
Submitted:
22 September 2023
Posted:
26 September 2023
You are already at the latest version
Abstract
Tomato pomace (TP), an agricultural industrial waste product from the tomato processing industry, was valorized by being used as a rearing substrates for Tenebrio molitor (L.). This study evaluated bran-based diets with increasing tomato pomace (0%, 27%, 41% and 100%). Protein sources, as brewer’s spent grain and yeast, are used in TP27 and TP41 diets, respectively, to ensure equal protein content to control diet (TP0). Results showed larval survival, larval and pupal weight no different between diets; however, growth time significantly increases in TP100 compared at all diets. The feed conversion rate progressively increases from 2.7 to 4.3, respectively from TP0 to TP100. Conversely, Lycopene and β-Carotene increases in the larvae. Fatty acid composition improves by increasing polyunsaturated fatty acids (mainly -Linoleic acid). Although the best nutritional quality was obtained in T100, the TP41 is the optimal diet for balance between larval performance and qualitative improvement of larvae. Therefore, tomato pomace is suitable for the formulation of mealworm diets, even in high dosages when supplemented with sustainable protein sources.
Keywords:
yellow mealworm
; edible insects
; by-products
; rearing substrates
; fatty acid
; nutraceutical
; antioxidant
; lycopene
; β-Carotene
1. Introduction
Use of by-products as growth substrate (from here below indicated as “diet”) allows to reduce production costs in rearing edible insects and increase their role as “bioconverters” in the circular economy [1,2]. This approach is mainly focused on Black Soldier Fly (Hermetia illucens L.; Diptera: Stratiomizidae) but extended to Yellow Mealworm (Tenebrio molitor L.; Coleoptera: Tenebrionidae) with the choice of suitable by-products and the correct formulation of diets. The diet affects the growth performance of T. molitor larvae or mealworms (MLW) [3] and the productivity of their adults [4]. Diet also has an impact on the nutraceutical composition of collected larvae [3,5]. It has frequently been thought that the latter ability would enhance the protein content and amino acid composition [6]. Diet affects the fatty acid composition [3,7,8] and a diet high in linseed has been shown to increase polyunsaturated fatty acids (PUFA) [9]. Diets have also been shown to improve calcium content and produce a beneficial Ca:P ratio [5,10,11].
Recently, the increase of antioxidant substances through diets has focused the interest of some researchers [12,13,14,15].
This approach is limited not only by the need to provide balanced diets for the nutritional requirements of the farmed insect but also by the anti-nutritional substances present in some by-products. Typical is the presence of polyphenols, as in olive pomace [16].
World tomato production is estimated at 34-42 million tonnes per year by the Word Processing Tomato Council (www.wptc.to). Tomato pomace (TP) is a byproduct of the tomato processing industry, consisting of 5–10% of the fresh weight of tomatoes [17]. It consists of peels, seed and, residual pulp [18]. The nutritional composition is influenced by the proportions of the different components and by the transformation process. Generally, its composition is made up of fiber (53.0%), sugars (25.7%), protein (19.3%), and fat (5.9%), on a dry weight basis [19]. It is also a source of carotenoids, especially lycopene and β-carotene in the peel [20]. Inclusion in animal feed has been tested for poultry [21], quail [22],ruminants [23], dairy cows [24], and lamb [17].
Classified among the phytochemicals, carotenoids as well as polyphenols [25] show marked antioxidant [26] and anti-inflammatory activity [27].
Due to these properties, several studies have associated their use with multiple health benefits, in particular α- and β-carotene and β-cryptoxanthin as a valuable source of vitamin A [28], lutein and β-carotene as positive adjuvants in many eye-related diseases, including cataracts and age-related macular degeneration [29], and finally lycopene and b-carotene as skin protectors from UV rays [30], adjuvans in the prevention of cancer [31] and heart health [32].
Use of this by-product to feed insects has been little investigated, although recently tomato was tested by H. illucens [33] and tomato pomace was included as a supplement (10% w/w) in diets for T. molitor [13]. A greater supplement would alter the initial nutritional composition of the diet; therefore, high-dose tomato pomace requires the formulation of specific isoproteic diets.
This work aims to evaluate diets assembled with increasing doses of tomato pomace, evaluating its influence on the growth performance and nutraceutical quality of the larvae to optimise the diet.
2. Materials and Methods
2.1. Yellow Mealworms Colony
Mealworms used in this work were reared at the insectarium of CIHEAM-Bari (in the Apulia region). T. molitor was fed a bran-based diet and yeast (ratio 95:5); the wet supplement was distributed twice a week with pieces of pumpkin (Cucurbita moschata, cv. Butternut). Rearing and subsequent experiments are conducted in a climatic room (28.0±1° C, 70±5% RH and 0L:24 D photoperiod).
2.2. Substrate composition/preparation
Four different by-products were used to formulate the tested diets. Local farms supplied tomato pomace as a by-product of the production of tomato sauce. The pomace consisted mainly of peels and seeds, while the presence of pulp was scarce. Bran was purchased from the mill (Molino “Cimminelli”, Montegiordano, CS, Italy) and derived from durum wheat milling. Brewer’s spent grain was supplied from small local brewers (Brewery “Jazz Beer”, Bernalda, MT, Italy). Zootechnical yeast, as a protein supplement, was purchased from Zabele Srl (Padova, PD, Italy). By-products were preliminarily dried at 60°C for 24 h in Food Dehydrator (COSORI, mod. CP267-FD-RXS, Anaheim, CA 92806, USA). Subsequently, matrices were sieved using a 2 mm manual sieve and the coarse part ground, ensuring homogeneity to avoid the influence of particle size [34]. The nutrient composition was determined on a representative sample using AOAC methods [35] (Table 1).
Diets were formulated with bran and increasing doses of tomato pomace. Mixtures also with brewer’s spent grain (TP27) or yeast (TP41) produced isoproteic diets, similar to the control diet (TP0). Finally, a diet with only tomato pomace (TP100) was tested. Table 2 reports the macro-nutrient and energy values (the latter calculated by conversion factors in Regulation (EU) 1169/2011, Annex XIV). All diets were assembled in the form of “cookies”, partly similar to previous procedures.[6,36], to avoid self-selection in mixed diets [37] and to facilitate the separation of the frass.
2.3. Experimental set-up
At the beginning of the experiment, six week-old larvae (28±1mg) were distributed in groups of 20 larvae [38] in plastic cups (bottom diameter 6 cm). Larvae were fed ad libitum with their respective diets and, wet supplements provided twice weekly. Complete randomization was applied to the experimental design, with 10 replicates/treatment and 20 larvae/replicate.
For each replicate, the final larval weight, the residual substrate, and the frass were weighed with an analytical scale (Mettler-Toledo, mod. B2002-S; precision ±0.1 mg).The collected larvae were starved for 48 h, blanched at 100°C for 5 min, and dried at 60 °C for 24 h in Food dehydrator in accordance with Melgar-Lalanne et al. [39]. Dried larvae, substrate, and frass were stored at -18 °C and powdered before chemical analysis. The latter was performed on three replicas per diet.
2.4. Mealworm growth perfomance
At the formation of the first pupa in the replica, were measured Larval survival (Equation (1)), Growth time (Equation (2)), Larval weight (Equation (3)), Pupal weight (Equation (4)), Feed Conversion Rate or FCR (Equation (5)) where FC represents the feed ingested excluding the wet supplement, Specific Growth Ratio or SGR (Equation (6)) where lFW and lIW represent final and initial fresh larval weight. The Efficiency of Conversion of Ingested feed or ECI (Equation (7)) and Efficiency of Conversion of Digested feed or ECD (Equation (8)) [40] were calculated considering the weight gained (WG), the feed consumed (FC) and Frass as dry weight.
Larval survival (%) = n. initial larvae/n. final larvae and pupae x 100
Growth time (d) = n. days between start of experiment and emergence of first pupa
Larval weight (mg) = weight larvae collected/n. larvae collected
Pupal weight (mg) = weight pupae collected/n. pupae collected
FCR = FC/WG
SGR (% day-1) = 100 x (lnFW-lnIW)/days
ECI (%) = [WG/(FC] x 100
ECD (%) = [WG/(FC-Frass)] x 100
2.5. Carotenoids analysis
Carotenoids were extracted from feed, mealworms and feces. According to Leni et al. [33], extraction was conducted with some modifications. Four hundred g of homogenized sample was mixed with 10 ml of hexane/ethanol/acetone (50:25:25) extraction mixture containing 0.1% ascorbic acid. After stirring at 200 rpm for 1 h on ice and under subdued light (Universal Table Shaker 709), the samples were centrifuged at 2,800 rpm at 4 °C for 20 min (Laborfuge 400R –Heraeus Instruments), and the supernatant was separated from the pellet. Separation and quantification of licopene and β-carotene were conducted according to Anthon and Barret [41], with some modifications. Distilled water (1.5 ml per 10 ml of extract) was added to the extract to cause phase separation. After stirring for 1 min under subdued light, the samples were centrifuged at 2,800 rpm at 4 °C for 10 min, and the upper hexane phase was recovered and used for spectrophotometric carotenoids quantification (Multiscan Go Spectrophotometer). Samples were read at 503 and 444 nm. The Concentration of Lycopene and β-Carotene was calculated using the following Equations (9) and (10):
where:
C lycopene (mg/kg) = (6.95 ×Abs.503 – 1.59 ×Abs.444)× 0.55 ×537×V/W
C β-carotene (mg/kg) = (9.38 × Abs.444 – 6.70 ×Abs.503) ×0.55 ×537V/W
- 0.55 = The final hexane layer volume ratio to the volume of mixed solvents added for hexane:acetone:ethanol (2:1:1)
- W (mg) = The weight of sample analyzed
- V (ml) = The volume of mixed solvents added
- 537 = The molecular weights of lycopene and β-carotene(g/mole).
2.6. Lipid analysisin mealworms
The defatting process of mealworm powder (MLWP) was performed according to Gkingali et al. [42] with some modifications. In brief, a three-step extraction procedure using n-hexane removed the fat from the MLWP. The sample was first mixed with n-hexane at a ratio of 1:5 w/v. The mixture was then shaken for 1 hour at 150 rpm (25 °C) using a rotary shaker (Universal Table Shaker 709). Following a centrifugation of the resultant slurry at 8,500g for 10 min at 10 °C, the organic phase in the supernatant was decanted. The supernatant is separated from the sediment and stored separately. The process was twice repeated, adding more hexane to the sediment each time. Supernatants were collected in a pre-weighed round-bottom flask, and the n-hexane was evaporated using a rotary evaporator (Steroglass Rotary Evaporator InstrumentsKentron-Strike 202). The final sediment was left to stand at room temperature to eliminate any remaining solvent. The resultant defatted larvae powder (DLP) and lipid extract were kept in a freezer at -18 C° until used. The oil extraction yield (or Crude Fat %) was calculated according to Equation (11) [43]:
Oil extraction yield (%) = [mass of extracted fat(g)/solids of the initial sample (g)] × 100
Lipids extracted from MLWP were directly trans esterified by producing fatty acid methyl esters (FAME) by applying the technique described by Tasselli et al [44]. The separation of FAMEs was carried out using an Agilent GC7890A gas chromatograph out fitted with a split-splitless injector and a flame ionisation detector (FID) at the settings specified by Di Fidio et al. [45]. The retention times of the fatty acids were compared to FAME standards (Sigma-Aldrich), and their percentage was estimated using the combined area of the present peaks.
The fatty acid profile data were processed by deriving indices. Indices of atherogenicity (AI) and thrombogenicity (IT) were calculated using Equations (12) and (13) previously described by Ulbricht and Southgate [46]:
Theoxidizability (COX) value was calculated using the following Equation (14) previously described to Fatemi et al. [47]
The Hypocholesterolemic/Hypercholesterolemic (HH) ratio was calculated as reported by Santos-Silva et al. [48] using the following Equation (15):
The Unsaturated index (UI) was calculated was calculated using the following Equation (16) previously described to Chen et al.[49]:
UI = [(1 × %monoenoics) + (2 × %dienoics) + (3 × % trienoics) + (4 × % tetraenoics) + (5 × % pentaenoics) + (6 ×% hexaenoics)]
2.7. Protein analysis
An alkaline protein extraction was performed on the DLP samples using the protocol made by Zhao et al. with minor modifications [50]. One g of sample was treated with 15 ml of 0.250 M NaOH at 40°C under agitation for 1 h in a thermostatic orbital shaker (ThermoScientific Forma, model 420) and centrifuged at 8,500 rpm for 30 min at 4°C for 3 times. The extraction procedure was repeated three more times in total. The supernatant and gel layer from all extractions were pooled and used for the quantification. The determination of crude protein content was performed according to Kotsou et al., with modifications [14]. Teen µL of the appropriately diluted supernatants pooled was transferred to the wells of a 96-well plate, and 200 µL of diluted Bradford reagent was added and then shaken for 30 seconds in a plate reader. The plate was incubated for 10 minutes at room temperature in the dark. The absorbance was measured at 595 nm with a Multiscan Go Spectrophotometer. A standard calibration curve was prepared using bovine serum albumin.
2.8. Statistical analysis
Larval performance data were initially submitted for normality and homogeneity of variance tests. A one-way ANOVA was applied at FCR, SRG, and ECI values, followed by Tukey-Kramer HDS test post-hoc to identify the differences between the diets. Alternatively, the non-parametric Kruskal–Wallis test and pairwise multiple comparisons with Bonferroni correction were applied to the other measured parameters.Significance was assumed at p< 0.05.All data were statistically processed by SPSS software version 26.0 (IBM Corporation, Armonk, NY, USA). The data of qualitative and quantitative analysis are shown as the mean values ± standard deviation (SD), from three replicas.
3. Results
3.1. Larval performances
Larval survival was close to 100% in all diets tested. No statistically significant differences were found between diets (H = 1.05; df = 3; p=0.788), including the tomato pomace diet (TP100) (Table 3).
Values of larval growth time showed significant differences between diets (H = 21.23; df = 3; p<0.000). The presence of tomato pomace increased the growth time by 5-6 days in the TP27 and TP41 diets, but these longer periods were not significantly different compared to the 32 days of the control diet (TP0). In contrast, the TP100 diet recorded growth times double (+33.4 days)that of the control and significantly longer than the other diets.
At harvest, the mean larval weight was not significantly different between diets (H = 6.41; df = 3; p=0.093).The lowest weight (91.0 mg) was achieved by the TP100 diet, and the highest weight (109.0 mg) by the larvae of the TP41 diet. The analysis of the weights of the first pupa showed significant differences between diets (H = 12.61; df = 3; p<0.006), although significant differences were found only between the TP41 (127.0 mg) and TP100 (101.0 mg) diets (Table 3).
3.2. Efficiency indicators
Generally, the utilization efficiency of the tested diets decreased with increasing doses of tomato pomace in the diets. The Feed Conversion Rate was significantly different between diets (F = 73.2; df = 3, 36; p< 0.001) with minimum values (FCR= 2.7) in the control, without tomato pomace, and progressively increasing until reaching the maximum in T100 (FRC=4.3) (Table 4).
The Specific Growth Ratio significantly differed between diets (F = 34.1; df = 3, 36; p <0.001). The highest values were obtained with the control diet (4.9%), while slightly lower values were recorded with the two diets mixed with tomato pomace (TP27 and TP41). The TP100 diet achieved significantly lower values (2.5%).
The Efficiency of Conversion of Ingested feed significantly differed between diets (F = 85.8; df = 3.36; p < 0.001). The ECI values decreased significantly as the amount of tomato pomace increased in the tested diets (Table 4).
The Efficiency of Conversion of Digested feed was significantly different between diets (H = 27.3; df = 3; p <0.001). There was no significant difference between the TP0 and TP41 diets, while the TP27 diet showed significantly higher ECD values (42.8%). Finally, significantly higher values than all other diets were obtained in the TP100 diet, with an ECD of 65.9% (Table 4).
3.3. Licopene and β-carotene Quantification
The values of carotenoids, lycopene and β-carotene, present in feed (diets), stored in mealworms, and excreted with frass, have been collected in Table 5. In feed, the addition of tomato pomace shows a proportional increase in the amount of lycopene, and β-carotene compared to the control: from 2.66 ug/g TP0 to 179.75 ug/g TP100 for lycopene; from 0.30 ug/g TP0 to 241.5 ug/g TP100 for β-carotene. Furthermore, an inversion of the lycopene/β-carotene (L/C) ratio between feeds is also evident. While the TP0 diet shows an L/C value >1 (8.8), all the supplemented feeds (TP21, TP47, and TP100) show a value < 1 (~ 0.5).Values referring to the larvae highlighted a general tendency towards the accumulation of both analyzed carotenoids. In the TP47 and TP100 diets, compared to TP0, there are incremental signals of both lycopene (0.61 ug/g and 1.19 ug/g against 0.08 ug/g) and β-carotene (2.56 ug/g and 7.28 ug/g versus 1.43 ug/g). However, the larvae are richer in β-carotene than in lycopene. In larvae, the increase of both carotenoids is evident only at TP27 (maximum at TP100), unlike the progressive increase observed in diets. The lycopene content in frass varies from 0.70 ug/g in the TP0 to 39.67ug/g in the TP100, while the B-carotene content varies from 12.09 ug/g in the TP0 to 147.46 ug/g in the TP100; so as seen for the larvae, the frass is richer in β-carotene than in lycopene.
3.4. Larval nutritional value
Figure 1 shows the fat extracted (% w/w of MLW powder) from larvae fed different diets enriched with tomato pomace (from 27 to 100%). In our results, the control larvae (TP0) contained 30.76% crude fat. Adding tomato pomace to the feed in percentages of 27% (TP27) does not generate any difference compared to the control diet. However, when tomato pomace was present at 41% (TP41) and 100% (TP100), there was a decrease in the fat content of the larvae. Specifically, TP41 larvae contained 19.6% less fat than control larvae, while TP100 larvae contained as much as 67.9% less fat. Total fatty acid content (TFA) analysis also shows a decreasing trend in values compared to TP0 (81%). In particular, TP41larvae contained about 77% TFA, while TP100 contained 53%, following the trend of crude fat.
The fatty acid composition of mealworms shows a total of 7 fatty acids detected and measured in all treatments (Table 6). In TP0 larvae, the main unsatured fatty acids(UFAs) found were oleic acid (OA) (50.2%), followed by linoleic acid (LA) (25%), while of the satured fatty acids (SFAs), the most abundant was palmitic acid (PA) (15%).While introducing a diet supplementation of tomato pomace produced no significant change in the amount of SFAs compared to the control, it produced a significant qualitative and quantitative change in UFAs. In fact, in diets TP27, TP41, and TP100, there is a decreasing trend in the percentage of oleic acid (OA), which falls from 50% (TP0) to 26%, and a simultaneous significant increase in PUFAs. The content of linoleic acid increases from 25% (TP0) to 40%, while that of linolenic acid varies greatly from 0.42% to 2.68%.The PUFA:SFA ratio, calculated to assess our samples' cardiovascular health benefits, is higher in all 3 case studies (TP27, TP41, and TP100) than the control TP0 larvae (1.18). Their PUFA:SFA ratio shows an increasing trend, with values between 1.35 (TP27) and 2.30 (TP100). The influence of diet on the n-6/n-3 ratio highlights that all larvae fed with tomato pomace supplementation significantly reduced their n-6/n-3 ratio. The greatest 70% reduction was obtained with the TP100 diet, followed by 46% of the TP41, compared to the TP0, which had an n-6/n-3 ratio of 61.5.
3.4.1. Lipid quality indices
Diet had a direct influence on both COX and UI. As shown in Table 7, in all cases of tomato-fed larvae (TP27, TP41, and TP100), there was a substantial increase in the COX value compared to the TP0 value, equal to 3.2. The TP41 and TP100 diets, in particular, increased by +25% and +80% respectively. The UI data also follow an increasing trend; in fact, compared to the control values TP41 and TP100, they show an increase of +7.7% and +25% respectively. Among the lipid quality indices related to the incidence of coronary heart disease, IA, IT, and HH do not appear to be significantly influenced by diet, except for the TP100 diet. Larvae fed with 100% tomato pomace showed IA and IT decreased, respectively by -20% and -12% compared to the control, and HH increased by +18% compared to the control.
Indices abbreviations: COX (Calculated oxidizability value); IT (Indices of thrombogenicity); IA (Indice of atherogenicity); HH (Hypocholesterolemic/Hypercholesterolemic ratio); UI (Unsaturation index)
3.4.2. Crude proteine
Larvae fed on control, TP27, and TP41 diets showed similar crude protein content (Figure 2). Their values vary from 47.3% to 49.2%, expressed on the DM of larvae defatted. In contrast, crude protein in larvae on the TP100 diet decreased at 42%.
4. Discussion
The use of by-products of vegetable origin in the feeding of edible insects must consider the presence of toxic secondary metabolites or with a negative influence on their development. The major secondary metabolite of tomato fruit is the glycoalkaloid tomatine, known for its repellent and antifeeding effects [51,52]. This abounds in unripe fruit and degrades with ripening [53], so it is not a limiting factor in the use of tomato pomace. The ripe fruit contains polyphenolic compounds such as gallic acid, chlorogenic acid, caffeic acid, rutin, kaempfero-3-O-glucoside, naringin, quercetin, and naringenin [54]. Some of these compounds have shown a negative and dose-dependent influence on the feeding of Lepinotarsa decemlineata (Say) [55]. Others, such as chlorogenic acid, have a repellent and antifeeding effect [56] or, like quercitin, they can be phagostimulant or phagodeterrent according to the doses [57]. As far as we know, it is not known whether the industrial tomato transformation process degrades the secondary metabolites, but the absence of mortality in the TP100 diet suggests their absence or presence at non-lethal doses. This is contrary to the hypothesis of Ruschioni et al. [16] in the presence of high mortality in larvae fed on olive pomace.
The nutritional composition is another important element in the use of by-products. The tomato pomace used in this study is particularly poor in macro-nutrients, probably because it derives from a very efficient industrial process [19]. Its content in protein (P 9.5%), carbohydrate (C 8.9%), and lipids (L 3.2%) is unfavorable in carbohydrates when compared to optimal compositions such as P 20-25%, C 65-75%,and L 3- 12% [36] or P 19.9-22.8%, C 67.3-71.5% and L 8.6-10% [6].
TP's protein to carbohydrate (P:C) ratio is 1:0.9 and was similar to the 1:1 ratio, considered the best among those evaluated by Rho and Lee [50]. However, these authors tested adults on synthetic diets and reported the “tendency to prioritise the regulation of carbohydrate intake over that of protein intake” [58]. More specifically, the ratio would be 1:1.6 for males and 1:1.3 for females [59]. This would indicate the need to supplement TP with carbohydrates. However, the TP100 diet results indicate that protein and carbohydrate deficiency had a more prevalent effect on their P:C ratio. TP100 larvae had lower protein content than larvae from TP0, TP27, and TP41 isoproteic diets, in contrast to the expected maximum protein accumulation with a ratio between 1:1 and 2:1 [58].
The TP100 diet has the lowest energy value (236.6 kcal/100 g) and the TP0 diet has the highest (318.3 kcal/100 g), but they all have lower energy values than the poorest diet (353 kcal/100 g) reported by other authors [16]. The low energy value of TP is determined by the high fiber content (67.1%), considered unfavorable if more than 5-10% in diets for T. molitor [60].
A comparison of the diets tested showed limited differences in larval performance. Major and minor larval and pupal weights were obtained in the TP41 and TP100 diets, respectively; however, only the pupal weight in the TP41 diet was significantly higher. The significant increase in larval growth time in the TP100 diet (twice as much as compared to the control) is a potential limitation in using pure TP due to the consequent increase in breeding costs. The increase in larval growth times can be mainly attributed to the low protein content, as there are no differences between isoproteic diets. This hypothesis is in agreement with the reduction of larval growth times observed in diets richer in proteins [61].
Results on the use of diets have highlighted the significant increase in the FCR value as the dose of TP increases. This result is expected in TP100 as a remedy to compensate for the low concentration of nutrients [58]. Furthermore, the increase in FCR values in isoproteic diets suggests a positive correlation to the fiber content and a negative correlation to the energy value of the diet. Our FCR values (from 2.7 to 4.3 for TP0 and TP100, respectively) are slightly higher than some control diets, such as wheat bran and yeast (FCR=2.3) [38], chicken feed (FCR=1.57), and wheat bran (FCR=2.08) [3]. It is important to point out that the FCR value (3.8) recorded in the TP41 diet is similar to 3.44 of the commercial diet for T. molitor used by van Broekhoven et al [62], however, with similar protein values (16.5 and 17.1%, respectively).
The tested diets have SRG values in line with the seed clearing process by-products values (2.7 to 7.2% day-1) [38], but lower than the mixed diets (8.2 to 11.9% day-1) [6]. The particularly low value (2.5% day-1) of the TP100 diet is probably influenced by the long larval growth time.
The ECI values significantly decreased from 15.4% to 9.8% with the increase of the TP in the diets; they are lower than commercial diet (18.96%) [62] and chicken feed (almost 22%) [3]. However, we find our results better comparable to Kroncke and Benning [6] and Morales- Ramos et al. [63] for the similar mode of diet administration. Administration through "cockies" greatly reduces self-selection in mixed diets, thus reducing the possibility of self-reducing the negative impact of unbalanced diets or fibres.In this case, our ECI values fall within the range 5.5-18.4% described by Kroncke and Benning [6] and higher than the range 7-10% [63], where the two best diets have ECI values similar to the TP100 diet (with 9.8%).
The ECD values (30.0 to 65.9%) were higher compared to the values (17 to 20%) found in larval density tests [64]. The limited knowledge on ECD and the high value of the TP100 diet suggest more studies, hypothesizing better conversion of the digested diet if it is poor in nutrients.
The positive influence of tomato pomace on larval quality is evidenced by carotenoids and fatty acid composition. The degree of accumulation of carotenoids observed in the larvae is very low if correlated with that contained in the substrate and in the faeces. This reduced efficiency of larval accumulation against an evident enrichment in frass carotenoids is in agreement with other data present in the literature on mealworms fed with former foodstuffs [65] and H. illucens fed with agri-food by-products (ground and coarse tomato) [33]. The use of commercial β-carotene supplements administered to insects shows larval accumulation values comparable to our results [66], as well as MLW fed with leaves of Moringa oleifera (Lam.) [15] or with carrot pomace [67].
The lycopene content in the larvae appears to be much lower than the β-carotene content accumulated in the substrate and faeces. This data is in agreement with the reduction of the mass balance also found for other non-provitamin A carotenoids such as zaexanthin and lutein in T.molitor [65] and in H. illucens [33] probably due to bioconversion phenomena by the 'insect (β-cyclase and carotene-9′,10′-monooxygenase) [68,69] or by the gut microbial community [70].
Of great interest is the observation of the effect of diet on the quantity and quality of lipids and FAs. In the MLW, lipids are second only to proteins in quantity [71]. TP0 larvae show a CF value (~30.8%) that is very comparable with other previously published data in which there is great variability in its concentration (from 22% to 42%) [14,43,72]. Insects and, in particular, MLW are equipped with a sophisticated enzymatic kit (Elongase and Desaturase) that allows them to synthesise de novo fatty acids and, in particular, PUFAs [9,73,74].
Furthermore, they can modulate the degree of lipid accumulation and change their profile in FA depending on the developmental stage, sex, growth environment, and especially the type of feed used [75,76,77,78,79,80].
Larvae fed with 100% TP showed a reduction in the percentage of CF compared to the control. This decrease agrees with many studies showing how caloric restriction, total carbohydrate intake in the diet, and, in particular, the addition of fatty acid and carotenoid supplements affect fat synthesis and accumulation in insect larvae and other animals [15,59,62,81,82]. Low levels of intramuscular fat are found in lamb-fed diets enriched in lycopene [83] and in pork fed diets, rich in linoleic acid [84], and supplemented with 15% TP [85].
The FAs composition of TP0 larvae also confirms it as one of the most abundant sources of OA, PA, and especially linoleic acid (LA) compared to other animal sources that are rich in fatty acids and especially omega-6 such as chicken fat and egg yolk [86]. LA and linolenic acid (ALA) are PUFAs defined as 'essential' for the human body, which is unable to synthesise them [87] and are therefore essential for human health and, in particular, for the prevention of cardiovascular disease, one of the leading causes of death worldwide [9].
The increase in the amount of ω-3 in feeds due to TP led to a general increase in PUFAs, especially LA and ALA, and to a decrease in both OA and ω-6/ω-3 ratios in especially TP41 and TP100 diets (-45%, -70%). Diets high in the ω-3/ω-6 ratio cause an increase in PUFA and ω-3/ω-6 ratios in larvae [88], and can modulate the activity of both [89] Δ-12 desaturase, which converts ω-9 oleic acids into ω-6 linoleic acids [90], and Elongase (TmElo1 and TmElo2) involved in the synthesis of PUFAs [74].
Our results are also in agreement with other data where MLW are fed feed supplemented with linseed, grape seeds, and winery waste sludge and show a reduction in MUFA content [9,12]. In contrast, diets with distillery by-products (grape pomace, exhausted grape marcs, grape skin pulp [12], or sunflower [91]) produced a significant increase in MUFAs and, in particular, OA, while the inclusion of olive pomace in the feed composition did not affect the FA composition of body lipids [16]. Our results are also in agreement with the improvement in the quality and quantity of PUFAs previously obtained with the addition of fish oil [92]. All this emphasizes how physiological mechanisms of MLW adaptation play a key role in the quality of the lipid profile of larvae on par with diets [62,65,88,93].
Incorporating TP into feeds also increased the wholesomeness of mealworms for human and animal consumption, as indicated by the lipid indices obtained. The increase in COX emphasizes the positive influence of diets rich in PUFAs on the stability and shelf life of by-products obtainable from MLW [14,94,95], while the increase in UI, comparable to some macroalgae (Hypnea esperi, Gracilaria fergusonii, Codium vermilara) [49], shows the strong impact of diet on increasing the percentage of high-quality PUFAs useful for reducing the risk of heart disease [96], preventing and managing type 2 diabetes, insulin resistance [97], osteoarthritis [98] and neurological disorders [99].
The absence of adverse effects on the IA, IT, and HH indices, however, makes these larvae comparable to other diets applied to MLW [15,89] and other valid novel foods such as brown seaweed, whose consumption produces the best results for human health as it has a positive effect against cardiovascular diseases [100].
Therefore, the use of an optimal TP-based diet can support the production of mealworms with higher nutraceutical value. Whole larvae and mealworm oil can be commercial products with specific health characteristics since the presence of carotenoids and the best chemical composition are crucial for the future use of this insect as feed and food [95,101].
5. Conclusions
Tomato pomace is a by-product rich in fiber, nutritionally poor and unbalanced. Its use in high doses in mealworm diets must include other sources of carbohydrates and proteins. In this study, the optimal diet was assembled with wheat bran (50%), tomato pomace (41%), and yeast (9%).This diet had no negative impact on larval performance and increased the content of carotenoids and polyunsaturated fatty acids. Further studies should point to the replacement of yeast with a cheaper protein source. The use of pure tomato pomace further increases the content of lycopene and β-carotene in the larvae and doubles the PUFA values. However, such a poor diet doubles the larval growth time and reduces the protein and fat content of the larvae. The latter use could only find application in the economic valorization of the greater nutraceutical value of the larvae.
Author Contributions
Conceptualization, F.B. and V.V.; methodology, F.B., A.S., F.L. and M.L.V.; validation, F.B., A.S., F.L. and M.L.V.; formal analysis, F.B., A.S. and F.L.; investigation, F.B., A.S., F.L. and M.L.V.; resources, F.B., A.S. and F.L.; data curation, F.B., A.S., F.L. and M.L.V.; writing—original draft preparation, F.B., A.S.; writing—review and editing, F.B., A.S. and F.L.; supervision, F.B.; project administration, F.B. and V.V.; funding acquisition, F.B. and V.V. All authors have read and agreed to the published version of the manuscript.
Funding
This research was partially funded by Project CoRoSect (Cognitive Robotic System for Digitalized and Networked (Automated) Insect Farms). H2020-E.U. 2.1.1. - INDUSTRIAL LEADERSHIP - Leadership in enabling and industrial technologies - Information and Communication Technologies (ICT). (Grant agreement No 101016953).
Data Availability Statement
All data from this experiment are contained in the article.
Acknowledgments
The authors thank Dr. Rosalia Viti for technical support in Insectarium CIHEAM-B and Mr. Vito Valerio (ENEA- R. C. Trisaia) for fatty acids analysis.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Cadinu, L.A.; Barra, P.; Torre, F.; Delogu, F.; Madau, F.A. Insect Rearing: Potential, Challenges, and Circularity. Sustainability 2020, 12, 4567. [Google Scholar] [CrossRef]
- Jensen, H.; Elleby, C.; Domínguez, I.P.; Chatzopoulos, T.; Charlebois, P. Insect-Based Protein Feed: From Fork to Farm. J. Insects as Food Feed 2021, 7, 1219–1233. [Google Scholar] [CrossRef]
- Bordiean, A.; Krzyżaniak, M.; Aljewicz, M.; Stolarski, M.J. Influence of Different Diets on Growth and Nutritional Composition of Yellow Mealworm. Foods 2022, 11, 3075. [Google Scholar] [CrossRef] [PubMed]
- El Deen, S.N.; Lamaj, F.; Verrastro, V.; Al Bitar, L.; Baldacchino, F. Effects of Two Diets on Adults’ Survival and Productivity in Mass-Rearing of Tenebrio Molitor (Coleoptera: Tenebrionidae). J. Insects as Food Feed 2021, 7, 1149–1157. [Google Scholar] [CrossRef]
- Noyens, I.; Schoeters, F.; Van Peer, M.; Berrens, S.; Goossens, S.; Van Miert, S. The Nutritional Profile, Mineral Content and Heavy Metal Uptake of Yellow Mealworm Reared with Supplementation of Agricultural Sidestreams. Sci. Rep. 2023, 13, 11604. [Google Scholar] [CrossRef]
- Kröncke, N.; Benning, R. Influence of Dietary Protein Content on the Nutritional Composition of Mealworm Larvae (Tenebrio Molitor L.). Insects 2023, 14, 261. [Google Scholar] [CrossRef]
- Riekkinen, K.; Väkeväinen, K.; Korhonen, J. The Effect of Substrate on the Nutrient Content and Fatty Acid Composition of Edible Insects. Insects 2022, 13, 590. [Google Scholar] [CrossRef]
- Dreassi, E.; Cito, A.; Zanfini, A.; Materozzi, L.; Botta, M.; Francardi, V. Dietary Fatty Acids Influence the Growth and Fatty Acid Composition of the Yellow Mealworm Tenebrio Molitor (Coleoptera: Tenebrionidae). Lipids 2017, 52, 285–294. [Google Scholar] [CrossRef]
- FRANCARDI, V.; CITO, A.; FUSI, S.; FUSI, S.; BOTTA, M.; DREASSI, E. LINSEED TO INCREASE N-3 FATTY ACIDS IN TENEBRIO MOLITOR (COLEOPTERA TENEBRIONIDAE). Redia 2017, 73–76. [Google Scholar] [CrossRef]
- Anderson, S.J. Increasing Calcium Levels in Cultured Insects. Zoo Biol. 2000, 19, 1–9. [Google Scholar] [CrossRef]
- Latney, L. V.; Toddes, B.D.; Wyre, N.R.; Brown, D.C.; Michel, K.E.; Briscoe, J.A. Effects of Various Diets on the Calcium and Phosphorus Composition of Mealworms (Tenebrio Molitor Larvae) and Superworms (Zophobas Morio Larvae). Am. J. Vet. Res. 2017, 78, 178–185. [Google Scholar] [CrossRef] [PubMed]
- Brai, A.; Vagaggini, C.; Pasqualini, C.; Poggialini, F.; Tarchi, F.; Francardi, V.; Dreassi, E. Use of Distillery By-Products as Tenebrio Molitor Mealworm Feed Supplement. J. Insects as Food Feed 2023, 9, 611–623. [Google Scholar] [CrossRef]
- Brai, A.; Poggialini, F.; Trivisani, C.I.; Vagaggini, C.; Tarchi, F.; Francardi, V.; Dreassi, E. Efficient Use of Agricultural Waste to Naturally Fortify Tenebrio Molitor Mealworms and Evaluation of Their Nutraceutical Properties. J. Insects as Food Feed 2023, 9, 599–610. [Google Scholar] [CrossRef]
- Kotsou, K.; Chatzimitakos, T.; Athanasiadis, V.; Bozinou, E.; Adamaki-Sotiraki, C.; Rumbos, C.I.; Athanassiou, C.G.; Lalas, S.I. Waste Orange Peels as a Feed Additive for the Enhancement of the Nutritional Value of Tenebrio Molitor. Foods 2023, 12, 783. [Google Scholar] [CrossRef] [PubMed]
- Kotsou, K.; Chatzimitakos, T.; Athanasiadis, V.; Bozinou, E.; Rumbos, C.I.; Athanassiou, C.G.; Lalas, S.I. Enhancing the Nutritional Profile of Tenebrio Molitor Using the Leaves of Moringa Oleifera. Foods 2023, 12, 2612. [Google Scholar] [CrossRef] [PubMed]
- Ruschioni, S.; Loreto, N.; Foligni, R.; Mannozzi, C.; Raffaelli, N.; Zamporlini, F.; Pasquini, M.; Roncolini, A.; Cardinali, F.; Osimani, A.; et al. Addition of Olive Pomace to Feeding Substrate Affects Growth Performance and Nutritional Value of Mealworm (Tenebrio Molitor L.) Larvae. Foods 2020, 9, 317. [Google Scholar] [CrossRef]
- Fondevila, M.; Guada, J.A.; Gasa, J.; Castrillo, C. Tomato Pomace as a Protein Supplement for Growing Lambs. Small Rumin. Res. 1994, 13, 117–126. [Google Scholar] [CrossRef]
- Heuzé V.; Tran G.; Hassoun P.; Bastianelli D.; Lebas F. Tomato Pomace, Tomato Skins and Tomato Seeds. Feed. a Program. by INRAE, CIRAD, AFZ FAO 2021.
- Del Valle, M.; Cámara, M.; Torija, M.-E. Chemical Characterization of Tomato Pomace. J. Sci. Food Agric. 2006, 86, 1232–1236. [Google Scholar] [CrossRef]
- Knoblich, M.; Anderson, B.; Latshaw, D. Analyses of Tomato Peel and Seed Byproducts and Their Use as a Source of Carotenoids. J. Sci. Food Agric. 2005, 85, 1166–1170. [Google Scholar] [CrossRef]
- Mansoori, B.; Modirsanei, M.; Radfar, M.; Kiaei, M.M.; Farkhoy, M.; Honarzad, J. Digestibility and Metabolisable Energy Values of Dried Tomato Pomace for Laying and Meat Type Cockerels. Anim. Feed Sci. Technol. 2008, 141, 384–390. [Google Scholar] [CrossRef]
- Reda, F.M.; Madkour, M.; El-Azeem, N.A.; Aboelazab, O.; Ahmed, S.Y.A.; Alagawany, M. Tomato Pomace as a Nontraditional Feedstuff: Productive and Reproductive Performance, Digestive Enzymes, Blood Metabolites, and the Deposition of Carotenoids into Egg Yolk in Quail Breeders. Poult. Sci. 2022, 101, 101730. [Google Scholar] [CrossRef] [PubMed]
- Marcos, C.N.; de Evan, T.; Molina-Alcaide, E.; Carro, M.D. Nutritive Value of Tomato Pomace for Ruminants and Its Influence on In Vitro Methane Production. Animals 2019, 9, 343. [Google Scholar] [CrossRef] [PubMed]
- Weiss, W.P.; Frobose, D.L.; Koch, M.E. Wet Tomato Pomace Ensiled with Corn Plants for Dairy Cows. J. Dairy Sci. 1997, 80, 2896–2900. [Google Scholar] [CrossRef] [PubMed]
- Nesci, S.; Spagnoletta, A.; Oppedisano, F. Inflammation, Mitochondria and Natural Compounds Together in the Circle of Trust. Int. J. Mol. Sci. 2023, 24, 6106. [Google Scholar] [CrossRef] [PubMed]
- Fiedor, J.; Burda, K. Potential Role of Carotenoids as Antioxidants in Human Health and Disease. Nutrients 2014, 6, 466–488. [Google Scholar] [CrossRef] [PubMed]
- Rubin, L.P.; Ross, A.C.; Stephensen, C.B.; Bohn, T.; Tanumihardjo, S.A. Metabolic Effects of Inflammation on Vitamin A and Carotenoids in Humans and Animal Models. Adv. Nutr. 2017, 8, 197–212. [Google Scholar] [CrossRef]
- Grune, T.; Lietz, G.; Palou, A.; Ross, A.C.; Stahl, W.; Tang, G.; Thurnham, D.; Yin, S.; Biesalski, H.K. β-Carotene Is an Important Vitamin A Source for Humans. J. Nutr. 2010, 140, 2268S–2285S. [Google Scholar] [CrossRef]
- de Almeida Torres, R.J.; dos Anjos Ferreira, A.L.; Luchini, A.; de Almeida Torres, R.J.; Correa, C.R. The Role of Non-Enzymatic Antioxidants on Age-Related Macular Degeneration. Front. Drug Chem. Clin. Res. 2022, 5, 1–19. [Google Scholar]
- Heinrich, U.; Wiebusch, M.; Tronnier, H.; Gärtner, C.; Eichler, O.; Sies, H.; Stahl, W. Supplementation with β-Carotene or a Similar Amount of Mixed Carotenoids Protects Humans from UV-Induced Erythema. J. Nutr. 2003, 133, 98–101. [Google Scholar] [CrossRef]
- Palozza, P.; Simone, R.E.; Catalano, A.; Mele, M.C. Tomato Lycopene and Lung Cancer Prevention: From Experimental to Human Studies. Cancers (Basel) 2011, 3, 2333–2357. [Google Scholar] [CrossRef]
- Jacques, P.F.; Lyass, A.; Massaro, J.M.; Vasan, R.S.; D’Agostino Sr, R.B. Relationship of Lycopene Intake and Consumption of Tomato Products to Incident CVD. Br. J. Nutr. 2013, 110, 545–551. [Google Scholar] [CrossRef] [PubMed]
- Leni, G.; Maistrello, L.; Pinotti, G.; Sforza, S.; Caligiani, A. Production of Carotenoid-Rich Hermetia Illucens Larvae Using Specific Agri-Food by-Products. J. Insects as Food Feed 2023, 9, 171–181. [Google Scholar] [CrossRef]
- NASER EL DEEN, S.; SPRANGHERS, T.; BALDACCHINO, F.; DERUYTTER, D. The Effects of the Particle Size of Four Different Feeds on the Larval Growth of Tenebrio Molitor (Coleoptera: Tenebrionidae). Eur. J. Entomol. 2022, 119, 242–249. [Google Scholar] [CrossRef]
- AOAC (Association of Official Analytical Chemists) No Title. Off. Methods Anal. 1990, Virginia, Arlington.
- Morales-Ramos, J.A.; Rojas, M.G.; Shapiro-llan, D.I.; Tedders, W.L. Use of Nutrient Self-Selection as a Diet Refining Tool in Tenebrio Molitor (Coleoptera: Tenebrionidae). J. Entomol. Sci. 2013, 48, 206–221. [Google Scholar] [CrossRef]
- RHO, M.S.; Pum LEE, K. Nutrient-Specific Food Selection Buffers the Effect of Nutritional Imbalance in the Mealworm Beetle, Tenebrio Molitor (Coleoptera: Tenebrionidae). Eur. J. Entomol. 2015, 112, 251–258. [Google Scholar] [CrossRef]
- Rumbos, C.I.; Bliamplias, D.; Gourgouta, M.; Michail, V.; Athanassiou, C.G. Rearing Tenebrio Molitor and Alphitobius Diaperinus Larvae on Seed Cleaning Process Byproducts. Insects 2021, 12, 293. [Google Scholar] [CrossRef] [PubMed]
- Melgar-Lalanne, G.; Hernández-Álvarez, A.; Salinas-Castro, A. Edible Insects Processing: Traditional and Innovative Technologies. Compr. Rev. Food Sci. Food Saf. 2019, 18, 1166–1191. [Google Scholar] [CrossRef]
- Waldbauer, G.B. The Consumption and Utilization of Food by Insects. Adv. Insect Physiolgy 1968, 5, 229–288. [Google Scholar]
- Anthon, G.; Barrett, D.M. STANDARDIZATION OF A RAPID SPECTROPHOTOMETRIC METHOD FOR LYCOPENE ANALYSIS. Acta Hortic. 2007, 111–128. [Google Scholar] [CrossRef]
- Gkinali, A.-A.; Matsakidou, A.; Paraskevopoulou, A. Characterization of Tenebrio Molitor Larvae Protein Preparations Obtained by Different Extraction Approaches. Foods 2022, 11, 3852. [Google Scholar] [CrossRef] [PubMed]
- Laroche; Perreault; Marciniak; Gravel; Chamberland; Doyen Comparison of Conventional and Sustainable Lipid Extraction Methods for the Production of Oil and Protein Isolate from Edible Insect Meal. Foods 2019, 8, 572. [CrossRef] [PubMed]
- Tasselli, G.; Filippucci, S.; Borsella, E.; D’Antonio, S.; Gelosia, M.; Cavalaglio, G.; Turchetti, B.; Sannino, C.; Onofri, A.; Mastrolitti, S.; et al. Yeast Lipids from Cardoon Stalks, Stranded Driftwood and Olive Tree Pruning Residues as Possible Extra Sources of Oils for Producing Biofuels and Biochemicals. Biotechnol. Biofuels 2018, 11, 147. [Google Scholar] [CrossRef] [PubMed]
- Fidio, N. Di; Liuzzi, F.; Mastrolitti, S.; Albergo, R.; Bari, I. De Single Cell Oil Production from Undetoxified Arundo Donax L. Hydrolysate by Cutaneotrichosporon Curvatus. J. Microbiol. Biotechnol. 2019, 29, 256–267. [Google Scholar] [CrossRef] [PubMed]
- Ulbricht, T.L.V.; Southgate, D.A.T. Coronary Heart Disease: Seven Dietary Factors. Lancet 1991, 338, 985–992. [Google Scholar] [CrossRef] [PubMed]
- Fatemi, S.H.; Hammond, E.G. Analysis of Oleate, Linoleate and Linolenate Hydroperoxides in Oxidized Ester Mixtures. Lipids 1980, 15, 379–385. [Google Scholar] [CrossRef]
- Santos-Silva, J.; Bessa, R.J.; Santos-Silva, F. Effect of Genotype, Feeding System and Slaughter Weight on the Quality of Light Lambs. Livest. Prod. Sci. 2002, 77, 187–194. [Google Scholar] [CrossRef]
- Chen, J.; Liu, H. Nutritional Indices for Assessing Fatty Acids: A Mini-Review. Int. J. Mol. Sci. 2020, 21, 5695. [Google Scholar] [CrossRef]
- Zhao, X.; Vázquez-Gutiérrez, J.L.; Johansson, D.P.; Landberg, R.; Langton, M. Yellow Mealworm Protein for Food Purposes - Extraction and Functional Properties. PLoS One 2016, 11, e0147791. [Google Scholar] [CrossRef]
- NISHA, R.; RAJAVEL, D.S. Effect of Tomatine on Termites Odontotermes Wallonensis (Wasmann) Vis-a-Vis Antifeedant and Repellent Activity. Int. J. Plant Prot. 2016, 9, 97–101. [Google Scholar] [CrossRef]
- Lu, F.M.; Chu, Y.I. Antifeeding Effects Of-Tomatine on Larvae of the Diamondback Moth (Plutella Xylostella L. CHINESE J. Entomol. 1992. [Google Scholar]
- Friedman, M. Analysis of Biologically Active Compounds in Potatoes (Solanum Tuberosum), Tomatoes (Lycopersicon Esculentum), and Jimson Weed (Datura Stramonium) Seeds. J. Chromatogr. A 2004, 1054, 143–155. [Google Scholar] [CrossRef] [PubMed]
- Zanfini, A.; Franchi, G.; Massarelli, P.; Corbini, G.; Dreassi, E. PHENOLIC COMPOUNDS, CAROTENOIDS AND ANTIOXIDANT ACTIVITY IN FIVE TOMATO (LYCOPESICON ESCULENTUM MILL.) CULTIVARS. Ital. J. FOOD Sci. 2017, 29. [Google Scholar]
- Pavela, R. The Feeding Effect of Polyphenolic Compounds on the Colorado Potato Beetle [Leptinotarsa Decemlineata (Say)]. Pest Technol. 2007, 1, 81–84. [Google Scholar]
- Kumar, M.; Umesh, K.P.; Pandey, P.P.; Firake, D.M.; Pandit, S.S. Eggplant{\textquoteright}s Foliar Chlorogenic Acid Provides Resistance against the Tropical Armyworm. bioRxiv 2023. [Google Scholar] [CrossRef]
- Diaz Napal, G.N.; Palacios, S.M. Bioinsecticidal Effect of the Flavonoids Pinocembrin and Quercetin against Spodoptera Frugiperda. J. Pest Sci. (2004) 2015, 88, 629–635. [Google Scholar] [CrossRef]
- Rho, M.S.; Lee, K.P. Geometric Analysis of Nutrient Balancing in the Mealworm Beetle, Tenebrio Molitor L. (Coleoptera: Tenebrionidae). J. Insect Physiol. 2014, 71, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Rho, M.S.; Lee, K.P. Balanced Intake of Protein and Carbohydrate Maximizes Lifetime Reproductive Success in the Mealworm Beetle, Tenebrio Molitor (Coleoptera: Tenebrionidae). J. Insect Physiol. 2016, 91–92, 93–99. [Google Scholar] [CrossRef]
- Li, L.; Stasiak, M.; Li, L.; Xie, B.; Fu, Y.; Gidzinski, D.; Dixon, M.; Liu, H. Rearing Tenebrio Molitor in BLSS: Dietary Fiber Affects Larval Growth, Development, and Respiration Characteristics. Acta Astronaut. 2016, 118, 130–136. [Google Scholar] [CrossRef]
- Morales Ramos, J.; Rojas, M.-G.; Shapiro Ilan, D.; TEDDERS, W. Developmental Plasticity in Tenebrio Molitor (Coleoptera: Tenebrionidae): Analysis of Instar Variation in Number and Development Time under Different Diets. J. Entomol. Sci. 2010, 45, 75–90. [Google Scholar] [CrossRef]
- van Broekhoven, S.; Oonincx, D.G.A.B.; van Huis, A.; van Loon, J.J.A. Growth Performance and Feed Conversion Efficiency of Three Edible Mealworm Species (Coleoptera: Tenebrionidae) on Diets Composed of Organic by-Products. J. Insect Physiol. 2015, 73, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Morales-Ramos, J.A.; Rojas, M.G.; Kelstrup, H.C.; Emery, V. Self-Selection of Agricultural By-Products and Food Ingredients by Tenebrio Molitor (Coleoptera: Tenebrionidae) and Impact on Food Utilization and Nutrient Intake. Insects 2020, 11, 827. [Google Scholar] [CrossRef] [PubMed]
- Morales-Ramos, J.A.; Rojas, M.G. Effect of Larval Density on Food Utilization Efficiency of Tenebrio Molitor (Coleoptera: Tenebrionidae). J. Econ. Entomol. 2015, 108, 2259–2267. [Google Scholar] [CrossRef]
- Mattioli, S.; Paci, G.; Fratini, F.; Dal Bosco, A.; Tuccinardi, T.; Mancini, S. Former Foodstuff in Mealworm Farming: Effects on Fatty Acids Profile, Lipid Metabolism and Antioxidant Molecules. LWT 2021, 147, 111644. [Google Scholar] [CrossRef]
- Finke, M.D. Complete Nutrient Content of Four Species of Commercially Available Feeder Insects Fed Enhanced Diets during Growth. Zoo Biol. 2015, 34, 554–564. [Google Scholar] [CrossRef] [PubMed]
- Rovai, D.; Ortgies, M.; Amin, S.; Kuwahara, S.; Schwartz, G.; Lesniauskas, R.; Garza, J.; Lammert, A. Utilization of Carrot Pomace to Grow Mealworm Larvae (Tenebrio Molitor). Sustainability 2021, 13, 9341. [Google Scholar] [CrossRef]
- Cobbs, C.; Heath, J.; Stireman, J.O.; Abbot, P. Carotenoids in Unexpected Places: Gall Midges, Lateral Gene Transfer, and Carotenoid Biosynthesis in Animals. Mol. Phylogenet. Evol. 2013, 68, 221–228. [Google Scholar] [CrossRef]
- Mein, J.R.; Dolnikowski, G.G.; Ernst, H.; Russell, R.M.; Wang, X.-D. Enzymatic Formation of Apo-Carotenoids from the Xanthophyll Carotenoids Lutein, Zeaxanthin and β-Cryptoxanthin by Ferret Carotene-9′,10′-Monooxygenase. Arch. Biochem. Biophys. 2011, 506, 109–121. [Google Scholar] [CrossRef]
- Poveda, J. Insect Frass in the Development of Sustainable Agriculture. A Review. Agron. Sustain. Dev. 2021, 41, 5. [Google Scholar] [CrossRef]
- Errico, S.; Spagnoletta, A.; Verardi, A.; Moliterni, S.; Dimatteo, S.; Sangiorgio, P. Tenebrio Molitor as a Source of Interesting Natural Compounds, Their Recovery Processes, Biological Effects, and Safety Aspects. Compr. Rev. Food Sci. Food Saf. 2022, 21, 148–197. [Google Scholar] [CrossRef]
- Ravzanaadii, N.; Kim, S.-H.; Choi, W.-H.; Hong, S.-J.; Kim, N.-J. Nutritional Value of Mealworm, Tenebrio Molitor as Food Source. Int. J. Ind. Entomol. 2012, 25, 93–98. [Google Scholar] [CrossRef]
- Sánchez-Muros, M.-J.; Barroso, F.G.; Manzano-Agugliaro, F. Insect Meal as Renewable Source of Food for Animal Feeding: A Review. J. Clean. Prod. 2014, 65, 16–27. [Google Scholar] [CrossRef]
- Zheng, T.; Li, H.; Han, N.; Wang, S.; Hackney Price, J.; Wang, M.; Zhang, D. Functional Characterization of Two Elongases of Very Long-Chain Fatty Acid from Tenebrio Molitor L. (Coleoptera: Tenebrionidae). Sci. Rep. 2017, 7, 10990. [Google Scholar] [CrossRef] [PubMed]
- Oonincx, D.G.A.B.; Finke, M.D. Nutritional Value of Insects and Ways to Manipulate Their Composition. J. Insects as Food Feed 2021, 7, 639–659. [Google Scholar] [CrossRef]
- Rumpold, B.A.; Schlüter, O.K. Nutritional Composition and Safety Aspects of Edible Insects. Mol. Nutr. Food Res. 2013, 57, 802–823. [Google Scholar] [CrossRef]
- Van Huis, A.; Van Itterbeeck, J.; Klunder, H.; Mertens, E.; Halloran, A.; Muir, G.; Vantomme, P. Edible Insects: Future Prospects for Food and Feed Security. FAO For. Pap. FAO, Rome, Italy. 2013, 171. Available online: http//www.fao.Org/3/i3253e/i3253e.pdf.
- Bjørge, J.D.; Overgaard, J.; Malte, H.; Gianotten, N.; Heckmann, L.-H. Role of Temperature on Growth and Metabolic Rate in the Tenebrionid Beetles Alphitobius Diaperinus and Tenebrio Molitor. J. Insect Physiol. 2018, 107, 89–96. [Google Scholar] [CrossRef]
- Hoc, B.; Genva, M.; Fauconnier, M.-L.; Lognay, G.; Francis, F.; Caparros Megido, R. About Lipid Metabolism in Hermetia Illucens (L. 1758): On the Origin of Fatty Acids in Prepupae. Sci. Rep. 2020, 10, 11916. [Google Scholar] [CrossRef]
- van Dooremalen, C.; Ellers, J. A Moderate Change in Temperature Induces Changes in Fatty Acid Composition of Storage and Membrane Lipids in a Soil Arthropod. J. Insect Physiol. 2010, 56, 178–184. [Google Scholar] [CrossRef]
- Arrese, E.L.; Soulages, J.L. Insect Fat Body: Energy, Metabolism, and Regulation. Annu. Rev. Entomol. 2010, 55, 207–225. [Google Scholar] [CrossRef]
- GEORGESCU, B.; STRUȚI, D.; PĂPUC, T.; CIGHI, V.; BOARU, A. Effect of the Energy Content of Diets on the Development and Quality of the Fat Reserves of Larvae and Reproduction of Adults of the Black Soldier Fly, Hermetia Illucens (Diptera: Stratiomyidae). Eur. J. Entomol. 2021, 118, 297–306. [Google Scholar] [CrossRef]
- Jiang, H.; Wang, Z.; Ma, Y.; Qu, Y.; Lu, X.; Guo, H.; Luo, H. Effect of Dietary Lycopene Supplementation on Growth Performance, Meat Quality, Fatty Acid Profile and Meat Lipid Oxidation in Lambs in Summer Conditions. Small Rumin. Res. 2015, 131, 99–106. [Google Scholar] [CrossRef]
- Hernández-López, S.H.; Rodríguez-Carpena, J.G.; Lemus-Flores, C.; Grageola-Nuñez, F.; Estévez, M. Avocado Waste for Finishing Pigs: Impact on Muscle Composition and Oxidative Stability during Chilled Storage. Meat Sci. 2016, 116, 186–192. [Google Scholar] [CrossRef] [PubMed]
- Biondi, L.; Luciano, G.; Cutello, D.; Natalello, A.; Mattioli, S.; Priolo, A.; Lanza, M.; Morbidini, L.; Gallo, A.; Valenti, B. Meat Quality from Pigs Fed Tomato Processing Waste. Meat Sci. 2020, 159, 107940. [Google Scholar] [CrossRef] [PubMed]
- Costa, S.; Pedro, S.; Lourenço, H.; Batista, I.; Teixeira, B.; Bandarra, N.M.; Murta, D.; Nunes, R.; Pires, C. Evaluation of Tenebrio Molitor Larvae as an Alternative Food Source. NFS J. 2020, 21, 57–64. [Google Scholar] [CrossRef]
- Mercola, J.; D’Adamo, C.R. Linoleic Acid: A Narrative Review of the Effects of Increased Intake in the Standard American Diet and Associations with Chronic Disease. Nutrients 2023, 15, 3129. [Google Scholar] [CrossRef] [PubMed]
- Fasel, N.J.; Mène-Saffrané, L.; Ruczyński, I.; Komar, E.; Christe, P. Diet Induced Modifications of Fatty-Acid Composition in Mealworm Larvae (Tenebrio Molitor). J. Food Res. 2017, 6, 22–31. [Google Scholar] [CrossRef]
- Rossi, G.; Mattioli, S.; Rondoni, G.; Bosco, A.D.; Servili, M.; Castellini, C.; Conti, E. Characterisation of Fatty Acid Profiles of Tenebrio Molitor Larvae Reared on Diets Enriched with Edible Oils. J. Insects as Food Feed 2022, 8, 901–912. [Google Scholar] [CrossRef]
- Brandstetter, B.; Ruther, J. An Insect with a Delta-12 Desaturase, the Jewel Wasp Nasonia Vitripennis, Benefits from Nutritional Supply with Linoleic Acid. Sci. Nat. 2016, 103, 40. [Google Scholar] [CrossRef] [PubMed]
- Amoedo, N.D.; Punzi, G.; Obre, E.; Lacombe, D.; De Grassi, A.; Pierri, C.L.; Rossignol, R. AGC1/2, the Mitochondrial Aspartate-Glutamate Carriers. Biochim. Biophys. Acta - Mol. Cell Res. 2016, 1863, 2394–2412. [Google Scholar] [CrossRef] [PubMed]
- Romero-Lorente, M.-Á.; Fabrikov, D.; Montes, J.; Morote, E.; Barroso, F.G.; Vargas-García, M. del C.; Varga, Á.T.; Sánchez-Muros, M.-J. Pre-Treatment of Fish By-Products to Optimize Feeding of Tenebrio Molitor L. Larvae. Insects 2022, 13, 125. [Google Scholar] [CrossRef]
- Lawal, K.G.; Kavle, R.R.; Akanbi, T.O.; Mirosa, M.; Agyei, D. Enrichment in Specific Fatty Acids Profile of Tenebrio Molitor and Hermetia Illucens Larvae through Feeding. Futur. Foods 2021, 3, 100016. [Google Scholar] [CrossRef]
- Otero, P.; Gutierrez-Docio, A.; Navarro del Hierro, J.; Reglero, G.; Martin, D. Extracts from the Edible Insects Acheta Domesticus and Tenebrio Molitor with Improved Fatty Acid Profile Due to Ultrasound Assisted or Pressurized Liquid Extraction. Food Chem. 2020, 314, 126200. [Google Scholar] [CrossRef] [PubMed]
- Jeon, Y.-H.; Son, Y.-J.; Kim, S.-H.; Yun, E.-Y.; Kang, H.-J.; Hwang, I.-K. Physicochemical Properties and Oxidative Stabilities of Mealworm (Tenebrio Molitor) Oils under Different Roasting Conditions. Food Sci. Biotechnol. 2016, 25, 105–110. [Google Scholar] [CrossRef] [PubMed]
- Oppedisano, F.; Mollace, R.; Tavernese, A.; Gliozzi, M.; Musolino, V.; Macrì, R.; Carresi, C.; Maiuolo, J.; Serra, M.; Cardamone, A.; et al. PUFA Supplementation and Heart Failure: Effects on Fibrosis and Cardiac Remodeling. Nutrients 2021, 13, 2965. [Google Scholar] [CrossRef] [PubMed]
- Fedor, D.; Kelley, D.S. Prevention of Insulin Resistance by N-3 Polyunsaturated Fatty Acids. Curr. Opin. Clin. Nutr. Metab. Care 2009, 12, 138–146. [Google Scholar] [CrossRef] [PubMed]
- Oppedisano, F.; Bulotta, R.M.; Maiuolo, J.; Gliozzi, M.; Musolino, V.; Carresi, C.; Ilari, S.; Serra, M.; Muscoli, C.; Gratteri, S.; et al. The Role of Nutraceuticals in Osteoarthritis Prevention and Treatment: Focus on n-3 PUFAs. Oxid. Med. Cell. Longev. 2021, 2021, 1–12. [Google Scholar] [CrossRef]
- Hashimoto, K. Role of Soluble Epoxide Hydrolase in Metabolism of PUFAs in Psychiatric and Neurological Disorders. Front. Pharmacol. 2019, 10. [Google Scholar] [CrossRef]
- Paiva, L.; Lima, E.; Neto, A.I.; Marcone, M.; Baptista, J. Health-Promoting Ingredients from Four Selected Azorean Macroalgae. Food Res. Int. 2016, 89, 432–438. [Google Scholar] [CrossRef]
- Son, Y.-J.; Choi, S.Y.; Hwang, I.-K.; Nho, C.W.; Kim, S.H. Could Defatted Mealworm (Tenebrio Molitor) and Mealworm Oil Be Used as Food Ingredients? Foods 2020, 9, 40. [Google Scholar] [CrossRef]
Figure 1.
Fat and fatty acid content in mealworms fed on different diets.Values are are expressed as means ± standard deviation of three independent analyses. (n = 3).
Figure 1.
Fat and fatty acid content in mealworms fed on different diets.Values are are expressed as means ± standard deviation of three independent analyses. (n = 3).
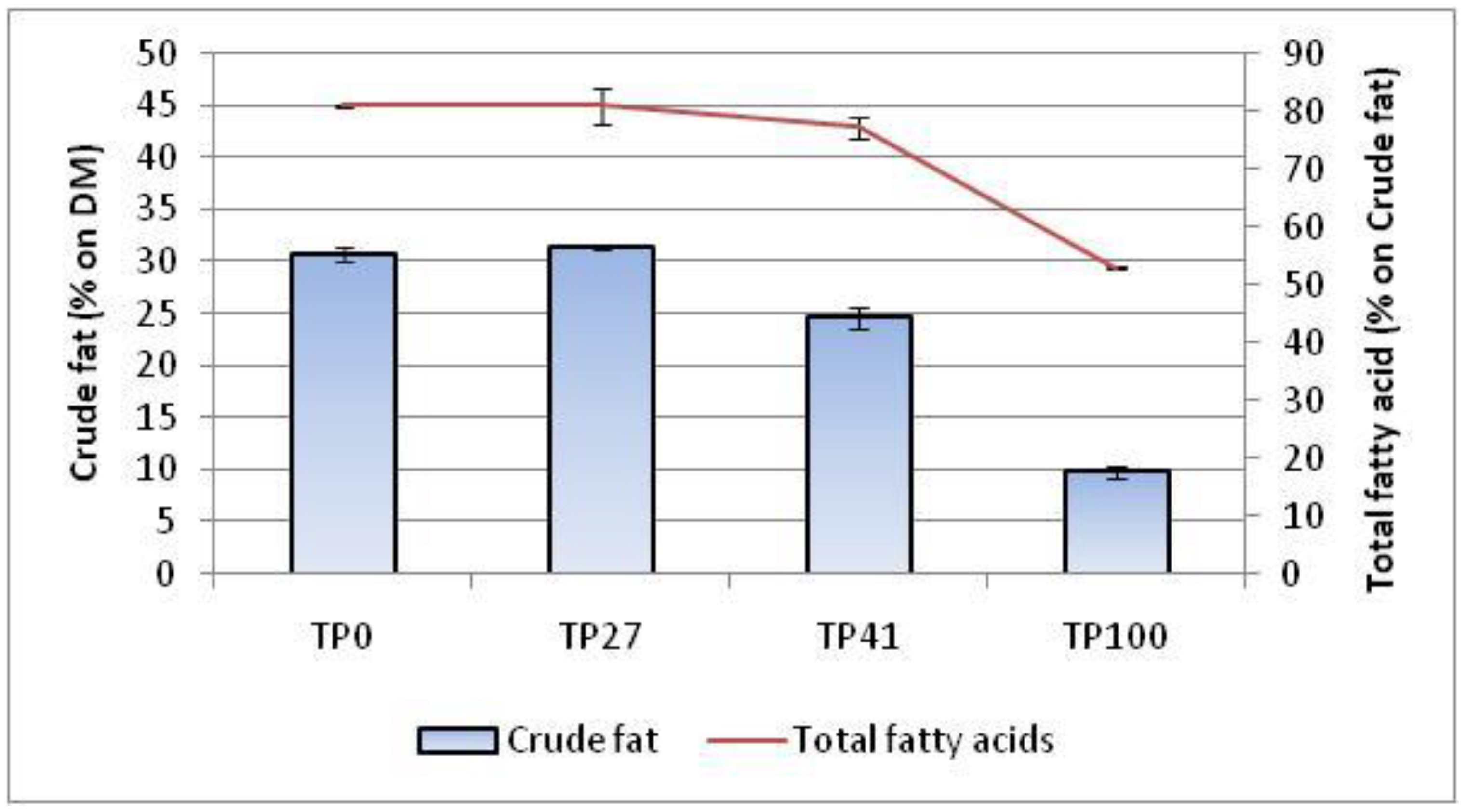
Figure 2.
Crude protein quantification in defatted larvae fed on different diets. Values are reported as means ± standard deviation of three independent analyses. (n = 3).
Figure 2.
Crude protein quantification in defatted larvae fed on different diets. Values are reported as means ± standard deviation of three independent analyses. (n = 3).
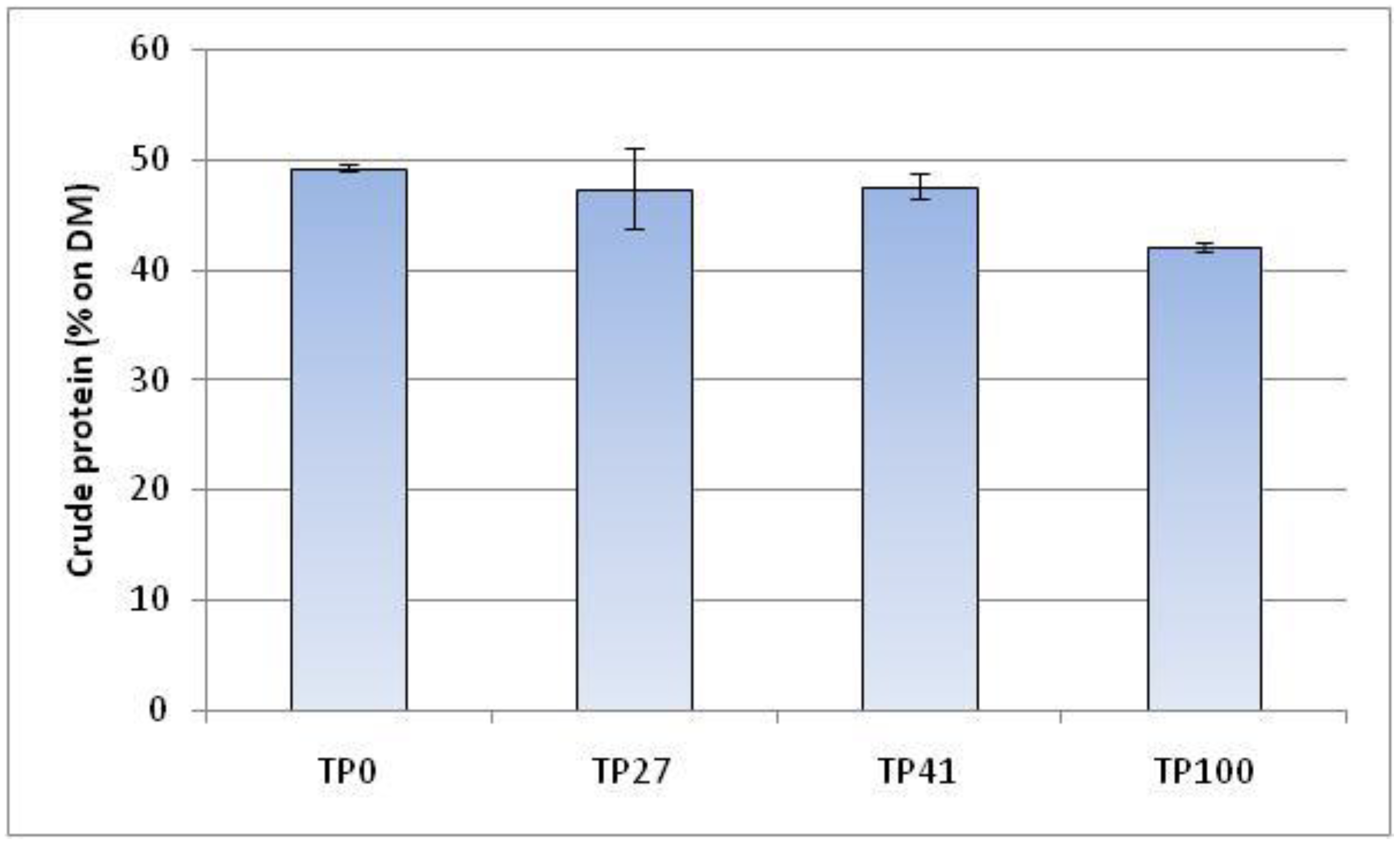

Table 1.
Nutrient composition of by-products preliminarily conditioned (%DM).
| By-product | Dry Matter | Crude protein | Crude Fat | Crude fiber | Ash | Carbohydrate |
|---|---|---|---|---|---|---|
| Bran | 91.2 | 16.7 | 6.5 | 36.1 | 4.2 | 30.2 |
| Tomato pomace | 92.1 | 9.5 | 3.2 | 67.1 | 3.9 | 8.9 |
| Brewer’s spent grain | 93.4 | 24.7 | 4.8 | 42 | 2.6 | 24 |
| Yeast | 93.0 | 47.6 | 2.4 | 6.8 | 8.0 | 13.8 |
Table 2.
Diets composition and nutritional values* (% DM).
| Diet | Bran (%) |
Tomato pomace (%) |
Brewer’s spent grain (%) |
Yeast (%) |
Protein value (%) |
Carbohydrate (%) |
P:C | Crude fiber (%) |
Fat (%) | Energy (kcal (100 g) |
|---|---|---|---|---|---|---|---|---|---|---|
| TP0 | 100 | - | - | - | 16.7 | 30.2 | 1:1.8 | 36.1 | 6.5 | 318.3 |
| TP27 | 50 | 27 | 23 | - | 16.6 | 23.0 | 1:1.4 | 45.8 | 5.21 | 296.9 |
| TP41 | 50 | 41 | - | 9 | 16.5 | 20.0 | 1:1.2 | 46.2 | 4.78 | 281.4 |
| TP100 | - | 100 | - | - | 9.5 | 8.9 | 1:0.9 | 67.1 | 3.2 | 236.6 |
* calculated values
Table 3.
Results of larval performances between diets.
| Diet | Survival (%) |
Growth time (d) | Larval Weight (mg) | Pupal Weight (mg) |
|---|---|---|---|---|
| TP0 | 99.5±1.6 | 32.0±5.5 a | 100.0±14.1 | 117.5±14.0 ab |
| TP27 | 100.0±0.0 | 38.5±3.9 a | 104.0±9.7 | 114.0±13.5 ab |
| TP41 | 99.5±1.6 | 37.7±5.0 a | 109.0±11.0 | 127.0±20.2 b |
| TP100 | 99.5±1.6 | 63.4±18.5 b | 91.0±17.9 | 101.0±9.9 a |
Mean ± standard deviation values with the same letter, within columns, are not significantly different (Kruskal–Wallis test and Pairwise multiple comparisons with Bonferroni correction), at α = 0.05.
Table 4.
Efficiency indicators of diets tested.
| Diet | FCR | SRG (% day-1) |
ECI (%) |
ECD (%) |
|---|---|---|---|---|
| TP0 | 2.7±0.2 a | 4.9±0.7 a | 15.4±1.1 a | 34. 6±3.3 a |
| TP27 | 3.2±0.1 b | 4.1±0.4 b | 13.1±0.5 b | 42.8±4.5 b |
| TP41 | 3.8±0.3 c | 4.3±0.4 b | 10.8±0.7 c | 30.0±6.3 a |
| TP100 | 4.3±0.4 d | 2.5±0.7 c | 9.8±1.0 d | 65.9±12.7 c |
Mean±standard deviation values with the same letter, within coloums, are not significantly different (Tukey-Kramer HDS test for FCR, SRG and ECI; Pairwise multiple comparisons with Bonferroni correctionfor ECD), at α = 0.05.
Table 5.
Licopene and β-Carotene Quantification1.
| Diet | Feed | Mealworm | Frass | |||
|---|---|---|---|---|---|---|
| Lycopene (ug/g) |
β-Carotene (ug/g) | Lycopene (ug/g) |
β-Carotene (ug/g) | Lycopene (ug/g) |
β-Carotene (ug/g) | |
| TP0 | 2.66±0.24 | 0.30±0.07 | 0.08±0.06 | 1.43±0.99 | 0.70±0.04 | 12.09±0.26 |
| TP27 | 22.68±0.79 | 45.30±1.95 | 0.08±0.01 | 1.11±0.16 | 12.44±0.35 | 50.99±0.57 |
| TP41 | 52.43±1.71 | 95.09±0.70 | 0.61±0.33 | 2.56±0.76 | 24.10±0.32 | 76.32±1.24 |
| TP100 | 179.75±2.74 | 241.47±2.53 | 1.19±0.27 | 7.28±0.06 | 39.67±1.56 | 147.46±4.61 |
1 Values are reported on dry mass and are expressed as means ± standard deviation of three independent analyses. (n = 3).
Table 6.
Fatty acid profile of the lipid extract of MLW powder fed different diets (% TFA)1.
| Fatty Acid (%) | Diets | |||||
|---|---|---|---|---|---|---|
| Common Name | Lipid number | TP0 | TP27 | TP41 | TP100 | |
| Caprilic acid | C8:0 | 0 | 0 | 0 | 0 | |
| Capric acid | C10:0 | 0 | 0 | 0 | 0 | |
| Lauric acid | C12:0 | 0 | 0 | 0 | 0 | |
| Myristic acid | C14:0 | 3.68±0.04 | 3.81±0.03 | 3.42±0.12 | 2.66±0.06 | |
| Palmitic acid | C16:0 | 15.60±0.04 | 14.8±0.09 | 15.28±1.13 | 13.80±0.09 | |
| Palmitoleic acid | C16:1 | 1.56±0.27 | 4.19±0.03 | 1.44±0.29 | 1.00±0.01 | |
| Stearic acid | C18:0 | 2.89±0.20 | 3.16±0.08 | 3.38±0.17 | 5.59±0.05 | |
| Oleic acid | C18:1 | 50.20±0.20 | 44.7±0.28 | 42.87±0.229 | 26.20±1.46 | |
| α-Linoleic acid | C18:2n-6 | 25.70±0.31 | 28.9±0.32 | 32.61±1.24 | 48.10±1.27 | |
| α-Linolenic acid | C18:3n3 | 0.42±0.04 | 0.51±0.09 | 0.99±0.11 | 2.68±0.08 | |
| Arachidic acid | C20:0 | 0 | 0 | 0 | 0 | |
| Behenic acid | C22:0 | 0 | 0 | 0 | 0 | |
| Erucic acid | C22:1 | 0 | 0 | 0 | 0 | |
| Lignoceric acid | C24:0 | 0 | 0 | 0 | 0 | |
| Σ SFA | 22.14±0.07 | 21.76±0.19 | 18.71±1.25 | 22.02±0.19 | ||
| Σ MUFA | 51.75±0.19 | 48.84±0.32 | 44.32±0.52 | 27.22±1.46 | ||
| Σ PUFA | 26.10±0.19 | 29.40±0.40 | 33.60±1.35 | 50.8±1.4 | ||
| Σ UFA | 77,9±0,08 | 78,2±0,27 | 77,9±0,39 | 78±0,20 | ||
| PUFA : SFA ratio | 1.18 | 1.35 | 1.79 | 2.30 | ||
| MUFA:PUFA ratio | 1.98 | 1.66 | 1.32 | 0.53 | ||
| ω6 : ω3 ratio | 61.47 | 56.13 | 32.99 | 17.95 | ||
1 Values are reported on % Total Fatty Acid (TFA) and are expressed as means ± standard deviation of three independent analyses. (n = 3).
Table 7.
Lipid quality indices of TFA obtained from MLW fed different diets1.
| Index | Diets | |||
|---|---|---|---|---|
| TP0 | TP27 | TP41 | TP100 | |
| COX | 3.2±0.43 | 3.53±0.18 | 4.01±0.3 | 5.79±0.14 |
| IT | 0.55±0.05 | 0.54±0.03 | 0.53±0.05 | 0.48±0.01 |
| IA | 0.39±0.06 | 0.38±0.01 | 0.37±0.02 | 0.31±0.02 |
| HH | 3.96±0.21 | 3.98±0.55 | 4.09±0.31 | 4.68±0.24 |
| UI | 104.4±0.78 | 108.1±0.92 | 112.5±0.15 | 131.4±0.91 |
1 Values are expressed as means ± standard deviation of three independent analyses. (n = 3).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



