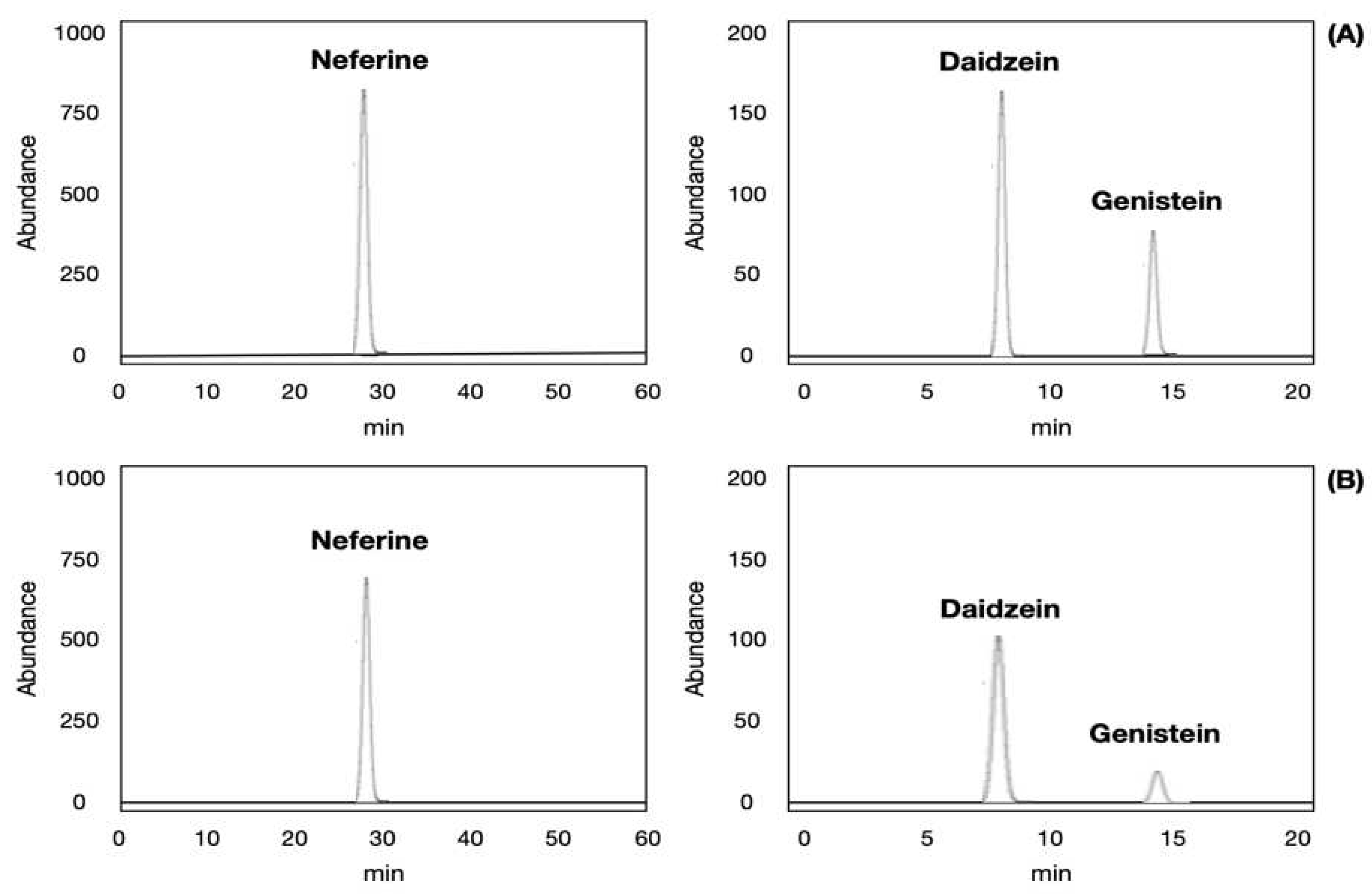

Table 1.
The limit of detection (LOD), limit of quantification (LOQ) and linearity (R2) and calibration equations for validation of the neferine in lotus, daidzein and genistein in soybean matrices using HPLC-UV analysis.
Table 1.
The limit of detection (LOD), limit of quantification (LOQ) and linearity (R2) and calibration equations for validation of the neferine in lotus, daidzein and genistein in soybean matrices using HPLC-UV analysis.
| Matrix type | neferine in lotus and daidzein and genistein in soybean matrices | ||||
|---|---|---|---|---|---|
| LOD (μg/kg)1) |
LOQ (μg/kg)2) |
Linearity3) | Calibration equation | ||
| Lotus matrix (n = 3) |
Neferine | 0.12 | 0.36 | R2=0.9967 |
y=0.0021x+0.007 |
| Soybean matrix (n = 3) |
Daidzein | 0.08 | 0.24 | R2=0.9958 | y=0.0012x-0.008 |
| Genistein | 0.09 | 0.27 | R2=0.9942 | y=0.0015x-0.005 | |
1) LOD is a signal-to-noise ratio (S/N) = 3.3. 2) LOQ is a signal-to-noise ratio (S/N) = 10. 3)Mathematical relationship between neferine, daidzein, genistein concentrations and result of chromatography analysis that can be represented as a straight line (n=3).
Table 2.
Recovery (%)1) for validation of the neferine in lotus, daidzein and genistein in soybean matrices using HPLC-UV analysis.
Table 2.
Recovery (%)1) for validation of the neferine in lotus, daidzein and genistein in soybean matrices using HPLC-UV analysis.
| Matrix type | Conc. | 3 (μg/kg) | 10 (μg/kg) | 40 (μg/kg) |
|---|---|---|---|---|
| Lotus matrix (n = 3) |
Neferine | 82.95±0.67 | 102.75±0.56 | 99.54±0.32 |
| Soybean matrix (n = 3) |
Daidzein | 93.02±2.45 | 111.46±5.67 | 86.72±0.11 |
| Genistein | 96.75±3.64 | 107.65±8.65 | 104.77±0.96 |
1) Recovery (%) was evaluated with 3,10, and 40 μg/kg concentration of spiked neferine in lotus and daidzein, genistein in soybean matrices and are shown as mean ± relative standard deviation (n=3).
Table 3.
Precision (%) for validation of the neferine in lotus, daidzein and genistein in soybean matrices using HPLC-UV analysis.
Table 3.
Precision (%) for validation of the neferine in lotus, daidzein and genistein in soybean matrices using HPLC-UV analysis.
| Matrix type | Conc. | Intraday (n=3) | Interday (n=3) |
|---|---|---|---|
| Precision (%)1) | Precision (%) | ||
| Lotus matrix (n = 3) |
Neferine | 0.24 | 0.65 |
| Soybean matrix (n = 3) |
Daidzein | 1.26 | 3.42 |
| Genistein | 6.82 | 4.67 |
1) Precision (%) was evaluated with CV (coefficient of variation, %) = (standard deviation/mean) × 100.
Table 4.
The concentrations of neferine in lotus, daidzein and genistein in soybean matrices using HPLC-UV analysis.
Table 4.
The concentrations of neferine in lotus, daidzein and genistein in soybean matrices using HPLC-UV analysis.
|
Sample name |
Concentrations (μg/kg)1) | |||
|---|---|---|---|---|
| Neferine | Daidzein | Genistein | Total | |
| Sample 1 | 1.02±0.25 | 1.67±0.86 | 1.11±0.05 | 3.80±1.16 |
| Sample 2 | 0.78±0.11 | 2.45±0.06 | 1.43±0.09 | 4.66±0.26 |
| Sample 3 | 0.66±0.05 | 2.31±0.09 | 1.67±0.08 | 4.64±0.22 |
| Sample 4 | 2.21±0.05 | 0.49±0.07 | 1.90±0.76 | 4.60±0.88 |
| Sample 5 | 0.56±0.23 | 0.41±0.07 | 2.03±0.87 | 3.00±1.17 |
| Sample 6 | 0.47±0.01 | 0.56±0.09 | 2.11±0.78 | 3.14±0.88 |
| Sample 7 | 0.45±0.15 | 1.25±0.99 | 2.78±0.93 | 4.48±2.07 |
| Sample 8 | 1.43±0.17 | 1.56±0.32 | 3.11±0.45 | 6.10±0.94 |
| Sample 9 | 0.87±0.14 | 2.78±0.54 | 0.78±0.32 | 4.52±1.00 |
| Sample 10 | 0.98±0.05 | 3.04±0.11 | 0.54±0.02 | 4.56±0.18 |
| Sample 11 | 0.76±0.34 | 2.93±0.54 | 0.32±0.01 | 4.01±0.89 |
| Sample 12 | 0.88±0.07 | 0.67±0.32 | 0.88±0.75 | 2.43±1.14 |
| Sample 13 | 1.05±0.78 | 0.44±0.11 | 1.02±0.99 | 2.51±1.88 |
| Sample 14 | 1.34±0.54 | 0.78±0.05 | 1.11±0.45 | 3.23±1.04 |
| Sample 15 | 1.97±0.08 | 1.56±0.99 | 1.75±0.03 | 5.28±1.10 |
| Sample 16 | 2.06±0.04 | 0.86±0.54 | 1.32±0.41 | 4.24±0.99 |
| Sample 17 | 2.21±0.03 | 0.54±0.07 | 1.05±0.55 | 3.80±0.65 |
| Sample 18 | <LOQ | 0.32±0.05 | 2.97±0.66 | 3.29±0.71 |
| Sample 19 | <LOQ | 0.27±0.08 | 3.23±0.77 | 3.50±0.85 |
| Sample 20 | <LOQ | 0.67±0.09 | 0.99±0.66 | 1.66±0.75 |
| Sample 21 | <LOQ | 0.99±0.06 | 1.92±0.43 | 2.91±0.49 |
| Sample 22 | <LOQ | 1.76±0.11 | 1.54±0.78 | 3.30±0.89 |
| Sample 23 | <LOQ | 2.01±0.07 | 2.03±0.97 | 4.04±1.04 |
| Sample 24 | <LOQ | 2.26±1.02 | 2.11±0.45 | 4.37±1.47 |
| Sample 25 | <LOQ | 3.02±0.96 | 3.02±0.86 | 6.04±1.82 |
| Sample 26 | <LOQ | 2.05±0.87 | 0.87±0.05 | 2.92±0.92 |
| Sample 27 | 0.55±0.08 | <LOQ | 0.45±0.03 | 1.00±0.11 |
| Sample 28 | 1.54±0.07 | <LOQ | 0.54±0.08 | 2.08±0.15 |
| Sample 29 | 1.23±0.56 | <LOQ | 1.25±0.07 | 2.48±0.63 |
| Sample 30 | 1.86±0.59 | <LOQ | 1.11±0.09 | 2.97±0.68 |
| Sample 31 | <LOQ | 3.01±0.67 | 1.56±0.06 | 4.57±0.73 |
| Sample 32 | <LOQ | 3.33±0.09 | 0.88±0.54 | 4.21±0.63 |
| Sample 33 | <LOQ | 2.06±0.08 | 0.55±0.09 | 2.61±0.17 |
| Sample 34 | <LOQ | 2.54±0.10 | 1.23±0.04 | 3.77±0.14 |
| Sample 35 | 0.56±0.03 | 1.45±0.67 | <LOQ | 2.01±0.70 |
| Sample 36 | 0.32±0.21 | 1.67±0.54 | <LOQ | 1.99±0.75 |
| Sample 37 | 0.45±0.09 | 2.55±0.43 | <LOQ | 3.00±0.52 |
| Sample 38 | 0.78±0.06 | 2.11±0.07 | <LOQ | 2.89±0.13 |
| Sample 39 | <LOQ | 0.99±0.06 | 1.78±0.32 | 2.77±0.38 |
| Sample 40 | <LOQ | 1.54±0.09 | 1.02±0.89 | 2.56±0.98 |
| Sample 41 | <LOQ | 1.23±0.41 | 1.65±0.32 | 2.88±0.73 |
| Sample 42 | 1.87±0.09 | 0.67±0.04 | <LOQ | 2.54±0.13 |
| Sample 43 | 1.56±0.76 | 0.78±0.09 | <LOQ | 2.34±0.85 |
| Sample 44 | 1.32±0.04 | 0.45±0.04 | <LOQ | 1.77±0.08 |
| Sample 45 | 1.13±0.75 | 0.67±0.08 | <LOQ | 1.80±0.83 |
| Sample 46 | 0.97±0.66 | 1.54±0.07 | <LOQ | 2.51±0.73 |
| Sample 47 | 0.45±0.08 | 2.02±0.78 | <LOQ | 2.47±0.86 |
| Sample 48 | 0.76±0.32 | 2.11±0.54 | <LOQ | 2.87±0.86 |
| Sample 49 | 1.05±0.33 | 2.34±0.65 | <LOQ | 3.39±0.98 |
| Sample 50 | 1.15±0.64 | 2.11±0.08 | <LOQ | 3.26±0.72 |
| Sample 51 | 1.78±0.56 | 2.34±0.54 | <LOQ | 4.12±1.10 |
| Sample 52 | 1.43±0.07 | 0.54±0.02 | <LOQ | 1.97±0.09 |
| Sample 53 | 1.32±0.56 | 0.87±0.06 | <LOQ | 2.19±0.62 |
| Sample 54 | 1.88±0.97 | 0.45±0.02 | <LOQ | 2.33±0.99 |
| Sample 55 | 1.32±0.54 | 0.34±0.08 | <LOQ | 1.68±0.62 |
| Sample 56 | 1.42±0.68 | 0.92±0.09 | <LOQ | 2.34±0.77 |
| Sample 57 | <LOQ | 1.05±0.45 | 1.24±0.07 | 2.29±0.52 |
| Sample 58 | <LOQ | 1.52±0.88 | 1.43±0.05 | 2.95±0.93 |
| Sample 59 | <LOQ | 2.11±0.32 | 1.25±0.68 | 3.36±1.00 |
| Sample 60 | <LOQ | 1.76±0.54 | 1.67±0.07 | 3.43±0.61 |
| Sample 61 | <LOQ | 2.01±0.66 | 1.54±0.16 | 3.55±0.82 |
| Sample 62 | <LOQ | 3.42±0.09 | 2.02±0.47 | 5.44±0.56 |
| Sample 63 | <LOQ | 2.11±0.78 | 2.33±0.87 | 4.44±1.65 |
| Sample 64 | <LOQ | 0.75±0.07 | 1.98±0.07 | 2.73±0.14 |
| Sample 65 | <LOQ | 0.41±0.08 | 1.67±0.54 | 2.08±0.62 |
| Sample 66 | 1.65±0.08 | 0.76±0.43 | <LOQ | 2.41±0.51 |
| Sample 67 | 1.23±0.05 | 0.88±0.02 | <LOQ | 2.11±0.07 |
| Sample 68 | 1.01±0.43 | 0.99±0.05 | <LOQ | 2.00±0.48 |
| Sample 69 | 2.45±0.09 | 1.05±0.23 | <LOQ | 3.50±0.32 |
| Sample 70 | 2.11±0.04 | 1.23±0.08 | <LOQ | 3.34±0.12 |
| Overall average | 0.78±0.20 | 1.40±0.29 | 1.01±0.27 | 3.19±0.76 |
1) All samples were replicated three times and are expressed with mean ± standard deviation. 2) <LOQ = less than lower limit of quantification.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



