
Submitted:
14 September 2023
Posted:
20 September 2023
You are already at the latest version
Abstract
The cultivation of Hevea brasiliensis, the primary commercial source of natural rubber, is strongly impacted by South American Leaf Blight (SALB) disease, caused by the fungus Pseudocercospora ulei. Various management strategies have been implemented, including the selection of resistant genotypes and the identification of escape zones. This study evaluated the growth, early yield, and resistance to SALB of nine Colombian elite genotypes from the ECC-100 series and IAN 873 clone (control) in a large-scale clone trial in a low SALB pressure zone in the Colombian Amazon, 2017 - 2020. Favorable early performance was evident, although there was a significant increase in the severity and sporulation of P. ulei over time, especially in the ECC 35, ECC 60, and IAN 873 genotypes. However, these scores represented a low susceptibility. Genotypes with higher resistance to SALB demonstrated greater growth and early yield compared to higher susceptible genotypes. The ECC 64, ECC 73, ECC 90, ECC 25, and ECC 29 genotypes were more desirable in low SALB pressure zones due to their higher resistance and early performance. It is important to highlight that this research contributes to the selection of new SALB-resistant Colombian genotypes of H. brasiliensis in the growth stage. Therefore, it is necessary to evaluate their productivity in the mature stage and long-term resistance to SALB, before recommending and promoting their commercial adoption in the Colombian Amazon.
Keywords:
Hevea brasiliensis
; Pseudocercospora ulei
; South American Leaf Blight
; Genetic resistance
; early performance
; clone selection
1. Introduction
Hevea brasilienses (Willd. ex A.Juss.) Müll.Arg., is the primary commercial source of natural rubber (NR) and supports the livelihoods of approximately 20 million people worldwide [1]. This resource is essential as a raw material in the manufacturing of over 50,000 products across various industrial sectors, including healthcare, construction, and automotive. In the latter sector, its usage is particularly prominent in the production of high-strength tires and rubber products [2,3].
Between 1961 and 2020, there has been a remarkable increase in the global average yield of rubber cultivation, rising from 0.5 to 1.2 tons per hectare and annual production has surged from 2.1 to 14.8 million tons [4]. This increase has been achieved through the implementation of advanced agronomic practices, the careful selection of highly productive and disease-resistant genotypes, the identification of suitable cultivation areas and the expansion of agricultural land dedicated to rubber tree cultivation [5,6]. The scarcity of other commercial sources of NR offering comparable quality and production capacity has justified research into various aspects of H. brasiliensis cultivation [7,8]. In contrast, rubber tree cultivation faces various phytosanitary limitations that, if not addressed in a timely manner, can lead to significant economic losses. In South America, the primary disease affecting H. brasiliensis is South American Leaf Blight (SALB), caused by the fungus Pseudocercospora ulei (Henn.) [9]. This disease results in repeated defoliations, reduced latex production, and in severe cases, it can lead to the death of the plant [10], thereby limiting production in the Amazon region [11].
Chemical management of SALB is possible, however, due to the height of the trees (20-30 m) and the required treatment frequency, it becomes economically unfeasible and raises significant ecological and public health concerns [12]. For the management of SALB, the primary approaches implemented have been the selection of fungus-resistant clones [13] and the identification of escape zones (areas where rubber grows with lower fungal pressure) [1]. These strategies have been applied in Colombia and the Amazon region, where the production of natural rubber has been severely impacted by SALB incidence [14,15,16].
According to FAO data [4], the yield of rubber tree plantations in Colombia reaches 1.6 tons per hectare surpassing the global average of 1.2 tons per hectare but falling below the averages of plantations in Mexico, Guatemala and India, where average yields reach 4.0, 2.8, and 2.1 tons per hectare, respectively. In the Amazon, despite the introduction of genotypes with good resistance and yield characteristics, the cultivation continues to be heavily impacted by the disease in the Amazon region, which diminishes productivity [17,18]. Therefore, it is necessary to select new high-yield clones that are higher resistant to SALB.
With the establishment of the rubber tree clone trials network in Latin America [1], clones/genotypes that exhibit partial or complete resistance to SALB have been identified [19]. Previous researches conducted in the Colombian Amazon has identified superior genotypes in terms of performance, quality and adaptability to climatic variations [16,20]. For the Elite Caquetá Colombia (ECC) series, Sterling et al. [15] observed favorable early growth in some genotypes in non-SALB escape zones. However, those genotypes with high resistance showed lower growth rates compared to the more susceptible genotypes. Therefore, further research is necessary for the final selection and commercial adoption of resistant genotypes with desirable productivity traits under high and low SALB pressure.
In Colombia, there has been an effort to expand the genetic base of rubber tree by assessing the growth, physiology, latex production, and response to SALB of introduced clones and genotypes from the ECC-100 series in small-scale and large-scale clone trials, under non-SALB escape conditions [15]. However, in escape zones or areas with low disease pressure, it is expected that the environmental response of these genotypes in agronomic and phytosanitary terms will be more favorable. Therefore, the objective of this study was to evaluate the growth, early yield and SALB resistance of nine Colombian elite genotypes from the ECC-100 series, and IAN 873 (control) under low disease pressure in the Colombian Amazon region.
2. Results
2.1. Climate
Between July 2017 and June 2020, the study area recorded an average monthly precipitation of 209.5 mm and an annual average precipitation of 2514.6 mm. The average annual temperature was 25.1°C. The maximum temperature ranged from 28.5°C in June and July 2019 to 34.1°C in January 2020, while the minimum temperature ranged from 21.84°C in December 2019 to 23.6°C in February 2020. The average temperature was of 8.7°C, with an average maximum temperature of 31.6°C and an average minimum temperature of 22.7°C. The lowest relative humidity was recorded in January 2020 with a value of 69.3%, while the highest occurred in June 2019 with a value of 88.8%, and the annual average was of 81.5%. The dew point ranged from 19.4°C in January 2020 to 22.9°C in May 2018 (Figure 1).
2.2. Effects of the Fixed Factors
The AIC criterion was minimized in the UNH model for all measured variables, except for the TR, for which the ARH1 model performed better in terms of AIC criteria. For BIC, the best model was UNH for the CTpt, AT1 and TR variables. The ARH1 model was superior for the BIC criterion in AT2, and CSH model was the best for the BIC ing ST (Table 1). Based on the above, the unstructured model with heterogeneous variances (UNH) was the best model. Consequently, significant differences were observed between the genotypes, between the years and in the genotype*year interaction (p < 0.05) (Table 2).
2.3. Growth and Early Yield
The annual tree growth data are shown in Table 3. Significant differences were found between the 10 genotypes (p < 0.05). The higher mean values of CTpt were observed in the ECC 64, ECC 25, and ECC 83 genotypes, where the ECC 64 genotype had the highest average of CTpt over the three years of evaluation (p < 0.05). The ECC 25 genotype showed higher means than the other genotypes in years 2 and 3 (p < 0.05), where the ECC 64 and ECC 25 genotypes had the greater vigor (Table 3).
The CTo ranged from 25.00 cm (ECC 35) to 27.92 cm (ECC 64), where the ECC 64 genotype was significantly higher than the other genotypes (p < 0.05). The ECC 64, ECC 90, and ECC 25 genotypes had higher values of CTo, with 27.92 cm, 26.94 cm and 26.86 cm, respectively (Table 4).
In the early tapping phase, the ECC 25, ECC 64, and ECC 83 genotypes had higher means of dry rubber yield (DRY), with 5.57, 3.76 and 3.30 g.tree-1.tap-1, respectively. These values were higher than IAN 873 (p < 0.05). On the other hand, the ECC 29, ECC 35, and ECC 66 genotypes showed lower DRY means, with values of 0.74, 0.42 and 1.09 g.tree−1.tap−1, respectively, although these did not differ significantly from IAN 873, which had a mean of 1.88 g.tree−1.tap−1 (Table 4).
The dry rubber content (DRC) was significantly higher in the IAN 873, ECC 25, ECC 73, ECC 29, ECC 64 and ECC 66 genotypes, with means of 32.83, 32.45, 32.42, 31.44, 31.38 and 20.75%, respectively. The ECC 35 and ECC 90 genotypes had lower DRC values (Table 4).
2.4. Resistance to SALB
According to Figure 2, there was a low SALB incidence in the first year, but in years 2 and 3, there was a significant increase in susceptibility to P. ulei. In young leaves, mean scores of AT1 and TR were lower than 1. Some genotypes had mean scores > 1 and > 3 for AT2 and ST, respectively. In terms of susceptibility, it was evident that the ECC 35 genotype had the highest SALB mean scores, while ECC 73 had the lowest scores.
For AT1, the ECC 64, ECC 29 and ECC 90 genotypes had lower attack intensity, with AT1 < 1. Despite the significant increase in attack for the third year, the scores were lower than 1 (Figure 2a). The TR means were statistically different between genotypes, with a significant increase in the second and third years. The ECC 64, ECC 90 and ECC 29 genotypes were the less susceptible, with TR = 0. On the other hand, the ECC 60, ECC 83 and ECC 35 genotypes showed higher sporulation intensities, especially in the third year. However, TR means < 1 were observed in all genotypes over the years (Figure 2b).
In mature leaves, a significant increase in the AT2 means were observed between the genotypes and between the years. The ECC 35 genotype was the most susceptible with an average AT2 score > 1, followed by IAN 873, ECC 83, ECC 60, ECC 25 and ECC 66. However, the highest score was observed in the third year, where ECC 35 had the highest severity scores (AT2 > 2), and was significantly superior to other genotypes in all years. Furthermore, for the second year, ECC 35 exhibited the highest susceptibility to SALB. On the other hand, the ECC 64, ECC 73, ECC 29 and ECC 90 genotypes were the less susceptible, with an average AT2 < 1 (Figure 2c).
With respect to the sexual phase of P. ulei, it was found that the ECC 35 and IAN 873 genotypes had higher formation of stroma (ST > 2). In the third year, ECC 35 was the most susceptible, with mean ST scores > 3, followed by IAN 873, ECC 60, ECC 25, ECC 66, ECC 83 and ECC 60 (all, ST > 2). In the second year, ECC 25, ECC 83 and ECC 60 had mean ST scores > 2. Overall, ECC 64 was the least susceptible, with an average ST < 1. The ECC 73, ECC 64 and ECC 90 genotypes had mean ST scores < 1, and were the least susceptible in the third year (Figure 2d).
The Co-Inertia analysis between the data matrices did not show significant covariation (p > 0.05). This indicated that the co-variation structure between early performance and SALB resistance was not entirely similar to the structures obtained in the individual analyses of each variable matrix. However, the first two axes of the Co-Inertia analysis explained 88.36% and 11.1% of the total variability (Figure 3a and b), indicating that the simultaneous analysis of both matrices was suitable for explaining the observed ordering pattern in the 10 genotypes of H. brasiliensis.
The most susceptible genotypes positioned itself in the lower-left quadrant of Figure 3c, characterized by lower vigor, and lower yield (DRY and DRC) (Figure 3d), as well as higher scores of AT2 and ST. Additionally, a positive association was observed between the ECC 60 genotype and AT1 and TR variables (i.e., a higher susceptibility in young leaves) (Figure 3e). On the other hand, the ECC 64 genotype showed higher values of CTo and CTpty3 (Figure 3d). This genotype had a greater vigor and lower SALB susceptibility, positioning itself in the lower-right quadrant of Figure 3c. The ECC 25, ECC 29, ECC 73, ECC 64 and ECC 90 genotypes were located towards the right side of the plot and had the best results in terms of growth and early yield, along with low susceptibility to SALB.
3. Discussion
This study evaluated the resistance to SALB, growth, and early yield of nine Colombian elite genotypes of H. brasiliensis from the ECC-100 series (Elite Caquetá Colombia) and IAN 873 (control) under conditions of low SALB pressure.
The climatic characteristics of the study area (i.e., average temperature of 25.1°C, average relative humidity in the driest months ranging from 69 to 75%, and precipitation of 2514.6 mm) were typical of regions with low SALB pressure [9,16,21,22,23]. These climatic parameters are unfavorable for the production of P. ulei spores [24], but they are optimal for rubber tree cultivation [9,19,22,25].
For the three years of the study, between November and March, there was a reduction in precipitation and relative humidity, along with an increase in temperature compared to the period from April to October. According to Sterling et al. [15], these climatic variations are optimal for the growth of H. brasiliensis in areas with no escape from SALB, resulting in more vigorous trees in the first years after planting. On the other hand, Rivano et al. [19] found that climatic variations allowed the foliage recovery of some clones during the dry season and under suitable phytosanitary conditions, they were not affected by SALB in a suboptimal rubber cultivation area in Ecuador. Furthermore, Sterling et al. [16] found that the climatic conditions in San Vicente del Caguán contribute to SALB resistance for introduced rubber tree genotypes from Brazil, Peru and Guatemala. Our results support the idea that these climatic conditions are favorable, promoting better vigor and greater resistance to SALB for Colombian rubber tree genotypes.
In this study, the circumference of trunk (CT) was higher than that reported in previous studies in the three first years of growth [16,22]. In escape SALB conditions, Sterling et al. [16], found CTpt values that ranged from 6.50 to 8.61 cm in the first year of planting. In contrast, in non-escape SALB conditions, Sterling et al. [15] with ECC-100 series genotypes reported CT values that ranged from 3.64 to 5.98 cm in the first year. Our results showed CTpt values between 8.73 and 10.69 cm in the first year, higher than that reported in the Colombian Amazon region.
In this study, in the second year, the average CTpt values ranged from 13.33 to 20.44 cm similar to that reported by Sterling et al. [26]. For the third year, the average CTpt values ranged from 20.23 cm to 27.48 cm, which was higher than that reported by Gireesh et al. [27] in India, where 20 promising genotypes had a girth between 15.3 and 23.0 cm. The ECC 64 and ECC 25 genotypes were the most vigorous, with an average annual trunk circumference increase of 7 to 8 cm, similar to Rivano et al. [19] that observed an 8 cm increase in circumference for the two more vigorous clones in a suboptimal rubber cultivation zone in Ecuador. This demonstrates the high potential of ECC-100 series genotypes for zones with low SALB pressure.
The rubber tree immaturity period ranges from four to eight years before conventional tapping is applied. Genotypes with high CT in the early years of planting reach the tapping period faster, allowing for early exploitation and early cost recovery for tree maintenance [27,28,29]. Therefore, one of the goals of rubber tree breeders is to identify genotypes that combine high yield and vigorous growth in early evaluation [30]. In this regard, the Colombian elite genotypes evaluated in this study showed a high CT in the first three years, similar to the highest-performing clones evaluated by Rivano et al. [19] and superior to Gireesh et al. [27].
Climate conditions and clonal variation have a direct impact on the phenotypic expression of H. brasiliensis [31,32]. This indicates that genotypes performance depends not only on the genetic nature of the trees but also on the environmental variations of the planting site [33]. In this study, a greater growth of ECC-100 series genotypes was observed compared to previous studies conducted in areas with favorable conditions for SALB [15]. These findings highlight the importance of considering both the selection of suitable genotypes and environmental conditions to maximize the performance of H. brasiliensis.
The dry rubber content (DRC) is an important parameter for assessing latex quality, as it is one of the main components that directly or indirectly contribute to rubber yield, and it determines the amount of rubber in a latex sample [34,35]. In this study, genotypes had a DRC that ranged from 26.24% to 32.83%. These results are lower than that reported by Mendoza-Vargas and Jiménez-Forero [36] (i.e., DRC of 34.3%) in a promising clonal collection from the FX series in Cundinamarca, Colombia. Furthermore, the percentages obtained are lower than that reported by Ajith et al. [37] in two locations in India (i.e., DRC ranged from 35.23% to 48.9%). The DRC values of IAN 873 (control) were also lower than that reported by Quesada-Méndez et al. [38] in genotypes with three years of planting (i.e., DRC of 40.91%), and lower than Mendoza-Vargas and Jiménez-Forero [36] (i.e., DRC of 41.3%).
The differences observed in DRC between the genotypes are mainly due to variations in the colloidal composition of latex, which depend on soil conditions, climate, tapping duration, diseases, and plant material. These factors primarily affect the DRC composition. Overall, the DRC values range between 27 and 45% [39,40]. This indicate that the results of our study fall within the optimal composition ranges.
Higher DRC values represent greater productivity in terms of dry rubber yield. However, elevated percentages also imply an increase in the viscosity of the colloidal latex composition, which can lead to blockages in the laticifer mantle and obstruction in latex flow from the tapping panel [34,41], which can reduce dry rubber yield (DRY). In this study, DRY values ranged from 0.42 to 5.57 g.tree−1.tap−1, which are lower than that reported by Ajith et al. [37] (i.e., DRY between 2.88 and 54.5 g.tree−1.tap−1).
Assessing the temporal dynamics of SALB resistance is crucial in determining optimal areas for crop establishment [15]. This study revealed different susceptibility patterns depending on the year and genotype. The severity of the attack on both young and mature leaves increased significantly over the years, especially in the third year and in the ECC 35, ECC 60 and IAN 873 genotypes. These results are consistent with Sterling et al. [15], where the ECC 35 and ECC 60 genotypes were the most affected in non-escape SALB areas. Furthermoe, the ECC 64, ECC 73, ECC 90, ECC 25 and ECC 29 genotypes showed a lower susceptibility, positively correlating with a higher vigor and early yield. These results differ from Sterling et al. [15], where the genotypes with higher resistance did not necessarily exhibit greater vigor.
With respect to asexual or conidial sporulation, it occurs on young leaves, and during this phase, conidia can disperse for short distances during dry periods [42], but it doesn't ensure disease survival [43]. However, in very rainy seasons, conidia can be particularly abundant [42]. In this study, the San Vicente del Caguán site is characterized by low precipitation and well-defined dry periods (i.e., semi-humid warm climate), which is unfavorable for the production of P. ulei conidia (i.e., low TR scores).
In relation to sexual sporulation, which provides quantitative information about the intensity of ascospore on mature leaves [44], the attack severity increased in the second and third years. In the second year, a moderate level of attack was evident for all genotypes, with the exception of the ECC 29 and ECC 64 genotypes, which had lower attack levels. In the third year, there was an increase in susceptibility; however, moderate attack levels were still recorded for all genotypes, with the exception of the ECC 35 genotype, which had severe attacks. Overall, we observed an increase in the ascospores production over the years. However, the ECC 64, ECC 73, ECC 90, ECC 25 and ECC 29 genotypes were the less susceptible. The SALB scores in the first year are similar to that reported by Rivano et al. [45] in the CDC 312, FDR 4575, FDR 5597 and MDF 180 clones, which were classified as completely resistant, and in the third year, the SALB scores are similar to that reported by Sterling et al. [20].
Considering that clones with high-yield and SALB-resistant is rare [46], this dual choice criteria is very important in rubber tree breeding programs. Based on these results, the ECC 64, ECC 73, ECC 90, ECC 25 and ECC 29 genotypes are considered potential candidates for establishment in areas with low SALB pressure in the Colombian Amazon, due to these genotypes have low susceptibility to SALB and high performance in the first three years of growth.
4. Materials and Methods
4.1. Study Area
4.1.1. Location
The study was conducted from July 2017 to June 2020 on the Parcela No. 12 farm, located in the Buenos Aires rural settlement area, in the Municipality of San Vicente del Caguán, Caquetá, Colombia. This site is located in the southern of the Colombian Amazon ( 02°01'42.62" north and 74°54'38.95" west, at an elevation of 346 meters above sea level. According to the classification by IGAC [47], the landscape in the study zona is hilly, characterized by hills with heights of less than 300 meters and slopes ranging from 7 to 12 percent. The region is a zone with a low SALB pressure, making it a promising region for the rubber cultivation [18,21].
4.1.2. Soils
The soils in Caquetá have unfavorable characteristics such as high compaction, low fertility and infiltration, poor drainage, shallow depth and a pH range between 4.5 and 5.8. Furthermore, these soils have high levels of aluminum saturation and low contents of Sodium (Na), Phosphorus (P), Potassium (K) and Magnesium (Mg) [47].
The study area had a very acidic pH of 5.05, an organic matter (OM) content of 1.21%, and an organic carbon (OC) content of 0.715%. Saturation levels were 17.51% (Ca), 1.18% (K), 4.99% (Mg) and 2.23% (Na). Elemental contents were as follows: Iron (Fe) (161.59 mg kg−1), Manganese (Mn) (4.27 mg kg−1), Copper (Cu) (1.71 mg kg−1), Zinc (Zn) (0.54 mg kg−1), Boron (B) (0.19 mg kg−1), Phosphorus (P) (0.84 mg kg−1), Sulfur (S) (6.91 mg kg−1) and total Nitrogen (N) (0.06%).
4.1.3. Climate
The climatic conditions in the study area correspond to a semi-humid warm climate, typical of the tropical wet region. It records an average temperature of 25.4°C, a relative humidity of 82.3% and an annual precipitation of 2503 mm [47]. The climate follows a monomodal regime with a transition to a dry period from July to October, followed by a dry period with less rain between November and February, and finally, a rainy period from March to June [48].
Microclimatic data for the study area were collected with a Portable Microclimate Station (Decagon Devices Inc, USA), from which monthly averages were calculated between July 2017 and June 2020 (Figure 1).
4.2. Experimental Design and Plot Management
A large-scale clonal trial (LSCT) [49], was established using a randomized complete block design (RCBD) with four replications (Fisher blocks). Each treatment consisted of planting 60 trees in single rows, with a planting distance of 7.0 x 3.0 meters, equivalent to a planting density of 476 trees per hectare and the total area of the LSCT was 5.04 hectares. The measured plot unit was 1260 square meters, containing 60 trees organized in 3 rows of 20 trees [15,16,20]
With respect to the management of the experimental plots, the fertilization plan employed by Sterling et al. [50] was applied every six months. Thus, a compound fertilizer [N (15%), P2O5 (15%), K2O (15%), CaO (2.2%), and SSO4 (1.7%)] was used at a rate of 150 g per plant. Minor elements included [N (8%), P2O5 (5%), CaO (18%), MgO (6%), S (1.6%), B (1%), Cu (0.14%), Mo (0.005%), and Zn (2.5%)], applied at a rate of 75 g per plant. Additionally, organic compost was applied at a dosage of 1000 g per plant. Weed control was carried out every three months and no phytosanitary controls were performed [15,16,20].
4.3. Plant material
Nine Colombian elite H. brasiliensis genotypes from the ECC-100 series (Elite Caquetá Colombia) were evaluated: ECC 25, ECC 29, ECC 35, ECC 60, ECC 64, ECC 66, ECC 73, ECC 83 and ECC 90. These genotypes were obtained from asexual propagation (cloning) of plus trees of H. brasiliensis from rubber-producing farms in Caquetá, Colombia, through natural cross-pollination and were characterized using molecular markers [15].
4.4. Evaluations and Data Analysis
4.4.1. Growth and early yield
The annual trunk circumference at 1.2 m above the ground was recorded during the pre-tapping phase (CTpt) of all trees, starting from July 2017 to June 2020, and in the early tapping phase in the third year (CTpty3). Then, the CT mean was calculated only for the trees that reached the threshold for extraction (CT ≥ 25 cm) at 1.2 m above the ground at the opening of the tapping panel (circumference of the trunk at opening, CTo) of the trees suitable for early tapping in the third year after planting, modified from Silva et al. [54].
The dry rubber yield (DRY) (g.tree−1.tap−1) was recorded during the first six months of tapping, following the methods described by Meenakumari et al. [55] and Silva et al. [54]. The dry rubber content (DRC) as a percentage was measured according to described by Lacote et al. [56], and Mendoza-Vargas and Jiménez-Forero [36].
4.4.2. Resistance to SALB
The resistance of H. brasiliensis to SALB was assessed monthly in 50% of the trees in each plot (30 trees) between July 2017 and June 2020, using a method adapted from Rivano et al. [45]. The data were analyzed by year. To evaluate resistance to P. ulei, both young and mature leaves (stages C and D, respectively) were examined [16].
The severity of the attack on young leaves (AT1) and mature leaves (AT2) was assessed using a scale adapted by Rivano et al. [45] based on the scale proposed by Chee and Holliday [57]. This scale defines five categories based on the percentage of leaf surface with symptoms or signs of the disease: 0 represents no attack (<1%); 1 represents minor attack (1-5%); 2 represents moderate attack (6-15%); 3 represents severe attack (16-30%) and 4 represents very severe attack (>30%).
In young leaves, the intensity of conidial sporulation (asexual phase of the fungus) on the lesions formed on the leaf surface at stage C was visually assessed using the variable "type of reaction" (TR). This variable is based on a scale from 1 to 6 proposed by Junqueira et al. [58] and adapted by Mattos et al. [59]. The scores on this scale are as follows: 1, Necrotic lesions without spores; 2, Non-necrotic lesions without spores; 3, Very scarce and heterogeneous sporulation on the underside of the leaf; 4, High sporulation, heterogeneous, or partial on the underside of the leaf; 5, Very high and uniform sporulation covering the entire lesion on the underside of the leaf; 6, Very high sporulation covering the entire lesion on both the underside and upper side of the leaf.
The intensity of sexual sporulation of the fungus on the upper side of the leaves at stage D was assessed using the variable "stroma density" (ST). A scale proposed by Rivano et al. [45] was used, defining four scores based on the number of stromata per leaflet: 0, absence of stromata; 1, fewer than 5 stromata; 2, between 5 and 10 stromata; 3, between 11 and 30 stromata; and 4, more than 30 stromata. For all these variables, the most severely affected leaves on each plant were evaluated and the average of each variable was calculated for each replicate of each genotype [20].
4.4.3. Statistical analysis
A linear mixed-effects model (LME) was used to analyze the fixed factors genotype, year and their interaction for the variables CTpt, AT1, AT2, TR and ST. QQ-plots of the model residuals were used to assess the normality assumption and fitted-plot (residual vs. predicted) for the homogeneity of variances. A transformation was applied to the AT1, AT2, TR and ST variables to normalize the data, using the formula: (value + 0.5)1/2. The effect of genotype on the growth variables was evaluated at the end of the evaluation cycle. Block and plot effects associated with genotypes within blocks were considered as random effects. Residual variance was modeled to adjust the heteroscedasticity by genotype, and residual correlation for successive observations was considered using models for longitudinal data: Compound Symmetry with Heterogeneous Variances (CSH), First-order Autoregressive with Heterogeneous Variances (ARH1), Continuous First-order Autoregressive with Heterogeneous Variances (CARH1) and Unstructured with Heterogeneous Variances (UNH) [60]. Akaike (AIC), Bayesian (BIC) and log-likelihood (Log Lik) criteria were used to select the structure of variances and residual correlations [20,60].
To analyze the effect of genotype on variables related to early yield (CTo, DRY, and DRC), a linear mixed-effects model (LME) with independent errors was used. The analyses were performed with the 'lme' function (Linear Mixed-Effects Models) in the 'nlme' package (Linear and Nonlinear Mixed Effects Models) [61] in the R language software, version 3.5.1 [62] and the interface in InfoStat v. 2018 [63]. Differences between means of the variables were analyzed using Fisher's LSD test (α = 0.05).
To explore the relationships between performance (growth and early yield) and SALB resistance in the 10 genotypes, two matrices were constructed with each group of variables, and a Principal Component Analysis (PCA) was performed as an ordination method. The results of the ordination of the two groups of variables were carried out with a co-inertia analysis with the Monte Carlo test [64]. The software used for this analysis was ADE-4 package [65], in R 3.5.1 [62].
5. Conclusions
The results of this study support the idea that, under favorable climatic conditions (i.e., zones with low SALB pressure), the genotypes from the ECC-100 series had higher yields and lower SALB susceptibility, and, therefore, could be successfully established in regions such as Colombian Amazon. This research contributes with the development of more effective management strategies based on genetic resistance to SALB.
The most genotypes showed low susceptibility to P. ulei. Over time, the impact of the disease increased significantly; however, when compared to other studies, the susceptibility level was very low (partial resistance to SALB, with means scores of TR < 2 and ST < 3). Furthermore, the genotypes with lower susceptibility to SALB expressed higher early performance indices (CTpt > 23 cm, DRY > 3 g.tree− 1 .tap− 1 and DRC > 31% in the 3th year after planting). The optimal early responses in yield, growth and SALB resistance reported in this study, are of interest for the selection of new desirable H. brasiliensis genotypes for the Amazon region. However, it is necessary to analyzed the productive performance and SALB resistance during the crop maturity period, before generating a commercial recommendation under the Colombian Amazon region conditions.
Author Contributions
Conceptualization, L.C.G.-R. and A.S..; methodology, L.C.G.-R., A.S., and J.A.F.R.; software, A.S. and H.M.-R.; validation, L.C.G.-R. and A.S.; formal analysis, L.C.G.-R., and A.S..; investigation, L.C.G.-R., A.S., H.M.-R. and J.A.F.R.; resources, L.C.G.-R. and A.S.; data curation, L.C.G.-R. and A.S.; writing—original draft preparation, L.C.G.-R., A.S., and H.M.-R.; writing—review and editing, , L.C.G.-R., A.S. and H.M.-R.; visualization, L.C.G.-R., A.S. and H.M.-R.; supervision, L.C.G.-R., and A.S.; project administration, L.C.G.-R., and A.S.; funding acquisition, L.C.G.-R., and A.S.. All authors have read and agreed to the published version of the manuscript.
Funding
This research is part of the “Ampliación de la base genética de caucho natural, Caquetá, Amazonía” project, funded by Fondo de Ciencia, Tecnología e Innovación FCTeI—SGR, Contract 59/2013 Instituto Amazónico de Investigaciones Científicas SINCHI—Gobernación del Caquetá—the Universidad de la Amazonía—the Asociación de Reforestadores y Cultivadores de Caucho del Caquetá Asoheca.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are available from the authors upon request.
Acknowledgments
The authors thank ASOHECA, the owners of the experimental farms, and the project’s technical personnel, who made the field work possible.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Rivano, F.; Mattos, C.R.R.; Cardoso, S.E.A.; Martinez, M.; Cevallos, V.; Le Guen, V.; Garcia, D. Breeding Hevea Brasiliensis for Yield, Growth and SALB Resistance for High Disease Environments. Ind. Crops Prod. 2013, 44, 659–670. [Google Scholar] [CrossRef]
- Mazlan, S.; Jaafar, N.; Wahab, A.; Sulaiman, Z.; Rajandas, H.; Zulperi, D. Major Diseases of Rubber ( Hevea Brasiliensis ) in Malaysia. Pertanika J. Sch. Res. Rev. 2019, 5, 10–21. [Google Scholar]
- Prabhakaran, K.P. The Agronomy and Economy of Important Tree Crops of the Developing World; Nair, K.P.P., Ed.; Elsevier: New Delhi, India, 2010; ISBN 978-0-12-384677-8.
- FAO FAOSTAT; http://www.fao.org/faostat/, 2023;
- Sant’Anna, I. de C.; Gouvêa, L.R.L.; Martins, M.A.; Scaloppi Junior, E.J.; de Freitas, R.S.; Gonçalves, P. de S. Genetic Diversity Associated with Natural Rubber Quality in Elite Genotypes of the Rubber Tree. Sci. Rep. 2021, 11, 1–10. [Google Scholar] [CrossRef]
- Truong, V. Van; Lam, L. Van; Thuy, L.M.; Dinh, H.D.; Thao, N.T.; Rivallan, R.; Biet, H. Van; Guen, V. Le Population Genetic Structure of a Thousand Rubber Tree Accessions from Wild Rondônia Populations Conserved in Vietnam. Genet. Resour. Crop Evol. 2020, 67, 475–487. [Google Scholar] [CrossRef]
- Maiden, N.A.; Syd Ali, N.; Ahmad, K.; Atan, S.; Wong, M.Y. Growth and Physiological Responses of Hevea Brasiliensis to Rigidoporus Microporus Infection. J. Rubber Res. 2022, 25, 213–221. [Google Scholar] [CrossRef]
- Malmonge, J.A.; Camillo, E.C.; Moreno, R.M.B.; Mattoso, L.H.C.; McMahan, C.M. Comparative Study on the Technological Properties of Latex and Natural Rubber from Hancornia Speciosa Gomes and Hevea Brasiliensis. J. Appl. Polym. Sci. 2008, 111, 2986–2991. [Google Scholar] [CrossRef]
- Jaimes, Y.; Rojas, J.; Cilas, C.; Furtado, E.L. Suitable Climate for Rubber Trees Affected by the South American Leaf Blight (SALB): Example for Identification of Escape Zones in the Colombian Middle Magdalena. Crop Prot. 2016, 81, 99–114. [Google Scholar] [CrossRef]
- Lieberei, R. South American Leaf Blight of the Rubber Tree (HeveaSpp.): New Steps in Plant Domestication Using Physiological Features and Molecular Markers. Ann. Bot. 2007, 100, 1125–1142. [Google Scholar] [CrossRef]
- da Hora Júnior, B.T. Da; de Macedo, D.M.; Barreto, R.W.; Evans, H.C.; Mattos, C.R.R.; Maffia, L.A.; Mizubuti, E.S.G. Erasing the Past: A New Identity for the Damoclean Pathogen Causing South American Leaf Blight of Rubber. PLoS One 2014, 9. [Google Scholar] [CrossRef]
- Le Guen, V.; Garcia, D.; Mattos, C.R.R.; Doaré, F.; Lespinasse, D.; Seguin, M. Bypassing of a Polygenic Microcyclus Ulei Resistance in Rubber Tree, Analyzed by QTL Detection. New Phytol. 2007, 173, 335–345. [Google Scholar] [CrossRef]
- Sobha, S.; Rekha, K.; Uthup, T.K.; Abstract Biotechnological Advances in Rubber Tree (Hevea Brasiliensis Muell. Arg.) Breeding. In Advances in Plant Breeding Strategies: Industrial and Food Crops; Al-Khayri, J.M., Jain, S.M., Johnson, D. V, Eds.; Springer Nature Switzerland: Switzerland, 2019; Vol. 6, pp. 179–236 ISBN 9783030232658.
- Sterling, A.; Pimentel-Parra, G.A.; Virguez-Díaz, Y.R.; Suárez-Córdoba, Y.D.; Hoyos-Duarte, J.D.; Fonseca-Restrepo, J.A. Long-Term Resistance in Promising Rubber Tree Genotypes as a Breeding Source for Improving South American Leaf Blight Management under High Disease Incidence in the Colombian Amazon. Crop Prot. 2021, 150. [Google Scholar] [CrossRef]
- Sterling, A.; Galindo-Rodríguez, L.C.; Suárez-Córdoba, Y.D.; Velasco-Anacona, G.; Andrade-Ramírez, T.; Gómez-Torres, A.K. Early Assessing Performance and Resistance of Colombian Rubber Tree Genotypes under High South American Leaf Blight Pressure in Amazon. Ind. Crops Prod. 2019, 141. [Google Scholar] [CrossRef]
- Sterling, A.; Martínez-Viuche, E.J.; Suárez-Córdoba, Y.D.; Agudelo-Sánchez, A.A.; Fonseca-Restrepo, J.A.; Andrade-Ramírez, T.K.; Virguez-Díaz, Y.R. Assessing Growth, Early Yielding and Resistance in Rubber Tree Clones under Low South American Leaf Blight Pressure in the Amazon Region, Colombia. In Industrial Crops and Products; Elsevier B.V., 2020; Vol. 158.
- Sterling, A.; Rodríguez, C.H. Valoración de Nuevos Clones de Hevea Brasiliensis Con Proyección Para La Amazonia Colombiana: Fases de Pre y Post-Sangría Temprana En El Caquetá; Sterling, A., Rodríguez, C.H., Eds.; Instituto Amazónico de Investigaciones Científicas - SINCHI: Bogotá (Colombia), 2020; ISBN 978-958-5427-25-9. [Google Scholar]
- Sterling, A.; Rodríguez, C.H. Estrategias de Manejo Para Las Principales Enfermedades y Plagas Del Cultivo Del Caucho Con Énfasis En La Amazonia Colombiana.; Instituto Amazónico de Investigaciones Científicas - SINCHI: Florencia Caquetá, 2018; ISBN 9789585427082. [Google Scholar]
- Rivano, F.; Vera, J.; Cevallos, V.; Almeida, D.; Maldonado, L.; Flori, A. Performance of 10 Hevea Brasiliensis Clones in Ecuador, under South American Leaf Blight Escape Conditions. Ind. Crops Prod. 2016, 94, 762–773. [Google Scholar] [CrossRef]
- Sterling, A.; Martínez-Viuche, E.J.; Pimentel-Parra, G.A.; Suárez-Córdoba, Y.D.; Fonseca-Restrepo, J.A.; Virguez-Díaz, Y.R. Dynamics of Adaptive Responses in Growth and Resistance of Rubber Tree Clones under South American Leaf Blight Non-Escape Conditions in the Colombian Amazon. Ind. Crops Prod. 2019, 141. [Google Scholar] [CrossRef]
- Castañeda, A. Zonas Aptas Para El Cultivo de Caucho En Colombia; Serie Técnica No. 39 - CONIF: Bogotá, Colombia, 1997;
- Correa-Pinilla, D.E.; Gutiérrez-Vanegas, A.J.; Gil-Restrepo, J.P.; Martínez-Atencia, J.; Córdoba-Gaona, O. de J. Agroecological and South American Leaf Blight Escape Zones for Rubber Cultivation in Colombia. Agron. J. 2022, 114, 2830–2844. [Google Scholar] [CrossRef]
- Da Silva, K.R.; Cecílio, R.A.; Xavier, A.C.; Pezzopane, J.R.M.; Garcia, E.G. de O. Zoneamento Edafoclimático Para a Cultura Da Seringueira No Espírito Santo. 2013, 1–12.
- Guyot, J.; Condina, V.; Doaré, F.; Cilas, C.; Sache, I. Segmentation Applied to Weather-Disease Relationships in South American Leaf Blight of the Rubber Tree. Eur. J. Plant Pathol. 2010, 126, 349–362. [Google Scholar] [CrossRef]
- Rivano, F.; Maldonado, L.; Simbaña, B.; Lucero, R.; Gohet, E.; Cevallos, V.; Yugcha, T. Suitable Rubber Growing in Ecuador: An Approach to South American Leaf Blight. Ind. Crops Prod. 2015, 66, 262–270. [Google Scholar] [CrossRef]
- Sterling, A.; Suárez, J.C.; Rodríguez, C.H.; Caicedo, D.F.; Salas, Y.M.; Virguez, Y.R. Crecimiento Inicial de Clones Promisorios de Hevea Brasiliensis (Willd. Ex A. Juss.) Müll. Arg. En Sistema Agroforestal En Caquetá, Colombia. Colomb. For. 2015, 18, 175. [Google Scholar] [CrossRef]
- Gireesh, T.; Meenakumari, T.; Mydin, K.K. Fast Track Evaluation and Selection of Hevea Brasiliensis Clones from a Clonal Nursery. Ind. Crops Prod. 2017, 103, 195–201. [Google Scholar] [CrossRef]
- Gonçalves, P.D.S.; Bortoletto, N.; Furtado, E.L. Desempenho de Clones de Seringueira Da Série IAC 300 Selecionados Para a Região Noroeste Do Estado de São Paulo ( 1 ) Material e Métodos. 2001, 589–599.
- Vrignon-Brenas, S.; Gay, F.; Ricard, S.; Snoeck, D.; Perron, T.; Mareschal, L.; Laclau, J.P.; Gohet, É.; Malagoli, P. Nutrient Management of Immature Rubber Plantations. A Review. Agron. Sustain. Dev. 2019, 39. [Google Scholar]
- Gouvêa, L.R.L.; Silva, G.A.P.; Verardi, C.K.; Oliveira, A.L.B.; Gonçalves, P. de S. Simultaneous Selection of Rubber Yield and Girth Growth in Young Rubber Trees. Ind. Crops Prod. 2013, 50, 39–43. [Google Scholar] [CrossRef]
- Gonçalves, P. de S.; Silva, M. de A.; Gouvêa, L.R.L.; Scaloppi Junior, E.J. Genetic Variability for Girth Growth and Rubber Yield in Hevea Brasiliensis. Sci. Agric. 2006, 63, 246–254. [Google Scholar] [CrossRef]
- Vélez-Sánchez, D.F.; Nieto-Rodríguez, V.M. Desempeño Inicial de Clones de Caucho En El Municipio de Puerto Carreño, Vichada, Colombia. Colomb. For. 2013, 16, 186–199. [Google Scholar]
- Vinod, K.K.; Rajeswari, M.J.; Pothen, J.; Krishnakumar, A.K.; Sethuraj, M.R. Performance Analysis for Wintering Pattern in Hevea Brasiliensis Clones; 1996; Vol. 9;
- Quesada-Méndez, I.; Aristizábal-Gutiérrez, F.; Montoya-Castaño, D. Characterization of Two Latex Parameters from Hevea Brasiliensis (Willd. Ex A. Juss.) Müll. Arg. Clones in the Colombian Altillanura. Colomb. For. 2012, 15, 139. [Google Scholar] [CrossRef]
- Sethuraj, M.R. Yield Components in Hevea Brasiliensis. In Natural Rubber Biology, Cultivation and Technology; Sethuraj, M.R., Mathew, N.M., Eds.; Developments in Crop Science; Elsevier: Kerala, India, 1992; Volume 23, pp. 137–163. [Google Scholar]
- Mendoza-Vargas, L.P.; Jiménez-Forero, J.A. Effect of Inclusion of Anticoagulants during the Benefit of Hevea Brasiliensis (Willd. Ex a.Juss.) Müll. Arg. Colomb. For. 2018, 21, 91–101. [Google Scholar] [CrossRef]
- Ajith, S. V.; Nair, N.N.; Sathik, M.B.M.; Meenakumari, T. Breeding for Low-Temperature Stress Tolerance in Hevea Brasiliensis: Screening of Newly Developed Clones Using Latex Biochemical Parameters. J. Rubber Res. 2021, 24, 819–828. [Google Scholar] [CrossRef]
- Quesada-Méndez, I.; Aristizábal-Gutiérrez, F. Characterization of Two Latex Parameters from Hevea Brasiliensis (Willd. Ex A. Juss.) Müll. Arg. Clones in the Colombian Altillanura. Colomb. For. 2012, 15, 139–146. [Google Scholar]
- Cáceres, S.A.P.; Gauthier-Maradei, P. Thermogravimetric Analysis as a New Method to Determine of Total Solid Content (TSC) and Dry Rubber Content (DRC) of Natural Latex. Rev. ION 2012, 57–65. [Google Scholar]
- Támara, A.M.; Reyes, H. Physicochemical Analysis and Characterization of the Hevea Brasiliensis Species. Entre Cienc. e Ingienería 2015, 18, 9–15. [Google Scholar]
- Jacob, J..; Prévöt, J.C.; Roussel, D.; Lacrotte, R.; Serres, E.; d’Auzac, J.; Eschbach, J.M.; Omont, H. Yield Limiting Factors for Latex Yield of Hevea. In Physiology of rubber tree latex. The laticiferous cell and latex : a model of cytoplasm; D’Auzac, J., Jacob, J.-L., Chrestin, H., Eds.; CRC Press: Florida, United States, 1989; Vol. 1, pp. 345–397 ISBN 0849348935.
- Guyot, J.; Condina, V.; Doaré, F.; Cilas, C.; Sache, I. Role of Ascospores and Conidia in the Initiation and Spread of South American Leaf Blight in a Rubber Tree Plantation. Plant Pathol. 2014, 63, 510–518. [Google Scholar] [CrossRef]
- Guyot, J.; Le Guen, V. A Review of a Century of Studies on South American Leaf Blight of the Rubber Tree. Plant Dis. 2018, 1–14. [Google Scholar] [CrossRef]
- Rivano, F.; Martinez, M.; Cevallos, V.; Cilas, C. Assessing Resistance of Rubber Tree Clones to Microcyclus Ulei in Large-Scale Clone Trials in Ecuador: A Less Time-Consuming Field Method. Eur. J. Plant Pathol. 2010, 126, 541–552. [Google Scholar] [CrossRef]
- Rivano, F.; Martinez, M.; Cevallos, V.; Cilas, C. Assessing Resistance of Rubber Tree Clones to Microcyclus Ulei in Large-Scale Clone Trials in Ecuador: A Less Time-Consuming Field Method. Eur. J. Plant Pathol. 2010, 126, 541–552. [Google Scholar] [CrossRef]
- Le Guen, V.; Garcia, D.; Mattos, C.; Fouet, O.; Doaré, F.; Condina, V.; Seguin, M. A Newly Identified Locus Controls Complete Resistance to Microcyclus Ulei in the Fx2784 Rubber Clone. Tree Genet. Genomes 2013, 9, 805–812. [Google Scholar] [CrossRef]
- Instituto Geográfico Agustín Codazzi (IGAC) Estudio General de Suelos y Zonificación de Tierras Departamento de Caquetá Escala 1.100.000; Imprenta Nacional de Colombia: Bogotá (Colombia), 2014.
- Instituto Geográfico Agustín Codazzi -IGAC Caquetá. Características Geodráficas; IGAC: Bogotá (Colombia), 2010; ISBN 978-958-8323-37-4.
- Clément-Demange, A.; Nicolas, D.; Legnaté, H.; Rivano, F.; Le Guen, V.; Gnagne, M.; Chapuset, T. Hévéa: Stratégies de Sélection. Plant. Rech. Développement 1995, 2, 5–14. [Google Scholar]
- Sterling, A.; Andrade-Ramírez, T.K.; Clavijo-Arias, E.A.; Guaca-Cruz, L.; Celis-Hernández, M.D.; Noriega, J.E.; Torres-Barrero, R.; Suárez-Salazar, J.C. Evaluación de Los Rasgos de Crecimiento, Nutricionales y Fisiológicos de Genotipos Caquetenses Selección ECC 1 de Caucho (Hevea Brasiliensis) En Campos Clonales a Gran Escala En El Caquetá. In Valoración de nuevos clones de Hevea brasiliensis con proyección para la Amazonia colombiana: fases de pre y post-sangría temprana en el Caquetá; Sterling, A., Rodríguez, C.H., Eds.; Instituto Amazónico de Investigaciones Científicas SINCHI: Bogotá, Colombia, 2020; pp. 127–152 ISBN 978-958-5427-25-9.
- Sterling, A.; Rodríguez, C.H. Nuevos Clones de Cuacho Natural Para La Amazonía Colombiana :Enfasís En La Resistencia Al Mal Suramericano de Las Hojas (Mycrocyclus Ulei); Sterling, A., Rodríguez, C.H., Eds.; 1st ed.; Instituto Amazónico de Investigaciones Científicas SINCHI: Bogotá, Colombia, 2011; ISBN 978-958-8317-68-7.
- 2015; 52. Confederación Cauchera Colombiana (CCC) Estado Actual Del Gremio Cauchero Colombiano; Bogotá, Colombia, 2015;
- Sterling, A.; Rodríguez, C.H. Estrategias de Manejo Para Las Principales Enfermedades y Plagas Del Cultivo Del Caucho Con Énfasis En La Amazonia Colombiana; Instituto Amazónico de Investigaciones Científicas - SINCHI: Florencia, Colombia, 2018; ISBN 978-958-5427-08-2.
- Silva, G.A.P.; Gouvêa, L.R.L.; Verardi, C.K.; Oliveira, A.L.B. de; Gonçalves, P. de S. Annual Growth Increment and Stability of Rubber Yield in the Tapping Phase in Rubber Tree Clones: Implications for Early Selection. Ind. Crops Prod. 2014, 52, 801–808. [Google Scholar] [CrossRef]
- Meenakumari, T.; Meenattoor, J.R.; Thirunavoukkarasu, M.; Vinod, K.K.; Krishan, B.; Gireesh, T.; Thomas, V.; Mydin, K.K.; Jacob, J. Dynamics of Long-Term Adaptive Responses in Growth and Rubber Yield among Hevea Brasiliensis Genotypes Introduced to a Dry Sub-Humid Climate of Eastern India. Ind. Crops Prod. 2018, 119, 294–303. [Google Scholar] [CrossRef]
- Lacote, R.; Gabla, O.; Obouayeba, S.; Eschbach, J.M.; Rivano, F.; Dian, K.; Gohet, E. Long-Term Effect of Ethylene Stimulation on the Yield of Rubber Trees Is Linked to Latex Cell Biochemistry. F. Crop. Res. 2010, 115, 94–98. [Google Scholar] [CrossRef]
- Chee, K.H.; Holliday, P. South American Leaf Blight of Hevea Rubber. Malaysian Rubber Res. Dev. Board Monogr. 1986, 13, 50. [Google Scholar]
- Junqueira, N.T. V; Chaves, G.M.; Zambolim, L.; Gasparotto, L.; Alfenas, A.C. Variabilidade Fisiológica de Microcyclus Ulei. Fitopatol. Bras. 1986, 11, 823–833. [Google Scholar]
- Mattos, C.; Garcia, D.; Pinard, F.; Le Guen, V. Variabilidade de Isolados de Microcyclus Ulei No Sudeste de Bahia. Fitopatol. Bras. 2003, 5, 502–507. [Google Scholar]
- Verbeke, G.; Molenberghs, G. Linear Mixed Models For Longitudinal Data. 2010.
- Pinheiro, J.; Bates, D.; Saikat, D. Deepayan Sarkar and the R Development Core Team. Nlme: Linear and Nonlinear Mixed Effects Models. R Packag. 2013, 3, 1–109. [Google Scholar]
- R. Core Team R: A Language and Environment for Statistical Computing. 2018.
- Di Rienzo, J.A.; Casanoves, F.; Balzarine, M.G.; Gonzales, L.; Tablada, M.; Robledo, C.W. InfoStat Versión 2018 2018.
- Dolédec, S.; Chessel, D. Co-inertia Analysis: An Alternative Method for Studying Species–Environment Relationships. Freshw. Biol. 1994, 31, 277–294. [Google Scholar] [CrossRef]
- Thioulouse, J.; Chessel, D.; Dolédec, S.; Olivier, J.-M. ADE-4 a Multivariate Analysis and Graphical Display Software. Stat. Comput. 1997, 7. [Google Scholar] [CrossRef]
Figure 1.
Monthly precipitation, T-max and T-min (°C): maximum and minimum temperature; RH (%): Relative humidity (San Vicente del Caguán, Caquetá, Colombia), between July 2017 and June 2020.
Figure 1.
Monthly precipitation, T-max and T-min (°C): maximum and minimum temperature; RH (%): Relative humidity (San Vicente del Caguán, Caquetá, Colombia), between July 2017 and June 2020.
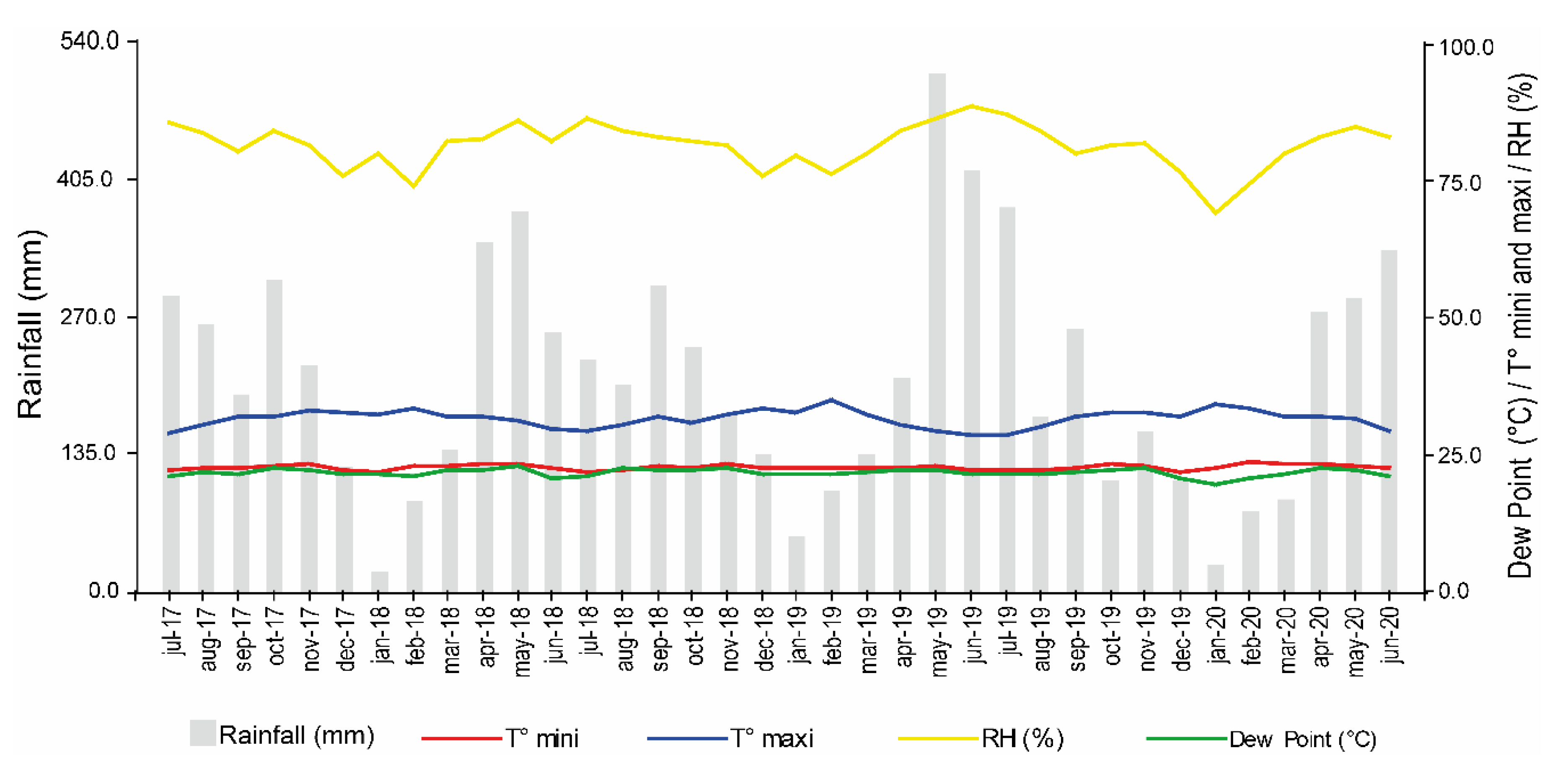
Figure 2.
Results of susceptibility to SALB over three years after planting. The following parameters were evaluated: (a) Attacks of P. ulei on young leaves at stage C (AT1). (b) Type of reaction (TR) to P. ulei on young leaves at stage C. (b) Attacks of P. ulei on mature leaves at stage D (AT2). (d) Stromal density on mature leaves at stage D (ST). Means between the genotypes followed by the same letter in the same year do not differ statistically (Fisher's LSD test, p < 0.05).
Figure 2.
Results of susceptibility to SALB over three years after planting. The following parameters were evaluated: (a) Attacks of P. ulei on young leaves at stage C (AT1). (b) Type of reaction (TR) to P. ulei on young leaves at stage C. (b) Attacks of P. ulei on mature leaves at stage D (AT2). (d) Stromal density on mature leaves at stage D (ST). Means between the genotypes followed by the same letter in the same year do not differ statistically (Fisher's LSD test, p < 0.05).
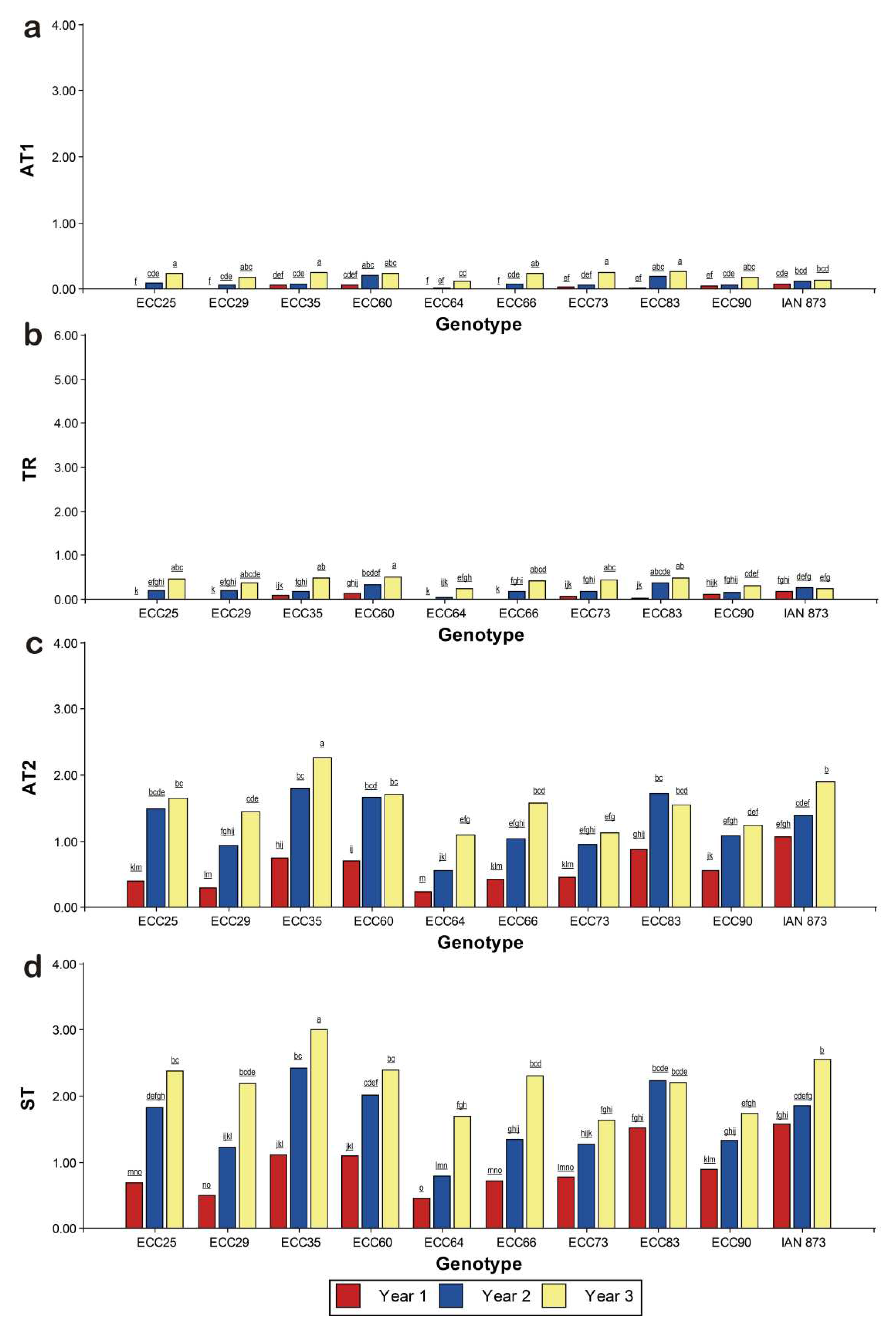
Figure 3.
Co-Inertia analysis between SALB resistance of 10 genotypes of Hevea brasiliensis: a. PCA (Principal Component Analysis); b. Captured Inertia (%); c. Genotype ordination plot; d. Growth and early yield projection; e. SALB resistance projection.
Figure 3.
Co-Inertia analysis between SALB resistance of 10 genotypes of Hevea brasiliensis: a. PCA (Principal Component Analysis); b. Captured Inertia (%); c. Genotype ordination plot; d. Growth and early yield projection; e. SALB resistance projection.
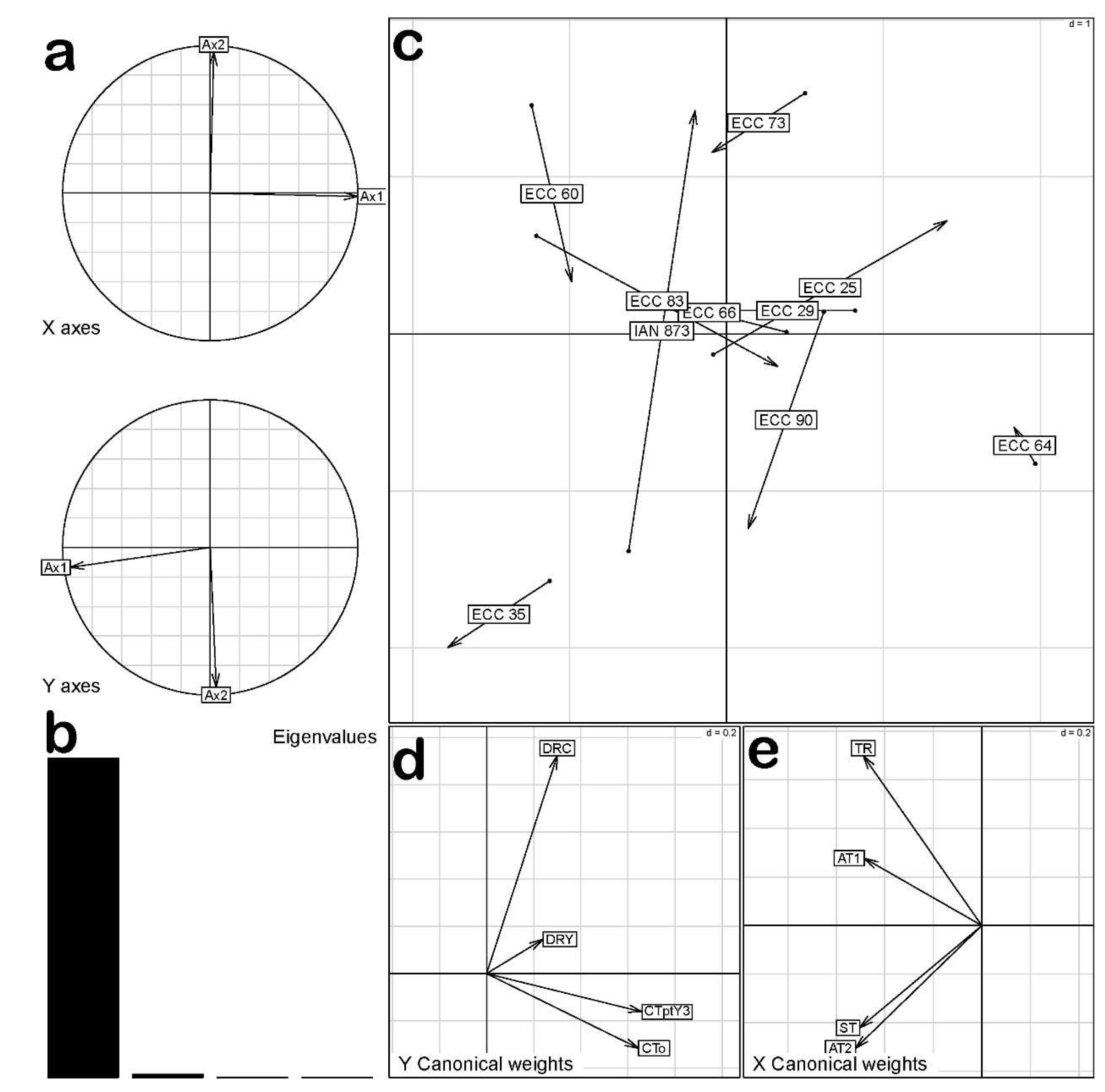

Table 1.
Selection criteria for the longitudinal data analysis model for assessing of the growth and resistance to SALB.
Table 1.
Selection criteria for the longitudinal data analysis model for assessing of the growth and resistance to SALB.
| Model | Compound symmetric model with heterogeneous variances (CSH) | First order autoregressive model with heterogeneous variances (ARH1) | First order continuous autoregressive model with heterogeneous variances (CARH1) | Unstructured model with heterogeneous variances (UNH) | |||||||||||
| Variable | log Lik | AIC | BIC | log Lik | AIC | BIC | log Lik | AIC | BIC | log Lik | AIC | BIC | |||
| CTpt | -175.38 | 420.77 | 508.26 | -170.39 | 410.77 | 498.27 | -170.39 | 410.77 | 498.27 | -165.18 | 404.35 | 496.84 | |||
| AT1 | 159.78 | -235.6 | -130.6 | 159.97 | -236 | -131 | 159.97 | -235.95 | -131 | 161.4 | -234.8 | -124.81 | |||
| AT2 | 63.18 | -56.36 | 31.13 | 63.29 | -56.59 | 30.91 | 63.29 | -56.59 | 30.91 | 64.22 | -54.44 | 38.06 | |||
| ST | 48.8 | -27.59 | 59.9 | 48.68 | -27.37 | 60.13 | 48.68 | -27.36 | 60.13 | 49.65 | -25.3 | 67.2 | |||
| TR | 97.9 | -125.8 | -38.31 | 97.1 | -124.2 | -36.7 | 97.1 | -124.19 | -36.7 | 101.28 | -128.57 | -36.07 | |||
| log Lik, Log-likelihood; AIC, Akaike criterion; BIC, Bayesian criterion (Verbeke and Molenberghs, 2000). | |||||||||||||||
| CTpt, circumference of the trunk in the pre-tapping phase; AT1, attack severity on young leaves; AT2, attack severity on mature leaves; TR, type of reaction on young leaves (conidial sporulation intensity, TR); ST, stroma density on mature leaves [16,20,44]. | |||||||||||||||
Table 2.
Analysis of variance (ANOVA) in growth, SALB resistance and early yield for effects genotype, year and genotype*year interaction.
Table 2.
Analysis of variance (ANOVA) in growth, SALB resistance and early yield for effects genotype, year and genotype*year interaction.
| Variable | DF1a | DF2b | DF3c | Genotype | Year | Genotype x Year |
| CTpt | 9 | 2 | 18 | 4.42 (0.0001) | 714.72 (<0.0001) | 2.23 (0.0074) |
| AT1 | 9 | 2 | 18 | 5.35 (<0.0001) | 89.39 (<0.0001) | 2.98 (0.0004) |
| AT2 | 9 | 2 | 18 | 16.19 (<0.0001) | 256.90 (<0.0001) | 2.68 (0.0012) |
| ST | 9 | 2 | 18 | 19.30 (<0.0001) | 215.16 (<0.0001) | 3.08 (0.0002) |
| TR | 9 | 2 | 18 | 3.50 (0.0010) | 116.21 (<0.0001) | 2.09 (0.0125) |
| DRC | 9 | - | - | 10.14 (<0.0001) | - | - |
| DRY | 9 | - | - | 18.85 (<0.0001) | - | - |
| CTo | 9 | - | - | 5.69 (<0.0001) | - | - |
| CTpt, circumference of the trunk in the pre-tapping phase; AT1, attack severity on young leaves; AT2, attack severity on mature leaves; TR, type of reaction on young leaves (conidial sporulation intensity, TR); ST, stroma density on mature leaves [16,20,44]. DRC dry rubber content, DRY dry rubber yield, CTo circumference of the trunk at opening [16,55] | ||||||
| DF1a Degrees of freedom for the factor genotype. | ||||||
| DF2b Degrees of freedom for the factor year. | ||||||
| DF3c Degrees of freedom for interaction genotype x year.-Not applicable | ||||||
Table 3.
Annual circumference of the trunk (cm) in the pre-tapping phase (CTpt) for 10 genotypes of Hevea brasiliensis.
Table 3.
Annual circumference of the trunk (cm) in the pre-tapping phase (CTpt) for 10 genotypes of Hevea brasiliensis.
| Genotype | Year 1 | Year 2 | Year 3 |
| ECC 25 | 9.37 ± 0,49akb | 18.33 ± 0.88def | 25.75 ± 1.24ab |
| ECC 29 | 9.43 ± 0,49k | 16.98 ± 0.88fgh | 22.80 ± 1.24bc |
| ECC 35 | 7.78 ± 0.49l | 13.33 ± 0.88i | 20.23 ± 1.24cde |
| ECC 60 | 9.03 ± 0.49k | 14.89 ± 0.88hi | 21.44 ± 1.24c |
| ECC 64 | 10.69 ± 0.49j | 20.44 ± 0.88cd | 27.48 ± 1. 24a |
| ECC 66 | 8.73 ± 0.49kl | 15.31 ± 0.88ghi | 22.54 ± 1.24bc |
| ECC 73 | 9.44 ± 0.49k | 16.66 ± 0.88fgh | 22.84 ± 1.24bc |
| ECC 83 | 9.62 ± 0.49 jk | 17.31 ± 0.88efg | 23.61 ± 1.24bc |
| ECC 90 | 9.02 ± 0.49k | 15.83 ± 0.88gh | 23.63 ± 1.24bc |
| IAN 873 | 9.82 ± 0.49jk | 16.28 ± 0.88fgh | 22.41 ± 1.24bc |
| a Standard error. | |||
| b Means in each column with the same letter do not differ statistically (Fisher’s LSD test, p < 0.05). | |||
Table 4.
Annual circumference of the trunk (cm) in the opening phase (CTo), and yield-related traits in the early tapping phase for 10 genotypes of Hevea brasiliensis.
Table 4.
Annual circumference of the trunk (cm) in the opening phase (CTo), and yield-related traits in the early tapping phase for 10 genotypes of Hevea brasiliensis.
| Genotype | CTo (cm) | DRY (g.tree−1 .tap−1) | DRC (%) |
| ECC 25 | 26.86 ± 0. 35abb | 5.57 ± 0. 43a | 32.45 ± 0. 44a |
| ECC 29 | 26.49 ± 0. 37b | 0.74 ± 0.18c | 31.44 ± 2. 29a |
| ECC 35 | 25.00 ± 2. 45b | 0.42 ± 1.14c | 26.24 ± 0.76b |
| ECC 60 | 25.03 ± 0.93b | 1.21 ± 0.43c | 30.43 ± 0.70ab |
| ECC 64 | 27.92 ± 0. 26a | 3.76 ± 0.66b | 31.38 ± 1.01a |
| ECC 66 | 25.54 ± 0. 78b | 1.09 ± 0.43c | 30.75 ± 1. 10a |
| ECC 73 | 25.62 ± 0. 56b | 2.23 ± 0.43bc | 32.42 ± 1. 18a |
| ECC 83 | 26.09 ± 0. 45b | 3.30 ± 0.24b | 30.53 ± 1.98ab |
| ECC 90 | 26.94 ± 0. 57b | 2.96 ± 0.81bc | 28.83 ± 3.05ab |
| IAN 873 | 25.47 ± 0. 60b | 1.88 ± 0.28c | 32.83 ± 0. 10a |
| CTo circumference of the trunk at opening (time of commencement of tapping or 3th year), DRY dry rubber yield, DRC dry rubber content (all in the early tapping). a Standard error. b Means in each column with the same letter do not differ statistically (Fisher’s LSD test, p < 0.05) | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



