
Submitted:
18 September 2023
Posted:
19 September 2023
You are already at the latest version
Abstract
We have identified a new human voltage-gated potassium channel (hKv1.3) blocker, NnK-1, in the jellyfish Nemopilema nomurai, based on its genomic information. The gene sequence encoding NnK-1 contains 5,408 base pairs, with five introns and six exons. The coding sequence of the NnK-1 precursor is 894 nucleotides long and encodes 297 amino acids, containing five presumptive ShK-like peptides. An electrophysiological assay demonstrated that the chemically synthesized fifth peptide, NnK-1, is an effective hKv1.3 blocker. A multiple sequence alignment with cnidarian Shk-like peptides, which have Kv1.3-blocking activity, revealed that four residues (3Asp, 25Lys, 33Lys, and 34Thr) of NnK-1, together with six cysteine residues, are conserved. Therefore, we hypothesize that these four residues are crucial for the binding of the toxins to voltage-gated potassium channels.
Keywords:
toxin
; venom
; jellyfish
; Cnidaria
; genomic information
; electrophysiology
1. Introduction
Cnidaria is a representative group of venomous marine animals. Various kinds of potential toxin proteins and peptides have been reported in cnidarians after transcriptomic and/or proteomic analyses, and have been reviewed [1,2,3,4]. Functional assays of some toxin components deduced with transcriptomic analyses have been undertaken with chemically synthesized materials rather than purified toxin components [5]. Technological advances in genome and transcriptome sequencing, bioinformatics, and the chemical synthesis of peptides and proteins provide unprecedented opportunities to isolate potential pharmaco-medical compounds from venoms. Nemopilema nomurai venom has great potential utility in this regard because its genomic information is available [6], and its venom extracts have shown various therapeutic properties, including antimetastatic [7] and anticancer effects [8]. Therefore, it is highly likely that the jellyfish venom also contains many types of ion channel blockers.
The activation of the voltage-gated potassium channel Kv1.3 in human T and B lymphocytes is related to the development of autoimmune diseases [9,10]. Therefore, Kv1.3 has become a therapeutic target for the treatment of these diseases [11,12,13].
Among the effective potassium channel blockers, an ShK-186 analogue was the first candidate drug to display clinically useful traits [14,15]. The ShK peptide was identified in the sea anemone Stichodactyla helianthus [16]. In brief, it consists of 35 amino acid residues, including six cysteines bridged by three disulfide bonds [17], and blocks voltage-dependent potassium channels. Since its discovery, other toxic ShK-like peptides have been detected and characterized in other sea anemone species [18,19,20,21] and corals [5,22], but not in jellyfish.
Recently, we identified seven ShK-like peptide precursor genes in the genomic information of the jellyfish N. nomurai [6]. In the present study, we describe the structure of an open reading frame (ORF) among these seven precursor genes and its deduced amino acid sequence, which contains five putative ShK-like peptides. The fifth peptide, NnK-1 was chemically synthesized and we investigated whether it has voltage-gated-potassium-channel-blocking activity. For this purpose, we used an internal ribosome entry site (IRES)-containing vector to separately express the human Kv1.3 (hKv1.3) gene and the enhanced green fluorescent protein (EGFP) gene to avoid generating an EGFP–Kv1.3 fusion protein. The current amplitudes at voltage increments of +50 mV were significantly reduced in cells expressing synthetic NnK-1. Our findings contribute to the characterization of toxins with potential voltage-gated-ion-channel-blocking functions, which could be developed for therapeutic applications.
2. Results and Discussion
2.1. Genomic DNA and transcript sequences of NnK-1 precursor gene
The whole gene sequence of the NnK-1 precursor contains 5,408 base pairs with six distinct exons, and the classical 5’ donor (GT) and 3’ acceptor (AG) splice sites are present at each exon/intron boundary (Figure 1).
A transcript encoding an ShK-like peptide was predicted. Figure 2 shows the sequence structure of 894 base pairs, encoding the 297 amino acid residues of an ORF. Five presumptive ShK-like peptides (printed in bold) were detected in the protein. All five peptides have mono- or dibasic amino acid residues at their upstream of N-termini and downstream of C-termini, which are responsible for peptide precursor conversion [23].
One ORF encodes one ShK-like peptide in sea anemone species. However, multiple ShK-like peptides are tandemly arranged in a single coding sequence in the jellyfish. Therefore, a more efficient process for producing cysteine-rich peptide toxins may have evolved at some point during the evolutionary history of the jellyfish, possibly through gene duplication.
2.2. Voltage-gated potassium channel blockade function of NnK-1
We constructed pAAV–CMV–hKv1.3–IRES2–EGFP to allow the activity of Kv1.3 to be measured separately without interference by green fluorescent protein (GFP). After the transfection of pAAV–CMV–hK1.3–IRES2–EGFP, the fluorescence of GFP was highly visible (Figure 3A), and the currents from HEK293 cells expressing AAV–CMV–hK1.3–IRES2–EGFP were clearly recorded when the specified voltage steps were applied, in contrast to untransfected HEK293 cells (Figure 3B and 3C). The current–voltage (IV) recorded from HEK cells expressing hKv1.3 showed a significant difference in the interaction between expressed hKv1.3 and voltage steps (Figure 3C). The hKv1.3 current amplitudes at 50 mV were significantly higher than those of untransfected HEK293 cells (Figure 3D). The hKv1.3 current was significantly reduced by treatment with 0.01, 1, or 100 nM synthetic ShK peptide, which was first identified in the sea anemone S. helianthus [16] (Figure 3E–G, ****P < 0.001), and by treatment with 0.01, 1, or 100 nM Psora4, a pan-Kv1.3 blocker (Figure 3H–3J, *P < 0.05). Importantly, the hKv1.3 current amplitude was also significantly reduced by the application of 0.01, 1, or 100 nM NnK-1 (Figure 3K–3M, *P < 0.05, ***P < 0.001), suggesting that NnK-1 is a candidate inhibitor of hKv1.3
2.3. Structural similarity between sea anemone ShKs and jellyfish NnK-1
The amino acid sequences of the voltage-gated potassium channel blockers identified in sea anemones (ShK, BgK, HmK, AeK, AsKs, and OsPTx2b) and that in the jellyfish (NnK-1) were compared (Figure 4). The aspartic acid (D) in the third position of the peptide is conserved in the ShK analogues from both the sea anemones and jellyfish. The two amino acid residues (KT) located just before the fifth cysteine are also conserved in all sequences. However, the positions of the two potential key binding residues (KY) [24] are conserved in the sea anemone peptides but not in the jellyfish peptide. Therefore, we suggest that these four residues (3Asp, 25Lys, 33Lys, and 34Thr in NnK-1) are essential for the binding of the toxin to voltage-gated potassium channels.
In contrast, the Shk-like peptides PcShK3 and AmAMP1 have been identified in corals. PcShK3 exerts both neuro- and cardioprotective effects in zebrafish [5], and AmAMP1 has antimicrobial activity [22]. Moreover, peptides with a cysteine-rich ShK motif that are expressed in neurons have been identified in Nematostella vectensis, a sea anemone model organism, together with a peptide expressed in nematocysts [25]. The position of aspartic acid (D) is conserved in ShK peptides with both toxic and other functions (Figure 5). However, the KT motif is only conserved in the ShK peptides classified as toxins (Figure 5).
In conclusion, we have successfully identified and characterized a new voltage-gated potassium channel blocker in the jellyfish. We have also shown that using synthetic peptides based on genomic information, rather than purified peptides, is an efficient way to identify biomaterials that can be developed for therapeutic application in the treatment of human diseases.
3. Materials and Methods
3.1. In silico identification of ShK-like peptide genes in N. nomurai
The ShK domains (PF01549.26) of N. nomurai genes were identified with the protein families database (Pfam ver. 34.0). We confirmed the number of cysteine residues in the ShK domains and the presence of basic residues within the 10 amino acids flanking the ShK domains. Finally, we manually selected the ShK-like peptides to be synthesized.
3.2. Peptide synthesis
NnK-1 was synthesized with a standard solid-phase methodology, followed by trifluoroacetic acid–anisole cleavage and high-performance liquid chromatographic purification. Three disulfide bridges formed between Cys1–Cys38, Cys11–Cys31, and Cys20–Cys35. The molecular weight of the synthetic peptide was confirmed with matrix-assisted laser desorption/ionization-time of flight (MALDI-TOF)–mass spectrometry (MS). The peptide was synthesized by Pepmic (Suzhou, China).
3.3. Cloning hKv1.3 cDNA and construction of hKv1.3 expression vector
The primer set used to amplify the hKv1.3 gene is provided in Table 1. Amplification was performed in the Veriti™ 96-Well Fast Thermal Cycler (Applied Biosystems, Waltham, MA, USA), with a thermal cycling program consisting of predenaturation for 3 min at 95 °C, followed by 30 cycles of denaturation for 30 s at 95 °C, annealing for 30 s at 58 °C, and extension for 30 s at 72 °C, with a final extension for 3 min at 72 °C. A Human cDNA Clone Set (OriGene Technologies GmbH, Herford, Germany) was used as the template. MCS (Multi-cloning site) digested from pAAV-MCS vector (Cell Biolabs, San Diego, CA, USA) was ligated into pIRES2-EGFP vector (Clontech Laboratories, Mountain View, CA, USA) to form pAAV–CMV–IRES2–EGFP. The PCR products were ligated into the pAAV–CMV–IRES2–EGFP vector at the NheI and SalI sites to construct pAAV–CMV–hK1.3–IRES2–EGFP.
3.4. Preparation of hKv1.3-vector-transformed HEK293 cells
HEK293 cells were transfected with pAAV–CMV–hK1.3–IRES2–EGFP (2 μg) using Effectene Transfection Reagent (Qiagen, Hilden, Germany) for 12–18 h. The transfectants (1 × 105/ml) were seeded onto 12 mm coverslips coated with 0.01 mg/ml poly-D-lysine for 2–3 h.
3.5. Current recording
HEK293 cells expressing AAV–CMV–hK1.3–IRES2–EGFP were reseeded onto coverslips. The HEK293 transfectants were monitored by their green fluorescence and used as patch cells. Patch pipettes were filled with internal solution (140 mM potassium gluconate, 10 mM HEPES, 0.2 mM ATP, 0.06 mM GTP). External solution [130 mM NaCl, 10 mM HEPES, 3 mM KCl, 1.5 mM D-glucose, 10 mM sucrose, 24 mM CaCl2, 1.5 mM MgCl2·6H2O, osmolarity 320 mmol/kg (pH 7.2)] was used as the basic buffer. Psora4, ShK, or NnK-1 (0, 0.01, 1, or 100 nM) was bath-applied for 2 min to block the currents recorded from HEK293 cells expressing AAV–CMV–hK1.3–IRES2–EGFP in voltage increments ranging from −130 to 50 mV, in increments of 20 mV and 0.6 s. Whole-cell patch recordings from cultured cortical neurons under voltage-clamp conditions (holding potential of –70 mV) were made with the Multiclamp 700B microelectrode amplifier (Automate Scientific Inc., Berkeley, CA, USA) digitized with a Digidata 1322A data acquisition system (Molecular Devices Limited, San Jose, CA, USA). In this study, all electrophysiological data from cultured cells were obtained at a temperature of 23–25 °C, maintained with the CL-100 Temperature Controller (Warner Instruments LLC, Hamden, CT, USA).
Author Contributions
YJK and DHW, electrophysiological analysis; YJ, NL, and HW, data analyses and manuscript preparation; SEL, vector construction; JK and JPC, bioinformatic analyses; DHW and SY, study design, manuscript preparation, and supervision.
Funding
This work was supported by the Marine Biotics Project (202110469) funded by the Ministry of Oceans and Fisheries, the Korea Institute of Toxicology (KIT) Research Program (1711159828) and the R&D Convergence Program of the National Research Council of Science and Technology (CAP-18-02-KRIBB) of the Republic of Korea.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The data that support the findings of this study are available in the figures of the article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Jouiaei, M.; Yanagihara, A.A.; Madio, B.; Nevalainen, T.J.; Alewood, P.F.; Fry, B.G. Ancient Venom Systems: A Review on Cnidaria Toxins. Toxins (Basel) 2015, 7, 2251–2271. [Google Scholar] [CrossRef]
- Brinkman, D.L.; Jia, X.; Potriquet, J.; Kumar, D.; Dash, D.; Kvaskoff, D.; Mulvenna, J. Transcriptome and venom proteome of the box jellyfish Chironex fleckeri. BMC Genomics 2015, 16, 407. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, I.; Hwang, D.H.; Lee, H.; Yoon, W.D.; Chae, J.; Han, C.H.; Yum, S.; Kang, C.; Kim, E. Proteomic Analysis of Novel Components of Nemopilema nomurai Jellyfish Venom: Deciphering the Mode of Action. Toxins (Basel) 2019, 11. [Google Scholar] [CrossRef]
- Wang, C.; Wang, B.; Wang, B.; Wang, Q.; Liu, G.; Wang, T.; He, Q.; Zhang, L. Unique Diversity of Sting-Related Toxins Based on Transcriptomic and Proteomic Analysis of the Jellyfish Cyanea capillata and Nemopilema nomurai (Cnidaria: Scyphozoa). J Proteome Res 2019, 18, 436–448. [Google Scholar] [CrossRef] [PubMed]
- Liao, Q.; Gong, G.; Siu, S.W.I.; Wong, C.T.T.; Yu, H.; Tse, Y.C.; Radis-Baptista, G.; Lee, S.M. A Novel ShK-Like Toxic Peptide from the Transcriptome of the Cnidarian Palythoa caribaeorum Displays Neuroprotection and Cardioprotection in Zebrafish. Toxins (Basel) 2018, 10. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.M.; Weber, J.A.; Lee, N.; Park, S.G.; Cho, Y.S.; Bhak, Y.; Lee, N.; Jeon, Y.; Jeon, S.; Luria, V.; et al. The genome of the giant Nomura's jellyfish sheds light on the early evolution of active predation. BMC Biol 2019, 17, 28. [Google Scholar] [CrossRef]
- Lee, H.; Bae, S.K.; Kim, M.; Pyo, M.J.; Kim, M.; Yang, S.; Won, C.K.; Yoon, W.D.; Han, C.H.; Kang, C.; et al. Anticancer Effect of Nemopilema nomurai Jellyfish Venom on HepG2 Cells and a Tumor Xenograft Animal Model. Evid Based Complement Alternat Med 2017, 2017, 2752716. [Google Scholar] [CrossRef]
- Choudhary, I.; Lee, H.; Pyo, M.J.; Heo, Y.; Chae, J.; Yum, S.S.; Kang, C.; Kim, E. Proteomic Investigation to Identify Anticancer Targets of Nemopilema nomurai Jellyfish Venom in Human Hepatocarcinoma HepG2 Cells. Toxins (Basel) 2018, 10. [Google Scholar] [CrossRef]
- Wulff, H.; Knaus, H.G.; Pennington, M.; Chandy, K.G. K+ channel expression during B cell differentiation: Implications for immunomodulation and autoimmunity. J Immunol 2004, 173, 776–786. [Google Scholar] [CrossRef]
- Beeton, C.; Wulff, H.; Standifer, N.E.; Azam, P.; Mullen, K.M.; Pennington, M.W.; Kolski-Andreaco, A.; Wei, E.; Grino, A.; Counts, D.R.; et al. Kv1.3 channels are a therapeutic target for T cell-mediated autoimmune diseases. Proc Natl Acad Sci U S A 2006, 103, 17414–17419. [Google Scholar] [CrossRef]
- Chhabra, S.; Chang, S.C.; Nguyen, H.M.; Huq, R.; Tanner, M.R.; Londono, L.M.; Estrada, R.; Dhawan, V.; Chauhan, S.; Upadhyay, S.K.; et al. Kv1.3 channel-blocking immunomodulatory peptides from parasitic worms: Implications for autoimmune diseases. FASEB J 2014, 28, 3952–3964. [Google Scholar] [CrossRef] [PubMed]
- Chandy, K.G.; Norton, R.S. Peptide blockers of K(v)1.3 channels in T cells as therapeutics for autoimmune disease. Curr Opin Chem Biol 2017, 38, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Perez-Verdaguer, M.; Capera, J.; Serrano-Novillo, C.; Estadella, I.; Sastre, D.; Felipe, A. The voltage-gated potassium channel Kv1.3 is a promising multitherapeutic target against human pathologies. Expert Opin Ther Targets 2016, 20, 577–591. [Google Scholar] [CrossRef]
- Shen, B.; Cao, Z.; Li, W.; Sabatier, J.M.; Wu, Y. Treating autoimmune disorders with venom-derived peptides. Expert Opin Biol Ther 2017, 17, 1065–1075. [Google Scholar] [CrossRef] [PubMed]
- Tarcha, E.J.; Olsen, C.M.; Probst, P.; Peckham, D.; Munoz-Elias, E.J.; Kruger, J.G.; Iadonato, S.P. Safety and pharmacodynamics of dalazatide, a Kv1.3 channel inhibitor, in the treatment of plaque psoriasis: A randomized phase 1b trial. PLoS ONE 2017, 12, e0180762. [Google Scholar] [CrossRef]
- Castaneda, O.; Sotolongo, V.; Amor, A.M.; Stocklin, R.; Anderson, A.J.; Harvey, A.L.; Engstrom, A.; Wernstedt, C.; Karlsson, E. Characterization of a potassium channel toxin from the Caribbean Sea anemone Stichodactyla helianthus. Toxicon 1995, 33, 603–613. [Google Scholar] [CrossRef]
- Tudor, J.E.; Pallaghy, P.K.; Pennington, M.W.; Norton, R.S. Solution structure of ShK toxin, a novel potassium channel inhibitor from a sea anemone. Nat Struct Biol 1996, 3, 317–320. [Google Scholar] [CrossRef]
- Gendeh, G.S.; Young, L.C.; de Medeiros, C.L.; Jeyaseelan, K.; Harvey, A.L.; Chung, M.C. A new potassium channel toxin from the sea anemone Heteractis magnifica: Isolation, cDNA cloning, and functional expression. Biochemistry 1997, 36, 11461–11471. [Google Scholar] [CrossRef]
- Cotton, J.; Crest, M.; Bouet, F.; Alessandri, N.; Gola, M.; Forest, E.; Karlsson, E.; Castaneda, O.; Harvey, A.L.; Vita, C.; et al. A potassium-channel toxin from the sea anemone Bunodosoma granulifera, an inhibitor for Kv1 channels. Revision of the amino acid sequence, disulfide-bridge assignment, chemical synthesis, and biological activity. Eur J Biochem 1997, 244, 192–202. [Google Scholar] [CrossRef]
- Minagawa, S.; Ishida, M.; Nagashima, Y.; Shiomi, K. Primary structure of a potassium channel toxin from the sea anemone Actinia equina. FEBS Lett 1998, 427, 149–151. [Google Scholar] [CrossRef]
- Krishnarjuna, B.; Villegas-Moreno, J.; Mitchell, M.L.; Csoti, A.; Peigneur, S.; Amero, C.; Pennington, M.W.; Tytgat, J.; Panyi, G.; Norton, R.S. Synthesis, folding, structure and activity of a predicted peptide from the sea anemone Oulactis sp. with an ShKT fold. Toxicon 2018, 150, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Mason, B.; Cooke, I.; Moya, A.; Augustin, R.; Lin, M.F.; Satoh, N.; Bosch, T.C.G.; Bourne, D.G.; Hayward, D.C.; Andrade, N.; et al. AmAMP1 from Acropora millepora and damicornin define a family of coral-specific antimicrobial peptides related to the Shk toxins of sea anemones. Dev Comp Immunol 2021, 114, 103866. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, T.W. Cellular peptide processing after a single arginyl residue. Studies on the common precursor for pancreatic polypeptide and pancreatic icosapeptide. J Biol Chem 1987, 262, 5093–5099. [Google Scholar] [CrossRef] [PubMed]
- Finol-Urdaneta, R.K.; Belovanovic, A.; Micic-Vicovac, M.; Kinsella, G.K.; McArthur, J.R.; Al-Sabi, A. Marine Toxins Targeting Kv1 Channels: Pharmacological Tools and Therapeutic Scaffolds. Mar Drugs 2020, 18. [Google Scholar] [CrossRef]
- Sachkova, M.Y.; Landau, M.; Surm, J.M.; Macrander, J.; Singer, S.A.; Reitzel, A.M.; Moran, Y. Toxin-like neuropeptides in the sea anemone Nematostella unravel recruitment from the nervous system to venom. Proc Natl Acad Sci U S A 2020, 117, 27481–27492. [Google Scholar] [CrossRef]
Figure 1.
Organization of the Nemopilema nomurai NnK-1 precursor gene, which contains six distinct exons (green).
Figure 1.
Organization of the Nemopilema nomurai NnK-1 precursor gene, which contains six distinct exons (green).
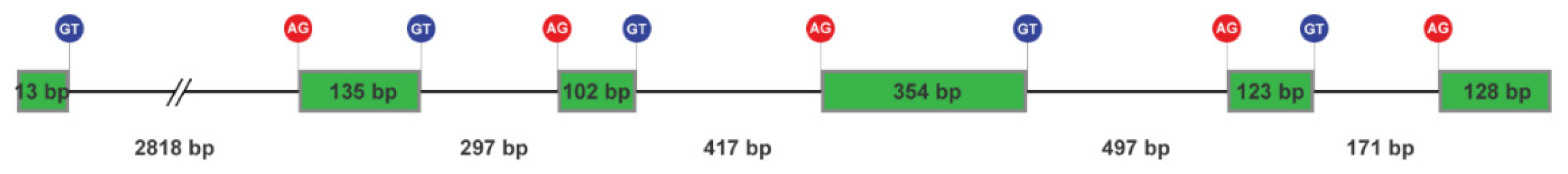
Figure 2.
Transcript sequence of the NnK-1-encoding gene and deduced amino acid sequence. The amino acid sequence of five ShK-like peptides is printed in bold. NnK-1 is printed in red.
Figure 2.
Transcript sequence of the NnK-1-encoding gene and deduced amino acid sequence. The amino acid sequence of five ShK-like peptides is printed in bold. NnK-1 is printed in red.
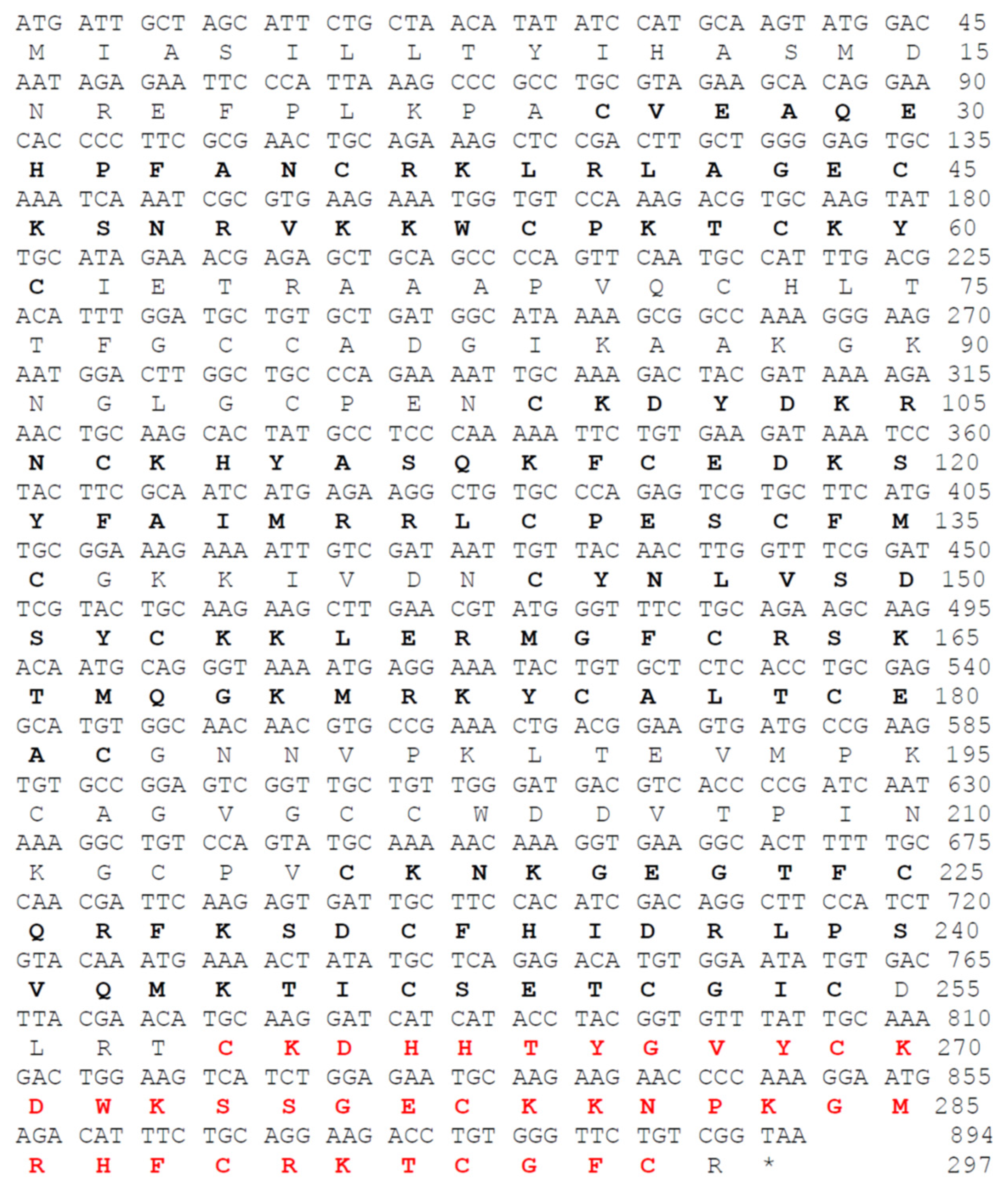
Figure 3.
Inhibitory effect of NnK-1 on the activation of hKv1.3 channels. (A) Bright-field images of glass pipettes used for patch clamp of HEK293 cells (upper panels) and fluorescence microscopic images of cells (lower panel) without (left panel) and with GFP (right panel), showing the expression of AAV–CMV–hKv1.3–IRES2–EGFP. Scale bar, 50 µm. (B) Current amplitudes ranged from −130 to 50 mV, in increments of 20 mV (10 steps) without (left) and with hKv1.3 (right). Inset indicates 10 voltage steps. (C) Representative current–voltage (IV) curves without (○) and with hKv1.3 (●). Two-way ANOVA, interaction between voltage and group F(9, 150) = 4.01, P < 0.0001; voltage F(9, 150) = 5.790, P < 0.0001; group F(1, 150) = 22.08, P < 0.0001. (D) Summary bar graphs of current amplitudes without and with hKv.1.3 expression at 50 mV. Unpaired t test, *P < 0.05. (E) hKv1.3-mediated currents at 50 mV in the presence of the indicated concentrations of ShK (0, 0.01, 1, 100 nM). (F) Representative IV curves in the presence of ShK (0, 0.01, 1, 100 nM). Two-way ANOVA, interaction between voltage and group F(27, 120) = 1.13, P = 0.32; voltage F(9, 120) = 33.32, P < 0.0001; group F(3, 120) = 5.62, P < 0.01. (G) Summary bar graphs of current amplitudes at 50 mV in the presence of ShK (0, 0.01, 1, 100 nM). One-way ANOVA, F(3, 12) = 50.36, P < 0.0001, followed by Dunnett’s post hoc test, ****P < 0.0001, 0 vs 0.01 nM, 0 vs 1 nM, 0 vs 100 nM. (H) hKv1.3-mediated currents at 50 mV in the presence of the indicated concentrations of Psora4 (0, 0.01, 1, 100 nM). (I) Representative IV curves in the presence of Psora4 (0, 0.01, 1, 100 nM). Two-way ANOVA, interaction between voltage and group F(27, 120) = 2.71, P < 0.001; voltage F(9, 120) = 61.44, P < 0.0001; group F(3, 120) = 17.49, P < 0.0001. (J) Summary bar graphs of current amplitudes at 50 mV in the presence of Psora4 (0, 0.01, 1, 100 nM). One-way ANOVA, F(3, 12) = 5.15, P < 0.05, followed by Dunnett’s post hoc test, *P < 0.05, 0 vs 1 nM, 0 vs 100 nM. (K) Representative traces in the presence of the indicated concentrations of NnK-1 (0, 0.01, 1, 100 nM). (L) Representative IV curves in the presence of NnK-1 (0, 0.01, 1, 100 nM). Two-way ANOVA, the interaction between voltage and group F(27, 160) = 3.04, P < 0.0001; voltage F(9, 160) = 79.08, P < 0.0001; group F(3, 160) = 13.27, P < 0.0001. (M) Summary bar graphs of current amplitudes at 50 mV in the presence of NnK-1 (0, 0.01, 1, 100 nM). One-way ANOVA, F(3, 16) = 12.8, P < 0.001, followed by Dunnett’s post hoc test, *P < 0.05, 0 vs 0.01 nM, ***P < 0.001, 0 vs 1 nM, 0 vs 100 nM. Data are means ± SEM.
Figure 3.
Inhibitory effect of NnK-1 on the activation of hKv1.3 channels. (A) Bright-field images of glass pipettes used for patch clamp of HEK293 cells (upper panels) and fluorescence microscopic images of cells (lower panel) without (left panel) and with GFP (right panel), showing the expression of AAV–CMV–hKv1.3–IRES2–EGFP. Scale bar, 50 µm. (B) Current amplitudes ranged from −130 to 50 mV, in increments of 20 mV (10 steps) without (left) and with hKv1.3 (right). Inset indicates 10 voltage steps. (C) Representative current–voltage (IV) curves without (○) and with hKv1.3 (●). Two-way ANOVA, interaction between voltage and group F(9, 150) = 4.01, P < 0.0001; voltage F(9, 150) = 5.790, P < 0.0001; group F(1, 150) = 22.08, P < 0.0001. (D) Summary bar graphs of current amplitudes without and with hKv.1.3 expression at 50 mV. Unpaired t test, *P < 0.05. (E) hKv1.3-mediated currents at 50 mV in the presence of the indicated concentrations of ShK (0, 0.01, 1, 100 nM). (F) Representative IV curves in the presence of ShK (0, 0.01, 1, 100 nM). Two-way ANOVA, interaction between voltage and group F(27, 120) = 1.13, P = 0.32; voltage F(9, 120) = 33.32, P < 0.0001; group F(3, 120) = 5.62, P < 0.01. (G) Summary bar graphs of current amplitudes at 50 mV in the presence of ShK (0, 0.01, 1, 100 nM). One-way ANOVA, F(3, 12) = 50.36, P < 0.0001, followed by Dunnett’s post hoc test, ****P < 0.0001, 0 vs 0.01 nM, 0 vs 1 nM, 0 vs 100 nM. (H) hKv1.3-mediated currents at 50 mV in the presence of the indicated concentrations of Psora4 (0, 0.01, 1, 100 nM). (I) Representative IV curves in the presence of Psora4 (0, 0.01, 1, 100 nM). Two-way ANOVA, interaction between voltage and group F(27, 120) = 2.71, P < 0.001; voltage F(9, 120) = 61.44, P < 0.0001; group F(3, 120) = 17.49, P < 0.0001. (J) Summary bar graphs of current amplitudes at 50 mV in the presence of Psora4 (0, 0.01, 1, 100 nM). One-way ANOVA, F(3, 12) = 5.15, P < 0.05, followed by Dunnett’s post hoc test, *P < 0.05, 0 vs 1 nM, 0 vs 100 nM. (K) Representative traces in the presence of the indicated concentrations of NnK-1 (0, 0.01, 1, 100 nM). (L) Representative IV curves in the presence of NnK-1 (0, 0.01, 1, 100 nM). Two-way ANOVA, the interaction between voltage and group F(27, 160) = 3.04, P < 0.0001; voltage F(9, 160) = 79.08, P < 0.0001; group F(3, 160) = 13.27, P < 0.0001. (M) Summary bar graphs of current amplitudes at 50 mV in the presence of NnK-1 (0, 0.01, 1, 100 nM). One-way ANOVA, F(3, 16) = 12.8, P < 0.001, followed by Dunnett’s post hoc test, *P < 0.05, 0 vs 0.01 nM, ***P < 0.001, 0 vs 1 nM, 0 vs 100 nM. Data are means ± SEM.
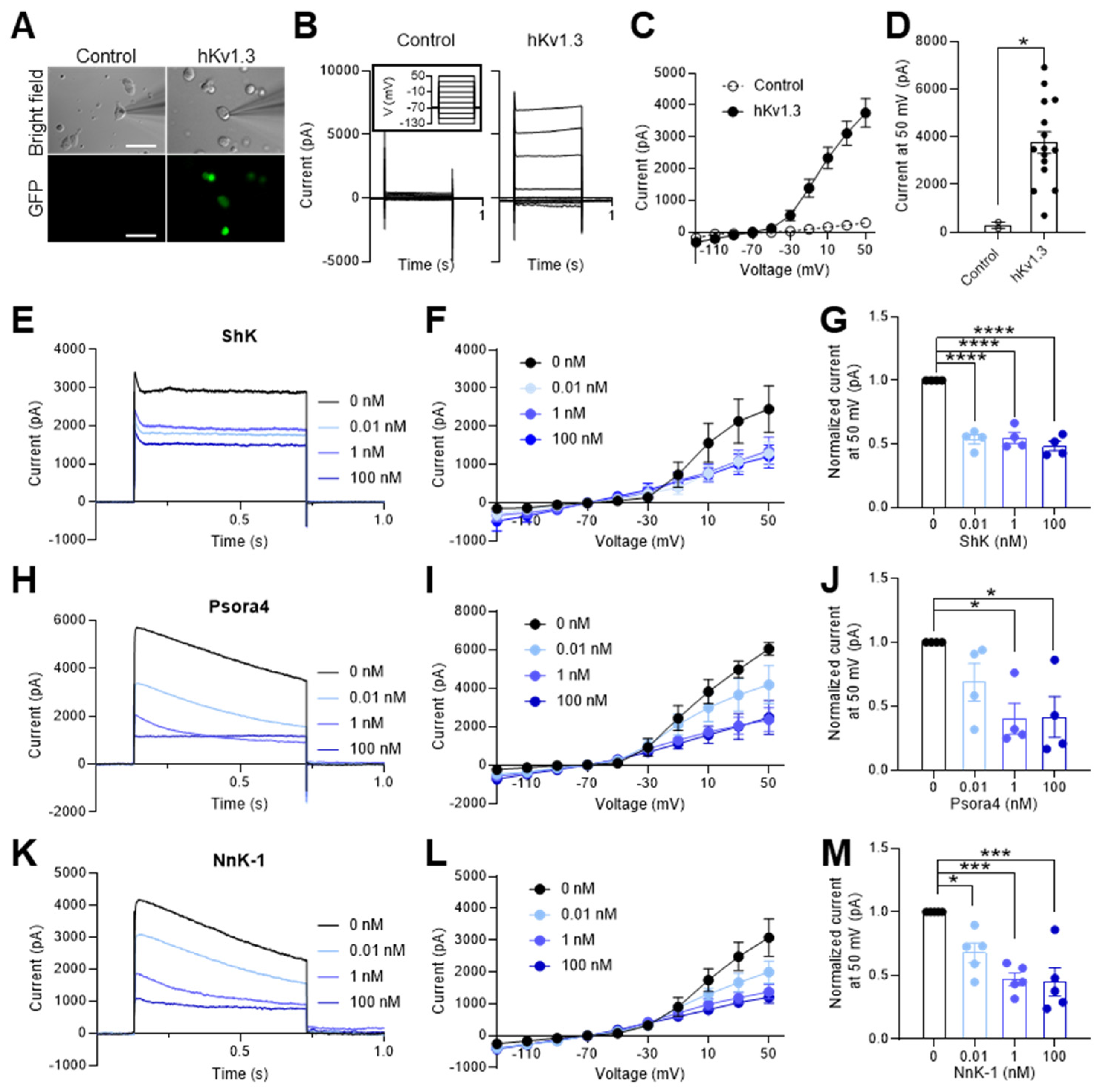
Figure 4.
Schemes follow the same formatting. Multiple sequence alignment of six sea anemone peptides and one jellyfish peptide, all of which show Kv1.3-blocking activity. ShK (from Stichodactyla helianthus), BgK (from Bunodosoma granulifera), HmK (from Heteractis magnifica) Aek (from Actinia equina), AsKs (from Anemonia sulcata), OsPtx2b (from Oulactis sp.), AEtxK1 (from Anemonia erythraea), and NnK-1 (from Nemopilema nomurai).
Figure 4.
Schemes follow the same formatting. Multiple sequence alignment of six sea anemone peptides and one jellyfish peptide, all of which show Kv1.3-blocking activity. ShK (from Stichodactyla helianthus), BgK (from Bunodosoma granulifera), HmK (from Heteractis magnifica) Aek (from Actinia equina), AsKs (from Anemonia sulcata), OsPtx2b (from Oulactis sp.), AEtxK1 (from Anemonia erythraea), and NnK-1 (from Nemopilema nomurai).
Figure 5.
Multiple sequence alignment of six ShK peptides. ShK, NnK-1 and ShK-like 1 peptide (from Nematostella vectecsis) are known to be toxins. ShK-like 2 (from N. vectecsis) is expressed in neurons, PcShK3 (from Palythoa cariboeorum) has neuro- and cardioprotective functions, and AmAMP1 (from Acropora millepora) has antimicrobial activity.
Figure 5.
Multiple sequence alignment of six ShK peptides. ShK, NnK-1 and ShK-like 1 peptide (from Nematostella vectecsis) are known to be toxins. ShK-like 2 (from N. vectecsis) is expressed in neurons, PcShK3 (from Palythoa cariboeorum) has neuro- and cardioprotective functions, and AmAMP1 (from Acropora millepora) has antimicrobial activity.
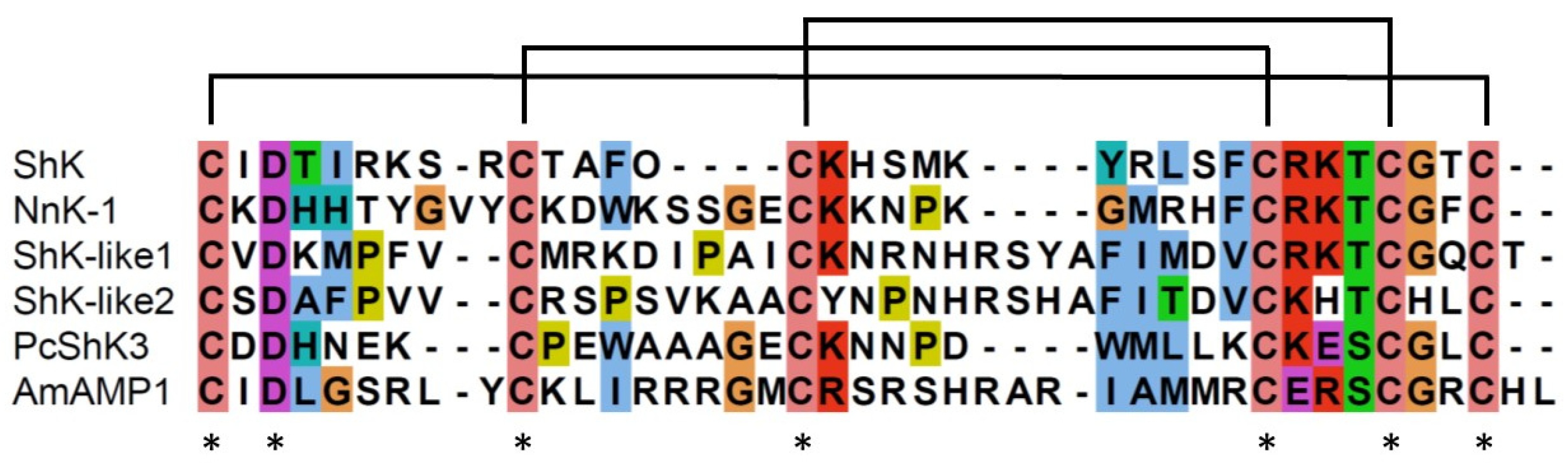

Table 1.
Primer set used to amplify the hKV1.3 gene with polymerase chain reaction.
| Primer | Sequence |
|---|---|
| Sal1-Kv1.3-R | 5’-TTTGTCGACCTAAACATCGGTGAATATCTTTT-3’ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



