Submitted:
19 September 2023
Posted:
20 September 2023
You are already at the latest version
Abstract
Due to the expansion of applications of rare earth elements (REE) in various technological pro-cesses, increasing amounts of these metals pollute the environment, including the marine one. Very little is known about the bioaccumulation and toxicity of REE in marine organisms. In the present work, we assessed the contents of these metals, including yttrium and scandium, in rhizomes and leaves of the widespread seagrass Zostera noltei and in the nearby sediments from the Black Sea coast. The total REE content in the sediments was found to be much higher than in Z. noltei. The order of decrease in the major REE contents in the sediments and the seagrass rhizomes was identical, except for La and Y: La was the most abundant in the sediments and Y in the rhizomes. The contents of all REE in rhizomes of Z. noltei were 1.5–10 times higher than in leaves. The greatest difference in the REE contents was found for the minor heavy elements (Sm–Lu). Translocation coefficients for Sc and the minor elements (excluding Tb) from sediments to rhizomes and from rhizomes to leaves turned out to be pairwise equal, which indicate the similarity of the REE translocation mechanisms. Comparing our results with the literature data, it is possible to conclude that the seagrass Z. noltei does not have an advantage in the REE accu-mulation over marine macroalgae. However, large coastal deposits of this seagrass after storms allow it to consider as a possible resource of REE.
Keywords:
rare earth elements
; seagrass Zostera noltei
; Black Sea
1. Introduction
Environmental pollution with various types of toxic inorganic, organic and organometallic compounds is one of the most serious environmental issues in the world. Rare earth element pollution in the marine environment is an emerging problem that has unpredictable consequences for marine ecosystems.
According to the definition of International Union of Theoretical and Applied Chemistry (IUPAC), rare earth elements (REE) are a group of 17 elements, including 15 lanthanoids (metals from lanthanum to lutetium), scandium and yttrium [1]. All rare earth elements are widespread but scattered in the earth’s crust, and they are found in more than 200 minerals, such as carbonates, silicates, and phosphates. The exception is the radioactive promethium, which is extremely rare in nature as it has no stable or long-lived isotopes [2]. Due to their unique physical and chemical properties, REE are of decisive importance for the development of a multitude of industrial technologies and products encompassing the range from cell phones and TVs to LEDs and wind turbines. These elements are indispensable components of magnetic, optical, and electronic devices used in the defense and aerospace industries for the production of unmanned aerial vehicles, guided missiles, laser guidance devices for satellite communications, etc. [3-5]. REEs are used in agriculture as supplements to fertilizers to improve crop growth and production and thus are accumulated in the soil [6,7]. As the use of rare earth elements increases, their release into the biosphere will inevitably grow [8,9].
There are a large number of studies on adverse effects of notoriously toxic elements, such as As, Pb, Cd and Hg [10,11] on biota. However, many other elements, including rare earth elements, are increasingly used in industry, and dumping of huge amounts of waste contributes to their ingress into the soil and groundwater. The background REE levels in waters vary greatly and depend mainly on local geology. Under natural conditions, REE are transferred only in small amounts through groundwater and the atmosphere, but their widespread use in recent times has increased the REE pollution and created new routes of REE bioaccumulation (in plants, animals and humans). Use of modern analytical methods, such as inductively coupled plasma mass spectrometry (ICP-MS), helps improve our understanding of reactivity and mobility of REE in the near-surface environment, their bioavailability and possible risks to human health [12-14]. At the same time, the information on REE effects on human health is very limited at present [15,16].
The determination of REE in marine aquatic organisms is an important aspect of studying the distribution of these elements in the environment. Currently, the REE levels in flora and fauna of aquatic systems are not considered hazardous, but with the increasing use of these elements, the situation may change for the worse in the near future [17]. High degrees of REE bioaccumulation were found in phytoplankton [18] and in mussel shells in rivers of Europe and North America [19], proving their bioavailability and ability to accumulate along the food chain. A possible risk to the environment lies in the bioavailability of REEs and their tendency to fractionate depending on environmental conditions, especially on the water pH and the nature of the solids [17]. Sediment characteristics such as Fe content can limit the transport of rare earth elements due to complexation reactions [20]. Currently, there is evidence of elevated levels of REE in various natural systems, for example, of an anomalously high contents of Ce mainly in the warm surface layer of the Black Sea [21] and Gd near densely populated areas with developed medical care facilities due to the use of Gd as a contrast agent in magnetic resonance imaging [17].
The quantitation of REE in marine biota is important for determining their levels in the environment. Along with other macrophytes, seagrasses are an important link in the matter and energy cycles in coastal ecosystems [22,23]. Therefore, seagrasses can serve as efficient bioindicators of REE pollution in the marine environment [24]. Moreover, they can be useful in the REE extraction from natural waters and wastewater with the prospect of REE recycling and water bioremediation [25,26].
Seagrasses act as important natural archivers of anthropogenic activity. By accumulating and immobilizing contaminants from sediments, they can be used as an indicator of short- and long-term changes in metal fluxes to the coastal ecosystem [27-29]. There is currently very little information on interactions between seagrasses and pollutants, including REE, as well as estimates of the REE transfer from sediments to seagrass [29]. The purpose of this work is to study the distribution of REE in rhizomes and leaves of the seagrass Zostera noltei as a possible indicator of REE pollution in coastal areas of the Black Sea and a potential agent for the REE recovery and bioremediation.
2. Materials and Methods
The seagrass Zostera noltei was collected in Kazachya Bay (Black Sea coast of Crimea) in June at a depth of 0.5–1 m (Figure 1). After removal of visible epiphytes, the leaves and rhizomes of Z. noltei were rinsed with distilled water, crushed and dried to constant weight at 105 °C. For the REE quantitation, 20 mg samples of dry biomass were used. The sediment was sampled from different, randomly selected, nearby sites and thoroughly mixed to eliminate local heterogeneities in the content of elements. The sediment was also dried to constant weight, ground in a porcelain mortar, and sifted through a sieve with a mesh size of 0.5 mm. All samples were prepared in 4–5 replicates.
The sample digestion was performed for two hours using analytical-grade nitric acid purified by sub-boiling distillation in an acid purification system DST-1000 (Savillex, USA) in capped PTFE tubes in an autoclave at 2.2 bar. The digested samples were diluted with deionized water to 1000 mL·g–1 d.w. The REE concentration was measured on an ICP-MS instrument PlasmaQuant® MS Elite (Analytik Jena, Germany) using multielement standards IV-ICPMS-71A,D (Inorganic Ventures, USA). The limits of REE detection on this instrument at this dilution are no higher than 0.03 µg·kg–1 d.w. [30]. Quality assurance and quality control of this analysis was provided by trace element quantitation in certified reference materials ERM®-CD200 (seaweed Fucus vesiculosus) and BCR®-670 (duckweed Lemna minor) after digestion in nitric acid according to the above procedure.
Each analyte was measured in five replications, five scans per replicate. The plasma flow was 9.0 L·min–1, and the RF power was 1.25 kW. The dwell time for each analyte was 50 ms, one point per peak in the peak-hopping mode. To ensure the insignificance of polyatomic interferences, collision reaction interface (CRI) was used as well. In the CRI, hydrogen with the flow rate 40 mL·min–1 was the skimmer gas. No internal standard was used. The signal drift was taken into account by measuring the element concentrations in the diluted standard IV-ICPMS-71A,D after every fifth sample.
Statistical analysis of the measurement results was carried out using one-way ANOVA in PAST 4.12 [31]. The post hoc pairwise comparison was performed using Tukey’s test. In case of heterogeneity of variances detected in Levene’s test, the averages were compared according to the Welch F-test and pairwise Games-Howell test in Matlab 8.2.0 [32]. Hierarchical cluster analysis was performed using PAST 4.12, with the Euclidean distance as the similarity measure and unweighted pair-group average (UPGMA) as the pairwise clustering algorithm.
To estimate whether or not REE in the sediments originate mainly from the rock weathering, the enrichment factor (EF) was calculated as follows [33]:
where the subscript “s” denotes sediments and “UCC” refers to the average values in the upper continental crust. The upper continental crust values [34,35] were taken as reference because the corresponding average values represent the average source rock composition [36]. In Eq. (1), REE are normalized to the aluminium content as the ratio of REE to Al in the weathering products is approximately the same as in the source rock [37].
EF = (REE/Al)s / (REE/Al)UCC
The REE transfer from sediments to the different parts of the seagrass and the translocation from rhizomes to leaves are characterized with the transfer factors (TF) and translocation factor (TF’), respectively, which can be expressed in a straightforward manner as
where “seagrass” stands for either seagrass rhizomes or leaves, and
where the subscripts “l” and “r” denote leaves and rhizomes, respectively. TF>1 indicates the REE bioaccumulation in a marine organism, and the greater TF, the more mobile and bioaccessible element forms are in the sediment environment [38].
TF = Cseagrass / Cs
TF’ = Cl / Cr
3. Results and discussion
3.1. REE contents in the seagrass and sediments
The REE contents in sediments were much higher than those in different parts of the seagrass (Table 1). The contents of the light REE in Z. noltei rhizomes decreased in the following order: Y > La > Ce > Sc > Nd > Pr. For Z. noltei leaves, this order was different: Sc > La > Ce > Y > Pr > Nd. In sediments, the Nd content exceeded that of Sc, and the sequence changed to La > Y > Ce > Nd > Sc > Pr. Thus, the elements in rhizomes and sediments, except for La and Y, had the same decreasing order. A similar order of the element decrease was observed in Bohai Bay situated west of the Bohai Sea, but the major element was Ce [39]. The coastal region surrounding Bohai Bay is one of China’s most densely populated and industrialized areas, where approximately 97% of REE are produced, and industrial and agricultural use of the elements is growing rapidly [40]. Compared to this region, the concentration of REE in the Black Sea, according to our data, was lower by a factor of 2 for La and 8-11 for all other REEs [39], which indicates a relatively low concentration of elements in the sediments of the Crimean coast of the Black Sea.
On the whole, the abundance of all REE in rhizomes was 1.5-10 times higher than in leaves. Moreover, the contents of heavy REE (Sm–Lu) differed by a factor of 7–8 (Table 1). Among all REE, the highest content was noted for Y in rhizomes of Z. noltei. In sediments, the highest content was registered for La. In the general case, the order of the REE decrease in different objects was as follows: sediments > rhizomes > leaves. The exceptions were Sc and La in rhizomes and leaves, where abundances of these elements did not differ significantly.
Cluster analysis of the REE contents in the seagrass (Figure 2) showed the division of elements into two groups: major (Sc, Y, La–Nd) and minor (Sm–Lu). Major REE are characterized by the levels of hundreds to thousands µg·kg–1 in rhizomes and tens to hundreds µg·kg–1 in leaves. Minor REE have the values of tens to hundreds µg·kg–1 in rhizomes and up to ten µg·kg–1 in leaves. The abundance of REE in marine organisms, excluding filter feeders, is due to the element contents in sediments and suspended particles rather than to the amount of REE dissolved in seawater [41]. In seawater, light REE are to a greater extent absorbed by organisms or adsorbed on their surfaces given that small ionic radii of heavy REE allow them to remain in solution due to the formation of stable complexes [17]. There is a small percentage of free ionic REE, mainly light REE, in the solution, which are easily incorporated into organisms [17,42] and may potentially have serious biological consequences. Another explanation for the fractionation of REE in seawater is associated with diatoms [43], which absorb these elements in the sea surface layer and transport them to the deeper waters. Light REE tends to be bound to carbonates and oxides and are thus more accumulated than heavy REE in the surficial layer of sediments. This may serve as an explanation for the enrichment of the composition of ocean waters with heavy REEs.
Contemporary studies show that REE are present in almost all parts of the human body in low concentrations and may play an important biological role [44]. On the other hand, the REE concentration in drinking water exceeding the ordinary levels by a factor of seven may pose a potential risk to health [18]. There is a lack of data on the content of REE in terrestrial and marine organisms, especially in the latter. Therefore, the role of REE in marine organisms requires further research. One of a few reviews on REE in aquatic biota [17] summarized data on REE in various marine organisms, such as algae, mollusks, crustaceans, corals and fish. Low REE levels were found in wild corals and even lower ones were in fish. The highest content of these elements was found in algae; in addition, algae can accurately indicate the provenance of REE, seawater or sediments [45]. For example, the macroalga Gracilaria gracilis can efficiently absorb and trap REE from wastewater with low REE concentrations in it [46]. The REE accumulation in marine organisms may depend on trophic level; however, biomagnification does not appear to occur [17].
The total REE content in the seagrass Z. noltei from the Black Sea is 4–20 times lower than in the seagrass Halodule wrightii collected in Todos os Santos Bay on the east coast of Brazil [29]. It is noteworthy that the total REE content in sediments on the coast of Brazil was only 1.2–2 times higher than in the plant. This indicates that the ability of Z. noltei to accumulate REE is relatively low. Unlike Z. noltei, no significant difference in the REE contents was registered between roots and leaves of H. wrightii. The authors of [29] also concluded that seagrasses and their tissues have a low potential for REE bioaccumulation as the highest REE values in sediments at one of the stations did not correspond to the highest levels of REE accumulation in the plants.
In another seagrass species, Cymodocea nodosa, the REE contents were significantly higher in leaves and not in the underground parts of the plant, with the light REE (La, Ce, Pr, Nd) being 1.7 times higher in abundance [33]. The contents of La–Lu in C. nodosa leaves were an order of magnitude higher than those obtained in the present study. In the underground parts, the REE contents were comparable to our data with the exception of Ce, Nd, Sm and Gd, which were also higher in C. nodosa. The contents of Pr–Lu, except for Tb, in sediments were 5.5–7 times higher, and the contents of Ce and La were 9 and 1.5 times higher, respectively. It is noteworthy that the REE in rhizomes of Z. noltei were almost equal to those in C. nodosa, with the abundance in sediments being significantly lower.
The difference in the REE contents in seagrasses can be explained, among other possible reasons, by different physicochemical properties of the sediments since the REE absorption through roots is affected by the concentration of iron oxyhydroxide, pH and redox potential in the rhizosphere and cation exchange capacity of sediments. Organic and inorganic ligands also play an important role in the REE uptake by plant roots and REE speciation, and affinity for plasma membrane affects uptake of individual elements [47]. There are two different patterns of the REE distribution: in plants and in sediments, and the contents of REE in plants do not correlate with those in the rhizosphere [47] probably due to the plant control of REE uptake [48].
Some elements play an important role in the life of organisms, and they are referred to as essential macro- and micronutrients. In some cases, biochemical functions of trace elements are extremely complex as they involve interactions with other elements and can have dose-dependent effects of different signs and intensities [49]. To date, the biological role of REE remains unknown, but it was shown in some studies that they can have both negative and positive influence on animals [50,51]. As a result, it has been shown that REEs are involved in the activity of a wide range of enzymes; it was also discovered that rare earth elements, namely Ce3+ and La3+, are necessary for some bacteria to promote methanol oxidation reaction [52]. The biochemical action of rare earth elements is largely due to the closeness of their ionic radii to those of other, essential elements. For example, the Ca2+ cation can be replaced by trivalent lanthanide ions at calcium binding sites in biological molecules [51]. The largest amount of Ca2+ is contained in the plant cell wall in the form of calcium pectinate. By replacing Ca2+, REE form complexes with pectin, and their trivalency gives them a much higher charge-to-volume ratio, which means that they have a much higher affinity than Ca2+ for these binding sites [53,54]. REE are assimilated to a greater extent compared to other non-essential elements. For example, La3+ and Eu3+ were found in membranes of chloroplasts, mitochondria, cytoplasm and nuclei [47,55]. REE are evenly distributed in membranes of chloroplasts and thylakoids, where most of them are associated with the photosystem II complex [54].
3.2. REE enrichment and anomalies in sediments
Enrichment factors (EF) calculated according to Eq. (1) are shown in Figure 3. Heavy REE demonstrates considerable enrichment compared to primordial rock composition. They are above the natural variability ranges [56] whereas light REE are within or below (Ce, Nd) the geochemical limits. There are two other facets in the REE enrichment pattern that need to be emphasized. One of them is the clear manifestation of the “inverse Oddo-Harkins” effect: the elements that are more abundant in the crust demonstrate smaller enrichment factor and vice versa. The other one is a trend of the EF increase with the atomic number.
The “inverse Oddo-Harkins” pattern can be explained by the smoothened REE distribution in complex natural objects such as bottom sediments, which bear the imprint of a random interplay of various mineralogical, biological and chemical processes that affect their composition, with smoothing out the differences in the REE abundances. The EF growth in heavy REE can be the consequence of these processes as well, but it can also result from multiple cycles of element adsorption from the water column, enriched with heavy REE, by suspended organics (including phytoplankton), its subsequent sedimentation, resuspension and remobilization [57]. Because the sediments are not enriched with light REE with respect to the continental rock, the anthropogenic contribution to the REE accumulation in Kazachya Bay can be considered negligible.
Anomalies of Ce and Eu are important indicators of biogeochemical conditions in the environment [33,36]. Their most characteristic implication is the information on the surrounding redox conditions. A relative depletion in Ce, when insoluble CeO2 is reduced to soluble Ce3+, represents an example of the negative Ce anomaly and indicates the reductive conditions, whereas the relative Ce enrichment is the positive Ce anomaly that suggests the oxidative environment. The Eu anomalies are opposite to those of Ce in terms of reaction to redox conditions and are typically considered in mineralogy. In the reductive conditions, ions of Eu2+ substitute for Ca2+ in some minerals such as feldspars in hydrothermal, metamorphic and magmatic processes and form positive Eu anomaly, whereas in oxidative conditions Eu occurs in the form of Eu3+ ions, which do not have such an ability, and negative or no Eu anomaly is observed. The element anomalies can be calculated using the following relationships [33]:
and
where CeN, LaN, PrN, EuN, SmN and GdN are the mean UCC-normalized values.
δCe = CeN / (LaN × PrN)1/2
δEu = EuN / (SmN × GdN)1/2
In our case, δEu = 1.26 did not essentially differ from the values for the other REE calculated according to similar relationships (0.73–1.57), which implies the absence of any Eu anomaly. However, δCe = 0.32 was substantially lower, and this suggests the existence of negative Ce anomaly in the sediment. This anomaly is also seen in the minimum in the Ce enrichment factor in Figure 3. The negative Ce anomaly indicates the reductive environment in sediments of Kazachya Bay, which is due to the decay of organic material settling to the seafloor. A similar anomaly was detected in other sediments in reductive environments [33,58].
3.3. REE transfer and translocation in the seagrass
Transfer factors associated with the transfer from sediment to rhizomes and leaves (TFr and TFl) and the rhizomes – leaves translocation factors (TF’) are presented in Table 1. All the factors are below 1, which confirm the low efficiency of REE uptake by the plant. The sediment – rhizomes transfer factors are up to an order of magnitude higher than the sediment – leaves transfer factors, with the efficiency of the REE transfer to leaves decreasing in the following order: Sc >> Pr > Lu > Ce > Tb > Tm > Yb > Dy = Er > Ho > Eu = Gd > La > Nd = Sm > Y. This fact is at odds with the data of the work [33], where the higher REE transfer to leaves of the seagrass Cymodocea nodosa was detected.
Consider the values reciprocal to TF’ and TFr, namely the ratios of REE contents in rhizomes and leaves of Z. noltei and in the sediment and rhizomes (Figure 4). For Sc and minor REE, except for Tb, these ratios turned out to be almost pairwise equal, which fact implies the similarity of the mechanisms of their accumulation and translocation. To explain this phenomenon, it can be assumed that the differences in the REE content (∆C) between sediments and rhizomes, rhizomes and leaves are proportional to their content (C) and differences in some factor (∆q) equal for the sediment – rhizomes and rhizomes – leaves interfaces:
∆C = kC∆q
In the small differences approximation, one can pass to the differential form of Eq (4), whence it follows that
where Cs is the content in the sediment, and it is seen that equal intervals of q will change the content by an equal factor.
C = Cs exp(kq)
It is likely that the q factor is related to the local pH value (more precisely, to the difference between local pH and pH of pore water), which goes down from the pore water to the juice of the plant leaves. It is well known that the pH decrease increases the solubility of sparingly soluble salts of REE and, consequently, reduces their ability to accumulate. REE can be accumulated in seagrass tissues by binding to plant matrix carboxyl or phosphate groups close in the binding energy to carbonates and phosphates in sediments; alternatively, REE can deposit on the corresponding inorganic microgranules in tissues. This assumption finds indirect confirmation in the high correlation coefficients with calcium (median r = 0.99), which was also noted in the algae of the genus Cystoseira [59].
The changes in the Cs/Cr ratio (Figure 3) represent the well-known tendency towards an increase in the solubility of lanthanide phosphates and carbonates with the atomic number increase [60,61]. The reduced values of Cr/C1 for La–Nd and Tb can be explained by the substitution for yttrium in rhizomes and/or the formation of specific soluble complexes of these elements in leaves. On the whole, the seagrass Z. noltei does not have an advantage in the accumulation of REE compared to other native macrophytes (Cystoseira spp.) [59] due to the low translocation efficiency through the root system, which was also noted for the South American species of the seagrass Halodule wrightii [29].
3.4. Seagrass wrack as a potential source of REE
Availability and extractability of REE from various natural sources other than minerals is an important issue associated with the commercial use of REE in the future [62]. Currently, the ever-increasing number of applications of REE and high added value of REE-containing products have led to an increase in the REE production volumes. Therefore, research on minimizing industrial production losses and more cost-effective extraction of REE with the least environmentally-unfriendly consequences is in an increasing demand [63]. As a result of processing algal and seagrass biomass, including storm-cast deposits (beach wrack), it is possible to produce various types of biofuels, including biodiesel, bioethanol, biogas, biohydrogen and other valuable products [64]. As storm-cast beach wreck of algae and seagrass can be also a source of REE and bioremediate industrial wastewater, further research in this direction is strongly needed.
The Black Sea has extensive natural resources of macrophytes, including seagrasses. In late summer and early autumn, seagrasses shed their leaves and after each storm their mass deposits appear on the coastline (Figure 5). They are most widespread in Kerch Strait and in Tendra, Dzharylgach, Yegorlyk and Karkinit bays.
An assessment of storm-cast deposits of the seagrass as a possible source of REE can be made. According to our data, the average REE concentration in seagrass leaves is 0.4 mg·kg–1 w.w. After only one storm, the seagrass biomass washed ashore can be estimated at 10 tons per linear kilometer of the coast of the southern part of Karkinit Bay [65]. This biomass contains approximately 4 kg of rare earth elements. Estimating the reserves of the deposits of the seagrass on the Black Sea shoreline at hundreds of thousands of tons [66], one can arrive at REE reserves in the coastal seagrass deposits of up to 100 tons. However, a comprehensive environmental analysis is clearly needed for the large-scale use of the deposits of macrophytes for the extraction of REE and other useful materials from them.
4. Conclusions
Various factors in marine ecosystems (carbon source and content, particle size distribution, pH, salinity) play a fundamental role in the biogeochemical cycles of REE and determine their bioavailability and accumulation in sediments. Studies in local ecosystems make a significant contribution to the understanding of these processes. As the example, the significantly lower REE concentrations in Z. noltei compared to their content in the sediments demonstrate the low ability of the seagrass to accumulate these elements, which feature may also be related to the characteristics of marine sediments in this area.
Ratios of the contents of Sc and minor REE, except for Tb, in rhizomes and leaves of Z. noltei and in sediments and rhizomes turned out to be very close, which probably indicate the similarity of mechanisms of the REE accumulation and translocation and are possibly associated with the difference between the local pH value and the pH of pore waters. This work represents the first quantitative determination of a complete set of rare earth elements (except for promethium) in the seagrass Z. noltei and in the bottom sediments surrounding it. The obtained results can be used to further study REE in the Black Sea ecosystems and to monitor these elements in the marine environments.
Author Contributions
Conceptualization, V.R.; Data curation, E.G., S.K.; Methodology, S.K., N.B.; Validation, S.K.; Writing—original draft, S.K., E.G.; Writing—review and editing, V.R., S.B. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Russian Science Foundation grant #23-24-00494 (https://rscf.ru/project/23-24-00494/).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Acknowledgments
We gratefully acknowledge the service of the Spectrometry and Chromatography core facility within IBSS RAS for the ICP-MS analysis and are thankful to A.M. Toichkin for his help in collecting seagrass samples.
Conflicts of Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
- Damhus, T.; Hartshorn, R.M.; Hutton, A.T. Nomenclature of Inorganic Chemistry: IUPAC Recommendations; RSC Publishing: Cambridge, UK, 2005; p. 51. [Google Scholar]
- Castor, S.B.; Hedrick, J.B. Rare earth elements. Industrial minerals and rocks 2006, 7, 769–792. [Google Scholar]
- Balaram, V. Rare earth elements: A review of applications, occurrence, exploration, analysis, recycling, and environmental impact. Geosci. Front. 2019, 10, 1285–1303. [Google Scholar] [CrossRef]
- Du, X.; Graedel, T.E. Global in-use stocks of the rare earth elements: a first estimate. Environ. Sci. Technol. 2011, 45, 4096–4101. [Google Scholar] [CrossRef] [PubMed]
- Sinharoy, A.; Lens, P. Environmental technologies to treat rare earth element pollution: Principles and Engineering; IWA Publishing: London, 2022. [Google Scholar] [CrossRef]
- Brown, P.H.; Rathjen, A.H.; Graham, R.D.; Tribe, D.E. Rare earth elements in biological systems. In Handbook on the physics and chemistry of rare earths; Gschneidner, K.A., Eyring, L., Eds.; Elsevier Science Publishers: Amsterdam, 1990; Volume 13, pp. 423–452. [Google Scholar] [CrossRef]
- Tyler, G. Rare earth elements in soil and plant systems: a review. Plant Soil 2004, 267, 191–206. [Google Scholar] [CrossRef]
- Loell, M.; Reiher, W.; Felix-Henningsen, P. Contents and bioavailability of rare earth elements in agricultural soils in Hesse (Germany). J. Plant Nutr. Soil Sci. 2011, 174, 644–654. [Google Scholar] [CrossRef]
- Das, N.; Das, D. Recovery of rare earth metals through biosorption: an overview. J. Rare Earths 2013, 31, 933–943. [Google Scholar] [CrossRef]
- Sparks, D.L. Toxic metals in the environment: the role of surfaces. Elements 2005, 1, 193–197. [Google Scholar] [CrossRef]
- Reddy, V.M.; Babu, K.S.; Balaram, V.; Satyanarayanan, M. Assessment of the effects of municipal sewage, immersed idols and boating on the heavy metal and other elemental pollution of surface water of the eutrophic Hussainsagar lake (Hyderabad, India). Environ. Monit. Assess. 2012, 184, 1991–2000. [Google Scholar] [CrossRef]
- Wei, B.; Li, Y.; Li, H.; Yu, J.; Ye, B.; Liang, T. Rare earth elements in human hair from a mining area of China. Ecotoxicol. Environ. Saf. 2013, 96, 118–123. [Google Scholar] [CrossRef]
- Ali, S.H. Social and environmental impact of the rare earth industries. Resources 2014, 3, 123–134. [Google Scholar] [CrossRef]
- Vysetti, B. Current advances in the miniaturization of analytical instruments—Applications in cosmochemistry, geochemistry, exploration, and environmental sciences. Spectroscopy 2016, 31, 40–44. [Google Scholar] [CrossRef]
- RIVM Report 601501011: Maximum permissible concentrations and negligible concentrations for rare earth elements (REEs); Sneller F.E.C.; Kalf D.F.; Weltje L.; Van Wezel A.P., Eds.; Bilthoven, Netherlands, 2000; 60 p. (National Institute of Public Health and Environmental Protection RIVM).
- Al-Rimawi, F.; Kanan, K.; Qutob, M. Analysis of different rare metals, rare earth elements, and other common metals in groundwater of South West Bank/Palestine by ICP/MS-Data and health aspects. J. Environ. Protect. 2013, 4, 1157–1164. [Google Scholar] [CrossRef]
- Neira, P.; Romero-Freire, A.; Basallote, M.D.; Qiu, H.; Cobelo-García, A.; Canovas, C.R. Review of the concentration, bioaccumulation, and effects of lanthanides in marine systems. Front. Mar. Sci. 2022, 9, 920405. [Google Scholar] [CrossRef]
- Khan, A.M.; Bakar, N.K.A.; Bakar, A.F.A.; Ashraf, M.A. Chemical speciation and bioavailability of rare earth elements (REEs) in the ecosystem: a review. Environ. Sci. Pollut. Res. 2017, 24, 22764–22789. [Google Scholar] [CrossRef]
- Merschel, G.; Bau, M. Rare earth elements in the aragonitic shell of freshwater mussel corbicula fluminea and the bioavailability of anthropogenic lanthanum, samarium and gadolinium in river water. Sci. Total Environ. 2015, 533, 91–101. [Google Scholar] [CrossRef]
- Armand, R.; Cherubini, C.; Tuduri, J.; Pastore, N.; Pourret, O. Rare earth elements in French stream waters - revisiting the geochemical continental cycle using FOREGS dataset. J. Geochem. Explor. 2015, 157, 132–142. [Google Scholar] [CrossRef]
- Schijf, J.; de Baar, H.J.W. Rare earth element exchange through the Bosporus: The black Sea as a net source of REEs to the Mediterranean Sea. Geochim. Cosmochim. Acta 1995, 59, 3503–3509. [Google Scholar] [CrossRef]
- den Hartog, C. Seagrasses and seagrass ecosystem, an appraisal of the research approach. Aquat. Bot. 1979, 7, 105–117. [Google Scholar] [CrossRef]
- Schlacher-Hoenlinger, M.A.; Schlacher, T.A. Differential accumulation patterns of heavy metals among the dominant macrophytes of a Mediterranean seagrass meadow. Chemosphere 1998, 37, 1511–1519. [Google Scholar] [CrossRef]
- Govers, L.L.; Lamers, L.P.; Bouma, T.J.; Eygensteyn, J.; de Brouwer, J.H.; Hendriks, A.J.; Huijbers, C.M.; van Katwijk, M.M. Seagrasses as indicators for coastal trace metal pollution: a global meta-analysis serving as a benchmark, and a Caribbean case study. Environ. Pollut. 2014, 195, 210–217. [Google Scholar] [CrossRef]
- Deniz, F. Bioremediation potential of waste biomaterials originating from coastal Zostera marina L. meadows for polluted aqueous media with industrial effluents. Prog. Biophys. Mol. Biol. 2019, 145, 78–84. [Google Scholar] [CrossRef]
- Lee, G.; Suonan, Z.; Kim, S.H.; Hwang, D.-W.; Lee, K.-S. Heavy metal accumulation and phytoremediation potential by transplants of the seagrass Zostera marina in the polluted bay systems. Mar. Pollut. Bull. 2019, 149, 110509. [Google Scholar] [CrossRef]
- Serrano, O.; Lavery, P.S.; Bongiovanni, J.; Duarte, C.M. Impact of seagrass establishment, industrialization and coastal infrastructure on seagrass biogeochemical sinks. Mar. Environ. Res. 2020, 160, 104990. [Google Scholar] [CrossRef]
- Serrano, R.; Gras, L.; Giménez-Casalduero, F.; del-Pilar-Ruso, Y.; Grindlay, G.; Mora, J. The role of Cymodocea nodosa on the dynamics of trace elements in different marine environmental compartments at the Mar Menor Lagoon (Spain). Mar. Pollut. Bull. 2019, 141, 52–60. [Google Scholar] [CrossRef]
- Sena, I.C.M.; Souza, L.A.; Patire, V.F.; Arias-Ortiz, A.; Creed, J.C.; Cruz, I.; Hatje, V. Environmental settings of seagrass meadows control rare earth element distribution and transfer from soil to plant compartments. Sci. Total Environ. 2022, 843, 157095. [Google Scholar] [CrossRef]
- Chemnitzer, R. Strategies for achieving the lowest possible detection limits in ICP-MS. Spectroscopy 2019, 34, 12–16. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Mégevand, P. Games-Howell post-hoc test for one-way ANOVA. Available online: https://github.com/pierremegevand/games_howell (accessed on 17 September 2023).
- Komar, D.; Smuc, N.R.; Belak, Z.L.; Matešić, S.; Lojen, S.; Kniewald, G.; Vrhovnik, P.; Dolenec, T.; Dolenec, M. Geochemical characteristics and distribution of rare earth elements in makirina bay sediments (n. Dalmatia, Republic of Croatia). Geologica Macedonica 2014, 28, 127–137. [Google Scholar]
- Taylor, S.R.; McLennan, S.M. The Continental Crust: Its Composition and Evolution; Blackwell: Oxford, 1985. [Google Scholar]
- Cotton, S.A. Scandium, yttrium & the lanthanides: Inorganic & coordination chemistry. In Encyclopedia of Inorganic Chemistry; Wiley: New York, 2006; the lanthanides: Inorganic &. [Google Scholar] [CrossRef]
- Mao, L.; Mo, D.; Yang, J.; Guo, Y.; Lv, H. Rare earth elements geochemistry in surface floodplain sediments from the Xiangjiang River, middle reach of Changjiang River, China. Quaternary International 2014, 336, 80–88. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, C.L. Riverine composition and estuarine geochemistry of particulate metals in China – weathering features, anthropogenic impact and chemical fluxes. Estuarine, Coastal and Shelf Science 2002, 54, 1051–1070. [Google Scholar] [CrossRef]
- Dean, J.R. Bioavailability, Bioaccessibility and Mobility of Environmental Contaminants; Wiley: London, 2007. [Google Scholar]
- Zhang, Y.; Gao, X. Rare earth elements in surface sediments of a marine coast under heavy anthropogenic influence: The Bohai Bay, China. Est. Coast. Shelf Sci. 2015, 164, 86–93. [Google Scholar] [CrossRef]
- Xie, H.G. Advance of application of rare earth elements in agriculture in China. Chin. Sci. Bull. 1991, 36, 561–564. [Google Scholar]
- Akagi, T.; Edanami, K. Sources of rare earth elements in shells and soft-tissues of bivalves from Tokyo bay. Mar. Chem. 2017, 194, 55–62. [Google Scholar] [CrossRef]
- Li, J.X.; Zheng, L.; Sun, C.J.; Jiang, F.H.; Yin, X.F.; Chen, J.H.; Han, B.; Wang, X. Study on ecological and chemical properties of rare earth elements in tropical marine organisms. Chin. J. Anal. Chem. 2016, 44, 1539–1546. [Google Scholar] [CrossRef]
- Akagi, T. Rare earth element (REE)-silicic acid complexes in seawater to explain the incorporation of REEs in opal and the “leftover” REEs in surface water: New interpretation of dissolved REE distribution profiles. Geochim. Cosmochim. Acta 2013, 113, 174–192. [Google Scholar] [CrossRef]
- Wieszczycka, K.; Staszak, K.; Wozniak-Budych, M.J.; Jurga, S. Lanthanides and tissue engineering strategies for bone regeneration. Coord. Chem. Rev. 2019, 388, 248–267. [Google Scholar] [CrossRef]
- Fu, F.; Akagi, T.; Yabuki, S.; Iwaki, M.; Ogura, N. Distribution of rare earth elements in seaweed: Implication of two different sources of rare earth elements and silicon in seaweed. J. Phycology 2000, 36, 62–70. [Google Scholar] [CrossRef]
- Jacinto, J.; Henriques, B.; Duarte, A.C.; Vale, C.; Pereira, E. Removal and recovery of critical rare elements from contaminated waters by living Gracilaria gracilis. J. Hazard. Mater. 2018, 344, 531–538. [Google Scholar] [CrossRef]
- Kovařiková, M.; Tomášková, I.; Soudek, P. Rare earth elements in plants. Biol. Plant. 2019, 63, 20–32. [Google Scholar] [CrossRef]
- Tyler, G.; Olsson, T. Rare earth elements in forest-floor herbs as related to soil conditions and mineral nutrition. Biol. Trace Element Res. 2005, 106, 177–191. [Google Scholar] [CrossRef]
- Gumienna-Kontecka, E.; Rowińska-Zyrek, M.; Luczkowski, M. The role of trace elements in living organisms. In Recent Advances in Trace Elements; Chojnacka, K., Saeid, A., Eds.; Wiley-Blackwell, 2018; pp. 177–206. [Google Scholar] [CrossRef]
- Blinova, I.; Lukjanova, A.; Muna, M.; Vija, H.; Kahru, A. Evaluation of the potential hazard of lanthanides to freshwater microcrustaceans. Sci. Total Environ. 2018, 642, 1100–1107. [Google Scholar] [CrossRef]
- Jalali, J.; Lebeau, T. The role of microorganisms in mobilization and phytoextraction of rare earth elements: A review. Front. Environ. Sci. 2021, 9. [Google Scholar] [CrossRef]
- Hibi, Y.; Asai, K.; Arafuka, H.; Hamajima, M.; Iwama, T.; Kawai, K. Molecular structure of La3+-induced methanol dehydrogenase-like protein in Methylobacterium radiotolerans. J. Biosci. Bioeng. 2011, 111, 547–549. [Google Scholar] [CrossRef] [PubMed]
- Evans, C.H. Interesting and useful biochemical properties of lanthanides. Trends Biochem. Sci. 1983, 8, 445–449. [Google Scholar] [CrossRef]
- Lai, Y.; Wang, Q.; Yang, L.; Huang, B. Subcellular distribution of rare earth elements and characterization of their binding species in a newly discovered hyperaccumulator Pronephrium simplex. Talanta 2006, 70, 26–31. [Google Scholar] [CrossRef]
- Gao, Y.; Zeng, F.; Yi, A.; Ping, S.; Jing, L. Research of the entry of rare earth elements Eu3+ and La3+ into plant cell. Biol. Trace Element Res. 2003, 91, 253–265. [Google Scholar] [CrossRef]
- Grousset, F.E.; Quetel, C.R.; Thomas, B.; Donard, O.F.X.; Lambert, C.E.; Guillard, F.; Monaco, A. Anthropogenic vs. lithogenic origins of trace elements (As, Cd, Pb, Rb, Sb, Sc, Sn, Zn) in water column particles: northwestern Mediterranean Sea. Mar. Chem. 1995, 48, 291–310. [Google Scholar] [CrossRef]
- McLennan, S.M. Rare earth elements in sedimentary rocks: influence of provenance and sedimentary processes. In Geochemistry and Mineralogy of Rare Earth Elements; Lipin B.R., McKay G.A., Eds.; Mineral Soc. Am., Washington, 1989; pp. 169–200. [CrossRef]
- Hannigan, R.; Dorval, E.; Jones, C. The rare earth element chemistry of estuarine surface sediments in the Chesapeake Bay. Chem. Geol. 2010, 272, 20–30. [Google Scholar] [CrossRef]
- Ryabushko, V.I. , Gureeva, E.V.; Kapranov, S.V.; Bobko, N.I.; Prazukin, A.V.; Nekhoroshev, M.V. Rare earth elements in brown algae of the genus Cystoseira (Phaeophyceae) (Black Sea). Eur. J. Phycol. 2022, 57, 433–445. [Google Scholar] [CrossRef]
- Caro, P.; Lemaître-Blaise, M.; Trombe, F. Identification et solubilités des phases solides à l’équilibre sous une atmosphère de gaz carbonique dans les systèmes temaires oxydes de terres rares-gaz carbonique-eau. Comptes rendus hebdomadaires des séances l’Académie des sciences. Série C 1968, 267, 1594–1597. [Google Scholar]
- Liu, X.; Byrne, R.H. Rare earth and yttrium phosphate solubilities in aqueous solution. Geochim. Cosmochim. Acta 1997, 61, 1625–1633. [Google Scholar] [CrossRef]
- Tse, P.-K. China’s rare-earth industry 2011, U.S. Geological Survey Open-File Report 2011–1042. Available online: https://pubs.usgs.gov/of/2011/1042 (accessed on 17 September 2023).
- Goecke, F.; Zachleder, V.; Vítová, M. Rare earth elements and algae: physiological effects, biorefinery and recycling. Algal Biorefineries 2015, 2, 339–363. [Google Scholar] [CrossRef]
- Behera, S.; Singh, R.; Arora, R.; Kumar, S.N.; Shukla, M.; Kumar, S. Scope of algae as third generation biofuels. Front. Bioeng. Biotechnol. 2015, 2. [Google Scholar] [CrossRef] [PubMed]
- Sadogurskiy, S.Y.; Belich, T.V.; Sadogurskaya, S.A. The Red Data Book of Crimea and macrophytes stormwrack: how to deal with it? Plant Biology and Horticulture: theory, innovation 2022, 3, 43–49. (In Russian) [Google Scholar] [CrossRef]
- Milchakova, N.A. On the status of seagrass communities in the Black Sea. Aquat. Bot. 1999, 65, 21–32. [Google Scholar] [CrossRef]
Figure 1.
Map of the sampling area (inset). The sampling site is denoted by black circle.
Figure 2.
Dendrogram of the Euclidean distances of log-transformed REE contents in rhizomes and leaves of Zostera noltei.
Figure 2.
Dendrogram of the Euclidean distances of log-transformed REE contents in rhizomes and leaves of Zostera noltei.
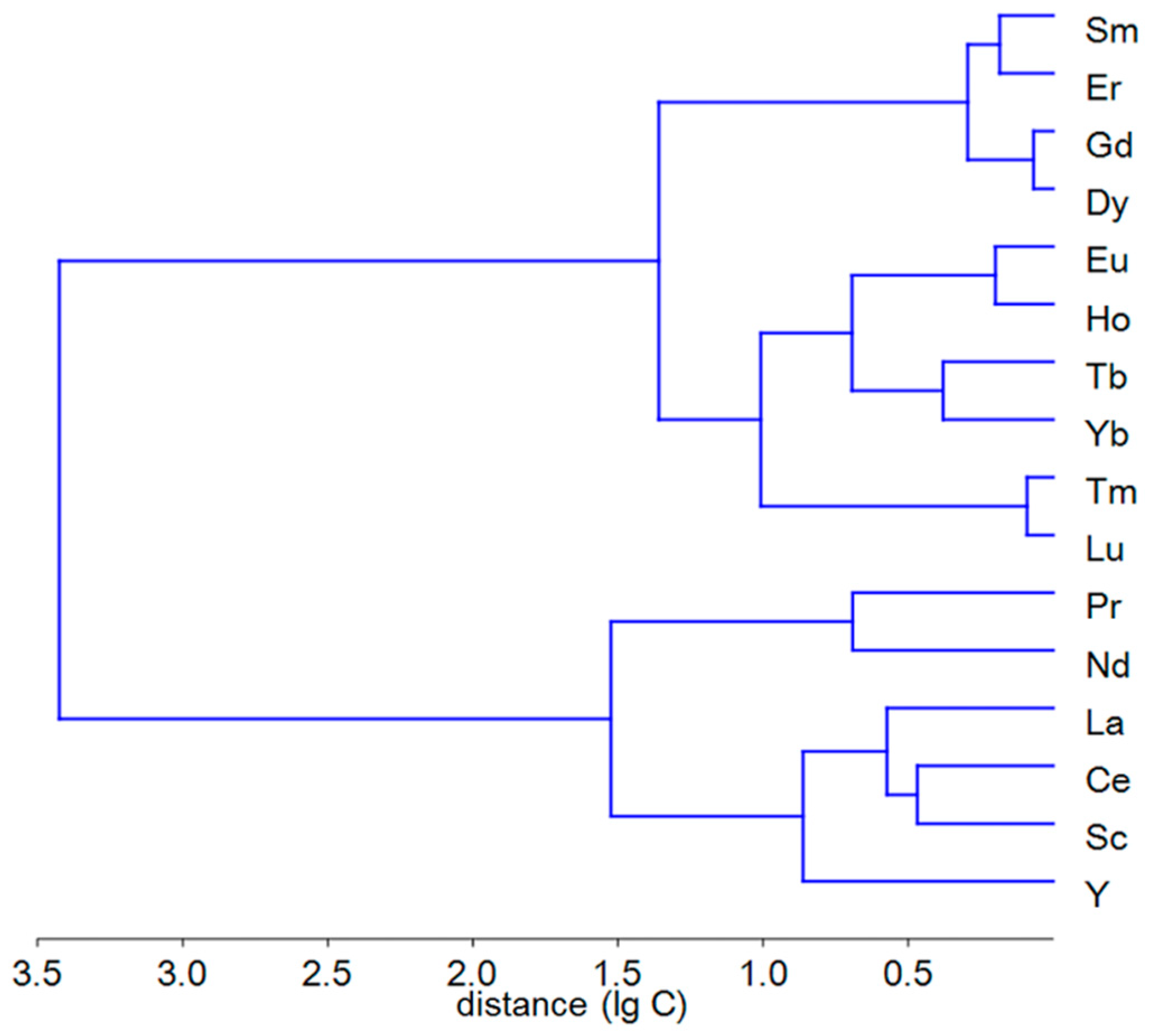
Figure 3.
Enrichment factors of REE in the sediments in Kazachya Bay with respect to the upper continental crust.
Figure 3.
Enrichment factors of REE in the sediments in Kazachya Bay with respect to the upper continental crust.
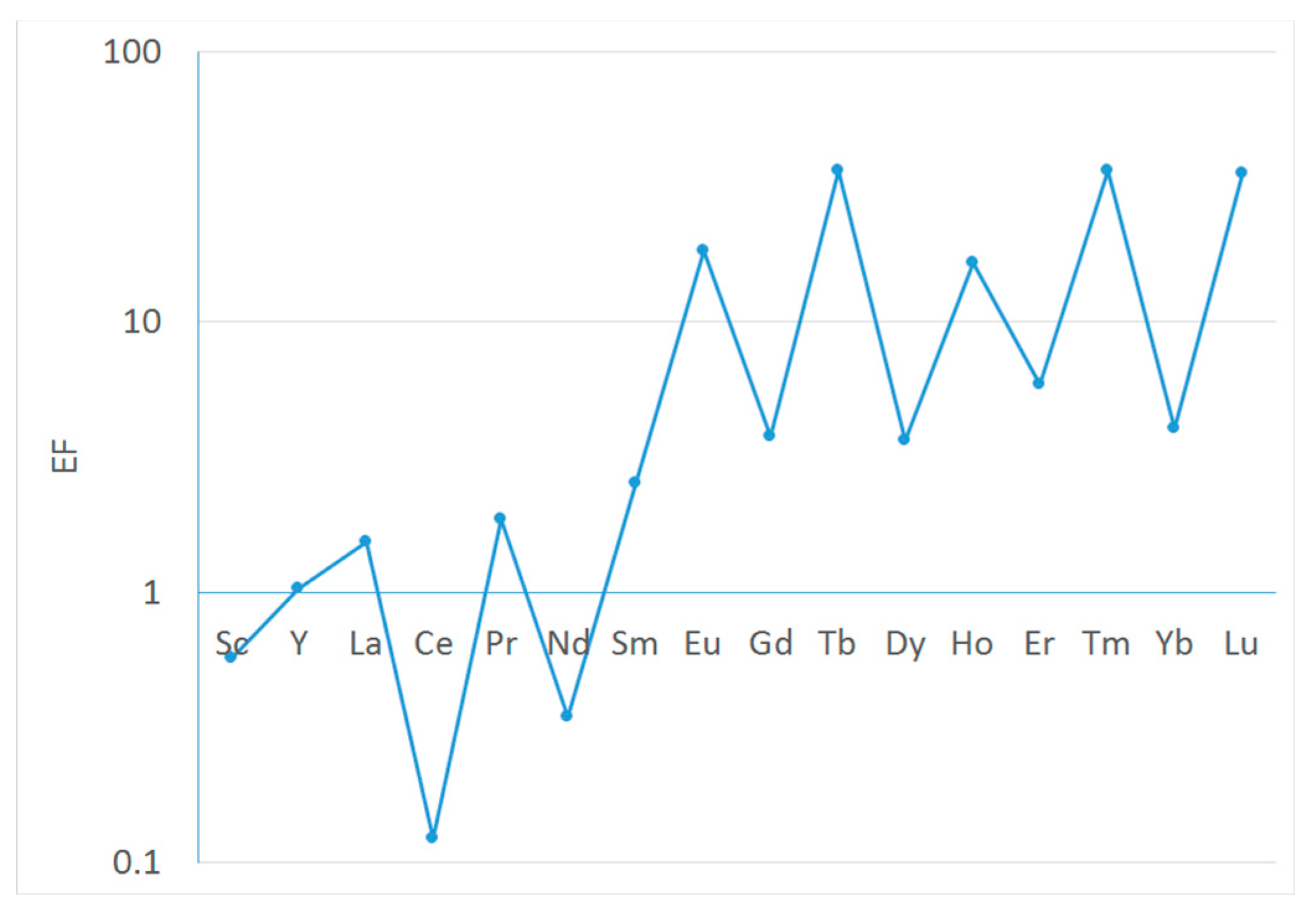
Figure 4.
Ratios between the REE contents in sediments and rhizomes (Cs/Cr) and in rhizomes and leaves (Cr/Cl) of Zostera noltei.
Figure 4.
Ratios between the REE contents in sediments and rhizomes (Cs/Cr) and in rhizomes and leaves (Cr/Cl) of Zostera noltei.
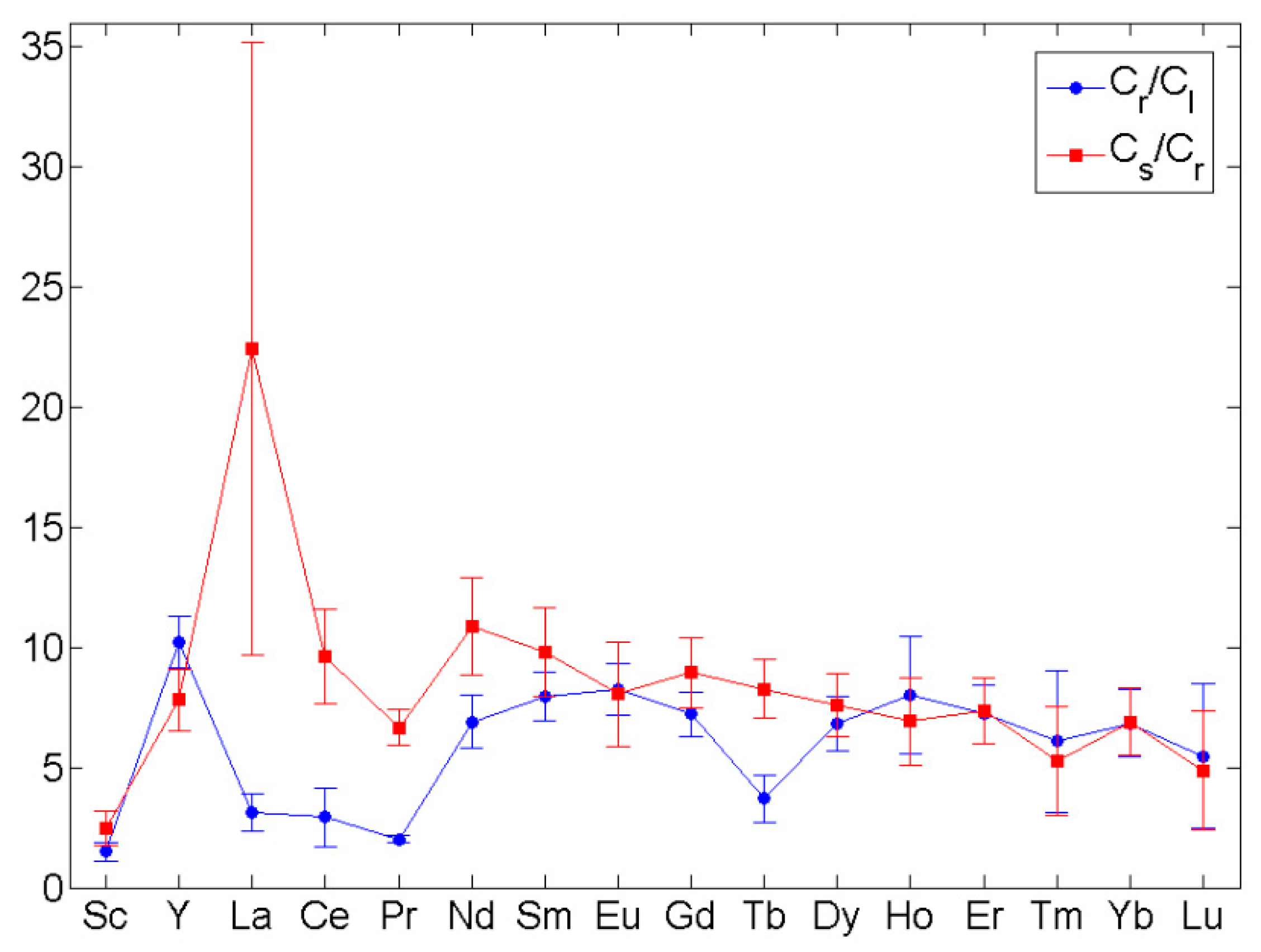
Figure 5.
Seagrass deposits on the Black Sea coast after storm in (a) Kazachya Bay, (b) Karkinit Bay near the Portovoye settlement.
Figure 5.
Seagrass deposits on the Black Sea coast after storm in (a) Kazachya Bay, (b) Karkinit Bay near the Portovoye settlement.


Table 1.
REE contents (in μg·kg–1 d.w.) in leaves and rhizomes of Zostera noltei and in sediments: mean ± SD; transfer factors for rhizomes (TFr) and leaves (TFl); and the rhizomes – leaves translocation factors (TF’). The different upper subscript letters denote significant differences between the element contents in the objects of research: a < b < c.
Table 1.
REE contents (in μg·kg–1 d.w.) in leaves and rhizomes of Zostera noltei and in sediments: mean ± SD; transfer factors for rhizomes (TFr) and leaves (TFl); and the rhizomes – leaves translocation factors (TF’). The different upper subscript letters denote significant differences between the element contents in the objects of research: a < b < c.
| Z. noltei (leaves) | Z. noltei (rhizomes) | Sediments | TFr | TFl | TF’ | |
|---|---|---|---|---|---|---|
| Sc | 373±22a | 560±15a | 1382±77b | 0.405 | 0.270 | 0.666 |
| Y | 153±9a | 1562±18b | 12221±646c | 0.128 | 0.012 | 0.098 |
| La | 243±26a | 761±112a | 17074±614b | 0.045 | 0.014 | 0.320 |
| Ce | 221±9a | 648±25b | 6232±89c | 0.104 | 0.035 | 0.341 |
| Pr | 87±3a | 174±11b | 1162±18c | 0.150 | 0.075 | 0.499 |
| Nd | 38±3a | 267±49b | 2894±37c | 0.092 | 0.013 | 0.145 |
| Sm | 8.2±0.4a | 65±6b | 635±19c | 0.102 | 0.013 | 0.126 |
| Eu | 2.6±0.4a | 21.7±1.3b | 175±6c | 0.124 | 0.015 | 0.122 |
| Gd | 10.4±0.3a | 75±6b | 671±16c | 0.112 | 0.015 | 0.138 |
| Tb | 6±1a | 22±3b | 184±8c | 0.121 | 0.033 | 0.270 |
| Dy | 11±1a | 72±7b | 551±27c | 0.131 | 0.019 | 0.146 |
| Ho | 2.4±0.3a | 19±1b | 131±4c | 0.144 | 0.018 | 0.125 |
| Er | 7.3±0.6a | 53±4b | 386±13c | 0.136 | 0.019 | 0.139 |
| Tm | 1.5±0.1a | 9±2b | 49±2c | 0.190 | 0.031 | 0.164 |
| Yb | 5.1±0.7a | 35±2b | 241±13c | 0.145 | 0.021 | 0.146 |
| Lu | 1.7±0.2a | 9.3±3.4b | 45±2c | 0.206 | 0.038 | 0.183 |
| ΣREE | 1171.2 | 4353 | 44033 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



