Submitted:
19 September 2023
Posted:
20 September 2023
You are already at the latest version
Abstract
No-till and cereal-legume intercropping have been recognized as favorable cropping practices to increase crop yields while maintaining soil quality in arid and semiarid environment, but the bio-logical mechanisms are poorly understood. The present study was to determine the response of soil properties, enzyme activities, and microbial community diversity and composition in mono- and inter-cropping under conventional and no-tillage conditions. We initiated a field experiment in Wuwei, a typical arid area of China, in 2014. Soil was sampled in August 2022 and, yields, soil properties, enzyme activities, and the microbial community diversity and composition were de-termined in the maize and pea strips in inter- and mono-cropping systems. Results revealed that the maize and pea strips in the no-till intercropping significantly increased yields, total and or-ganic carbon stocks, decreased NO3--N, and obtained the highest total and organic P in the soil. The α- and β-diversity of archaea and eukaryotes were significantly affected by planting patterns, while α- and β-diversity of the bacterial community were significantly affected by tillage practices. Both no-tillage and intercropped maize significantly increased the abundance of archaea phylum Thaumarchaeota and bacterial phylum Nitrospirae, benefiting nitrogen fixation of intercropped pea from the atmosphere under the no-tillage cereal/legume intercropping. No-till intercropping was conducive to the accumulation of organic carbon, while decreasing the abundance of Prote-obacteria, Acidobacteria, and Verrucomicrobia. Limited soil enzyme activities (ACP, ALP, DP, NAG, BG, AG, CB) led to decreases in organic carbon turnover and utilization. Intercropping al-tered soil microbial community diversity and composition due to changes in soil properties and enzyme activities. These findings suggest that no-tilled cereal-legume intercropping is a sustaina-ble cropping practice for improving soil properties and enhancing microbial (archaea, bacterial, Eukaryota) diversity, but the long-term persistence is not conducive to rapid turnover of soil nu-trients due to limited enzyme activities.
Keywords:
no-tillage
; intercropping
; microbial community
; soil properties
; soil enzyme activities
1. Introduction
In many parts of the world, intensified agricultural production has been adopted as a key strategy to increase food production since ‘Green Revolution’, but this has also caused soil micro-ecological environment damage [1], reducing soil productivity [2], and increasing greenhouse gas emissions [3] that contributes to the climate extremes [4]. In recent years, cereal-legume intercropping has shown the great potential to increase grain production while benefiting soil and environment in modern agricultural systems [1,5]. In the arid irrigated areas of northwestern China where annual total solar radiation is abundant but water resource is scarce [6,7], intercropping maize with pea (Figure 1) has shown significant benefits to food security through a sustainable production system. With the emphasis on the ecological environment, no-tillage has been shown greater potential for soil carbon sequestration, the improvement of soil quality [8,9,10,11], and the sustainability of crop productivity, compared to conventional tillage [9,10,12,13]. No-tillage is shown to decrease soil pH, but increase soil bulk density, mean weight diameter, water-stable aggregate, the soil enzyme activity of β-glucosidase, acid phosphatase, and urease [12,13]. Additionally, no-till with plastic mulching has been increasingly used to lower soil CO2 emissions, lower labor inputs, and decrease soil disturbance along with the inhabitation of soil microbial community functions over traditional systems [14,15,16].
No-tillage in association with intercropping has been adopted worldwide as a promising approach to increase food production while improving soil quality. In the semiarid region of Brazil, intercropping with minimum tillage was used as an alternative form of soil management to maintain or increase the SOC fractions [17]. In the Northern Great Plains of North America, no-till with cropping diversification has been considered a sustainable practice to robust the system’s productivity and resilience [18,19], while reducing environmental footprints [1,3,20]. In the northernmost part of South Africa, the concentrations of organic C, P and K, and C: N ratio were found significantly higher in no-tilled sorghum/cowpea intercropping than in conventional tillage [21]. Intercropping combined with conservation practices makes the distribution and penetration of the residual roots into the soil, contributing to the improvement of the soil physical properties and soil structure [22], due to improved C sequestration and storage [23,24] and reduced CO2 emissions [25]. The effect of no-tillage intercropping on soil physicochemical properties has been well documented, however, the biological characteristics and the mechanisms of microbial involvement are less researched, especially in the northwest arid area of China where intercropping is widely practiced.
Soil microbial activities play an important role in nutrient mobilization and mineralization which are essential for plant growth [11,26,27,28], with nutrients and water sharing among crops commonly detected in legume–cereal intercropping [29,30]. Studies have shown that intercropping can significantly alter the composition and structure of functional microbial communities to reduce CO2 emissions [7] Bacterial community composition and diversity reflect changes in soil properties and enzyme activities better than those of fungi in the intercropping system [22], due to changes in microbes shifting soil properties, like soil pH, nutrient contents, and enzyme activities [31]. Moreover, no-tillage changes the metabolic capabilities of soil microbial communities by altering the soil physiochemical properties, offering significant benefits in conserving soil and water and increasing SOC [32,33]. However, the microbiological mechanisms of the effect of no-till intercropping on nutrient cycling and microbial community attributes are poorly understood. The objective of the present study was to determine the soil properties, enzyme activities, and the microbial community diversity and composition in response to monoculture and intercropping in long-term conventional and no-tillage systems in a semi-arid area.
2. Materials and Methods
2.1. Study site
The experiment was conducted since 2014 at the Agricultural Research and Education Station of Gansu Agricultural University in Wuwei (37°56′ N, 102°38′ E, altitude 1,520 m), Gansu, China. The mean annual temperature is 7.2℃, the accumulated>0℃ temperature is 3513℃ and>10℃ is 2985℃, the frost-free period is 156 days. The mean annual precipitation is 155 mm, and evaporation is greater than 2400 mm. The soil type is sandy loam Aridisols at the experiment site.
2.2. Experimental design
The long-term field trial was laid out in randomized blocks in a split-plot design with the main plots being two tillage practices: conventional tillage (CT) and no-tillage (NT), and the split plots being three planting patterns: maize (Zea mays L.) monoculture (M), pea (Pisum sativum L.) monoculture (P), and maize intercropping pea (M/P). Each treatment was replicated in four blocks. Each plot was 7 m long and 9 m wide. Plastic film mulching was applied to each plot at sowing. In CT plots, plastic film was removed at crop harvest before the plot was plowed. New film was applied to the plot at the following spring sowing time. In NT plots, the mulching film was applied at sowing and was kept in the field after crop was harvested for reuse the following season(s). For both the sole and intercrops, the planting densities of sole maize and sole pea were 90,000 and 800,000 plants hm-2, with a row spacing of 40 and 20 cm, respectively. In the M/P planting pattern, the M and P was in a 4:3 ratio for the area sharing, with planting density of 52,000 and 760,000 plants hm-2, respectively. The maize variety was ‘Xianyu 335′, and the pea variety was ‘Longpea 1′. In each growing season, plots were irrigated with 465 mm ha-1 year-1. The total N application rate was 360 kg N ha-1 for maize and 135 kg N ha-1 for pea. Phosphorus fertilizer was applied as the base fertilizer following a N:P ratio of 2:1.
2.3. Soil sampling and soil physicochemical properties measurement
Soils were sampled in the sole maize (M), sole pea (P), intercropped maize (IM), and intercropped pea (IP) system from conventional tillage (CT) and no-tillage (NT) treatments, with a total of 32 samples (4 planting patterns x 2 tillage x 4 replicates). In each plot, five bulk soil cores were taken between the rows to the 0-30 cm soil depth using a soil auger (5 cm in diameter) on 20th August 2022. The five soil cores per plot were homogenized into a composite sample for each treatment, and subsequently sieved through a 2 mm mesh to remove plant residues and debris. The composite soil sample was divided into three parts: one was immediately stored on ice and transported to a laboratory for DNA extraction within 24 h post sampling, one was kept fresh for the determination of soil microbial biomass, enzyme activities, and the third one was air-dried for the determination of soil moisture, pH, electrical conductivity, total carbon (TC), organic carbon (OC), and inorganic carbon (IC), and total, organic and inorganic nitrogen (TN, ON, and IN, respectively) in the facilities of the State Key Laboratory of aridland crop science, Gansu Agricultural University. The activities of extracellular enzymes, included acid phosphatase (ACP), alkaline phosphatase (ALP), phosphodiesterase (DP), N-acetyl-β-glucosaminidase (NAG), cellobiohydrolase (CB), and β-glucosidase (BG), α-glucosidase (AG), and β-xylosidase (BX) were measured using 4-methylumbelliferone-linked fluorogenic substrates in modified universal buffer with appropriate pH value [34].
2.4. Soil DNA extraction and metagenomics sequencing
Total raw DNA was extracted from 0.5 g of fresh soil using the PowerSoil DNA Isolation Kit (Mo Bio Laboratories, Carlsbad, CA, USA). The concentration and purity of the raw DNA samples were tested with a 1% agarose gel (1% AGE, 100 V/40 min), and qualified samples were stored at -80℃ for further analysis. Metagenomic libraries were size-selected to fragment lengths of 350 bp. The fragment length range was determined using Agilent 2100 High Sensitivity DNA Assay with Agilent 2100 Bioanalyzer (Agilent Technologies, Palo Alto, CA). Q-PCR method was used to quantify the effective concentration of the library (effective concentration > 3nM) to ensure the quality of the library. Totally, 32 metagenomic DNA libraries were generated. The size-selected libraries were sequenced using 2 x 150 bp paired-end chemistry (PE150) on an Illumina NovaSeq 6000 platform (Novogene Bioinformatics Technology Co. Ltd, Beijing, China). The raw sequencing data were submitted to NCBI Sequence Read Archive (SRA) under bioproject accession number PRJNA1017654.
2.5. Metagenomic assembly
The raw reads were quality trimmed by removing adapters (overlap ≥15bp)), ambiguous reads (“N” ≥ 10%), and low-quality reads (quality value ≤ 40) using trimmomatic [35]. The high-quality paired-end reads were assembled into contigs using MEGAHIT (version 1.1.3) [36]. The quality of contigs was assessed using QUAST. Basic information statistics of Scaftigs of each sample assembly result (≥ 500bp) showed in Table S1. DIAMOND software was used to search predicted genes against from the National Center for Biotechnology Information (NCBI) non-redundant protein database using BLASTP (best hit with e-value cutoff of 1e-5) [37].
2.6. Statistical and bioinformatics analysis
Statistical analyses were conducted using R version 4.2.3. For α-diversity determination, the Shannon index was calculated using the ‘vegan’ package in R [38], and species evenness was calculated using Pielou evenness [39]. The effects of different treatments on soil microbial diversity was determined using permutational multivariate ANOVA (PermANOVA) with the “lme4” and “lmerTest” packages in R. Principal component analysis (PCA) and adonis analysis based on Bray-Curtis dissimilarity matrices were to test the effects of treatments on the microbial community using the ‘vegan’, ‘pairwise’, ‘ade4′ and ‘ggplot2′ packages in R [40]. The microbial co-occurrence networks analysis were performed in Gephi (v. 9.3) [41]. The linear discriminant analysis (LDA) effect size (LEfSe) including the distribution of LDA scores of different species (tax with score > 4) and the cladogram [42]. Spearman correlation analysis and heatmaps of correlation were used to determine the relationships among net area yield, soil properties, and taxonomic and functional diversity using the ‘linkET’, ‘vegan’, and ‘dplyr’ packages in R [43]. In addition, spearman correlation analysis and the heatmaps of correlation were used to determine the relationships between yield, soil properties, and microbial communities using the function ‘pheatmap’ in R.
3. Results
3.1. Yields, soil physicochemical characters and soil enzyme activities
No-tillage improved the number of 1000-seed weight and grain yields of maize and pea in inter-and mono-cropping compared with conventional tillage (Table 1). Maize and pea intercropping increased the number of grains per pod, 1000-seed weight, grain yields, and net area yields whatever in the no-tillage or conventional tillage. The highest yields observed in maize and pea intercropping under the no-tillage condition (Table 1).
No-tillage increased soil pH, TC, OC, TN, NH4+-N, and OP significantly, while decreasing soil EC and NO3--N as compared to conventional tillage (Table 2). There was a significant tillage by planting-pattern interaction in affecting soil physiochemical properties, with the NTIM and NTIP treatments increasing TC and OC and decreasing NO3--N as compared to the CTIM and CTIP treatments. Additionally, the NTIM treatment increased soil TP and OP but decreased AP as compared to CTIM.
Planting patterns affected soil enzyme activities significantly (Table 3). Compared with the maize or pea monocropping, maize-pea intercropping decreased DP, NAG, AG, and CB activities. The BX activity was the lowest in the maize strip of the M/P intercropping, whereas the ALP and BG activities were the lowest in the NTIM systems.
3.2. Diversity and structure of soil taxonomic microbial community
Species annotation results showed that the maize-pea intercropping improved the α-diversity (Shannon index and Pielou evenness) of archaea and eukaryote communities at the phylum level as compared to the corresponding monocropping (Table S1). Also, no-tillage significantly enhanced the Shannon index and Pielou evenness of the bacterial community over conventional tillage, with the α-diversity of the bacterial community distinctly higher in the NTIM treatment than in the CTIM treatment. Moreover, planting pattern significantly affected the β-diversity of archaea (Figure 2a), bacterial (Figure 2b) and eukaryotes (Figure 2c) communities, while tillage practices significantly affected the β-diversity of bacterial community (Figure 2b). In the diversity distribution patterns, intercropped maize and pea had a distinct separation from the corresponding monocrops (Figure 2a–c). The co-occurrence networks analysis showed that an increasing trend of taxonomic complexity from the CT (nodes = 131; total linkages = 490) to NT (nodes = 138; total linkages = 573) (Figure 2d,e). The NT network exhibited a higher degree of modularity (0.569) and had more negative (linkakes = 193) and positive (linkages = 380) associations than the NB network (modularity = 0.860; positive linkages = 358, negative linkages = 132) (Figure 2d,e).
The analysis of microbial community structure at the phyla level showed that the main archaea phyla were p_Thaumarchaeota (94%), eukaryote phyla were p_Ascomycota (56%), p_Basidiomycota, p_Chytridiomycota, p_Mucoromycota and there were p_Proteobacteria (42%), p_Acidobacteria, p_Actinobacteria, p_Gemmatimonadetes, p_Candidatus Rokubacteria, p_Chloroflexi, p_Bacteroidetes, p_Nitrospirae, p_Planctomycetes, p_Cyanobacteria, p_Verrucomicrobia, p_Firmicutes for bacteria (Figure 3b). No-tillage significantly increased the archaea p_Thaumarchaeota and bacterial p_Chloroflexi, and p_Nitrospirae, while significantly decreased bacterial p_Proteobacteria, p_Gemmatimonadetes, and p_Firmicutes compared with conventional tillage (Tables S2 and S4). The abundance of eukaryote p_Mucoromycota and the bacterial p_Proteobacteria, p_Acidobacteria were significantly higher in maize treatments than in pea treatments in both intercropping and monocropping under the long-term conventional and no-tillage conditions (Tables S2–S4). Moreover, the maize strips of intercropping significantly enhanced the abundance of archaea p_Thaumarchaeota, and maize and pea strips of the intercropping significantly increased the abundance of p_Nitrospirae compared to monocropping (Figure 3b, Tables S2 and S4).
Additionally, the cladogram (Figure 3a) and LDA scores of different species (Figure 3c) showed that the abundance of the unclassified species differed substantially between treatments at the class, order, and family levels. the abundance of the unclassified_Actinobacteria in the NTIM treatment; p_Candidatus Rokubacteria, p_Acidobacteria, o_unclassified_Betaproteobateria, o_Chitinophagales, o_unclassified_bacteria in the NTM treatment; o_Nitrospirales in the NTIP treatment; p_Thaumarchaeota, p_Nitrososphaerota_o_Nitrososphaerales, p_Actinomycetes, p_Chloroflexi, p_Actinomycetot, g_Kocuria in the NTP treatment; p_Proteobacteria for CTIM, and g_Uncassified_Deltaproteobacteria in the CTIP treatment; Unclasified_Gemmatimonadetes at class, order, family and genus levels, g_Sphingomonas, s_Sphingomonas_jaspsi, s_Sphingomonas_sp_URHD0057, f_Nocardioidaceae, g_Nocardioides in the CTP treatment were significantly different (Figure 3a,c).
3.3. Soil properties drive the soil microbial community difference in inter/mono-cropping systems under conventional and no-tillage
The α-diversity of the archaea community was significantly related to yield, soil IC, OC, and NH4+-N, while the α-diversity of the eukaryote community was significantly related to soil IC and SW, NAG (Figure 4a). Soil pH and yield was significantly negatively related to the soil EC and ALP; OC was significantly positively related with TC, TN, and NH4+-N, while significantly negatively related with IC; SW, NO3--N. The C:N ratio in the soil was significantly negatively related to the soil enzyme activities (Figure 4b).
Yield and soil pH were mainly correlated to eukaryote phyla Mucoromycota, and bacterial phyla Bacteroidetes (Figure 5). TC, OC and NH4+-N were negatively related to bacterial phyla Proteobacteria, Acidobacteria, Bacteroidetes, Frimicutes, Verrucomicrobia, Candidate division NC10, Candidate division Zixibacteria, Elusimicrobia, Candidatus Omnitrophica, Candidatus Dadabacteria, Candidatus Glassbacteria, and Rhodothermaeota, while were positively related to Actinobacteria and Nitrospirae (Figure 5). Additionally, the heatmap showed the soil enzyme activities were positively related to bacterial phyla (proteobacteria, acidobacteria, verrucomicrobia, Candidate division Zixibacteria), while it was significantly negatively related to the archaea phyla (Candidatus Bathyarchaeota, Candidatus Diapherotrites, Candidatus Thorarchaeota, Crenarchaeota, Thaumarchaeota) and eukaryote phyla (Basidiomycota, Mucoromycota, Zoopagomycota) and bacterial phyla (Actinobacteria, Chlorofiexi, Nitrospirae, Deinococcus_Thermus).
4. Discussion
4.1. Yields, and soil properties in response to no-till intercropping system
Both of no-tillage and intercropping improved the grain yields of maize and pea, and higher yields of maize and pea resulted in the highest total yield observed in M/P treatments under the no-tillage condition in this study. As reported that the intercropped maize and pea strips under the no-tillage with plastic mulch were as effective as conventional tillage with plastic mulch to accumulate dry matter and achieve a favorable yield outcome [44]. Soil physiochemical properties are commonly considered the indicators of soil quality, capable of sustaining plant productivity and maintaining soil water and fertility. In this study, no-tillage significantly increased soil pH, TC, OC, TN, NH4+-N, and OP, while decreasing soil EC and NO3--N. These results disagree with the decreased soil pH observed in some other no-tillage related research [13]. Some studies show that no-tillage, as compared to conventional tillage, increased the activity of β-glucosidase, acid phosphatase, and urease [12], but these effects were not observed in our study. The differences are probably due to various reasons but we suggest that crop fertilization and history of soil tillage may have played a role in influencing enzyme activities. Our results showed that intercropping decreased the DP, NAG, AG, and CB activities compared with the monocropping system. These enzymes are the direct expression of the microorganism in microbial metabolic requirements and utilizing available nutrients [45]. However, intercropping system did not reduce soil nutrients and, in fact, improved soil NH4+-N and NO3--N. The decrease in enzyme activities may be due to the increase of SW, NH4+-N, NO3--N, and the C:N ratio in our study. Also, we found that TC and OC were significantly higher in the NTIM and NTIP treatments than in the CTIM and CTIP treatments; this may be due to the distribution and penetration of the plant roots into the soil profile under no-till management, contributing to the improvement of the soil physical properties and soil structure [17], especially for C sequestration [23,24]. Our results suggest that the intercropping systems with minimum soil tillage are able to maintain or increase SOC stocks, in agreement with previous reports [22]. Also, from the perspective of the intricate microbially mediated soil C cycling, soil extracellular enzymes catalyse maythe may have limited SOM decomposition [46]. Additionally, the lowered AP in the NTIM treatment may be due to the losses of particulate P and dissolved P with NT as compared to CT [47]. The highest soil TP and OP in the NTIM treatment was, which correlation to higher yield, from the perspective of the low efficiency of organic P mineralization, probably due to the decrease in available phosphorus. The ALP and BX were restricted in the maize strip of M/P intercropping system under the no-tillage condition in our study. Also, alkaline phosphatase plays an important role in influencing organophosphorus components in alkaline soil due to the association of organophosphorus with soil microorganisms [48].
4.2. The diversity of soil microbial community in response to no-till intercropping system
In this study, planting patterns significantly affected the β-diversity of archaea, eukaryote, and bacterial community, while tillage practices significantly affected the β-diversity of bacterial community. Others have found that no-tillage increased soil bacterial diversity without significant changes in fungal diversity [49]. At the same time, our study showed no-tillage significantly enhanced the Shannon index and Pielou evenness of bacterial community, exhibited more complex co-occurrence networks, and showed distinct higher bacterial community α-diversity than conventional tillage, especially in the maize strips of intercropping system. Such diversity indicates the steadiness and adaptability of microbial communities against soil disturbances, which are important for maintaining ecosystem functioning [50]. Moreover, the Shannon index and Pielou evenness of soil archaea and eukaryote communities in the intercropped pea and maize strips were improved compared with monocropping system, benefiting ecosystem’s functions and services [51]. In particular, the α-diversity of the archaea community was significantly related to yield, soil IC, OC, and NH4+-N, while the α-diversity of the eukaryote community was significantly related to soil IC and SW, NAG. Significant, positive effects of SOC on bacterial alpha diversity is reported by others [22]. Collectively, our results revealed that soil nutrient cycling processes were driven by microbial community diversity and compositions, and the magnitude of the effect varied planting patterns with the M/P intercropping influencing the cycling intensity more significantly than the monoculture due to enzyme activities.
4.3. The compositions of soil microbial community in response to no-till intercropping system
The abundance of eukaryote Mucoromycota and bacterial Proteobacteria, Cidobacteria were significantly higher in maize than in pea fields whether intercropping or monocropping under the long-term conventional and no-tillage conditions, which positively related to yield. It is reported that maize and pea strips in intercropping and monocropping differ in microbial communities [7] because of soil microbial activities that play an important role in nutrient mobilization and mineralization [29]. TC, OC, and NH4+-N were positively related to Actinobacteria and Nitrospirae, while were negatively related to Proteobacteria and Acidobacteria; this was probably due to no-tilled intercropping promoting underground interspecies interactions [13,52].
No-tillage significantly increased archaea Thaumarchaeota and bacterial Chloroflexi and Nitrospirae, while significantly decreased bacterial Proteobacteria, Gemmatimonadetes, and Firmicutes compared with conventional tillage, which was attributable to the higher soil OC and NH4+-N in the no-tillage. Cai, et al. [53] showed that SOC had a significant positive correlation with the relative abundance of Chloroflexi, in agreement with our results, but OC was negatively related to Proteobacteria and Acidobacteria in our study. Xu, et. al. [7] indicated the mitigation of CO2 emissions by suppressing the growth of Proteobacteria and Bacteroidota but promoting the growth of Chloroflexi. According to previous research, archaea Thaumarchaeota contain a complete pathway for carbon fixation, and a member of Cenarchaeum symbiosome is regarded as the most energetically efficient carbon fixation pathway [54,55,56], and their activity is closely related to N2O production [57,58]. Proteobacteria is a large bacterial phylum with many classes containing bacteria of various ecological types and nutritional characteristics [59]. The adaptability of Gemmatimonadetes to the soil environment was related to the carbon and nitrogen environment [60]. Firmicutes were significantly different in the proportion of conventional tillage (33%) and no-tillage (6%), respectively [61]. A meta-analysis synthesized that no-tillage had little effect on Proteobacteria, Chloroflex, Firmicute, and Bacteroides, but significantly increased the abundance of Acidobacteria and decreased Actinobacteria [49]. Also, it is possible that the changes in the abundance of bacterial communities between treatments differed substantially leading to inconsistent or even opposite results where soil sampling was taken at the different stages [7,62]. Therefore, the mechanisms underlying the maintenance of biodiversity and functional changes of these microbes in the agro-ecosystems’ functioning need further investigation.
Intercropping altered soil microbial community diversity and composition, with soil bacteria reflecting the changes in yield, soil properties and enzyme activities more significant than archaea and Eukaryota in our study. Studies have confirmed that bacterial composition can be stimulated by enzyme activities and soil properties with considerable change under cereal-legume intercropping systems [22,63]. Furthermore, we found that intercropping maize significantly enhanced the abundance of Thaumarchaeota and Actinobacteria. Bao, et al. [64] provided evidence that non-dominant Actinobacteria played a vital ecophysiological role in plant residue decomposition, which were more abundant in soils than other media, especially in alkaline soils and soils rich in organic matter, where they constitute an important part of the microbial population [54]. The distribution and penetration of the plant roots into the soil provide the C source [24], which can be absorbed and utilized by Actinobacteria [23]. Additionally, intercropping maize and pea significantly increased the abundance of Nitrospirae with higher yield compared to monocropping maize and pea. Moreover, ammonia-oxidizing archaea (AOA) and ammonia-oxidizing bacteria (AOB) co-participate in the ammonia oxidation process, which is the first step and a key rate-limiting step in the nitrification process [65]. The members of the archaeal Thaumarchaeota and bacteria (g_Nitrosomonas, g_Nitrobacter or g_Nitrospira belong to p_Nitrospirae) are ammonia oxidizers [66]. There are evidences that nitrogen fixation in the cereal/legume intercropping system provided by intercropped legumes promote nutrient sharing between cereal and legume crops [5].
Intercropping maize significantly decreased the abundance of Proteobacteria, Acidobacteria, and Verrucomicrobia, especially in no-tillage, due to the positive effect on soil enzyme activities (ACP, ALP, DP, NAG, BG, AG, CB). This was partly due to higher accumulation of SOC contributed by the increased input of vegetation production (quantity and quality) by intercrops. Rumpel and Kögel-Knabner [24] found that root exfoliations contained a certain number of difficult-to-degrade components, such as lignin and polyphenols, which accounted for a considerable proportion of SOC stocks. Among these, SOM has certain protection and stability mechanisms affected by biological and abiotic factors [67,68]. Soil aggregates can physically protect organic carbon from decomposition by affecting microbial community structure and limiting oxygen diffusion and nutrient circulation [69]. We found that no-till intercropping system was highly conducive to the accumulation of organic carbon, however, lacking the biological turnover and utilization of organic carbon; this was probably attributable to enhanced soil aggregates (which was not measured in our study). No-till practices also lower CO2 emissions from the soil due to minimum soil disturbance inhabiting soil microbial community activities as compared to traditional tillage practices [14,15,16].
5. Conclusions
The no-tilled, maize and pea strip intercropping system (i.e., the NTIM treatment) significantly increased yield, TC and OC, decreased NO3--N, and achieved the highest soil TP and OP among the treatments evaluated. These changes were significantly correlated with soil microbial community diversities, compositions, and soil enzyme activities. The α- and β-diversity of archaea and eukaryotes were significantly affected by planting patterns, while α- and β-diversity of the bacterial community were significantly affected by tillage practices. Soil properties and enzyme activities affected soil bacterial community characteristics more significantly than affecting archaea and eukaryote communities under the long-term no-tillage condition. Both no-tillage and intercropping maize significantly increased the abundance of archaea Thaumarchaeota and bacterial Nitrospirae, benefiting biological nitrogen fixation from the atmosphere for the no-tilled cereal/legume intercropping system. No-till intercropping system is conducive to the accumulation of organic carbon, while decreasing the abundance of Proteobacteria, Acidobacteria, and Verrucomicrobia significantly. Lack of soil enzyme activities (ACP, ALP, DP, NAG, BG, AG, CB) led to reduced biological turnover and organic carbon utilization.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: taxonomic alpha diversity based on absolute abundance homogenization matrix at phylum levels in the different treatments; Table S2: the abundance of archaea communities at the phylum level; Table S3: the abundance of Eukaryote communities at the phylum level; Table S4: the abundance of bacterial communities at the phylum level.
Author Contributions
PL contributed to investigation, performing experiments, methodology, statistical analysis, writing of the original draft. CZ contributed to investigation, performing experiments, resources, funding acquisition, supervision, and writing, reviewing, and editing of the manuscript. WY, HF, FZ, AY, FH contributed to reviewing and editing of the manuscript. All authors contributed to writing the paper and approved the final version of the manuscript.
Funding
This research was funded by the Gansu Province Natural Science Foundation key project (21JR7RA802), the National Natural Science Foundation of China (31860363), and the key talent project of Gansu Province, the Scientific Research Startup Funds for Openly-recruited Doctors of Gansu Agricultural University (GAU-KYQD-2021-10), the Research Program Sponsored by the State Key Laboratory of Aridland Crop Science of China (GSCS-2023-11).
Data Availability Statement
The original contributions presented in the study are included in the article.
Acknowledgments
We would like to thank Joann, Whalen for her constructive comments on the results of the study.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- Liu, C.; Plaza-Bonilla, D.; Coulter, J. A.; Kutcher, H. R.; Beckie, H. J.; Wang, L.; Floc’h, J.-B.; Hamel, C.; Siddique, K. H. M.; Li, L.; Gan, Y. Diversifying Crop Rotations Enhances Agroecosystem Services and Resilience. Advances in Agronomy 2022, 173, 299–335. [Google Scholar] [CrossRef]
- Zhang, X.; Dong, Z.; Wu, X.; Gan, Y.; Chen, X.; Xia, H.; Kamran, M.; Jia, Z.; Han, Q.; Shayakhmetova, A.; Siddique, K. H. M. Matching Fertilization with Water Availability Enhances Maize Productivity and Water Use Efficiency in a Semi-Arid Area: Mechanisms and Solutions. Soil and Tillage Research 2021, 214, 105164. [Google Scholar] [CrossRef]
- Liu, C.; Cutforth, H.; Chai, Q.; Gan, Y. Farming Tactics to Reduce the Carbon Footprint of Crop Cultivation in Semiarid Areas. A Review. Agronomy for Sustainable Development 2016, 36, 1–16. [Google Scholar] [CrossRef]
- Burney, J. A.; Davis, S. J.; Lobell, D. B. Greenhouse Gas Mitigation by Agricultural Intensification. Proceedings of the National Academy of Sciences of the United States of America 2010, 107, 12052–12057. [Google Scholar] [CrossRef] [PubMed]
- Chai, Q.; Nemecek, T.; Liang, C.; Zhao, C.; Yu, A.; Coulter, J. A.; Wang, Y.; Hu, F.; Wang, L.; Siddique, K. H. M.; Gan, Y. Integrated Farming with Intercropping Increases Food Production While Reducing Environmental Footprint. Proc Natl Acad Sci U S A 2021, 118, e2106382118. [Google Scholar] [CrossRef] [PubMed]
- Gou, Z.; Yin, W.; Asibi, A. E.; Fan, Z.; Chai, Q.; Cao, W. Improving the Sustainability of Cropping Systems Via Diversified Planting in Arid Irrigation Areas. Agronomy for Sustainable Development 2022, 42, 88. [Google Scholar] [CrossRef]
- Xu, K.; Hu, F.; Fan, Z.; Yin, W.; Niu, Y.; Wang, Q.; Chai, Q. Delayed Application of N Fertilizer Mitigates the Carbon Emissions of Pea/Maize Intercropping Via Altering Soil Microbial Diversity. Front Microbiol 2022, 13, 1002009. [Google Scholar] [CrossRef]
- St. Luce, M.; Lemke, R.; Gan, Y.; McConkey, B.; May, W.; Campbell, C.; Zentner, R.; Wang, H.; Kroebel, R.; Fernandez, M.; Brandt, K. Diversifying Cropping Systems Enhances Productivity, Stability, and Nitrogen Use Efficiency. Agronomy Journal 2020, 112, 1517–1536. [Google Scholar] [CrossRef]
- Wang, J.; Zou, J. No-Till Increases Soil Denitrification Via Its Positive Effects on the Activity and Abundance of the Denitrifying Community. Soil Biology and Biochemistry 2020, 142, 107706. [Google Scholar] [CrossRef]
- Knapp, S.; van der Heijden, M. G. A. A Global Meta-Analysis of Yield Stability in Organic and Conservation Agriculture. Nat Commun 2018, 9, 3632. [Google Scholar] [CrossRef]
- Luo, Z.; Gan, Y.; Niu, Y.; Zhang, R.; Li, L.; Cai, L.; Xie, J. Soil Quality Indicators and Crop Yield under Long-Term Tillage Systems. Experimental Agriculture 2017, 53, 497–511. [Google Scholar] [CrossRef]
- Carlos, F. S.; Schaffer, N.; Marcolin, E.; Fernandes, R. S.; Mariot, R.; Mazzurana, M.; Roesch, L. F. W.; Levandoski, B.; de Oliveira Camargo, F. A. A Long-Term No-Tillage System Can Increase Enzymatic Activity and Maintain Bacterial Richness in Paddy Fields. Land Degradation & Development 2021, 32, 2257–2268. [Google Scholar] [CrossRef]
- Li, Y.; Li, Z.; Cui, S.; Zhang, Q. Trade-Off between Soil Ph, Bulk Density and Other Soil Physical Properties under Global No-Tillage Agriculture. Geoderma 2020, 361, 114099. [Google Scholar] [CrossRef]
- Guo, Y.; Yin, W.; Chai, Q.; Fan, Z.; Hu, F.; Fan, H.; Zhao, C.; Yu, A.; Coulter, J. A. No Tillage with Previous Plastic Covering Increases Water Harvesting and Decreases Soil CO2 Emissions of Wheat in Dry Regions. Soil and Tillage Research 2021, 208, 104883. [Google Scholar] [CrossRef]
- Guo, Y.; Yin, W.; Fan, Z.; Hu, F.; Fan, H.; Zhao, C.; Yu, A.; Chai, Q.; Coulter, J. A. No-Tillage with Reduced Water and Nitrogen Supply Improves Water Use Efficiency of Wheat in Arid Regions. Agronomy Journal 2020, 112, 578–591. [Google Scholar] [CrossRef]
- Boeckx, P.; Van Nieuland, K.; Van Cleemput, O. Short-Term Effect of Tillage Intensity on N2o and Co2 Emissions. Agronomy for Sustainable Development 2011, 31, 453–461. [Google Scholar] [CrossRef]
- Maia, S. M. F.; Otutumi, A. T.; Mendonça, E. d. S.; Neves, J. C. L.; Oliveira, T. S. d. Combined Effect of Intercropping and Minimum Tillage on Soil Carbon Sequestration and Organic Matter Pools in the Semiarid Region of Brazil. Soil Research 2019, 57, 266–275. [Google Scholar] [CrossRef]
- Niu, Y.; Bainard, L. D.; May, W. E.; Hossain, Z.; Hamel, C.; Gan, Y. Intensified Pulse Rotations Buildup Pea Rhizosphere Pathogens in Cereal and Pulse Based Cropping Systems. Front Microbiol 2018, 9, 1909. [Google Scholar] [CrossRef]
- Li, J.; Huang, L.; Zhang, J.; Coulter, J. A.; Li, L.; Gan, Y. Diversifying Crop Rotation Improves System Robustness. Agronomy for Sustainable Development 2019, 39, 38. [Google Scholar] [CrossRef]
- Liu, K.; Bandara, M.; Hamel, C.; Knight, J. D.; Gan, Y. Intensifying Crop Rotations with Pulse Crops Enhances System Productivity and Soil Organic Carbon in Semi-Arid Environments. Field Crops Research 2020, 248, 107657. [Google Scholar] [CrossRef]
- Mogale, E. T.; Ayisi, K. K.; Munjonji, L.; Kifle, Y. G. Biological Nitrogen Fixation of Cowpea in a No-Till Intercrop under Contrasting Rainfed Agro-Ecological Environments. Sustainability 2023, 15, 2244. [Google Scholar] [CrossRef]
- Gong, X.; Liu, C.; Li, J.; Luo, Y.; Yang, Q.; Zhang, W.; Yang, P.; Feng, B. Responses of Rhizosphere Soil Properties, Enzyme Activities and Microbial Diversity to Intercropping Patterns on the Loess Plateau of China. Soil and Tillage Research 2019, 195, 104355. [Google Scholar] [CrossRef]
- Cong, W. F.; Hoffland, E.; Li, L.; Six, J.; Sun, J. H.; Bao, X. G.; Zhang, F. S.; Van Der Werf, W. Intercropping Enhances Soil Carbon and Nitrogen. Global change biology 2015, 21, 1715–1726. [Google Scholar] [CrossRef] [PubMed]
- Rumpel, C.; Kögel-Knabner, I. Deep Soil Organic Matter—a Key but Poorly Understood Component of Terrestrial C Cycle. Plant and Soil 2011, 338, 143–158. [Google Scholar] [CrossRef]
- Hu, F.; Feng, F.; Zhao, C.; Chai, Q.; Yu, A.; Yin, W.; Gan, Y. Integration of Wheat-Maize Intercropping with Conservation Practices Reduces Co2 Emissions and Enhances Water Use in Dry Areas. Soil and Tillage Research 2017, 169, 44–53. [Google Scholar] [CrossRef]
- Li, Y.; Laterrière, M.; Lay, C.-Y.; Klabi, R.; Masse, J.; St-Arnaud, M.; Yergeau, É.; Lupwayi, N. Z.; Gan, Y.; Hamel, C. Effects of Arbuscular Mycorrhizal Fungi Inoculation and Crop Sequence on Root-Associated Microbiome, Crop Productivity and Nutrient Uptake in Wheat-Based and Flax-Based Cropping Systems. Applied Soil Ecology 2021, 168, 104136. [Google Scholar] [CrossRef]
- Wang, L.; Gan, Y.; Bainard, L. D.; Hamel, C.; St-Arnaud, M.; Hijri, M. Expression of N-Cycling Genes of Root Microbiomes Provides Insights for Sustaining Oilseed Crop Production. Environmental Microbiology 2020, 22, 4545–4556. [Google Scholar] [CrossRef]
- Bazghaleh, N.; Hamel, C.; Gan, Y.; Tar’an, B.; Knight, J. D. Genotypic Variation in the Response of Chickpea to Arbuscular Mycorrhizal Fungi and Non-Mycorrhizal Fungal Endophytes. Canadian Journal of Microbiology 2018, 64, 265–275. [Google Scholar] [CrossRef]
- Chen, J.; Arafat, Y.; Wu, L.; Xiao, Z.; Li, Q.; Khan, M. A.; Khan, M. U.; Lin, S.; Lin, W. Shifts in Soil Microbial Community, Soil Enzymes and Crop Yield under Peanut/Maize Intercropping with Reduced Nitrogen Levels. Applied Soil Ecology 2018, 124, 327–334. [Google Scholar] [CrossRef]
- Lai, H.; Gao, F.; Su, H.; Zheng, P.; Li, Y.; Yao, H. Nitrogen Distribution and Soil Microbial Community Characteristics in a Legume–Cereal Intercropping System: A Review. Agronomy for Sustainable Development 2022, 12, 1900. [Google Scholar] [CrossRef]
- Ablimit, R.; Li, W.; Zhang, J.; Gao, H.; Zhao, Y.; Cheng, M.; Meng, X.; An, L.; Chen, Y. Altering Microbial Community for Improving Soil Properties and Agricultural Sustainability During a 10-Year Maize-Green Manure Intercropping in Northwest China. Journal of Environmental Management 2022, 321, 115859. [Google Scholar] [CrossRef] [PubMed]
- Moraru, P. I.; Rusu, T. Soil Tillage Conservation and Its Effect on Soil Organic Matter, Water Management and Carbon Sequestration. Journal of Food, Agriculture & Environment 2010, 8, 309–312. [Google Scholar]
- Liu, C.; Li, L.; Xie, J.; Coulter, J. A.; Zhang, R.; Luo, Z.; Cai, L.; Wang, L.; Gopalakrishnan, S. Soil Bacterial Diversity and Potential Functions Are Regulated by Long-Term Conservation Tillage and Straw Mulching. Microorganisms 2020, 8, 836. [Google Scholar] [CrossRef]
- Turner, B. L. Variation in Ph Optima of Hydrolytic Enzyme Activities in Tropical Rain Forest Soils. Appl. Environ. Microbiol. 2010, 76, 6485–6493. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A. M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Li, D.; Liu, C.-M.; Luo, R.; Sadakane, K.; Lam, T.-W. Megahit: An Ultra-Fast Single-Node Solution for Large and Complex Metagenomics Assembly Via Succinct De Bruijn Graph. Bioinformatics 2015, 31, 1674–1676. [Google Scholar] [CrossRef]
- Buchfink, B.; Xie, C.; Huson, D. H. Fast and Sensitive Protein Alignment Using Diamond. Nature methods 2015, 12, 59–60. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D. “Vegan: Community Ecology Package. R Package Version 2.4-5.” 2017. Available online: https://CRAN.R-project.org/package=vegan.
- Pielou, E. C. The Measurement of Diversity in Different Types of Biological Collections. J Theor Biol 1966, 13, 131–144. [Google Scholar] [CrossRef]
- Peng, M.; Jia, H.; Wang, Q. The Effect of Land Use on Bacterial Communities in Saline-Alkali Soil. Curr Microbiol 2017, 74, 325–333. [Google Scholar] [CrossRef]
- Bastian, M.; Heymann, S.; Jacomy, M. Gephi: An Open Source Software for Exploring and Manipulating Networks. Proceedings of the international AAAI conference on web and social media 2009, 3, 361–362. [Google Scholar] [CrossRef]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W. S.; Huttenhower, C. Metagenomic Biomarker Discovery and Explanation. Genome biology 2011, 12, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Sunagawa, S.; Coelho, L. P.; Chaffron, S.; Kultima, J. R.; Labadie, K.; Salazar, G.; Djahanschiri, B.; Zeller, G.; Mende, D. R.; Alberti, A. Structure and Function of the Global Ocean Microbiome. Science 2015, 348, 1261359. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Chai, Q.; Cao, W.; Whalen, J. K.; Zhao, L.; Cai, L. No-Tillage Reduces Competition and Enhances Compensatory Growth of Maize (Zea Mays L.) Intercropped with Pea (Pisum Sativum L.). Field Crops Research 2019, 243, 107611. [Google Scholar] [CrossRef]
- Wei, H.; Wang, L.; Hassan, M.; Xie, B. Succession of the Functional Microbial Communities and the Metabolic Functions in Maize Straw Composting Process. Bioresour Technol 2018, 256, 333–341. [Google Scholar] [CrossRef] [PubMed]
- García-Palacios, P.; Chen, J. Emerging Relationships among Soil Microbes, Carbon Dynamics and Climate Change. Functional Ecology 2022, 36, 1332–1337. [Google Scholar] [CrossRef]
- Daryanto, S.; Wang, L.; Jacinthe, P. A. Meta-Analysis of Phosphorus Loss from No-Till Soils. J Environ Qual 2017, 46, 1028–1037. [Google Scholar] [CrossRef]
- Nannipieri, P.; Giagnoni, L.; Landi, L.; Renella, G. Role of Phosphatase Enzymes in Soil, In Phosphorus in Action: Springer, Berlin, Heidelberg., 2011; pp.215-243. [CrossRef]
- Li, Y.; Song, D.; Liang, S.; Dang, P.; Qin, X.; Liao, Y.; Siddique, K. H. M. Effect of No-Tillage on Soil Bacterial and Fungal Community Diversity: A Meta-Analysis. Soil and Tillage Research 2020, 204, 104721. [Google Scholar] [CrossRef]
- Wagg, C.; Schlaeppi, K.; Banerjee, S.; Kuramae, E. E.; van der Heijden, M. G. Fungal-Bacterial Diversity and Microbiome Complexity Predict Ecosystem Functioning. Nature communications 2019, 10, 4841. [Google Scholar] [CrossRef]
- Wagg, C.; Bender, S. F.; Widmer, F.; van der Heijden, M. G. Soil Biodiversity and Soil Community Composition Determine Ecosystem Multifunctionality. Proc Natl Acad Sci U S A 2014, 111, 5266–5270. [Google Scholar] [CrossRef]
- Mathew, R. P.; Feng, Y.; Githinji, L.; Ankumah, R.; Balkcom, K. S. Impact of No-Tillage and Conventional Tillage Systems on Soil Microbial Communities. Applied and Environmental Soil Science 2012, 2012, 1–10. [Google Scholar] [CrossRef]
- Cai, L.; Guo, Z.; Zhang, J.; Gai, Z.; Liu, J.; Meng, Q.; Liu, X. No Tillage and Residue Mulching Method on Bacterial Community Diversity Regulation in a Black Soil Region of Northeastern China. Plos One 2021, 16, e0256970. [Google Scholar] [CrossRef] [PubMed]
- Barka, E. A.; Vatsa, P.; Sanchez, L.; Gaveau-Vaillant, N.; Jacquard, C.; Meier-Kolthoff, J. P.; Klenk, H. P.; Clement, C.; Ouhdouch, Y.; van Wezel, G. P. Taxonomy, Physiology, and Natural Products of Actinobacteria. Microbiol Mol Biol Rev 2016, 80, 1–43. [Google Scholar] [CrossRef] [PubMed]
- Hallam, S. J.; Konstantinidis, K. T.; Putnam, N.; Schleper, C.; Watanabe, Y.-i.; Sugahara, J.; Preston, C.; de la Torre, J.; Richardson, P. M.; DeLong, E. F. Genomic Analysis of the Uncultivated Marine Crenarchaeote Cenarchaeum Symbiosum. Proceedings of the National Academy of Sciences of the United States of America 2006, 103, 18296–18301. [Google Scholar] [CrossRef] [PubMed]
- Könneke, M.; Schubert, D. M.; Brown, P. C.; Hügler, M.; Standfest, S.; Schwander, T.; Schada von Borzyskowski, L.; Erb, T. J.; Stahl, D. A.; Berg, I. A. Ammonia-Oxidizing Archaea Use the Most Energy-Efficient Aerobic Pathway for CO2 Fixation. Proceedings of the National Academy of Sciences of the United States of America 2014, 111, 8239–8244. [Google Scholar] [CrossRef] [PubMed]
- Santoro, A. E.; Buchwald, C.; McIlvin, M. R.; Casciotti, K. L. Isotopic Signature of N2o Produced by Marine Ammonia-Oxidizing Archaea. Science Advances 2011, 333, 1282–1285. [Google Scholar] [CrossRef]
- Baker, B. J.; De Anda, V.; Seitz, K. W.; Dombrowski, N.; Santoro, A. E.; Lloyd, K. G. Diversity, Ecology and Evolution of Archaea. Nat Microbiol 2020, 5, 887–900. [Google Scholar] [CrossRef]
- Mhete, M.; Eze, P. N.; Rahube, T. O.; Akinyemi, F. O. Soil Properties Influence Bacterial Abundance and Diversity under Different Land-Use Regimes in Semi-Arid Environments. Scientific African 2020, 7, e00246. [Google Scholar] [CrossRef]
- Cederlund, H.; Wessén, E.; Enwall, K.; Jones, C. M.; Juhanson, J.; Pell, M.; Philippot, L.; Hallin, S. Soil Carbon Quality and Nitrogen Fertilization Structure Bacterial Communities with Predictable Responses of Major Bacterial Phyla. Applied soil ecology 2014, 84, 62–68. [Google Scholar] [CrossRef]
- Aslam, Z.; Yasir, M.; Yoon, H. S.; Jeon, C. O.; Chung, Y. R. Diversity of the Bacterial Community in the Rice Rhizosphere Managed under Conventional and No-Tillage Practices. Journal of Microbiology 2013, 51, 747–756. [Google Scholar] [CrossRef]
- Wang, Z.; Li, Y.; Li, T.; Zhao, D.; Liao, Y. Tillage Practices with Different Soil Disturbance Shape the Rhizosphere Bacterial Community Throughout Crop Growth. Soil and Tillage Research 2020, 197, 104501. [Google Scholar] [CrossRef]
- Yu, L.; Luo, S.; Gou, Y.; Xu, X.; Wang, J. Structure of Rhizospheric Microbial Community and N Cycling Functional Gene Shifts with Reduced N Input in Sugarcane-Soybean Intercropping in South China. Agriculture, Ecosystems, Environment 2021, 314, 107413. [Google Scholar] [CrossRef]
- Bao, Y.; Dolfing, J.; Guo, Z.; Chen, R.; Wu, M.; Li, Z.; Lin, X.; Feng, Y. Important Ecophysiological Roles of Non-Dominant Actinobacteria in Plant Residue Decomposition, Especially in Less Fertile Soils. Microbiome 2021, 9, 1–17. [Google Scholar] [CrossRef]
- Beman, J. M.; Popp, B. N.; Francis, C. A. Molecular and Biogeochemical Evidence for Ammonia Oxidation by Marine Crenarchaeota in the Gulf of California. The ISME Journal 2008, 2, 429–441. [Google Scholar] [CrossRef] [PubMed]
- Nicol, G. W.; Schleper, C. Ammonia-Oxidising Crenarchaeota: Important Players in the Nitrogen Cycle? Trends Microbiol 2006, 14, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Lützow, M. v.; Kögel-Knabner, I.; Ekschmitt, K.; Matzner, E.; Guggenberger, G.; Marschner, B.; Flessa, H. Stabilization of Organic Matter in Temperate Soils: Mechanisms and Their Relevance under Different Soil Conditions - a Review. European Journal of Soil Science 2006, 57, 426–445. [Google Scholar] [CrossRef]
- von Lützow, M.; Kögel-Knabner, I.; Ekschmitt, K.; Flessa, H.; Guggenberger, G.; Matzner, E.; Marschner, B. Som Fractionation Methods: Relevance to Functional Pools and to Stabilization Mechanisms. Soil Biology and Biochemistry 2007, 39, 2183–2207. [Google Scholar] [CrossRef]
- Zhang, X.; Xin, X.; Zhu, A.; Yang, W.; Zhang, J.; Ding, S.; Mu, L.; Shao, L. Linking Macroaggregation to Soil Microbial Community and Organic Carbon Accumulation under Different Tillage and Residue Managements. Soil and Tillage Research 2018, 178, 99–107. [Google Scholar] [CrossRef]
Figure 1.
Maize intercropping pea in field strips—an increasingly adopted cropping system in the arid northwestern China (Photo taken at Wuwei county, Gansu province, China, in 2018).
Figure 1.
Maize intercropping pea in field strips—an increasingly adopted cropping system in the arid northwestern China (Photo taken at Wuwei county, Gansu province, China, in 2018).

Figure 2.
Principal component analysis of soil microbial diversity of (a) archaea, (b) bacteria, and (c) eukaryota, based on absolute abundance homogenization matrix at phylum level; Microbial co-occurrence networks based on taxonomic composition constructed for (d) the conventional tillage and (e) no-tillage treatments. The treatment abbreviations are: CTM, CTP, CTIM, CTIP were the maize monoculture, pea monoculture, maize in intercropping (Maize/Pea), and pea in intercropping (Maize/Pea) in conventional tillage system and NTM, NTP, NTIM, NTIP were the maize monoculture, pea monoculture, maize in intercropping (Maize/Pea), and pea in intercropping (Maize/Pea) in no tillage system, respectively.
Figure 2.
Principal component analysis of soil microbial diversity of (a) archaea, (b) bacteria, and (c) eukaryota, based on absolute abundance homogenization matrix at phylum level; Microbial co-occurrence networks based on taxonomic composition constructed for (d) the conventional tillage and (e) no-tillage treatments. The treatment abbreviations are: CTM, CTP, CTIM, CTIP were the maize monoculture, pea monoculture, maize in intercropping (Maize/Pea), and pea in intercropping (Maize/Pea) in conventional tillage system and NTM, NTP, NTIM, NTIP were the maize monoculture, pea monoculture, maize in intercropping (Maize/Pea), and pea in intercropping (Maize/Pea) in no tillage system, respectively.
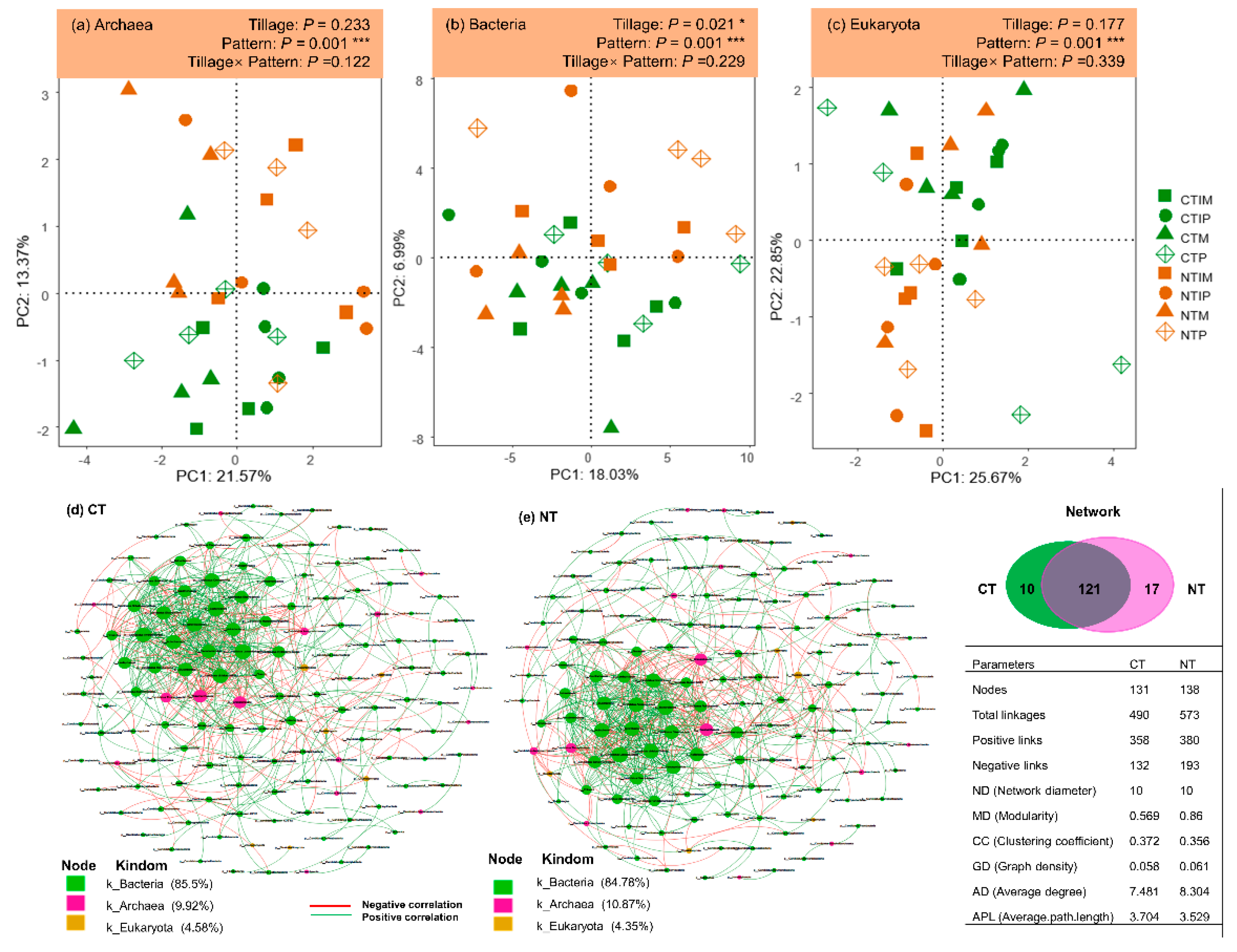
Figure 3.
(a) The circles radiating from the inside out represent taxonomic levels from phylum to genus (or species), and each small circle at a different classification level represents a classification at that level, and the size of the small circle diameter is proportional to the size of the relative abundance. (b) The composition of archaea, bacteria, and fungi at phylum levels in different treatments. (c) The distribution of LDA scores of different species, the color represents the corresponding groups, and the length of the bar represents the contribution of different species (LDA Score>4).
Figure 3.
(a) The circles radiating from the inside out represent taxonomic levels from phylum to genus (or species), and each small circle at a different classification level represents a classification at that level, and the size of the small circle diameter is proportional to the size of the relative abundance. (b) The composition of archaea, bacteria, and fungi at phylum levels in different treatments. (c) The distribution of LDA scores of different species, the color represents the corresponding groups, and the length of the bar represents the contribution of different species (LDA Score>4).
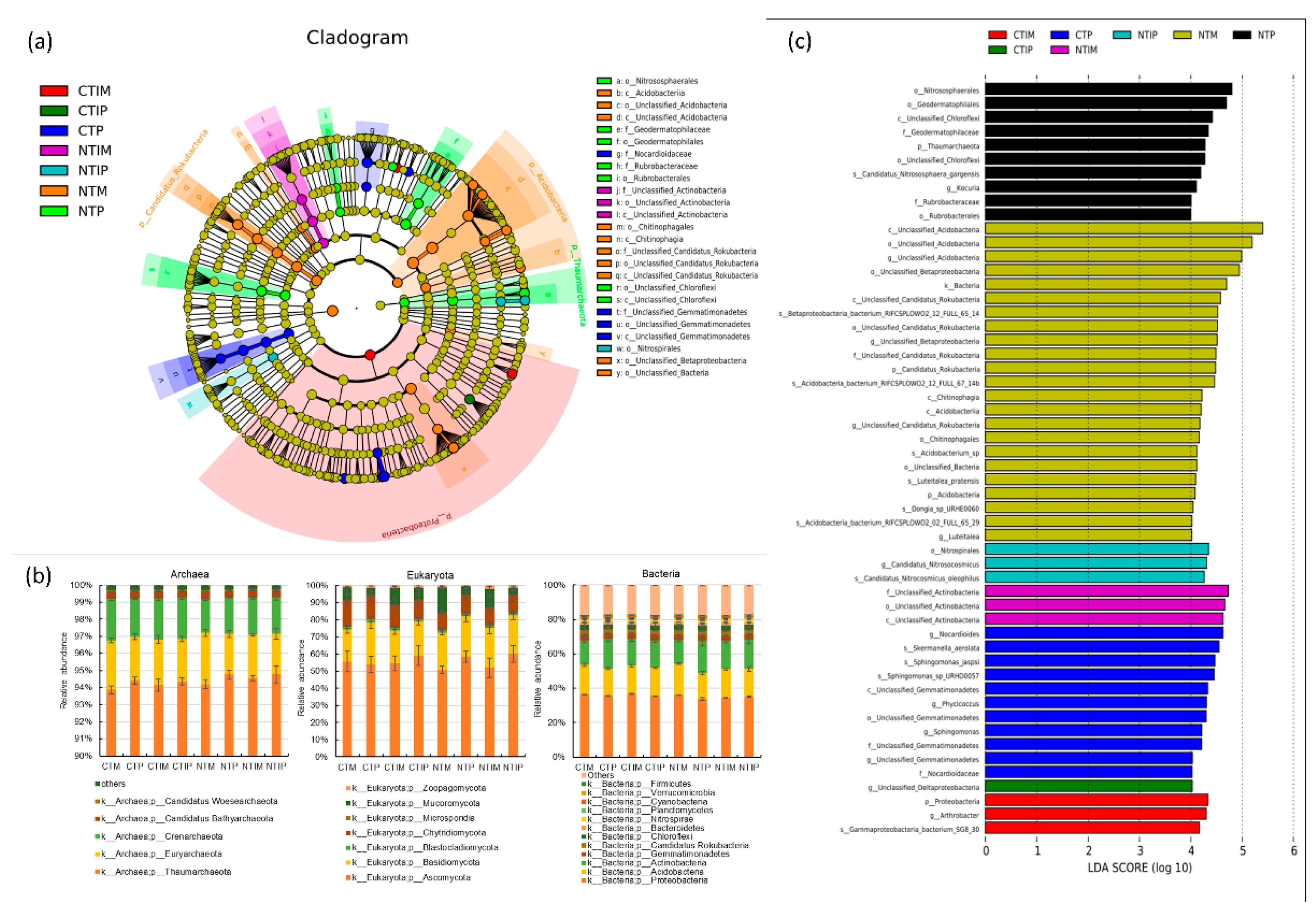
Figure 4.
Correlations of (a) microbial taxonomic and functional diversity with yield and soil chemical properties, and (b) among soil chemical properties, including soil moisture (SW), pH, electrical conductivity (EC), total carbon (TC), inorganic carbon (IC), organic carbon (OC), total phosphorus (TP), inorganic phosphorus (IP), organic phosphorus (OP), total nitrogen (TN), ammonium nitrogen (HN4+-N), nitrate nitrogen (NO3--N), the ratio of TC to TN, the ratio of TC and TP, and the ratio of TN and TP. In (g), the colored circles represented significance at the 0.05 level.
Figure 4.
Correlations of (a) microbial taxonomic and functional diversity with yield and soil chemical properties, and (b) among soil chemical properties, including soil moisture (SW), pH, electrical conductivity (EC), total carbon (TC), inorganic carbon (IC), organic carbon (OC), total phosphorus (TP), inorganic phosphorus (IP), organic phosphorus (OP), total nitrogen (TN), ammonium nitrogen (HN4+-N), nitrate nitrogen (NO3--N), the ratio of TC to TN, the ratio of TC and TP, and the ratio of TN and TP. In (g), the colored circles represented significance at the 0.05 level.
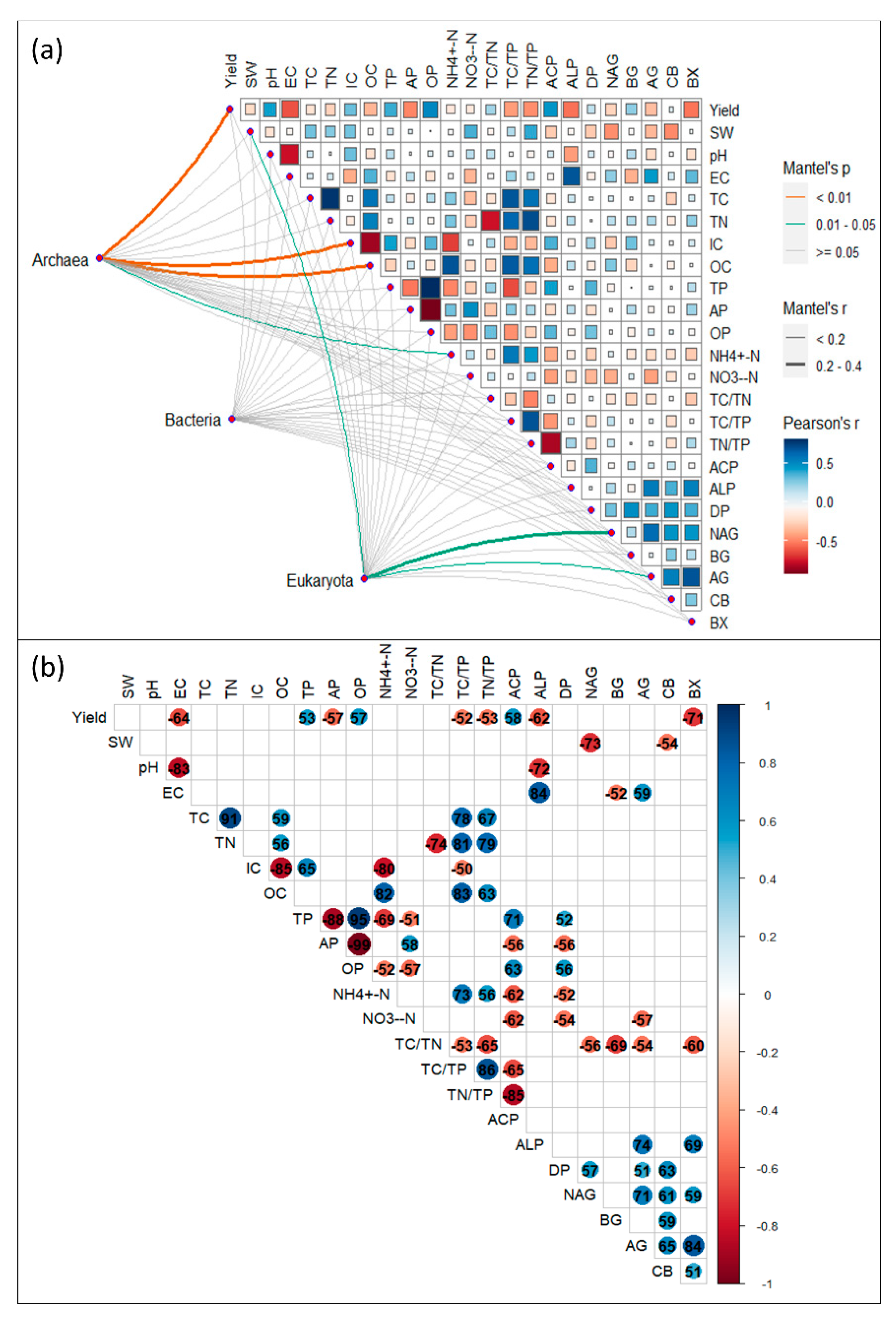
Figure 5.
Correlation analysis of yield and soil chemical properties with (a) archaea, (b) bacteria and (c) fungi at phylum levels. Asterisks indicate a significant association between the abundance of a given functional gene and particular soil chemical properties: “*” P<0.05, “**” P<0.01, and “***” P<0.001.
Figure 5.
Correlation analysis of yield and soil chemical properties with (a) archaea, (b) bacteria and (c) fungi at phylum levels. Asterisks indicate a significant association between the abundance of a given functional gene and particular soil chemical properties: “*” P<0.05, “**” P<0.01, and “***” P<0.001.
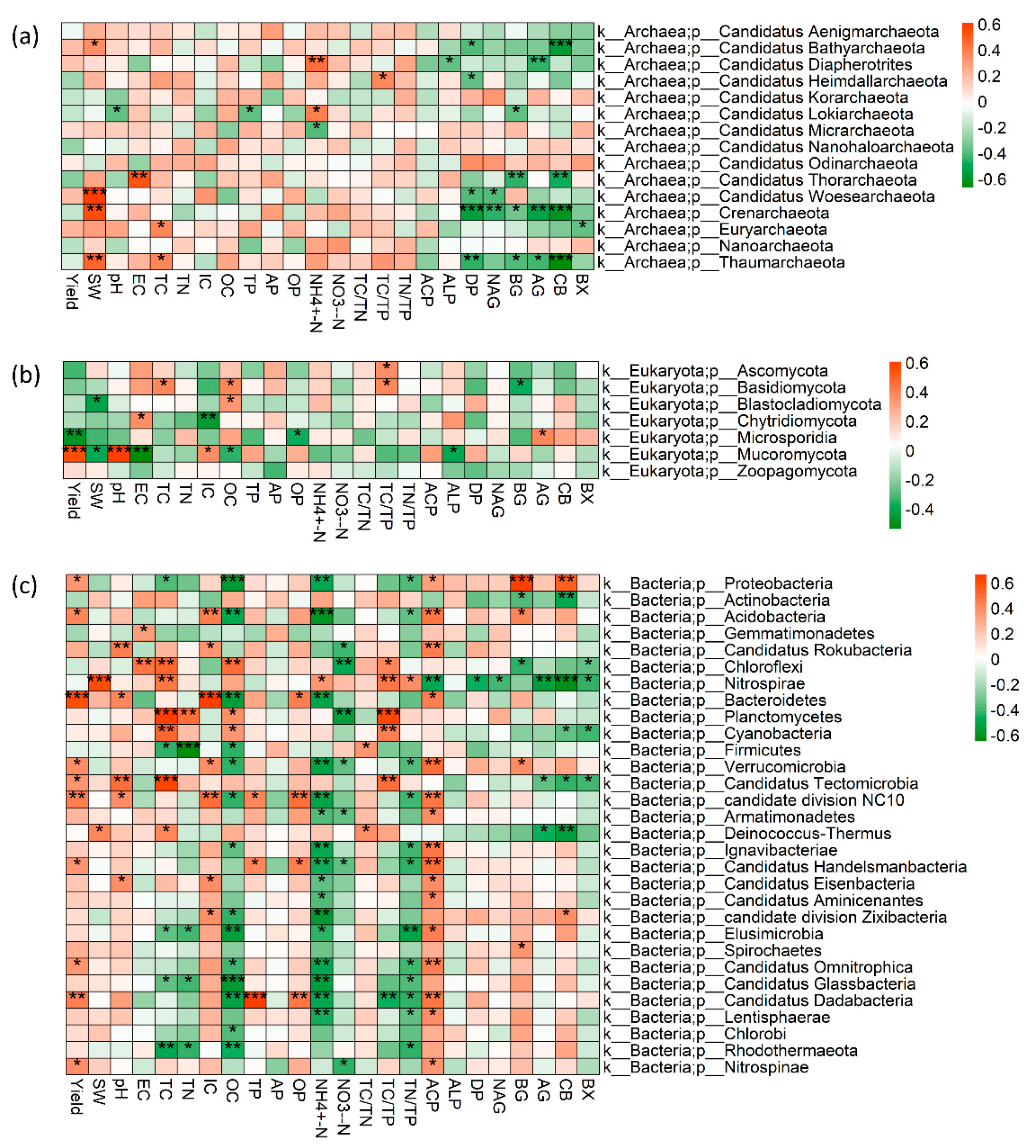

Table 1.
Yields of pea and maize in inter-/mono-cropping systems under the conditions of conventional tillage and no-tillage.
Table 1.
Yields of pea and maize in inter-/mono-cropping systems under the conditions of conventional tillage and no-tillage.
| Treatments | Pea | Maize | Total of pea and maize (kg·hm-2) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Tillage | Pattern | Number of pods per plant | Number of grains per pod | 1000-seed weight (g) | Yield (kg hm-2) |
Net area yields (kg hm-2) |
Number of spikes per square meter | Kernel number per spike | 1000-seed weight (g) | Yield (kg·hm-2) |
Net area yields(kg·hm-2) | ||
| NT | P | 9.3a† | 6.0bc | 211.2c | 3955.03a | 3955.03c | 3955.03±74e | ||||||
| M | 7.5b | 684a | 480.5a | 14041.47c | 14041.47c | 14041.47±368c | |||||||
| M/P | 8.0bc | 6.7a | 242.1a | 2393.63c | 5684.88a | 7.5b | 684a | 480.5a | 12983.87bc | 22426.68a | 15377.50±353a | ||
| CT | P | 8.3ab | 5.3c | 198.3d | 3667.50b | 3667.5d | - | - | - | - | 3667.50±47e | ||
| M | - | - | - | - | 7.6ab | 607c | 421.1c | 13302.47b | 13302.47d | 13302.47±177d | |||
| M/P | 7.3c | 6.3a | 224.2b | 2205.10d | 5237.11b | 7.5b | 659b | 445.6b | 12545.30c | 21669.15b | 14750.40±267b | ||
| Tillage | 0.076 | 0.122 | 0.000 *** | 0.000 *** | 0.000 *** | 0.017 * | 0.001 ** | 0.001 ** | 0.026 * | 0.006 ** | 0.002 ** | ||
| Pattern | 0.021 * | 0.020 * | 0.000 *** | 0.000 *** | 0.000 *** | 0.000 *** | 0.000 *** | 0.000 *** | 0.003 ** | 0.000 *** | 0.000 *** | ||
| Tillage* Pattern | 0.694 | 0.58 | 0.466 | 0.293 | 0.180 | 0.040 * | 0.883 | 0.184 | 0.507 | 0.967 | 0.435 | ||
† Different letters in the same column indicated the significant difference between different treatments. P, M, M/P were the pea monoculture, maize monoculture, maize and pea intercropping; NT and CT were no-tillage and conventional tillage, respectively. “*” P<0.05, “**” P<0.01, and “***” P<0.001.
Table 2.
Soil chemical properties in maize and pea intercropping under the conditions of conventional tillage and no-tillage.
Table 2.
Soil chemical properties in maize and pea intercropping under the conditions of conventional tillage and no-tillage.
| Treatment | SW (%) | pH | EC (us m-1) | TC (g kg-1) |
TN (g kg-1) |
OC (g kg-1) |
HN4+-N (mg kg-1) |
NO3—N (mg kg-1) |
TP (mg kg-1) |
AP (mg kg-1) |
OP (mg kg-1) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Tillage | |||||||||||
| CT | 11.8±0.64 | 8.19±0.02b† | 129.75±10.61a | 10.59±0.12b | 0.51±0.01b | 3.32±0.24b | 5.3±0.1b | 4.67±0.24a | 130.72±1.73 | 93.72±3.49 | 37±4.62b |
| NT | 12.51±0.7 | 8.26±0.03a | 112.33±5.93b | 11.64±0.12a | 0.57±0.01a | 4.61±0.37a | 5.54±0.11a | 4.31±0.14b | 132.75±2.87 | 88.01±3.47 | 44.74±5.38a |
| Pattern | |||||||||||
| M | 10.48±0.35b† | 8.24±0.06 | 116.5±12.23b | 10.97±0.24 | 0.55±0.02 | 3.37±0.18b | 5.12±0.11b | 3.9±0.11c | 136.27±1.66 | 82.94±6.09b | 53.33±6.4a |
| P | 10.84±0.51b | 8.18±0.03 | 154.83±12.67a | 11.27±0.13 | 0.55±0.01 | 5.33±0.51a | 5.71±0.19a | 4.28±0.12bc | 125.68±3.3 | 100.63±0.87a | 25.04±3.45b |
| IM | 11.97±0.66b | 8.28±0.01 | 95±3.77b | 10.95±0.3 | 0.52±0.01 | 3.62±0.16b | 5.5±0.07a | 4.59±0.3ab | 133.62±3.84 | 86.07±5.12b | 47.55±8.1a |
| IP | 15.33±0.58a | 8.2±0.02 | 117.83±4.01b | 11.27±0.41 | 0.55±0.03 | 3.56±0.62b | 5.35±0.12ab | 5.17±0.29a | 131.38±3.12 | 93.82±3.34ab | 37.56±4.72ab |
| Tillage* Pattern | |||||||||||
| CTM | 10.49±0.54 | 8.11±0.02d | 143.67±2.73b | 10.47±0.14c | 0.5±0.02cd | 3.42±0.12d | 5.03±0.21c | 3.76±0.17d | 137.48±3.48ab | 75.74±3.52c | 61.73±4.57ab |
| CTP | 10.95±1.14 | 8.15±0.06cd | 178.33±8.65a | 11.19±0.12b | 0.55±0.01bc | 4.29±0.24bc | 5.4±0.16bc | 4.02±0.01cd | 127.78±1.57abc | 102.28±0.38a | 25.5±1.32d |
| CTIM | 11.12±1.19 | 8.28±0.02ab | 87.33±2.73e | 10.3±0.06c | 0.5±0.02cd | 3.36±0.26d | 5.58±0.11ab | 5.22±0.08a | 127.27±1.81bc | 96.4±4.82ab | 30.87±3.99cd |
| CTIP | 14.65±0.78 | 8.24±0.02bc | 109.67±2.73cd | 10.4±0.12c | 0.49±0.03d | 2.22±0.2e | 5.19±0.19bc | 5.66±0.05a | 130.37±3.84abc | 100.46±2.88ab | 29.91±3.79cd |
| NTM | 10.48±0.57 | 8.37±0.05a | 89.33±1.45e | 11.46±0.12b | 0.59±0.02ab | 3.33±0.39d | 5.21±0.11bc | 4.04±0.11cd | 135.06±0.46abc | 90.13±11ab | 44.93±10.65bc |
| NTP | 10.74±0.07 | 8.22±0.04bc | 131.33±13.25b | 11.36±0.26b | 0.56±0.03abc | 6.36±0.41a | 6.02±0.23a | 4.54±0.05bc | 123.58±6.9c | 98.99±0.98ab | 24.59±7.59d |
| NTIM | 12.82±0.09 | 8.28±0.01ab | 102.67±2.19de | 11.6±0.14b | 0.53±0.01bcd | 3.88±0.04c | 5.41±0.09bc | 3.97±0.21d | 139.98±5.47a | 75.74±0.98c | 64.23±5.84a |
| NTIP | 16.02±0.77 | 8.17±0.02cd | 126±2.52bc | 12.14±0.23a | 0.61±0.01a | 4.89±0.3b | 5.5±0.09b | 4.68±0.4b | 132.4±5.74abc | 87.18±1.84ab | 45.22±6.18bc |
| PremANOVA: Pr(>F) | |||||||||||
| Tillage | 0.200 | 0.018 * | <0.001 *** | <0.001 *** | <0.001 *** | 0.097 | 0.049 * | 0.009 ** | 0.495 | 0.081 | 0.043 * |
| Pattern | <0.001 *** | 0.067 | <0.001 *** | 0.098 | 0.303 | 0.152 | 0.012 * | <0.001 *** | 0.110 | 0.004 ** | <0.001 *** |
| Tillage*Pattern | 0.499 | 0.001 ** | <0.001 *** | 0.001** | 0.042 * | 0.559 | 0.133 | <0.001 *** | 0.210 | 0.007 ** | 0.001** |
†Different letters in the same column indicated the significant difference between the treatments. CTM, CTP, CTIM, CTIP were the maize monoculture, pea monoculture, maize in intercropping (Maize/Pea), and pea in intercropping (Maize/Pea) in conventional tillage system and NTM, NTP, NTIM, NTIP were the maize monoculture, pea monoculture, maize in intercropping (Maize/Pea), and pea in intercropping (Maize/Pea) in no-tillage system, respectively. “*” P<0.05, “**” P<0.01, and “***” P<0.001.
Table 3.
Soil enzyme activities in maize and pea intercropping under the conditions of conventional tillage and no-tillage.
Table 3.
Soil enzyme activities in maize and pea intercropping under the conditions of conventional tillage and no-tillage.
| Treatment | ACP | ALP | DP | NAG | BG | AG | CB | BX |
|---|---|---|---|---|---|---|---|---|
| Tillage | ||||||||
| CT | 111.98±2.9 | 119.49±8.22 | 14.37±1.23 | 16.44±1.37 | 37.84±5.32 | 4.17±0.65 | 15.01±1.19 | 11.1±1.41 |
| NT | 108.45±5.41 | 107.37±5.67 | 15.07±0.99 | 16.66±2.07 | 39.06±10.3 | 3.63±0.56 | 12.28±1 | 10.06±1.14 |
| Pattern | ||||||||
| M | 120.43±5.72a† | 113.59±6.02ab | 19.08±1.91a | 20.73±2.59a | 54.15±16.88 | 5.49±0.8a | 17.13±1.64a | 13±0.61a |
| P | 105.43±4.31ab | 129.03±14.1a | 13.1±1.06b | 20.15±2.26a | 23.97±5.09 | 4.62±1.07ab | 13.2±1.06ab | 11.48±1.35a |
| IM | 115.87±7a | 93.09±6.01b | 13.13±0.85b | 12.35±1.17b | 39.86±10.84 | 2.08±0.3c | 12.31±1.98b | 5.73±1.09b |
| IP | 99.13±3.39b | 118.01±7.85ab | 13.59±0.73b | 12.97±1.35b | 35.82±8.38 | 3.42±0.31bc | 11.95±1.01b | 12.12±2.18a |
| Tillage*Pattern | ||||||||
| CTM | 122.52±1.57 | 123.52±7.15bc | 19.44±3.36 | 20.11±1.14 | 37.4±5.29abc | 5.36±0.73 | 18.59±2.62 | 12.64±0.53 |
| CTP | 113.35±4.12 | 153.35±18.39a | 12.26±1.63 | 19.29±3.09 | 24.65±8.93bc | 6.26±1.56 | 14.81±0.86 | 13.28±2.18 |
| CTIM | 106.42±7.66 | 99.26±11.76cd | 12.87±1.06 | 13.96±1.91 | 63.95±2.71ab | 2.13±0.65 | 15.59±2.69 | 6.74±1.31 |
| CTIP | 105.62±3.92 | 101.84±4.18cd | 12.92±1.07 | 12.4±2.17 | 25.38±0.93bc | 2.95±0.39 | 11.04±1.43 | 11.76±4.85 |
| NTM | 118.35±12.52 | 103.65±5.6cd | 18.71±2.6 | 21.35±5.64 | 70.9±33.41a | 5.62±1.63 | 15.68±2.12 | 13.37±1.2 |
| NTP | 97.51±3.64 | 104.71±8.05cd | 13.94±1.49 | 21±3.9 | 23.3±7bc | 2.97±0.75 | 11.58±1.5 | 9.69±1.05 |
| NTIM | 125.31±9.84 | 86.93±2.08d | 13.38±1.55 | 10.74±0.81 | 15.76±0.31c | 2.04±0.14 | 9.03±1.25 | 4.73±1.81 |
| NTIP | 92.64±0.06 | 134.18±5.37ab | 14.27±1.04 | 13.55±2 | 46.26±15.52abc | 3.88±0.35 | 12.85±1.49 | 12.48±0.3 |
| PremANOVA: Pr(>F) | ||||||||
| Tillage | 0.449 | 0.080 | 0.608 | 0.918 | 0.903 | 0.376 | 0.054 | 0.484 |
| Pattern | 0.019* | 0.010** | 0.015* | 0.017* | 0.225 | 0.007** | 0.047* | 0.013* |
| Tillage*Pattern | 0.065 | 0.004** | 0.921 | 0.825 | 0.047* | 0.104 | 0.201 | 0.660 |
†Different letters in the same column indicated the significant difference between different treatments. CTM, CTP, CTIM, CTIP were the maize monoculture, pea monoculture, maize in intercropping (Maize/Pea), and pea in intercropping (Maize/Pea) in conventional tillage system and NTM, NTP, NTIM, NTIP were the maize monoculture, pea monoculture, maize in intercropping (Maize/Pea), and pea in intercropping (Maize/Pea) in no-tillage system, respectively. The soil enzyme activities including (acid phosphatase, alkaline phosphatase, phosphodiesterase, N-acetyl-β-glucosaminidase, and β-glucosidase, α-glucosidase, cellobiohydrolase, and β-xylosidase following by the abbreviations for each of the enzymes ACP, ALP, DP, NAG, BG, AG, CB, BX. “*” P<0.05, “**” P<0.01, and “***” P<0.001.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



