
Submitted:
16 September 2023
Posted:
19 September 2023
You are already at the latest version
Abstract
Cancer is second leading cause of death, worldwide. Lung cancer is the leading cause of cancer-related mortality. Plant-based therapeutics and herbal medicine have played a vital role in the development of several anti-cancerous agents, and has been used to reduce the severe side effects of chemotherapy as well. Since the anti-lung cancer properties of the plant Medicago. orbicularis are not explored yet, we identified its phytochemical composition and investigated the anti-oxidant, anti-hemolytic, and anti-cancerous properties of extracts of this plant in A549 human lung adenocarcinoma cells. Results show that all parts of M. orbicularis (stems, leaves, and fruits) exhibit remarkable anti-oxidant and hemolytic activities. In addition, all extracts showed a dose-dependent anti-cancerous cytotoxic activity against A549 cells; with fruit extracts being the most potent. This cytotoxic effect could be related, at least partly, to the induction of apoptosis, where M. orbicularis fruit extracts activated Caspase-3 and PARPP-1, and reduced the ratio of anti-apoptotic BCL-2/ pro-apoptotic BAX, thereby promoting cellular death. Furthermore, the use of M. orbicularis, in combination with a conventional chemotherapeutic agent, cisplatin, was assessed. Indeed, combination of cisplatin and M. orbicularis fruit extracts was more cytotoxic and induced more aggregation of A549 cells than either treatment alone. GC-MS analysis and total polyphenol and flavonoid content determination indicated that M. orbicularis is rich in compounds that have anti-cancerous effects. M. orbicularis may be a potential source of anti-cancerous agents to manage progression of lung cancer and its resistance to therapy.
Keywords:
Medicago orbicularis
; Lung cancer
; Herbal medicine
; Antioxidants
; hemolytic
; cisplatin
; A549 cells
I. Introduction
Cancer remains the second leading cause of disability and death worldwide where around 1 in 6 deaths are related to cancer. In 2020, around 19 million new cancer cases and almost 10 million cancer-related deaths were recorded globally [1].
Lung cancer is a major public health concern, with significant impact on individuals and communities worldwide. Lung cancer has a high incidence rate; it was the second most commonly diagnosed cancer in 2020. Lung cancer is the leading cause of cancer-related deaths (around 1.8 million deaths or 18% of global cancer-related deaths in 2020) [1]. With a 21% 5-year survival rate, lung cancer has one of the most poor prognoses [2].
Depending on its progression, or the presence of metastasis, treatment of lung cancer will differ. Surgical resection, combination chemotherapy, and stereotactic body radiation are considered as the main modes of treatment. A significant obstacle in lung cancer therapy is the resistance that develops to chemo- and radio-therapy. Following therapy, resistance can lead to the relapse of lung cancer around 6 months post-therapy, in many patients [3,4]. Cisplatin is a cytotoxic chemotherapeutic agent used to treat several cancers including lung cancer [5]. The standard of treatment for limited-stage lung cancer remains chemotherapy using four to six cycles of cisplatin and etoposide and concurrent radiation therapy. However, 60-70% of patients are at extensive-stage lung cancer and require a different treatment regimen; also involving cisplatin [4,6]. Nevertheless, these treatments can become refractory in patients. When relapse takes place lung cancer is usually refractory to treatment and has poor prognosis due to the limited availability of therapeutic options [3,4]. As a result, there is an urgent demand for new treatment options and alternative therapeutic modalities that can treat lung cancer and circumvent its chemoresistance. Herbal medicine represents a promising alternative in this regard.
Herbal medicine and phytotherapy, medicinal treatment based on plants or herbs and their extracts, have played a central role in the development of several anti-cancer agents such as Paclitaxel, Camptothecin, and Vincristine [7,8,9,10,11]. Herbal medicine can be utilized as an assistant therapeutic modality as well. It can enhance the response rate to chemotherapy as well as radiotherapy, help overcome resistance to therapy, decrease the severity of side-effects caused by chemotherapy and radiotherapy, and enhance the quality of life and survival of cancer patients [12,13,14,15]. Relatedly, herbal preparations have been used as well to reduce the severe side effects of chemotherapy, including cancer-related fatigue [7]. Herbal preparations and herbal active compounds act as anti-cancerous and anti-metastatic agents through several mechanisms, including scavenging of reactive oxygen species (ROS), modulation of epithelial to mesenchymal transition (EMT), impairment of angiogenesis, modification of the expression and activity of matrix metalloproteinases (MMPs), among others [16,17].
The Fabaceae family of plants includes a large number of domesticated species which are harvested as crops for human and animal consumption as well as oils, fuel, fertilizers, and medicinal and agricultural varieties [18]. Medicago is a genus of the Fabaceae family and it comprises more than 83-87 different species of flowering plants [18,19,20]. Twenty of these species are herbaceous perennials, 63 are herbaceous annuals and only 2 are shrubs [20]. Medicago orbicularis L. Bartal. (common names: black disk, button medick, or button clover) is a species of the Medicago genus indigenous to Eurasia and North Africa [19]. The plant is distributed throughout the Mediterranean basin mainly in Palestine, Lebanon, Syria, Algeria, Spain, France, Italy and Greece, in Middle East countries such as Iraq and Iran, and several other countries [18]. M. orbicularis is a winter annual plant which flowers in Spring and early Summer. Its stems are leafy, stipules are fimbriate, leaflets are oval non-hairy with toothed margins, flowers are orange-yellow, seed pods are flat, coiled and lack spines [19].
M. orbicularis phytochemical composition has not been investigated. But, other Medicago species have been reported to be rich in phytochemicals, including flavonoids, carotenoids, saponins, phenolic acids, and phytoestrogens [21,22,23]. These phytochemicals have been demonstrated to exhibit anticancer activities through several mechanisms including apoptosis, antiproliferative, immunomodulatory, and anti-inflammatory activities, and the capacity to modulate oxidative stress [24,25,26,27,28]. It is well recognized that antioxidant activity is one mechanism through which medicinal plants may exert a therapeutic effect [27,28,29,30,31,32]. Close-by species such as M. polymorpha, M. sativa, M. arabica, or M. truncatula were shown to possess a robust antioxidant activity [22,23]; an indication to the potential anti-oxidant and therapeutic value of M. orbicularis. Güleç and Kültür reported a traditional therapeutic use of M. orbicularis in Turkey for heart diseases [33]. However, there are no studies currently on the antioxidant potential, therapeutic applications or anti-cancerous properties of M. orbicularis. Therefore, we assessed the antioxidant capacity, hemolytic properties, phytochemical composition and the effects of M. Orbicularis on cell proliferation, cell aggregation, and cell migratory potential against A549 human lung cancer cells. Additionally, the combinatorial effect of a treatment using both cisplatin and M. orbicularis on A549 cells was tested.
II. Materials and Methods
Cell Culture and Reagents
Human lung adenocarcinoma cell line A549 and primary normal neonatal fibroblast (HDFn) were obtained from the American Type Culture Collection (ATCC; Manassas, VA, USA). Cells were maintained in a humidified chamber at 37 °C and 5% CO2 in DMEM (Dulbecco’s Modified Eagle’s Medium; cat# D0819 Sigma-Aldrich Co., St. Louis, MO, USA) supplemented with 10% fetal bovine serum (FBS) (cat# F9665, Sigma-Aldrich) and 100U/ml penicillin and 0.1 mg/ml streptomycin (cat# P4333, Sigma-Aldrich). Cells were passaged by trypsinization when they reached 90% confluence.
Plant Collection and Extraction
Fresh Medicago orbicularis L. Bartal was collected from Almat, Byblos, Lebanon between May-June 2021. A voucher specimen was stored in the herbarium of the Faculty of Pharmacy, Lebanese University, Beirut, Lebanon and was authenticated by Professor George Tohme, an experienced taxonomist and herbalist. Medicago orbicularis L. Bartal: Kingdom Plantae; Phylum Tracheophyta Class Magnoliopsida; Order Fabales; Family Fabaceae; Genus Medicago L. (Index Kewensis; https://powo.science.kew.org/taxon/urn:lsid:ipni.org:names:506322-1).
Extracts were obtained from leaves, stems and fruits of M. orbicularis. In brief, the different plant parts were cleaned, air-dried in the shade at room temperature, ground to powder, and kept in plastic containers, away from light, heat and moisture. Then, 10 grams of the powder were mixed with 100 ml of 70% aqueous ethanol, and the mixture was placed in a reciprocating shaker and continuously agitated at 150rpm for 3 to 4 days. The solutions were filtered and concentrated by a rotary evaporator at 40°C and then lyophilized using a lyophilizer. The lyophilized extracts were stored at -20°C until their use. At the time of use, the extracts were dissolved in 70% ethanol.
DPPH (α, α-diphenyl-β-picrylhydrazyl) Antioxidant Activity
The antioxidant free radical scavenging ability of the ethanolic extracts of the different parts of M. orbicularis (leaves, fruits and stems) was measured using the DPPH radical scavenging assay which was performed as described [34]. DPPH changes color from intense purple to pale-yellow due to the donation of a hydrogen atom, if the plant extract has radical scavenging activity. For M. orbicularis ethanolic extracts, 1 ml of different concentrations of the ethanolic plant extracts (50, 100, 150, 200, 300 or 500 μg/ml) was mixed with an equal volume of DPPH (cat# D9312, Sigma-Aldrich Co.) solution (0.15 mM in ethanol). The blank consisted of 1 ml of DPPH solution and 1 ml of 70% ethanol. Mixed samples were then kept in the dark for 30 min and the OD was measured at a wavelength of 515 nm using a spectrophotometer. Ascorbic acid, a potent antioxidant, was employed as a positive control and reference. The DPPH scavenging activity of each concentration of the extracts was calculated using the formula:
In vitro Cytotoxicity Assay
In vitro cytotoxicity of M. orbicularis extracts was assessed using MTT assay which is a reduction assay that measures cellular metabolic activity and is reflective of cell viability. A549 and neonatal fibroblast cells were seeded in a 96-well tissue culture plate at a density of 0.7x104 cells per well in DMEM culture medium containing 10% FBS and penicillin/ streptomycin. After an overnight incubation, cells were treated with different concentrations (50, 100, 150, 200, 300 or 500 μg/ml) of M. orbicularis extracts for 24, 48 and 72 h. As a control, cells were treated with a concentration of ethanol (vehicle-control cells) equal to the concentration of ethanol in the extract-treated cells. Following the treatment, MTT reagent (3-(4,5-Dimethylthiazol-2-yl)-2,5-Diphenyl Tetrazolium Bromide; cat# M5655, Sigma-Aldrich Co.) was added for 3 hours until a purple formazan precipitate was formed. Media were removed gently, and an isopropanol-HCl solvent was added to dissolve the formazan precipitates. The Optical Density (OD) was measured by an ELISA plate reader at a wavelength of 595 nm. The percentage of cell viability of the treated cells was calculated using the following formula:
The IC50 (half-maximal inhibitory concentration) value, was determined by extrapolation from the cell killing curve to determine the concentration of the extract that induced 50% cell death.
Cisplatin (Sigma-Aldrich Co.), dissolved in DMSO, was used as a positive control of the cytotoxic effect of M. orbicularis parts extracts on A549 cells. A549 cells were treated for 24, 48 and 72 hours with increasing concentrations of cisplatin (2.5, 5, 10 and 20 μg/ml) and compared to the DMSO treated vehicle-control cells. IC50 values of cisplatin in A549 cells were determined at the three time points. Data are displayed as the mean ± SEM of three independent experiments.
Cell Aggregation Assay
Seeded A549 cells were detached using 2mM EDTA PBS (calcium- and magnesium-free). Cells were pelleted and washed with PBS, and then resuspended in 1ml PBS including or not 100 or 150 μg/ml of M. orbicularis fruit extract alone, 5 μg/ml cisplatin alone, or a combination of both and kept shaking on a rocker for 60 minutes at 37°C in a cell culture incubator. Cells were fixed with 1ml of 1% formaldehyde and imaged using an inverted microscope (Leica Microsystems GmbH, Wetzlar, Germany). Percentage of aggregation was calculated using the formula: ; where Nt is the number of single cells in treated wells and Nc is number of single cells in the vehicle control cells.
Assay of Hemolysis and Anti-hemolytic activity
Hemolytic activity assay was performed as previously described [35]. Erythrocytes were obtained after separation from plasma by centrifugation at 2500 rpm for 10 min at 4°C starting from fresh sheep blood. Briefly, RBCs were washed three times with 1X PBS then pelleted and resuspended in PBS to obtain a 5% suspension of RBCs. Then, 50 μL of different concentrations (10, 20, 40, 50 and 100 μg/ml) of ethanolic extract of fruits of M. orbicularis were added to 1 ml of the RBC suspension. The mixture was incubated at 37°C for 1.5 hours, then the suspension was centrifuged at 2,500 rpm for 10 min at 4°C and the OD measured at 540 nm using a spectrophotometer. Ethanol concentrations equal to those present in the fruit extracts were added to blood and used as a negative control, 1% SDS as a positive control of hemolysis of RBCs, and 1X PBS as the blank. Hemolytic levels were calculated as follows:
Anti-hemolytic activity assessment was performed as previously published [36]. Briefly, the same steps were performed as in the hemolysis assay except that H2O2 was added to induce hemolysis. After adding M. orbicularis extracts to the RBC suspension for 20 min, RBCs were incubated with 350 μL of 30% H2O2 at 37°C for 1.5 h. The suspension was centrifuged at 2500 rpm for 10 min at 4°C and OD at 540 nm was determined. Thirty % H2O2 was used as a positive control, and 1X PBS as the blank. Anti-hemolytic levels were calculated according to the following equation:
Scratch/ Wound-Healing Assay
A549 cells were grown in 12-well tissue culture plates until confluence. A scrape was made through the confluent monolayer using a sterile yellow 200 μL pipette tip. The culture medium was then removed, the cells were washed twice with PBS (Sigma-Aldrich, St. Louis, MO, USA) to remove cellular debris, and incubated at 37℃ in fresh medium in the presence or absence of the indicated concentrations of M. orbicularis extracts. Photomicrographs of the scratch were obtained at baseline (0 h) and for the next 4 to 12 h using an inverted microscope (objective 10×). The width of the wound was expressed as the average ±SEM of the difference between the measurements taken at time zero and the corresponding time points. Ethanol was used as a vehicle control.
Western Blotting Analysis
A549 cells were seeded in a 6-well plate at a density of 2.8x105 cells per well and cultured for 24 h. The cells were treated with extracts of M. orbicularis for 48 h and then the cells were washed twice with PBS and lysed using a lysis buffer containing 2% SDS, 60 mM Tris lysis buffer (pH 6.8), and protease inhibitors and centrifuged at 10,000 g for 10 min. The protein concentration of the supernatants was determined using the Bradford protein assay kit (Biorad, Hercules, CA, USA) and 25 μg of protein lysates were resolved on 10% SDS-PAGE before being transferred to a polyvinylidene difluoride membrane (Immobilon PVDF; Biorad, Hercules, CA, USA). The membranes were then blocked for 1 h at room temperature with 5% non-fat dry milk in TBST (TBS and 0.05% Tween 20). Immunodetection was performed by incubating the membrane overnight with specific primary antibodies at 4℃. Primary antibodies included rabbit IgG anti-full length human poly-adenosine diphosphate (ADP) ribose polymerase-1 (PARP-1) (Cell Signaling Technologies, CST 9542; dilution 1/1,000), rabbit anti-human proCaspase 3 (Santa Cruz, sc-7148; dilution 1/1,000), rabbit anti-human B-cell lymphoma 2 (BCL-2) (Abcam, ab32124; dilution 1/5000), rabbit anti-Bcl-2 associated X protein (BAX) (Abcam, ab32503; dilution 1/2,000), and rabbit anti-GAPDH (Abcam, ab181602; dilution 1/10,000). Membranes were washed and incubated with a horseradish peroxidase (HRP)-conjugated goat anti-rabbit IgG secondary antibody (Abcam, ab6721; dilution 1/5,000) for 1 h followed by washing in TBST. Immunoreactive bands were detected using ECL substrate kit (Thermo Scientific, Rockford, IL, USA), and membranes were scanned using the Chemidoc imaging system (Bio-Rad). The intensity of the obtained bands was quantified using ImageJ software (NIH, MD, USA). All bands were normalized to GAPDH, which was used as a loading control.
Spectroscopic determination of total phenolic content
Total phenolic compounds content (TPC) was determined using the Folin–Ciocalteu colorimetric oxidation/reduction-based reaction as previously described [37], with slight modifications. Briefly, 100µl of diluted fruit extracts of M. orbicularis were added to 500µl of Folin–Ciocalteu reagent, followed by the addition of 400µl of saturated sodium carbonate solution (2%). The oxidation products of the reaction show a blue color with a broad light with an absorption maximum of 765 nm. OD 765 of the different mixtures was measured, against a blank, using a spectrophotometer. A calibration curve was prepared using a strong antioxidant, gallic acid, and the results were expressed as gallic acid equivalents (GAE) in milligrams per gram of the extract.
Spectroscopic determination of total flavonoids content
The total flavonoids content (TFC) of M. orbicularis fruit extracts was determined by the aluminium chloride colorimetric method as previously detailed [38]. AlCl3 forms a flavonoid–aluminium, complex that has a maximum OD at 510 nm. Briefly, a 100µl aliquot of appropriately diluted fruit extracts of M. orbicularis were added to 400µl of distilled water. Then, 400µl of 5% NaNO2 were added, followed by 300µl of 10% AlCl3. After 6 min of incubation 200µl of 1M NaOH were added to the mixture followed by 240µl of distilled water. The absorbance of the resulting pink solution was determined at 510 nm versus a blank (water). A calibration curve was prepared using a known flavonoid, quercetin. The curve was used to calculate the amount of flavonoids in the plant extracts which was expressed as quercetin equivalents in mg/g dry weight of the plant part.
Gas Chromatography/ Mass Spectrometry (GC/MS)
The phytochemical compounds of M. Orbicularis fruit extracts were analyzed using GC/MS analysis. A Perkin Elmer Clarus 680 (Perkin Elmer, USA) system attached to a triple quadrupole mass spectrometer was used. Chromatography was conducted on a hydrophobic capillary column RTxi-5 Sil MS column (30 m × 0.25 mm ID × 0.25 µm) using an injection volume of 10 µl, a flow rate of 1.5 mL/min, a pressure of 23.1 KPa, and an average velocity of 0.2 sec. The temperatures of the source and the interface were 200 °C and 280 °C, respectively. The initial temperature was set at 80 °C for 2 min, increased to 250 °C at 15 °C/min, and raised to 280 °C at 15 °C/min (held for 12 min). Identification of phytochemicals in extracts was carried out by comparing the obtained retention indices with those of chemical compounds in the database of the National Institute of Standards and Technology (NIST).
Statistical Analysis
Results were evaluated for statistical difference using One-way ANOVA followed by Bonferroni test to calculate P values. Data are presented as mean ± standard error of the mean (SEM) and a p-value of P <0.05 was considered as statistically significant. Statistical analysis was performed using GraphPad Prism 8 software (GraphPad Software Inc., San Diego, CA, USA).
III. Results
M. orbicularis plant ethanolic extracts exhibit remarkable antioxidant properties
The study commenced by testing the anti-oxidant capacity of M. orbicularis, as this was not addressed in previous studies. The antioxidant activity of the different parts of M. orbicularis was measured using the DPPH radical scavenging assay. Ethanolic extracts from all parts of M. orbicularis exhibited significant free radical scavenging activity which was concentration dependent (Figure 1). M. orbicularis leaves ethanolic extracts had the highest antioxidant potential (17% free radical scavenging activity at 200 μg/ml), followed by M. orbicularis fruits ethanolic extracts while M. orbicularis stems ethanolic extracts showed the lowest antioxidant activity (Figure 1).
M. orbicularis plant parts ethanolic extracts reduce the viability of A549 lung adenocarcinoma cells
Antioxidant ability usually correlates with the antiproliferative effects of plant extracts [34,39,40]. Therefore, the antiproliferative effects of extracts from different aerial parts of M. orbicularis were tested against A549 human lung adenocarcinoma cells. A549 cells were treated with ethanolic extracts (50, 100, 150, 200 μg/ml) of M. orbicularis plant parts (leaves, fruits and stems) for 24, 48, and 72 h, and their antiproliferative effects were determined using the metabolism-dye-based MTT assay (Figure 2). All plant extracts reduced the number of metabolically active A549 cells in a concentration and time dependent manner (Figure 2). M. orbicularis leaves extracts at the concentration of 100 μg/ml caused a significant (P< 0.05) decrease in the number of A549 cells starting at 48 hours, compared to control vehicle-treated cells (Figure 2A). M. orbicularis fruits extract on the other hand significantly (P< 0.0001) reduced A549 cell number at the same starting concentration of 100 μg/ml, but at the earlier time point of 24 hours (Figure 2B). Figure 2C, shows that M. orbicularis stems ethanolic extracts required a higher concentration (150 μg/ml) to cause a significant (P< 0.0001) decrease in A549 cell number, at the later time point of 72 hours. These data show that all M. orbicularis parts ethanolic extracts can reduce the viability of A549 cells. Furthermore, M. orbicularis fruits ethanolic extracts seem to be the most potent at decreasing the viability of A549 cells.
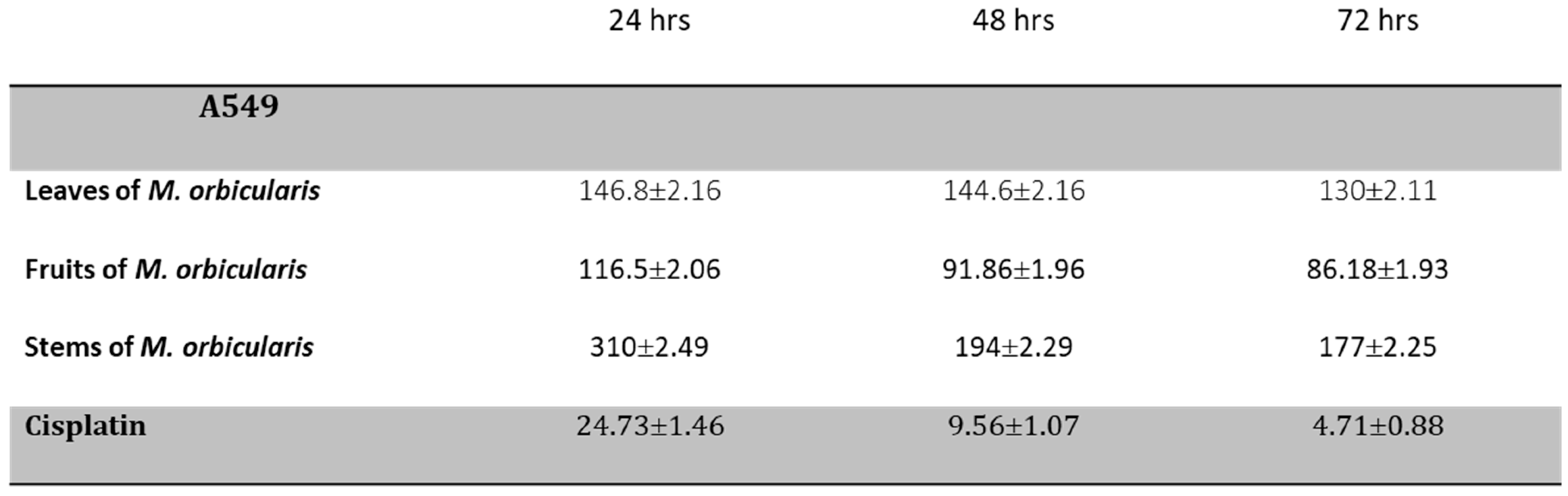
Calculation of IC50 values of the different plant extracts confirmed the time- and concentration-dependent cytotoxic effect of M. orbicularis extracts. Table 1 reveals that indeed M. orbicularis fruits ethanolic extracts have the lowest IC50 value of A549 cell viability (116.5± 2.06 μg/ml at 24 hours, 91.86±1.96 μg/ml at 48 hours, and 86.18±1.93 μg/ml at 72 hours). Additionally, A549 cells were treated with the chemotherapeutic agent, cisplatin, which potently decreases A549 cell viability (Table 1). M. orbicularis fruits ethanolic extracts show a decent inhibitory effect when compared with cisplatin (around 10 times higher IC50 at 48 hours). Given that M. orbicularis fruits ethanolic extracts were the most effective at reducing lung cancer cell viability, we decided to perform the next experiments using M. orbicularis ethanolic fruit extracts.
- Table 1. IC50 values (in μg/ml) of ethanolic extracts of parts of M. orbicularis used to treat A549 cells for 24, 48 and 72 hours. The IC50 values of cisplatin in A549 cells are also listed for comparison.
The antiproliferative effects of M. orbicularis ethanolic extracts appear to be specific to cancer cells. M. orbicularis fruits ethanolic extracts did not decrease the number of normal human neonatal fibroblast cells, even at the highest tested concentration of 200 μg/ml (Figure 2D). In comparison, the same concentration of the fruits extracts caused more than 90% decrease in the number of A549 cells at 48 h, compared to control vehicle-treated cells (Figure 2B).
M. orbicularis fruit ethanolic extracts induce apoptosis of A549 lung adenocarcinoma cells
Herbal extracts have been reported to act by inducing apoptosis in cancer cells [16]. To assess whether M. orbicularis reduced the viability of A549 cells by inducing apoptosis, we treated A549 cells with M. orbicularis fruit extracts for 48 h and used Western blotting to evaluate the protein levels of apoptosis effector proteins PARP-1 and proCaspase 3, the anti-apoptotic protein BCL-2 and the pro-apoptotic protein BAX, hallmark proteins of apoptosis. Activation of Caspase 3 is a central irreversible event in the process of apoptosis. Once activated Caspase 3 will drive apoptosis by proteolytically digesting its substrates including DNA and proteins like PARP-1 [41]. Initiation of apoptosis involves an increase in protein levels of the pro-apoptotic protein BAX, which acts by forming channels in the mitochondrial membrane, leading to the release of cytochrome C, initiating apoptosis. On the other hand, BCL-2 is an anti-apoptotic protein which prevents the release of cytochrome C from the mitochondrial membrane, thereby inhibiting apoptosis [42]. Figure 3 shows a significant decrease (P < 0.01) in the protein levels of full length PARP-1 and proCaspase 3 in A549 cells treated with M. orbicularis fruit extracts. This decrease seems to be dose dependent at 100 and 150 g/ml concentrations of M. orbicularis fruit extracts. In addition, the protein levels of BAX have decreased while BCL-2 protein levels have increased leading to a decrease in the protein ratio of BCL-2/BAX, a ratio that determines the balance between anti- versus pro-apoptotic events. Indeed, the ratio of BCL-2/ BAX was dose dependently and significantly (P < 0.05), decreased by treatment of A549 cells with 150 g/ml of M. orbicularis fruit extracts. These results indicated that M. orbicularis fruit extracts dose dependently induced apoptosis in A549 cells and that apoptosis may be responsible, at least in part, for M. orbicularis-induced cell death of A549 cells.
Ethanolic extracts of fruits of M. orbicularis reduce the migration of A549 lung adenocarcinoma cells
Cell migration is critical for proper development as well as physiological processes in adult tissues. Cell migration is also elevated in certain pathological states such as cancer. Increased cell migration may lead to enhanced cancer metastasis contributing to a malignant cancer phenotype. Non-small cell lung cancer cells (NSCLC) such as A549 cells are notorious for their highly invasive and migration potentials. We tested the ability of the ethanolic extracts of M. orbicularis fruits to inhibit A549 cell migration using a wound healing scratch assay. Figure 4 shows the outcome of the wound healing scratch assay where M. orbicularis fruits inhibited migration of A549 cells. A549 cells treated with 100 and 150 μg/ml have traveled less distance to cover the scratched area than control cells, 12 h following scratching. Figure 4B shows a significant anti-migration activity (P < 0.05) due to treatment of A549 cells with 150 μg/ml of M. orbicularis fruit extracts. This shows that M. orbicularis may attenuate metastatic properties of A549 cells by inhibiting cell migration, a hallmark of metastasis.
Ethanolic extracts of M. orbicularis fruits exhibit potent antihemolytic properties
One reason behind preference of herbal preparations and plant extracts over synthetic drugs is the general public conception that they are of natural source and therefore “safe” [30]. A drug safety profile includes the ability to safely administer a drug or herbal preparation by systemic infusion into a patient through the blood. Thus, it is empirical to test if extracts of M. orbicularis can impact the hemolysis of red blood cells (RBCs). A hemolysis assay was performed to test if M. orbicularis fruits ethanolic extracts can induce hemolysis of RBCs. Ethanolic extracts of M. orbicularis fruits did not cause hemolysis of RBCs (data not shown).
This result prompted us to test the ability of the ethanolic extracts from the fruits of M. orbicularis to protect RBCs against hemolysis which usually results from the use of chemotherapeutic agents in cancer therapy. To this end, an anti-hemolysis assay was performed. Indeed, pretreatment of RBCs with different concentrations (40, 50, or 100 μg/ml) of the ethanolic extracts from the fruits of M. orbicularis protected RBCs from H2O2-induced hemolysis (Figure 5). Figure 5 shows that 40, 50, or 100 μg/ml of ethanolic extracts of M. orbicularis fruits significantly inhibited hemolysis of RBCs. For example, at 100 μg/ml fruit extracts of M. orbicularis caused a significant 75.65 % inhibition of RBCs hemolysis. This result sheds the light on the safety of fruit extracts of M. orbicularis and underscores the potential use of these extracts in combination therapy with classical therapeutic agents, known to induce RBCs hemolysis.
Ethanolic extracts of fruits of M. orbicularis enhance cisplatin-induced cytotoxicity in A549 lung cancer cells
Cisplatin has potent anti-proliferative effects against A549 cells [43], and the anti-proliferative effect of cisplatin were confirmed in this study. The IC50 values of cisplatin-induced reduction of viability of A549 cells were 24.73, 9.56 and 4.72 μg/ml at 24, 48 and 72 hours, respectively (Table 1 and Figure 6A). These values are comparable to the values previously reported in the literature [43,44].
Many types of cancer including lung cancer develop resistance to chemotherapy and as a result they usually relapse following chemotherapy with cisplatin or other chemotherapeutic agents [45]. Combination therapy can help to overcome the chemoresistance that develops against chemotherapeutic agents. When combined with chemotherapy, plant extracts with anti-cancerous properties may offer a way of overcoming chemoresistance and cancer relapse. This is because plant preparations usually have a different mechanism of action than chemotherapeutic agents [16]. Therefore, we assessed the effect of a combination treatment using ethanolic extracts of M. orbicularis fruits on cisplatin-induced killing of A549 cells. A549 cells were treated for 48 h with a low-dose of cisplatin (5 μg/ml) in combination with 100 μg/ml of M. orbicularis fruits extracts. Cell viability assays showed that tat these concentrations, cisplatin alone and M. orbicularis fruits extracts alone have moderate cytotoxicity levels (Table 1, Figure 2B, and Figure 6A). Hence, any enhanced cytotoxic effects can be noticed using these chosen concentrations. For example, the cell viability of A549 decreased significantly (P< 0.0001) from 85.9 % in A549 cells treated with cisplatin alone to 25.8% in cells treated with the combination of cisplatin and M. orbicularis fruits extracts (Figure 6B). In addition, the combination treatment enhanced the cytotoxic effect of M. orbicularis fruits extracts where the A549 cell viability decreased from 34.4% in cells treated with M. orbicularis fruits extracts alone to 25.8% in the combination treatment (Figure 6B). These results showed that M. orbicularis fruits ethanolic extracts sensitized A549 cells to the cytotoxic effects of cisplatin.
Ethanolic extracts of fruits of M. orbicularis augment cisplatin-induced aggregation of A549 lung cancer cells
Cisplatin is a standard chemotherapeutic drug used to manage different types of cancers [5], including lung cancer [46]. When cancer cells undergo metastasis, they lose their ability to adhere together in order to invade other organs [47]. When a cancer is treated with chemotherapeutic drugs, cell-cell adhesion usually increases in order to reduce metastasis [48]. Chemoresistance of cancer cells to drugs such as cisplatin results in altered cell adhesion patterns, permitting cancer cells to metastasize even under treatment [49]. Therefore, we next sought to examine the effect of M. orbicularis fruit extracts on the cell–cell adhesion of A549 cells. Toward this, a cell aggregation assay was performed, where A549 cells were incubated, while shaking in a non-adherent cell culture plastic plate, in the absence or presence of the extract alone, cisplatin alone, or the extract combined with cisplatin. Figure 7 shows that after 1 h of treatment of A549 cells with 100 and 150 μg/ml of M. orbicularis fruit extracts, the percentages of cell aggregation significantly (P<0.001) increased by 47 % and 65 %, respectively, compared to control cells (Figure 6 A, B). However, the percentage of cell aggregation of A549 is 44 % after treatment with cisplatin (5 μg/ml) alone and significantly increased to 71.5 % after the co-treatment with cisplatin (5 μg/ml) and M. orbicularis (150 μg/ml) (Figure 5A, B). These results showed that the M. orbicularis extract significantly augmented cisplatin-induced aggregation of A549 lung cancer cells aggregation.
Total polyphenols and flavonoids contents of M. orbicularis fruits ethanolic extracts
The anticancer properties of plant extracts are frequently attributed to their rich phytochemical content [29,50]. Results of this study have indicated that M. orbicularis fruits ethanolic extracts have strong antioxidant effects. These effects could be mediated by polyphenol and flavonoids [26,51,52] of M. orbicularis fruits extracts among other phytochemicals. Furthermore, the phytochemicals of M. orbicularis have not been previously identified. In order to characterize the phytochemical contents of ethanolic extract of M. orbicularis fruits, we measured the total phenolics and flavonoids contents of M. orbicularis fruits extracts. Indeed, the total phenolics content was 1.1583± 0.00005 mg GAE/g of dry matter of Medicago orbicularis fruits. Total flavonoids content was 0.0246± 0.00003 mg quercetin equivalents/ g of dry matter of Medicago orbicularis fruits. These results show that Medicago orbicularis fruits are rich in phenolics and flavonoids and may explain the anticancer properties of Medicago orbicularis fruits against A549 cells.
Identification of phytochemical composition of M. orbicularis fruits ethanolic extracts by GC/MS
To further characterize the phytochemical contents of M. orbicularis fruits, GC/MS analysis was used Figure 8. Table 2 lists 18 of the major compounds of M .orbicularis fruits as identified by GC/MS, by comparing their mass spectral fragmentation patterns to those of known compounds listed in the NIST library.
IV. Discussion
Cancer is a complex and devastating disease which affects millions of lives worldwide. Lung cancer, particularly non-small cell lung cancer, has the highest mortality rate among malignant tumors across the globe [43]. Current treatments for lung cancer such as chemotherapy and radiotherapy are associated with considerable toxicity and other side effects. In addition, lung cancer patients develop resistance to therapy and suffer from lung cancer recurrence as early as 6 months following therapy [3,4]. In this regard, there is a revived research interest in using herbal- and plant-based therapies as sources of anti-cancer bioactives. Several herbal remedies have been shown to have efficacy and minimal toxicity and side effects for disease treatment [12,13]. In addition, plants and their phytochemicals are being used as starting scaffolds for developing more potent anticancer agents [15,30]. Historically, plants or herbs and their extracts were used for the development of several important anti-cancer agents such as Paclitaxel, Camptothecin, and Vincristine [7,8,9,10,11,30]. Furthermore, several intricate biochemical pathways are simultaneously dysregulated in cancer [6,15], and herbal-based or herbal-derived remedies can target several molecular pathways and can become an alternative or complementary treatment to conventional cancer therapies [7,12,13,14,15,53,54]. Relatedly, multiple recent in vitro and in vivo studies demonstrated that concurrent use of natural products with conventional treatments (chemo- and radio-therapy) can synergistically sensitize tumors to therapy, enhancing therapeutic efficacy and reducing toxicities [15]. Many studies have documented the promising use of herbal plant extracts against cancer cell lines in vitro and in vivo in animal models of cancer, and are now being tested in clinical trials. These studies have shown that herbal-based remedies have antioxidant, antihemolytic, and apoptosis-inducing effects. They can also impact cell proliferation, aggregation and adhesion, migration, and metastasis [16,17,55].
Medicago orbicularis L. Bartal is an understudied plant species as far as its therapeutic effects are concerned. Except for a study reporting a traditional use of M. orbicularis for heart diseases management in Turkey [33], there are no reports on the therapeutic applications or phytochemical composition of M. orbicularis. Notably, M. orbicularis anti-cancerous activities have not been explored yet [18]. It was therefore pertinent to study phytochemical composition, antioxidant capacity, antihemolytic properties, and the effects of M. orbicularis on cell proliferation and apoptosis, cell aggregation, and cell migratory potential against human lung adenocarcinoma A549 cells, in addition to the combinatorial effect of a treatment of both the chemotherapeutic agent cisplatin and M. orbicularis on A549 cells.
There are no studies currently on the anti-oxidant potential of M. orbicularis, but other species of the genus Medicago such as M. polymorpha, M. sativa, M. arabica, and M. truncatula were shown to have potent anti-oxidant activities and to be rich in phytochemicals [22,23]. Since anti-oxidant activity is often times accompanied by therapeutic effects [27,28,29,30,31,32], this study was initiated with an evaluation of the antioxidant activity of the ethanolic extract of different plant parts of M. orbicularis including its leaves, fruits and stems. All plant parts of M. orbicularis showed anti-oxidant potential and the leaves showed the highest antioxidant activity.
To test if the anti-oxidant capacity of M. orbicularis is accompanied by an anti-cancerous effect, we tested the cytotoxic effect of M. orbicularis (leaves, fruits and stems) on A549 human lung adenocarcinoma cells. All M. orbicularis plant parts inhibited proliferation of A549 cells, but M. orbicularis fruits exhibited the highest reduction of viability of A549 cells and were chosen to perform the rest of the experiments of the study. The National Cancer Institute (NCI, USA) considers an IC50 of 30 μg/mL to indicate a strong cytotoxic activity and a promising candidate for the further purification of a crude extract, and an IC50 of 31–200 μg/ml to indicated moderate cytotoxicity [56]. The IC50 of the fruit extracts at 72 h of treatment was 86.18± 1.93 μg/mL, indicating that this crude extract has moderate cytotoxicity. To date, no other study has reported the cytotoxicity levels of M. orbicularis against cancerous cells. Importantly, the cytotoxic effects of the fruit extracts were showed selectivity to A549 cancerous cells and did not affect human neonatal fibroblast cells even at high concentrations. This indicated that M. orbicularis fruits extracts may not have side effects when used in vivo.
Plant bioactives can decrease viability of cancer cells through several mechanism including induction of apoptosis [7,12,13,14,15,53,54]. Cytotoxic agents can cause an increase in the expression of the pro-apoptotic protein BAX that makes pores in the mitochondrial membrane leading to the release of cytochrome C [57]. Release of cytochrome C initiates the execution phase of apoptosis and activates Caspase 9 which cleave pro-Caspase 3 into active Caspase 3, the main executer effector caspase [58,59]. Active Caspase 3 can cleave many protein substrates [60], such as caspase activated DNAse that fragments genomic DNA [61] and PARP-1, to promote apoptosis [59]. PARP-1 is required for DNA repair where it adds poly (ADP ribose) polymers to acceptor proteins to promote repair of DNA damage. PARP-1 cleavage by Caspase 3 inactivates PARP-1 leading to reduction of DNA repair and promotion of apoptosis [58,59,62,63]. These events are recognized as the intrinsic apoptotic pathway. The anti-apoptotic protein BCL-2 can inhibit the release of cytochrome C into the cytoplasm, thereby attenuating the intrinsic apoptotic pathway [59,64]. Western blotting analysis of lysates of A549 cells treated with 100 and 150 g/ml M. orbicularis fruit extract indicated that M. orbicularis fruit extracts activated Caspase-3, inactivated PARP-1, and lowered the expression of BCL-2 and increased the expression of BAX, suggesting the activation of the intrinsic apoptotic pathway. The results showed a significant decrease in the protein levels of full-length PARP-1 and proCaspase 3 in A549 cells in a dose-dependent manner. Moreover, there was a dose dependent decrease in the BCL-2/BAX ratio. This is the first report showing that M. orbicularis-induced death of A549 cells is mediated, at least partly, by the intrinsic apoptotic machinery. Other mechanisms of cell death, such as autophagy, necroptosis, or pyroptosis, may also be induced by M. orbicularis, and need to be tested in future studies.
As part of the safety profile of a cancer drug, its systemic infusion through the blood of a patient should not cause any hemolysis of red blood cells [65]. M. orbicularis fruit extracts did not cause any hemolysis of RBCs. Even more, the fruits extracts protected RBCs against hemolysis. This attested to the possible safety of these extracts in future in vivo or clinical studies.
Metastasis is the major culprit behind cancer-associated mortality [66,67,68]. Cell migration is a key requirement for the metastatic process. Although cell migration is essential in many physiological processes such as wound repair, tissue formation, and proper immune response, its deregulation is required for the initial steps of cancer metastasis as cells spread away from the primary tumor site. Cell migration involves the downregulation of cell junction and adhesion proteins and proteolysis of ECM proteins by metalloproteinases (MMPs) [67,69] . We assessed the effect of the fruit extract on migration of A549 using a wound healing assay. Non-cytotoxic concentrations of the fruit extracts of M. orbicularis inhibited migration of A549 cells in a concentration-dependent manner. This indicated that M. orbicularis fruits could attenuate the malignant phenotype of A549 cells by inhibiting cell migration and consequently metastasis. This result invites future investigation of the ability of M. orbicularis to inhibit the invasive potential of A549 cells through its action on MMPs, and proteins of the process of epithelial-to-mesenchymal transition (EMT), a hall mark of carcinogenesis and metastasis [27,28,67,68,70].
In order for cancer cells to migrate and metastasize, they should lose their adhesion to neighboring cells allowing them to migrate and invade at secondary tumor sites and organs [27,28,67,68,70]. Cancer therapeutics can act by strengthening cell adhesion and aggregation to prevent cell migration and metastasis [71]. In this study, an aggregation assay showed that M. orbicularis fruits significantly enhanced cell aggregation, signifying that the extract enhanced adhesion of A549 cells. This result further attests to the ability of M. orbicularis to attenuate the malignant phenotype of A549 cells. Future, studies should focus on elucidating the molecular mechanisms underlying this finding, include examination of cell adhesion proteins, connexins gap junction proteins, and EMT markers such as E-cadherin, N-cadherin, vimentin, Snail, among others [72].
Apart from their direct suppressive effects on carcinogenesis and cancer metastasis, herbal remedies and plant extracts have been used to complement conventional therapies since they can target several molecular pathways other than those targeted by conventional therapy, and therefore may overcome refractoriness that develops against cancer therapeutics [7,12,13,14,15,53,54]. When combined with chemotherapy, plant-based therapies can enhance response to chemotherapy, help overcome resistance to therapy, decrease the severity of side-effects of chemotherapy including cancer-related fatigue [7,12,13,14,15]. Chemotherapeutic agents, including cisplatin can become refractory in lung cancer patients, and as result the cancer may relapse with poor prognosis [3,4]. Combination of cisplatin with a multitargeted herbal remedy may circumvent this problem. In this study, the cytotoxic effect of cisplatin alone on A549 cells was confirmed. The IC50 of cisplatin with A549 that we obtained are similar to those reported in the literature [43]. In order to determine the impact of a combination treatment on cisplatin-induced death of A549 cells, we applied a co-treatment of cisplatin and the ethanolic extract of M. orbicularis fruits. The results showed that the combination treatment significantly augmented cisplatin-induced decrease in cell viability of A549 cells, when compared to either the extract alone, or cisplatin alone. These results show that M. orbicularis may be a source of a complementary therapy for lung cancer, in combination with chemotherapy. The combination treatment showed a similar result when tested on aggregation of A549 cells. Co-treatment of M. orbicularis fruits extracts and cisplatin showed that the combination treatment significantly augmented the increase in aggregation of A549 cells, when compared to either the extract alone, or cisplatin alone. Taken together, these results support M. orbicularis as a source for development of anti-lung cancer drug candidates that can complement chemotherapy.
M. orbicularis phytochemical composition has not been defined yet. But, other Medicago species have been reported to be rich in phytochemicals [21,22,23], which have anticancer activities [24,25,26,27,28]. Natural polyphenols are plant secondary metabolites which have two or more phenol rings. Polyphenols health benefits include being antioxidant, antidiabetic, cardioprotective, neuroprotective. Relatedly, phenolics have been reported to have strong anticancer effects through various mechanisms including removal of cancer cells, inhibition of cell cycle, and induction of apoptosis, inhibition of metastasis among others [52]. Flavonoids are natural polyphenols with documented anticancer and antioxidant properties. Flavonoids have been reported to inhibit carcinogenesis by suppressing oxidative stress through their antioxidant activities [26,51]. Our results indicated that M. orbicularis fruits ethanolic extracts have strong antioxidant and cytotoxic effects against A549 cells. These effects could be mediated by polyphenols and flavonoids of M. orbicularis fruits extracts among other phytochemicals. In this study, Medicago orbicularis fruits were shown to be rich in phenolics and flavonoids, which may explain the anti-oxidant and anticancer properties of Medicago orbicularis fruits against A549 cells.
To further evaluate the phytochemical composition and establish a more comprehensive profile of the phytochemical constituents of Medicago orbicularis fruits, GC-MS analysis was conducted. We identified 18 prominent peaks that correspond to bioactive compounds of Medicago orbicularis fruits. Medicago orbicularis ethanol extract was found to be a complex mixture of various classes of phytochemicals, including flavonoids, alkaloids, diterpenes, triterpenes, sesquiterpenes, sterols, alcohols, aldehydes, fatty acids, and hydrocarbons. Among the diverse array of phytochemicals present in the extract, several compounds were correlated with therapeutic properties, including anti-cancerous activities. For example, lupeol and its derivatives, which were the most frequently identified compounds, have been recently demonstrated to possess a diverse array of pharmacological activities [73]. These activities included anticancer, antimicrobial, and antidiabetic effects, with certain lupeol derivatives exhibiting greater potency than lupeol [74]. Future investigation should focus on more comprehensive assessment of the plant extracts of M. orbicularis, using more advanced techniques such as preparative HPLC followed by LC-MS/MS or NMR, to identify the bioactives responsible for the therapeutic effects of crude extracts from this plant.
In conclusion, this study shows for the first time that the hydroalcoholic extract of M. orbicularis has anti-oxidant, anti-hemolytic, and cytotoxic properties against A549 lung cancer cells. The decrease in cell viability could be at least partly mediated by the intrinsic pathway of apoptosis, involving PARP-1, Caspase 3, and BCL-2/BAX. M. orbicularis was also able to suppress cell migration and enhance cell aggregation of A549 cells. These activities could be attributed to the phytochemicals present in M. orbicularis such as polyphenols and falvonoids, and the other compounds that were identified by GC/MS. These properties place M. orbicularis as potential source of new candidate effective natural agents for the treatment of the lung cancer.
Author Contributions
Conceptualization: SN, Experimental design: SN, FK, and AS; Formal analysis: AS, FK, and SN did analysis and interpretation of data; Resources: SN, FK, and AS; Performed experiments: IO, NA, and FS; Writing: original draft preparation, AS, IO, FS; Writing: review and editing, all authors; Supervision: SN, FK, and AS; Funding acquisition, SN, FK, and AS. All authors read and approved the final manuscript.
Funding
This work was supported by the a grant from the Lebanese University to SN and student grants number QUST-1-BRC-2022-315; QUST-1-BRC-2022-316, QUST-1-BRC-2023-836; and QUST-1-BRC-2023-846 to AS. Publication fees APC were covered by Qatar National Library (QNL).
Acknowledgement
The authors acknowledge the help they received to perform the spectrophotometric assays and GC/MS analysis, which were accomplished in the Central Laboratories Unit, Qatar University.
Conflict of Interest
The authors declare that there are no conflicts of interest.
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J Clin 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- National Cancer Institute. Cancer Stat Facts: Lung and Bronchus Cancer. 2023.
- Rossi, A.; Sacco, P.C.; Sgambato, A.; Casaluce, F.; Santabarbara, G.; Palazzolo, G.; Maione, P.; Gridelli, C. Optimal drugs for second-line treatment of patients with small-cell lung cancer. Expert Opin Pharmacother 2016, 17, 969–976. [Google Scholar] [CrossRef] [PubMed]
- Rudin, C.M.; Ismaila, N.; Hann, C.L.; Malhotra, N.; Movsas, B.; Norris, K.; Pietanza, M.C.; Ramalingam, S.S.; Turrisi, A.T., 3rd; Giaccone, G. Treatment of Small-Cell Lung Cancer: American Society of Clinical Oncology Endorsement of the American College of Chest Physicians Guideline. J Clin Oncol 2015, 33, 4106–4111. [Google Scholar] [CrossRef] [PubMed]
- Dasari, S.; Tchounwou, P.B. Cisplatin in cancer therapy: molecular mechanisms of action. Eur J Pharmacol 2014, 740, 364–378. [Google Scholar] [CrossRef] [PubMed]
- Almquist, D.; Mosalpuria, K.; Ganti, A.K. Multimodality Therapy for Limited-Stage Small-Cell Lung Cancer. J Oncol Pract 2016, 12, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Kwon, C.Y.; Lee, B.; Kim, K.I.; Lee, B.J. Herbal medicine on cancer-related fatigue of lung cancer survivors: Protocol for a systematic review. Medicine (Baltimore) 2020, 99, e18968. [Google Scholar] [CrossRef] [PubMed]
- Fridlender, M.; Kapulnik, Y.; Koltai, H. Plant derived substances with anti-cancer activity: from folklore to practice. Front Plant Sci 2015, 6, 799. [Google Scholar] [CrossRef]
- Buyel, J.F. Plants as sources of natural and recombinant anti-cancer agents. Biotechnol Adv 2018, 36, 506–520. [Google Scholar] [CrossRef]
- Cragg, G.M.; Newman, D.J. Plants as a source of anti-cancer agents. J Ethnopharmacol 2005, 100, 72–79. [Google Scholar] [CrossRef]
- Shaik, B.B.; Katari, N.K.; Jonnalagadda, S.B. Role of Natural Products in Developing Novel Anticancer Agents: A Perspective. Chem Biodivers 2022, 19, e202200535. [Google Scholar] [CrossRef]
- Konkimalla, V.B.; Efferth, T. Evidence-based Chinese medicine for cancer therapy. J Ethnopharmacol 2008, 116, 207–210. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.Y.J.; Meng, M.; Liu, Y.; Su, T.; Kwan, H.Y. Medicinal herbs and bioactive compounds overcome the drug resistance to epidermal growth factor receptor inhibitors in non-small cell lung cancer. Oncol Lett 2021, 22, 646. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Li, S.; Li, C.; Li, T.; Huang, Y. Remodeling tumor microenvironment with natural products to overcome drug resistance. Front Immunol 2022, 13, 1051998. [Google Scholar] [CrossRef] [PubMed]
- Nisar, S.; Masoodi, T.; Prabhu, K.S.; Kuttikrishnan, S.; Zarif, L.; Khatoon, S.; Ali, S.; Uddin, S.; Akil, A.A.; Singh, M. , et al. Natural products as chemo-radiation therapy sensitizers in cancers. Biomed Pharmacother 2022, 154, 113610. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Jeong, D.; Song, M.; Kim, B. Recent Advances in Anti-Metastatic Approaches of Herbal Medicines in 5 Major Cancers: From Traditional Medicine to Modern Drug Discovery. Antioxidants (Basel) 2021, 10. [Google Scholar] [CrossRef] [PubMed]
- Anwar, S.; Malik, J.A.; Ahmed, S.; Kameshwar, V.A.; Alanazi, J.; Alamri, A.; Ahemad, N. Can Natural Products Targeting EMT Serve as the Future Anticancer Therapeutics? Molecules 2022, 27. [Google Scholar] [CrossRef] [PubMed]
- Zitouna, N.; Marghali, S.; Gharbi, M.; Haddioui, A.; Trifi-Farah, N. Sequence divergence of microsatellites for phylogeographic assessment of Moroccan Medicago species. Genet Mol Res 2014, 13, 1548–1562. [Google Scholar] [CrossRef]
- Morshedi, Z.; Assadi, M.; Small, E.; Dehshiri, M.M.; Mehregan, I. Systematic Studies on Populations of Medicago orbicularis (L.) Bartal: Molecular, Morphological and Ecological Characterizations. Journal of Genetic Resources 2022, 8, 178–187. [Google Scholar] [CrossRef]
- Karam, N.; Choueiry, Z.; Al-Beyrouthy, J.; Shehadeh, A.; Chalak, L.; Yazbek, M. Phenotypic diversity of Medicago crop wild relatives growing in Lebanon. Genetic Resources and Crop Evolution 2023, 70, 1487–1499. [Google Scholar] [CrossRef]
- Bora, K.S.; Sharma, A. Phytochemical and pharmacological potential of Medicago sativa: a review. Pharm Biol 2011, 49, 211–220. [Google Scholar] [CrossRef]
- Khan, M.I.; Asad, S.; Zaman, G.; Rehman, H.; Rehman, S.; Iqbal, A.; Ullah, A.; Ullah, I.; Ali, S. Antioxidant And Cytotoxic Activities Of Crude Methanolic Extract Of Medicago Polymorpha. IOSR Journal of Pharmacy 2013, 3, 32–37. [Google Scholar]
- Usman, M.; Khan, W.R.; Yousaf, N.; Akram, S.; Murtaza, G.; Kudus, K.A.; Ditta, A.; Rosli, Z.; Rajpar, M.N.; Nazre, M. Exploring the Phytochemicals and Anti-Cancer Potential of the Members of Fabaceae Family: A Comprehensive Review. Molecules 2022, 27. [Google Scholar] [CrossRef] [PubMed]
- Choudhari, A.S.; Mandave, P.C.; Deshpande, M.; Ranjekar, P.; Prakash, O. Phytochemicals in Cancer Treatment: From Preclinical Studies to Clinical Practice. Front Pharmacol 2019, 10, 1614. [Google Scholar] [CrossRef] [PubMed]
- Anifowose, S.O.; Alqahtani, W.S.N.; Al-Dahmash, B.A.; Sasse, F.; Jalouli, M.; Aboul-Soud, M.A.M.; Badjah-Hadj-Ahmed, A.Y.; Elnakady, Y.A. Efforts in Bioprospecting Research: A Survey of Novel Anticancer Phytochemicals Reported in the Last Decade. Molecules 2022, 27. [Google Scholar] [CrossRef] [PubMed]
- Slika, H.; Mansour, H.; Wehbe, N.; Nasser, S.A.; Iratni, R.; Nasrallah, G.; Shaito, A.; Ghaddar, T.; Kobeissy, F.; Eid, A.H. Therapeutic potential of flavonoids in cancer: ROS-mediated mechanisms. Biomed Pharmacother 2022, 146, 112442. [Google Scholar] [CrossRef] [PubMed]
- Mesmar, J.; Abdallah, R.; Hamade, K.; Baydoun, S.; Al-Thani, N.; Shaito, A.; Maresca, M.; Badran, A.; Baydoun, E. Ethanolic extract of Origanum syriacum L. leaves exhibits potent anti-breast cancer potential and robust antioxidant properties. Front Pharmacol 2022, 13, 994025. [Google Scholar] [CrossRef] [PubMed]
- AlKahlout, A.; Fardoun, M.; Mesmar, J.; Abdallah, R.; Badran, A.; Nasser, S.A.; Baydoun, S.; Kobeissy, F.; Shaito, A.; Iratni, R.; et al. Origanum syriacum L. Attenuates the Malignant Phenotype of MDA-MB231 Breast Cancer Cells. Front Oncol 2022, 12, 922196. [Google Scholar] [CrossRef]
- Imran Khan, M. Antioxidant And Cytotoxic Activities Of Crude Methanolic Extract Of Medicago Polymorpha. IOSR Journal of Pharmacy (IOSRPHR) 2013, 03, 32–37. [Google Scholar] [CrossRef]
- Shaito, A.; Thuan, D.T.B.; Phu, H.T.; Nguyen, T.H.D.; Hasan, H.; Halabi, S.; Abdelhady, S.; Nasrallah, G.K.; Eid, A.H.; Pintus, G. Herbal Medicine for Cardiovascular Diseases: Efficacy, Mechanisms, and Safety. Front Pharmacol 2020, 11, 422. [Google Scholar] [CrossRef]
- Alsamri, H.; Athamneh, K.; Pintus, G.; Eid, A.H.; Iratni, R. Pharmacological and Antioxidant Activities of Rhus coriaria L. (Sumac). Antioxidants (Basel) 2021, 10. [Google Scholar] [CrossRef]
- Visnevschi-Necrasov, T.; Barreira, J.C.; Cunha, S.C.; Pereira, G.; Nunes, E.; Oliveira, M.B. Advances in isoflavone profile characterisation using matrix solid-phase dispersion coupled to HPLC/DAD in Medicago species. Phytochem Anal 2015, 26, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Güleç, M. , Erarslan, Z. B. & Kültür, Ş.. The Medicinal Plants Traditionally Used Against Cardiovascular Diseases In Türkiye. International Journal of Traditional and Complementary Medicine Research 2023, 4, 81–96. [Google Scholar] [CrossRef]
- NASREDDINE, S.; MCHEIK, M.; KHALIL, M.; El-Rashed, Z.; DAHER, A.; KHALIFE, A. The antioxidant, antibacterial, antihemolytic and epithelial ovarian cancer antiproliferative activities of the lebanese plant salvia libanotica. Asian Journal of Science and Technology 2018, 09, 8695–8703. [Google Scholar]
- Malagoli, D. A full-length protocol to test hemolytic activity of palytoxin on human erythrocytes. Invertebrate Survival Journal 2007, 4, 92–94. [Google Scholar]
- James, O.; Alewo, I.M. In vitro Antihemolytic Activity of Gymnema Sylvestre Extracts Against Hydrogen Peroxide (H2O2) Induced Haemolysis in Human Erythrocytes. American Journal of Phytomedicine and Clinical Therapeutics 2014, 2, 861–869. [Google Scholar]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. American journal of Enology and Viticulture 1965, 16, 144–158. [Google Scholar] [CrossRef]
- Chang, C.-C.; Yang, M.-H.; Wen, H.-M.; Chern, J.-C. Estimation of total flavonoid content in propolis by two complementary colorimetric methods. Journal of food and drug analysis 2002, 10. [Google Scholar]
- Katarina, Š.; Živković, J.; Zdunić , G.; Gođevac, D.; Đorđević, B.; Dojčinović, B.; Đorđević, N. Phenolic and mineral profiles of four Balkan indigenous apple cultivars monitored at two different maturity stages. Journal of food composition and analysis 2014, v. 35, pp. 101-111-2014 v.2035 no.2012. [CrossRef]
- Rolim, P.M.; Fidelis, G.P.; Padilha, C.E.A.; Santos, E.S.; Rocha, H.A.O.; Macedo, G.R. Phenolic profile and antioxidant activity from peels and seeds of melon (Cucumis melo L. var. reticulatus) and their antiproliferative effect in cancer cells. Brazilian Journal of Medical and Biological Research 2018, 51. [Google Scholar] [CrossRef]
- Li, J.; Yuan, J. Caspases in apoptosis and beyond. Oncogene 2008, 27, 6194–6206. [Google Scholar] [CrossRef]
- Cory, S.; Roberts, A.W.; Colman, P.M.; Adams, J.M. Targeting BCL-2-like Proteins to Kill Cancer Cells. Trends Cancer 2016, 2, 443–460. [Google Scholar] [CrossRef]
- Tseng, C.Y.; Lin, C.H.; Wu, L.Y.; Wang, J.S.; Chung, M.C.; Chang, J.F.; Chao, M.W. Potential Combinational Anti-Cancer Therapy in Non-Small Cell Lung Cancer with Traditional Chinese Medicine Sun-Bai-Pi Extract and Cisplatin. PloS one 2016, 11, e0155469. [Google Scholar] [CrossRef] [PubMed]
- Omairi, I.; Kobeissy, F.; Nasreddine, S. Anti-Oxidant, Anti-Hemolytic Effects of Crataegus aronia Leaves and Its Anti- Proliferative Effect Enhance Cisplatin Cytotoxicity in A549 Human Lung Cancer Cell Line. Asian Pacific journal of cancer prevention : APJCP 2020, 21, 2993–3003. [Google Scholar] [CrossRef] [PubMed]
- De Giorgi, U.; Casadei, C.; Bergamini, A.; Attademo, L.; Cormio, G.; Lorusso, D.; Pignata, S.; Mangili, G. Therapeutic challenges for cisplatin-resistant ovarian germ cell tumors. Cancers 2019, 11, 1584. [Google Scholar] [CrossRef] [PubMed]
- Hotta, K.; Matsuo, K.; Ueoka, H.; Kiura, K.; Tabata, M.; Tanimoto, M. Meta-analysis of randomized clinical trials comparing cisplatin to carboplatin in patients with advanced non–small-cell lung cancer. Journal of Clinical Oncology 2004, 22, 3852–3859. [Google Scholar] [CrossRef] [PubMed]
- Leber, M.F.; Efferth, T. Molecular principles of cancer invasion and metastasis. International journal of oncology 2009, 34, 881–895. [Google Scholar] [PubMed]
- Larzabal, L.; El-Nikhely, N.; Redrado, M.; Seeger, W.; Savai, R.; Calvo, A. Differential effects of drugs targeting cancer stem cell (CSC) and non-CSC populations on lung primary tumors and metastasis. PloS one 2013, 8, e79798. [Google Scholar] [CrossRef] [PubMed]
- Maubant, S.; Cruet-Hennequart, S.; Poulain, L.; Carreiras, F.; Sichel, F.; Luis, J.; Staedel, C.; Gauduchon, P. Altered adhesion properties and alpha v integrin expression in a cisplatin-resistant human ovarian carcinoma cell line. International journal of cancer 2002, 97, 186–194. [Google Scholar] [CrossRef]
- Pareek, A.; Pant, M.; Gupta, M.M.; Kashania, P.; Ratan, Y.; Jain, V.; Pareek, A.; Chuturgoon, A.A. Moringa oleifera: An Updated Comprehensive Review of Its Pharmacological Activities, Ethnomedicinal, Phytopharmaceutical Formulation, Clinical, Phytochemical, and Toxicological Aspects. International Journal of Molecular Sciences 2023, 24, 2098. [Google Scholar] [CrossRef]
- Seglab, F.; Hamia, C.; Khacheba, I.; Djeridane, A.; Yousfi, M. High in vitro antioxidant capacities of Algerian Cleome arabica leaves’ extracts. Phytothérapie 2021, 19, 16–24. [Google Scholar] [CrossRef]
- Bhosale, P.B.; Ha, S.E.; Vetrivel, P.; Kim, H.H.; Kim, S.M.; Kim, G.S. Functions of polyphenols and its anticancer properties in biomedical research: a narrative review. Transl Cancer Res 2020, 9, 7619–7631. [Google Scholar] [CrossRef]
- Winslow, L.C.; Kroll, D.J. Herbs as medicines. Archives of internal medicine 1998, 158, 2192–2199. [Google Scholar] [CrossRef] [PubMed]
- Ho, J.W.; Leung, Y.; Chan, C. Herbal medicine in the treatment of cancer. Current Medicinal Chemistry-Anti-Cancer Agents 2002, 2, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Vong, C.T.; Chen, H.; Gao, Y.; Lyu, P.; Qiu, L.; Zhao, M.; Liu, Q.; Cheng, Z.; Zou, J. , et al. Naturally occurring anti-cancer compounds: shining from Chinese herbal medicine. Chinese Medicine 2019, 14. [Google Scholar] [CrossRef] [PubMed]
- Suffness, M.P., J.M. Assays related to cancer drug discovery. In Methods in plant biochemistry: assays for bioactivity, Hostettmann, K., Ed. Academic Press: London, 1991; Vol. 6, pp. 71-133.
- Chipuk, J.E.; Kuwana, T.; Bouchier-Hayes, L.; Droin, N.M.; Newmeyer, D.D.; Schuler, M.; Green, D.R. Direct activation of Bax by p53 mediates mitochondrial membrane permeabilization and apoptosis. Science 2004, 303, 1010–1014. [Google Scholar] [CrossRef]
- Wolf, B.B.; Green, D.R. Suicidal tendencies: apoptotic cell death by caspase family proteinases. J Biol Chem 1999, 274, 20049–20052. [Google Scholar] [CrossRef]
- Brentnall, M.; Rodriguez-Menocal, L.; De Guevara, R.L.; Cepero, E.; Boise, L.H. Caspase-9, caspase-3 and caspase-7 have distinct roles during intrinsic apoptosis. BMC Cell Biol 2013, 14, 32. [Google Scholar] [CrossRef]
- Fernandes-Alnemri, T.; Litwack, G.; Alnemri, E.S. CPP32, a novel human apoptotic protein with homology to Caenorhabditis elegans cell death protein Ced-3 and mammalian interleukin-1 beta-converting enzyme. J Biol Chem 1994, 269, 30761–30764. [Google Scholar] [CrossRef] [PubMed]
- Widłak, P. The DFF40/CAD endonuclease and its role in apoptosis. Acta Biochim Pol 2000, 47, 1037–1044. [Google Scholar] [CrossRef]
- Kaufmann, S.H.; Desnoyers, S.; Ottaviano, Y.; Davidson, N.E.; Poirier, G.G. Specific proteolytic cleavage of poly(ADP-ribose) polymerase: an early marker of chemotherapy-induced apoptosis. Cancer Res 1993, 53, 3976–3985. [Google Scholar]
- Chaitanya, G.V.; Steven, A.J.; Babu, P.P. PARP-1 cleavage fragments: signatures of cell-death proteases in neurodegeneration. Cell Commun Signal 2010, 8, 31. [Google Scholar] [CrossRef]
- Adams, J.M.; Cory, S. The BCL-2 arbiters of apoptosis and their growing role as cancer targets. Cell Death Differ 2018, 25, 27–36. [Google Scholar] [CrossRef]
- Mayer, B.; Bartolmäs, T.; Yürek, S.; Salama, A. Variability of Findings in Drug-Induced Immune Haemolytic Anaemia: Experience over 20 Years in a Single Centre. Transfus Med Hemother 2015, 42, 333–339. [Google Scholar] [CrossRef]
- Talmadge, J.E.; Fidler, I.J. AACR centennial series: the biology of cancer metastasis: historical perspective. Cancer Res 2010, 70, 5649–5669. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Settleman, J. EMT, cancer stem cells and drug resistance: an emerging axis of evil in the war on cancer. Oncogene 2010, 29, 4741–4751. [Google Scholar] [CrossRef] [PubMed]
- Thiery, J.P.; Lim, C.T. Tumor dissemination: an EMT affair. Cancer Cell 2013, 23, 272–273. [Google Scholar] [CrossRef] [PubMed]
- Trepat, X.; Chen, Z.; Jacobson, K. Cell migration. Compr Physiol 2012, 2, 2369–2392. [Google Scholar] [CrossRef] [PubMed]
- El-Hajjar, L.; Jalaleddine, N.; Shaito, A.; Zibara, K.; Kazan, J.M.; El-Saghir, J.; El-Sabban, M. Bevacizumab induces inflammation in MDA-MB-231 breast cancer cell line and in a mouse model. Cell Signal 2019, 53, 400–412. [Google Scholar] [CrossRef] [PubMed]
- Abduljauwad, S.N.; Ahmed, H.U. Enhancing cancer cell adhesion with clay nanoparticles for countering metastasis. Sci Rep 2019, 9, 5935. [Google Scholar] [CrossRef] [PubMed]
- Jalaleddine, N.; El-Hajjar, L.; Dakik, H.; Shaito, A.; Saliba, J.; Safi, R.; Zibara, K.; El-Sabban, M. Pannexin1 Is Associated with Enhanced Epithelial-To-Mesenchymal Transition in Human Patient Breast Cancer Tissues and in Breast Cancer Cell Lines. Cancers (Basel) 2019, 11. [Google Scholar] [CrossRef]
- Liu, K.; Zhang, X.; Xie, L.; Deng, M.; Chen, H.; Song, J.; Long, J.; Li, X.; Luo, J. Lupeol and its derivatives as anticancer and anti-inflammatory agents: Molecular mechanisms and therapeutic efficacy. Pharmacological research 2021, 164, 105373. [Google Scholar] [CrossRef]
- Sharma, N.; Palia, P.; Chaudhary, A.; Verma, K.; Kumar, I. A review on pharmacological activities of lupeol and its triterpene derivatives. Journal of Drug Delivery and Therapeutics 2020, 10, 325–332. [Google Scholar] [CrossRef]
Figure 1.
Ethanolic extracts of M. orbicularis exhibit antioxidant properties. The percentages of radical scavenging activity of different concentrations (50, 100, 150, 300 and 500 μg/ml) of leaves, fruits, and stems of M. orbicularis ethanolic extracts were measured by DPPH radical scavenging assay. Data are presented as the mean ± SEM of three independent experiments (n=3). ** denotes P<0.01, *** denotes P<0.001, and **** denotes P<0.0001.
Figure 1.
Ethanolic extracts of M. orbicularis exhibit antioxidant properties. The percentages of radical scavenging activity of different concentrations (50, 100, 150, 300 and 500 μg/ml) of leaves, fruits, and stems of M. orbicularis ethanolic extracts were measured by DPPH radical scavenging assay. Data are presented as the mean ± SEM of three independent experiments (n=3). ** denotes P<0.01, *** denotes P<0.001, and **** denotes P<0.0001.
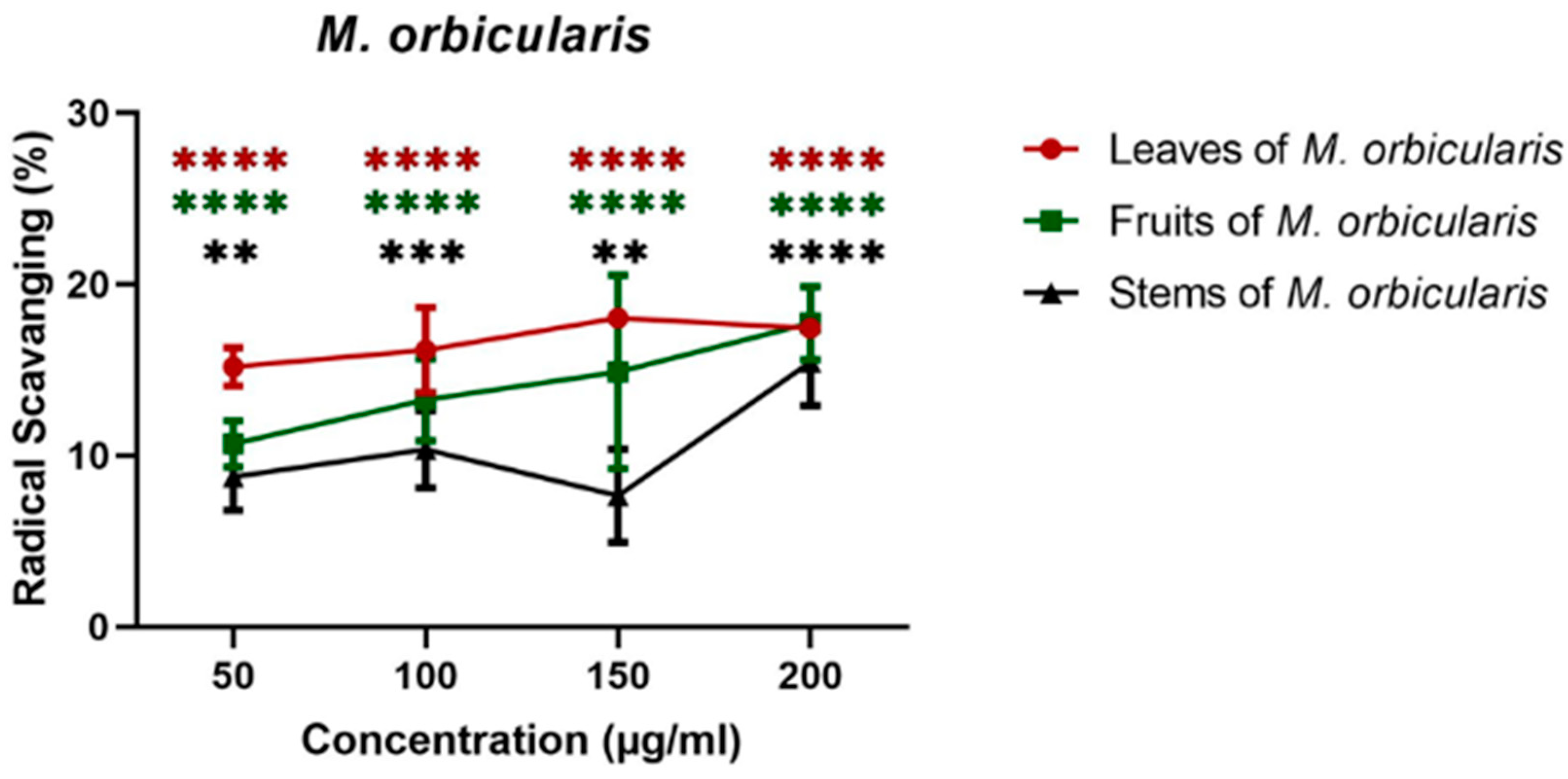
Figure 2.
M. orbicularis plant parts ethanolic extracts reduce the viability of A549 cells. MTT assay was used to measure cell viability of A549 cells treated for 24, 48 and 72 h with increasing concentrations (50, 100, 150, 200 μg/ml) of M. orbicularis plant parts ethanolic extracts. A, B and C show the results of MTT cell cytotoxicity assay of A549 cells treated with the indicated concentrations of ethanolic plant extracts of M. orbicularis leaves, fruits and stems, respectively. D shows the cell viability of neonatal fibroblast cells treated with the indicated concentrations of M. orbicularis fruits ethanolic extract. Viability of treated cells was compared to the control (ctr) vehicle-treated cells. Data are displayed as the mean ± SEM of three independent experiments (n=3). * P< 0.05, **P<0.01, ***P<0.001, ****P<0.0001.
Figure 2.
M. orbicularis plant parts ethanolic extracts reduce the viability of A549 cells. MTT assay was used to measure cell viability of A549 cells treated for 24, 48 and 72 h with increasing concentrations (50, 100, 150, 200 μg/ml) of M. orbicularis plant parts ethanolic extracts. A, B and C show the results of MTT cell cytotoxicity assay of A549 cells treated with the indicated concentrations of ethanolic plant extracts of M. orbicularis leaves, fruits and stems, respectively. D shows the cell viability of neonatal fibroblast cells treated with the indicated concentrations of M. orbicularis fruits ethanolic extract. Viability of treated cells was compared to the control (ctr) vehicle-treated cells. Data are displayed as the mean ± SEM of three independent experiments (n=3). * P< 0.05, **P<0.01, ***P<0.001, ****P<0.0001.
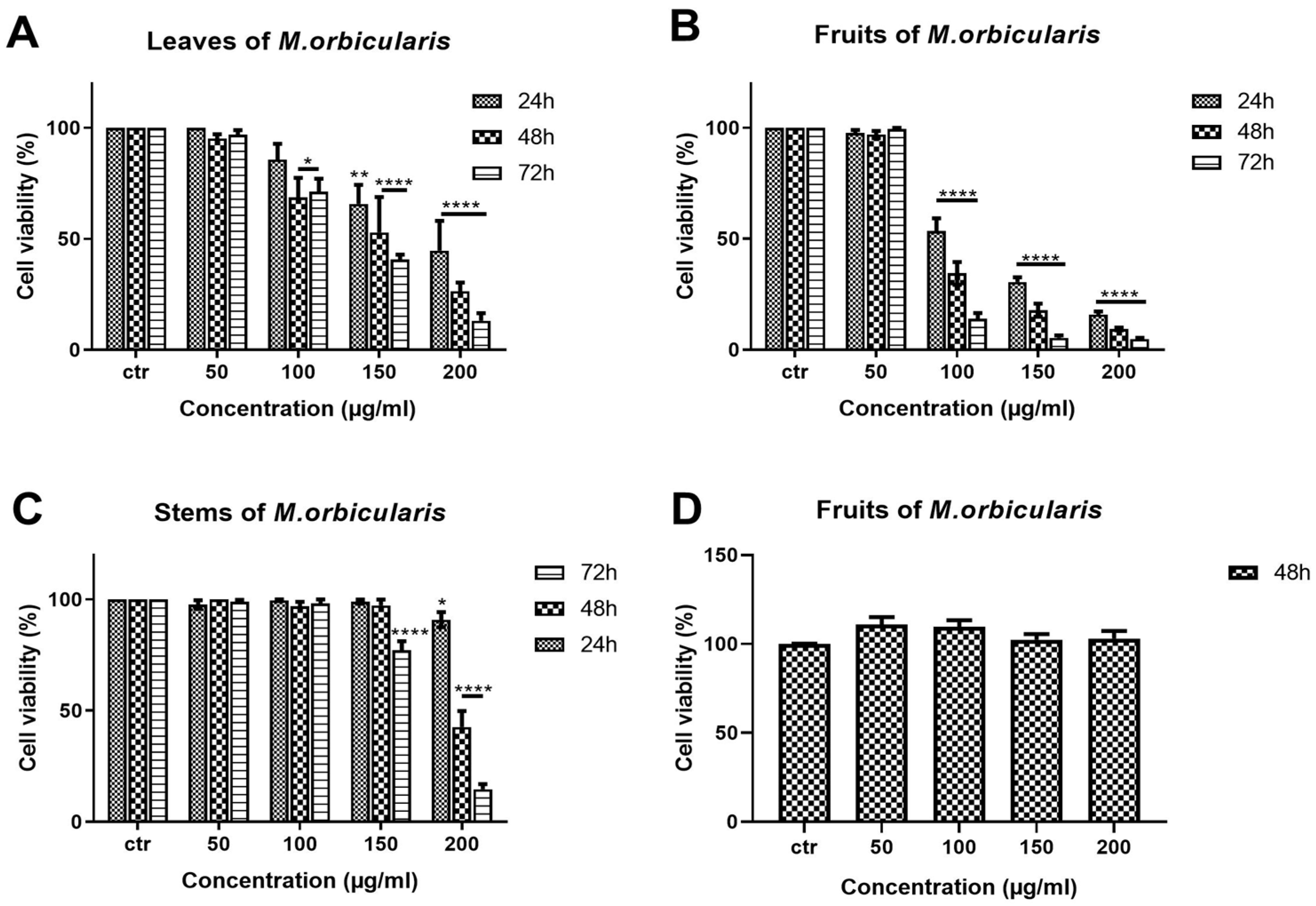
Figure 3.
M. orbicularis fruit ethanolic extracts induce apoptosis of A549 cells. (A) Representative western blot of protein lysates from A549 cells treated 100 and 150 g/ml of M. orbicularis fruit extracts for 48 h. Levels of proteins were normalized to GAPDH protein levels. (B) Quantification of images in (A). Intensity of protein bands were quantified by ImageJ software and normalized to intensity of bands of GAPDH protein. The ratio is expressed in arbitrary units. Data are presented as the mean ± SEM of 3 independent experiments. .
Figure 3.
M. orbicularis fruit ethanolic extracts induce apoptosis of A549 cells. (A) Representative western blot of protein lysates from A549 cells treated 100 and 150 g/ml of M. orbicularis fruit extracts for 48 h. Levels of proteins were normalized to GAPDH protein levels. (B) Quantification of images in (A). Intensity of protein bands were quantified by ImageJ software and normalized to intensity of bands of GAPDH protein. The ratio is expressed in arbitrary units. Data are presented as the mean ± SEM of 3 independent experiments. .

Figure 4.
M. orbicularis fruits ethanolic extracts inhibit migration of A549 cells. (A) A confluent monolayer of A549 cells was wounded by scratching with a pipette tip. The cells were then incubated with the indicated concentrations of M. orbicularis fruits extracts. After 12 h, the scratch was photographed using an inverted microscope and then measured and analyzed. Values represent the fold change in migration compared to the control vehicle-treated cells. (B) Quantification of the micrographs in (A). Bar graphs represent the distance (μm) migrated by the cells after 12 h of treatment. Values represent the average of three independent experiments (n=3) and are represented as mean ± SEM. * P < 0.05.
Figure 4.
M. orbicularis fruits ethanolic extracts inhibit migration of A549 cells. (A) A confluent monolayer of A549 cells was wounded by scratching with a pipette tip. The cells were then incubated with the indicated concentrations of M. orbicularis fruits extracts. After 12 h, the scratch was photographed using an inverted microscope and then measured and analyzed. Values represent the fold change in migration compared to the control vehicle-treated cells. (B) Quantification of the micrographs in (A). Bar graphs represent the distance (μm) migrated by the cells after 12 h of treatment. Values represent the average of three independent experiments (n=3) and are represented as mean ± SEM. * P < 0.05.
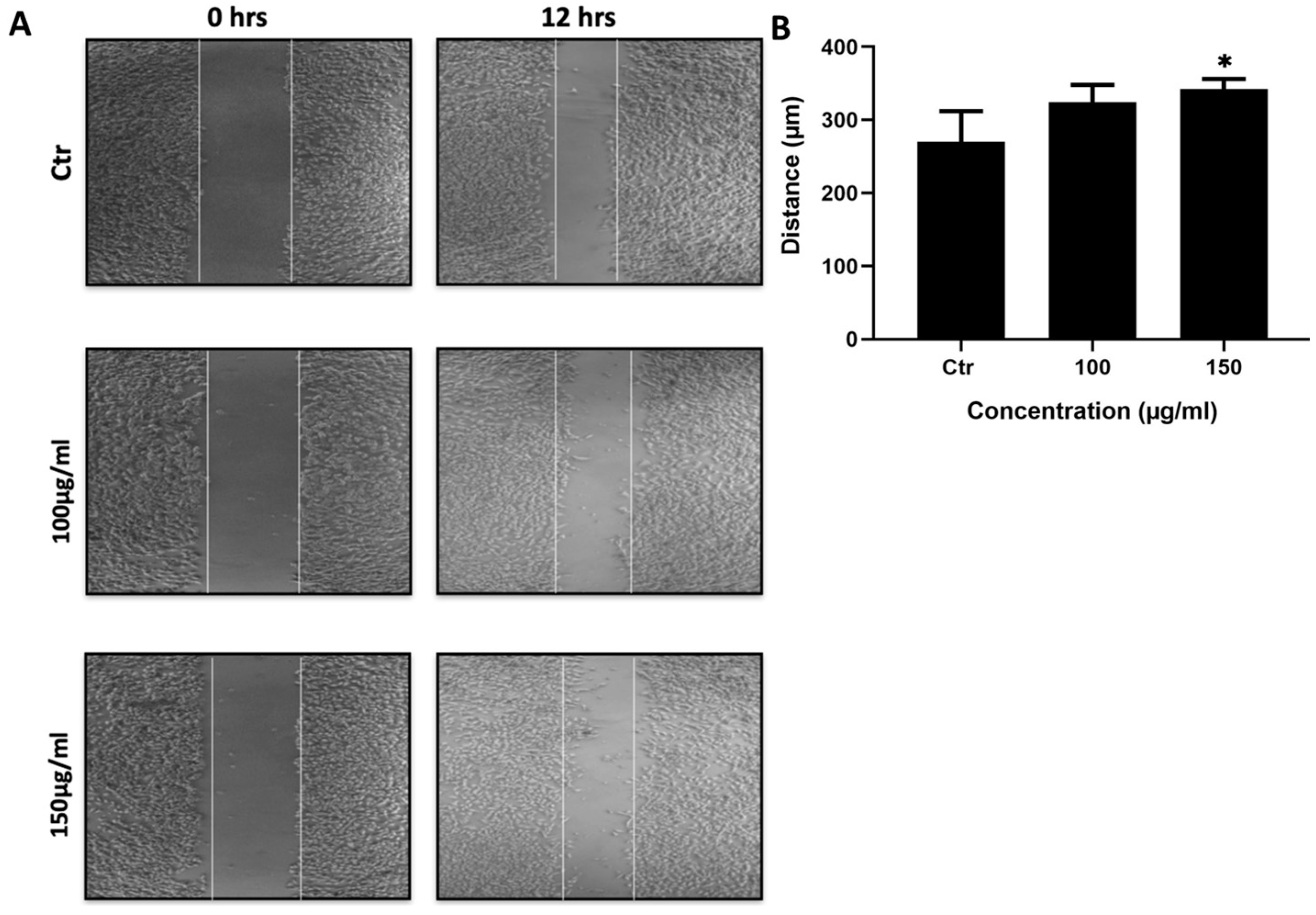
Figure 5.
M. orbicularis fruits ethanolic extracts show potent anti-hemolytic activity. Erythrocytes were pretreated with different (40, 50 and 100 μg/ml) concentrations of ethanolic extracts from the fruits of M. orbicularis. H2O2 was then added to induce hemolysis. Data represent % inhibition of hemolysis compared to control (Ctr) vehicle-treated cells. Data are displayed as the mean ± SEM of three independent experiments; **** denotes P<0.0001.
Figure 5.
M. orbicularis fruits ethanolic extracts show potent anti-hemolytic activity. Erythrocytes were pretreated with different (40, 50 and 100 μg/ml) concentrations of ethanolic extracts from the fruits of M. orbicularis. H2O2 was then added to induce hemolysis. Data represent % inhibition of hemolysis compared to control (Ctr) vehicle-treated cells. Data are displayed as the mean ± SEM of three independent experiments; **** denotes P<0.0001.
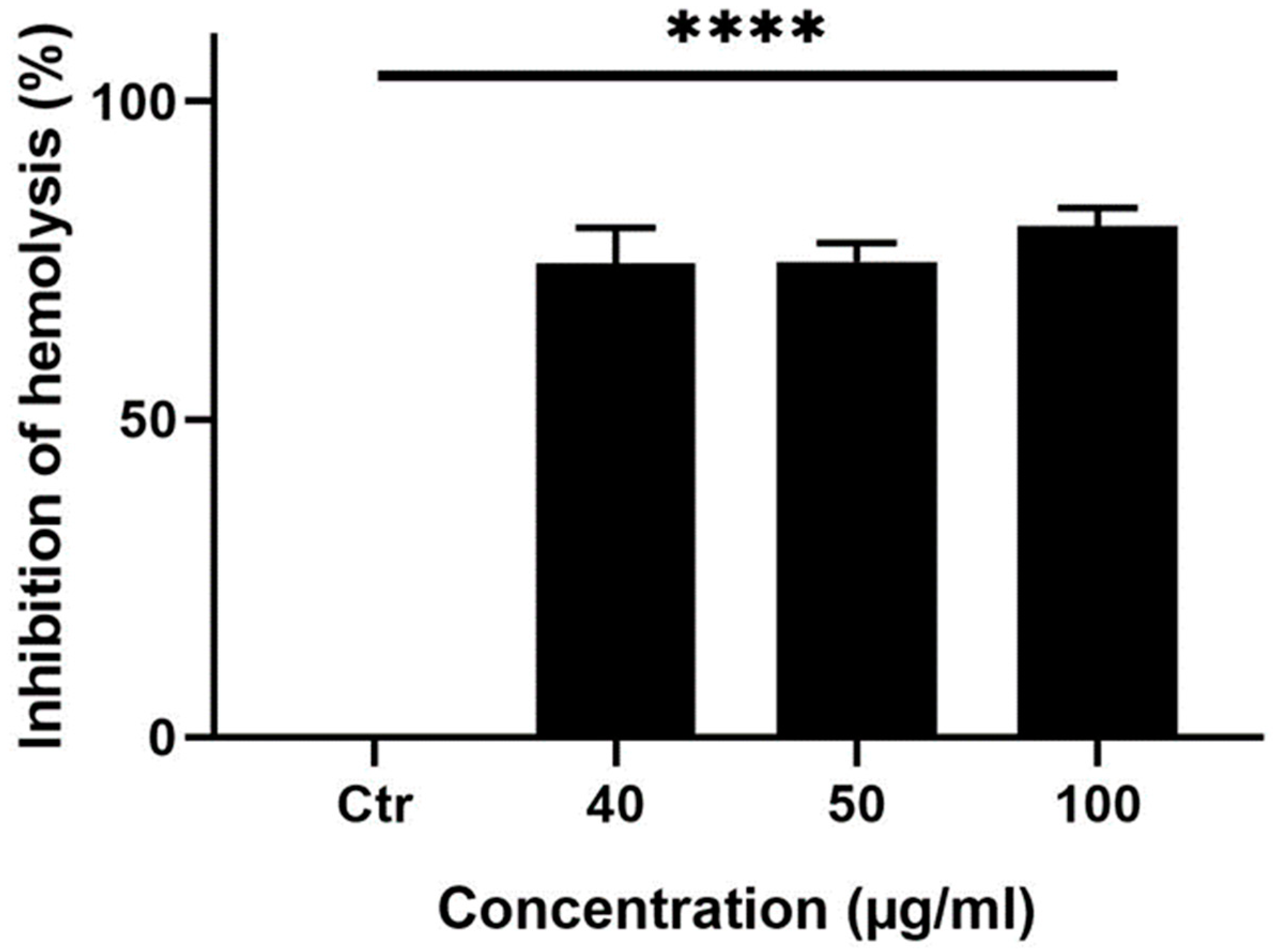
Figure 6.
M. orbicularis enhance cisplatin-induced cytotoxicity in A549 lung cancer cells. (A) Cell viability of A549 cells treated with the indicated concentrations of cisplatin was measured using MTT assay. Data are displayed as % cell viability relative to control (Ctr) cells. (B) A549 cells were treated with fruit extracts of M. orbicularis (100 μg/ml) alone or combined with 5 μg/ml cisplatin (Cis) for 48 h, and then assayed for cell viability using MTT assay. Bar graphs represent % cell viability relative to control (Ctr) cells. Data are presented as the mean ± SEM of three independent experiments (n=3). Significant difference from control: **** p< 0.0001; significant difference from cisplatin treatment alone: #### denotes P< 0.0001; and significant difference from M. orbicularis alone:ⱡ denotes P< 0.05. M. orbicularis F. denotes M. orbicularis fruits extracts.
Figure 6.
M. orbicularis enhance cisplatin-induced cytotoxicity in A549 lung cancer cells. (A) Cell viability of A549 cells treated with the indicated concentrations of cisplatin was measured using MTT assay. Data are displayed as % cell viability relative to control (Ctr) cells. (B) A549 cells were treated with fruit extracts of M. orbicularis (100 μg/ml) alone or combined with 5 μg/ml cisplatin (Cis) for 48 h, and then assayed for cell viability using MTT assay. Bar graphs represent % cell viability relative to control (Ctr) cells. Data are presented as the mean ± SEM of three independent experiments (n=3). Significant difference from control: **** p< 0.0001; significant difference from cisplatin treatment alone: #### denotes P< 0.0001; and significant difference from M. orbicularis alone:ⱡ denotes P< 0.05. M. orbicularis F. denotes M. orbicularis fruits extracts.
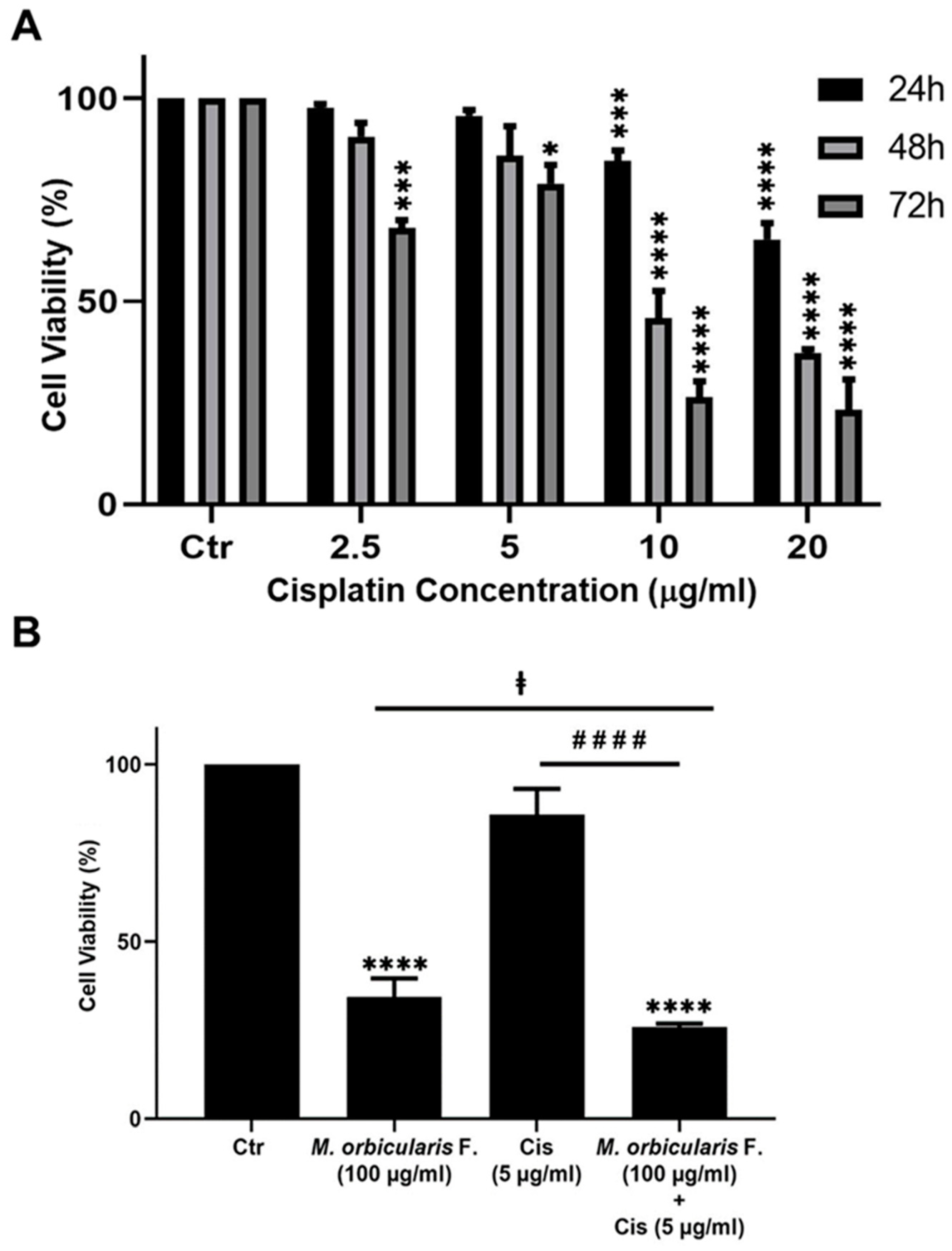
Figure 7.
M. orbicularis augments cisplatin-induced aggregation of A549 lung cancer cells. (A) A549 cells were incubated with M. orbicularis alone (100 or 150 μg/ml) or combined with 5 μg/ml cisplatin (Cis), and then subjected to cell aggregation assay as described in the materials and methods. (B) Quantification of the data in (A). Micrographs of cells were taken after 1h of treatment and the percentage of cell-cell aggregations was measured using the following equation: % aggregation= (1– Nt/Nc) x 100, where Nt is the number of single cells in the control and Nc is the number of single cells in the treated sample. Data represent the mean ± SEM of three independent experiments (n=3). Significant difference from control: **** P< 0.0001; significant difference from Cisplatin treatment alone: # denotes P< 0.005; and significant difference from M. orbicularis extracts alone: ⱡ denotes P< 0.05. M. orbicularis F. denotes M. orbicularis fruits extracts.
Figure 7.
M. orbicularis augments cisplatin-induced aggregation of A549 lung cancer cells. (A) A549 cells were incubated with M. orbicularis alone (100 or 150 μg/ml) or combined with 5 μg/ml cisplatin (Cis), and then subjected to cell aggregation assay as described in the materials and methods. (B) Quantification of the data in (A). Micrographs of cells were taken after 1h of treatment and the percentage of cell-cell aggregations was measured using the following equation: % aggregation= (1– Nt/Nc) x 100, where Nt is the number of single cells in the control and Nc is the number of single cells in the treated sample. Data represent the mean ± SEM of three independent experiments (n=3). Significant difference from control: **** P< 0.0001; significant difference from Cisplatin treatment alone: # denotes P< 0.005; and significant difference from M. orbicularis extracts alone: ⱡ denotes P< 0.05. M. orbicularis F. denotes M. orbicularis fruits extracts.
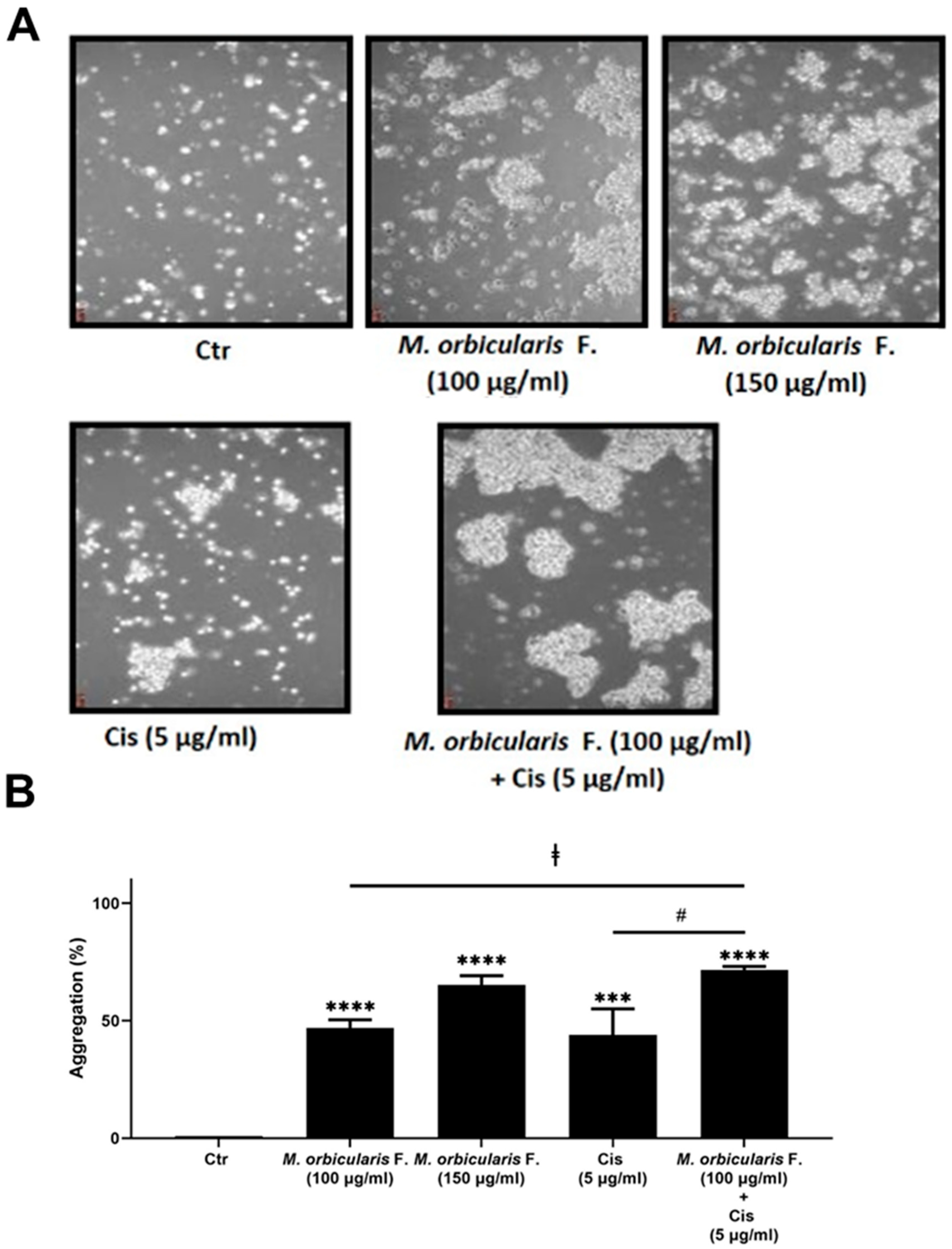
Figure 8.
GC/MS chromatogram of M. orbicularis fruit ethanolic extract displaying the elution profile of phytochemicals listed in Table 2. The detected phytochemicals are arranged according to their retention time, compound name, and molecular weight.
Figure 8.
GC/MS chromatogram of M. orbicularis fruit ethanolic extract displaying the elution profile of phytochemicals listed in Table 2. The detected phytochemicals are arranged according to their retention time, compound name, and molecular weight.
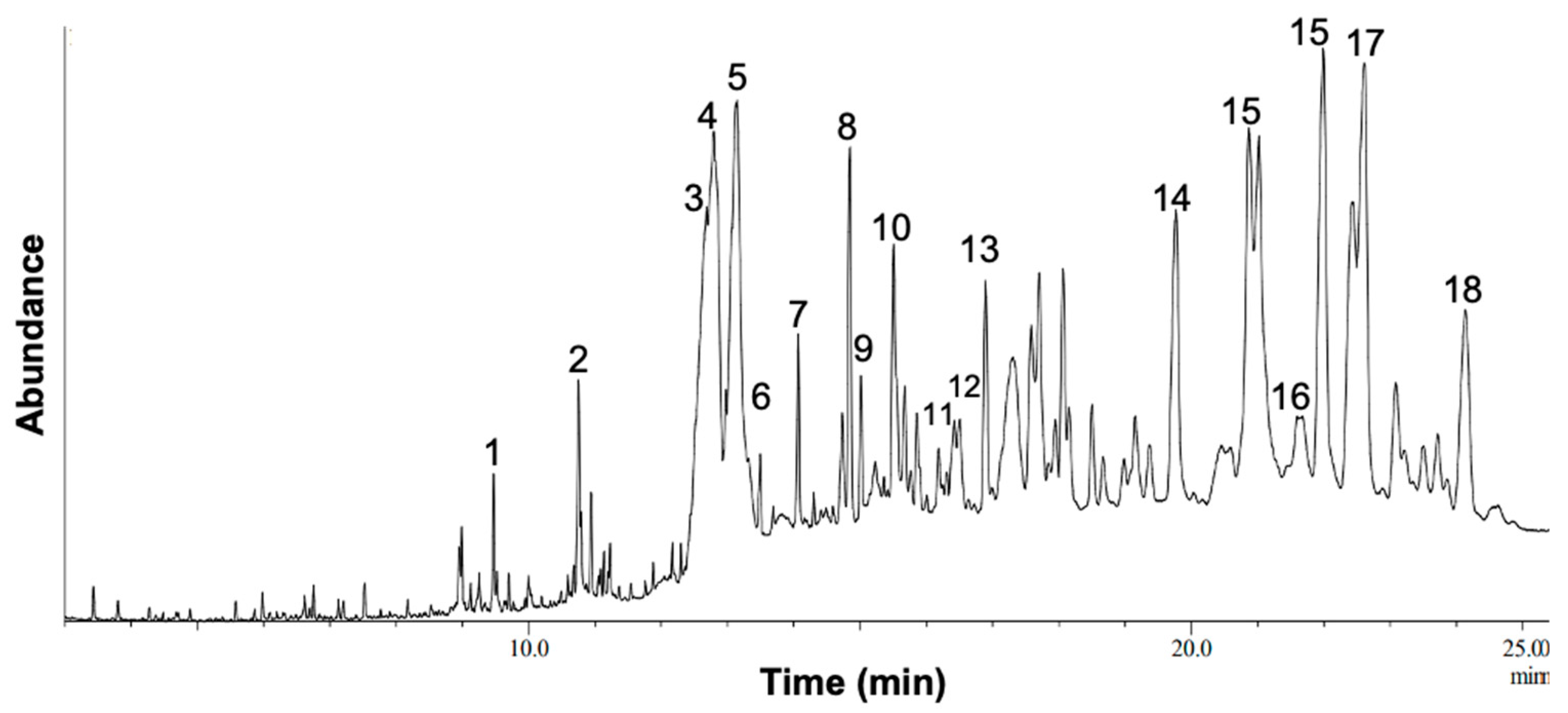

Table 2.
Results of GC-MS analysis of M. orbicularis fruits extracts. The Table presents the name of the identified compounds, their chemical nature and retention time RT(min), and their molecular formula and molecular weight (MW). The listed compounds have highest similarity indices to candidates in the NIST database.
Table 2.
Results of GC-MS analysis of M. orbicularis fruits extracts. The Table presents the name of the identified compounds, their chemical nature and retention time RT(min), and their molecular formula and molecular weight (MW). The listed compounds have highest similarity indices to candidates in the NIST database.
| S.N0 | Name of the Compound | CompoundNature | RT(min) | Molecular Formula |
MW |
|---|---|---|---|---|---|
| 1 | 7-Hexadecenal, (Z)- | Unsaturated aldehyde | 9.530 | C16H30O | 238 |
| 2 | Octadecanamide, N-(2-hydroxyethyl)- | Fatty acid | 10.95 | C20H41NO2 | 327 |
| 3 | Lup-20(29)-en-3-ol, acetate, (3.beta.) | Triterpenes | 12.690 | C32H52O2 | 468 |
| 4 | 6-methoxy-2-methyltetrahydroquinoline | Quinolines | 12.88 | C32H50N6O2 | 550 |
| 5 | 12-Oleanen-3-yl acetate, (3.alpha.) - | Triterpenoids | 13.19 | C32H52O2 | 468 |
| 6 | Trismethoxyresveratrol, trans- | Polyphenol resveratrol | 13.49 | C17H18O3 | 270 |
| 7 | Linoleic acid ethyl ester | Fatty acid | 14.03 | C20H36O2 | 308 |
| 8 | Lup-20(29)-en-3-one | Triterpenoid | 14.86 | C30H48O | 424 |
| 9 | 4-Nitrophenyl laurate | 4-Nitrophenyl esters | 15.15 | C18H27NO4 | 321 |
| 10 | Lupeol, trifluoroacetate | Triterpene alcohol | 15.69 | C32H49F3O2 | 522 |
| 11 | N-Methyl-pseudotomatidine diacetate | Alkaloids | 16.74 | C32H51NO4 | 513 |
| 12 | Pyridine, 1-acetyl-1,2,3,4-tetrahydro-5-(2-piperidinyl)- | Piperidine alkaloid. | 16.91 | C12H20N2O | 208 |
| 13 | Meristic acid | Fatty acid | 17.19 | C14H28O2 | 228 |
| 14 | Oxirane | Rrganic compound | 19.74 | C18H36O | 268 |
| 15 | gamma-Sitosterol | Sterol | 20.91 | C29H50O | 414 |
| 16 | beta-Amyrone | Sesquiterpene | 21.62 | C30H48O | 424 |
| 17 | Lupeol | Triterpene alcohol | 22.89 | C30H50O | 426 |
| 18 | Dihydroniloticin diacetate | Flavonoids | 24.35 | C34H54O5 | 542 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



