
Submitted:
20 September 2023
Posted:
21 September 2023
You are already at the latest version
Abstract
Cobalt (Co) is a transition metal and is classified as a beneficial metal for plants, but its benefits for plants remain obscure. Cobalt has been reported to negatively affect plant physiochemical processes both at higher (toxic) and lower (deficient) applied levels. High concentrations of cobalt in plant organs cause irreversible changes to the plant cells, mainly via enhanced production of reactive radicals. Moreover, Co and its compound play an essential role in humans as they are the central atoms of cobalamin, a co-enzyme precursor whose absence causes anemia. Hence, the optimum Co supply to plants is critical for customary plant metabolic workings. Henceforth, monitoring Co behavior in the soil-plant-human system is highly imperative.
This review highlighted the latest literature on (i) Co in soil and plants; (ii) its mobility and phyto-availability in soil; (iii) phytouptake and translocation towards shoot tissues; (iv) toxic and deficient effects of Co on plants; (v) plant detoxification mechanisms under increased Co levels inside plants; and (v) its role in the human body. For this purpose, 1026 plant observations from literature data were analyzed related to Co biogeochemical behavior in the soil-plant system. The data analysis revealed an overall increase in 567 observations and a decrease in 381 observations. However, these general trends in plant responses vary significantly for different types of plant species and physiological attributes. Overall, the current review delineates an updated and critical representation of the biogeochemical behavior of Co in the soil-plant-human system, supported by up-to-date 9 main tables, 14 supporting tables, and 3 figures. The authors believe that the literature presented here can be of great interest to scientists, researchers, policymakers, and graduate-level students.
Keywords:
Cobalt
; toxicity
; plants
; detoxification
; micronutrient
; macronutrients
; anti-oxidative enzymes
1. Introduction
Cobalt (Co) is classified as a transition/heavy/trace metal. It is considered a beneficial component to the metabolism of all animals, including humans, and the prokaryotes [1]. This metal is a key constituent of cobalamin, also identified as vitamin B12, which is required for human and animal nutrition [2]. Despite beneficial physiological and biological roles in legumes, Co has not been classified as an essential nutrient for plant growth [3]. In legumes, Co is used by microorganisms to fix atmospheric nitrogen [3]. Cobalt is also reported to have some beneficial effects on plant growth [4]. Hence an optimum supply of Co can be crucial to withstand the regular metabolic functions without showing toxicity or deficiency symptoms. Nevertheless, Co can be highly toxic to plant metabolism above threshold levels. Thus, Co renders a severe threat to plant physiological and growth attributes at high levels in soils. Keeping in view the beneficial and toxic effects of Co on plants, it is worthwhile to study its biogeochemical behavior in soil-plant system.
Cobalt is considered somewhat mobile in soil. Owing to its potential mobility in soil, Co is easily up taken by plant roots. Inside plants, Co is considered immobile or relatively less mobile, therefore, elevated levels of Co are reported in root tissues. High concentrations of cobalt in plants are noxious and damaging to physiological, and biochemical functions in plants [5]. Cobalt affects the availability of essential nutrients, disrupts the structural integrity of chloroplasts, limits the net photosynthesis process, and decreases the iron-containing enzyme activity [3,6]. Studies have revealed numerous toxicity symptoms to Co stress, including leaf chlorosis, and reduced plant [3]. Cobalt stress also induces enhanced levels of reactive radicles such as reactive oxygen species (ROS), thereby initiating and triggering numerous chain reactions and causing damage to various essential macromolecules such as lipids, proteins, and DNA [7]. Under excess Co exposure, the degradation of enzymatic machinery took place leading to severe phytotoxicity [7]. In addition, Co possesses a high binding potential with sulfhydryl groups by interacting with their thiol group (-SH). This interaction generally causes glutathione (GSH) depletion, thereby limiting radicle scavenging potential of plants.
To cope with Co and other toxic metal toxicity, plants are equipped with a series of tolerance and adaptation mechanisms. Firstly, plants minimize metal/Co transfer to roots by secreting organic acids and making them immobile and stable in soil [7]. However, the amount of absorbed Co is detoxified by binding via numerous ligands and sequestration in different cellular compartments such as vacuoles [8]. Moreover, the stimulation of enzymatic antioxidant and enhanced production of organic ligands inside plant cells [GSH and phytochelatins (PCs)] is one the most effective mechanism to detoxify and tolerate Co as well as Co-induced ROS [7,8]. Nevertheless, different plant species vary greatly with respect to their potential to tolerate Co stress and scavenge ROS.
Considering the beneficial and toxic roles of Co on plant biochemistry, it is imperative to understand the environmental levels of Co and its behavior in soil-plant system. However, there is limited data on plant biochemistry under Co stress compared to other trace metals such as lead, cadmium, arsenic, zinc, chromium, etc. Moreover, there is scarcity of review articles which critically elucidate and summarize the prevailing trends/comparisons among the physicochemical roles of Co below and above toxic levels in plants. Hence, data-analysis of latest scientific literature related to biogeochemistry of Co in soil-plant interaction can be of great importance. This review critically reveals latest findings about (i) Co mobility/phytoavailability in soil; (ii) soil-plant-human transfer of Co; (iii) Co toxicity to plants; (iv) Co deficiency in plants (v) Co detoxification mechanisms in plants; and (v) human health effects of Co.
2. Cobalt minerals and concentration in soil
Cobalt is a naturally occurring compound is a constituent of numerous naturally occurring minerals such as hydrates, sulfides, and oxides. Commonly occurring Co ores include erythrite [Co3(AsO4)2.8H2O)], cobaltite (CoAsS), linnealite (Co3S4), smaltite (Co(Ni)As3), safflorite (CoAs2), and spherocobaltite (CoCO3) [9]. Cobalt chalcophiles manifest themselves to exist as sulfide ore in the Earth’s crust, while its siderophility form has limited affinity to sulfur and oxygen. Cobalt is also present in silicate layers of Earth’s crust indicating the lithophilic properties. Alkaline igneous rocks are considered to be the largest reservoirs of Co. The Co concentration in soil profiles and its distribution is mainly dependent upon the soil forming processes, therefore, diffing from soils of various agroclimatic regions. The Mediterranean Basin countries in Europe contain a higher level of Co compared to Northern Europe due to the presence of ophiolitic rocks (ultramafic and mafic) [10].
Cobalt is considered a rarely occurring trace metal in the earth’s crust (33rd most abundant element in nature) [11]. The natural Co content of the soil is up to 40 mg/kg [12]. The mean level of cobalt in world soils is 8 mg/kg [11]. However, its contents may differ from 0.1 to 100 mg/kg in different soils [11]. Cobalt average levels reported in different soils include 5.5 mg/kg for sandy podsol soils, 4.4 mg/kg for organic soils, 10–12 mg/kg in dark brown clay soils, limestone soils, and fen soils [11]. Studies from different regions have reported different ranges of Co in soil such as 16.5–26.8 mg/kg in Egyptian soils, 116 mg/kg in soil of Japan, 122 mg/kg in Australian ferralsols, and 12,700 mg/kg in the United States [11,13].
The maximum allowable level of Co in the soil is 50 mg/kg [14]. The exceeding Co concentration from permissible limit in the soil primarily results from both natural and human activities, late being more prominent. The continuous use of Co in various industrial applications has enhanced its concentration to toxic levels at several sites in recent years. Major anthropogenic sources of Co in soil include mining of ores, metal industrial activities, electronic waste recycling, painting, pigment production, agrochemicals (fertilizers and pesticides), and sewage sludge dumping [13,14,15].
According to the USGS, a total of 1,665,000 metric tons of Co has been mined during 2010-2022 with a mean value of 128,077. In 2022, about 190,000 metric tons of Co was mined and used in different industrial processes (Figure 1). Moreover, Congo, Russia, Canada, Australia, Cuba, Madagascar, the Philippines, Zambia, Papua New Guinea, New Caledonia, and South Africa are among the top producers of cobalt. These countries also possess the major reserves of Co, Congo being at the top with 3500000 metric tons. The mining processes have several folds increased the concertation of cobalt in surface soil. Cobalt concentrations in the soil of different countries of the world collected from mining, smelting, and industrial areas are presented in Table 1.
2.1. Cobalt speciation and bioavailability in soil: role of soil characteristics
Chemical speciation (oxidation forms) of a metal in soil governs its biogeochemical behavior in the soil-plant continuum [16,17]. Recent data believe that compared to total soil contents, chemical speciation of a metal(loid) is a better indicator of its mobility and phytoavailability in the soil as well as the uptake and translocation in plants [18]. Cobalt exists in the environment in four major oxidation forms (+1 to +4) [14]. Most often, Co exists as +2, +3, and rarely as a +1-oxidation state.
The Eh potential and pH are the two most crucial soil dynamics that influence the speciation of Co metal in soils [15]. In soil, cobalt frequently transforms from Co2+ to Co3+ primarily due to Mn oxides. Different redox processes, such as (i) Co2+ to Co3+ oxidation at the surface of oxides (ii) Mn4+ to Mn3+ reduction in the oxide crystal lattice, and (iii) Mn4+ or Mn3+ by Co3+ replacement, govern the sorption of Co on Mn and Fe oxides [19]. The amount of Co that is absorbed by the minerals Mn and Fe increases when soil pH rises. However, the soil redox potential has a major role in controlling the sorption of Co on Mn/Fe oxides. In soil solutions, Co exists mostly in the ionic forms Co3+, Co2+, Co(OH)3, CoOH+, and complex compounds as Co(OH)2° [20].
Generally, 95% of the total Co concentration in soil remains immobile [11]. In majority of the soils, Co level in soil pore water varies from 0.3 to 87 μg/L. Cobalt is adsorbed on different soil constituents (e.g., clay minerals) due to chemical methods and ion exchange processes [15]. In this regard, montmorillonite has a higher sorption capacity for Co than other clay minerals. Application of organic fertilizers in the soil causes little increase in Co concentration in soil pore water. The complexation of Co by different chelates in soil also enhances its soil mobility.
By increasing soil acidity, cobalt mobility in soil rises [21]. Similarly, Anaerobic soil conditions can increase the mobility of Co soil [21]. The suppression of valence bonding with Mn and Fe that causes this interaction. Due to the divalent Co and Mn ions' low potentials for complex formation, this contact does not, however, cause the outer hydration to split [22]. Co mobility in soil is also enhanced by cobalt in chelate-complexed soil. High quantities of Co can be retained in soils with high levels of organic matter and Fe/Mn oxides. Fe/Mn oxides' strong retention/binding and adsorption capability for trivalent and divalent Co, as well as their improved adsorption on organic materials, are associated with this process [23].
Additionally, the kind and quantity of other ligands/ions have a role in the association of Co in soil. Where the Fe level was high, the Co content in the B-soil horizons remained higher. Co binding with Mn predominates over other variables governing Co geochemistry in soils with high Mn levels. It is shown that whereas Fe/Mn oxides may collect Co from 70 to 390 mg/kg, Fe nodules can accumulate Co up to 300 mg/kg. High Mn soil may deposit > 20,000 mg/kg Co [24].
The effect of soil organic matter (OM) varies and is influenced by the pH of the soil. Although Co concentrations in soil with high OM content are typically lower, Chernozems had greater Co concentrations of up to 50 mg/kg [25]. Co sorption on Fe-coated Si compounds is stimulated by a number of organic ligands, although increased Co mobility was seen in the absence of Fe hydroxides [26]. The kind of soil OM has a significant impact on the Co mobility. Co organic chelates are thought to be quite mobile.
3. Metal analysis of literature data: Data collection and development of database
To trace out the possible trends of Co transfers in the soil-plant system and associated effects on plant physiology/biochemistry, a database was developed considering (i) Co content in soil (ii) soil plant transfer, the (iii) effect of Co on plant micronutrients and macronutrients (iv) Co compartmentation in plant tissues (root, shoot) (v) effect of Co on plant growth parameters and (vi) enzymes activation under toxic Co levels. So as to accomplish this, literature information regarding the aforementioned Co features was gathered from peer-reviewed papers. In the soil-plant system, 1026 response variables relating to Co behavior were gathered (Table 2).
Using these response variables, an overall effect of Co on plants was calculated using “the percentage change in response variable”. This “percentage change in response variables” was calculated using the following equation:
These 1026 observations found 567 increases, 381 decreases, and 1 with no impact on various plant variables/parameters (Table 2). The soil-plant system's CO behavior is not straightforward, according to meta-analysis. Depending on a number of variables, including the physicochemical characteristics of the soil, the amount of Co that has been applied, and the kind of plant species, it can both decrease and enhance certain variables. The entire trend in the soil-plant system was better understood because of this data analysis.
3.1. Molecular understanding of Co uptake by plants
Sessile plants can absorb elements either actively or passively from soils. Plants typically have Co concentrations of 0.1 to 10 mg/kg dry weight [14]. Plants absorb cobalt from soil in a variety of ways, mostly as di- and trivalent cations. Co uptake by plant roots has an active transport mechanism in higher plants. The enrichment of Co in the soil raises the concentration of cobalt in plants, according to multiple prior studies. This represents the functions of Co concentration in solution and plant absorption of its mobile component. Plant absorption of Co is influenced by its complexation with organic and inorganic ligands and chemical speciation. According to Bhattacharyya, et al. [27], co-occurring in OM, carbonate, and residual fractions of cow dung manure (CDM) and municipal solid waste compost (MSWC) does not correlate with its levels in grains or rice straw. However, rice is easily able to absorb co-occurring in Mn and Fe oxides of CDM and MSWC. Similar to this, there is a favorable correlation between the concentration of AAAc-EDTA soluble Co compounds and agricultural plants [28].
Cobalt uptake by plants greatly varies with the type of plant species. For example, Co was hardly detected in radish whereas a considerable amount was noticed in the shoot of the green beans. Leafy vegetables like Spinacia oleracea, Lactuca sativa, and Brassica oleracea have relatively high cobalt concentrations (>0.6 ppm), whereas fodder crops have a much wider range (0.6 to 3.5 ppm).
After being absorbed by plant roots, cobalt is either transferred to plant shoots or sequestered by the vacuole of root cells. Although poor mobility of Co inside plants limits its transfer toward shoots, roots absorb cobalt through an active transport process, and its distribution in plants may include different chemical molecules [29,30,31].
Mobility of Co within the plant leaf tissue is low and mostly it accumulates in plant roots, but free Co ions can be transported to aerial parts. Page and Feller [32] studied 57Co transportation in wheat plants and found that 80% of the 57Co was retained in the roots after four days of culture 50% of Co remained in the roots after 50 days and the remaining was translocated towards shoot. The data analysis of the current study revealed an overall 49% accumulation in plant roots and 51% in aerial parts based on 33 observations of different studies (Table 3, Table S1). However, the % accumulation ranges 14-94% in roots and 6-86% in aerial parts. This proposes that there exists great variation in Co partitioning between root and shoot tissues. Moreover, data analysis revealed that plants have an effective mechanism to uptake and transfer from root to shoot.
This relative distribution of cobalt in plant roots and aerial tissues greatly varies with the type of plant species. For example, in wheat crops, Co was dispersed in the leaves exactly in the same way as Ca and K through a parallel veins system [29]. Whereas in tomato leaves Co was not dispersed in a similar way as in wheat crop. The authors proposed that the variation in plant sequestration and defense system for Co can be a possible reason for varied distribution among different plant species. It has also been found that Co ions in higher plants bind with the roots and translocate within the body through passive transport. Cobalt ions move through the plasma membrane enter the cell and might be translocated within the whole plant with the help of IRT1 transporters [33]. IRT-1 can also transport Co inside cortical cells and vacuoles of the root epidermal thereby sequestrating Co in most outer layers of the roots.
There haven't been any reports of particular Co transporters to yet. Mobile Co can be translocated toward the shoot by FPN1 after being loaded into the xylem [34]. Cobalt is chelated with ligands before being transferred to shoots. Since phytochelatins have a low affinity for it, Co-S bonds are not expected to be the ligands. According to the Collins and Kinsela [35], in wheat or tomato plants, Co was complexed with carboxylic acids and moved from the roots to the shoots. Other ligands include non-proteinogenic amino acids like nicotinamide and histidine as well as citrate or malate.
Distribution, transportation, and uptake of Co are mainly dependent on species and governed by several mechanisms (Figure 2). Intracellular affinity, binding sites, and transport proteins mediate the Co ion uptake through the plasma membrane. Co transport within cell membranes is facilitated by a number of distinct transmembrane proteins. The 1B-ATPase (HMA) family has been described among the different known heavy metal transporters, and this family is involved in the transportation of both divalent (Co2+/Cd2+/Zn2+/Pb2+) and monovalent (Ag+/Cu+) heavy metals. [36]. Similarly, HMA2 and HMA3 belong to the P1B2 sub-group and transport the divalent metals such as Co2+ and Cd2+ based upon the phylogenetic analysis, HMA5 fits in the P1B1 subclass specific for monovalent ions (Cu+) [37]. HMA2 is limited in the pericycle and contributes to the movement of metals from root to shoot by loading divalent heavy metals, such as Co, into the xylem [38], HMA3 and HMA5, however, trap monovalent Co into vacuoles, which helps with metal tolerance [39].
The AtHMA3 which belongs to the P-type ATPase family is located in the plant vacuolar membrane and is involved in heavy metal transportation to the vacuole [39] hence enhancing plant tolerance. Previous studies have revealed CPX involvement in Co ion coordination during its transport [40]. Also unexplored is the metal specificity of these ATPase proteins. Studies on Synechocystis sp. have shown that the Co2+ tolerance was decreased and intracellular Co2+ levels were elevated in the presence of the ATPase-encoding gene deletion.
3.2. Beneficial effects of Co on plants
Uncertainty surrounds the direct involvement of cobalt in plant metabolism. Insufficient evidence exists for both Co's direct impact on plants and its bioactive form [1]. Moreover, the mechanisms of Co-mediated beneficial effects on plants are not well-elucidated. Some studies have reported a positive effect of Co on plants at low applied levels [11]. Several enzyme and coenzyme processes require cobalt. It is now widely accepted that it assists legumes fix nitrogen fast [3]. Other advantages include the development of the stem, the lengthening of the coleoptile, the creation of buds, the promotion of plant growth, and the yield when exogenously carried out [3,11,33]. According to several research, Co is essential for plants to produce ethylene. The free-living Azotobacter bacteria, which belong to the genus Rhizobium, use cobalt as one of their coenzymes to bind nitrogen from the air [41] because redox reactions and the production of nucleoproteins both depend on the production of vitamin B12. Through its interactions with Fe, Ni, and Zn, cobalt plays a significant role in preserving plant cellular homeostasis.
Cobalt is a useful metal for leguminous plants in terms of metabolism, growth, and the formation of root nodules [33]. Similarly, it improves the development of salinized plants by increasing leaf water potential in comparison to untreated plants. Plant height, branch number, fruit number, and anthocyanin and flavonoid content of Hibiscus sabdariffa rose considerably following application of Co at 20-40 mg/kg [42]. When increasing levels of Co (50, 100, 150, 200, and 250 mg/kg) were given to maize plants, the root length, shoot height, and number of cobs and seeds per plant increased, but these parameters decreased as the concentration reached 100 mg Co/kg and above [43]. Cobalt at 10 mg/kg significantly increased the growth, bulb yields, bulb length, and bulb quality of two onion cultivars, including nutritional and essential oil levels. Bulb diameter and weight were considerably larger in the experimental treatment than in the control treatment [44], However, Co doses greater than 10 mg/kg greatly decreased the pro-inflammatory effects.
Explanations for enhanced plant growth vary, however the following can be summarized: (1) enhanced abiotic stress tolerance, (2) activation of antioxidative enzymes, (3) active metal substitution, and (4) hormesis. Furthermore, Co has been shown to alleviate the effects of drought, salt, and heavy metal stressors, ensuring that plant development is not inhibited by the activation of various enzymes and coenzymes in the presence of Co [11,15].
The data analysis of the literature showed an overall 11% and 2% increase, respectively in plant pigment contents (based on 151 observations) and growth parameters (214 observations) (Table 4, Tables S2–S4). This reveals that application of Co to plants can enhance plant pigment contents and the growth parameters.
3.3. Cobalt deficiency in plants
Plants respond to Co concentrations in soil in the same way as they do to other critical micronutrients: at low concentrations, it supports plant development, while at greater concentrations, it causes phytotoxicity [3]. However, it differs from other helpful elements in that plants demonstrate Co shortcoming when soil Co availability is severely limited. Leaf chlorosis and necrosis, growth retardation, and reduced seed germination and crop yield are all symptoms of cobalt deficiency, which is comparable to N deficiency in plants [45,46] (Table 5). Cobalt-deficient legumes have smaller plant size, smaller and pale-yellow leaves, and smaller pods when compared to non-plants [3]. Furthermore, Co-deficient plants may exhibit nitrogen deficit symptoms. Root development is also hampered by a general decrease in root volume and length. Cobalt shortage reduces methionine synthesis, reducing protein synthesis and contributing to smaller-sized bacteroid. Co-deficient foods include sweet lupin, ground almonds, navy beans, and soy.
3.4. Toxic effects of excess cobalt on the plants
Heavy metals including Co have been recognized for their toxic effects on plant biochemical attributes. Cobalt is not considered an essential metal for plants but rather designated as a beneficial element [47]. Plant toxicity to cobalt is rare in natural soils, but occurs when plants grow in Co-contaminated soils. Higher concentrations of cobalt in plant tissues can cause impairment to cell membranes, which results in biomass reduction, chlorosis, reduction in growth, and uptake of nutrients and water, and hence induce cellular toxicity [6,48,49,50,51]. Excess Co exposure causes chlorosis (pale-white coloring) on young leaves of leguminous plants and interveinal chlorosis (scattered chlorosis) on young leaves of tomatoes [3]. However, the main plant reaction to greater Co concentrations is interveinal chlorosis, which has been connected to Fe chlorosis. Phytotoxicity study of Co concentration in Hordeum Vulgare L., Lycopersicon esculentum L., and Brassica napus L. has revealed the adverse effect on plant shoot and biomass growth [51]. According to Sree, et al. [52], Co may reduce the activity of enzymes involved in the production of chlorophyll intermediates such 5-aminolevulinic acid and protoporphyrin, resulting in a decrease in net photosynthetic activities.
Plants with higher Co concentrations had lower Fe concentrations, protein, chlorophyll, and CAT activity in their leaves. Jayakumar et al. [5] showed the increased chlorophyll concentration at lower Co levels. Higher Co levels limit the Fe incorporation in protoporphyrin which causes a decrease in chlorophyll content. This was reinforced by the fact that high concentrations of other heavy metals, including Co [53], caused chlorosis in plants, which was comparable to the chlorosis caused by Fe shortage. Excess Co can limit transpiration and water potential rate substantially. Cauliflower leaves exhibited diffusive resistance and greater water content when exposed to high Co concentrations [6].
Higher concentration of Co also affects root growth by hindering the cell division and retarding the nutrient and water uptake. Reduced germination was also observed in wheat seeds due to the higher Co exposure [54]. Similar findings of reduction in germination due to Co exposure had been stated in several crops such as Vigna mungo [55], and ragi [56]. All of these elements, when combined, can cause phytotoxicity and drastically limit plant development. However, phytotoxicity varies according to plant species and Co concentration in plant organs.
The data analysis revealed both increase and decrease in plant pigment and growth parameters under Co exposure. Out of 151 observations, 92 showed a decrease while 54 showed an increase in plant pigments (Table 4). In the case of plant growth parameters, 106 observations demonstrated a decrease and 103 represented an increase. The intensity of increase and decrease was, respectively 94% and 1587% for pigments and 169% and 214% for plant growth factors. Hence, Co exposure can mediate both positive and negative effects on plant growth. Therefore, it is highly necessary to monitor Co soil levels, possible soil-plant transfer index, accumulation in different plant tissues, and toxicity.
3.5. Cobalt effects on plant micronutrients
Except for Fe, increasing Co levels in faba bean from 5 to 20 ppm considerably boosted the concentration of micronutrients. Such Co treatments, on the other hand, led in a steady drop in plant Fe content in the roots and shoots [57]. The steady reduce in plant Fe content was driven by increasing Co concentration in wheat grains [58]. whenever cobalt was applied to wheat plants, the concentrations of Cu, Mn, and Zn in grains increased significantly as compared to untreated plants [59]. These findings are consistent with those of Aziz, et al. [60], who discovered that a modest quantity of Co had a strong promoter impact on the Cu, Zn, and Mn concentrations of coriander plants.
3.6. Cobalt effects on plant macronutrients
Excessive soil Co level might impact plant absorption of other necessary elements, in addition to decreasing plant growth and development [15,61]. Soil pollution with Co-produced a reduction in the quantity of K, P, Na, Mg, and mainly Ca in the aboveground sections of oats in a research by Wyszkowski, et al. [62], Soil pollution with Co reduced the level of K, P, Na, Mg, and, in particular, Ca in the aboveground sections of oats. Low Co dosages (10-20 mg Co/kg soil) showed a minor influence on macronutrient composition in spring barley in another experiment [63]. High dosages (320 mg Co/kg soil) enhanced the concentration of all macronutrients, particularly Ca, Na, and N, in the plant's aboveground portions [64] discovered a significant rise in P content in plants when exposed to Co.. Chaudhari, et al. [65] discovered that increasing Co levels beyond 10.0 mg kg-1 resulted in a decrease in the promoter impact on macronutrients (N, P, and K), except when Co was applied in the oxide form, which marginally increased the N, P, and K content. These findings are consistent with those of Gad and Kandil [66], who discovered that Co content in wheat plants had a favorable influence on the status of all minerals. Basu, et al. [67] discovered that co-treatment increased nutrient absorption by groundnut seeds when compared to the control.
When compared to the control, 12 mg kg-1 of Co had a substantial primitive influence on the plant macronutrient (N, P, and K) content of soybean with all sources of N2 fertilizers [57]. Increasing Co content in plant medium by more than 12 mg kg-1, on the other hand, resulted in a considerable drop in these nutritive element quantities. This drop appeared to be connected to the Co concentration. Co also has an adverse effect on the movement of P, S, Cu, Mn, and Zn from roots to shoots [6].
We calculated the effect of Co on the uptake of other elements. Data analysis of 148 observations revealed an overall 46 increase in element contents in the presence of Co (Table 6, Table S5-S6). The effect varied from -48% to 2316% with 43 and 105 observations showing, respectively decrease and increase in element contents. Hence, Co co-presence can affect the uptake of different elements by plants, both positively and negatively. Generally, the effect greatly varied for different elements, plants, and soil types.
Moreover, the presence of other elements also mediates an overall increase in Co plant uptake by 53% (range -32% to 612%) (Table 6). Out of 119 observations, 18 showed a decrease while 101 showed an increase in Co phytouptake.
3.7. Cobalt-induced oxidative stress and lipid peroxidation
Excess cobalt is known to function as a catalyst in the formation of hazardous reactive oxygen species such as superoxide (O2), hydroxyl radical (OH), hydrogen peroxide (H2O2), and alkoxy radical (RO) [8]. The generation of increased levels of ROS in plants is thought to be the most damaging and early impact of heavy metal poisoning. The Fenton reaction produces a hydroxyl radical (OH), which promotes lipid peroxidation and degrades the integrity and fluidity of cell membranes [68]. ROS induce oxidative damage to biological macromolecules such as nucleic acids, lipids, proteins, and so on [69,70]. Excess Co also causes proline buildup and has the potential to disrupt antioxidative defense mechanisms [71]. According to Catalani, et al. [72], Co ions are capable of inducing the formation of ROS in vitro and in vivo. Cobalt catalyzes the formation of OH from H2O2 in a Fenton-type reaction. Cobalt exposure can produce oxidative stress, lower glutathione levels, increase oxidized glutathione levels, promote hexose monophosphate shunt, and cause DNA free-radical damage [73,74]. The mechanisms of action include co-interaction with the mitochondria and ROS generation, which may result in mitochondrial neuropathies. It is believed that mitochondrial dysfunction can be impacted by specific mutations in mitochondrial DNA and aggravated by a variety of environmental variables. The data analysis confirmed that Co-mediated enhanced ROS production (average 59% increase with a range of -35% to 352). Out of 35 observations, 11 showed a decrease and 24 exhibited an increase in ROS contents by Co. Hence, it is anticipated that Co presence in plant tissues mediates increased production of ROS. However, it is not fully explored whether this increase always induced negative effects or not. Indeed, ROS plays several beneficial roles inside plants up to their optimal level. However, these radicles become highly toxic when present at supra-optimal levels. Therefore, a Co-induced increase in ROS contents can cause both beneficial and negative effects, depending on the ROS contents and the defense mechanism of the plant.
It is well-established that enhanced ROS production is associated with peroxidation of lipids. The data analysis of the current study supported the hypothesis by showing an overall 47% increase (range, -88% to 436%) in LPO after Co exposure (Table 7, Table S7-S8). Data analysis showed that Co exposure caused an increase in LOP by 82 observations and a decrease by 72 observations. Hence, Co-mediated overproduction of ROS may cause LPO in plants.
3.8. Cobalt genotoxicity
Cobalt is implicated in the creation of ROS in plants, which can interact with plant DNA directly or indirectly to cause strand breaks. According to Faisal, et al. [75], CoO-NPs may have indirectly caused DNA damage in eggplant, as demonstrated by an increase in ROS, mitochondrial membrane damage, and an enhanced esterase level. So far, in vitro studies on Co nanoparticle toxicity has revealed DNA damage [76], oxidative stress [77,78], genotoxic effects [79] and inflammatory responses [80]. Cobalt is a genotoxic metal both in vitro and in vivo, and can potentially cause inhibition of DNA repair and oxidative stress [81,82,83,84]. The genomic instabilities in RAPD (randomly amplified polymorphic DNA) patterns are said to indicate DNA damage caused by genotoxins at Co doses of 5 mM, 10 mM, and 20 mM. SDS-PAGE examination of protein profiles revealed similar alterations [85]. When organisms are exposed to high levels of Co, it directly causes DNA damage, DNA-protein cross-links, and sister chromatid exchange. Furthermore, Co disrupts the DNA repair machinery (Leonard et al., 1998). Co-exposed seeds of various Fabaceae plants revealed individual polymorphism in a research on V. faba plants to evaluate the influence of Co-treatment on polymorphism [86]. However, Co genotoxicity is still a contentious issue.
3.9. Antioxidant enzymes
Plants have an effective tolerance and detoxifying system for dealing with increased levels of ROS caused by biotic and abiotic stress [87,88,89]. Cobalt effect on plant antioxidative enzyme activities is contradictory and both their inhibition and induction have been found in different plant species such as Brassica napus [7], dark tea [4], Glycine max [90], Vigna radiata L. [91], Brassica oleracea [6,49,50,92] and Lycopersicon esculentum L. [50]. In a study, a reduction in the CAT activity which coincided with a significant elevation in the H2O2 content in mustard leaves [93]. The reduced CAT activity under Co stress does not induce the scavenging of H2O2 in mustard leaves. The results confirmed prior findings using co-treated tomato [50], spinach [49], and cabbage [94]. Furthermore, the reduction in enzyme activity may be due to enzyme deactivation caused by direct interaction with metal ions or by higher amounts of damaging ROS [95].
Cobalt, at the right amounts, can activate antioxidative enzymes, minimizing ROS-caused damage. The mitogen-activated protein kinase (MAPK) cascades are crucial pathways in the response of plants to a variety of stressors [96]. Heavy metals such as Hg, Zn, Cd, and Cu have been found to participate in plant signal transduction through MAPK in Medicago sativa [97] and Oryza sativa [98,99]. Despite our expertise and data on Co toxicity and detoxification procedures, there is still a scarcity of information on Co-induced signal transduction pathways. The role of MAPKs (activation of the 44 and 46 kDa MAPKs) in signal transduction in response to excess Co through ROS cannot be ruled out. Several prior studies in various plant species have found evidence of MAPK activation by heavy metals and ROS [97,100]. As a result, the total relevance of the MAPK signaling pathways in the response to heavy metal stress remains unknown. The data analysis of the current study (155 observations) revealed an overall 122% enhanced antioxidant activities with a range of -99% to 3750% (Table 8, Table S9-14). However, 72 observations revealed an increase while 82 observations highlighted a decrease in the activities of antioxidants. Thus, Co generally enhances antioxidant activities, but it may also decrease antioxidant activities. However, the effect depends on the type of enzyme, plant, plant tissue, and the applied Co level.
To check the effect of Co exposure on different types of antioxidant enzymes, a meta-analysis was also performed for each antioxidant. The overall effect was 456%, 64%, 49%, 49%, and -7%, respectively for APX, CAT, GR, POD, and SOD (Table 9, Table S9-S13). This revealed a great variation regarding the effect of Co exposure on the activities of different enzymes. Different enzymes are involved in different reactions to mediate the conversion of different types of ROS. Hence, exposure to CO can differently affect the activities of enzymes.
3.10. Non-enzymatic antioxidants
Co exposure alters the quantity of non-enzymatic antioxidants in addition to antioxidants. Non-enzymatic antioxidants, such as phytochelatins and glutathione, have been shown to scavenge ROS in the presence of heavy metals [101,102]. The role of cysteine as a modifying element in Co stress is particularly intriguing. The activation of the GSH production pathway in response to metal stress is essential for the synthesis of Cys-residual rich peptides, phytochelatins (PC), which naturally bind to and detoxify the detrimental toxic metals. One of the heavy metals that does not activate PC-synthase is cobalt [103], and the activation caused by Co is quite little [104]. Citrate and cysteine concentrations were observed to be greater in the non-accumulators Silene cucubalus and Rauvolfia serpentina, as well as the hyperaccumulator Crotolaria cobalticola, following Co exposure. Free cysteine was found to be involved in Co complexation in plant cells using size exclusion chromatography [105]. A recent research found that Co treatments significantly increased PC, GSH, and GSSG contents in both roots and shoots of the two barley genotypes when compared to controls [106].
Data analysis observed a Co-mediated increase in the activities of non-enzymatic antioxidants (average 100%, range -33% to 1900%) (Table 8, Table S9-S14). Overall, 58 observations showed an increase while 7 observations showed a decrease in the activities of non-enzymatic antioxidants. Hence, like enzymatic antioxidants, the activities of non-enzymatic antioxidants also increased under Co exposure in plants. This increase can ultimately induce some beneficial or tolerance effects inside plants due to the role of different types of antioxidants in various biochemical processes/reactions involved in plant homeostasis/tolerance as well as growth improvement.
3.11. Cobalt in the human body
Co has a wide range of effects on people. Cobalt is essential for the production of vitamin B12 in humans [107]. An adult person typically carries 1 mg of Co, with vitamin B12 accounting for 85% of it. According to Leyssens, et al. [108], 0.1 mol concentration of Co in vitamin B12 forms provides the human body with the appropriate quantity of Co. Cobalt intake by humans varies between 5 and 50 μg day−1 and is mainly in the form of inorganic Co. The maximum Co intake up to 1800 µmol does not cause any negative effect. Another manifestation of the deficiency of Co in humans is disturbed functions of the osseous nervous systems and alimentary (Figure 3) [109]. Although, there is no suggested dietary threshold of Co for humans, but its deficiency can induce pernicious anemia. It is reported that the total body contents of Co in humans are 1.1 and 1.5 mg [110].
Its excess concentration can cause hyperglycemia, gastric disturbance, and polycythemia, and lead to reproductive changes. Cobalt and its compound have higher allergic potential and cause allergic dermatitis. The disease symptoms due to higher levels of Co include heart and lung disease and allergic reactions [111,112,113]. Other symptoms of excess Co exposure include headache, high/low blood pressure, diarrhea, and frequent vomiting. High Co concentration also affects the immunological system [112,114,115]. Cobalt can also mediate significant allergenic problems. interstitial lung disease, asthma, and alveolitis [115].
Intensification of the above diseases is mostly through the plants near industrial sites where human exposure to harmful levels of Co is much greater than at any other site. The frequent use of instruments with high Co admixtures has a considerable impact on the induction of allergic responses, particularly on the hands and skin. Workers and carpenters are particularly vulnerable to harmful metal exposure [116]. Concerns have been raised concerning the long-term effects of exposure to compounds containing tungsten carbide (80-95%) in matrices containing Ni (0-5%) and Co (5-20%) [117,118,119,120,121].
The harmful effects of Co on humans are majorly due to the intake of Co-contaminated water and food. Mainly, pregnant women can accumulate a significant Co quantity in their bodies from food consumption, which ultimately affects the growing fetus. Chan-Hon-Tong, et al. [122] studied pregnant women and showed that women who had higher blood Co levels were eating only fish in comparison to women who were eating milk products, fruits, soups, and sweats.
Cobalt is a proven carcinogen in humans; however, it is only weakly genotoxic. Cobalt produces DNA breaks, changes in DNA basis, and sister chromatid exchanges in human aneuploidy and lymphocytes but not chromosomal abnormalities [123] or a substantial increase in DNA migration when the comet' test was used [124]. In HeLa cells, cobalt causes DNA strand breaks, a smaller proportion of 6-thioguanine-resistant mutants, and a higher frequency of sister chromatid exchanges than in V79 Chinese hamster cells [30]. Cobalt compounds help to produce sticky bridges in binucleate cells and anaphase, as well as fragmentation and chromatin bridges. High Co concentration inhibits RNA synthesis and reduces DNA and RNA quantities, most likely through altering the activities of a large number of exo- and endonucleases [31].
When Co interacts with other mutagens, its genotoxicity may become more dangerous. In the presence of H2O2, cobalt causes alterations in the DNA of chromatin isolated from grown human cells [125]. Cobalt inhibits DNA repair after it has been damaged by genotoxic substances such as X-rays, ultraviolet light, and alkylating compounds [126,127] even at lower cytotoxic content [128].
4. Conclusions
The current review highlighted the biogeochemical behavior of Co in the soil-plant-human continuum. Cobalt and its compounds are beneficial for plants and essential for animals and humans. In the recent era, natural and anthropogenic sources have enhanced the environmental pollution of Co as well as its associated ecological risks. Cobalt is greatly used in several industrial processes, which caused an 116% increase in the mining of this metal worldwide during 2010–2022. Cobalt is a rare metal in soil and mainly exists in divalent form. Cobalt is partially mobile in soil and has low phytoavailability. The mobility and solubility of Co in the soil system and its phytouptake are controlled by various factors, such as plant species and soil properties.
Although Co is not essential for plants, it mediates numerous beneficial effects on them at low applied levels. However, high levels of Co can mediate numerous toxic effects, leading to decreased plant growth. Cobalt is capable of overproducing ROS and inducing oxidative stress in plants. Nevertheless, plants possess tolerance and detoxification mechanisms comprising enzymatic and non-enzymatic antioxidants to scavenge overproduction of ROS. Cobalt also plays a key role in human metabolism. However, a high intake of Co can cause numerous clinical symptoms in humans. Hence, an optimum supply of Co to plants, animals, and humans is necessary for a healthy life.
4.1. Future perspectives
This study finds the following research gaps about Co in the soil-plant-human system:
- Understanding regarding Co phytouptake/accumulation and the mechanism of its translocation inside the plant are still unknown.
- Various physiological pathways in the plant roots that control Co immobilization in plants is still not fully unraveled.
- Cobalt transport inside the plants by specific transporters and its binding with various transporters need to be explored.
- The mechanism behind Co-mediated beneficial and toxic effects in plants needs further research. The deficiency symptoms and underlying processes are still unveiled.
- Cobalt-induced overproduction of reactive radicles needs further exploration.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
References
- Srivastava, P.; Bolan, N.; Casagrande, V.; Benjamin, J.; Adejumo, S.A.; Sabir, M.; Farooqi, Z.-u.-R. ; Saifullah. Chapter 5 - Cobalt in soils: sources, fate, bioavailability, plant uptake, remediation, and management. In Appraisal of Metal(loids) in the Ecosystem, Kumar, V., Sharma, A., Setia, R., Eds.; Elsevier: 2022; pp. 81-104.
- Arango-Guerra, P.; Lopez-Gonzalez, R. Cobalamin (vitamin B12) deficiency: A reversible cause of myelopathy and pancytopenia to never forget. Neurol. Perspect. 2023, 3. [Google Scholar] [CrossRef]
- Kumar, V.; Srivastava, A.K.; Suprasanna, P. Plant Nutrition and Food Security in the Era of Climate Change; Elsevier BV: Amsterdam, NX, Netherlands, 2022; ISBN 9780128229163. [Google Scholar]
- Zhou, H.; Chen, Y.; Wang, Z.; Xie, C.; Ye, D.; Guo, A.; Xie, W.; Xing, J.; Zheng, M. Preparation, characterization and antioxidant activity of cobalt polysaccharides from Qingzhuan Dark Tea. Heliyon 2023, 9, e15503. [Google Scholar] [CrossRef] [PubMed]
- Jayakumar, K.; Rajesh, M.; Ganesh, K.S.; Vijayarengan, P. Cobalt alters the nodulation, leg-haemoglobin content and antioxidant potential of horse gram (Dolichos biflorus). Intl J Mod Biochem 2013, 2, 15–23. [Google Scholar]
- Chatterjee, J.; Chatterjee, C. Phytotoxicity of cobalt, chromium and copper in cauliflower. Environ. Pollut. 2000, 109, 69–74. [Google Scholar] [CrossRef]
- Ali, S.; Gill, R.A.; Ulhassan, Z.; Zhang, N.; Hussain, S.; Zhang, K.; Huang, Q.; Sagir, M.; Tahir, M.B.; Gill, M.B.; et al. Exogenously applied melatonin enhanced the tolerance of Brassica napus against cobalt toxicity by modulating antioxidant defense, osmotic adjustment, and expression of stress response genes. Ecotoxicol. Environ. Saf. 2023, 252, 114624. [Google Scholar] [CrossRef]
- Gao, J.-J.; Peng, R.-H.; Zhu, B.; Tian, Y.-S.; Xu, J.; Wang, B.; Fu, X.-Y.; Han, H.-J.; Wang, L.-J.; Zhang, F.-J.; et al. Enhanced phytoremediation of TNT and cobalt co-contaminated soil by AfSSB transformed plant. Ecotoxicol. Environ. Saf. 2021, 220, 112407. [Google Scholar] [CrossRef]
- Shahid, M. Biogeochemical behavior of heavy metals in soil-plant system. 2017.
- Albanese, S.; Sadeghi, M.; Lima, A.; Cicchella, D.; Dinelli, E.; Valera, P.; Falconi, M.; Demetriades, A.; De Vivo, B. GEMAS: Cobalt, Cr, Cu and Ni distribution in agricultural and grazing land soil of Europe. J. Geochem. Explor. 2015, 154, 81–93. [Google Scholar] [CrossRef]
- Tsamo, C.; Yerima, N.E.; Mua, E.N. Evaluation of the transport and mobility of Co(II) in soils from agricultural, waste dump and an automobile repair shop sites in Bambili-Cameroon. Environ. Chem. Ecotoxicol. 2021, 4, 29–36. [Google Scholar] [CrossRef]
- Kosiorek, M.; Wyszkowski, M. Remediation of Cobalt-Contaminated Soil Using Manure, Clay, Charcoal, Zeolite, Calcium Oxide, Main Crop (Hordeum vulgare L.), and After-Crop (Synapis alba L.). Minerals 2020, 10, 429. [Google Scholar] [CrossRef]
- Rawat, K.S.; Kumar, R.; Singh, S.K. Topographical distribution of cobalt in different agro-climatic zones of Jharkhand state, India. Geol. Ecol. Landscapes 2018, 3, 14–21. [Google Scholar] [CrossRef]
- Bidast, S.; Golchin, A.; Baybordi, A.; Naidu, R. Effects of Fe oxide-based nanoparticles on yield and nutrient content of corn in Cobalt-contaminated soils. Environ. Technol. Innov. 2022, 26, 102314. [Google Scholar] [CrossRef]
- Jiang, M.; Wang, K.; Wang, Y.; Zhao, Q.; Wang, W. Technologies for the cobalt-contaminated soil remediation: A review. Sci. Total. Environ. 2021, 813, 151908. [Google Scholar] [CrossRef] [PubMed]
- Shahid, M. Effect of soil amendments on trace element-mediated oxidative stress in plants: Meta-analysis and mechanistic interpretations. J. Hazard. Mater. 2020, 407, 124881. [Google Scholar] [CrossRef] [PubMed]
- Shahid, M.; Dumat, C.; Khalid, S.; Schreck, E.; Xiong, T.; Niazi, N.K. Foliar heavy metal uptake, toxicity and detoxification in plants: A comparison of foliar and root metal uptake. J. Hazard. Mater. 2017, 325, 36–58. [Google Scholar] [CrossRef] [PubMed]
- Shahid, M.; Pinelli, E.; Dumat, C. Review of Pb availability and toxicity to plants in relation with metal speciation; role of synthetic and natural organic ligands. J. Hazard. Mater. 2012, 219-220, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Fleming, G. Aspects of the soil chemistry of cobalt. The Significance of Trace Elements in Solving Petrogenetic Problems and Controversies". Ed. SS Augustithis 1983, 731-743.
- Kabata-Pendias, A.; Sadurski, W. Trace elements and compounds in soil. Elements and Their Compounds in the Environment: Occurrence, Analysis and Biological Relevance, Second Edition 2004, 79-99.
- Narendrula, R.; Nkongolo, K.K.; Beckett, P. Comparative Soil Metal Analyses in Sudbury (Ontario, Canada) and Lubumbashi (Katanga, DR-Congo). Bull. Environ. Contam. Toxicol. 2011, 88, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Hiller, E.; Jurkovič. ; Šutriepka, M. Metals in the Surface Sediments of Selected Water Reservoirs, Slovakia. Bull. Environ. Contam. Toxicol. 2010, 84, 635–640. [Google Scholar] [CrossRef]
- Dávila-Rangel, J.I.; Solache-Ríos, M. Sorption of cobalt by two Mexican clinoptilolite rich tuffs zeolitic rocks and kaolinite. J. Radioanal. Nucl. Chem. 2006, 270, 465–471. [Google Scholar] [CrossRef]
- Kabata-Pendias, A. Trace elements in soils and plants; CRC press: 2010.
- Protasova, N.; Kopayeva, M. Trace and dispersed elements in soils of Russian Plateau. Pochvovedeniye 1985, 1, 2937. [Google Scholar]
- Brooks, S.C.; Herman, J.S. Rate and extent of cobalt sorption to representative aquifer minerals in the presence of a moderately strong organic ligand. Appl. Geochem. 1998, 13, 77–88. [Google Scholar] [CrossRef]
- Bhattacharyya, P.; Chakrabarti, K.; Chakraborty, A.; Tripathy, S.; Kim, K.; Powell, M. Cobalt and nickel uptake by rice and accumulation in soil amended with municipal solid waste compost. Ecotoxicol. Environ. Saf. 2008, 69, 506–512. [Google Scholar] [CrossRef] [PubMed]
- Sillanpää, M.; Jansson, H. ; Food; Nations, A.O.o.t.U. Status of Cadmium, Lead, Cobalt, and Selenium in Soils and Plants of Thirty Countries; Food and Agriculture Organization of the United Nations: 1992.
- Bakkaus, E.; Gouget, B.; Gallien, J.-P.; Khodja, H.; Carrot, F.; Morel, J.L.; Collins, R. Concentration and distribution of cobalt in higher plants: The use of micro-PIXE spectroscopy. Nucl. Instruments Methods Phys. Res. Sect. B: Beam Interact. Mater. Atoms 2005, 231, 350–356. [Google Scholar] [CrossRef]
- Barysas, D.; Cesniene, T.; Balciuniene, L.; Vaitkuniene, V.; Rancelis, V. Genotoxicity of Co2+ in plants and other organisms. Biologija 2002, 1, 58–63. [Google Scholar]
- Palit, S.; Sharma, A.; Talukder, G. Effects of cobalt on plants. Bot. Rev. 1994, 60, 149–181. [Google Scholar] [CrossRef]
- Page, V.; Feller, U. Selective Transport of Zinc, Manganese, Nickel, Cobalt and Cadmium in the Root System and Transfer to the Leaves in Young Wheat Plants. Ann. Bot. 2005, 96, 425–434. [Google Scholar] [CrossRef]
- Mahey, S.; Kumar, R.; Sharma, M.; Kumar, V.; Bhardwaj, R. A critical review on toxicity of cobalt and its bioremediation strategies. SN Appl. Sci. 2020, 2, 1–12. [Google Scholar] [CrossRef]
- Morrissey, J.; Baxter, I.R.; Lee, J.; Li, L.; Lahner, B.; Grotz, N.; Kaplan, J.; Salt, D.E.; Guerinot, M.L. The Ferroportin Metal Efflux Proteins Function in Iron and Cobalt Homeostasis in Arabidopsis. Plant Cell 2009, 21, 3326–3338. [Google Scholar] [CrossRef]
- Collins, R.N.; Kinsela, A.S. The aqueous phase speciation and chemistry of cobalt in terrestrial environments. Chemosphere 2010, 79, 763–771. [Google Scholar] [CrossRef]
- Migocka, M.; Kosieradzka, A.; Papierniak, A.; Maciaszczyk-Dziubinska, E.; Posyniak, E.; Garbiec, A.; Filleur, S. Two metal-tolerance proteins, MTP1 and MTP4, are involved in Zn homeostasis and Cd sequestration in cucumber cells. J. Exp. Bot. 2014, 66, 1001–1015. [Google Scholar] [CrossRef]
- Argüello, J.M. Identification of Ion-Selectivity Determinants in Heavy-Metal Transport P 1B -type ATPases. J. Membr. Biol. 2003, 195, 93–108. [Google Scholar] [CrossRef]
- Wong, C.K.E.; Cobbett, C.S. HMA P-type ATPases are the major mechanism for root-to-shoot Cd translocation in Arabidopsis thaliana. New Phytol. 2008, 181, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Morel, M.; Crouzet, J.; Gravot, A.; Auroy, P.; Leonhardt, N.; Vavasseur, A.; Richaud, P. AtHMA3, a P1B-ATPase Allowing Cd/Zn/Co/Pb Vacuolar Storage in Arabidopsis. Plant Physiol. 2008, 149, 894–904. [Google Scholar] [CrossRef] [PubMed]
- Argüello, J.M.; Eren, E.; González-Guerrero, M. The structure and function of heavy metal transport P1B-ATPases. BioMetals 2007, 20, 233–248. [Google Scholar] [CrossRef]
- Rancelis, V.; Cesniene, T.; Kleizaite, V.; Zvingila, D.; Balciuniene, L. Influence of cobalt uptake by Vicia faba seeds on chlorophyll morphosis induction, SOD polymorphism, and DNA methylation. Environ. Toxicol. 2011, 27, 32–41. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, A.K.; Kesavachandran, C.N.; Kumar, S. Evaluating risks of acquired clinical vulnerability among subjects exposed to E-waste. Rev Environ Contam Toxicol 2011, 214, 1–14. [Google Scholar] [CrossRef]
- Jaleel, C.A.; Changxing, Z.; Jayakumar, K.; Iqbal, M. Low Concentration of Cobalt Increases Growth, Biochemical Constituents, Mineral Status and Yield in Zea Mays. J. Sci. Res. 2008, 1, 128–137. [Google Scholar] [CrossRef]
- Abdel-Salam, A.I.; Attia, S.Y.; Mohamed, S.G.; El-Hosiny, F.I.; Sadek, M.; Rashad, M. Designing a hierarchical structure of nickel-cobalt-sulfide decorated on electrospun N-doped carbon nanofiber as an efficient electrode material for hybrid supercapacitors. Int. J. Hydrogen Energy 2023, 48, 5463–5477. [Google Scholar] [CrossRef]
- Hu, X.; Wei, X.; Ling, J.; Chen, J. Cobalt: An Essential Micronutrient for Plant Growth? Front. Plant Sci. 2021, 12. [Google Scholar] [CrossRef]
- Banerjee, A.; Roychoudhury, A. Chapter 18 - Beneficial aspects of cobalt uptake in plants exposed to abiotic stresses. In Frontiers in Plant-Soil Interaction, Aftab, T., Hakeem, K.R., Eds.; Academic Press: 2021; pp. 523-529.
- Marschner, H. Marschner's mineral nutrition of higher plants; Academic press: 2011.
- Chatterjee, J.; Chatterjee, C. AMELIORATION OF PHYTOTOXICITY OF COBALT BY HIGH PHOSPHORUS AND ITS WITHDRAWAL IN TOMATO. J. Plant Nutr. 2002, 25, 2731–2743. [Google Scholar] [CrossRef]
- Pandey, N.; Pathak, G.C.; Pandey, D.K.; Pandey, R. Heavy metals, Co, Ni, Cu, Zn and Cd, produce oxidative damage and evoke differential antioxidant responses in spinach. Braz. J. Plant Physiol. 2009, 21, 103–111. [Google Scholar] [CrossRef]
- Gopal, R.; Dube, B.K.; Sinha, P.; Chatterjee, C. Cobalt Toxicity Effects on Growth and Metabolism of Tomato. Commun. Soil Sci. Plant Anal. 2003, 34, 619–628. [Google Scholar] [CrossRef]
- Li, H.-F.; Gray, C.; Mico, C.; Zhao, F.-J.; McGrath, S.P. Phytotoxicity and bioavailability of cobalt to plants in a range of soils. Chemosphere 2009, 75, 979–986. [Google Scholar] [CrossRef] [PubMed]
- Sree, K.S.; Keresztes. ; Mueller-Roeber, B.; Brandt, R.; Eberius, M.; Fischer, W.; Appenroth, K.-J. Phytotoxicity of cobalt ions on the duckweed Lemna minor – Morphology, ion uptake, and starch accumulation. Chemosphere 2015, 131, 149–156. [Google Scholar] [CrossRef] [PubMed]
- El-Sheekh, M.; El-Naggar, A.; Osman, M.; El-Mazaly, E. Effect of cobalt on growth, pigments and the photosynthetic electron transport in Monoraphidium minutum and Nitzchia perminuta. Braz. J. Plant Physiol. 2003, 15, 159–166. [Google Scholar] [CrossRef]
- López–Luna, J.; Camacho–Martínez, M.; Solís–Domínguez, F.; González–Chávez, M.; Carrillo–González, R.; Martinez–Vargas, S.; Mijangos–Ricardez, O.; Cuevas–Díaz, M. Toxicity assessment of cobalt ferrite nanoparticles on wheat plants. J. Toxicol. Environ. Heal. Part A 2018, 81, 604–619. [Google Scholar] [CrossRef] [PubMed]
- Munzuroglu, O.; Geckil, H. Effects of Metals on Seed Germination, Root Elongation, and Coleoptile and Hypocotyl Growth in Triticum aestivum and Cucumis sativus. Arch. Environ. Contam. Toxicol. 2001, 43, 203–213. [Google Scholar] [CrossRef]
- Jayakumar, K.; Vijayarengan, P.; Changxing, Z.; Gomathinayagam, M.; Jaleel, C.A. Soil applied cobalt alters the nodulation, leg-haemoglobin content and antioxidant status of Glycine max (L.) Merr. Colloids Surfaces B: Biointerfaces 2008, 67, 272–275. [Google Scholar] [CrossRef]
- Kandil, H.; Farid, I.; El-Maghraby, A. Effect of Cobalt Level and Nitrogen Source on Quantity and Quality of Soybean Plant. J. Basic. Appl. Sci. Res. 2013, 3, 185–192. [Google Scholar]
- Kandil, H.; El-Maghraby, A. Impact of cobalt form and level addition on wheat plants (Triticum aestivum L. ): I. growth parameters and nutrients status. 2016, 9, 111–118. [Google Scholar]
- Basu, A.; Mustafiz, S.; Islam, M.R.; Bjorndalen, N.; Rahaman, M.S.; Chaalal, O. A COMPREHENSIVE APPROACH FOR MODELING SORPTION OF LEAD AND COBALT IONS THROUGH FISH SCALES AS AN ADSORBENT. Chem. Eng. Commun. 2006, 193, 580–605. [Google Scholar] [CrossRef]
- Aziz, E.; Gad, N.; Badran, N. Effect of cobalt and nickel on plant growth, yield and flavonoids content of Hibiscus sabdariffa L. Aust. J. Basic Appl. Sci. 2007, 1, 73–78. [Google Scholar]
- Elshamly, A.M.S.; Nassar, S.M.A. The Impacts of Applying Cobalt and Chitosan with Various Water Irrigation Schemes at Different Growth Stages of Corn on Macronutrient Uptake, Yield, and Water Use Efficiency. J. Soil Sci. Plant Nutr. 2023, 23, 2770–2785. [Google Scholar] [CrossRef]
- Wyszkowski, M.; Wyszkowska, J.; Radziemska, M. Macroelement content in oats (Avena Sativa L. ) cultivated on soils contaminated with copper, zinc, tin, cobalt and manganese. Ecological Chemisty Engennering A 2009, 16, 1387–1394. [Google Scholar]
- Kosiorek, M.; Wyszkowski, M. Content of macronutrients in oat (Avena sativa L.) after remediation of soil polluted with cobalt. Environ. Monit. Assess. 2019, 191, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, J.; Chatterjee, C. Management of phytotoxicity of cobalt in tomato by chemical measures. Plant Sci. 2003, 164, 793–801. [Google Scholar] [CrossRef]
- Chaudhari, S.R. Effect of Co Level and FYM on Growth and Yield of Fodder Sorghum. Int. J. Pure Appl. Biosci. 2017, 5, 618–622. [Google Scholar] [CrossRef]
- Gad, N.; Kandil, H. Maximizing the tolerance of wheat plants to soil salinity using cobalt I-growth and mineral composition. Journal of Applied Sciences Research 2011, 7, 1569–1574. [Google Scholar]
- Basu, M.; Mondal, P.; Datta, A.; Basu, T. Effect of Cobalt, Rhizobium and Phosphobacterium Inoculations on Growth Attributes of Summer Groundnut(Arachis hypogaea Linn). Environment and Ecology 2003, 21, 813–816. [Google Scholar]
- Shahid, M.; Pourrut, B.; Dumat, C.; Nadeem, M.; Aslam, M.; Pinelli, E. Heavy-Metal-Induced Reactive Oxygen Species: Phytotoxicity and Physicochemical Changes in Plants. Rev. Environ. Contam. Toxicol. 2014, 232, 1–44. [Google Scholar]
- Natasha, N.; Shahid, M.; Bibi, I.; Iqbal, J.; Khalid, S.; Murtaza, B.; Bakhat, H.F.; Farooq, A.B.U.; Amjad, M.; Hammad, H.M.; et al. Zinc in soil-plant-human system: A data-analysis review. Sci. Total. Environ. 2021, 808, 152024. [Google Scholar] [CrossRef]
- Menhas, S.; Yang, X.; Hayat, K.; Bundschuh, J.; Chen, X.; Hui, N.; Zhang, D.; Chu, S.; Zhou, Y.; Ali, E.F.; et al. Pleiotropic melatonin-mediated responses on growth and cadmium phytoextraction of Brassica napus: A bioecological trial for enhancing phytoremediation of soil cadmium. J. Hazard. Mater. 2023, 457, 131862. [Google Scholar] [CrossRef] [PubMed]
- Tourky, S.M.; Shukry, W.M.; Hossain, M.A.; Siddiqui, M.H.; Pessarakli, M.; Elghareeb, E.M. Cobalt enhanced the drought-stress tolerance of rice (Oryza sativa L.) by mitigating the oxidative damage and enhancing yield attributes. South Afr. J. Bot. 2023, 159, 191–207. [Google Scholar] [CrossRef]
- Catalani, S.; Rizzetti, M.; Padovani, A.; Apostoli, P. Neurotoxicity of cobalt. Human & experimental toxicology 2012, 31, 421–437. [Google Scholar]
- Valko, M.; Rhodes, C.J.; Moncol, J.; Izakovic, M.; Mazur, M. Free radicals, metals and antioxidants in oxidative stress-induced cancer. Chem. Biol. Interact. 2006, 160, 1–40. [Google Scholar] [CrossRef]
- Hoet, P.M.; Roesems, G.; Demedts, M.G.; Nemery, B. Activation of the hexose monophosphate shunt in rat type II pneumocytes as an early marker of oxidative stress caused by cobalt particles. Arch. Toxicol. 2002, 76, 1–7. [Google Scholar] [CrossRef]
- Faisal, M.; Saquib, Q.; Alatar, A.A.; Al-Khedhairy, A.A.; Ahmed, M.; Ansari, S.M.; Alwathnani, H.A.; Dwivedi, S.; Musarrat, J.; Praveen, S. Cobalt oxide nanoparticles aggravate DNA damage and cell death in eggplant via mitochondrial swelling and NO signaling pathway. Biol. Res. 2016, 49, 20. [Google Scholar] [CrossRef]
- Ponti, J.; Sabbioni, E.; Munaro, B.; Broggi, F.; Marmorato, P.; Franchini, F.; Colognato, R.; Rossi, F. Genotoxicity and morphological transformation induced by cobalt nanoparticles and cobalt chloride: an in vitro study in Balb/3T3 mouse fibroblasts. Mutagenesis 2009, 24, 439–445. [Google Scholar] [CrossRef]
- Monteiller, C.; Tran, L.; MacNee, W.; Faux, S.; Jones, A.; Miller, B.; Donaldson, K. The pro-inflammatory effects of low-toxicity low-solubility particles, nanoparticles and fine particles, on epithelial cells in vitro: the role of surface area. Occup. Environ. Med. 2007, 64, 609–615. [Google Scholar] [CrossRef]
- Papis, E.; Rossi, F.; Raspanti, M.; Dalle-Donne, I.; Colombo, G.; Milzani, A.; Bernardini, G.; Gornati, R. Engineered cobalt oxide nanoparticles readily enter cells. Toxicol. Lett. 2009, 189, 253–259. [Google Scholar] [CrossRef]
- Colognato, R.; Bonelli, A.; Ponti, J.; Farina, M.; Bergamaschi, E.; Sabbioni, E.; Migliore, L. Comparative genotoxicity of cobalt nanoparticles and ions on human peripheral leukocytes in vitro. Mutagenesis 2008, 23, 377–382. [Google Scholar] [CrossRef]
- Petrarca, C.; Perrone, A.; Verna, N.; Verginelli, F.; Ponti, J.; Sabbioni, E.; Di Giampaolo, L.; Dadorante, V.; Schiavone, C.; Boscolo, P. Cobalt nano-particles modulate cytokine in vitro release by human mononuclear cells mimicking autoimmune disease. International journal of immunopathology and pharmacology 2005, 19, 11–14. [Google Scholar]
- Hartwig, A.; Asmuss, M.; Ehleben, I.; Herzer, U.; Kostelac, D.; Pelzer, A.; Schwerdtle, T.; Bürkle, A. Interference by toxic metal ions with DNA repair processes and cell cycle control: molecular mechanisms. Environ. Health Perspect. 2002, 110, 797–799. [Google Scholar] [CrossRef] [PubMed]
- De Boeck, M.; Kirsch-Volders, M.; Lison, D. Cobalt and antimony: genotoxicity and carcinogenicity. Mutation Research/Fundamental and Molecular Mechanisms of Mutagenesis 2003, 533, 135–152. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Hudson, L.G.; Liu, K.J. Serial review: redox-active metal ions, reactive oxygen species, and apoptosis. Free Radic Biol Med 2004, 37, 582–593. [Google Scholar] [CrossRef]
- Unfried, K.; Albrecht, C.; Klotz, L.-O.; Von Mikecz, A.; Grether-Beck, S.; Schins, R.P. Cellular responses to nanoparticles: Target structures and mechanisms. Nanotoxicology 2007, 1, 52–71. [Google Scholar] [CrossRef]
- Erturk, F.A.; Ay, H.; Nardemir, G.; Agar, G. Molecular determination of genotoxic effects of cobalt and nickel on maize (Zea mays L.) by RAPD and protein analyses. Toxicol. Ind. Heal. 2012, 29, 662–671. [Google Scholar] [CrossRef]
- Rancelis, V.; Cesniene, T.; Zvingila, D.; Barysas, D.; Balciuniene, L.; Dapkuniene, S. Polymorphism of response to cobalt excess in individual Vicia faba plants. Environ. Exp. Bot. 2006, 55, 221–234. [Google Scholar] [CrossRef]
- Shahid, M. ; Natasha; Dumat, C. ; Niazi, N.K.; Xiong, T.T.; Farooq, A.B.U.; Khalid, S. Ecotoxicology of Heavy Metal (loid)-Enriched Particulate Matter: Foliar Accumulation by Plants and Health Impacts. Reviews of Environmental Contamination and Toxicology 2021, 253, 65–113. [Google Scholar]
- Natasha, N.; Shahid, M.; Khalid, S.; Bibi, I.; Naeem, M.A.; Niazi, N.K.; Tack, F.M.G.; Ippolito, J.A.; Rinklebe, J. Influence of biochar on trace element uptake, toxicity and detoxification in plants and associated health risks: A critical review. Crit. Rev. Environ. Sci. Technol. 2021, 52, 2803–2843. [Google Scholar] [CrossRef]
- Antoniadis, V.; Shaheen, S.M.; Levizou, E.; Shahid, M.; Niazi, N.K.; Vithanage, M.; Ok, Y.S.; Bolan, N.; Rinklebe, J. A critical prospective analysis of the potential toxicity of trace element regulation limits in soils worldwide: Are they protective concerning health risk assessment? - A review. Environ. Int. 2019, 127, 819–847. [Google Scholar] [CrossRef]
- Jayakumar, K.; Vijayarengan, P.; Changxing, Z.; Gomathinayagam, M.; Jaleel, C.A. Soil applied cobalt alters the nodulation, leg-haemoglobin content and antioxidant status of Glycine max (L.) Merr. Colloids Surfaces B: Biointerfaces 2008, 67, 272–275. [Google Scholar] [CrossRef] [PubMed]
- Tewari, R.K.; Kumar, P.; Sharma, P.N.; Bisht, S.S. Modulation of oxidative stress responsive enzymes by excess cobalt. Plant Sci. 2002, 162, 381–388. [Google Scholar] [CrossRef]
- Karuppanapandian, T.; Kim, W. Cobalt-induced oxidative stress causes growth inhibition associated with enhanced lipid peroxidation and activates antioxidant responses in Indian mustard (Brassica juncea L.) leaves. Acta Physiol. Plant. 2013, 35, 2429–2443. [Google Scholar] [CrossRef]
- Karuppanapandian, T.; Kim, W. Cobalt-induced oxidative stress causes growth inhibition associated with enhanced lipid peroxidation and activates antioxidant responses in Indian mustard (Brassica juncea L.) leaves. Acta Physiol. Plant. 2013, 35, 2429–2443. [Google Scholar] [CrossRef]
- Pandey, N.; Sharma, C.P. Effect of heavy metals Co 2+, Ni 2+ and Cd 2+ on growth and metabolism of cabbage. Plant Science 2002, 163, 753–758. [Google Scholar] [CrossRef]
- Cavalcanti, F.R.; Lima, J.P.M.S.; Ferreira-Silva, S.L.; Viégas, R.A.; Silveira, J.A.G. Roots and leaves display contrasting oxidative response during salt stress and recovery in cowpea. J. Plant Physiol. 2007, 164, 591–600. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Liu, Y. Activation of salicylic acid–induced protein kinase, a mitogen-activated protein kinase, induces multiple defense responses in tobacco. The Plant Cell 2001, 13, 1877–1889. [Google Scholar]
- Jonak, C.; Nakagami, H.; Hirt, H. Heavy Metal Stress. Activation of Distinct Mitogen-Activated Protein Kinase Pathways by Copper and Cadmium. Plant Physiol. 2004, 136, 3276–3283. [Google Scholar] [CrossRef]
- Yeh, C.-M.; Hung, W.-C.; Huang, H.-J. Copper treatment activates mitogen-activated protein kinase signalling in rice. Physiol. Plant. 2003, 119, 392–399. [Google Scholar] [CrossRef]
- Huang, T.-L.; Huang, H.-J. ROS and CDPK-like kinase-mediated activation of MAP kinase in rice roots exposed to lead. Chemosphere 2008, 71, 1377–1385. [Google Scholar] [CrossRef]
- Kovtun, Y.; Chiu, W.-L.; Tena, G.; Sheen, J. Functional analysis of oxidative stress-activated mitogen-activated protein kinase cascade in plants. Proc. Natl. Acad. Sci. 2000, 97, 2940–2945. [Google Scholar] [CrossRef]
- Shahid, M.; Niazi, N.K.; Rinklebe, J.; Bundschuh, J.; Dumat, C.; Pinelli, E. Trace elements-induced phytohormesis: A critical review and mechanistic interpretation. Crit. Rev. Environ. Sci. Technol. 2019, 50, 1984–2015. [Google Scholar] [CrossRef]
- Shabbir, Z.; Sardar, A.; Shabbir, A.; Abbas, G.; Shamshad, S.; Khalid, S.; Natasha; Murtaza, G. ; Dumat, C.; Shahid, M. Copper uptake, essentiality, toxicity, detoxification and risk assessment in soil-plant environment. Chemosphere 2020, 259, 127436. [Google Scholar] [CrossRef] [PubMed]
- Rauser, W.E. Phytochelatins and Related Peptides (Structure, Biosynthesis, and Function). Plant Physiol. 1995, 109, 1141–1149. [Google Scholar] [CrossRef] [PubMed]
- Vatamaniuk, O.K.; Mari, S.; Lu, Y.-P.; Rea, P.A. Mechanism of heavy metal ion activation of phytochelatin (PC) synthase blocked thiols are sufficient for PC synthase-catalyzed transpeptidation of glutathione and related thiol peptides. Journal of Biological Chemistry 2000, 275, 31451–31459. [Google Scholar] [CrossRef] [PubMed]
- Kleizaite, V.; Cesniene, T.; Rancelis, V. The use of cobalt-induced chlorophyll morphoses for studying Co2+ interactions with cysteine and SOD. Plant Sci. 2004, 167, 1249–1256. [Google Scholar] [CrossRef]
- Lwalaba, J.L.W.; Louis, L.T.; Zvobgo, G.; Richmond, M.E.A.; Fu, L.; Naz, S.; Mwamba, M.; Mundende, R.P.M.; Zhang, G. Physiological and molecular mechanisms of cobalt and copper interaction in causing phyto-toxicity to two barley genotypes differing in Co tolerance. Ecotoxicol. Environ. Saf. 2019, 187, 109866. [Google Scholar] [CrossRef]
- Dobrowolski, R.; Otto, M. Determination of nickel and cobalt in reference plant materials by carbon slurry sampling GFAAS technique after their simultaneous preconcentration onto modified activated carbon. J. Food Compos. Anal. 2012, 26, 58–65. [Google Scholar] [CrossRef]
- Leyssens, L.; Vinck, B.; Van Der Straeten, C.; Wuyts, F.; Maes, L. Cobalt toxicity in humans—A review of the potential sources and systemic health effects. Toxicology 2017, 387, 43–56. [Google Scholar] [CrossRef]
- Soylak, M.; Murat, I. Determination of Copper, Cobalt, Lead, and Iron in Table Salt by FAAS After Separation Using Violuric Acid and Multiwalled Carbon Nanotubes. Food Anal. Methods 2011, 5, 1003–1009. [Google Scholar] [CrossRef]
- Gad, S.C. Cobalt. In Reference Module in Biomedical Sciences; Elsevier: 2023.
- Chen, J.; Chen, C.; Wang, N.; Wang, C.; Gong, Z.; Du, J.; Lai, H.; Lin, X.; Wang, W.; Chang, X.; et al. Cobalt nanoparticles induce mitochondrial damage and β-amyloid toxicity via the generation of reactive oxygen species. NeuroToxicology 2023, 95, 155–163. [Google Scholar] [CrossRef] [PubMed]
- wiątkowska, I.; Akinfosile, O.J.; Badhe, R.V.; Barba, M.; Mathew, M.T.; Bijukumar, D. Chapter 7 - Hip implants and systemic cobalt toxicity: a comprehensive review with case studies. In Biomarkers of Hip Implant Function, Świątkowska, I., Ed.; Academic Press: 2023; pp. 205-247.
- Monnot, A.D.; Kovochich, M.; Bandara, S.B.; Wilsey, J.T.; Christian, W.V.; Eichenbaum, G.; Perkins, L.E.; Hasgall, P.; Taneja, M.; Connor, K.; et al. A hazard evaluation of the reproductive/developmental toxicity of cobalt in medical devices. Regul. Toxicol. Pharmacol. 2021, 123, 104932. [Google Scholar] [CrossRef] [PubMed]
- Garcia, M.D.; Hur, M.; Chen, J.J.; Bhatti, M.T. Cobalt toxic optic neuropathy and retinopathy: Case report and review of the literature. Am. J. Ophthalmol. Case Rep. 2020, 17, 100606. [Google Scholar] [CrossRef] [PubMed]
- Domingo, J.L. Cobalt in the environment and its toxicological implications. Rev Environ Contam Toxicol 1989, 108, 105–132. [Google Scholar] [CrossRef]
- Thyssen, J.P. Nickel and cobalt allergy before and after nickel regulation – evaluation of a public health intervention. Contact Dermat. 2011, 65, 1–68. [Google Scholar] [CrossRef]
- Kraus, T.; Schramel, P.; Schaller, K.H.; Zöbelein, P.; Weber, A.; Angerer, J. Exposure assessment in the hard metal manufacturing industry with special regard to tungsten and its compounds. Occup. Environ. Med. 2001, 58, 631–634. [Google Scholar] [CrossRef]
- Lison, D.; De Boeck, M.; Verougstraete, V.; Kirsch-Volders, M. Update on the genotoxicity and carcinogenicity of cobalt compounds. Occup. Environ. Med. 2001, 58, 619–625. [Google Scholar] [CrossRef]
- Scansetti, G.; Maina, G.; Botta, G.C.; Bambace, P.; Spinelli, P. Exposure to cobalt and nickel in the hard-metal production industry. International archives of occupational and environmental health 1998, 71, 60–63. [Google Scholar] [CrossRef]
- Wild, P.; Perdrix, A.; Romazini, S.; Moulin, J.-J.; Pellet, F. Lung cancer mortality in a site producing hard metals. Occup. Environ. Med. 2000, 57, 568–573. [Google Scholar] [CrossRef]
- Leodolter, K.; Refregier, M.; Schmidt, H.; Zidek, T.; Wild, P. A cohort mortalily and nested case-control study of French and Austrian talc workers. (Original Article). Occupational and Environmental Medicine 2002, 59, 98–106. [Google Scholar]
- Chan-Hon-Tong, A.; Charles, M.-A.; Forhan, A.; Heude, B.; Sirot, V. Exposure to food contaminants during pregnancy. Sci. Total. Environ. 2013, 458-460, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Beyersmann, D.; Hartwig, A. The genetic toxicology of cobalt. Toxicology and applied pharmacology 1992, 115, 137–145. [Google Scholar] [CrossRef] [PubMed]
- De Boeck, M.; Lison, D.; Kirsch-Volders, M. Evaluation of the in vitro direct and indirect genotoxic effects of cobalt compounds using the alkaline comet assay. Influence of interdonor and interexperimental variability. Carcinog. 1998, 19, 2021–2029. [Google Scholar] [CrossRef]
- Nackerdien, Z.; Kasprzak, K.S.; Rao, G.; Halliwell, B.; Dizdaroglu, M. Nickel(II)- and cobalt(II)-dependent damage by hydrogen peroxide to the DNA bases in isolated human chromatin. . 1991, 51, 5837–42. [Google Scholar] [PubMed]
- Hartwig, A.; Krüger, I.; Beyersmann, D. Mechanisms in nickel genotoxicity: the significance of interactions with DNA repair. Toxicol. Lett. 1994, 72, 353–358. [Google Scholar] [CrossRef] [PubMed]
- Hartwig, A. Carcinogenicity of metal compounds: possible role of DNA repair inhibition. Toxicol. Lett. 1998, 102-103, 235–239. [Google Scholar] [CrossRef]
- Hartwig, A. Current aspects in metal genotoxicity. BioMetals 1995, 8, 3–11. [Google Scholar] [CrossRef]
- Lago-Vila, M.; Arenas-Lago, D.; Rodríguez-Seijo, A.; Couce, M.L.A.; Vega, F.A. Cobalt, chromium and nickel contents in soils and plants from a serpentinite quarry. Solid Earth 2015, 6, 323–335. [Google Scholar] [CrossRef]
- Medyńska-Juraszek, A.; Bednik, M.; Chohura, P. Assessing the Influence of Compost and Biochar Amendments on the Mobility and Uptake of Heavy Metals by Green Leafy Vegetables. Int. J. Environ. Res. Public Heal. 2020, 17, 7861. [Google Scholar] [CrossRef]
- Evdokimov, A. Heavy metals in the organic soil horizon of the Northern taiga forests resulting from industrial air pollution. 169, 0101; 1. [Google Scholar] [CrossRef]
- Oladeji, S.; Saeed, M.D. Assessment of cobalt levels in wastewater, soil and vegetable samples grown along Kubanni stream channels in Zaria, Kaduna State, Nigeria. Afr. J. Environ. Sci. Technol. 2015, 9, 765–772. [Google Scholar] [CrossRef]
- Rawat, J.; Saxena, J.; Sanwal, P. Biochar: A Sustainable Approach for Improving Plant Growth and Soil Properties. In Biochar—An Imperative Amendment for Soil and the Environment; IntechOpen: London, UK, 2019. [Google Scholar] [CrossRef]
- Kim, H.W.; Seo, S.B.; Kang, S.Y.; Go, E.S.; Oh, S.S.; Lee, Y.; Yang, W.; Lee, S.H. Effect of flue gas recirculation on efficiency of an indirect supercritical CO2 oxy-fuel circulating fluidized bed power plant. Energy 2021, 227, 120487. [Google Scholar] [CrossRef]
- Sibeso, P.M. Assessment of potentially harmful arsenic, cadmium, cobalt and lead in soils and plants in Nomtsoub suburb, Tsumeb town. University of Namibia, 2020.
- Cappuyns, V.; Alian, V.; Vassilieva, E.; Swennen, R. pH Dependent Leaching Behavior of Zn, Cd, Pb, Cu and As from Mining Wastes and Slags: Kinetics and Mineralogical Control. Waste Biomass- Valorization 2013, 5, 355–368. [Google Scholar] [CrossRef]
- Rizo, O.D.; Castillo, F.E.; López, J.O.A.; Merlo, M.H. Assessment of Heavy Metal Pollution in Urban Soils of Havana City, Cuba. Bull. Environ. Contam. Toxicol. 2011, 87, 414–419. [Google Scholar] [CrossRef] [PubMed]
- Adagunodo, T.; Sunmonu, L.; Emetere, M. Heavy metals' data in soils for agricultural activities. Data Brief 2018, 18, 1847–1855. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.; Wang, E.; Sun, O.J. Can no-tillage stimulate carbon sequestration in agricultural soils? A meta-analysis of paired experiments. Agric. Ecosyst. Environ. 2010, 139, 224–231. [Google Scholar] [CrossRef]
- Jiao, X.; Teng, Y.; Zhan, Y.; Wu, J.; Lin, X. Soil Heavy Metal Pollution and Risk Assessment in Shenyang Industrial District, Northeast China. PLOS ONE 2015, 10, e0127736–e0127736. [Google Scholar] [CrossRef]
- Banza, C.L.N.; Nawrot, T.S.; Haufroid, V.; Decrée, S.; De Putter, T.; Smolders, E.; Kabyla, B.I.; Luboya, O.N.; Ilunga, A.N.; Mutombo, A.M.; et al. High human exposure to cobalt and other metals in Katanga, a mining area of the Democratic Republic of Congo. Environ. Res. 2009, 109, 745–752. [Google Scholar] [CrossRef]
- Krishna, A.K.; Govil, P.K. Soil Contamination Due to Heavy Metals from an Industrial Area of Surat, Gujarat, Western India. Environ. Monit. Assess. 2006, 124, 263–275. [Google Scholar] [CrossRef]
- van der Ent, A.; Erskine, P.; Sumail, S. Ecology of nickel hyperaccumulator plants from ultramafic soils in Sabah (Malaysia). Chemoecology 2015, 25, 243–259. [Google Scholar] [CrossRef]
- Ngole-Jeme, V.M.; Fantke, P. Ecological and human health risks associated with abandoned gold mine tailings contaminated soil. PLOS ONE 2017, 12, e0172517. [Google Scholar] [CrossRef]
- Pourret, O.; Lange, B.; Bonhoure, J.; Colinet, G.; Decrée, S.; Mahy, G.; Séleck, M.; Shutcha, M.; Faucon, M.-P. Assessment of soil metal distribution and environmental impact of mining in Katanga (Democratic Republic of Congo). Appl. Geochem. 2016, 64, 43–55. [Google Scholar] [CrossRef]
- Demková, L.; Árvay, J.; Bobuľská, L.; Hauptvogl, M.; Hrstková, M. Open mining pits and heaps of waste material as the source of undesirable substances: Biomonitoring of air and soil pollution in former mining area (Dubnik, Slovakia). Environmental Science and Pollution Research 2019, 26, 35227–35239. [Google Scholar] [CrossRef]
- Demková, L.; Árvay, J.; Bobuľská, L.; Tomáš, J.; Stanovič, R.; Lošák, T.; Harangozo, L.; Vollmannová, A.; Bystrická, J.; Musilová, J. Accumulation and environmental risk assessment of heavy metals in soil and plants of four different ecosystems in a former polymetallic ores mining and smelting area (Slovakia). Journal of Environmental Science and Health, Part A 2017, 52, 479–490. [Google Scholar] [CrossRef] [PubMed]
- Lotfy, H.; Amer, S.; Zaazaa, H.; Mostafa, N. Spectrophotometric methods for quantitative determination of binary mixture of naproxen sodium and domperidone maleate. Future 2013, 1, 1. [Google Scholar]
- Demková, L.; Jezný, T.; Bobuľská, L. Assessment of soil heavy metal pollution in a former mining area - before and after the end of mining activities. Soil Water Res. 2017, 12, 229–236. [Google Scholar] [CrossRef]
- Kosiorek, M.; Wyszkowski, M. Content of macronutrients in oat (Avena sativa L.) after remediation of soil polluted with cobalt. Environ. Monit. Assess. 2019, 191, 1–15. [Google Scholar] [CrossRef]
- Lwalaba, J.L.W.; Zvobgo, G.; Mwamba, T.M.; Louis, L.T.; Fu, L.; Kirika, B.A.; Tshibangu, A.K.; Adil, M.F.; Sehar, S.; Mukobo, R.P.; et al. High accumulation of phenolics and amino acids confers tolerance to the combined stress of cobalt and copper in barley (Hordeum vulagare). Plant Physiol. Biochem. 2020, 155, 927–937. [Google Scholar] [CrossRef]
- Kandil, H.; Gad, N.; Abdelhamid, M.T. Effects of different rates of phosphorus and molybdenum application on two varieties common bean of (Phaseolus vulgaris L. ). Journal of Agriculture and Food Technology 2013, 3, 8–16. [Google Scholar]
- Jaleel, C.A.; Manivannan, P.; Lakshmanan, G.; Gomathinayagam, M.; Panneerselvam, R. Alterations in morphological parameters and photosynthetic pigment responses of Catharanthus roseus under soil water deficits. Colloids Surfaces B: Biointerfaces 2008, 61, 298–303. [Google Scholar] [CrossRef]
- Jayakumar, K.; Jaleel, C.A.; Azooz, M. Impact of cobalt on germination and seedling growth of Eleusine coracana L. and Oryza sativa L. under hydroponic culture. Global Journal of Molecular Sciences 2008, 3, 18–20. [Google Scholar]
- Imtiyaz, S.; Veqar, Z.; Shareef, M. To Compare the Effect of Vibration Therapy and Massage in Prevention of Delayed Onset Muscle Soreness (DOMS). J. Clin. Diagn. Res. 2014, 8, 133–6. [Google Scholar] [CrossRef] [PubMed]
- Fulekar, M.H.; Jadia, C.D. PHYTOREMEDIATION: THE APPLICATION OF VERMICOMPOST TO REMOVE ZINC, CADMIUM, COPPER, NICKEL AND LEAD BY SUNFLOWER PLANT. Environ. Eng. Manag. J. 2008, 7, 547–558. [Google Scholar] [CrossRef]
- López-Moreno, M.L.; Avilés, L.L.; Pérez, N.G.; Irizarry, B. .; Perales, O.; Cedeno-Mattei, Y.; Román, F. Effect of cobalt ferrite (CoFe2O4) nanoparticles on the growth and development of Lycopersicon lycopersicum (tomato plants). Sci. Total. Environ. 2016, 550, 45–52. [Google Scholar] [CrossRef]
- Gopal, R.; Dube, B.K.; Sinha, P.; Chatterjee, C. Cobalt Toxicity Effects on Growth and Metabolism of Tomato. Commun. Soil Sci. Plant Anal. 2003, 34, 619–628. [Google Scholar] [CrossRef]
Figure 1.
Global annual mine production of cobalt (USGS 2023).
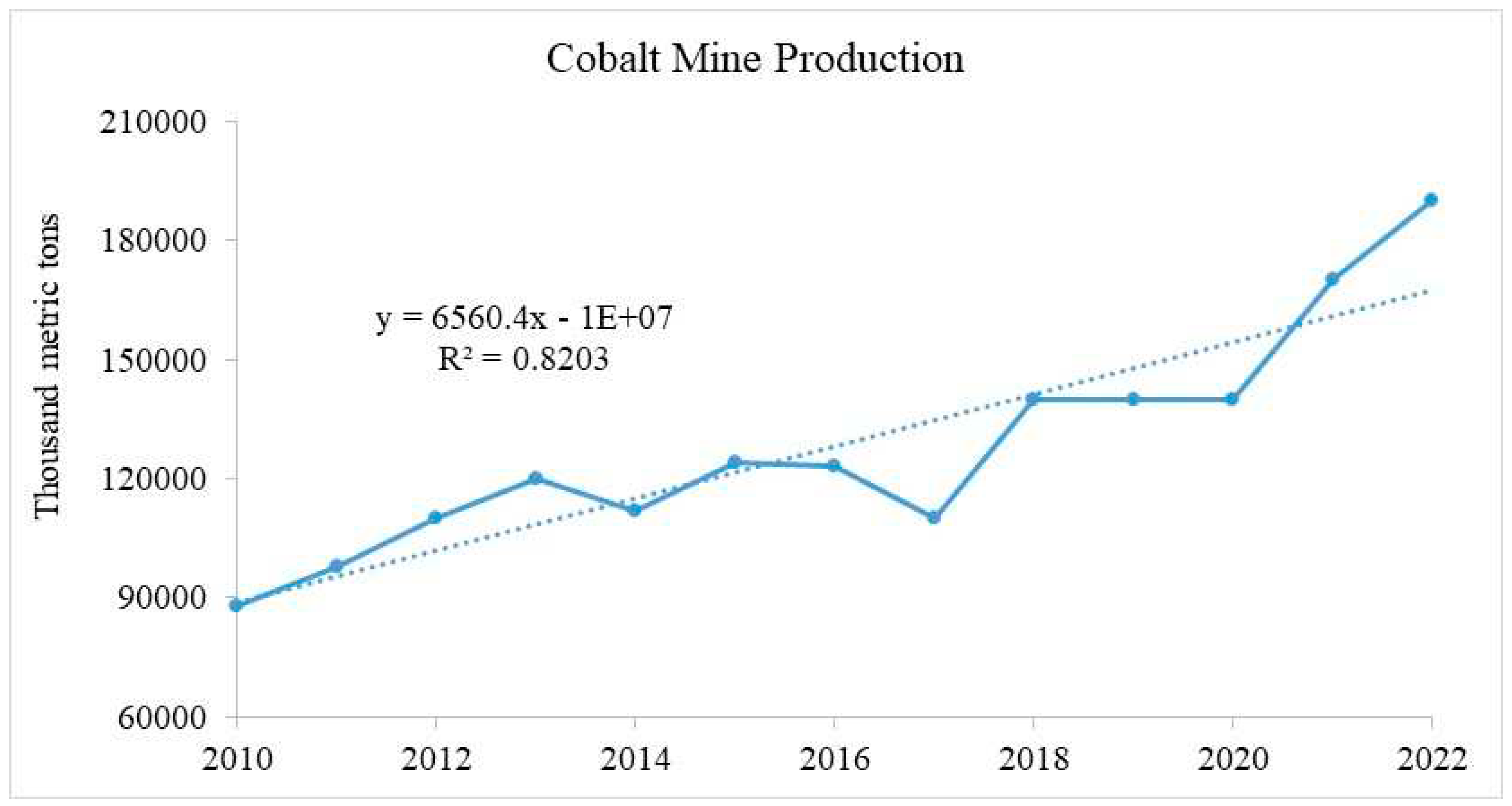
Figure 2.
Overall cobalt functions in soil-plant system.
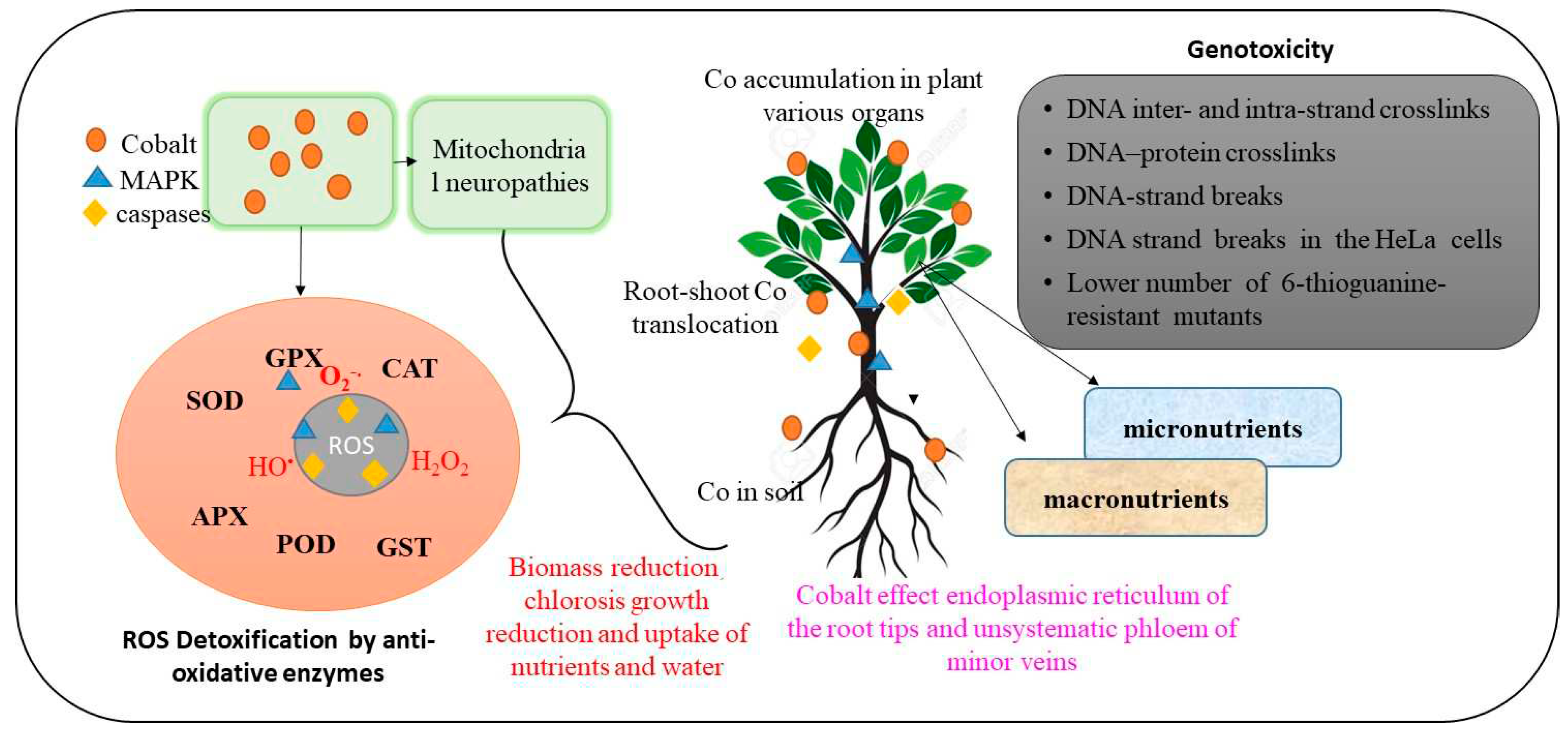
Figure 3.
Cobalt functions in human body.
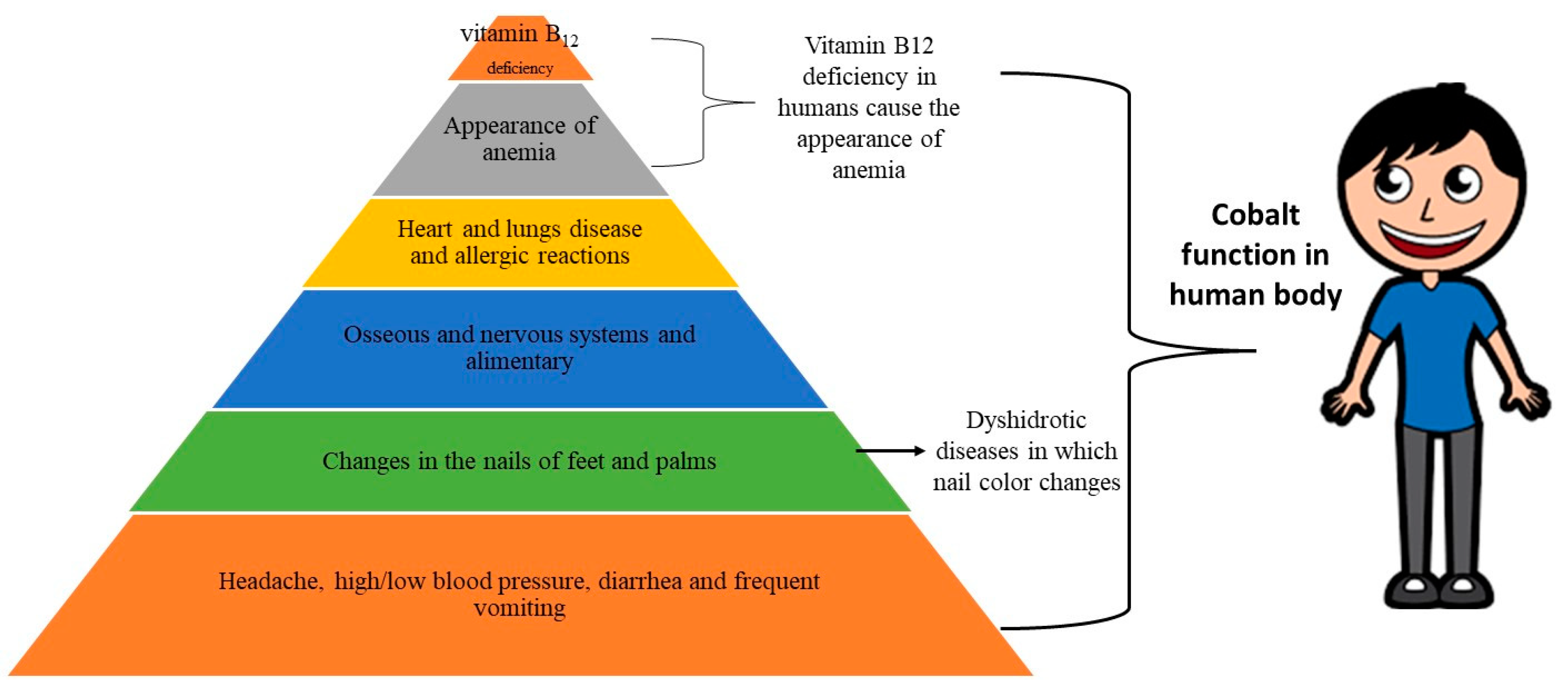

Table 1.
Cobalt contamination of soil in mining, smelting and industrial areas.
| Study area | Co level (mg/kg) | Source | Reference |
|---|---|---|---|
| Coruña, Spain | 147 | Industrial sites | [129] |
| Poland | 74 | Copper-Smelter | [130] |
| Northern taiga | 28.68 | Industrial sites | [131] |
| Nigeria | 3 | Industrial sites | [132] |
| India | 112.72 | - | [133] |
| South Korea | 253 | Power plant | [134] |
| Namibia | 6.3 | Mining area | [135] |
| Belgium | 255 | Manure or sewage sludge | [136] |
| Cuba | 9.2 | Industrial sites | [137] |
| Nigeria | 10 | Agriculture site | [138] |
| china | 21.7 | - | [139] |
| china | 12.91 | Industrial sites | [140] |
| Spain | 110 | Industrial sites | [129] |
| Katanga, Republic of Congo | 15.7 | Mining and refining site | [141] |
| Gujarat, Western India | 51.3 | Industrial area | [142] |
| Katanga, Republic of Congo | 5.72 | Mining and refining site | [141] |
| Northern Zambia | 6.5 | Mining and smelter | [143] |
| Krugersdorp, South Africa | 1679 | Mine tailing | [144] |
| Krugersdorp, South Africa | 1394 | Mine tailing | [144] |
| Katanga, Republic of Congo | 22,467 | Mining and ore processing | [145] |
| North-east part of Slovakia | 31 | Mining and smelting area | [146,147] |
| North greater Cairo, Egypt | 146 | Industrial and sewage effluent | [148] |
| Central Spiš, Europe | 27.2 | Mining and processing of copper | [149] |
Table 2.
Total number of observations collected from different published articles and used for the database development.
Table 2.
Total number of observations collected from different published articles and used for the database development.
| Observations | # |
| Total # of observations | 1026 |
| Negative observations | 381 |
| Positive observations | 567 |
| No effect | 1 |
| NA |
Table 3.
Mean, minimum and maximum Co concentration in shoot and roots of plants (full data available in Table S1).
Table 3.
Mean, minimum and maximum Co concentration in shoot and roots of plants (full data available in Table S1).
| Parameter | Root % | Shoot % |
| Mean | 49 | 51 |
| Minimum | 14 | 6 |
| Maximum | 94 | 86 |
| Total # of observations | 33 | 33 |
Table 4.
Mean and range of % decrease and increase in growth variables of plant roots and shoots (full data available in Table S2-S4).
Table 4.
Mean and range of % decrease and increase in growth variables of plant roots and shoots (full data available in Table S2-S4).
| Parameter | Pigments | Growth | Growth variables for different organs | |
|---|---|---|---|---|
| Shoot | Root | |||
| Mean | 11 | 2 | 3 | 7 |
| Minimum | -94 | -91 | -91 | -80 |
| Maximum | 1587 | 169 | 169 | 140 |
| Total # of observations | 151 | 214 | 131 | 52 |
| Negative observations | 92 | 106 | 52 | 23 |
| Positive observations | 54 | 103 | 79 | 29 |
| No effect | 5 | 5 | 0 | 0 |
Table 5.
Effect of different macro and micronutrients on Co uptake (full data available in Table S5-S6).
Table 5.
Effect of different macro and micronutrients on Co uptake (full data available in Table S5-S6).
| Parameter | Effect on Co uptake | Effect of Co on cations |
|---|---|---|
| Macronutrient | Micronutrients | |
| Mean | 53 | 46 |
| Minimum | -32 | -48 |
| Maximum | 612 | 2316 |
| Total # of observations | 119 | 148 |
| Negative observations | 18 | 43 |
| Positive observations | 101 | 105 |
| No effect | 0 | 0 |
Table 6.
Mean and range of % change in ROS and lipid peroxidation in the response of Co (full data available in Table S7-S8).
Table 6.
Mean and range of % change in ROS and lipid peroxidation in the response of Co (full data available in Table S7-S8).
| Parameter | ROS | LPO |
|---|---|---|
| Mean | 59 | 47 |
| Minimum | -35 | -88 |
| Maximum | 352 | 436 |
| Total # of observations | 35 | 73 |
| Negative observations | 11 | 22 |
| Positive observations | 24 | 50 |
| No effect | 0 | 1 |
Table 7.
Mean and range of % change in enzymes and non-enzymes activities in response to Co (full data available in Table S10-S14).
Table 7.
Mean and range of % change in enzymes and non-enzymes activities in response to Co (full data available in Table S10-S14).
| Parameter | Enzymes | Non-Enzymes |
|---|---|---|
| Mean | 122 | 100 |
| Minimum | -99 | -33 |
| Maximum | 3750 | 1900 |
| Total # of observations | 155 | 65 |
| Negative observations | 82 | 7 |
| Positive observations | 72 | 58 |
| No effect | 0 | 0 |
Table 8.
Mean and range of % change in enzymatic activities of plants in response to Co (full data available in Table S10-S13).
Table 8.
Mean and range of % change in enzymatic activities of plants in response to Co (full data available in Table S10-S13).
| Parameter | APX | CAT | SOD | GR | POD |
|---|---|---|---|---|---|
| Mean | 456 | 64 | -7 | 49 | 49 |
| Minimum | -39 | -99 | -52 | -75 | -59 |
| Maximum | 3750 | 800 | 156 | 300 | 217 |
| Total # of observations | 10 | 58 | 33 | 12 | 42 |
| Negative observations | 3 | 31 | 24 | 5 | 19 |
| Positive observations | 7 | 27 | 8 | 7 | 23 |
| No effect | 0 | 0 | 1 | 0 | 0 |
Table 9.
Effect of Co on plants (deficiency symptoms by various plant species).
| Co level | Organ | Plant species | Deficiency symptoms | Reference |
|---|---|---|---|---|
| 0, 20, 40, 80, 160, 320 mg/kg | Leaves | Avena sativa L. | Higher cobalt levels had significantly decreased the grain yield and straw and also biomass of its roots | [150] |
| 25, 50, 75, and 100 µM | Leaves | Hordeum vulgare | Increase in oxidative stress and reduction in plant pigment contents and the photosynthetic rate | [151] |
| 0, 4, 8, 12, 16 mg/kg | Leaves | Glycine max L. | Significant decrease in concentrations of nutritive elements (N, P, K). | [152] |
| 100, 200 mg/kg | Leaves | Arachis hypogaea | Decreased the various growth parameter (shoot and root length, leaf area, number of nodules, pigment contents starch, sugar, amino acid and protein contents of leaves, antioxidant enzyme such as CAT | [153] |
| 5, 10, 25, 50, 75 and 100 mg /L | Leaves | Eleusine coracana L. | Decrease the seedling growth and plant dry weight at higher cobalt level | [154] |
| 50, 100, 150 µM | Leaves | Glycine Max | Reduction in plant seed germination percentage, seedling growth, plant leaf area, and plant biomass | [155] |
| 5, 10, 20, 40, 50 mg/kg | Leaves | Helianthus annuus | Delay in seed germination, decrease plant shoot and root and shoot | [156] |
| 62.5, 125, 250, 500, 1000 µM | Leaves | Lycopersicon lycopersicum | decrease in plant enzymes activities | [157] |
| 0.05, 0.1, 0.2, 0.4, and 0.5 mM | Leaves | Lycopersicon esculentum | Chlorosis of young leaves, activity of CAT decreased and other enzymes such as peroxidase, acid phosphatase and ribonuclease increased | [158] |
| 0.05, 0.1, 0.2, 0.4, and 0.5 mM | Leaves | Lycopersicon esculentum | Reduction in biomass, reduction in plant chlorophyll content, nucleic acids, nitrogen and carbohydrate fractions, decrease in photosynthesis and CAT activity | [158] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



