
Submitted:
16 September 2023
Posted:
18 September 2023
You are already at the latest version
Abstract
The upsurge in and havoc being wrecked by antibiotic resistant bacteria makes urgent the need for efficacious alternatives to antibiotics. This study assessed the antibacterial efficacy of two isobutyl-cyanoacrylate nanoparticles (iBCA-NPs), D6O and NP30, against major bacterial pathogens of fish. In vivo tests on rainbow trout were preceded by tests of in vitro minimum inhibitory concentration (MIC) and minimum bactericidal concentration (MBC). NP30 exhibited higher efficacy, but both iBCA-NPs demonstrated dose-dependent and species-specific in vitro antibacterial properties against the bacterial isolates. Generally, Gram-negative bacteria were more resistant to the iBCA-NPs. Streptococcus iniae, Tenacibaculum maritimum, and Photobacterium damselae were particularly sensitive to both iBCA-NPs. Orally administered to rainbow trout, the iBCA-NPs produced relative gain rate and survival rates comparable to the control (P > 0.05). Condition factor, hepatosomatic and viscerosomatic indices of fish were indifferentiable (P > 0.05) between the iBCA-NP groups and the control. The iBCA-NPs caused no alteration in stress, oxidative stress (superoxide dismutase, SOD), plasma complement titre, and lysozyme activity. The results of this study suggest that D60 and NP30 may contribute to reducing the amount of antibiotics and chemotherapeutic agents being used in aquaculture.
Keywords:
antibacterial efficacy
; isobutyl-cyanoacrylate nanoparticles
; rainbow trout
; safety
1. Introduction
Aquaculture has become very important in increasing global access to fish [1,2], improving nutrition to the deprived populations of the world [3]. However, burden of infectious diseases has been identified as the greatest of all setbacks in the quest to sustainable global aquaculture growth [4,5], with losses exceeding 6 billion USD per annum [6]. More than 50 % of the most common devastating infectious diseases of fish are caused by bacteria [7,8].
In response, frequent and indiscriminate use of antibiotics have characterized aquaculture in many countries [9,10,11]. The consequences have been antibiotic resistance in several bacteria strains resulting in high numbers of diverse bacteria in aquacultural systems [12]. As a result, treatments for some common fish bacterial diseases become less successful as it is reported for furunculosis and edwardsiellosis, increasing losses to these diseases. Also, some fish bacterial pathogens including Streptococcus iniae, and Photobacterium damselae, which are zoonotic, carry antimicrobial resistant genes like the extended-spectrum beta-lactamases. These bacteria, hence, the resistant genes are transferrable to humans through contact and food [13]. Due to these, some countries have strict regulations completely prohibiting the use of some and overall reduction in the application of chemotherapeutic drugs in aquaculture [14,15]. Vaccines and probiotics are considered safer alternatives in preventing diseases in aquaculture but there remain difficulties [16,17]. The use of nanoparticles of high antimicrobial ability is considered the most modern, advanced, and promising approach to fish disease control in aquaculture [18]. Several metal nanoparticles have been found to exhibit strong antibacterial properties [19]. However, bioaccumulation in fish flesh, gene toxicity and induction of oxidative to fish have raised serious concerns about their use in aquaculture [20,21,22]. Preference is growing for nanoparticles which are biodegradable, easy to prepare and with no or very minimal toxicity threats [23]. Poly (alkyl cyanoacrylate) (PACA) have been used in medicine as adhesives and its nanoparticles as drug carriers for several decades [24,25,26]. However, antibacterial property and hence suitability of cyanoacrylate nanoparticles as antibiotic agents are recent discoveries [27]. Their advantages, including easy fabrication and functionalization, biocompatibility, and biodegradability, make them promising candidates for disease control including fish disease management in the aquaculture industry [26,28]. Cyanoacrylate nanoparticles have high affinity for the glycopeptides that make up the cell walls of bacteria. They attach to the cell walls, induce distortion of cell wall synthesis, leading to autolysis and lysis by mechanisms detailed by Shirotake [23]. Widyaningrum et al. [29] also reported a similar interaction between isobutyl cyanoacrylate nanoparticles, iBCA-NPs, and algae. Thus, unlike antibiotics, iBCA-NPs, like other nanoparticles, employ physical action against pathogens, hence, it is believed that the possibility of emergence of resistant bacteria is reduced. The antibacterial activity of cyanoacrylate nanoparticles remains a new research area. As much as we know, the application of cyanoacrylate nanoparticles in disease control in the field of fisheries has not yet been considered by anyone else. In this study, we investigated the in vitro antimicrobial activity of two iBCA-NPs (NP30 and D60) against twelve prominent fish disease pathogens. We also assessed the selection preference and safety of D60 and NP30 for the land-based aquacultural production of rainbow trout, [Onchorhyncus mykiss (Walbaum, 1792)], an important aquaculture species, when administered through feed.
2. Materials and Methods
2.1. Isobutyl Cyanoacrylate Nanoparticle Preparation
Two isobutyl cyanoacrylate nanoparticles, iBCA-NPs, (D60 and NP30) of different sizes prepared from isobutyl cyanoacrylate monomers were used in this work. They were prepared in the laboratories of Chikami Miltec Inc. (Kochi, Japan) and are part of a collection of patented products of the inventors (JP-A-2016-56481; JP-A-2017-81852). Their preparation is detailed in Figure 1, following the methods described by Widyaningrum et al. [29] with slight modifications. Briefly, monomer solutions were added dropwise into 0.2 % (v/v) 5 mol/L HCl in respective dispersants until the monomer reached 1.10 % (v/v) of the HCl solution under continuous stirring at 600 rpm at 25 °C. In the case of D60, 1.0% (w/v) dextran 60,000 (041-30525; Wako Pure Chemical Industries, Osaka, Japan) served as the dispersant. On the other hand, 1.25% (v/v) of non-ionic surfactant RHEODOL TW-L (Kao Corporation, Tokyo, Japan) and 1.0% (v/v) of anionic surfactant NEOPELEX G-15 (Kao Corporation, Tokyo, Japan) were the dispersants for NP30. After stirring for 2 h to polymerize and prevent particles agglomeration, 0.2 M NaOH was added drop wisely until the pH reached 7.0. The mixtures were stirred for an additional hour to generate solid nanoparticles. The solution was filtered to remove any debris in the solution, if present, using a 5 μm pore size Minisart® (2-5322-04-30; Sartorius, Goettingen, Germany). The synthesized iBCA-NPs were stored at 2 – 5 °C until further use.
2.1.1. Characterization of the Synthesized iBCA-NPs
Dispersed diameter values for each iBCA-NP were measured immediately after production using the dynamic light scattering (DLS) method. The final concentration of the iBCA-NPs was 10 mg/mL. The characterization analyses were conducted in triplicate on iBCA-NP suspensions. D60 was diluted by a factor of ten (10) for characterization. The effective surface charge (zeta potential) was determined via the Electrophoretic light scattering method, while particle size and distribution, and polydispersity index (PDI) of the particles were measured using the Dynamic light scattering (DLS) method with a Zetasizer (Nano ZS90; Malvern Instruments, Malvern, UK).
2.2. Bacterial Pathogens
Aliquots of twelve important fish pathogenic bacterial isolates from eight different genera (Aeromonas, Edwardsiella, Nocardia, Lactococcus, Photobacterium, Streptococcus, Tenacibaculum and Vibrio) were tested for their sensitivity to NP30 and D60. These bacterial pathogens were isolated from infected fish in different regions of Japan. All, except Tenacibaculum maritimum, were cultured in Brain-Heart Infusion nutrient broth (BHI, Difco Becton, Dickinson, and Company, USA). T. maritimum was grown in marine broth (Difco, Becton, Dickinson, and Company, USA). They were then cryopreserved at -80°C in equal volumes of their respective broths and of 40% glycerol at the Laboratory of Cell Structure and Function, Division of Marine Bioresource Science, Graduate School of Kuroshio Science, Kochi University, Nankoku, Kochi, Japan. Identification of the bacteria species included macroscopic examination of the pathological signs of infected fish, characteristic morphology on nutrient agar, Gram-staining and subsequent molecular confirmation.
Molecular identification of all the bacterial isolates was executed using 63f (5’-CAGG CCTAACACATGCAAGTC-3`) and 1387r (5’-CGGCGGWGTGTACAAGGC-3`) bacterial 16S rRNA primers [30] following Ahmed et al. [31] with minor modifications. Briefly, amplified PCR products were purified using EXOSAP-IT (Thermo Fisher Scientific) and then subjected to sequencing. Each nucleotide sequence was queried against the NCBI GenBank database by BLASTn [32] and homologous sequences with ≥ 97% similarity selected for identification of each bacteria isolate.
2.2.1. Bacterial Culture Medium for MIC Assay
The bacteria isolates included the typical and atypical Edwardsiella tarda strains [33,34], Nocardia seriolae, Lactococcus garvieae, Photobacterium damselae subsp. piscicida, Streptococcus iniae, T. maritimum and four Vibrio spp. (V. anguillarum, V. harveyi, V. parahaemolyticus and V. rotiferanus). All bacteria strains, except T. maritimum, were cultured in Tryptic Soy Broth, TSB, (Bacto, Becton, Dickinson, and Company, USA) and incubated in a shaking incubator at 100 rpm except for A. salmonicida subsp. salmonicida (120 rpm) and N. seriolae (180 rpm). Incubation temperature was 25oC, except for the two E. tarda strains (28oC), 2 % NaCl for all except for the two E. tarda strains, and A. salmonicida subsp. salmonicida, (where NaCl = 0.5 %). T. maritimum was cultured in marine broth (Difco, Becton, Dickinson, and Company, USA), incubated at 25oC, 180 rpm. The growth conditions for the bacterial strain were in accordance with the Revised Standard Methods of the Japanese Society of Antimicrobials for Animals (JSAA), for antimicrobials test against bacteria isolated from Animals [35].
2.3. MIC Measurement by Micro Liquid Dilution Method
Susceptibility tests were carried out by the standard broth microdilution method according to the performance standards for antimicrobial susceptibility testing by the Clinical Laboratory Standards Institute [36]. The minimum inhibitory concentration (MIC) of the iBCA-NPs were evaluated spectrophotometrically by the turbidimetric method. The bacteria, except T. maritimum, were cultured in tryptic soy broth (TSB) overnight (or longer, 96 hours for N. seriolae). T. maritimum was cultured in marine broth and incubated for 48 hours. 100 μl of 10 mg/ml of each, NP30 and D60, were double-fold serially diluted up to at least 0.078 mg/ml in 96-well plates under aseptic conditions and inoculated with bacterial inoculums adjusted to 1.0×105 CFU/ml. Blank controls consisted of 100 μl inoculum-free fresh broth + iBCA-NPs mixture while inoculated 100 μl nanoparticle-free fresh broth served as the positive control. The method was slightly modified for N. seriolae. The cells of N. seriolae form large aggregates in broth and strongly stick to surfaces like the 1.5 Eppendorf tubes and pipette tubes making it difficult to successfully inoculate using the CLSI (2019) prescribed method. Therefore, for this bacterial, the MIC assay used was 100 μl of bacterial inoculums adjusted to 1.0×105 CFU/ml in fresh broth inoculated into 100 μl of iBCA-NPs serially diluted with sterilized distilled water (total assay volume = 200 μl). The plates were incubated in a static incubator at recommended standard conditions of temperatures and time as indicated above. Optical densities at 620 nm (OD620) absorbance were measured for each MIC assay before and after incubation. If the MIC assay was more turbid, per OD620, after incubation, bacterial growth was implied, while lower or similar OD620 values as before incubation, indicated growth inhibition [37]. The lowest concentration at which each iBCA-NP inhibited at least 80 % growth of the test bacteria was considered the related MIC value [38], deduced from plots as exemplified in Figure 2. To assess the source of antibacterial activity, and role of the dispersants, the procedure was also executed with the dispersants of each iBCA-NP. Each assay was performed in triplicate, and the result that repeated twice or more was considered [31,39]. Percentage bacterial growth inhibition was computed using the formula:
The GI % for each bacteria strain at each concentration of each iBCA-NPs were determined and expressed as mean percentages ± standard error of the mean (SEM) in bar graphs showing the dose- dependent and species-specific antibacterial efficacy of each iBCA-NPs (Figure 3, Figure 4 and Figure 5).
2.3.1. Minimum Bactericidal Concentration (MBC) Assay
The MBC corresponding to each sensitive bacteria was the lowest concentration of each iBCA-NPs, which inhibited 99.9% of the bacterial growth on suitable growth agar plates of the bacteria strain [40,41]. This was determined by plating 50 μl of the bacteria-iBCA-NP mixture at the MIC and higher concentrations on the suitable nutrient agar for each. The plates were then incubated in a static incubator at recommended conditions as stated above. Absence of bacterial colonies on plates signified the lack of living bacteria and the bactericidal effect at that concentration. On the other hand, growth of bacterial colonies on plates indicated the presence of living bacteria and hence bacteriostatic effect at that concentration. The assay was performed in triplicate, and results repeated twice or more considered.
2.4. Selection Preference and Growth Performance of iBCA-NPs-Fed Rainbow Trout
In vivo tests are the ultimate determinants of the relevance of the iBCA-NPs, even if found to be efficacious in vitro. Long chain PACA and its nanoparticles are considered safe to body tissues and can be efficiently eliminated from the body by excretion [25,26,27]. We also found that although passive swimmers like the olive flounder (Paralichthys olivaceus) fared well, for active swimmers like the rainbow trout, immersion in the iBCA-NPs resulted in fatalities due to clogging except at very low concentrations (unpublished data). We therefore tested the selection preference, growth performance and safety of NP30 and D60 when administered orally through feed to rainbow trout. The basal diet was EP commercial feed grade 4 (Nisshin Marubeni Feed Co., Japan) for rainbow trout. Rainbow trout fry raised at a seed and nursery center in Ehime Prefecture were transported to the Freshwater Experiment Site of the Kochi University (Japan), where they were put on the EP commercial feed (Nisshin Marubeni Feed Co., Japan) in a 1.1-ton fiberglass reinforced plastic (FRP) tank until the experiments begun.
2.4.1. Experimental Design
The experimental setup included six flow-through 500 L fiberglass reinforced plastic (FRP) tanks demarcated into three groups, two replicates each (n = 2). Each tank contained 45 fish (mean weight 35.08 ± 0.46 g, mean length 14.18 ± 0.09 cm). The iBCA-NP groups (NP30 and D60 groups) were fed EP Feed supplemented with the respective iBCA-NP solutions at the maximum amount that could be added to the feed which was determined to be 3,571.4 mg(iBCA-NP)/kg feed weight. The control groups were fed EP Feed mixed with same volume of distilled water (as the iBCA-NPs). The fish were acclimatized in their respective new tanks on iBCA-NP-free basal feed for two weeks before the start of treatments. Water used was degassed and well aerated and water volume kept at 400 L for each tank. Tanks were maintained under natural light/day regimes. Water quality parameters were frequently recorded throughout the period of the study where rate of water exchange was 85.5 % per hour per tank, mean temperature was 18.56 ± 0.01 oC, PH (8.38 ± 0.17) and DO (9.80 ± 0.21 mgl-1). Fish in each tank were fed to satiation twice daily (morning and evening) for the period of the study (12 weeks). Daily feed intake per tank and feeding behavior were recorded.
2.4.2. Sampling
Five fish samples were randomly taken from each tank every 28-day and anesthetized with a lethal dose of 2-phenoxyethanol (Wako). 1ml blood from the caudal vein of each fish sample was aliquoted into labelled 1.5 ml Eppendorf tubes (Wako) using a heparinized syringe, and plasma obtained via centrifugation at 3,000 x G for 15 min at 4°C. The blood plasma collected was frozen at -80°C until use. Morphometrics (weight and length) were also measured for all groups every 28 days.
2.4.3. Growth Performance
The following equations were used to calculate the growth performance parameters:
2.4.4. Blood Plasma Lysozyme Activity and Complement Titer
Lysozyme activity in blood plasma was examined via the turbidimetric assay and lysis of the lysozyme-sensitive Gram-positive bacterium Micrococcus lysodeikticus (Sigma) according to [42]. For complement titer, sample plasma and 25 µl of sheep red blood cells (Japan Lamb) were suspended and allowed to react for 1 hour at room temperature. After completion of the reaction, 50 µl of EGTA, Mg2+, GGVB (glucose gelatin veronal buffer with 10 mM glycol ether diamine tetra acetic acid and 40 mM MgCl2) was added, and the mixture was centrifuged at 4°C for 3,000 × G for 3 minutes. 70 µl of the supernatant was added to a 96-well plate, and the absorbance value at 493 nm wavelength measured with a microplate reader (Thermo Scientific Multiskan FC). Complement titer, y, was computed using the following formula:
Complement titer y = {absorbance value of sample - CB (mechanical hemolysis, 25 µl of red blood cells + 75 µl of buffer solution) / (absorbance value of 100% hemolysis - absorbance value of CB) } × 100
The results were plotted to assess the comparative status of each group.
2.4.5. Stress and Oxidative Stress Biomarkers
Pollutants have significant effects on fish condition factor, state of fish liver and the viscera of fish [43,44,45]. We estimated the condition factor, hepatosomatic index and viscerasomatic index of fish in each treatment to ascertain fish health and condition. Indications of possible acute stress consequent to the iBCA-NP treatments were investigated via glucose concentration, which was found to rise with rising cortisol levels in response to stress in fish [46,47]. The glucose concentration in blood sera was measured at OD493 using the Mutarotase-GOD method according to Miwa et al. [48] via a commercially available Glucose C-II kit following (FUJIFILM Wako Pure Chemical). Superoxide dismutase (SOD) activity in sera was spectrophotometrically estimated at 493 nm by the xanthine oxidase method using the SOD Assay Kit- WST (Dojindo, Japan) following the manufacturer guideline.
2.5. Statistical Analysis
Results of the antimicrobial tests were in triplicates and presented as mean ± standard error of the mean (SEM, n = 3). Data from the in vivo growth test were subjected to statistical verification via using one-way ANOVA tests at 95 % confidence level. Tukeys posthoc test were used to confirm differences where differences occurred within the experimental groups. Data which violated the parametric assumptions were analyzed using the Welch F-test, Brown and Forythe and Games-Howell non-parametric tests. All statistical tests were performed using the IBM SPSS for WINDOWS (version 29.01.0).
3. Results
3.1. Physicochemical Characteristics of iBCA-NPs
Analysis of the zeta potential of the two iBCA-NPs showed that both D60 and NP30 were negatively charged (NP30 = -20 ~ -25 mV; D60 = -5 ~ -10 mV) There was no significant difference between concentrations. DLS showed that mean diameter of the iBCA-NPs ranged 150 – 200 nm for D60 and 20 – 40 nm for NP30. PDI and size distribution analyses showed that particles of both D60 and NP30 were largely monodisperse, with a uniform and rather narrow size distribution [PDI(D60) = 0.15 to 0.45; PDI (NP30) = 0.05 – 0.30].
3.2. Antibacterial Effect of iBCA-NPs
3.2.1. Minimum Inhibitory Concentration (MIC)
Both isobutyl cyanoacrylate nanoparticles, NP30 and D60, exhibited species-specific dose-dependent antibacterial activity against the tested fish bacteria pathogens (Table 1). However, NP30 was more efficacious, showing antibacterial activity against all twelve tested bacteria strains (Table 1; Figure S1). D60 was efficacious against 83.33 % (n = 10) of the strains. 33.33 % (n = 4) of the bacteria strains [S. iniae and L. garvieae (Gram-positive), P. damselae, and T. maritimus (Gram-negative)] were the most sensitive species (MIC = 78 or 156 µg/ml for both iBCA-NPs). Three Vibrio spp. (V. harveyi, V. parahaemolyticus, and V. rotiferanus) demonstrated the highest resistance. No MIC was obtained for D60 against V. harveyi, and V. parahaemolyticus. From our results, the dispersants showed no significant antibacterial activity against any of the bacteria isolates (Figure 2 & Figure S1).
3.2.2. Minimum Bactericidal Concentration (MBC)
This referred to the lowest concentration of each iBCA-NPs at which no bacteria growth was observed on nutrient agar plates. NP30 was bactericidal against all tested bacterial isolates while D60 was bactericidal against only two isolates, namely, T. maritimum and V. anguillarum. The antibacterial activity of D60 was thus dominantly bacteriostatic (Table 1). T. maritimum recorded the lowest MBC for both iBCA-NPs.
3.3. Growth Performance and Safety Evaluations in Rainbow Trout
The iBCA-NPs had no adverse effect on water quality parameters throughout the period of the studies. The following are the parameters taken: mean temperature was 18.56 ± 0.01 oC, PH (8.38 ± 0.17), DO (9.80 ± 0.21 mgl-1), total ammonium (0.42 ± 0.05 mgl-1), nitrite (0.02 ± 0.003 mgl-1) and nitrate (0.35 ± 0.04 mgl-1). Survival rate was highest (100 %) in the NP30 group but not significantly different (P > 0.05) from the others (98.89 %). Feeding aggression was relatively higher in the iBCA-NP groups than in the control. After 12 weeks of feeding, rainbow trout fed with iBCA-NPs (both NP30 and D60) supplemented feed showed relative gain rate, specific growth rate, and feed conversion ratio as good (P > 0.05) as fish in the control group, fed the commercial EP feed only (Table 2). Relative gain rate was steeply high for the first month but significantly declined (P < 0.001) in subsequent months for all groups. (Figure 7a). Generally, NP30-treated tanks showed slightly better growth performance, but there was no significant difference among the treatments (Table 2; P > 0.05). At the end of 8 weeks, feed conversion ratio (FCR) was significantly lower (P < 0.05; not shown) in the iBCA-NPs groups than in the control. However, FCR was not differentiable (P > 0.05) among treatments at the end of the study.
3.4. Innate Defense Capability
Macroscopic physical examinations of fish during each sampling time suggested healthy growth in all treatments. Lysozyme activity increased with age of rainbow trout. Lysozyme activity after the first four weeks were significantly lower (P < 0.05) than activity rate after the twelfth week (Figure 7b). The control group had the highest lysozyme activity (0.092 ± 0.031 U/ml) at the end of the eighth week but declined steadily to be the least (0.072 ± 0.01 U/ml) at the end of this study. On the other hand, lysozyme activity of the D60 group was the least (0.017 ± 001 U/ml) at the at first sampling time but consistently steadily increased to become the highest (0.089 ± 0.025 U/ml) at the end of the 12th week (Figure 7b). However, there was no significant difference among the treatments at any sampling time (P > 0.05)
Similarly, complement titer values were significantly lower (P < 0.05) for all groups (mean = 0.174 ± 0.02 U/ml) at the end of the 4th week than after 8 weeks (mean = 0.342 ± 0.033 U/ml). The control groups had the least complement titer after the first month (mean 0.158 ± 0.017 U/ml) while NP30 showed the highest rate of hemolysis at both sampling times, reaching 0.408 ± 0.023 U/ml at the end of eight weeks. However, there was no significant difference in rate of hemolysis among treatments at any sampling time (P > 0.05).
3.5. Stress-Related Biomarkers
Condition factor, hepatosomatic and viscerosomatic indices were consistently similar among groups and not significantly different (P > 0.05; Table 2). At the end of the first four weeks, glucose concentration was slightly higher in the D60 group, but at the end of the study, concentrations were strikingly similar for all treatments (Figure 7d). Overall rate of SOD activity was slightly higher in the iBCA-NP groups (control = 67.64 ± 0.37; D60 = 68.60 ± 0.53; NP30 = 69.84 ± 0.43 %) but there was no significant difference among treatments, including among replicates (Figure 7e). However, we found one fish with a severe internal tumor in a NP30 tank (Figure 7f).
4. Discussion
The characteristics of the cyanoacrylate nanoparticles, iBCA-NPs, (D60 and NP30) were very similar to iBCA-NPs (25 nm and 180 nm), respectively, described by Widyanigrum et al. [29]. This is because, in both instances, the monomers used were the same, and reagents and methods of formulation were very similar.
From the results of the antibacterial and bactericidal activity examined in this study, the isobutyl iBCA-NPs we studied demonstrated strong antibacterial activity against all the bacterial pathogens we examined (Table 1). NP30 was bactericidal against 100 % of the pathogens while D60 was bacteriostatic against most of them (Table 1). Shirotake, [27] reported high affinity of iBCA-NPs for glycoproteins and junctioning between iBCA-NPs and Gram-positive bacteria resulting in bacterial wall autolysis. The dispersant types played very important roles in differentiating between the sizes of the two iBCA-NPs and in preventing their agglomeration consistent with Guzman et al., [49]. However, the dispersants did not interfere with the antimicrobial effect of our iBCA-NPs, as none showed significant antibacterial against the bacterial strains. That is, from our results, the iBCA-NPs, not the dispersants, were the source of the antibacterial activities observed (Figure 2 & S1). Widyaningrum et al. [29] reported a similar finding, where the dispersants had no effects on algal cell walls. The massive global aquacultural losses to these bacteria species are well documented [50,51,52,53,54,55,56,57,58]. The cost of using antibiotics to control these aquacultural pathogens is enormous [11] and it will require several different types of vaccines to combat them. The importance of the efficacy of a single safer antimicrobial agent, iBCA-NP, to manage several bacterial strains cannot be overemphasized.
The comparatively higher antimicrobial activity of NP30 might be due to its minute particle size (mean size = 30 nm), increasing the surface area for attachment to the bacterial pathogens. This assertion agreed with Widyaningrum et al. [29] who found an inverse relationship between iBCA-NPs size and algal cell mortality at any given concentration. D60 particles were larger (mean size = 180 nm) resulting in a more viscous colloid (Figure 1b). The superior performance of NP30 might also be explained by the negative electrical charge effect between NP30 particles induced by its anionic dispersant. This effect is thought to keep particles uniformly distributed in solution and in constant motion, increasing contact with bacteria cells. The nanoparticle-size-antibacterial activity relationship has been widely studied. Consistent with our results, generally, the smaller the size, the higher the activity [59,60,61].
The presence of a lipopolysaccharide outer membrane and chromosomally encoded drug efflux mechanisms in Gram-negative bacilli is considered to confer comparative higher antibacterial resistance on them than on Gram-positive bacteria [62,63,64]. Consistent with this, Gram-negative bacteria were more resistant to our iBCA-NPs (Table 1). Importantly, although susceptibility of several Gram-positive bacteria, including multiple antibiotic resistant bacteria, to iBCA-NPs has been reported, all previous work reported no antibacterial activity of iBCA-NPs on Gram-negative baccilli [27,65]. The extreme sensitivity of P. damselae, and T. maritimus and indeed the susceptibility of other Gram-negative bacilli we tested was thus an interesting result. It is possible that differences in the iBCA-NPs preparation method and bacterial strains used accounted for the variations in results as it is known that differences in structure and composition between N-acetylglucosamine and N-acetylmuraminic acid may account for differences in antimicrobial resistance among Gram-negative bacteria [66,67]. The antimicrobial resistance of Aeromonas spp. and Vibrio spp., consistent with our results, are well documented [31,68,69,70,71,72,73]. On the other hand, although differences have been demonstrated in the mechanism of virulence and pathogenicity of the typical and atypical E. tarda strains [33,34], their antimicrobial susceptibility and resistance to both iBCA-NPs were very similar (Table 1). Their structural indistinguishability and molecular closeness might have accounted for the observation [74,75].
We observed increasing GI % with decreasing concentration for NP30 against T. maritimum (Figure 5 c) which we initially thought to be the display of the Eagle effect [38]. Divya et al., [76] reported similar results, where lower, rather than higher, concentrations of some of their chitosan nanoparticles showed higher growth inhibition. However, bactericidal properties were observed at both the higher NP30 concentrations (500 µl) and lower concentrations (until 31.25 µl). T. maritimum is a filamentous bacterium and possesses rhizoids with which gliding colonies adhere one-to-another creating a huge network of bacteria mass [54]. On the other hand, the iBCA-NPs assay transparency increased (decreasing OD620 value) by dilution. The effect of bacteria lysis and assay transparency on OD reading was suspected to have accounted for the inconsistency observed. As much as we know, this study presents the lowest concentrations of iBCA-NPs at which antibacterial activities have been observed [65]. If the in vitro efficacy of our iBCA-NPs is replicated in vivo, their relevance in the disease-burdened aquacultural industry may be enormous.
Growth performance and safety evaluation in rainbow trout: The treatments did not have adverse effects on water quality parameters [77] throughout the period of the study. In vitro, as well as in vivo, medium molecular weight cyanoacrylate nanoparticles like iBCA-NPs significantly degrade in 24 hours [24,25,28]. This, coupled with high rate of water exchange (85.5 % h-1) might have played a role in maintaining stable water quality parameters despite the heavy feed ration.
The growth test lasted for twelve weeks; a duration we considered long enough to reveal any effects the iBCA-NPs might have on rainbow trout. During this duration, rainbow trout were consistently fed with iBCA-NPs-supplemented feed at the highest possible concentration (3,571.4 mg/kg feed). At high concentrations, several metallic nanoparticles have been found to be toxic to fish, killing fish [47]. Growth inhibition was also reported in Cyprinus carpio by copper nanoparticles after thirty days of exposure [78]. On the contrary, growth performance was excellent in the iBCA-NPs groups as in the control and sometimes slightly better, but not significantly higher (P > 0.05), than the latter. The comparatively significant decline in gain rate (Figure 7a) after the first four weeks was consistent with documented growth trends of rainbow trout. Feed conversion efficiency is reportedly lower in older rainbow trout [79] which hypothesis was consistent with our results (results not shown). Fish condition factor, K, hepatosomatic and viscerosomatic indices (HIS & VSI respectively) are important indicators of fish health, metabolism and of energy reserves in fish [43,44,45]. The K, HSI and VSI values we obtained in this study are similar to the values reported for healthy fish in similar studies [80,81]. Therefore, like the growth performance parameters, these indices also suggested no toxic effects of the iBCA-NPs (Table 2).
Both the complement system and lysozymes are important components of the first barrier of fish against disease pathogens and are also important growth inhibition factors of bacterial agents. They also possess lytic activity against viruses and both Gram-positive and Gram-negative bacteria [82,83,84,85]. In addition, the complement system of fish is critical in the modulation of the adaptive immune response and has chemotaxic, opsonization, and pro-inflammatory functions, making it one of the most important factors of fish innate immune system [82,83,86]. Increased alternative complement hemolysis of foreign objects like sheep erythrocytes are indicative of improved immune response [82]. While the relationship between the level of lysozyme activity and immunity in fish remains unclear, generally, long term exposure to toxins and immunosupressants have decreasing effects on it. On the other hand, immunostimulants and dietary supplements like prebiotics are known to be associated with increasing lysozyme levels and fish complement titer, conferring protection against a variety of fish diseases [83,84,85]. In this study, lysozyme activity was significantly higher (P < 0.05) in the twelfth week than fourth week (Figure 7b). Similarly, complement titer was significantly higher in the eighth week than the fourth week (Figure 7c). Generally, the immune indicators increased with age suggesting improved immunity of rainbow trout with age, irrespective of treatment. This agrees with Sauer et al. [87] that age improves, not decline, fish health. Considering both indicators therefore, iBCA-NPs had no adverse effects on rainbow trout in this study.
The major concerns about nanoparticle use in aquaculture include, among others, oxidative damage to fish and impairment of many biomarkers. Metallic nanoparticle-induced oxidative stress has been reported by several publications [22,47,88,89,90,91]. Similar damages in fish have been reported in the use of carbon-based and green synthesized cadmium nanoparticles [92,93]. These damages have been proven to correlate with increased acute stress, signified by increased concentrations of blood cortisol and glucose in the Nile tilapia and the African catfish [47,93]. Toxic effects are fought off by fish by increased rates of glycolysis, resulting in increased glucose concentration [93]. In the current study, glucose concentration for all groups was consistently optimal (mean = 60.02 ± 0.76 mg/dl) [46,47,93] which meant that at 3,571.4 mg(iBCA-NP)/kg feed, the iBCA-NPs did not induce stress in rainbow trout. Compared to the liver, blood plasma estimates of oxidative stress are more accurate because the former has high activity of antioxidant enzymes to adapt to, and cope with oxidative stress [90]. Persistent or high oxidative stress may cause notable damages to proteins, decreasing SOD activity [90]. On the contrary, plasma SOD activity also confirmed that the SOD antioxidant defense systems of iBCA-NPs treated rainbow trout were as good as fish in the control group (Figure 7d).
A couple of pathways have been predicted for orally administered poly (alkyl cyanoacrylate), nanoparticles, PACA-NPs. According to Vauthier et al. [26] translocation via the Peyer’s patches in the ileum is the major pathway for drug-carrier PACA-NPs. Catalyzed by esterases from serum, lysosomes, and pancreatic juice, they are eliminated from the body by the kidneys in the forms of alkyl alcohol and poly-cyanoacrylic acid. Unlike shorter-chain derivatives, butyl derivatives and higher homologs of PACA are considered non-toxic to tissues [25] and safer than any other antimicrobial substances [27]. In agreement, from results of the current study, at 3,571.4 mg(iBCA-NP)/kg feed, iBCA-NPs produced excellent growth and innate immunity of rainbow trout was not compromised.
5. Conclusion
D60 and NP30 demonstrated high in vitro bactericidal and or bacteriostatic properties against both the Gram-positive and Gram-negative bacterial pathogens of fish diseases we tested. In vivo, we confirmed the safety of D60 and NP30 for the culture of fish by land-based aquaculture using rainbow trout, an industrially important salmonid species. It is expected that these results will be replicated irrespective of the in other systems of production. In vitro efficacy of antimicrobials may not always be replicated in vivo. Given that the in vitro tests have confirmed the antibacterial activity of the iBAC-NPs, with favorable selection by fish without any significant health implications, we would like to conduct tests to investigate its preventive and therapeutic effects in the future.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, M.G.A., K.M., M.H., K.I., F.B. and S-I.O; Methodology, M.G.A., K.M., M.H., K.I., F.B. and S-I.O; Software, M.G.A., K.I. and F.B.; Validation, M.G.A., K.M., M.H., K.I., F.B. and S-I.O; Formal Analysis, M.G.A., K.M., and M.H.; Investigation, M.G.A., K.M., M.H. and S-I.O.; Resources, M.G.A., K.I., F.B. and S-I.O; Data Curation, M.G.A., K.M., M.H. and S-I.O.; Writing – Original Draft Preparation, MGA.; Writing – Review & Editing, M.G.A., K.M., M.H. and S-I.O.; Supervision, S-I.O.; Project Administration, M.G.A., K.M., M.H., K.I., F.B. and S-I.O.; Funding Acquisition, M.G.A., K.I., F.B. and S-I.O.
Data Availability Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Acknowledgments
We express profound gratitude to the Japanese Government for the PhD Scholarship awarded Gabriel Mawuko Ahiable through MEXT. This work was also supported by JSPS Grant-in-Aid for Scientific Research C. Finally, we are very thankful to Chikami Miltec Inc., Kochi, Japan, for the supply of the nanoparticles for this work.
Conflicts of Interest
The authors declare that there is no conflict of interest.
References
- Cohen, J.T.; Bellinger, D.C.; Connor, W.E.; Kris-Etherton, P.M.; Lawrence, R.S.; Savitz, D.A.; Shaywitz, B.A.; Teutsch, S.M.; Gray, G.M. A Quantitative Risk-Benefit Analysis of Changes in Population Fish Consumption. Am. J. Prev. Med. 2005, 29, 325–334. [Google Scholar] [CrossRef] [PubMed]
- Pradeepkiran, J.A. Aquaculture Role in Global Food Security with Nutritional Value: A Review. Transl. Anim. Sci. 2019, 3, 903–910. [Google Scholar] [CrossRef] [PubMed]
- Garlock, T.; Asche, F.; Anderson, J.; Ceballos-Concha, A.; Love, D.C.; Osmundsen, T.C.; Pincinato, R.B.M. Aquaculture: The Missing Contributor in the Food Security Agenda. Glob. Food Secur. 2022, 32, 100620. [Google Scholar] [CrossRef]
- Sebastião, F. de A.; Lemos, E.G.M.; Pilarski, F. Validation of Absolute Quantitative Real-Time PCR for the Diagnosis of Streptococcus Agalactiae in Fish. J. Microbiol. Methods 2015, 119, 168–175. [Google Scholar] [CrossRef] [PubMed]
- Stentiford, G.D.; Sritunyalucksana, K.; Flegel, T.W.; Williams, B.A.P.; Withyachumnarnkul, B.; Itsathitphaisarn, O.; Bass, D. New Paradigms to Help Solve the Global Aquaculture Disease Crisis. PLoS Pathog. 2017, 13, e1006160. [Google Scholar] [CrossRef]
- Assefa, A.; Abunna, F. Maintenance of Fish Health in Aquaculture: Review of Epidemiological Approaches for Prevention and Control of Infectious Disease of Fish. Vet. Med. Int. 2018, 2018, 5432497. [Google Scholar] [CrossRef]
- Kibenge, F.S.B.; Godoy, M.G.; Fast, M.; Workenhe, S.; Kibenge, M.J.T. Countermeasures against Viral Diseases of Farmed Fish. Antiviral Res. 2012, 95, 257–281. [Google Scholar] [CrossRef]
- Surachetpong, W.; Roy, S.R.K.; Nicholson, P. Tilapia Lake Virus: The Story so Far. J. Fish Dis. 2020, 43, 1115–1132. [Google Scholar] [CrossRef]
- Dawood, M.A.O.; Koshio, S.; Abdel-Daim, M.M.; Van Doan, H. Probiotic Application for Sustainable Aquaculture. Rev. Aquac. 2019, 11, 907–924. [Google Scholar] [CrossRef]
- Van Doan, H.; Hoseinifar, S.H.; Ringø, E.; Ángeles Esteban, M.; Dadar, M.; Dawood, M.A.O.; Faggio, C. Host-Associated Probiotics: A Key Factor in Sustainable Aquaculture. Rev. Fish. Sci. Aquac. 2020, 28, 16–42. [Google Scholar] [CrossRef]
- Ventola, C.L. The Antibiotic Resistance Crisis: Part 1: Causes and Threats. P T Peer-Rev. J. Formul. Manag. 2015, 40, 277–283. [Google Scholar]
- Watts, J.E.M.; Schreier, H.J.; Lanska, L.; Hale, M.S. The Rising Tide of Antimicrobial Resistance in Aquaculture: Sources, Sinks and Solutions. Mar. Drugs 2017, 15, 158. [Google Scholar] [CrossRef] [PubMed]
- Bondad-Reantaso, M.G.; MacKinnon, B.; Karunasagar, I.; Fridman, S.; Alday-Sanz, V.; Brun, E.; Le Groumellec, M.; Li, A.; Surachetpong, W.; Karunasagar, I.; et al. Review of Alternatives to Antibiotic Use in Aquaculture. Rev. Aquac. n/a. [CrossRef]
- Abdelkhalek, N.K.M.; Eissa, I.A.M.; Ahmed, E.; Kilany, O.E.; El-Adl, M.; Dawood, M.A.O.; Hassan, A.M.; Abdel-Daim, M.M. Protective Role of Dietary Spirulina Platensis against Diazinon-Induced Oxidative Damage in Nile Tilapia; Oreochromis Niloticus. Environ. Toxicol. Pharmacol. 2017, 54, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Adel, M.; Dadar, M.; Khajavi, S.H.; Pourgholam, R.; Karimí, B.; Velisek, J. Hematological, Biochemical and Histopathological Changes in Caspian Brown Trout (Salmo Trutta Caspius Kessler, 1877) Following Exposure to Sublethal Concentrations of Chlorpyrifos. Toxin Rev. 2017, 36, 73–79. [Google Scholar] [CrossRef]
- Matsuura, Y.; Terashima, S.; Takano, T.; Matsuyama, T. Current Status of Fish Vaccines in Japan. Fish Shellfish Immunol. 2019, 95, 236–247. [Google Scholar] [CrossRef]
- Newaj-Fyzul, A.; Al-Harbi, A.H.; Austin, B. Review: Developments in the Use of Probiotics for Disease Control in Aquaculture. Aquaculture 2014, 431, 1–11. [Google Scholar] [CrossRef]
- Shaalan, M.; Saleh, M.; El-Mahdy, M.; El-Matbouli, M. Recent Progress in Applications of Nanoparticles in Fish Medicine: A Review. Nanomedicine Nanotechnol. Biol. Med. 2016, 12, 701–710. [Google Scholar] [CrossRef]
- Slavin, Y.N.; Asnis, J.; Hńfeli, U.O.; Bach, H. Metal Nanoparticles: Understanding the Mechanisms behind Antibacterial Activity. J. Nanobiotechnology 2017, 15, 1–20. [Google Scholar] [CrossRef]
- Khosravi-Katuli, K.; Prato, E.; Lofrano, G.; Guida, M.; Vale, G.; Libralato, G. Effects of Nanoparticles in Species of Aquaculture Interest. Environ. Sci. Pollut. Res. 2017, 24, 17326–17346. [Google Scholar] [CrossRef]
- Fajardo, C.; Martinez-Rodriguez, G.; Blasco, J.; Mancera, J.M.; Thomas, B.; De Donato, M. Nanotechnology in Aquaculture: Applications, Perspectives and Regulatory Challenges. Aquac. Fish. 2022, 7, 185–200. [Google Scholar] [CrossRef]
- Bacchetta, C.; Ale, A.; Simoniello, M.F.; Gervasio, S.; Davico, C.; Rossi, A.S.; Desimone, M.F.; Poletta, G.; López, G.; Monserrat, J.M.; et al. Genotoxicity and Oxidative Stress in Fish after a Short-Term Exposure to Silver Nanoparticles. Ecol. Indic. 2017, 76, 230–239. [Google Scholar] [CrossRef]
- Carlson, C.; Hussain, S.M.; Schrand, A.M.; Braydich-Stolle, L.K.; Hess, K.L.; Jones, R.L.; Schlager, J.J. Unique Cellular Interaction of Silver Nanoparticles: Size-Dependent Generation of Reactive Oxygen Species. J. Phys. Chem. B 2008, 112, 13608–13619. [Google Scholar] [CrossRef]
- Lenaerts, V.; Couvreur, P.; Christiaens-Leyh, D.; Joiris, E.; Roland, M.; Rollman, B.; Speiser, P. Degradation of Poly (Isobutyl Cyanoacrylate) Nanoparticles. Biomaterials 1984, 5, 65–68. [Google Scholar] [CrossRef] [PubMed]
- Leonard, F.; Kulkarni, R.K.; Brandes, G.; Nelson, J.; Cameron, J.J. Synthesis and Degradation of Poly (Alkyl α-Cyanoacrylates). J. Appl. Polym. Sci. 1966, 10, 259–272. [Google Scholar] [CrossRef]
- Vauthier, C.; Dubernet, C.; Fattal, E.; Pinto-Alphandary, H.; Couvreur, P. Poly(Alkylcyanoacrylates) as Biodegradable Materials for Biomedical Applications. Adv. Drug Deliv. Rev. 2003, 55, 519–548. [Google Scholar] [CrossRef]
- Shirotake, S. A New Cyanoacrylate Colloidal Polymer with Novel Antibacterial Mechanism and Its Application to Infection Control. J. Nanomedicine Biotherapeutic Discov. 2014, 04, 1. [Google Scholar] [CrossRef]
- Sulheim, E.; Baghirov, H.; von Haartman, E.; Bøe, A.; Åslund, A.K.O.; Mørch, Y.; Davies, C. de L. Cellular Uptake and Intracellular Degradation of Poly(Alkyl Cyanoacrylate) Nanoparticles. J. Nanobiotechnology 2016, 14, 1. [Google Scholar] [CrossRef]
- Widyaningrum, D.; Iida, D.; Tanabe, Y.; Hayashi, Y.; Kurniasih, S.D.; Ohama, T. Acutely Induced Cell Mortality in the Unicellular Green Alga Chlamydomonas Reinhardtii (Chlorophyceae) Following Exposure to Acrylic Resin Nanoparticles. J. Phycol. 2019, 55, 118–133. [Google Scholar] [CrossRef]
- Marchesi, J.R.; Sato, T.; Weightman, A.J.; Martin, T.A.; Fry, J.C.; Hiom, S.J.; Dymock, D.; Wade, W.G. Design and Evaluation of Useful Bacterium-Specific PCR Primers That Amplify Genes Coding for Bacterial 16S RRNA. Appl. Environ. Microbiol. 1998, 64, 795–799. [Google Scholar] [CrossRef]
- Ahmed, F.; Soliman, F.M.; Adly, M.A.; Soliman, H.A.M.; El-Matbouli, M.; Saleh, M. In Vitro Assessment of the Antimicrobial Efficacy of Chitosan Nanoparticles against Major Fish Pathogens and Their Cytotoxicity to Fish Cell Lines. J. Fish Dis. 2020, 43, 1049–1063. [Google Scholar] [CrossRef]
- Altschul, S.F.; Boguski, M.S.; Gish, W.; Wootton, J.C. Issues in Searching Molecular Sequence Databases. Nat. Genet. 1994, 6, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Matsuyama, T.; Kamaishi, T.; Ooseko, N.; Kurohara, K.; Iida, T. Pathogenicity of Motile and Non-Motile Edwardsiella Tarda to Some Marine Fish. Fish Pathol. 2005, 40, 133–135. [Google Scholar] [CrossRef]
- Nakamura, Y.; Takano, T.; Yasuike, M.; Sakai, T.; Matsuyama, T.; Sano, M. Comparative Genomics Reveals That a Fish Pathogenic Bacterium Edwardsiella Tarda Has Acquired the Locus of Enterocyte Effacement (LEE) through Horizontal Gene Transfer. BMC Genomics 2013, 14, 642. [Google Scholar] [CrossRef] [PubMed]
- Japanese Society of Antimicrobials for Animals The Determination Method of Minimum Inhibitory Concentration (MIC) of Antimicrobials against Bacteria Isolated from Animals (RevisedStandard Method of the Japanese Society of Antimicrobials for Animals in 2003). Proceeding Jpn. Soc. Antimicrob. Anim. 2003, 25, 63–73.
- Clinical and Laboratory Standards Institute Performance Standards for Antimicrobial Susceptibility Testing. CLSI Suppl. M100 Wayne PA CLSI 2019, 25.
- Vijayakumar, P.P.; Muriana, P.M. A Microplate Growth Inhibition Assay for Screening Bacteriocins against Listeria Monocytogenes to Differentiate Their Mode-of-Action. Biomolecules 2015, 5, 1178–1194. [Google Scholar] [CrossRef]
- Kowalska-Krochmal, B.; Dudek-Wicher, R. The Minimum Inhibitory Concentration of Antibiotics: Methods, Interpretation, Clinical Relevance. Pathog. Basel Switz. 2021, 10, 165. [Google Scholar] [CrossRef]
- Du, W.-L.; Niu, S.-S.; Xu, Y.-L.; Xu, Z.-R.; Fan, C.-L. Antibacterial Activity of Chitosan Tripolyphosphate Nanoparticles Loaded with Various Metal Ions. Carbohydr. Polym. 2009, 75, 385–389. [Google Scholar] [CrossRef]
- Du, W.-L.; Xu, Y.-L.; Xu, Z.-R.; Fan, C.-L. Preparation, Characterization and Antibacterial Properties against E. Coli K(88) of Chitosan Nanoparticle Loaded Copper Ions. Nanotechnology 2008, 19, 085707. [Google Scholar] [CrossRef]
- Qi, L.; Xu, Z.; Jiang, X.; Hu, C.; Zou, X. Preparation and Antibacterial Activity of Chitosan Nanoparticles. Carbohydr. Res. 2004, 339, 2693–2700. [Google Scholar] [CrossRef]
- Demers, N.E.; Bayne, C.J. The Immediate Effects of Stress on Hormones and Plasma Lysozyme in Rainbow Trout. Dev. Comp. Immunol. 1997, 21, 363–373. [Google Scholar] [CrossRef] [PubMed]
- Gupta, N.; Gupta, D.K.; Sharma, P.K. Condition Factor and Organosomatic Indices of Parasitized Rattus Rattus as Indicators of Host Health. J. Parasit. Dis. 2017, 41, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Ighwela, K.A.; Ahmad, A.B.; Abol-Munafi, A.B. The Selection of Viscerosomatic and Hepatosomatic Indices for the Measurement and Analysis of Oreochromis Niloticus Condition Fed with Varying Dietary Maltose Levels.
- Morado, C.N.; Araújo, F.G.; Gomes, I.D. The Use of Biomarkers for Assessing Effects of Pollutant Stress on Fish Species from a Tropical River in Southeastern Brazil. Acta Sci. Biol. Sci. 2017, 39, 431–439. [Google Scholar] [CrossRef]
- Hashiguchi, K.; Kawai, K.; Imajoh, M.; Oshima, S. Influence of Fresh Water Bathing on Stress Response of Yellowtail <I>Seriola Quinqueradiata</I>. Aquac. Sci. 2015, 63, 79–87. [Google Scholar] [CrossRef]
- Mahboub, H.H.; Khedr, M.H.E.; Elshopakey, G.E.; Shakweer, M.S.; Mohamed, D.I.; Ismail, T.A.; Ismail, S.H.; Abdel Rahman, A.N. Impact of Silver Nanoparticles Exposure on Neuro-Behavior, Hematology, and Oxidative Stress Biomarkers of African Catfish (Clarias Gariepinus). Aquaculture 2021, 544, 737082. [Google Scholar] [CrossRef]
- Miwa, I.; Okudo, J.; Maeda, K.; Okuda, G. Mutarotase Effect on Colorimetric Determination of Blood Glucose with -D-Glucose Oxidase. Clin. Chim. Acta Int. J. Clin. Chem. 1972, 37, 538–540. [Google Scholar] [CrossRef] [PubMed]
- Guzman, M.; Arcos, M.; Dille, J.; Rousse, C.; Godet, S.; Malet, L. Effect of the Concentration and the Type of Dispersant on the Synthesis of Copper Oxide Nanoparticles and Their Potential Antimicrobial Applications. ACS Omega 2021, 6, 18576–18590. [Google Scholar] [CrossRef] [PubMed]
- Agnew, W.; Barnes, A.C. Streptococcus Iniae: An Aquatic Pathogen of Global Veterinary Significance and a Challenging Candidate for Reliable Vaccination. Vet. Microbiol. 2007, 122, 1–15. [Google Scholar] [CrossRef]
- Eissa, A.E.; Abou-Okada, M.; Alkurdi, A.R.M.; El Zlitne, R.A.; Prince, A.; Abdelsalam, M.; Derwa, H.I.M. Catastrophic Mass Mortalities Caused by Photobacterium Damselae Affecting Farmed Marine Fish from Deeba Triangle, Egypt. Aquac. Res. 2021, 52, 4455–4466. [Google Scholar] [CrossRef]
- Handlinger, J.; Soltani, M.; Percival, S. The Pathology of Flexibacter Maritimus in Aquaculture Species in Tasmania, Australia. J. Fish Dis. 2003, 20, 159–168. [Google Scholar] [CrossRef]
- Kato, G.; Oka, K.; Matsumoto, M.; Kanemaru, M.; Yamamoto, M.; Sano, M. Prevalence of Infection with Nocardia Seriolae in Juvenile of Yellowtail Seriola Quinqueradiata Cultured in Owase Bay, Japan. Fish Pathol. 2020, 55, 1–7. [Google Scholar] [CrossRef]
- Mabrok, M.; Algammal, A.M.; Sivaramasamy, E.; Hetta, H.F.; Atwah, B.; Alghamdi, S.; Fawzy, A.; Avendaño-Herrera, R.; Rodkhum, C. Tenacibaculosis Caused by Tenacibaculum Maritimum: Updated Knowledge of This Marine Bacterial Fish Pathogen. Front. Cell. Infect. Microbiol. 2023, 12, 1068000. [Google Scholar] [CrossRef] [PubMed]
- Park, S.Y.; Han, J.E.; Kwon, H.; Park, S.C.; Kim, J.H. Recent Insights into Aeromonas Salmonicida and Its Bacteriophages in Aquaculture: A Comprehensive Review. J. Microbiol. Biotechnol. 2020, 30, 1443–1457. [Google Scholar] [CrossRef] [PubMed]
- Sanches-Fernandes, G.M.M.; Sá-Correia, I.; Costa, R. Vibriosis Outbreaks in Aquaculture: Addressing Environmental and Public Health Concerns and Preventive Therapies Using Gilthead Seabream Farming as a Model System. Front. Microbiol. 2022, 13. [Google Scholar] [CrossRef] [PubMed]
- Soltani, M.; Baldisserotto, B.; Hosseini Shekarabi, S.P.; Shafiei, S.; Bashiri, M. Lactococcosis a Re-Emerging Disease in Aquaculture: Disease Significant and Phytotherapy. Vet. Sci. 2021, 8, 181. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Zhang, X.-H. Edwardsiella Tarda: An Intriguing Problem in Aquaculture. Aquaculture 2014, 431, 129–135. [Google Scholar] [CrossRef]
- Álvarez-Chimal, R.; García-Pérez, V.I.; Álvarez-Pérez, M.A.; Tavera-Hernández, R.; Reyes-Carmona, L.; Martínez-Hernández, M.; Arenas-Alatorre, J.Á. Influence of the Particle Size on the Antibacterial Activity of Green Synthesized Zinc Oxide Nanoparticles Using Dysphania Ambrosioides Extract, Supported by Molecular Docking Analysis. Arab. J. Chem. 2022, 15, 103804. [Google Scholar] [CrossRef]
- Dong, Y.; Zhu, H.; Shen, Y.; Zhang, W.; Zhang, L. Antibacterial Activity of Silver Nanoparticles of Different Particle Size against Vibrio Natriegens. PloS One 2019, 14, e0222322. [Google Scholar] [CrossRef]
- Naqvi, Q.-U.-A.; Kanwal, A.; Qaseem, S.; Naeem, M.; Ali, S.R.; Shaffique, M.; Maqbool, M. Size-Dependent Inhibition of Bacterial Growth by Chemically Engineered Spherical ZnO Nanoparticles. J. Biol. Phys. 2019. [Google Scholar] [CrossRef]
- Breijyeh, Z.; Jubeh, B.; Karaman, R. Resistance of Gram-Negative Bacteria to Current Antibacterial Agents and Approaches to Resolve It. Molecules 2020, 25, 1340. [Google Scholar] [CrossRef]
- Epand, R.M.; Walker, C.; Epand, R.F.; Magarvey, N.A. Molecular Mechanisms of Membrane Targeting Antibiotics. Biochim. Biophys. Acta BBA - Biomembr. 2016, 1858, 980–987. [Google Scholar] [CrossRef]
- Li, X.-Z.; Plésiat, P.; Nikaido, H. The Challenge of Efflux-Mediated Antibiotic Resistance in Gram-Negative Bacteria. Clin. Microbiol. Rev. 2015, 28, 337–418. [Google Scholar] [CrossRef] [PubMed]
- Sarian, F.D.; Ando, K.; Tsurumi, S.; Miyashita, R.; Ute, K.; Ohama, T. Evaluation of the Growth-Inhibitory Spectrum of Three Types of Cyanoacrylate Nanoparticles on Gram-Positive and Gram-Negative Bacteria. Membranes 2022, 12, 782. [Google Scholar] [CrossRef] [PubMed]
- Nikolaidis, I.; Favini-Stabile, S.; Dessen, A. Resistance to Antibiotics Targeted to the Bacterial Cell Wall. Protein Sci. Publ. Protein Soc. 2014, 23, 243–259. [Google Scholar] [CrossRef] [PubMed]
- Yadav, A.K.; Espaillat, A.; Cava, F. Bacterial Strategies to Preserve Cell Wall Integrity Against Environmental Threats. Front. Microbiol. 2018, 9, 2064. [Google Scholar] [CrossRef]
- Baker-Austin, C. Chapter 6 - Antimicrobial Resistance in Vibrio Species. In Antimicrobial Resistance and Food Safety; Chen, C.-Y., Yan, X., Jackson, C.R., Eds.; Academic Press: San Diego, 2015; pp. 105–118. ISBN 978-0-12-801214-7. [Google Scholar]
- Guz, L.; Nowakiewicz, A.; Puk, K.; Zięba, P.; Gnat, S.; Matuszewski, Ł. Virulence and Antimicrobial Resistance Pattern of Aeromonas Spp. Colonizing European Pond Turtles Emys Orbicularis and Their Natural Environment. First Study from Poland. Anim. Open Access J. MDPI 2021, 11, 2772. [Google Scholar] [CrossRef]
- Gxalo, O.; Digban, T.O.; Igere, B.E.; Olapade, O.A.; Okoh, A.I.; Nwodo, U.U. Virulence and Antibiotic Resistance Characteristics of Vibrio Isolates From Rustic Environmental Freshwaters. Front. Cell. Infect. Microbiol. 2021, 11. [Google Scholar] [CrossRef]
- Loo, K.-Y.; Letchumanan, V.; Law, J.W.-F.; Pusparajah, P.; Goh, B.-H.; Ab Mutalib, N.-S.; He, Y.-W.; Lee, L.-H. Incidence of Antibiotic Resistance in Vibrio Spp. Rev. Aquac. 2020, 12, 2590–2608. [Google Scholar] [CrossRef]
- Trudel, M.V.; Vincent, A.T.; Attéré, S.A.; Labbé, M.; Derome, N.; Culley, A.I.; Charette, S.J. Diversity of Antibiotic-Resistance Genes in Canadian Isolates of Aeromonas Salmonicida Subsp. Salmonicida: Dominance of PSN254b and Discovery of PAsa8. Sci. Rep. 2016, 6, 35617. [Google Scholar] [CrossRef]
- Wu, L.; Tian, Y.; Pang, M.; Yang, Z.; Bao, H.; Zhou, Y.; Sun, L.; Wang, R.; Zhang, H. A Novel Vibriophage VB_VhaS_PcB-1G Capable of Inhibiting Virulent Vibrio Harveyi Pathogen. Aquaculture 2021, 542, 736854. [Google Scholar] [CrossRef]
- Reichley, S.R.; Ware, C.; Steadman, J.; Gaunt, P.S.; García, J.C.; LaFrentz, B.R.; Thachil, A.; Waldbieser, G.C.; Stine, C.B.; Buján, N.; et al. Comparative Phenotypic and Genotypic Analysis of Edwardsiella Isolates from Different Hosts and Geographic Origins, with Emphasis on Isolates Formerly Classified as E. Tarda, and Evaluation of Diagnostic Methods. J. Clin. Microbiol. 2017, 55, 3466–3491. [Google Scholar] [CrossRef] [PubMed]
- Costa, A.B.; Kanai, K.; Yoshikoshi, K. Serological Characterization of Atypical Strains of Edwardsiella Tarda Isolated from Sea Breams. Fish Pathol. 33, 265–274. [CrossRef]
- Divya, K.; Vijayan, S.; George, T.K.; Jisha, M.S. Antimicrobial Properties of Chitosan Nanoparticles: Mode of Action and Factors Affecting Activity. Fibers Polym. 2017, 18, 221–230. [Google Scholar] [CrossRef]
- Boyd, C.E.; Tucker, C.S. Pond Aquaculture Water Quality Management; Springer US: Boston, MA, 1998; ISBN 978-1-4613-7469-5. [Google Scholar]
- Zhao, J.; Wang, Z.; Liu, X.; Xie, X.; Zhang, K.; Xing, B. Distribution of CuO Nanoparticles in Juvenile Carp (Cyprinus Carpio) and Their Potential Toxicity. J. Hazard. Mater. 2011, 197, 304–310. [Google Scholar] [CrossRef] [PubMed]
- Azevedo, P. a.; Leeson, S.; Cho, C. y.; Bureau, D. p. Growth and Feed Utilization of Large Size Rainbow Trout (Oncorhynchus Mykiss) and Atlantic Salmon (Salmo Salar) Reared in Freshwater: Diet and Species Effects, and Responses over Time. Aquac. Nutr. 2004, 10, 401–411. [Google Scholar] [CrossRef]
- Zaineldin, A.I.; Hegazi, S.; Koshio, S.; Ishikawa, M.; Bakr, A.; El-Keredy, A.M.S.; Dawood, M.A.O.; Dossou, S.; Wang, W.; Yukun, Z. Bacillus Subtilis as Probiotic Candidate for Red Sea Bream: Growth Performance, Oxidative Status, and Immune Response Traits. Fish Shellfish Immunol. 2018, 79, 303–312. [Google Scholar] [CrossRef]
- Ighwela, K.A.; Ahmad, A.B.; Abol-Munafi, A.B. The Selection of Viscerosomatic and Hepatosomatic Indices for the Measurement and Analysis of Oreochromis Niloticus Condition Fed with Varying Dietary Maltose Levels. 2014, 3, 18–20. [Google Scholar]
- Bavia, L.; Santiesteban-Lores, L.E.; Carneiro, M.C.; Prodocimo, M.M. Advances in the Complement System of a Teleost Fish, Oreochromis Niloticus. Fish Shellfish Immunol. 2022, 123, 61–74. [Google Scholar] [CrossRef]
- Biller-Takahashi, J.D.; Urbinati, E.C. Fish Immunology. The Modification and Manipulation of the Innate Immune System: Brazilian Studies. An. Acad. Bras. Ciênc. 2014, 86, 1484–1506. [Google Scholar] [CrossRef]
- Saurabh, S.; Sahoo, P.K. Lysozyme: An Important Defence Molecule of Fish Innate Immune System. Aquac. Res. 2008, 39, 223–239. [Google Scholar] [CrossRef]
- Subbotkin, M.F.; Subbotkina, T.A. Effects of Environment and Physiological State of an Organism on the Activity and Content of Lysozyme in Fishes of the Family Cyprinidae: A Review. Inland Water Biol. 2018, 11, 184–194. [Google Scholar] [CrossRef]
- Holland, M.C.H.; Lambris, J.D. The Complement System in Teleosts. Fish Shellfish Immunol. 2002, 12, 399–420. [Google Scholar] [CrossRef] [PubMed]
- Sauer, D.J.; Heidinger, B.J.; Kittilson, J.D.; Lackmann, A.R.; Clark, M.E. No Evidence of Physiological Declines with Age in an Extremely Long-Lived Fish. Sci. Rep. 2021, 11, 9065. [Google Scholar] [CrossRef] [PubMed]
- Abdelazim, A.M.; Saadeldin, I.M.; Swelum, A.A.-A.; Afifi, M.M.; Alkaladi, A. Oxidative Stress in the Muscles of the Fish Nile Tilapia Caused by Zinc Oxide Nanoparticles and Its Modulation by Vitamins C and E. Oxid. Med. Cell. Longev. 2018, 2018, e6926712. [Google Scholar] [CrossRef]
- Farag, M.R.; Abo-Al-Ela, H.G.; Alagawany, M.; Azzam, M.M.; El-Saadony, M.T.; Rea, S.; Di Cerbo, A.; Nouh, D.S. Effect of Quercetin Nanoparticles on Hepatic and Intestinal Enzymes and Stress-Related Genes in Nile Tilapia Fish Exposed to Silver Nanoparticles. Biomedicines 2023, 11, 663. [Google Scholar] [CrossRef] [PubMed]
- García-Medina, S.; Galar-Martínez, M.; Cano-Viveros, S.; Ruiz-Lara, K.; Gómez-Oliván, L.M.; Islas-Flores, H.; Gasca-Pérez, E.; Pérez-Pastén-Borja, R.; Arredondo-Tamayo, B.; Hernández-Varela, J.; et al. Bioaccumulation and Oxidative Stress Caused by Aluminium Nanoparticles and the Integrated Biomarker Responses in the Common Carp (Cyprinus Carpio). Chemosphere 2022, 288, 132462. [Google Scholar] [CrossRef]
- Saddick, S.; Afifi, M.; Abu Zinada, O.A. Effect of Zinc Nanoparticles on Oxidative Stress-Related Genes and Antioxidant Enzymes Activity in the Brain of Oreochromis Niloticus and Tilapia Zillii. Saudi J. Biol. Sci. 2017, 24, 1672–1678. [Google Scholar] [CrossRef]
- Hood, E. Fullerenes and Fish Brains: Nanomaterials Cause Oxidative Stress. Environ. Health Perspect. 2004, 112, A568. [Google Scholar] [CrossRef]
- Ibrahim, A.Th.A.; Banaee, M.; Sureda, A. Genotoxicity, Oxidative Stress, and Biochemical Biomarkers of Exposure to Green Synthesized Cadmium Nanoparticles in Oreochromis Niloticus (L.). Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2021, 242, 108942. [Google Scholar] [CrossRef]
Figure 1.
(a) Preparation of the isobutyl cyanoacrylate (iBCA) nanoparticles used in this study (b) Plate comparing cloudiness of the iBCA nanoparticles solutions used in this study.
Figure 1.
(a) Preparation of the isobutyl cyanoacrylate (iBCA) nanoparticles used in this study (b) Plate comparing cloudiness of the iBCA nanoparticles solutions used in this study.
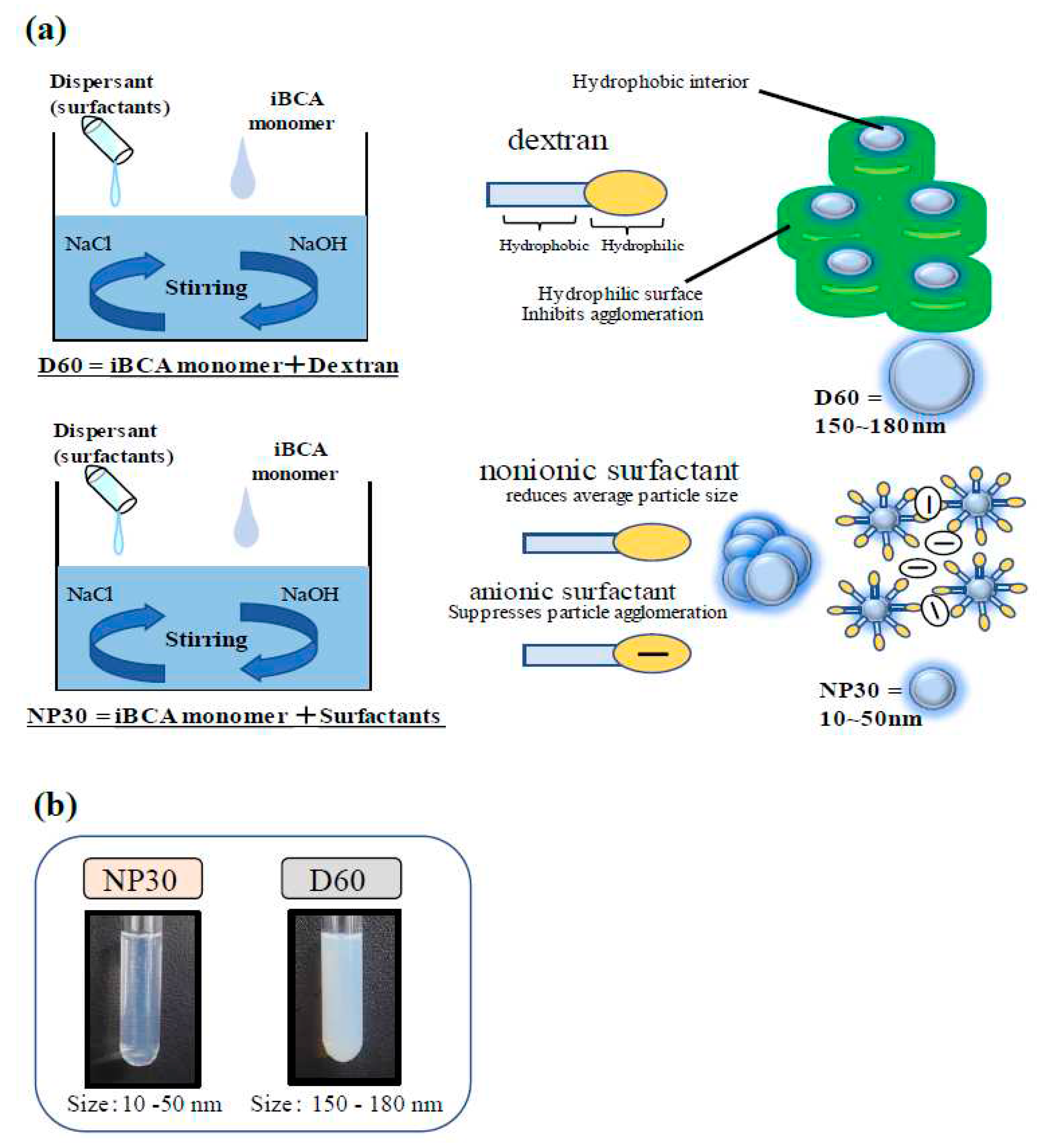
Figure 2.
Plot to determine MIC, showing bacterial inhibition after 24 hours (in NP30) vs bacterial growth (in Dispersant). From the plot, MIC of NP30 against E. tarda (fl) is 125 µg/ml.
Figure 2.
Plot to determine MIC, showing bacterial inhibition after 24 hours (in NP30) vs bacterial growth (in Dispersant). From the plot, MIC of NP30 against E. tarda (fl) is 125 µg/ml.
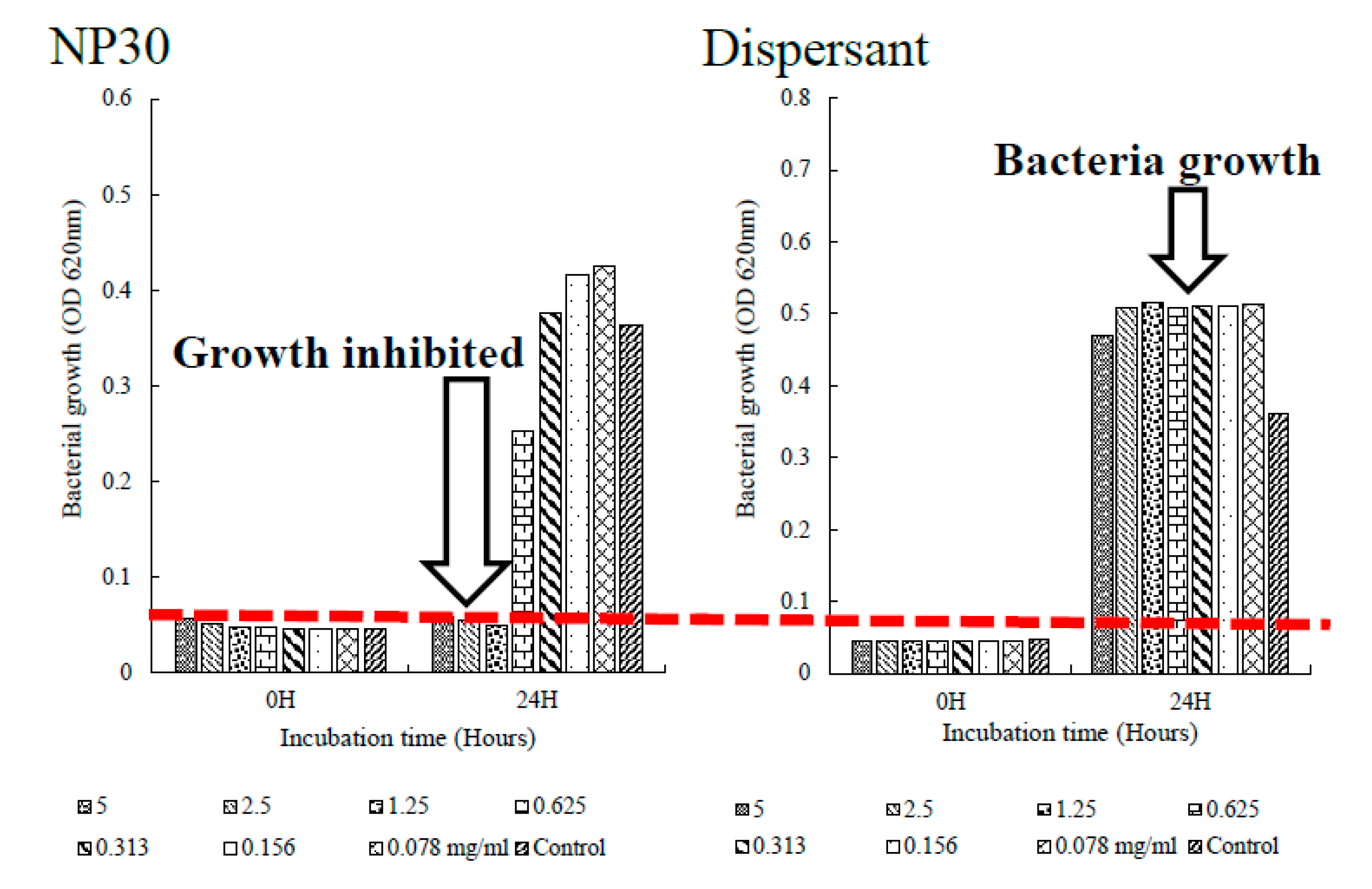
Figure 3.
Per cent growth inhibition (GI %) of Vibrio spp. at all growth inhibiting concentrations (a) GI % of Vibrio anguillarum for D60 (I) and NP30 (II), (b) GI % of Vibrio harveyi for D60 (I) and NP30 (II); (c) GI % of Vibrio parahaemolyticus for D60 (I) and NP30 (II); (d) GI % of Vibrio rotiferanus for D60 (I) and NP30 (II). All data were expressed as means ± SEM (n = 3).
Figure 3.
Per cent growth inhibition (GI %) of Vibrio spp. at all growth inhibiting concentrations (a) GI % of Vibrio anguillarum for D60 (I) and NP30 (II), (b) GI % of Vibrio harveyi for D60 (I) and NP30 (II); (c) GI % of Vibrio parahaemolyticus for D60 (I) and NP30 (II); (d) GI % of Vibrio rotiferanus for D60 (I) and NP30 (II). All data were expressed as means ± SEM (n = 3).
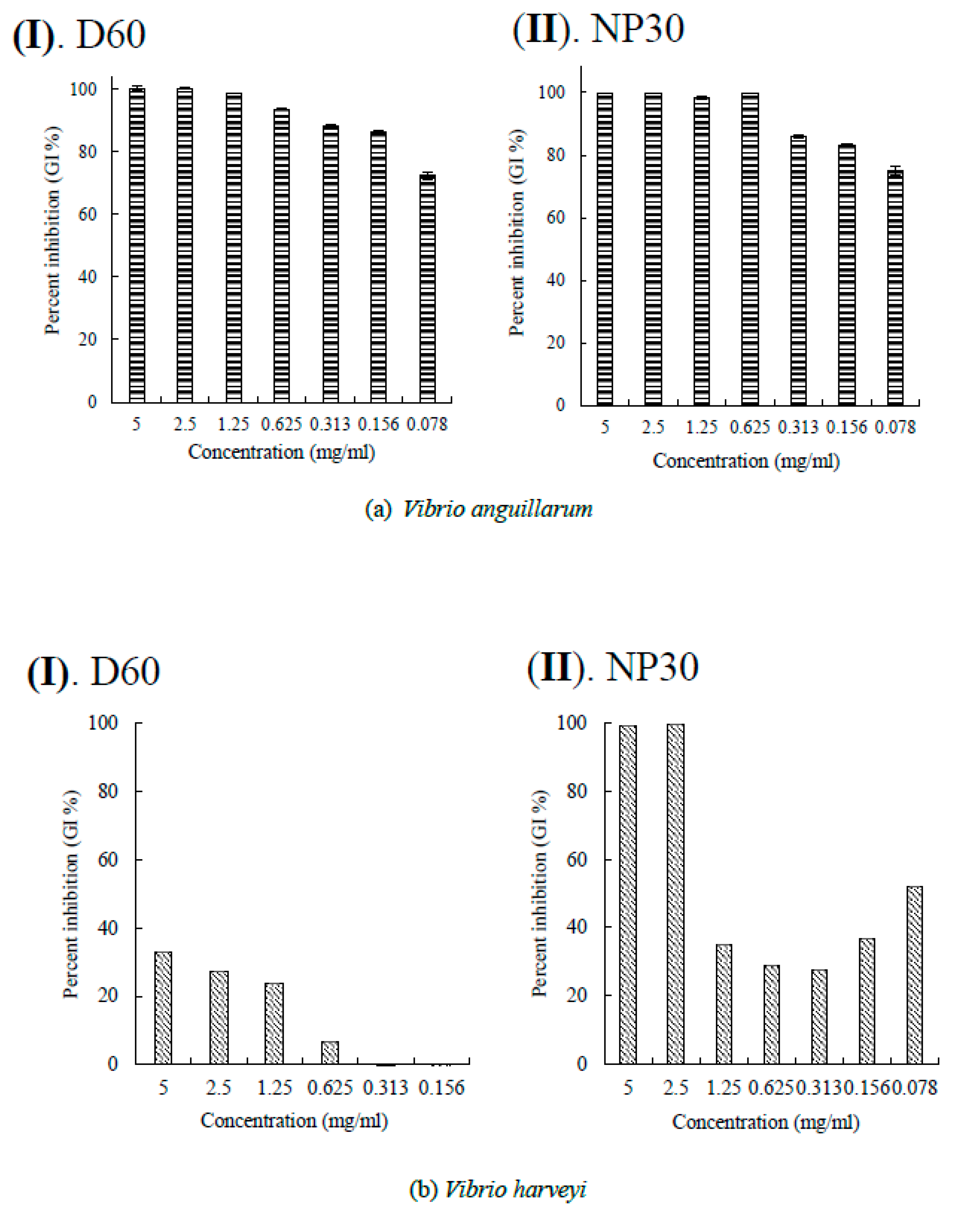
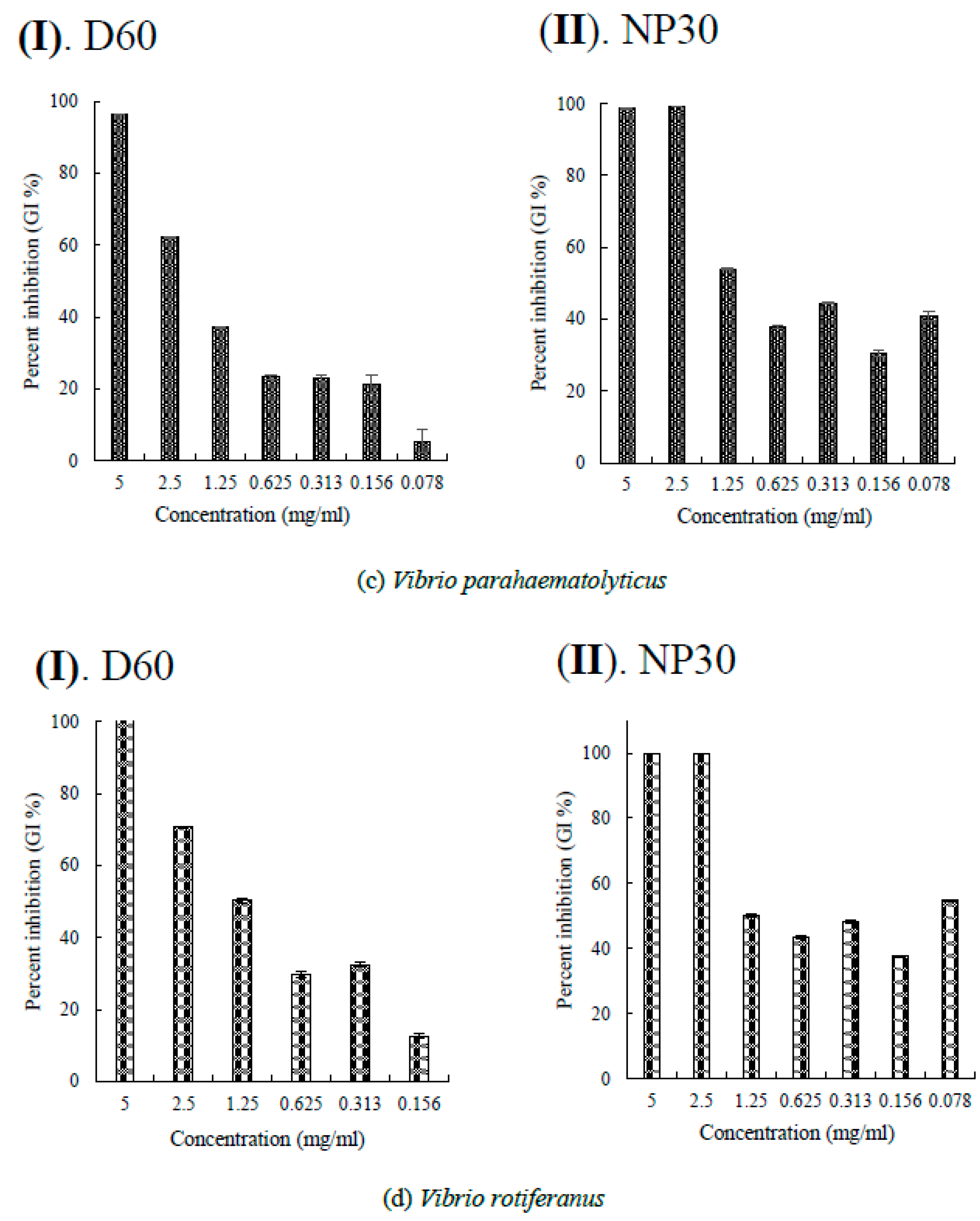
Figure 4.
Per cent growth inhibition (GI %) of some Gram-positive bacteria strains at all growth inhibiting concentrations observed (a) GI % of Lactococcus garvieae; (b) GI % of Nocardia seriolae; (c) GI % of Streptococcus iniae for D60 (I) and NP30 (II). All data were expressed as means ± SEM (n = 3).
Figure 4.
Per cent growth inhibition (GI %) of some Gram-positive bacteria strains at all growth inhibiting concentrations observed (a) GI % of Lactococcus garvieae; (b) GI % of Nocardia seriolae; (c) GI % of Streptococcus iniae for D60 (I) and NP30 (II). All data were expressed as means ± SEM (n = 3).
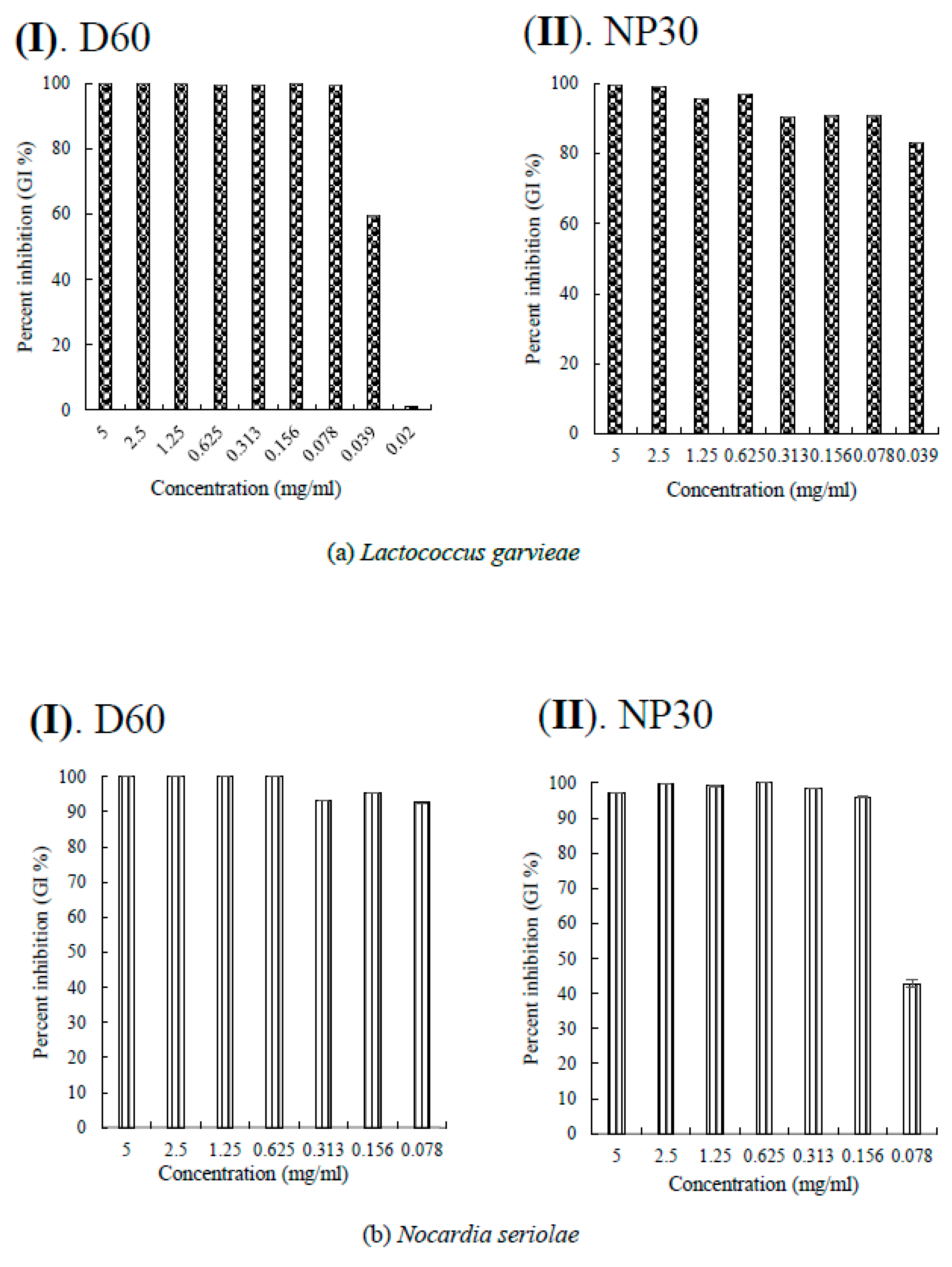
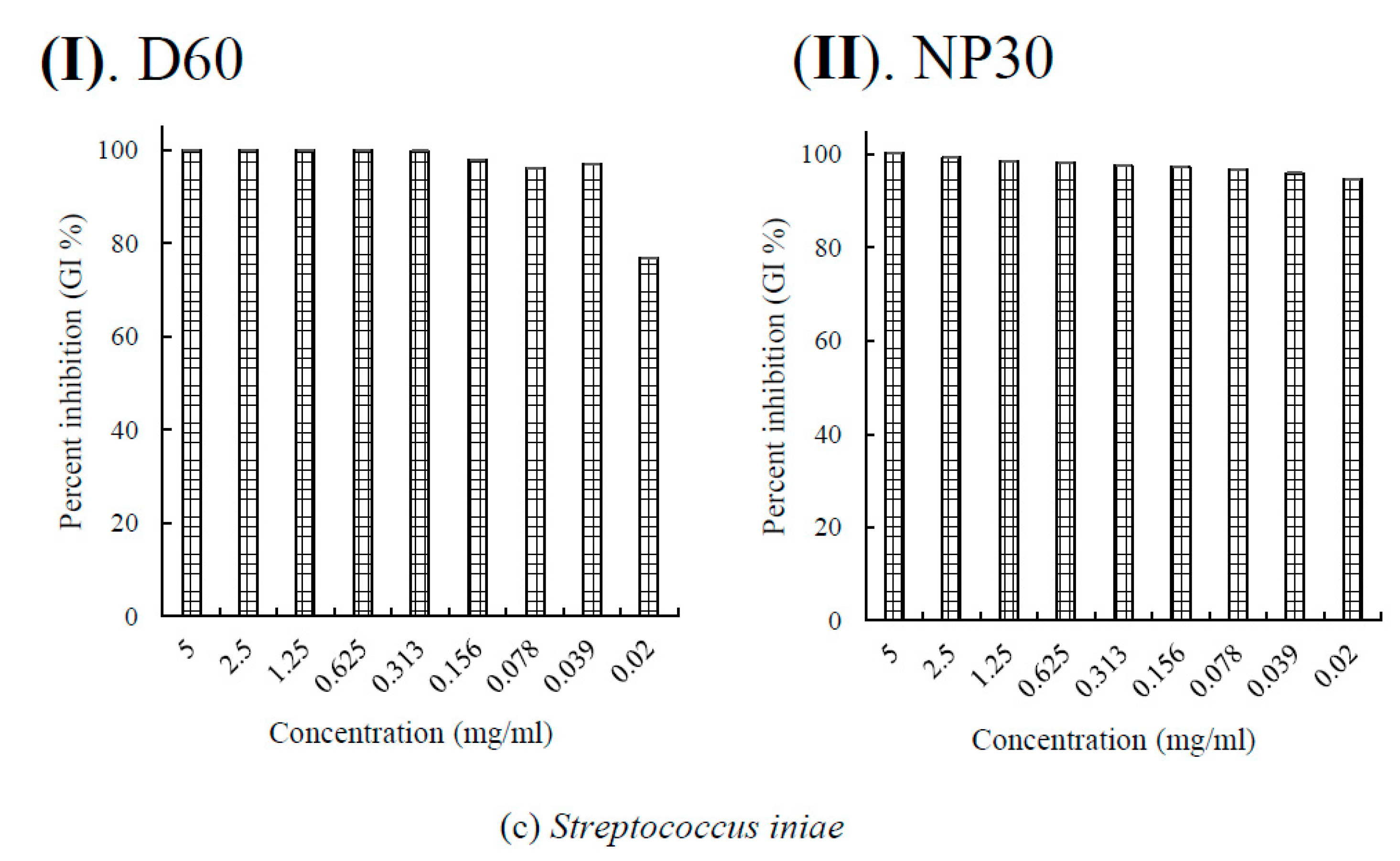
Figure 5.
Per cent growth inhibition (GI %) of some Gram-negative bacteria strains at all growth inhibiting concentrations observed (a) GI % of Flexibacter maritimus; (b) GI % of Photobacterium damselae subsp. piscicida; (c) GI % of Tericibaculum maritimum; (d) GI % of Aeromonas. salmonicida subsp. salmonicida; (e) GI % of Edwardsiella tarda (typical); (f) GI % of Edwardsiella tarda (atypical) for D60 (I) and NP30 (II). All data were expressed as means ± SEM (n = 3).
Figure 5.
Per cent growth inhibition (GI %) of some Gram-negative bacteria strains at all growth inhibiting concentrations observed (a) GI % of Flexibacter maritimus; (b) GI % of Photobacterium damselae subsp. piscicida; (c) GI % of Tericibaculum maritimum; (d) GI % of Aeromonas. salmonicida subsp. salmonicida; (e) GI % of Edwardsiella tarda (typical); (f) GI % of Edwardsiella tarda (atypical) for D60 (I) and NP30 (II). All data were expressed as means ± SEM (n = 3).
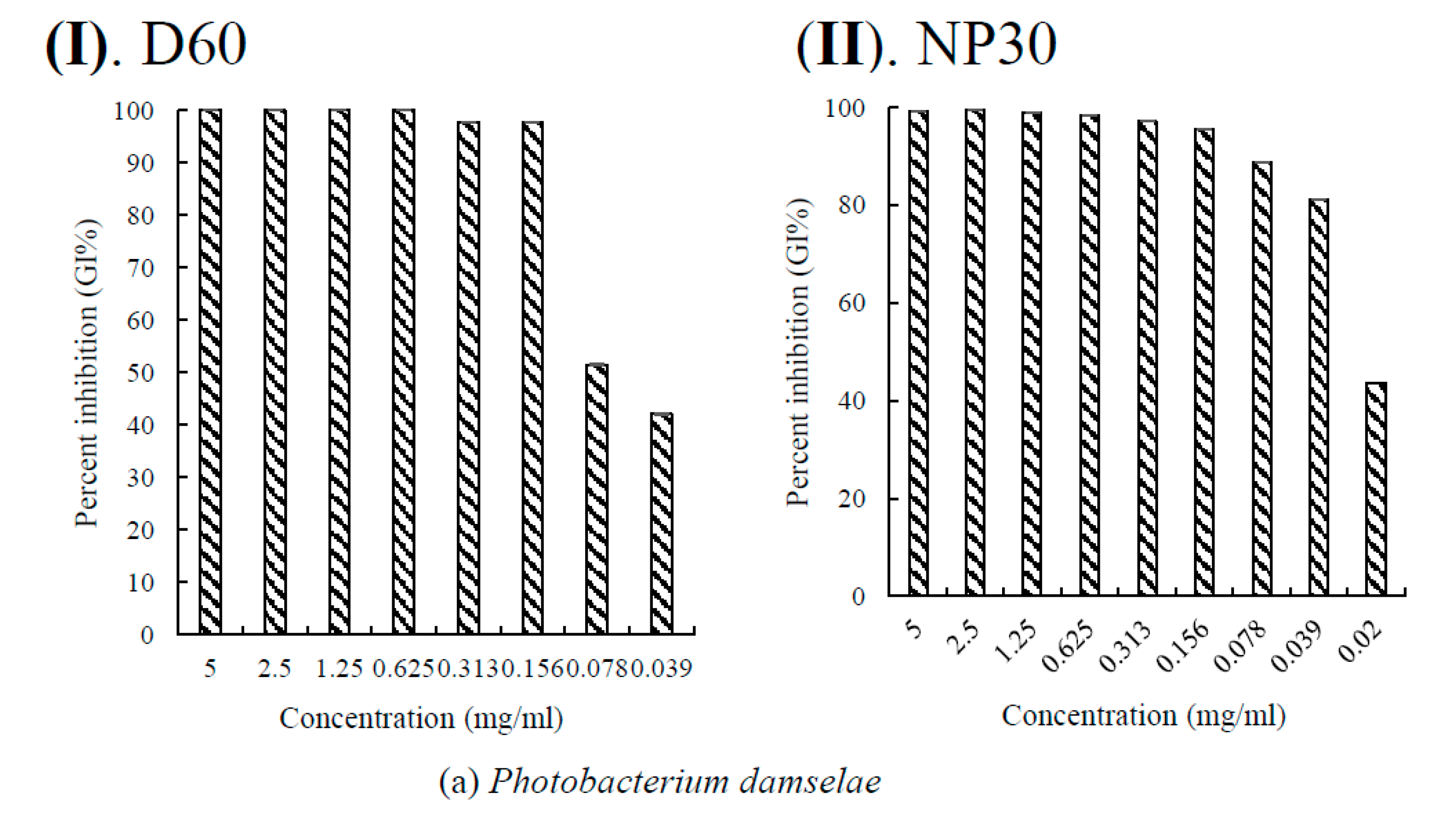
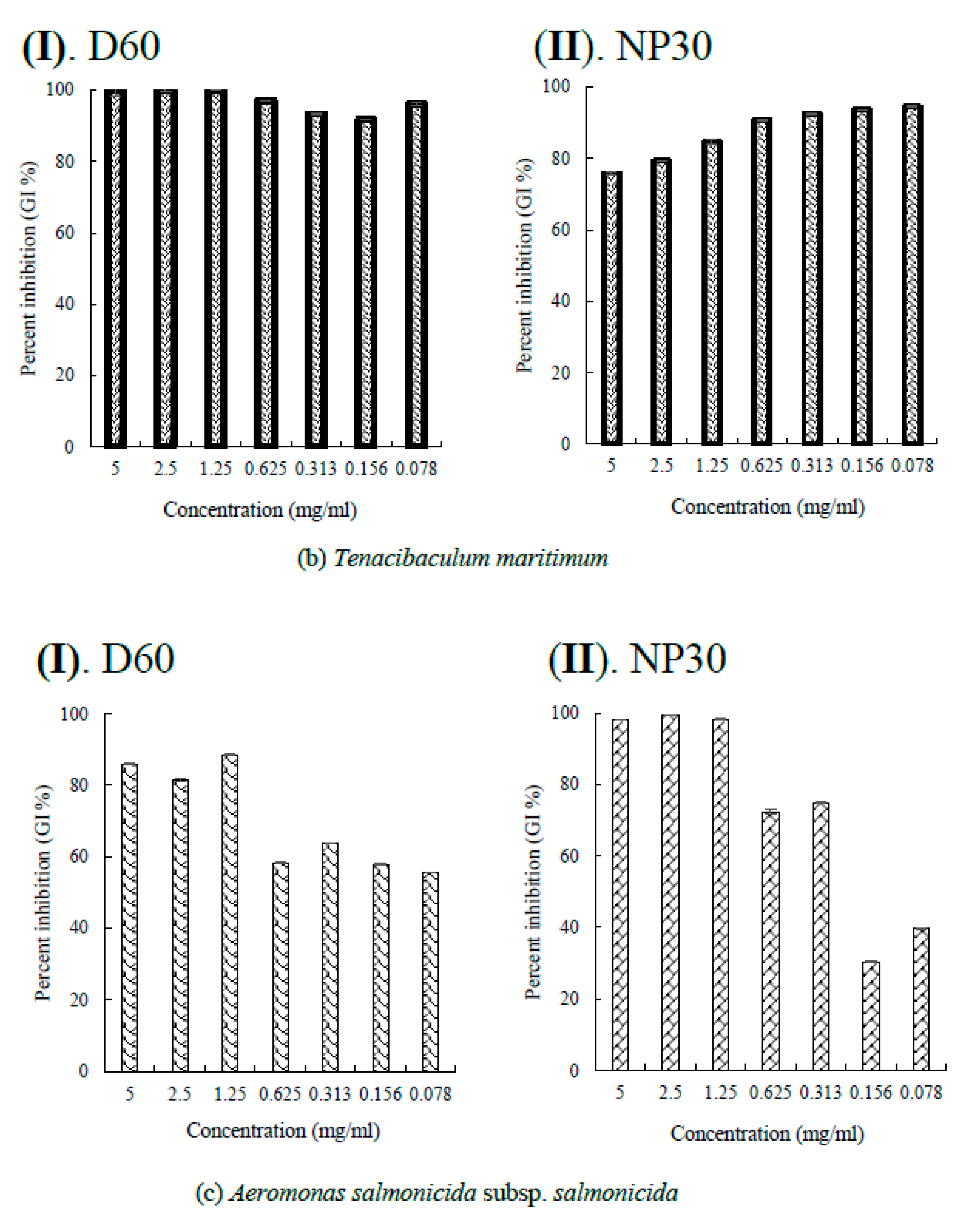
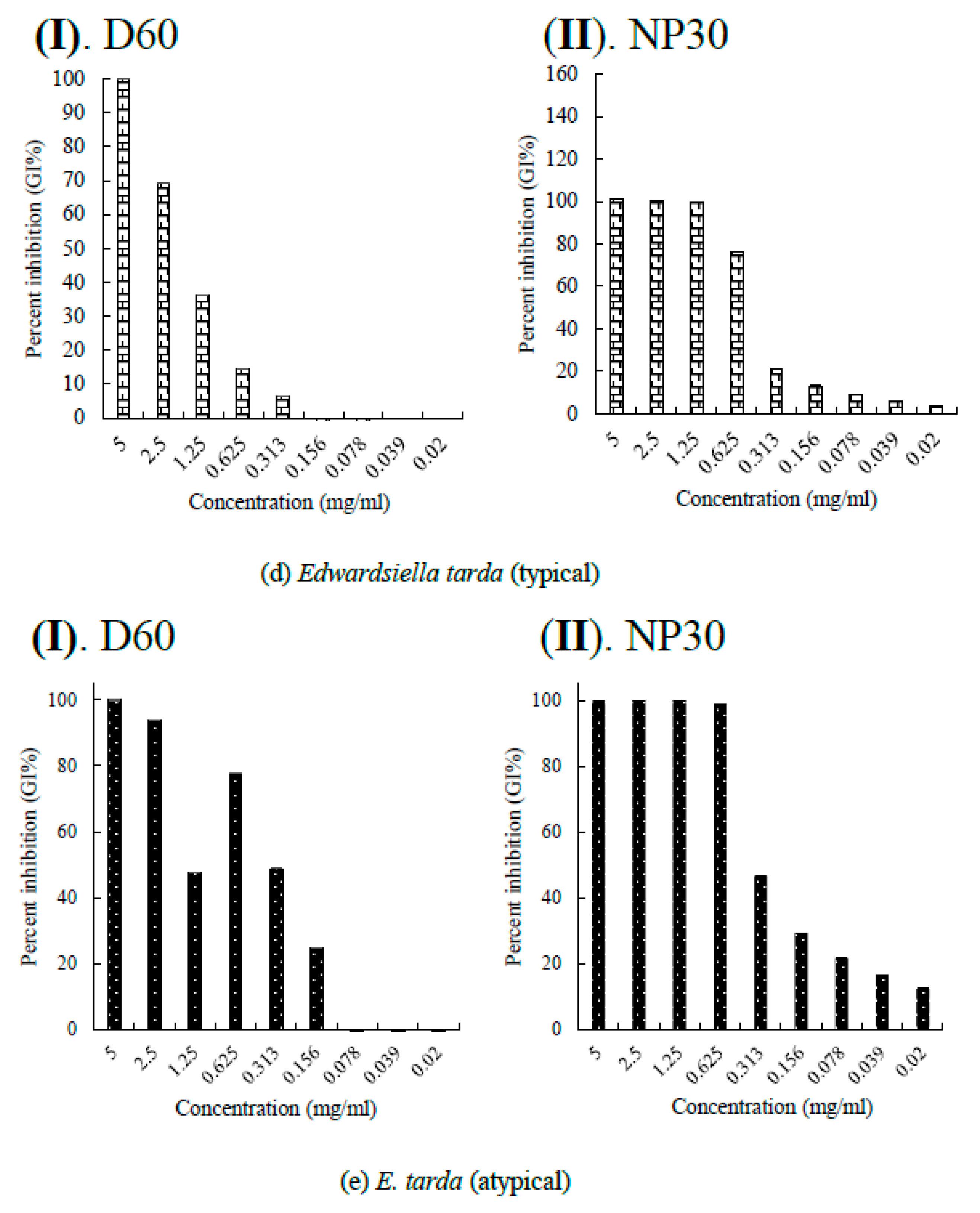
Figure 6.
Overall bacterial sensitivity via percent growth inhibition by (A) D60 and (B) NP30 at 0.5 mg/ml in descending order.
Figure 6.
Overall bacterial sensitivity via percent growth inhibition by (A) D60 and (B) NP30 at 0.5 mg/ml in descending order.

Figure 7.
Fish growth and safety parameters (a) Relative gain rate of O. mykiss over the study period (b) Blood plasma lysozyme activity at OD 493 nm (c) Plasma complement activity at OD 493 nm (d) Plasma Glucose concentrations; (e) Rate of plasma SOD inhibition (%) per treatment; (f) Plate comparing healthy rainbow trout and one with a tumor isolated from a NP30 (N = 1). All data were expressed as means ± SEM; where n = number of replicates per treatment and N = total number of fish for each test. Different letter superscripts indicate significant difference between treatments and or sampling time (P < 0.05). CT = Control group; M1, M2 & M3 = Months 1,2 & 3.
Figure 7.
Fish growth and safety parameters (a) Relative gain rate of O. mykiss over the study period (b) Blood plasma lysozyme activity at OD 493 nm (c) Plasma complement activity at OD 493 nm (d) Plasma Glucose concentrations; (e) Rate of plasma SOD inhibition (%) per treatment; (f) Plate comparing healthy rainbow trout and one with a tumor isolated from a NP30 (N = 1). All data were expressed as means ± SEM; where n = number of replicates per treatment and N = total number of fish for each test. Different letter superscripts indicate significant difference between treatments and or sampling time (P < 0.05). CT = Control group; M1, M2 & M3 = Months 1,2 & 3.
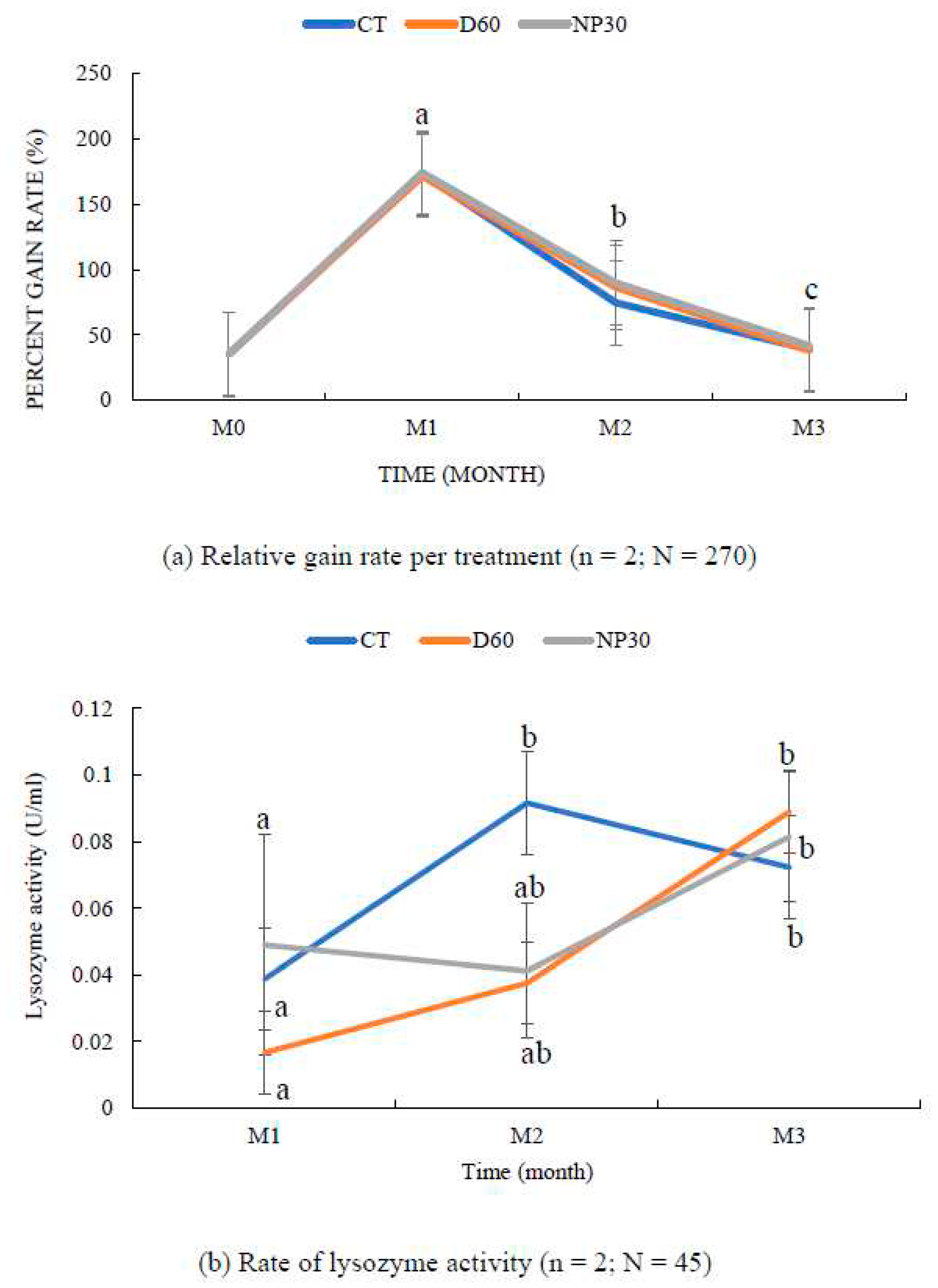
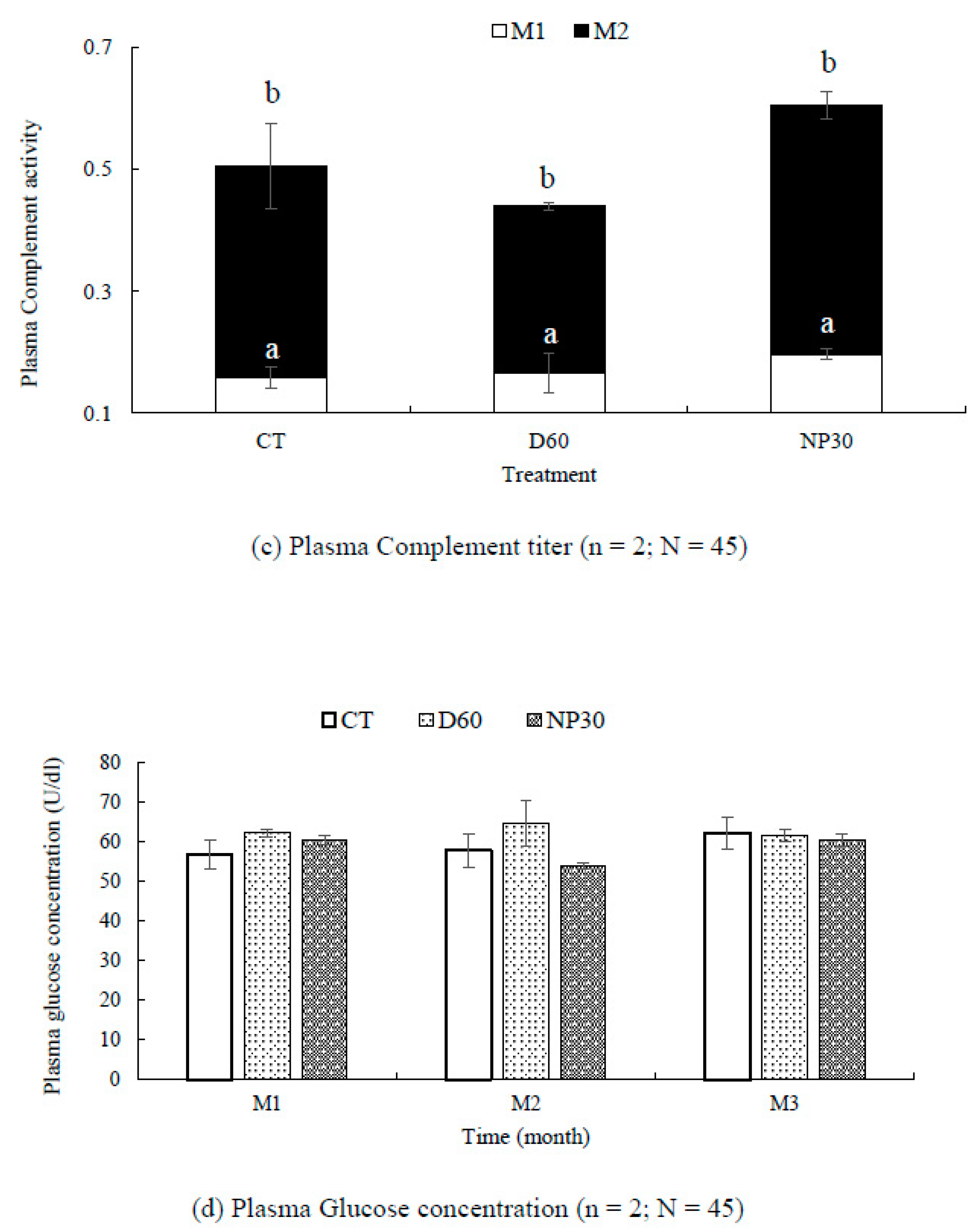
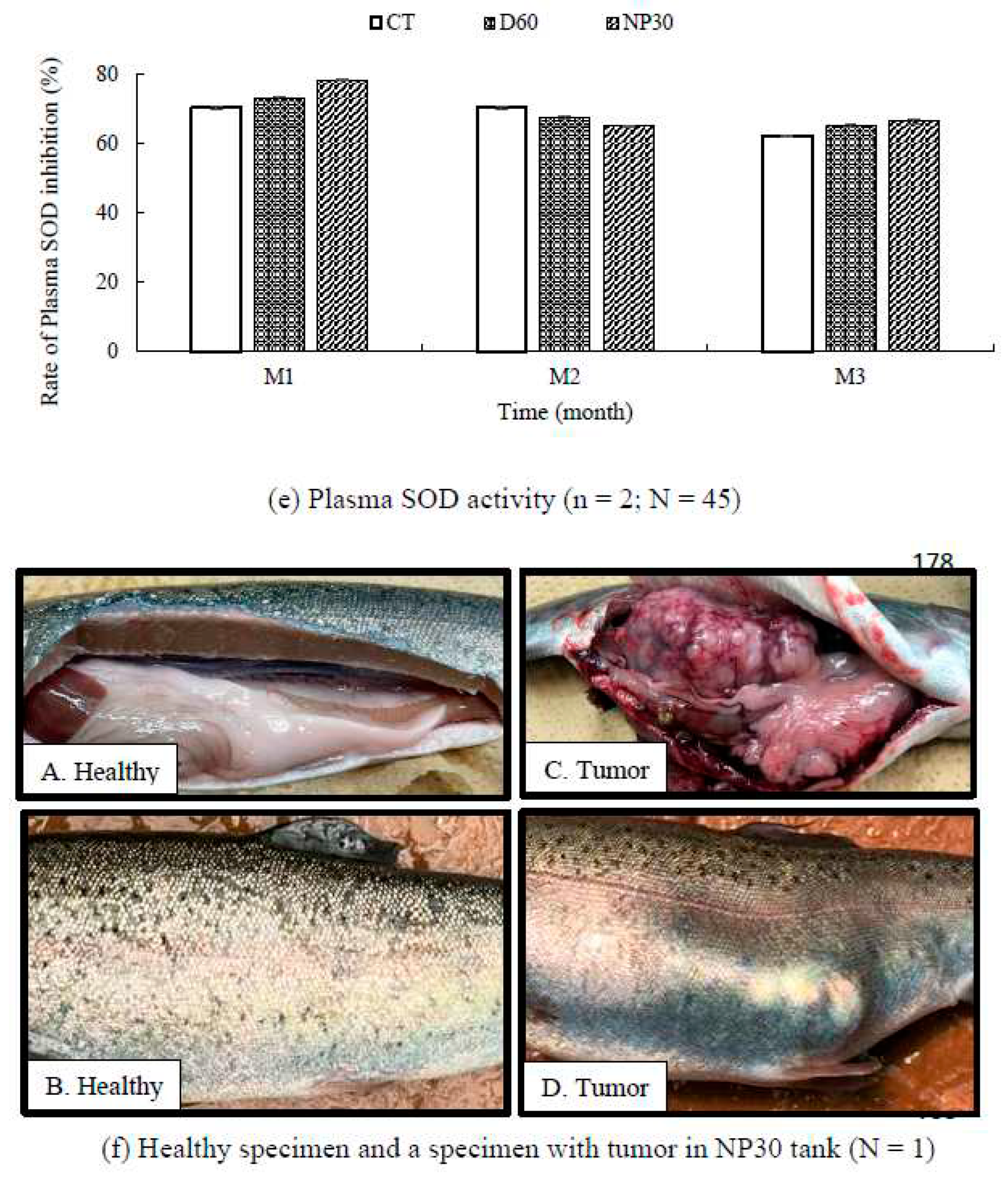

Table 1.
Antibacterial activity, of isobutyl cyanoacrylate nanoparticles, D60 (180 nm) and NP30 (30 nm) expressed as MIC and MBC against some fish bacterial pathogens.
Table 1.
Antibacterial activity, of isobutyl cyanoacrylate nanoparticles, D60 (180 nm) and NP30 (30 nm) expressed as MIC and MBC against some fish bacterial pathogens.
| Bacteria type | Species | NP30 (mg/ml) | D60 (mg/ml) | ||
|---|---|---|---|---|---|
| MIC | MBC | MIC | MBC | ||
| Gram-positive bacteria | Lactococcus garvieae | 0.156 | 5.00 | 0.078 | - |
| Streptococcus iniae | 0.078 | 0.078 | 0.078 | - | |
| Nocardia seriolae | 0.313 | 1.25 | 0.313 | - | |
| Gram-negative bacteria | Edwardsiella tarda (typical) | 1.25 | 2.50 | 5.00 | - |
| Edwardsiella tarda (atypical) | 1.25 | 2.50 | 5.00 | - | |
| Aeromonas salmonicida subsp. salmonicida | 1.25 | 2.50 | 5.00 | - | |
| Tenacibaculum maritimum | 0.078 | 0.313 | 0.078 | 0.625 | |
| Photobacterium damselae subsp. piscicida | 0.078 | 5.00 | 0.078 | - | |
| Vibrio anguillarium | 0.625 | 1.25 | 0.313 | 2.50 | |
| V. harveyi | 2.50 | 2.50 | - | - | |
| V. parahaemolyticus | 2.50 | 2.50 | 5.00 | - | |
| V. rotiferanus | 2.50 | 2.50 | 5.00 | - | |
MIC: minimum inhibitory concentration, MBC: minimum bactericidal concentration. (-): The value was not obtained at all doses examined in the current study.
Table 2.
Growth performance, feed utilization and health conditions of rainbow trout fed the test diets (with iBCA-NP – D60, NP30 or without any -Control).
Table 2.
Growth performance, feed utilization and health conditions of rainbow trout fed the test diets (with iBCA-NP – D60, NP30 or without any -Control).
| Parameters | Treatment | ||
|---|---|---|---|
| Control | D60 | NP30 | |
| Initial weight (g) | 35.05 ± 0.42 | 35.07 ± 0.42 | 35.12 ± 0.52 |
| Final weight (g) | 231.75 ± 6.61 | 244.76 ± 6.46 | 258.04 ± 7.65 |
| Relative gain rate (RGR %) | 561.20 ± 6.62 | 598.16 ± 40.48 | 634.78 ± 52.8 |
| Specific growth (% per day) | 2.25 ± 0.71 | 2.31 ± 0.7 | 2.37 ± 0.69 |
| Feed conversion ratio (FCR) | 1.51 ± 0.06 | 1.25 ± 0.31 | 1.26 ± 0.26 |
| Survival rate (%) | 98.89 ± 1.11 | 98.89 ± 1.11 | 100 ± 0.00 |
| Condition factor | 1.75 ± 0.02 | 1.92 ± 0.10 | 1.83 ± 0.06 |
| Hepatosomatic index | 1.03 ± 0.04 | 1.07 ± 0.05 | 1.06 ± 0.07 |
| Viscerosomatic index | 8.47 ± 0.09 | 8.46 ± 0.12 | 8.92 ± 0.46 |
Results were presented as means ± SEM (n = 2). In a row, means with the different letter superscripts were significantly different (P < 0.05). There was no significant difference among treatments for all parameters assessed. Condition factor = 100 × [fish weight/(fish length)^3]; Hepatosomatic index = 100 × (liver weight/fish weight); Viscerosomatic index = 100 × (viscera weight/fish weight).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



