
Submitted:
17 September 2023
Posted:
19 September 2023
You are already at the latest version
Abstract
The heavy-metal-associated (HMA) proteins are a class of PB1-Type ATPases related to the intracellular transport and detoxification of metals. However, due to a lack of information regarding the HMA gene family in the Cucurbitaceae family, a comprehensive genome-wide analysis of the HMA family was performed in ten Cucurbitaceae species: Citrullus amarus, Citrullus colocynthis, Citrullus lanatus, Citrullus mucosospermus, Cucumis melo, Cucumis sativus, Cucurbita maxima, Cucurbita moschata, Cucurbita pepo, and Legenaria siceraria. We identified 103 Cucurbit HMA proteins with various members, ranging from 8 (Legenaria siceraria) to 14 (Cucurbita pepo) across species. The phylogenetic and structural analysis confirmed that the Cucurbitaceae HMA protein family could be further classified into two major clades: Zn/Co/Cd/Pb and Cu/Ag. GO annotation-based subcellular localization analysis predicted that all HMA gene family members were localized on membranes. Moreover, the analysis of conserved motifs and gene structure (intron/exon) revealed the functional divergence between clades. Interspecies microsynteny analysis demonstrated that maximum orthologous genes were found between species of the Citrullus genera. Finally, nine candidates HMA genes were selected, and their expression analysis was carried out via qRT-PCR in root, leaf, flower, and fruit tissues of C. pepo under arsenic stress. The expression pattern of the CpeHMA genes showed a distinct pattern of expression in root and shoot tissues, with a remarkable expression of CpeHMA6 and CpeHMA3 genes from the Cu/Ag clade. Overall, this study provides insights into the functional analysis of the HMA gene family in Cucurbitaceae species and lays down the basic knowledge to explore the role and mechanism of the HMA gene family to cope with arsenic stress conditions.
Keywords:
HMA
; P1B-type ATPase
; Arsenic stress
; Arsenic transport
; Cucurbits
1. Introduction
Cucurbits are one of the major crop families with high economic value and are widely cultivated worldwide. Four genera, Cucurbita (squash, pumpkins), Cucumis (cucumbers, melons), Citrullus (watermelons), and Lagenaria (bottle gourd) are among the ten most economically significant vegetable crops in the world, while numerous others have regional significance [1]. Nevertheless, the global cucurbit growth and yield are adversely influenced by environmental stresses such as drought, salinity, and high concentrations of heavy metals and metalloids [2,3]. Arsenic (As) is a non-essential metalloid ubiquitous in the soil at low levels; however, agricultural soils are threatened by toxic contamination from anthropogenic activities, leading to excessive accumulation of arsenic [4]. Its presence in polluted environmental conditions such as groundwater and cropping soil causes severe threats to living organisms, including plants and, consequently humans [5,6]. Plants have developed various adaptation strategies to protect themselves from harmful environmental conditions, including accumulating and transporting heavy metals [7]. Membrane transport plays a vital role in heavy metal detoxification, allowing absorption and transport of many cations from the root to the shoot and redistribution among aerial parts [8,9]. Among the different membrane transporters, the P1B-type ATPase, also known as the heavy metal ATPase (HMAs), which belongs to the large P-type ATPase family, plays an important role in heavy metal transport [10,11,12]. HMAs transport essential metal ions required for plant growth and development, such as Cu2+ and Zn2+, and distribute non-essential heavy metal ions, including Cd2+, Co 2+, and Pb2+. Typical HMA proteins contain the E1–E2 ATPase domain and a haloacid dehalogenase-like hydrolase domain. Additionally, both sides of the N-terminal and C-terminal metal-binding sites may possess one or more soluble metal-binding domains (MBDs) that interact with or bind to specific metal ions [8,13,14]. The HMA domain is also located in P1B-type ATPases, which is a heavy metal-associated regulatory domain [15,16]. Based on metal substrate specificity, HMAs can be clustered into two major phylogenetic subclasses, namely, the Cu/Ag P1B-ATPase group and the Zn/Co/Cd/Pb P1B-ATPase group [17].
HMAs genes have been identified in model and non-model plants with a different number of genes and diversification patterns, including 8 in Arabidopsis thaliana [15], 9 in rice (Oryza sativa) [18], 12 in Populus trichocarpa [19], 20 in soybean (Glycine max) [20], 11 in Maize (Zea mays) [21], 11 in sorghum (Sorghum bicolor) [21], 9 in Barley (Hordeum vulgare) [22,23], 12 in Flax (Linum usitatissimum) [24], 31 in Brassica napus [25], 8 in Chinese pear (Pyrus bretschneideri) [26], 8 in mulberry (Morus alba) [27], 9 in Medicago truncatula [28] and 7 in Tartary buckwheat [29]. The functions of HMA genes have been comprehensively studied; for instance, in A. thaliana, AtHMA1 is involved in exporting Zn from the chloroplast [30], while the overexpression of AtHMA3 enhances tolerance and accumulation of Cd, Zn, Pb, and Co in plants [31]. Likewise, overexpression of SpHMA3 in Sedum plumbizinccicola has been reported to confer Cd hyper-tolerance [32]. Moreover, it has been shown that OsHMA5 is involved in the shoot translocation of Cu2+ in rice [33], whereas OsHMA1 and OsHMA3 are involved in the Zn and Cd transport, respectively [18]. A total of eight HMA proteins have been identified in Cucumber as a response to cadmium accumulation. Although, these proteins have not been further characterized, among them, CsHMA3 and CsHMA4 have been associated with the transportation of cadmium, lead, and zinc from the root to the stem [34].
Although HMAs genes play a vital role in heavy metal transmembrane trafficking in different plants [16]. To date, a single study has reported the function of P-type ATPase involved in active arsenic transport in Pteris vitatta [35]. Nevertheless, the identification and functional characterization of the HMAs gene family in Cucurbits species and their expression under As stress has not been previously evaluated. Thus, in this study, we performed a systematic genome-wide identification and analysis of the HMA family in Citrullus amarus, Citrullus colocynthis, Citrullus lanatus, Citrullus mucusospermus, Cucumis melo, Cucumis sativus, Cucurbita maxima, Cucurbita moschata, Cucurbita pepo and Legenaria siceraria. HMAs genes were identified and studied in terms of their chromosomal location and synteny, phylogeny, conserved motifs, structure, and expression profiles in different tissues of C. pepo under As stress. Therefore, our results provide insights for future investigations into the roles of HMA genes in Cucurbitaceous plants and other species.
2. Materials and Methods
2.1. Identification of HMA genes in Cucurbits
For the identification of members of the HMA gene family in Cucurbit species (Citrullus amarus, Citrullus colocynthis, Citrullus lanatus, Citrullus mucusospermus, Cucumis melo, Cucumis sativus, Cucurbita maxima, Cucurbita moschata, Cucurbita pepo and Legenaria siceraria), the Basic Local Alignment Search Tool (BLAST) of the Cucurbit genomics database v2 (CuGenDB; http://cucurbitgenomics.org/v2) was employed using all the Arabidopsis HMA genes as query [15,36]. To ensure the presence of conserved domains E1-E2 ATPase (IPR008250), Hydrolase (IPR041492), and HMA (IPR006121) in the identified HMA proteins, the Pfam database [37] and the NCBI Conserved Domain Database (https://www.ncbi.nlm.nih.gov/Structure/bwrpsb/bwrpsb) were used. Sequences with no HMA-related domains were excluded from further analysis. Moreover, genomic, coding sequence (CDS), and promoter regions sequences were downloaded from the CuGenDB for confirmed genes. Further, physiochemical parameters such as molecular weight, polypeptide length, and theoretical isoelectric point (pI) value were calculated using the ExPASy ProtParam software (http://web.expasy.org/protparam/). The identified HMA genes were named CamHMA1-CamHMA10; CcoHMA1-CcoHMA9; ClaHMA1-ClaHMA10; CmuHMA1-CmuHMA9; CmeHMA1-CmeHMA10; CsaHMA1-CsaHMA9; CmaHMA1-CmaHMA12; CmoHMA1-CmoHMA12; CpeHMA1-CpeHMA14 and LsiHMA1-LsiHMA8 using the prefix “Cam”, “Cco”, “Cla”, “Cmu”, “Cme”, “Csa”, “Cma”, “Cmo”, “Cpe” and “Lsi” for C. amarus, C. colocynthis, C. lanatus, C. mucusospermus, C. melo, C. sativus, C. maxima, C. moschata, C. pepo and L. siceraria, respectively, followed by “HMA” for heavy metal associated domain and lastly, the progressive number according to their chromosome number and chromosomal positions. A. thaliana HMA sequences were downloaded from the Arabidopsis Information Resource (TAIR, http://www.arabidopsis.org/index.jsp, release 10.0).
2.2. Chromosomal location and Gene Structure of HMA proteins
Physical chromosome location data for each HMA gene in the Cucurbit species was obtained from the CuGenDB database and then displayed by using the MapGene2Chromosome V2 (http://mg2c.iask.in/mg2c_v2.0/) in each chromosome where an HMA gene was found. Moreover, exon-intron gene structure analysis was carried out by submitting merged General Feature Format (GFF3) files of Cucurbitaceous plants to the Biosequence structure tool in TBtools [38].
2.3. Phylogenetic analysis, synteny analysis, and gene duplication events of the Cucurbit HMA family
The amino acid sequences of the Cucurbit species and Arabidopsis were imported into MEGA 7 [39], and multiple sequence alignment was performed using ClustalW [40]] with gap-open and gap-extension penalties of 10 and 0.1, respectively. Alignment was used to build a phylogenetic tree based on the neighbor-joining (NJ) method. After bootstrap analysis with 1000 replicates, the tree was exported into Newick format to display it by using iTOL software (http://itol.embl.de/index.shtml).
Syntenic relationship analysis of the HMA gene family between species of Citrullus, Cucumis and Cucurbita genera was carried out and visualized using TBTools with E-value < 1× 10-10 [38]. Non-synonymous (Ka), synonymous substitution (Ks), and Ka/Ks ratio for duplicated gene pairs were calculated in the Ka/Ks calculation tool (http://services.cbu.uib.no/tools/kaks) using the CDS of Cucurbits. The duplication date was estimated according to the following formula: Million Years Ago (MYA) = Ks/2λ, assuming a clock-like rate (λ) of 6.56 synonymous substitutions per 10−9 years [41].
2.4. Motif Analysis and Promoter Cis-Element Identification
HMA proteins were subjected to Multiple Em for Motif Elicitation (MEME) tool found in the MEME suite (https://meme-suite.org/meme/) to identify common and species-specific motifs in the Cucurbit proteins. The analysis was performed with a maximum number of motifs of ten and optimum motif width from 6 to 50. To identify motif function, discovered MEME motifs were searched in the ExPASy-PROSITE database using the ScanProsite tool (https://prosite.expasy.org/scanprosite/). Furthermore, promoter sequences (2000 bp upstream) from Cucurbit HMA genes were submitted to The PlantCARE database (http://bioinformatics.psb.ugent.be/webtools/plantcare/html) to analyze potential cis-regulatory elements within promoter sequences of the Cucurbit HMA genes.
2.5. Gene ontology (GO) annotation of HMA proteins
The functional annotation, including cellular component, molecular function, and biological process of HMA proteins, was performed using OmicsBox Software (https://www.biobam.com/omicsbox/). The amino acid sequences of HMA proteins were imported into the OmicsBox program to execute three steps: 1) BLASTp against the NCBI non-redundant protein database, 2) mapping and retrieval of GO terms associated with the BLAST results, and 3) annotation of GO terms associated with each query to relate the sequences to known protein function.
2.6. Expression pattern of the HMA family in Cucurbita pepo under different Cu treatments.
RNA-seq gene expression data of CpeHMA genes was retrieved from the NCBI GEO DataSets (Accesion: GSE173716) from a previously published work by [42]. The Fragments Per Kilobase transcripts per Million mapped reads (FPKM) expression values for root, leaf and pollen tissues from Cucurbita pepo exposed to copper oxide (CuO) nanoparticles, bulk CuO (100 mg Kg-1), and CuSO4 (320 mg Kg-1) were used to generate a heatmap and compare the expression of CpHMA1-14 genes identified by using the tydir and ggplot2 packages (https://ggplot2.tidyverse.org/).
2.7. Expression pattern by RT-qPCR of the HMA family in Cucurbita pepo under As treatment
Cucurbita pepo var. cylindrica “Golden” seeds were germinated directly in the soil. Seedlings were transferred to 0, 50, 100, and 200 µM As (V) soil treatments and irrigated with the same water concentration every other day. Roots, leaves, flowers, and fruit tissues were collected at the anthesis stage, as previously reported by [43,44]. Tissues were immediately frozen in liquid nitrogen and stored at -80 ºC. Further, total RNA was isolated from roots, leaves, flowers, and fruits of C. pepo according to the E.Z.N.A. Plant RNA Kit (Omega Bio-Tek) following manufacturer instructions. An equivalent concentration of total RNA of the different tissues was used to synthesize first-strand cDNA with the superscript First-strand synthesis system (Invitrogen). Equal cDNA concentration of samples was used for the qRT-PCR analysis. StepOnePlus Real-Time PCR system (Applied Biosystems, Foster City, CA, USA) was employed for the qRT-PCR analysis with a final reaction volume of 20 µl containing 1 μL of cDNA template, 2 μL of forward and reverse primer at 10 μM, 10 μL SYBR Green PCR Master (ROX) (Roche, Shanghai) and 7 μL of nuclease-free water. Three replicates per treatment were run to compute the average Ct values that were further analyzed by the 2 −ΔΔCt method [45]. The Actin gene was used as an endogenous control to normalize the relative gene expression. HMA Primers for qRT-PCR were designed in Primer3Plus software [46] and listed in Table S1.
3. Results
3.1. Identification of HMA genes in Cucurbits
A total of 103 genes potentially encoding HMA proteins were identified and classified: 10 from Citrullus amarus, 9 from Citrullus colocynthis, 10 from Citrullus lanatus, 9 from Citrullus mucusospermus, 10 from Cucumis melo, 9 from Cucumis sativus, 12 from Cucurbita maxima, 12 from Cucurbita moschata, 14 from Cucurbita pepo and 8 from Legenaria siceraria (Table 1). Moreover, amino acid length, molecular weight, and isoelectric point of Cucurbit HMA proteins were deducted from their protein sequences and are listed in Table S2. In general, the protein length of Cucurbit HMA proteins varied between 356 to 1251 amino acid residues. The molecular weight was determined to range from 37.8 KDa to 143.5 KDa, while the Isoelectric Point (pI) of ranged from 4.96 pH to 9.23 pH.
3.2. Chromosomal location and Gene structure of HMA Genes
HMA genes in Cucurbits were found to be located in various chromosomes with a wide distribution, with the exception of CpeHMA1 from C. pepo that was found on the unanchored scaffold. Nevertheless, not all Cucurbit species showed the presence of HMA genes in all of their chromosomes, as depicted in Figure 1. HMA genes were detected in Chromosomes 1, 2, 4, 5, 6, 10 from Citrullus plants; 2, 5, 7, 8, 9, 11 of C. melo; 1, 2, 4, 6 of C. sativus; 2, 3, 5, 7, 9, 10 of C. maxima and C. moschata; 5, 6, 10, 11, 14, 18, 19 of C. pepo and 1, 3, 4, 6, 9, 10, 11 of L. siceraria. C. amarus and L. siceraria have the highest number of HMA proteins in Chr1 with 3 and 2 members, respectively, while C. lanatus, C. maxima and C. moschata in Chr2 with 3, 4 and 4 members, respectively. Similarly, C. melo have the highest number of HMA proteins in Chr11 with 3 members while C. pepo in Chr5 with 4. Furthermore, similar quantity of members in not only one chromosomes of several species are present due tandem duplicates; C. colocynthis and C. sativus have the highest amount in Chr2 and Chr4 with 2 and 3 members each, respectively and C. mucosospermus have the same highest amount in Chr1 and Chr2 with 2 members each.
Next, we utilized a biosequence structure tool to produce gene structure schematic diagrams and assess the exon/intron arrangement of coding and genome sequences in HMA genes across Cucurbit species. Our analysis of the gene structures indicated significant variation in intron positions, lengths, and numbers across all species studied. Nonetheless, members most closely related shared similar exon/intron structures either according to the number of introns or exon length. Specifically, the number of exons present in Citrullus species ranged from 3 to 25, Cucumis species ranged from 3 to 17, Cucurbita species ranged from 5 to 19 and in L. siceraria ranged from 6 to 16. The detailed gene structure of the Cucurbit HMA genes is in Figure 2a. Further, we also observed that the location of the domains in the HMA proteins follows the forward pattern reported in Arabidopsis [Axelsen & Palgrem., 2001], i.e., HMA proteins in the clade of Cu/Ag showed HMA, E1-E2_ATPase, and hydrolase domains, while, for HMA proteins in the Zn/Co/Cd/Pb exhibited only the E1-E2_ATPase and hydrolase domains (Figure 2b).
3.3. Phylogenetic analysis, synteny analysis, and gene duplication events of the Cucurbit HMA gene family
To examine the phylogenetic relationships among the Cucurbit HMA proteins, an unrooted phylogenetic tree was constructed from alignments of the 103 full-length HMA sequences (Figure 3). The HMA proteins were classified into two major clades, the Zn/Co/Cd/Pb and the Cu/Ag. While the number of HMA genes in Citrullus, Cucumis and L. siceraria species were similar compared to Arabidopsis [36], Cucurbita species exhibited distinct, independent duplication events. Specifically, 4 paralog pairs (CmaHMA7-CmaHMA8, CmoHMA7-CmoHMA8, CpHMA1-CpHMA2 and CpeHMA9-CpeHMA10) were found.
The HMA genes from Citrullus, Cucumis and Cucurbita species were subjected to synteny and gene duplication analysis to further confirm the results from the phylogenetic tree (Figure 4). The syntenic analysis among the HMA genes of the three genera revealed collinearity among species; in the Citrullus genera specifically between C. amarus and C. lanatus with 10 events while in the Cucurbita genera between C. maxima and C. moschata with 12 events where chromosome/position/ HMA copy number are conserved. Conversely, although the Cucumis genera shows 9 events, HMA genes are not positional conserved.
In addition, gene duplication events of the HMA gene family were found in all species, Cucurbita genera being the highest. Interestingly, tandem duplication occurs in pairs labeled as CamHMA2-3, CamHMA4-5, CcoHMA2-3, ClaHMA3-4, CmuHMA3-4, CmeHMA9-10, CmaHMA1-2, CmaHMA7-8, CmoHMA1-2, CmoHMA7-8, CpeHMA4-5, CpeHMA9-10 while in C. sativus and L. siceraria no tandem duplications were found. The non-synonymous rate (Ka), synonymous rate (Ks), the Ka/Ks, and the duplication date using the Ks values of these pairs are presented in Table 2. In general, The Ka/Ks of tandem and segmental duplicates is less than 1, indicating they were under purifying selection. Moreover, the Ks of Citrullus duplication pairs range from 0.27 to 0.47 with divergent times that range from 21 to 36 MYA, similarly, in Cucumis and L. siceraria the Ks value range from 0.27 to 0.44 and 0.33 to 0.44. and the divergent times from 20 to 33 MYA and 24 to 34 MYA, respectively. Nevertheless, Cucurbita duplication pairs range Ks values from 0.01 to 0.53 and divergent times from 1.27 to 40.88 which means a spectrum range of both, synonymous mutation as well diversification time. Interestingly, the tandem duplicates CmaHMA7-8, CmoHMA7-8 and CpeHMA10-CpeHMA9 were speculated to diverge in recent time, < 3 MYA ago, in contrast to the Citrullus, Cucumis and L. siceraria duplicates that resulted >20 Mya.
3.4. Conserved Motif Analysis and Cis-elements of HMA proteins
We identified ten common conserved motifs in HMA proteins by analyzing motif composition with MEME motif analysis, as shown in Figure 5. Common motifs ranged in length from 29 to 50 aa. The position and number of motifs vary according to the substrate-specificity of Cu/Ag and Zn/Cd/Co/Pb clades (Figure S1). Furthermore, ScanProsite analysis indicated that most of the motifs in the Cucurbit HMA family were associated with common functions of ATPase, such as the E1-E2 ATPase phosphorylation site, which is integral to the ATPase function. Details of the ten conserved motifs in common and for each Cucurbit species are given in Table S3.
To identify common putative cis-elements that can influence the expression of Cucurbit HMA genes, 2000 bp DNA sequences upstream of the start codon (ATG) for the HMA proteins for each species were analyzed using the PlantCARE database. There were identified 20 common elements associated with environmental stresses and plant hormonal processes. Table 3 shows all the identified common regulatory elements for the Cucurbit HMA family. The Cis-regulatory elements associated with stress responses and possibly involved in the activation against heavy metals were TC-rich repeats, engaged in defense and stress response, LTR, associated with low-temperatures, TCA cis-elements, related to salicylic acid that have an attenuation against biotic and abiotic stresses, also associated with heavy metal toxicity.
3.5. GO annotation of HMA proteins
The GO analysis performed with Omics Box suggested the putative participation of HMA genes in multiple biological processes, molecular functions, and cellular component (Figure 6 and Table S4). For instance, all 103 HMA proteins identified were predicted to be found in the membrane as a cellular component. Likewise, all HMA proteins were associated with the ATP-hydrolysis and binding activity for molecular function, followed by ATPase-coupled cation transmembrane transport activity. HMA proteins were involved in copper, cadmium, zinc transport, homeostasis, and metal ion binding for biological processes.
3.6. Gene expression pattern of HMA genes in tissues of Cucurbita pepo under Cu and As treatments
We analyzed the expression profile of CPeHMA genes on the basis of their FPKM values from RNA-seq data under Cu stress (Table S5). An expression heatmap was used to visualize the specific gene expression patterns of each gene in root, leaf and pollen tissue under CuO nanoparticles, bulk CuO, and CuSO4 (Figure 7). In general, genes in clade Cu/Ag exhibited high expression levels, whereas genes in clade Zn/Cd/Co/Cu exhibited low expression levels. CpeHMA14, a member of the Cu/Ag clade, exhibited the highest expression among treatments and tissues, whereas CpeHMA12, a member of the Zn/Cd/Co/Pb clade, exhibited the lowest expression. The maximum fold-change expression levels of CpeHMA genes were detected in root and leaf tissues, while low or relative no expression was found in pollen tissues. Members of the Cu/Ag clade, CpeHMA7 and CpeHMA8, were highly expressed under bulk CuO and CuSO4 with a 21-fold and 14-fold change, respectively. On the other hand, the highest expressed gene for leaf was also a member of the Cu/Ag family, CpeHMA6, with 19- and 5-fold alterations under bulk CuSO4 and CuO, respectively. CpeHMA3 and CpeHMA8, members of the Cu/Ag clade, had the maximum expression levels in pollen under CuO NPs, with a 5- and 6-fold change, respectively.
We further evaluated the expression pattern of nine randomly selected C. pepo genes (CpeHMA2, CpeHMA3, CpeHMA5, CpeHMA6, CpeHMA8, CpeHMA10, CpeHMA11, CpeHMA12, and CpeHMA14) in root, leaf, flower and fruit tissues under 50 µM, 100 µM and 200 µM As treatment through RT-qPCR. Notably, no down-regulated genes were observed among all the evaluated HMA genes and tissues. However, genes belonging to the Cu/Ag clade exhibited significantly higher differential expression across all tissues (Figure 8). Interestingly, CpeHMA6 showed upregulation in all As treatments across all tissue types. CpeHMA3 displayed the highest expression in leaf tissue with a 6-fold upregulation under the 50 µM treatment. In roots, CpeHMA6, CpeHMA3, CpeHMA11, CpeHMA2, and CpeHMA8 exhibited differential expression with fold changes of 27, 9, 4, and 3, respectively, in the 200 µM treatment. Moreover, the flower tissue exhibited the most remarkable fold change pattern, as all the studied genes showed upregulation across all treatments. CpeHMA6 consistently exhibited significant expression with a 36-fold change under the 200 µM treatment, while CpeHMA2 and CpeHMA14 showed fold changes of >20 and >30, respectively, among different treatments. In contrast, the expression of the studied CpeHMA genes did not show significant differential expression in leaf and fruit tissues compared to root and flower tissues. The exception was CpeHMA6, which exhibited a notable upregulation of 3-fold under the 200 µM treatment in both leaf and fruit tissues.
4. Discussion
The HMA "heavy metal ATPase transporter" is a type of ATPase known as P1B-type ATPase. It belongs to the P-type ATPase family, comprising ion pumps utilizing energy from ATP hydrolysis to uptake, translocate, compartmentalize, and detoxify heavy metal ions within plant cells [9,16]. Although HMA members have been identified and analyzed in Arabidopsis [36] and several crops such as rice [18], soybean [20], and Populus [19], a comprehensive identification and characterization of this gene family in the Cucurbits have not been performed. In this study, a total of 103 heavy metal ATPase (HMA) proteins were identified across ten different Cucurbit species. The subsequent phylogenetic analysis of the HMA gene family revealed the division of HMA proteins into two distinct subfamilies (Zn/Co/Cd/Pb P1B-ATPase and the Cu/Ag P1B-ATPase) based on their structural and functional characteristics as described in previous studies [15,20,28]. Cucurbita plants, specially C. pepo with 14 members, exhibited higher abundance of HMA proteins, despite having smaller genome size (271.4 Mb for C. maxima, 269.9 Mb for C. moschata and 263Mb for C. pepo) [47,48] in comparison to Cucumis (375Mb for C. melo) [49] and Citrullus (425Mb for C. lanatus ) [50] indicating that genome size may not have a positive correlation with the number of HMA family members. The length of sequences and isoelectric points of proteins significantly varied, indicating a high degree of diversification among the HMA genes in cucurbits. Moreover, the domain structure of HMA genes from the significant clades was similar to the pattern shown in Arabidopsis [15]. Nevertheless, it is important to mention that all HMA Cucurbit genes possess a hydrolase domain since several HMA genes in other species have a lack or disruption of this domain, such as MtHMA8 in Medicago and ZmHMA9 in Zea mays [21,28]. This suggests that all Cucurbit HMA proteins may play an active role in metal transport due to ATP-hydrolysis-dependent mechanisms of energy required for transport [8,9]. Additionally, protein localization is the fundamental concept for understanding interactions at the systems level and the function of transporters is inextricably linked to their subcellular localization [51]. In Arabidopsis, AtHMA2 is expressed mainly in vascular tissues [52], however, previous studies in both Arabidopsis and rice has demonstrated that different HMAs exhibit diverse subcellular localizations [19,53]. In Cucurbits all HMA proteins were predicted to localize within the cell membrane. Membrane proteins play a vital role in regulating plant responses to heavy-metal stress, as they facilitate the transport of metals across membranes, thereby contributing to metal homeostasis and detoxification processes [54]. Expression of CsHMA3 and CsHMA4 were found to confer tolerance to Cd and Zn by metal efflux tolerance and accumulation of Cd and Pb through sequestration, proving being part of the the Zn/Co/Cd/Pb clade and suggesting its role in plant translocation from plasma membrane and bioaccumulation of these metals into the vacuoles [34]. However, experimental validation is needed to locate HMAs and understand the role in other Cucurbits.
Furthermore, an important feature of P1B-ATPases is the presence of soluble metal binding domains (MBDs) that regulate transport activity [55]. The conserved structure characteristics of two cysteines (CxxC) of the HMA domain give HMA genes the basic function of binding metal ions through thiol groups [56,57]. P1B-type ATPase are capable of driving the efflux out of cells of both essential transition metal ions (e.g., Zn2+, Cu+, and Co2+) and toxic metal ions (e.g., Ag+, Cd2+, Pb2+) contributing to their homeostasis maintenance [15,32]. Previous studies on members of the HMA gene family in Arabidopsis focused on heavy metal stress. Several genes, e.i., AtHMA4, AtHMA2, and AtHMA3, have been identified as Cd transporters involved in transporting Cd across the cell membrane and from the cytoplasm to the vacuole [10,11]. However, the molecular basis of HMA metal ion specificity remains unclear [58]. According to [8] HMA proteins appear to have functional roles in transporting manganese, iron, nickel, and other thiophilic heavy metals and metalloids such as arsenic. In plants, arsenic can easily enter through phosphate (P) transporters (arsenate) and aquaporin channels (arsenite), inhibiting plant growth and reducing crop yield [59,60,61]. After entering the plant, arsenic can be sequestered in the form of As-cysteine-rich peptides such as phytochelatins and then translocated into vacuoles mainly by ABC transporters subfamily C (ABCC) [62,63,64]. Nonetheless, different studies have reported alternative and independent arsenic transporters, such as the silicon transporters Ls1 and Ls2 that transport As (III) and the peptide transporter OsPTR7 associated with the translocation of methylated-As species in Oryza sativa [24,59,65,66].
Although HMA proteins have not been previously characterized in the arsenic transport, transcriptomic analysis have shown that P-type ATPase genes were up-regulated in roots and shoots of the hyper-accumulator Pteris vitatta in response to arsenic, which implies the role of P-type ATPase in the translocation of this metalloid [35]. Additionally, vacuolar proteomics showed that P-type ATPase were highly abundant compared to other metal transporters under arsenic stress. Likewise, previous studies have also reported the participation of non-elucidated transporters for arsenic in Pteris vitatta when treated with a mix of Ag-As due to the inhibition of the entrance, translocation, and the enhancement of As tolerance when Ag and As are supplemented simultaneously [67,68]. Therefore, metal homeostasis in plants must be regulated by several complex processes [54] and the collaboration of transporters in different tissues may play an important role in plant metal distribution [19].
Hence, we analyzed the gene expression levels from C. pepo genes (CpeHMA) in root, leaf and pollen from C. pepo under Cu treatments [42], a well-recognized HMA-related metal, and under arsenic stress in root, leaf, flower and fruit. Under either Cu or As, genes that belongs to Zn/Co/Cd/Pb clade; CpeHMA2, CpeHMA4, CpeHMA5, CpeHMA9, CpeHMA10, CpeHMA11, and CpeHMA12 exhibited low to no expression levels in all tissues, whereas genes in the Cu/Ag clade such as CpeHMA3, CpeHMA6, CpeHMA7, CpeHMA8 and CpeHMA14 in the Cu treatments and CpeHMA3, CpeHMA6, CpeHMA8, and CpeHMA14 in the As treatments were highly expressed. CpeHMA6 showed a high up-regulation in leaf under Cu treatment and in root and flower tissues under As stress. It was observed from the phylogenetic tree that CpHMA6 is orthologous to the AtHMA8 from Arabidopsis, which is related to the Cu transport through the thylakoid membrane [69,70,71]. Cu is an essential metal due its function as an enzyme cofactor for a number of physiological processes [72]. In the other hand, As (V) can act as a P analog in the phosphorylation process that occurs in the thylakoid membrane, leading to the disruption of the ATP production process and thus threatening the energy homeostasis of the cell [59,73].
Moreover, the CpeHMA3 gene, which is classified within the Cu/Ag clade, exhibited significant upregulation in response to both Cu and As treatments. Notably, the Cu/Ag clade lacks annotated HMA domains, a distinctive feature of this particular clade. The absence of highly conserved regions, particularly those associated with similar functionality observed in other species, can have an impact on both the affinity of the protein for various ionic metals and its inherent characteristics, such as heavy metal binding properties [13]. Moreover, CpeHMA8 is orthologue of AtHMA5 and OsHMA5 in Arabidopsis and rice, respectively, while CpeHMA14 is orthologue of AtHMA7. AtHMA5 is located in plasma membrane and is involved in the Cu translocation from roots to shoots or Cu detoxification of roots [74]. OsHMA5 is involved in loading Cu to the xylem of the roots and other organs [33]. Nevertheless, a study in Populus trichocarpa suggested that PtHMA5 may differ in the function from that of AtHMA5 and OsHMA5, where it was found to have a significant role in Ag detoxification in addition to Cu detoxification [19]. High expressed CpeHMA7 in root tissues under CuSO4 is orthologous of AtHMA7 also known as RAN1. The AtHMA7 gene has been recognized as an ATP-dependent copper transporter that interacts with the ethylene receptor ETR1, which is primarily found in the endoplasmic reticulum regulating plant growth and development [75]. Therefore, considering the high expression observed in CpeHMA3, CpeHMA6, CpeHMA7, CpeHMA8, and CpeHMA14 across the different tissues under Cu and As stress it is plausible to hypothesize that HMA proteins may play a role in the transportation and tolerance mechanisms of arsenic in Cucurbita pepo, similar to their involvement in copper transport. However, further study and confirmation are required to elucidate the specific molecular pathways by which these CpeHMA genes respond to arsenic stress.
Moreover, through the examination of cis-regulatory elements in the HMA gene family, it has been observed that all genes harbor multiple cis-elements associated with abiotic stress, including those related to heavy metals [27,76]. Other cis-elements identified were the ABRE, ARE elements, LTR, and TC-rich repeats, which are associated with various stress responses, such as abscisic acid stress, anaerobic induction, low-temperature stress, defense mechanisms, and oxidative stress, which may be involved in generating a response to arsenic-induced oxidative stress [26,28]. Consequently, these findings suggest that the HMA genes in Cucurbits may be activated and potentially play a role in responding to other stress conditions.
5. Conclusions
In plants, metal transporters play vital roles in distributing and transforming essential, non-essential, and even toxic metal ions. In this study, we performed a comprehensive analysis of the HMA gene family in ten Cucurbit species. A total of 103 HMA genes from species of the Cucurbitaceous family were characterized and classified into two distinct groups based on phylogenetic analysis and their structural characteristics. According to the findings, the Cucurbit HMA genes had conserved or divergent gene structures, protein motif patterns, and cis-regulatory elements. The expression profiles of CpeHMA genes in various tissues/organs of C. pepo in response to As stress indicate that the members of this gene family might be involved in transporting As metal ions across Cucurbit tissues. This information is valuable for functional investigation and understanding alternative molecular mechanisms in response to As stress in Cucurbits and other crops.
Supplementary Materials
The following supporting information can be downloaded at: Preprints.org, Figure S1: Position of the ten common motifs in protein sequences of Cucurbits. Sequence name is shown in the left side of each sequence and the legend of each motif with the typed sequence is presented below all sequences; Table S1: Primer sequences of selected CpeHMA genes evaluated by RTqPCR under Arsenic stress; Table S2: Basic information on the HMA gene family in ten Cucurbit species; Table S3: Common Conserved Motifs Present in HMA Family in Cucurbit species; Table S4: Gene ontology annotation results of HMA genes in Cucurbit species; Table S5: Normalized RNA-seq data from C. pepo under Cu stress.
Author Contributions
Conceptualization, Umesh K. Reddy and Nagamani Balagurusamy; Data curation, Gerardo Flores-Iga; Formal analysis, Gerardo Flores-Iga and Celeste Gracia-Rodriguez; Funding acquisition, Padma Nimmakayala and Umesh K. Reddy; Investigation, Gerardo Flores-Iga; Methodology, Gerardo Flores-Iga, Carlos Lopez-Ortiz and Celeste Gracia-Rodriguez; Project administration, Padma Nimmakayala and Nagamani Balagurusamy; Software, Gerardo Flores-Iga; Supervision, Carlos Lopez-Ortiz, Padma Nimmakayala, Umesh K. Reddy and Nagamani Balagurusamy; Validation, Gerardo Flores-Iga; Visualization, Nagamani Balagurusamy; Writing – original draft, Gerardo Flores-Iga and Carlos Lopez-Ortiz; Writing – review & editing, Carlos Lopez-Ortiz, Umesh K. Reddy and Nagamani Balagurusamy.
Funding
Please add: This research was funded by the National Institute of Food and Agriculture grant number 2023-38821-39586 and 2021-38821-34598.
Data Availability Statement
The data presented in this study its available in the article supplementary material.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Chomicki, Guillaume, Hanno Schaefer, and Susanne S Renner. "Origin and Domestication of Cucurbitaceae Crops: Insights from Phylogenies, Genomics and Archaeology." New Phytologist 226, no. 5 (2020): 1240-55. [CrossRef]
- Xie, Xiulan, Zhouqing He, Nifan Chen, Zizhong Tang, Qiang Wang, and Yi Cai. "The Roles of Environmental Factors in Regulation of Oxidative Stress in Plant." BioMed research international 2019 (2019). [CrossRef]
- ul Haq, Saeed, Abid Khan, Muhammad Ali, Abdul Mateen Khattak, Wen-Xian Gai, Huai-Xia Zhang, Ai-Min Wei, and Zhen-Hui Gong. "Heat Shock Proteins: Dynamic Biomolecules to Counter Plant Biotic and Abiotic Stresses." International journal of molecular sciences 20, no. 21 (2019): 5321. [CrossRef]
- Chen, Yanshan, Yong-He Han, Yue Cao, Yong-Guan Zhu, Bala Rathinasabapathi, and Lena Q Ma. "Arsenic Transport in Rice and Biological Solutions to Reduce Arsenic Risk from Rice." Frontiers in Plant Science 8 (2017): 268. [CrossRef]
- Podgorski, Joel, and Michael Berg. "Global Threat of Arsenic in Groundwater." Science 368, no. 6493 (2020): 845-50. [CrossRef]
- Garg, Neera, and Priyanka Singla. "Arsenic Toxicity in Crop Plants: Physiological Effects and Tolerance Mechanisms." Environmental chemistry letters 9 (2011): 303-21. [CrossRef]
- Raza, Ali, Ali Razzaq, Sundas Saher Mehmood, Xiling Zou, Xuekun Zhang, Yan Lv, and Jinsong Xu. "Impact of Climate Change on Crops Adaptation and Strategies to Tackle Its Outcome: A Review." Plants 8, no. 2 (2019): 34. [CrossRef]
- Smith, Aaron T, Kyle P Smith, and Amy C Rosenzweig. "Diversity of the Metal-Transporting P 1b-Type Atpases." JBIC Journal of Biological Inorganic Chemistry 19 (2014): 947-60. [CrossRef]
- Palmgren, Michael G, and Poul Nissen. "P-Type Atpases." Annual review of biophysics 40 (2011): 243-66.
- Mills, Rebecca F, Gerard C Krijger, Paul J Baccarini, JL Hall, and Lorraine E Williams. "Functional Expression of Athma4, a P1b-Type Atpase of the Zn/Co/Cd/Pb Subclass." The Plant Journal 35, no. 2 (2003): 164-76.
- Gravot, Antoine, Aurélie Lieutaud, Frédéric Verret, Pascaline Auroy, Alain Vavasseur, and Pierre Richaud. "Athma3, a Plant P1b-Atpase, Functions as a Cd/Pb Transporter in Yeast." FEBS letters 561, no. 1-3 (2004): 22-28. [CrossRef]
- Hussain, Dawar, Michael J Haydon, Yuwen Wang, Edwin Wong, Sarah M Sherson, Jeff Young, James Camakaris, Jeffrey F Harper, and Christopher S Cobbett. "P-Type Atpase Heavy Metal Transporters with Roles in Essential Zinc Homeostasis in Arabidopsis." The plant cell 16, no. 5 (2004): 1327-39. [CrossRef]
- Li, Jiaming, Minghui Zhang, Jian Sun, Xinrui Mao, Jingguo Wang, Hualong Liu, Hongliang Zheng, Xianwei Li, Hongwei Zhao, and Detang Zou. "Heavy Metal Stress-Associated Proteins in Rice and Arabidopsis: Genome-Wide Identification, Phylogenetics, Duplication, and Expression Profiles Analysis." Frontiers in Genetics 11 (2020): 477. [CrossRef]
- Zorrig, Walid, Chedly Abdelly, and Pierre Berthomieu. "The Phylogenetic Tree Gathering the Plant Zn/Cd/Pb/Co P1b-Atpases Appears to Be Structured According to the Botanical Families." Comptes Rendus Biologies 334, no. 12 (2011): 863-71. [CrossRef]
- Williams, Lorraine E, and Rebecca F Mills. "P1b-Atpases–an Ancient Family of Transition Metal Pumps with Diverse Functions in Plants." Trends in plant science 10, no. 10 (2005): 491-502. [CrossRef]
- Argüello, José M, Elif Eren, and Manuel González-Guerrero. "The Structure and Function of Heavy Metal Transport P 1b-Atpases." Biometals 20 (2007): 233-48. [CrossRef]
- Axelsen, Kristian B, and Michael G Palmgren. "Evolution of Substrate Specificities in the P-Type Atpase Superfamily." Journal of molecular evolution 46 (1998): 84-101. [CrossRef]
- Takahashi, Ryuichi, Khurram Bashir, Yasuhiro Ishimaru, Naoko K Nishizawa, and Hiromi Nakanishi. "The Role of Heavy-Metal Atpases, Hmas, in Zinc and Cadmium Transport in Rice." Plant signaling & behavior 7, no. 12 (2012): 1605-07. [CrossRef]
- Li, Dandan, Xuemei Xu, Xiaoqing Hu, Quangang Liu, Zhanchao Wang, Haizhen Zhang, Han Wang, Ming Wei, Hanzeng Wang, and Haimei Liu. "Genome-Wide Analysis and Heavy Metal-Induced Expression Profiling of the Hma Gene Family in Populus Trichocarpa." Frontiers in Plant Science 6 (2015): 1149. [CrossRef]
- Fang, Xiaolong, Lei Wang, Xiaojuan Deng, Peng Wang, Qibin Ma, Hai Nian, Yingxiang Wang, and Cunyi Yang. "Genome-Wide Characterization of Soybean P 1b-Atpases Gene Family Provides Functional Implications in Cadmium Responses." BMC genomics 17, no. 1 (2016): 1-15. [CrossRef]
- Zhiguo, E, LI Tingting, CHEN Chen, and WANG Lei. "Genome-Wide Survey and Expression Analysis of P1b-Atpases in Rice, Maize and Sorghum." Rice Science 25, no. 4 (2018): 208-17.
- Zhang, Chiran, Qianhui Yang, Xiaoqin Zhang, Xian Zhang, Tongyuan Yu, Yuhuan Wu, Yunxia Fang, and Dawei Xue. "Genome-Wide Identification of the Hma Gene Family and Expression Analysis under Cd Stress in Barley." Plants 10, no. 9 (2021): 1849. [CrossRef]
- Mills, Rebecca F, Kerry A Peaston, John Runions, and Lorraine E Williams. "Hvhma2, a P1b-Atpase from Barley, Is Highly Conserved among Cereals and Functions in Zn and Cd Transport." (2012). [CrossRef]
- Khan, Nadeem, Frank M You, Raju Datla, Sridhar Ravichandran, Bosen Jia, and Sylvie Cloutier. "Genome-Wide Identification of Atp Binding Cassette (Abc) Transporter and Heavy Metal Associated (Hma) Gene Families in Flax (Linum Usitatissimum L.)." BMC genomics 21 (2020): 1-14. [CrossRef]
- Li, Nannan, Hua Xiao, Juanjuan Sun, Shufeng Wang, Jingchao Wang, Peng Chang, Xinbin Zhou, Bo Lei, Kun Lu, and Feng Luo. "Genome-Wide Analysis and Expression Profiling of the Hma Gene Family in Brassica Napus under Cd Stress." Plant and Soil 426 (2018): 365-81. [CrossRef]
- Manzoor, Muhammad Aamir, Xi Cheng, Guohui Li, Xueqiang Su, Muhammad Abdullah, and Yongping Cai. "Gene Structure, Evolution and Expression Analysis of the P-Atpase Gene Family in Chinese Pear (Pyrus Bretschneideri)." Computational Biology and Chemistry 88 (2020): 107346. [CrossRef]
- Fan, Wei, Changying Liu, Boning Cao, Meiling Qin, Dingpei Long, Zhonghuai Xiang, and Aichun Zhao. "Genome-Wide Identification and Characterization of Four Gene Families Putatively Involved in Cadmium Uptake, Translocation and Sequestration in Mulberry." Frontiers in Plant Science 9 (2018): 879. [CrossRef]
- Ma, Yitong, Na Wei, Qiuxia Wang, Zhipeng Liu, and Wenxian Liu. "Genome-Wide Identification and Characterization of the Heavy Metal Atpase (Hma) Gene Family in Medicago Truncatula under Copper Stress." International Journal of Biological Macromolecules 193 (2021): 893-902. [CrossRef]
- Ye, Xueling, Changying Liu, Huiling Yan, Yan Wan, Qi Wu, Xiaoyong Wu, Gang Zhao, Liang Zou, and Dabing Xiang. "Genome-Wide Identification and Transcriptome Analysis of the Heavy Metal-Associated (Hma) Gene Family in Tartary Buckwheat and Their Regulatory Roles under Cadmium Stress." Gene 847 (2022): 146884. [CrossRef]
- Kim, Yu-Young, Hyunju Choi, Shoji Segami, Hyung-Taeg Cho, Enrico Martinoia, Masayoshi Maeshima, and Youngsook Lee. "Athma1 Contributes to the Detoxification of Excess Zn (Ii) in Arabidopsis." The Plant Journal 58, no. 5 (2009): 737-53. [CrossRef]
- Morel, Mélanie, Jérôme Crouzet, Antoine Gravot, Pascaline Auroy, Nathalie Leonhardt, Alain Vavasseur, and Pierre Richaud. "Athma3, a P1b-Atpase Allowing Cd/Zn/Co/Pb Vacuolar Storage in Arabidopsis." Plant physiology 149, no. 2 (2009): 894-904. [CrossRef]
- Liu, Huan, Haixia Zhao, Longhua Wu, Anna Liu, Fang-Jie Zhao, and Wenzhong Xu. "Heavy Metal Atpase 3 (Hma3) Confers Cadmium Hypertolerance on the Cadmium/Zinc Hyperaccumulator Sedum Plumbizincicola." New Phytologist 215, no. 2 (2017): 687-98. [CrossRef]
- Deng, Fenglin, Naoki Yamaji, Jixing Xia, and Jian Feng Ma. "A Member of the Heavy Metal P-Type Atpase Oshma5 Is Involved in Xylem Loading of Copper in Rice." Plant physiology 163, no. 3 (2013): 1353-62. [CrossRef]
- Feng, Shengjun, Yanghui Shen, Huinan Xu, Junyang Dong, Kexin Chen, Yu Xiang, Xianda Jiang, Chenjie Yao, Tao Lu, and Weiwei Huan. "Rna-Seq Identification of Cd Responsive Transporters Provides Insights into the Association of Oxidation Resistance and Cd Accumulation in Cucumis Sativus L." Antioxidants 10, no. 12 (2021): 1973. [CrossRef]
- Yan, Huili, Yiwei Gao, Lulu Wu, Luyao Wang, Tian Zhang, Changhua Dai, Wenxiu Xu, Lu Feng, Mi Ma, and Yong-Guan Zhu. "Potential Use of the Pteris Vittata Arsenic Hyperaccumulation-Regulation Network for Phytoremediation." Journal of hazardous materials 368 (2019): 386-96. [CrossRef]
- Axelsen, Kristian B., and Michael G. Palmgren. "Inventory of the Superfamily of P-Type Ion Pumps in Arabidopsis." Plant physiology 126, no. 2 (2001): 696-706. [CrossRef]
- Mistry, Jaina, Sara Chuguransky, Lowri Williams, Matloob Qureshi, Gustavo A Salazar, Erik L L Sonnhammer, Silvio C E Tosatto, Lisanna Paladin, Shriya Raj, Lorna J Richardson, Robert D Finn, and Alex Bateman. "Pfam: The Protein Families Database in 2021." Nucleic Acids Research 49, no. D1 (2020): D412-D19. [CrossRef]
- Chen, Chengjie, Hao Chen, Yi Zhang, Hannah R. Thomas, Margaret H. Frank, Yehua He, and Rui Xia. "Tbtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data." Molecular Plant 13, no. 8 (2020): 1194-202. [CrossRef]
- Kumar, Sudhir, Glen Stecher, and Koichiro Tamura. "Mega7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets." Molecular Biology and Evolution 33, no. 7 (2016): 1870-74. [CrossRef]
- Larkin, M.A., G. Blackshields, N.P. Brown, R. Chenna, P.A. McGettigan, H. McWilliam, F. Valentin, I.M. Wallace, A. Wilm, R. Lopez, J.D. Thompson, T.J. Gibson, and D.G. Higgins. "Clustal W and Clustal X Version 2.0." Bioinformatics 23, no. 21 (2007): 2947-48. [CrossRef]
- Moniz de Sá, M, and G Drouin. "Phylogeny and Substitution Rates of Angiosperm Actin Genes." Molecular Biology and Evolution 13, no. 9 (1996): 1198-212. [CrossRef]
- Marmiroli, Marta, Luca Pagano, Riccardo Rossi, Roberto De La Torre-Roche, Giovanni Orazio Lepore, Roberta Ruotolo, Gianluca Gariani, Valentina Bonanni, Simone Pollastri, and Alessandro Puri. "Copper Oxide Nanomaterial Fate in Plant Tissue: Nanoscale Impacts on Reproductive Tissues." Environmental science & technology 55, no. 15 (2021): 10769-83. [CrossRef]
- Obrero, Ángeles, Jose V. Die, Belén Román, Pedro Gómez, Salvador Nadal, and Clara I. González-Verdejo. "Selection of Reference Genes for Gene Expression Studies in Zucchini (Cucurbita Pepo) Using Qpcr." Journal of Agricultural and Food Chemistry 59, no. 10 (2011): 5402-11. [CrossRef]
- Obrero, Ángeles, Clara I. González-Verdejo, Jose V. Die, Pedro Gómez, Mercedes Del Río-Celestino, and Belén Román. "Carotenogenic Gene Expression and Carotenoid Accumulation in Three Varieties of Cucurbita Pepo During Fruit Development." Journal of Agricultural and Food Chemistry 61, no. 26 (2013): 6393-403. [CrossRef]
- Livak, Kenneth J., and Thomas D. Schmittgen. "Analysis of Relative Gene Expression Data Using Real-Time Quantitative Pcr and the 2−Δδct Method." Methods 25, no. 4 (2001): 402-08. [CrossRef]
- Untergasser, Andreas, Harm Nijveen, Xiangyu Rao, Ton Bisseling, René Geurts, and Jack A.M. Leunissen. "Primer3plus, an Enhanced Web Interface to Primer3." Nucleic Acids Research 35, no. suppl_2 (2007): W71-W74. [CrossRef]
- Sun, Honghe, Shan Wu, Guoyu Zhang, Chen Jiao, Shaogui Guo, Yi Ren, Jie Zhang, Haiying Zhang, Guoyi Gong, and Zhangcai Jia. "Karyotype Stability and Unbiased Fractionation in the Paleo-Allotetraploid Cucurbita Genomes." Molecular Plant 10, no. 10 (2017): 1293-306. [CrossRef]
- Montero-Pau, Javier, José Blanca, Aureliano Bombarely, Peio Ziarsolo, Cristina Esteras, Carlos Martí-Gómez, María Ferriol, Pedro Gómez, Manuel Jamilena, and Lukas Mueller. "De Novo Assembly of the Zucchini Genome Reveals a Whole-Genome Duplication Associated with the Origin of the Cucurbita Genus." Plant biotechnology journal 16, no. 6 (2018): 1161-71.
- Garcia-Mas, Jordi, Andrej Benjak, Walter Sanseverino, Michael Bourgeois, Gisela Mir, Víctor M. González, Elizabeth Hénaff, Francisco Câmara, Luca Cozzuto, Ernesto Lowy, Tyler Alioto, Salvador Capella-Gutiérrez, Jose Blanca, Joaquín Cañizares, Pello Ziarsolo, Daniel Gonzalez-Ibeas, Luis Rodríguez-Moreno, Marcus Droege, Lei Du, Miguel Alvarez-Tejado, Belen Lorente-Galdos, Marta Melé, Luming Yang, Yiqun Weng, Arcadi Navarro, Tomas Marques-Bonet, Miguel A. Aranda, Fernando Nuez, Belén Picó, Toni Gabaldón, Guglielmo Roma, Roderic Guigó, Josep M. Casacuberta, Pere Arús, and Pere Puigdomènech. "The Genome of Melon (<I>Cucumis Melo</I> L.)." Proceedings of the National Academy of Sciences 109, no. 29 (2012): 11872-77. [CrossRef]
- Guo, Shaogui, Jianguo Zhang, Honghe Sun, Jerome Salse, William J. Lucas, Haiying Zhang, Yi Zheng, Linyong Mao, Yi Ren, Zhiwen Wang, Jiumeng Min, Xiaosen Guo, Florent Murat, Byung-Kook Ham, Zhaoliang Zhang, Shan Gao, Mingyun Huang, Yimin Xu, Silin Zhong, Aureliano Bombarely, Lukas A. Mueller, Hong Zhao, Hongju He, Yan Zhang, Zhonghua Zhang, Sanwen Huang, Tao Tan, Erli Pang, Kui Lin, Qun Hu, Hanhui Kuang, Peixiang Ni, Bo Wang, Jingan Liu, Qinghe Kou, Wenju Hou, Xiaohua Zou, Jiao Jiang, Guoyi Gong, Kathrin Klee, Heiko Schoof, Ying Huang, Xuesong Hu, Shanshan Dong, Dequan Liang, Juan Wang, Kui Wu, Yang Xia, Xiang Zhao, Zequn Zheng, Miao Xing, Xinming Liang, Bangqing Huang, Tian Lv, Junyi Wang, Ye Yin, Hongping Yi, Ruiqiang Li, Mingzhu Wu, Amnon Levi, Xingping Zhang, James J. Giovannoni, Jun Wang, Yunfu Li, Zhangjun Fei, and Yong Xu. "The Draft Genome of Watermelon (Citrullus Lanatus) and Resequencing of 20 Diverse Accessions." Nature Genetics 45, no. 1 (2013): 51-58. [CrossRef]
- David, Rakesh, Caitlin S Byrt, Stephen D Tyerman, Matthew Gilliham, and Stefanie Wege. "Roles of Membrane Transporters: Connecting the Dots from Sequence to Phenotype." Annals of Botany 124, no. 2 (2019): 201-08. [CrossRef]
- Wong, Chong Kum Edwin, Renée S Jarvis, Sarah M Sherson, and Christopher S Cobbett. "Functional Analysis of the Heavy Metal Binding Domains of the Zn/Cd-Transporting Atpase, Hma2, in Arabidopsis Thaliana." New Phytologist 181, no. 1 (2009): 79-88.
- Leonhardt, Nathalie, Pierre Cun, Pierre Richaud, and Alain Vavasseur. "Zn/Cd/Co/Pb P1b-Atpases in Plants, Physiological Roles and Biological Interest." In Metal Toxicity in Plants: Perception, Signaling and Remediation, 227-48: Springer, 2011.
- Wu, Danxia, Muhammad Saleem, Tengbing He, and Guandi He. "The Mechanism of Metal Homeostasis in Plants: A New View on the Synergistic Regulation Pathway of Membrane Proteins, Lipids and Metal Ions." Membranes 11, no. 12 (2021): 984. [CrossRef]
- Smith, Aaron T., Dulmini Barupala, Timothy L. Stemmler, and Amy C. Rosenzweig. "A New Metal Binding Domain Involved in Cadmium, Cobalt and Zinc Transport." Nature Chemical Biology 11, no. 9 (2015): 678-84. [CrossRef]
- Arnesano, Fabio, Lucia Banci, Ivano Bertini, Simone Ciofi-Baffoni, Elena Molteni, David L. Huffman, and Thomas V. O'Halloran. "Metallochaperones and Metal-Transporting Atpases: A Comparative Analysis of Sequences and Structures." Genome Research 12, no. 2 (2002): 255-71. [CrossRef]
- Furukawa, Yoshiaki, Carolyn Lim, Takehiko Tosha, Koki Yoshida, Tomoaki Hagai, Shuji Akiyama, Shoji Watanabe, Kenta Nakagome, and Yoshitsugu Shiro. "Identification of a Novel Zinc-Binding Protein, C1orf123, as an Interactor with a Heavy Metal-Associated Domain." PLOS ONE 13, no. 9 (2018): e0204355. [CrossRef]
- Huang, Qingyu, Wenmin Qiu, Miao Yu, Shaocui Li, Zhuchou Lu, Yue Zhu, Xianzhao Kan, and Renying Zhuo. "Genome-Wide Characterization of Sedum Plumbizincicola&Nbsp;Hma Gene Family Provides Functional Implications in Cadmium Response." Plants 11, no. 2 (2022): 215. [CrossRef]
- Abbas, Ghulam, Behzad Murtaza, Irshad Bibi, Muhammad Shahid, Nabeel Khan Niazi, Muhammad Imran Khan, Muhammad Amjad, Munawar Hussain, and Natasha. "Arsenic Uptake, Toxicity, Detoxification, and Speciation in Plants: Physiological, Biochemical, and Molecular Aspects." International Journal of Environmental Research and Public Health 15, no. 1 (2018): 59.
- Garbinski, Luis D, Barry P Rosen, and Jian Chen. "Pathways of Arsenic Uptake and Efflux." Environment international 126 (2019): 585-97. [CrossRef]
- Li, Nannan, Jingchao Wang, and Won-Yong Song. "Arsenic Uptake and Translocation in Plants." Plant and Cell Physiology 57, no. 1 (2016): 4-13. [CrossRef]
- Bali, Aditi Shreeya, and Gagan Preet Singh Sidhu. "Arsenic Acquisition, Toxicity and Tolerance in Plants-from Physiology to Remediation: A Review." Chemosphere 283 (2021): 131050.
- Song, Won-Yong, Jiyoung Park, David G Mendoza-Cózatl, Marianne Suter-Grotemeyer, Donghwan Shim, Stefan Hörtensteiner, Markus Geisler, Barbara Weder, Philip A Rea, and Doris Rentsch. "Arsenic Tolerance in Arabidopsis Is Mediated by Two Abcc-Type Phytochelatin Transporters." Proceedings of the National Academy of Sciences 107, no. 49 (2010): 21187-92. [CrossRef]
- Song, Won-Yong, Tomohiro Yamaki, Naoki Yamaji, Donghwi Ko, Ki-Hong Jung, Miho Fujii-Kashino, Gynheung An, Enrico Martinoia, Youngsook Lee, and Jian Feng Ma. "A Rice Abc Transporter, Osabcc1, Reduces Arsenic Accumulation in the Grain." Proceedings of the National Academy of Sciences 111, no. 44 (2014): 15699-704.
- Yamaji, Naoki, Gen Sakurai, Namiki Mitani-Ueno, and Jian Feng Ma. "Orchestration of Three Transporters and Distinct Vascular Structures in Node for Intervascular Transfer of Silicon in Rice." Proceedings of the National Academy of Sciences 112, no. 36 (2015): 11401-06. [CrossRef]
- Tang, Zhong, Yi Chen, Fei Chen, Yuchen Ji, and Fang-Jie Zhao. "Osptr7 (Osnpf8. 1), a Putative Peptide Transporter in Rice, Is Involved in Dimethylarsenate Accumulation in Rice Grain." Plant and Cell Physiology 58, no. 5 (2017): 904-13. [CrossRef]
- Mathews, Shiny, Bala Rathinasabapathi, and Lena Q. Ma. "Uptake and Translocation of Arsenite by Pteris Vittata L.: Effects of Glycerol, Antimonite and Silver." Environmental Pollution 159, no. 12 (2011): 3490-95. [CrossRef]
- Nagarajan, V., and Stephen Ebbs. "Transport of Arsenite by the Arsenic Hyperaccumulating Brake Fern Pteris Vittata Is Inhibited by Monovalent Silver." Indian Journal of Plant Physiology 12 (2007): 312-16.
- Mayerhofer, Hubert, Emeline Sautron, Norbert Rolland, Patrice Catty, Daphné Seigneurin-Berny, Eva Pebay-Peyroula, and Stéphanie Ravaud. "Structural Insights into the Nucleotide-Binding Domains of the P1b-Type Atpases Hma6 and Hma8 from Arabidopsis Thaliana." PLOS ONE 11, no. 11 (2016): e0165666. [CrossRef]
- Sautron, Emeline, Cécile Giustini, ThuyVan Dang, Lucas Moyet, Daniel Salvi, Serge Crouzy, Norbert Rolland, Patrice Catty, and Daphné Seigneurin-Berny. "Identification of Two Conserved Residues Involved in Copper Release from Chloroplast P<Sub>Ib-1</Sub>-Atpases *." Journal of Biological Chemistry 291, no. 38 (2016): 20136-48. [CrossRef]
- Sautron, Emeline, Hubert Mayerhofer, Cécile Giustini, Danièle Pro, Serge Crouzy, Stéphanie Ravaud, Eva Pebay-Peyroula, Norbert Rolland, Patrice Catty, and Daphné Seigneurin-Berny. "Hma6 and Hma8 Are Two Chloroplast Cu+-Atpases with Different Enzymatic Properties." Bioscience Reports 35, no. 3 (2015). [CrossRef]
- Shingles, Richard, Larry E Wimmers, and Richard E McCarty. "Copper Transport across Pea Thylakoid Membranes." Plant physiology 135, no. 1 (2004): 145-51. [CrossRef]
- Gresser, M. J. "Adp-Arsenate. Formation by Submitochondrial Particles under Phosphorylating Conditions." Journal of Biological Chemistry 256, no. 12 (1981): 5981-83. [CrossRef]
- Kobayashi, Yuriko, Keishi Kuroda, Keisuke Kimura, Jennafer L Southron-Francis, Aya Furuzawa, Kazuhiko Kimura, Satoshi Iuchi, Masatomo Kobayashi, Gregory J Taylor, and Hiroyuki Koyama. "Amino Acid Polymorphisms in Strictly Conserved Domains of a P-Type Atpase Hma5 Are Involved in the Mechanism of Copper Tolerance Variation in Arabidopsis." Plant physiology 148, no. 2 (2008): 969-80. [CrossRef]
- Yang, Yanzhi, Chen Hao, Jianmei Du, Lei Xu, Zhonglong Guo, Dong Li, Huaqing Cai, Hongwei Guo, and Lei Li. "The Carboxy Terminal Transmembrane Domain of Spl7 Mediates Interaction with Ran1 at the Endoplasmic Reticulum to Regulate Ethylene Signalling in Arabidopsis." New Phytologist 236, no. 3 (2022): 878-92. [CrossRef]
- Freitas, Elinea O., Bruno P. Melo, Isabela T. Lourenço-Tessutti, Fabrício B. M. Arraes, Regina M. Amorim, Maria E. Lisei-de-Sá, Julia A. Costa, Ana G. B. Leite, Muhammad Faheem, Márcio A. Ferreira, Carolina V. Morgante, Elizabeth P. B. Fontes, and Maria F. Grossi-de-Sa. "Identification and Characterization of the Gmrd26 Soybean Promoter in Response to Abiotic Stresses: Potential Tool for Biotechnological Application." BMC Biotechnology 19, no. 1 (2019): 79. [CrossRef]
Figure 1.
Chromosomal locations of HMA genes in C. amarus (dark blue), C. colocynthis (yellow), C. lanatus (dark green), C. mucusospermus (red), C. melo (purple), C. sativus (brown), C. maxima (pink), C. moschata (gray), C. pepo (light green) and L. siceraria (light blue). Chromosome numbers are represented at the top of each chromosome. The left panel scale indicates the chromosome length in Mb.
Figure 1.
Chromosomal locations of HMA genes in C. amarus (dark blue), C. colocynthis (yellow), C. lanatus (dark green), C. mucusospermus (red), C. melo (purple), C. sativus (brown), C. maxima (pink), C. moschata (gray), C. pepo (light green) and L. siceraria (light blue). Chromosome numbers are represented at the top of each chromosome. The left panel scale indicates the chromosome length in Mb.
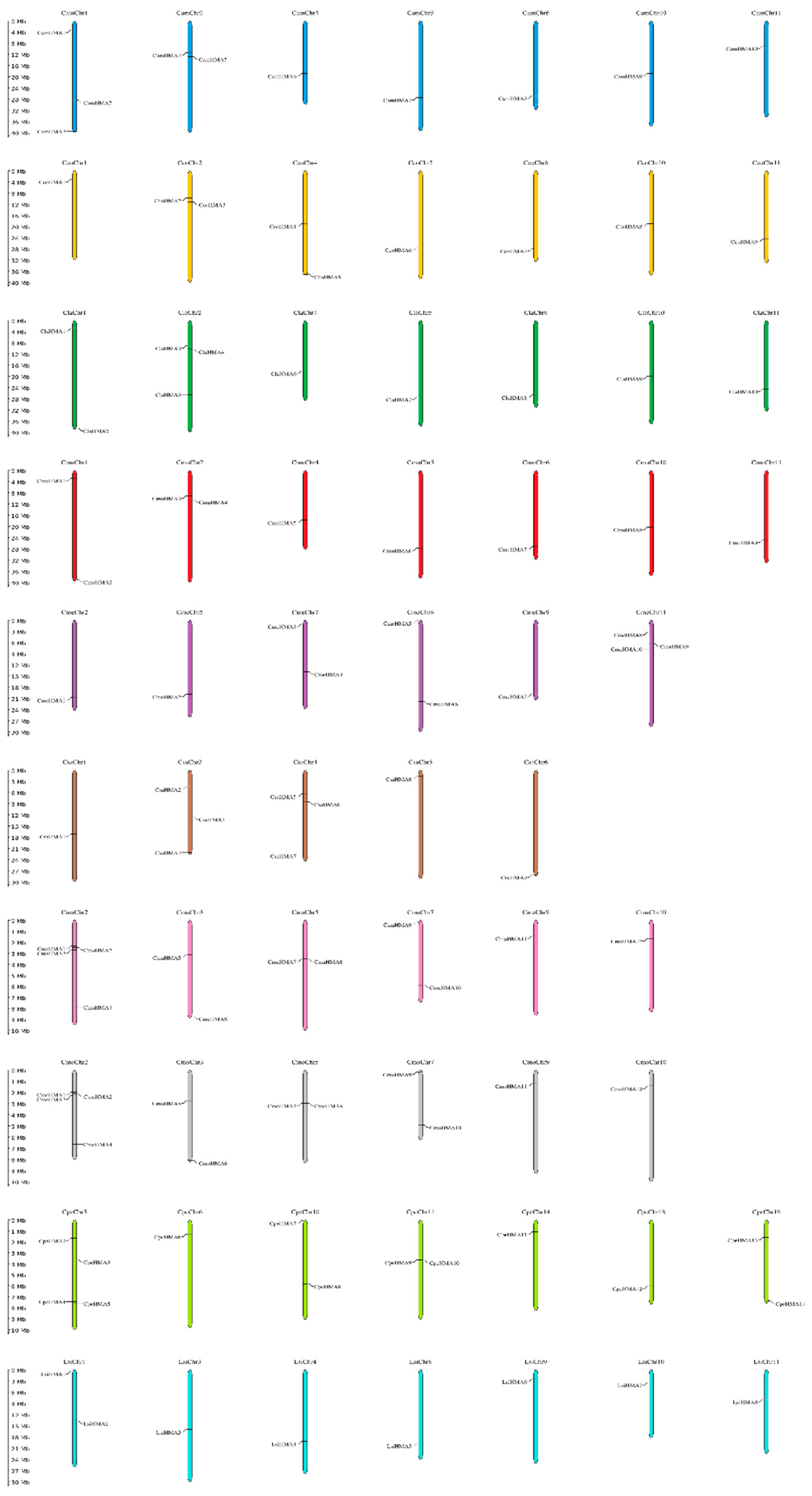
Figure 2.
Gene structure and protein domains of the HMA family in Cucurbit species. (a) Exon/intron structures of HMA genes. Exons are represented as orange boxes, introns as strings, and promoter UTRs in black. (b) Protein domain structure pattern of HMA gene family, different-colored rectangles represent different structural domains; the green rectangles represent E1–E2 ATPase; the yellow rectangle represents Hydrolase; the pink rectangle represents HMA; the green-dark rectangle represents E1–E2 ATPase superfamily. The gene name is in the left side of each sequence and the below scale indicates the length in kb and aa, respectively.
Figure 2.
Gene structure and protein domains of the HMA family in Cucurbit species. (a) Exon/intron structures of HMA genes. Exons are represented as orange boxes, introns as strings, and promoter UTRs in black. (b) Protein domain structure pattern of HMA gene family, different-colored rectangles represent different structural domains; the green rectangles represent E1–E2 ATPase; the yellow rectangle represents Hydrolase; the pink rectangle represents HMA; the green-dark rectangle represents E1–E2 ATPase superfamily. The gene name is in the left side of each sequence and the below scale indicates the length in kb and aa, respectively.
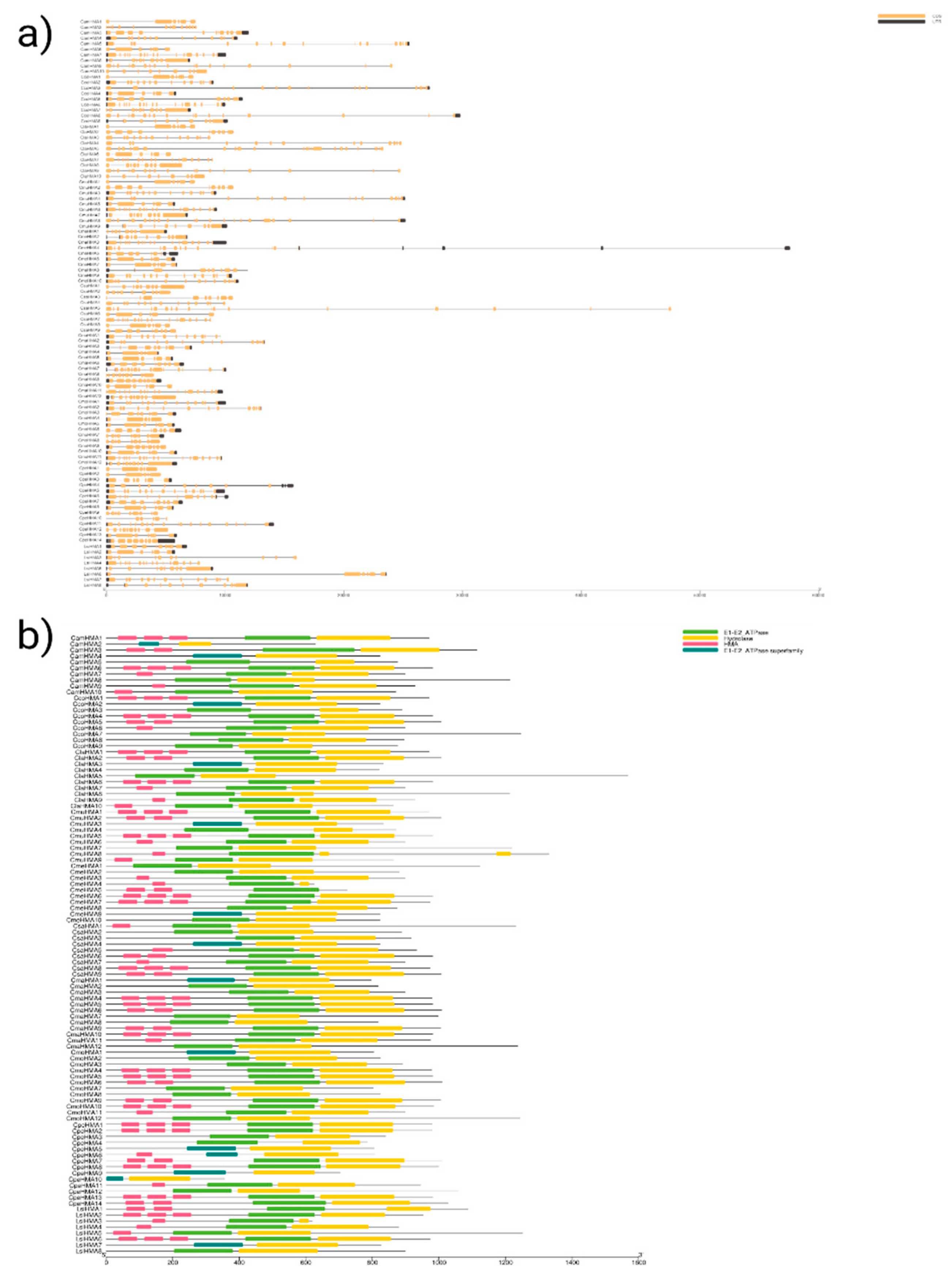
Figure 3.
Phylogenetic relationship of the HMA gene family. Phylogenetic analysis of Cucurbits and Arabidopsis was carried out by the neighbor-joining method with 1000 bootstrap. Based on genetic and functional studies, the phylogeny was divided in two major clades, P1B-ATPases, zinc (Zn)/cobalt (Co)/cadmium (Cd)/lead (Pb) group was highlighted in red and the copper (Cu)/silver (Ag) group was highlighted in green.
Figure 3.
Phylogenetic relationship of the HMA gene family. Phylogenetic analysis of Cucurbits and Arabidopsis was carried out by the neighbor-joining method with 1000 bootstrap. Based on genetic and functional studies, the phylogeny was divided in two major clades, P1B-ATPases, zinc (Zn)/cobalt (Co)/cadmium (Cd)/lead (Pb) group was highlighted in red and the copper (Cu)/silver (Ag) group was highlighted in green.
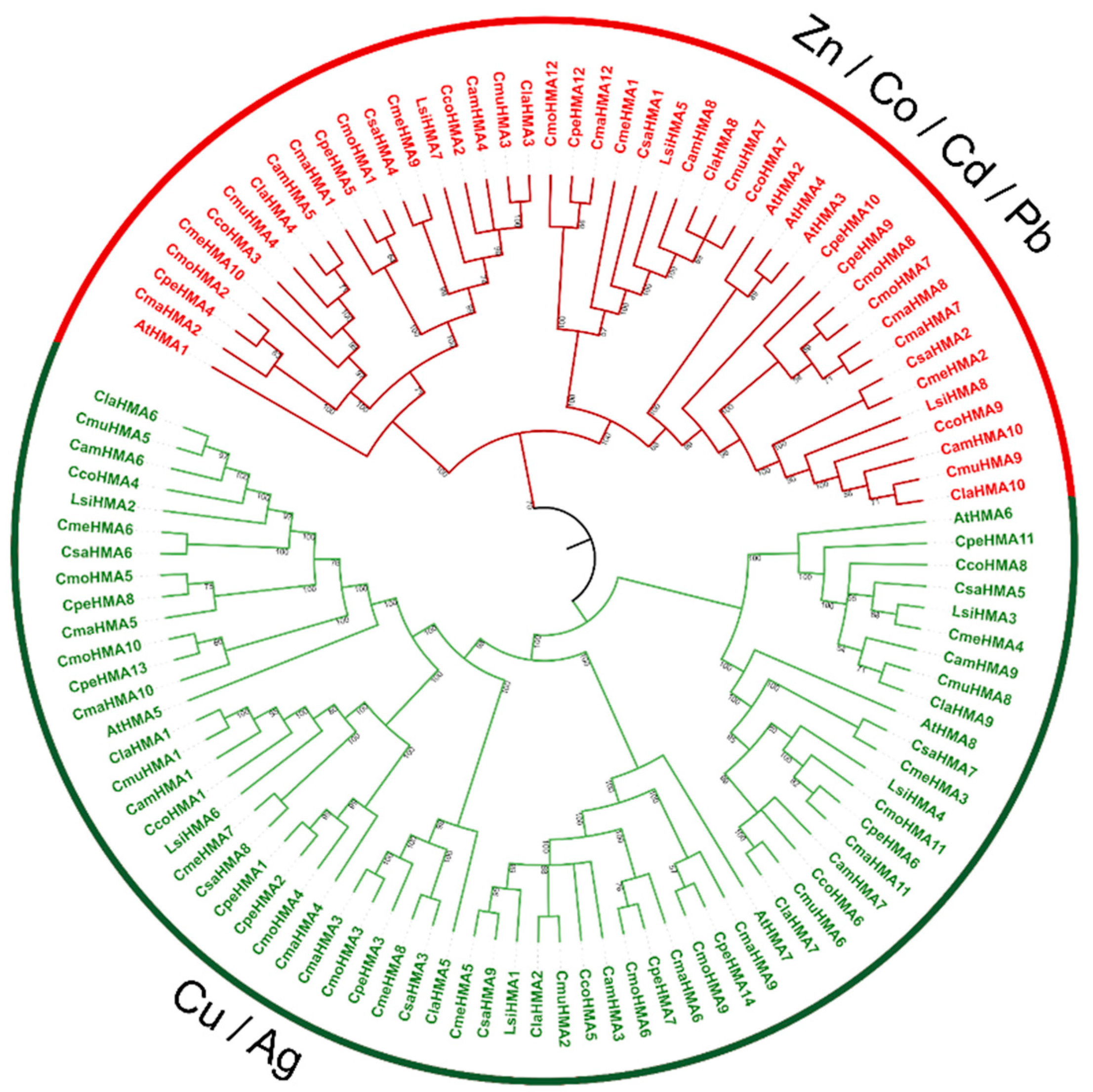
Figure 4.
Collinearity analysis between the HMA gene family in Cucurbitaceae species. a) Synteny between the Citrullus genera: Citrullus amarus (dark blue), Citrullus colocynthis (yellow), Citrullus lanatus (dark green) and Citrullus mucusospermus (red). b) Synteny between the Cucumis genera: Cucumis melo (purple) and Cucumis sativus (brown). c) Synteny between the Cucurbita genera: Cucurbita maxima (pink), Cucurbita pepo (light green) and Cucurbita moschata (gray). The collinearity analysis was visualized using TBTools with E-value < 1× 10-10.
Figure 4.
Collinearity analysis between the HMA gene family in Cucurbitaceae species. a) Synteny between the Citrullus genera: Citrullus amarus (dark blue), Citrullus colocynthis (yellow), Citrullus lanatus (dark green) and Citrullus mucusospermus (red). b) Synteny between the Cucumis genera: Cucumis melo (purple) and Cucumis sativus (brown). c) Synteny between the Cucurbita genera: Cucurbita maxima (pink), Cucurbita pepo (light green) and Cucurbita moschata (gray). The collinearity analysis was visualized using TBTools with E-value < 1× 10-10.
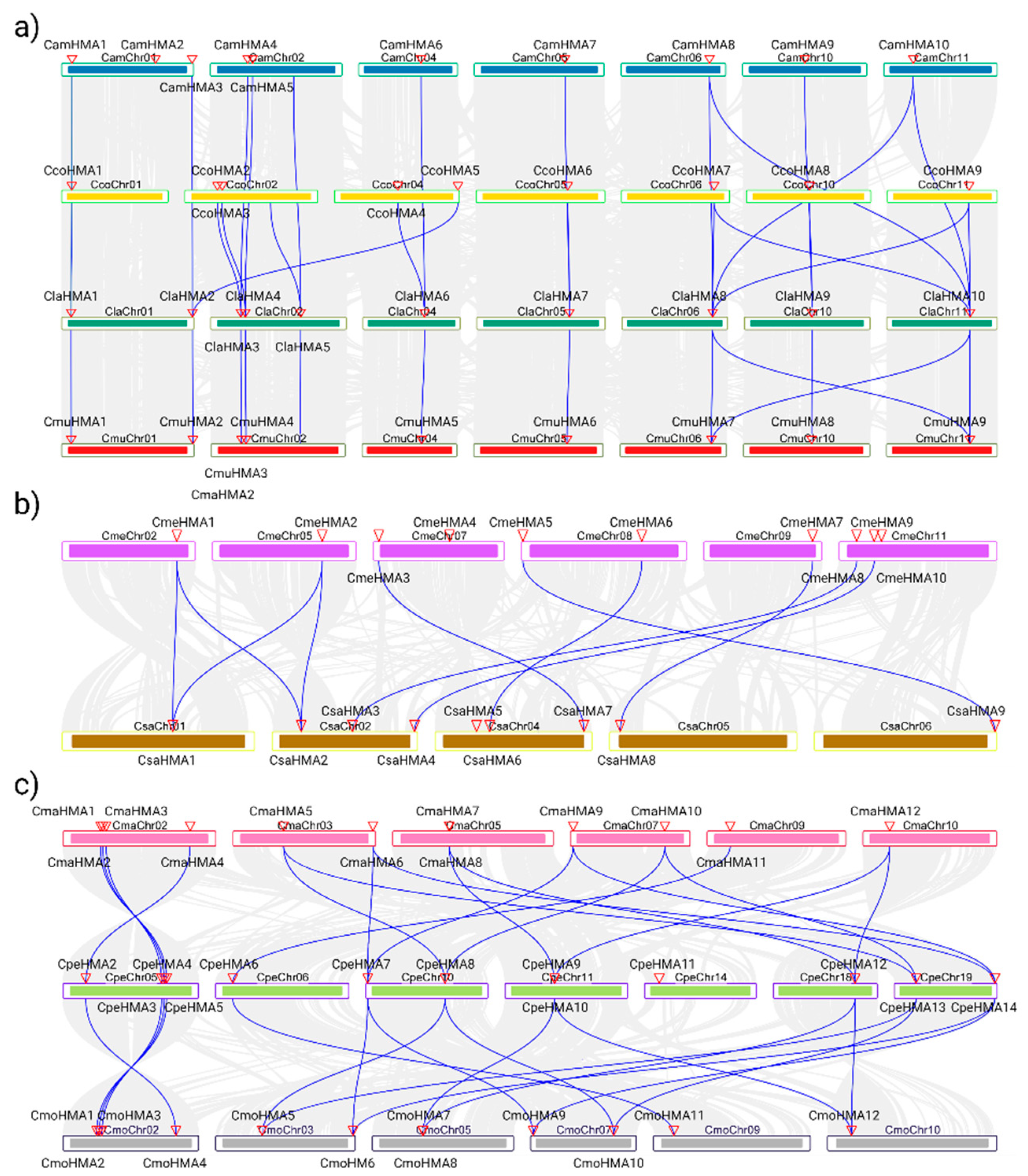
Figure 5.
Common conserved motifs of HMA proteins among Cucurbitaceae plants: C. amarus, C. colocynthis, C. lanatus, C. mucusospermus, C. melo, C. sativus, C. maxima, C. moschata, C. pepo, L. siceraria. The overall height of the stack indicates the degree of sequence conservation. The height of residues suggests the relative frequency of each residue at that position. Typed sequences of motifs are represented below each stack.
Figure 5.
Common conserved motifs of HMA proteins among Cucurbitaceae plants: C. amarus, C. colocynthis, C. lanatus, C. mucusospermus, C. melo, C. sativus, C. maxima, C. moschata, C. pepo, L. siceraria. The overall height of the stack indicates the degree of sequence conservation. The height of residues suggests the relative frequency of each residue at that position. Typed sequences of motifs are represented below each stack.
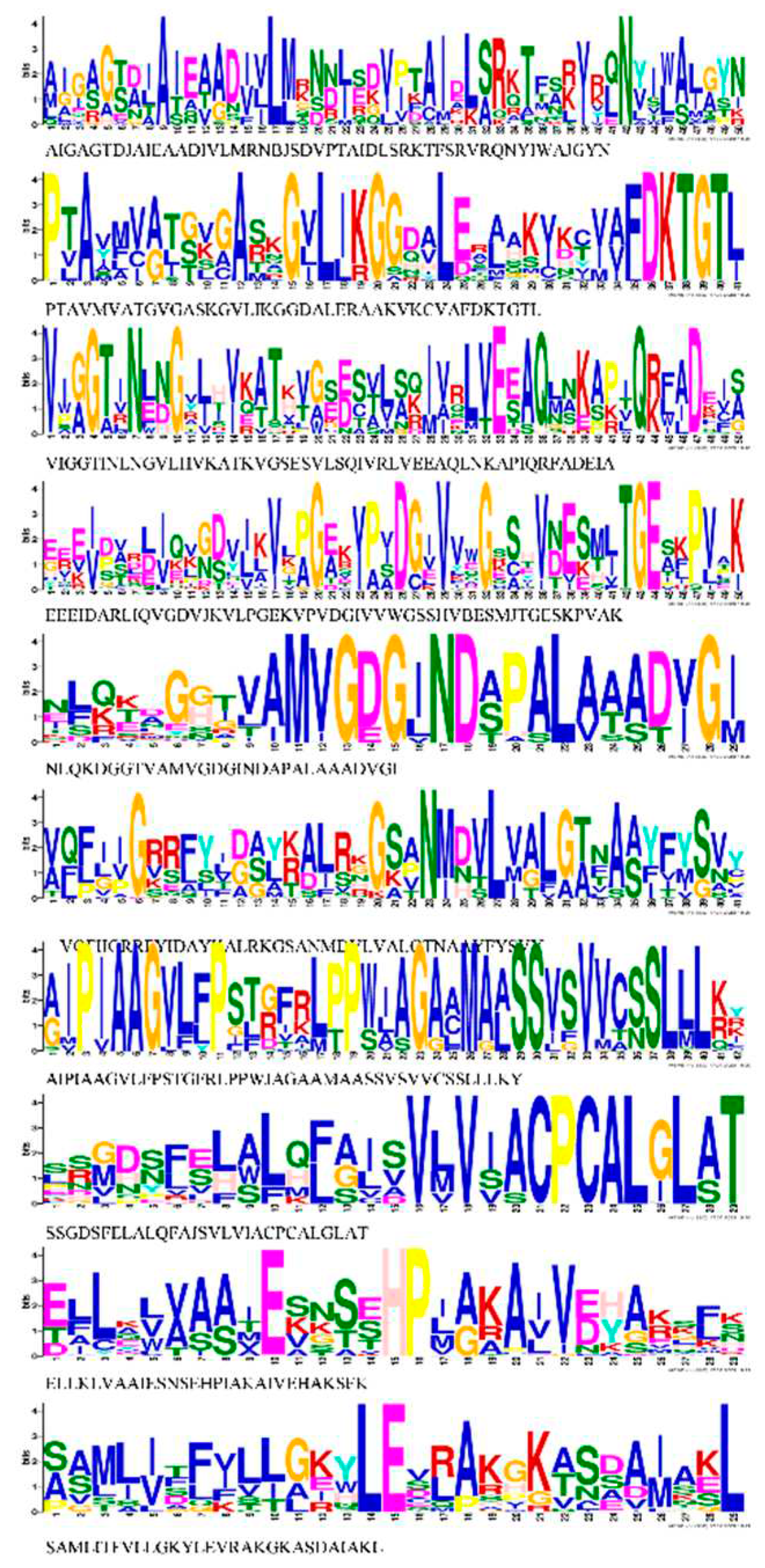
Figure 6.
Gene ontology analysis results for Cucurbit species. Cellular Component (CC), Molecular Function (MF) and Biological Processes (BP) were identified with the OmicsBox program. Specific results for genes of each species is found in Supplementary Table S4.
Figure 6.
Gene ontology analysis results for Cucurbit species. Cellular Component (CC), Molecular Function (MF) and Biological Processes (BP) were identified with the OmicsBox program. Specific results for genes of each species is found in Supplementary Table S4.
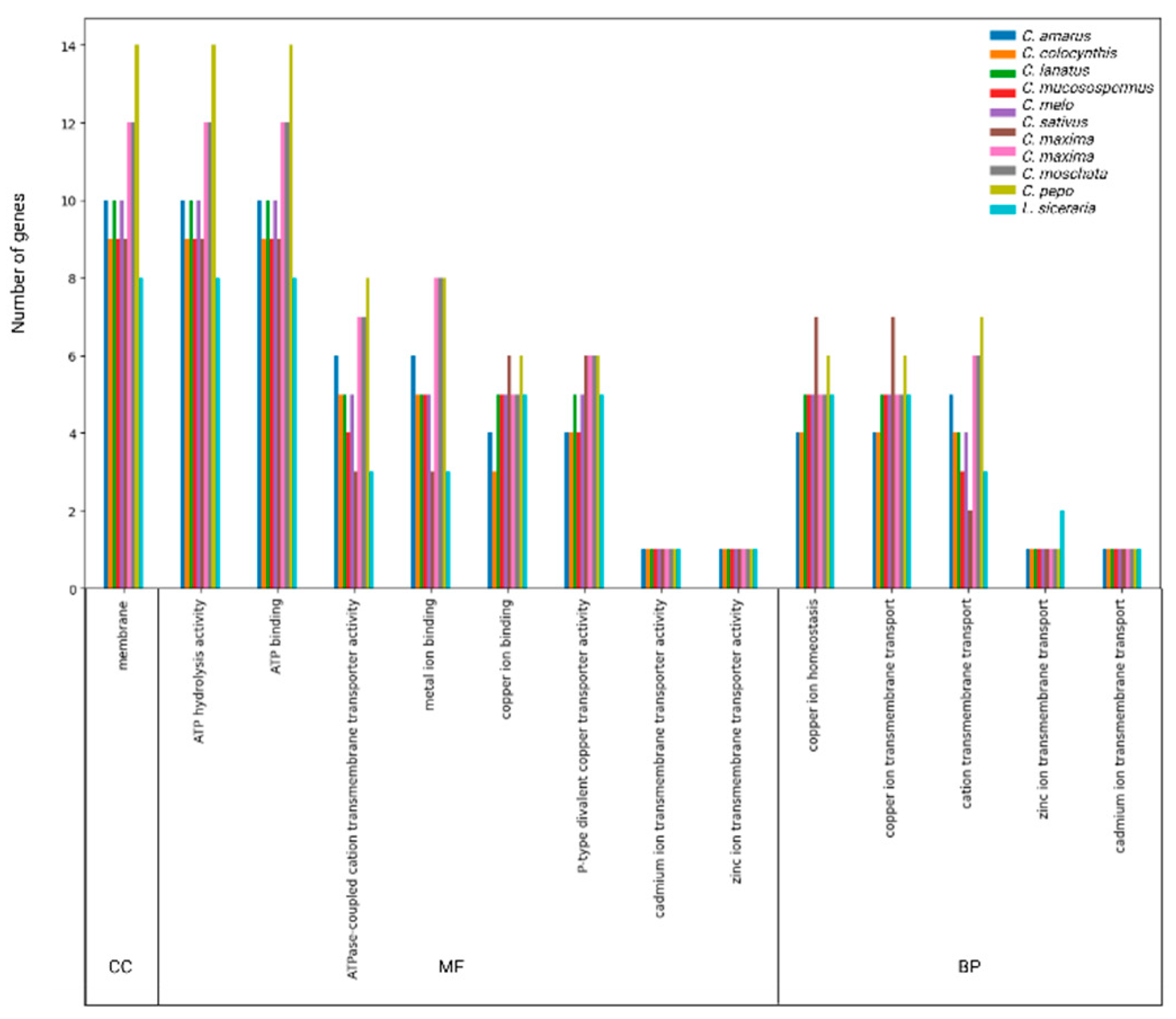
Figure 7.
Heat map of gene expression levels of HMA genes in Root, Leaf, and Pollen tissues of Cucurbita pepo exposed to CuO NPs, CuO bulk, and CuSO4 (Accesion: GSE173716). The bar right to the heat map represents normalized expression values in each treatment.
Figure 7.
Heat map of gene expression levels of HMA genes in Root, Leaf, and Pollen tissues of Cucurbita pepo exposed to CuO NPs, CuO bulk, and CuSO4 (Accesion: GSE173716). The bar right to the heat map represents normalized expression values in each treatment.
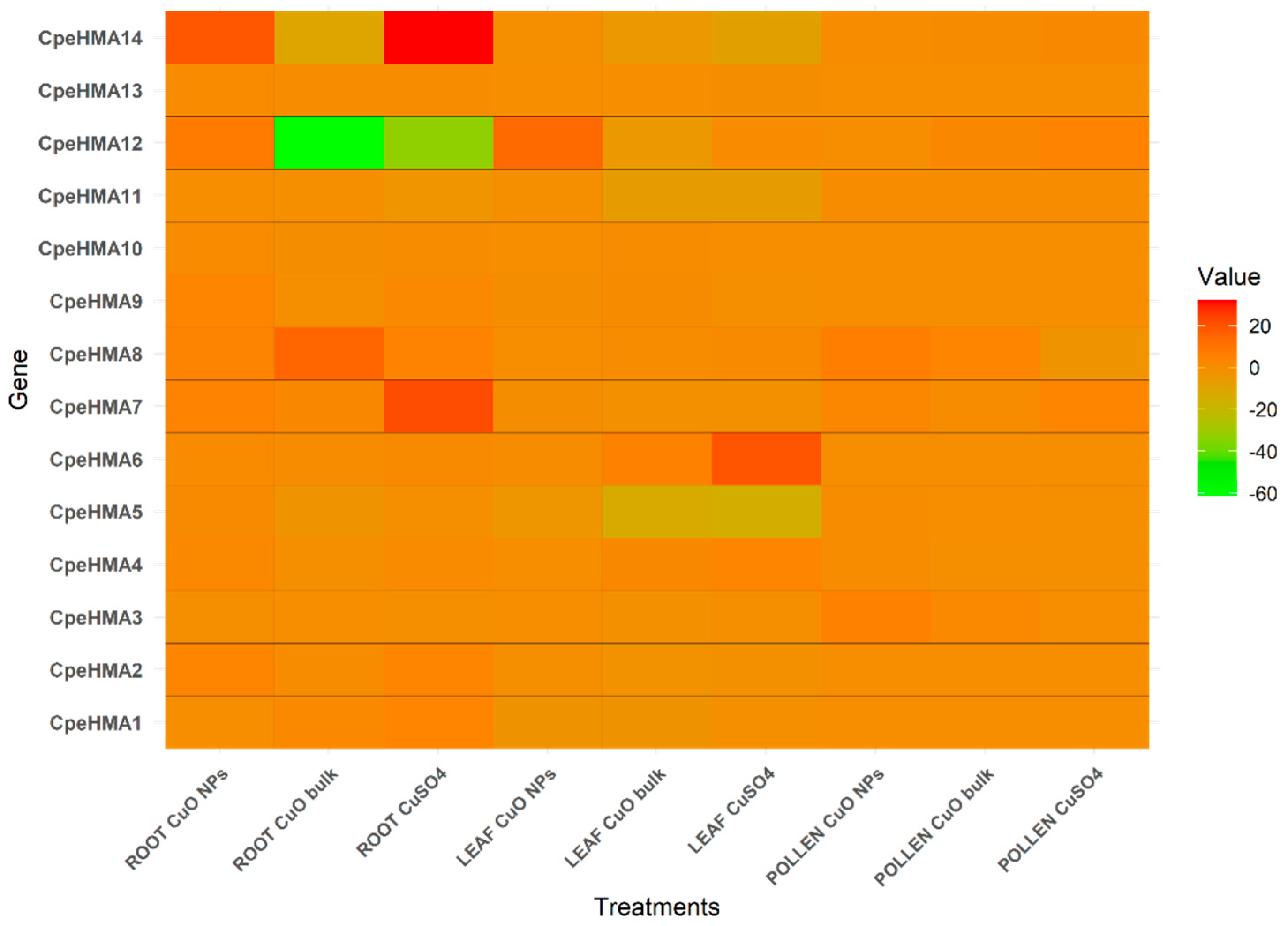
Figure 8.
Expression pattern of HMA genes from C. pepo in different tissues under As stress. Relative expression levels of the CpHMA genes in a) Root, b) Leaf, c) Flower, d) and Fruit of C. pepo plants treated with 50, 100, and 200 μM Arsenic in soil were determined by qRT-PCR. The gene expression level for each HMA gene in the control plants with no As was normalized to 1, as the 2^ΔΔCT method suggests. The results represent the means of the biological replicates with their standard deviation represented as error bars. “*”. “**” and “***” indicates genes statistically significantly differentially expressed between the treatment and the control using a t-test at the level of p ≤ 0.05, p ≤ 0.01, and p ≤ 0.001, respectively.
Figure 8.
Expression pattern of HMA genes from C. pepo in different tissues under As stress. Relative expression levels of the CpHMA genes in a) Root, b) Leaf, c) Flower, d) and Fruit of C. pepo plants treated with 50, 100, and 200 μM Arsenic in soil were determined by qRT-PCR. The gene expression level for each HMA gene in the control plants with no As was normalized to 1, as the 2^ΔΔCT method suggests. The results represent the means of the biological replicates with their standard deviation represented as error bars. “*”. “**” and “***” indicates genes statistically significantly differentially expressed between the treatment and the control using a t-test at the level of p ≤ 0.05, p ≤ 0.01, and p ≤ 0.001, respectively.

Table 1.
Number of protein members of the HMA family in Arabidopsis and Cucurbitaceous species and their distribution in the Zn/Cd/Co/Pb and Cu/Ag major clades.
Table 1.
Number of protein members of the HMA family in Arabidopsis and Cucurbitaceous species and their distribution in the Zn/Cd/Co/Pb and Cu/Ag major clades.
| Species | HMA clade | Total | |
| Zn/Cd/Co/Pb | Cu/Ag | ||
| Arabidopsis thaliana | 4 | 4 | 8 |
| Citrullus amarus | 4 | 6 | 10 |
| Citrullus colocynthis | 4 | 5 | 9 |
| Citrullus lanatus | 4 | 6 | 10 |
| Citrullus mucusospermus | 4 | 5 | 9 |
| Cucumis melo | 4 | 6 | 10 |
| Cucumis sativus | 3 | 6 | 9 |
| Cucurbita maxima | 5 | 7 | 12 |
| Cucurbita moschata | 5 | 7 | 12 |
| Cucurbita pepo | 5 | 9 | 14 |
| Legenaria siceraria | 3 | 5 | 8 |
Table 2.
Ka, Ks, Ka/Ks ratio and divergent time of the duplicated HMA genes in Cucurbitaceous plants.
Table 2.
Ka, Ks, Ka/Ks ratio and divergent time of the duplicated HMA genes in Cucurbitaceous plants.
| Species | Pair# | Gene names | Ka | Ks | Ka/Ks ratio | Duplication type | MYA1 |
|---|---|---|---|---|---|---|---|
| C. amarus | 1 | CamHMA2-CamHMA3 | 0.5081 | 0.4684 | 1.0846 | Tandem | 35.70 |
| 2 | CamHMA1-CamHMA6 | 0.1036 | 0.4185 | 0.2474 | Segmental | 31.90 | |
| 3 | CamHMA8-CamHMA10 | 0.1208 | 0.3328 | 0.3629 | Segmental | 25.36 | |
| 4 | CamHMA7-CamHMA9 | 0.2087 | 0.4635 | 0.4503 | Segmental | 35.33 | |
| 5 | CamHMA4-CamHMA5 | 0.0796 | 0.2789 | 0.2855 | Tandem | 21.25 | |
| C. colocynthis | 1 | CcoHMA1-CcoHMA4 | 0.1056 | 0.4402 | 0.2400 | Segmental | 33.55 |
| 2 | CcoHMA7-CcoHMA9 | 0.1192 | 0.3347 | 0.3562 | Segmental | 25.51 | |
| 3 | CcoHMA6-CcoHMA8 | 0.2019 | 0.4616 | 0.4374 | Segmental | 35.18 | |
| 4 | CcoHMA2-CcoHMA3 | 0.0823 | 0.2832 | 0.2907 | Tandem | 21.59 | |
| C. lanatus | 1 | ClaHMA2-ClaHMA8 | 0.3042 | 0.4742 | 0.6415 | Segmental | 36.14 |
| 2 | ClaHMA1-ClaHMA6 | 0.1059 | 0.4301 | 0.2463 | Segmental | 32.78 | |
| 3 | ClaHMA5-ClaHMA10 | 0.1174 | 0.3248 | 0.3615 | Segmental | 24.75 | |
| 4 | ClaHMA7-ClaHMA9 | 0.2128 | 0.4653 | 0.4573 | Segmental | 35.46 | |
| 5 | ClaHMA3-ClaHMA4 | 0.0832 | 0.2848 | 0.2922 | Tandem | 21.70 | |
| C. mucusospermus | 1 | CmuHMA1-CmuHMA5 | 0.1075 | 0.4420 | 0.2431 | Segmental | 33.69 |
| 2 | CmuHMA7-CmuHMA9 | 0.1212 | 0.3426 | 0.3538 | Segmental | 26.11 | |
| 3 | CmuHMA6-CmuHMA8 | 0.2076 | 0.4521 | 0.4591 | Segmental | 34.46 | |
| 4 | CmuHMA3-CmuHMA4 | 0.0847 | 0.2817 | 0.3009 | Tandem | 21.47 | |
| C. melo | 1 | CmeHMA6-CmeHMA7 | 0.1115 | 0.4288 | 0.2599 | Segmental | 32.68 |
| 2 | CmeHMA2-CmeHMA4 | 0.2848 | 0.3743 | 0.7608 | Segmental | 28.52 | |
| 3 | CmeHMA9-CmeHMA10 | 0.0771 | 0.2719 | 0.2835 | Tandem | 20.72 | |
| 4 | CmeHMA3-CmeHMA5 | 0.2542 | 0.3687 | 0.6894 | Segmental | 28.10 | |
| C. sativus | 1 | CsaHMA6-CsaHMA8 | 0.1077 | 0.4426 | 0.2434 | Segmental | 33.73 |
| 2 | CsaHMA1-CsaHMA2 | 0.1225 | 0.3678 | 0.3330 | Segmental | 28.03 | |
| 3 | CsaHMA5-CsaHMA7 | 0.2109 | 0.4253 | 0.4958 | Segmental | 32.41 | |
| 4 | CsaHMA4-CsaHMA9 | 0.3201 | 0.4293 | 0.7455 | Segmental | 32.72 | |
| C. maxima | 1 | CmaHMA7-CmaHMA8 | 0.0256 | 0.0308 | 0.8303 | Tandem | 2.35 |
| 2 | CmaHMA6-CmaHMA9 | 0.0272 | 0.1394 | 0.1951 | Segmental | 10.62 | |
| 3 | CmaHMA1-CmaHMA2 | 0.0703 | 0.2765 | 0.2543 | Tandem | 21.07 | |
| 4 | CmaHMA3-CmaHMA11 | 0.2830 | 0.4255 | 0.6651 | Segmental | 32.43 | |
| 5 | CmaHMA5-CmaHMA10 | 0.0186 | 0.1056 | 0.1762 | Segmental | 8.05 | |
| C. moschata | 1 | CmoHMA3-CmoHMA11 | 0.3381 | 0.5363 | 0.6304 | Segmental | 40.88 |
| 2 | CmoHMA1-CmoHMA2 | 0.0874 | 0.3327 | 0.2628 | Tandem | 25.36 | |
| 3 | CmoHMA5-CmoHMA10 | 0.0190 | 0.1179 | 0.1612 | Segmental | 8.99 | |
| 4 | CmoHMA6-CmoHMA9 | 0.0241 | 0.1136 | 0.2125 | Segmental | 8.66 | |
| 5 | CmoHMA7-CmoHMA8 | 0.0130 | 0.0167 | 0.7802 | Tandem | 1.27 | |
| C. pepo | 1 | CpeHMA4-CpeHMA5 | 0.0981 | 0.3070 | 0.3196 | Tandem | 23.39 |
| 2 | CpeHMA7-CpeHMA14 | 0.0294 | 0.1337 | 0.2198 | Segmental | 10.19 | |
| 3 | CpeHMA6-CpeHMA11 | 0.2511 | 0.4202 | 0.5975 | Segmental | 32.03 | |
| 4 | CpeHMA9-CpeHMA10 | 0.0170 | 0.0301 | 0.5649 | Tandem | 2.29 | |
| 5 | CpeHMA8-CpeHMA13 | 0.0183 | 0.1176 | 0.1562 | Segmental | 8.96 | |
| L. siceraria | 1 | LsiHMA5-LsiHMA8 | 0.1290 | 0.3793 | 0.3400 | Segmental | 28.91 |
| 2 | LsiHMA3-LsiHMA4 | 0.2196 | 0.4498 | 0.4881 | Segmental | 34.28 | |
| 3 | LsiHMA1-LsiHMA7 | 0.2933 | 0.3786 | 0.7748 | Segmental | 28.85 | |
| 4 | LsiHMA2-LsiHMA6 | 0.0856 | 0.3274 | 0.2615 | Segmental | 24.95 |
1million years ago.
Table 3.
Common putative cis-elements identified in the promoter sequences of HMA proteins genes in Cucurbit species.
Table 3.
Common putative cis-elements identified in the promoter sequences of HMA proteins genes in Cucurbit species.
| Cis-regulatory element | Expression pattern | Signal sequence |
| 3-AF1 binding site | light responsive element | TAAGAGAGGAA |
| AAGAA-motif | binding site in many light-regulated gene | GAAAGAA |
| ABRE | cis-acting element involved in the abscisic acid responsiveness | ACGTG |
| ABRE4 | early responsive to dehydration | CACGTA |
| ACE | cis-acting element involved in light responsiveness | CTAACGTATT |
| AE-box | part of a module for light response | AGAAACAA |
| AP-1 | cis-acting element for proline | TGAGTTAG |
| ARE | cis-acting regulatory element essential for the anaerobic induction | AAACCA |
| as-1 | transcriptional activation of several genes by auxin and/or salicylic acid; May be relevant to light regulation | TGACG |
| AT~TATA-box | Critical for accurate initiation of transcription | TATATA |
| AT1-motif | part of a light responsive module | AATTATTTTTTATT |
| ATCT-motif | part of a conserved DNA module involved in light responsiveness | AATCTAATCC |
| AT-rich element | binding site of AT-rich DNA binding protein (ATBP-1) | ATAGAAATCAA |
| AT-rich sequence | element for maximal elicitor-mediated activation (2copies) | TAAAATACT |
| AuxRR-core | cis-acting regulatory element involved in auxin responsiveness | GGTCCAT |
| Box 4 | part of a conserved DNA module involved in light responsiveness | ATTAAT |
| Box II | part of a light responsive element | TGGTAATAA |
| Box III | protein binding site | ATCATTTTCACT |
| CAAT-box | common cis-acting element in promoter and enhancer regions | CAAAT |
| CAT-box | cis-acting regulatory element related to meristem expression | GCCACT |
| CCAAT-box | MYBHv1 binding site | CAACGG |
| CGTCA-motif | cis-acting regulatory element involved in the MeJA-responsiveness | CGTCA |
| chs-CMA1a | part of a light responsive element | TTACTTAA |
| circadian | cis-acting regulatory element involved in circadian control | CAAAGATATC |
| DRE1 | expressed during late embryogenesis, induced by ABA | ACCGAGA |
| ERE | ethylene-induced activation | ATTTTAAA |
| GA-motif | part of a light responsive element | ATAGATAA |
| Gap-box | part of a light responsive element | CAAATGAAGA |
| GARE-motif | gibberellin-responsive element | TCTGTTG |
| GATA-motif | part of a light responsive element | AAGGATAAGG |
| G-box | cis-acting regulatory element involved in light responsiveness | CAGACGTGGCA |
| GCN4_motif | cis-regulatory element involved in endosperm expression | TGAGTCA |
| GT1-motif | light responsive element | GGTTAA |
| H-box | Essential for both light regulation and elicitor induction | CCTACCNNNNNNNCTNNNNA |
| HD-Zip 1 | element involved in differentiation of the palisade mesophyll cells | CAATAATTG |
| I-box | part of a light responsive element | TGATAATGT |
| LAMP-element | part of a light responsive element | CTTTATCA |
| L-box | part of a light responsive element | ATCCCACCTAC |
| LTR | cis-acting element involved in low-temperature responsiveness | CCGAAA |
| MBS | MYB binding site involved in drought-inducibility | CAACTG |
| MBSI | MYB binding site involved in flavonoid biosynthetic genes regulation | TTTTTACGGTTA |
| MRE | MYB binding site involved in light responsiveness | AACCTAA |
| MYB | Involved in regulation of genes that are responsive to water stress | CAACAG |
| MYB recognition site | Involved in regulation of genes that are responsive to water stress | CCGTTG |
| Myb-binding site | gibberellin-responsive element | CAACAG |
| MYB-like sequence | Involved in regulation of genes that are responsive to water stress | TAACCA |
| MYC | Related to cold and dehydration responsiveness | CAATTG |
| O2-site | cis-acting regulatory element involved in zein metabolism regulation | GATGACATGG |
| P-box | gibberellin-responsive element | CCTTTTG |
| Sp1 | light responsive element | GGGCGG |
| TATA | core promoter element around -30 of transcription start | TATAAAAT |
| TATA-box | core promoter element around -30 of transcription start | ATATAT |
| TATC-box | cis-acting element involved in gibberellin-responsiveness | TATCCCA |
| TCA | cis-acting element involved in salicylic acid responsiveness | TCATCTTCAT |
| TCA-element | cis-acting element involved in salicylic acid responsiveness | CCATCTTTTT |
| TCCC-motif | characteristic of the promoters activated in infected cells DE of root nodules | TCTCCCT |
| TC-rich repeats | cis-acting element involved in defense and stress responsiveness | GTTTTCTTAC |
| TCT-motif | part of a light responsive element | TCTTAC |
| TGACG-motif | cis-acting regulatory element involved in the MeJA-responsiveness | TGACG |
| TGA-element | auxin-responsive element | AACGAC |
| W box | involved in elicitor-responsive transcription of defense genes | TTGACC |
| WRE3 | RRE (R response element) | CCACCT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



