
Submitted:
25 September 2023
Posted:
26 September 2023
You are already at the latest version
Abstract
Water deficit (drought) is still one of the most important stress factors affecting not only yields, but also production quality. Breeders are focusing on breeding cultivars and crop lines that are more resistant to water shortages. In addition to breeding, for example, for the size of the roots and changes in the color of the leaves, focus on changing the morphotype i.e. long chaff, multirowed ear. It is predicted that wheat genotypes with a morphotype change should be more drought tolerant. Therefore, our research focused on the study of changes in the physiological parameters of wheat (cv. ´Bohemia´) with a changed morphotype (genotypes ´284-17´ [long chaff] and genotype ´29-17´ [multirowed ear]) in relation to exposure to drought. Pigment content, photosynthesis and transpiration rate, water potential were measured in plants irrigated and affected by water deficit. Water deficit was shown to have a statistically significant effect on transpiration rate, photosynthetic rate, water potential and total chlorophyll content. Furthermore, the influence of the genotype on the observed parameters and on the effect of water deficit was demonstrated. Due to the fact that cv. ´Bohemia´, increases during such periods of transpiration, so for this reason it is not suitable for dry areas. Genotype ́29-17 ́ appears promising with respect to drought resistance.
Keywords:
common wheat
; Triticum aestivum L.
; water deficit
; drought
; gas exchange
; fluorescence
; water regime
1. Introduction
Drought, which is caused by low water availability [1], is one of the most important factors limiting agricultural production, severely affecting crop yield [2,3]. As a result, it also affects food production and supply [3]. According to Sekhon et al. [4], drought is secondarily affected by higher air temperature and CO2 concentration, more intense solar radiation, low relative humidity and air flow. In general, drought stress reduces osmotic pressure and disrupts water potential in plant cells, causing oxidative stress and cell damage, reducing carbon fixation and primary metabolism [5,6,7].
These negative impacts result in reduced crop survival and yield. In response to drought stress, plants activate their drought response mechanisms such as morphological and structural changes [8], expression of drought tolerance genes, synthesis of hormones, and osmotic regulators to alleviate drought stress, alteration in primary [9] and secondary metabolism [10], and imbalance in nutrient uptake by maize [11].
The structural changes that are associated with a decrease in water potential include a reduction in leaf area, which is associated with lower availability of assimilates [12,13]. In wheat, according to Sewore et al. [14], there is significant flag leaf rolling due to drought. A similar phenomenon is observed phenomenon is observed in maize. The mentioned changes in metabolism are related to a decrease in water potential inside cells and tissues [15], which in turn results in stomatal closure.
According to Liu et al. [15] and Onyemaobi et al. [16], a decrease in turgor not only leads to stomatal closure, a decrease in photosynthesis, for example, in wheat [17] and tomato [18], and changes in enzyme activity, especially RUBISCO [19], but also to an increase in membrane permeability, denaturation and inactivation of proteins, metabolic disturbances, damage to the photosynthetic apparatus [20], and nutrient deficiency.
In barley leaves, drought reduces the density of stomata in leaves, which could be one of the signs of resistance to this stressor according to [21,22]. A similar phenomenon was also observed in wheat [23]. Furthermore, drought exposure leads to photoinhibition, where the actual efficiency of photosystem II (ΦPSII) is reduced at the same time [24] due to the reduction of RUBISCO activity. Furthermore, the chlorophyll content decreases and the ratio of chlorophyll a, b and carotenoids changes [25], which subsequently causes changes in photosynthetic function [26,27]. On the other hand, according to Yang et al. [9], a cascade of defense reactions is triggered, which includes the formation of specific proteins such as LEA, dehydrins, AQP, OSM proteins, etc.
Common wheat (Triticum aestivum L.) is the world's most widely grown cereal and is therefore also most affected by drought stress. One way to mitigate the effects of climate change is to find and select suitable stress-tolerant genotypes. Breeding for drought tolerance therefore requires the integration of different knowledge systems and methodologies from different disciplines of plant sciences [28,29].
One way of breeding is to change the morphotype of plants, especially the ear. This is a group of genes controlling the occurrence of so-called supernumerary spikelets, where, unlike in common wheat, more than one spikelet can grow from a single node of the ear spindle [30,31,32]. Multi-rowed ear in wheat is caused by the recessive gene WFZP-D (Wheat Frizzy Panicle) on the short arm of chromosome 2D [33,34]. Another trait is the long chaff, which occurs in two tetraploid species, Triticum polonicum L. (gene P1 on chromosome 7A) and T. ispahanicum Hesolt. (gene P2 on chromosome 7B), as well as in the hexaploid T. petropavlovskyi Udac. & Migusch [35,36].
It is hypothesized that wheat genotypes with morphotype change should be more drought tolerant. Therefore, our research focused on the study of changes in physiological parameters of wheat with altered morphotype in relation to drought exposure. The proposed hypotheses were whether there are genotypic differences to drought exposure, are plant physiological responses influenced by genotype, and does drought affect the physiological responses of juvenile wheat plants. The objective of this study was to determine genotypic differences in plant response to drought.
2. Materials and Methods
2.1. Plant Material and Experimental Conditions
Winter wheat plants were grown in 11 x 11 cm pots in a greenhouse experiment. The air temperature during the experiment was maintained at 25 °C during the day and 19 °C at night, with a natural light regime of 14 hours of light and 10 hours of darkness. The wheat plants were irrigated to a level of 70% of the substrate moisture by volume (150 ml of water per container). A 2:1 mixture of high organic matter garden substrate and silica sand was used as growing medium. The garden substrate was fine (with no more than 10% of particles above 10 mm), kept the environment airy, unsalted, with a pH of the aqueous solution between 5.5 and 6.5, and contained no weeds or pests. Furthermore, it contained 55% of combustible matter in the dried sample and a maximum of 5% of particles above 25 mm.
The experimental design included four variants (Table 1). The control (C) was irrigated regularly. The other three treatments were stressed by water deficit induced by gradual drying of the substrate: namely, (D1) 10 d no irrigated, 4 d watered, 10 d water deficit re-induced, 4 d watered; (D2) 10 d watered and then stress induced until the end of the experiment; (D3) 10 d stressed and then irrigated.
The effect of short-term water deficit was monitored in breeding material and near-isogenic lines of seeded wheat: (i)´Bohemia´; (ii) genotype ´284-17´ (long chaff); (iii) genotype ´29-17´ (multirowed ear). Seed was obtained from Agrotest Phyto, Inc. Kroměříž, Czech Republic. The experiment was established at the stage of plant development 16.BBCH and lasted 28 days, when the physiological parameters of the plants were measured: 0, 5, 10, 14,18, 22 and 28 days from the start of the experiment.
2.2. Pigments Contents
Pigment content was determined according to the methodology [37]. From wheat leaves, the following were extracted at 1 cm2 targets were taken. Targets were placed in plastic vials and the 1 ml of dimethylformamide (DMF; Merck KGaA, Darmstadt, Germany) was added. Within 24 hours, the pigments were extracted in cold, darkness under nonstop shaking. Twenty-four hours after collection, the samples were spectrophotometrically were analysed using a UV-Vis Evolution 2000 instrument (Thermo Fisher Scientific Inc., Waltham, MA, USA). As blank, pure dimethylformamide was used.
2.3. Gas Exchange Parameters
Gas exchange rate was measured by a non-destructive method using the Integrated fluorometer and gas exchange system – iFL (ADC Bioscientific Ltd., Hoddesdon, UK). Net photosynthetic rate (Pn) was measured on a photosynthetically mature leaf, in the central part of the leaf blade. Gas exchange rate and stomatal conductance (gs) were derived from Pn [38]. Gas exchange rate was measured in the morning (8-13 h CET) at an irradiance density of 650 μM m-2 s-1 and temperature of 25°C, according to the methodology of Kuklova et al. [38].
2.4. Parameters of Fluorescence
Chlorophyll fluorescence parameters were analysed in juvenile wheat plants. These were the maximum quantum yield of photosystem II (Fv/Fm), the ratio of maximum fluorescence to initial fluorescence (Fm/Fo), and the ratio of variable fluorescence to initial fluorescence (Fv/Fo). Integrated fluorometer and gas exchange system – iFL (ADC Bioscientific Ltd., Hoddesdon, UK) was used to measure fluorescence parameters. Chlorophyll fluorescence parameters were measured: Fo, Fm – minimum and maximum dark-adjusted fluorescence yield; Fv – variable fluorescence (Fv = Fm/Fo). These parameters were used to calculate Fv/Fo (potential photochemical efficiency) and Fv/Fm (maximum quantum efficiency of PSII) ratios, which are considered as indicators for the efficiency of PSII in primary photochemical reactions. The fluorescence parameters were measured on five selected fully expanded upper leaves. Leaves were acclimated to darkness for 30 min before measurements were made. The measurement time was 5 sec and the irradiance was 3000  M m-2 s-1.
M m-2 s-1.
M m-2 s-1.2.5. Water potential
Water potential (ψw) as the energy status of the water in the system was determined using the dewpoint with a water potential meter WP4C (Decagon Devices Inc., Pullman, WA, USA). Leaves were packed in plastic syringes and airtight sealed with parafilm. Then, samples were frozen at -18°C. After the thawing of samples at room temperature, drop of liquid was extracted from the syringe and used for measurements [39].
2.6. Statistical Analysis
The experiments used four independent biological replicates. Each replicate was a sample of plant material derived from a different pot. The variability of the differences in the parameters of interest for all treatments was tested using multivariate ANOVA (p < 0.05) and Tukey's post hoc test for significant differences between treatments. Data were analysed using Statistica 13.5 software (StatSoft, Tulsa, OK, USA). To test the dependence of physiological scores on trial variation and genotype, a linear regression was constructed using artificial variables at a significance level of α = 0.05. ´Bohemia´ control was chosen as the baseline group. The R software (v 4.0.5, The R Foundation, Vienna, Austria) was used to calculate the model.
3. Results
3.1. Pigments Contents
Changes in the content of photosynthetically active pigments were studied in juvenile wheat plants as a function of genotype and water deficit. The results obtained are presented in Table 2. It shows that the effect of genotype on the content of pigments in leaves was demonstrated, with the lowest content of total chlorophylls and carotenoids in cv. ´Bohemia´ (6.893 nM cm-2 and 1.246 nM cm-2, respectively). On the other hand, genotype ´284-17´ had the highest pigment content. It had a total chlorophyll content of 14.945 nM cm-2 and carotenoid content of 2.486 nM cm-2.
Further, the effect of variant on chlorophyll content was confirmed as the stressed variants showed lower chlorophyll values compared to the control. The highest reduction in chlorophyll content due to water deficit was found in variant D2, where the content was 1.685 nM cm-2 (14.30%) lower compared to the control. The least reduction in chlorophyll content was observed in variant D3, where the chlorophyll content was inconclusively lower by 0.313 nM cm-2 (2.66%) compared to the control. In the case of carotenoids, their content was conclusively the highest in variant D1 (2.013 nM cm-2) and the lowest in D3 (1.784 nM cm-2). No significant differences in carotenoid content were found between D3, D2 and C.
The highest reduction in chlorophyll content due to water deficit was found in genotype ´284-17´, with a reduction of 2.901 nM cm-2 or 17.46% in variant D2. Similarly, the chlorophyll content of this variant was reduced in the ´Bohemia´ and genotype ´29-17´, with a reduction of 0.736 nM or 10.15% in the case of the ´Bohemia´ and 1.236 nM cm-2 or 10.51% in the case of genotype ´29-17´. The lowest reduction in total chlorophyll content was found in variant D3, where the pigment content decreased in the following order depending on the influence of genotype: genotype ´284-17´ (1.007 nM cm-2; 6.06 %) > genotype ´29-17´ (0.412 nM cm-2; 3.50 %) > ´Bohemia´ (0.013 nM cm-2; 0.18 %).
The carotenoid content was clearly influenced by variant and genotype, see Table 2. In the ´Bohemia´, the carotenoid content increased in the stressed D2 variant, where the carotenoid content was 0.249 nM cm-2 or 20.61% higher compared to the control. On the other hand, their content decreased by 0.116 nM cm-2 (9.60 %) in the D1 variant. No significant difference was found between control and D3 variant plants. On the other hand, in genotype ´29-17´, the carotenoid content decreased due to drought, with the most significant decrease observed in variant D2, which showed a decrease of 0.317 nM cm-2 (12.03%). On the other hand, the lowest decrease was found in variant D3 (0.297 nM cm-2; 2.66%). On the other hand, an inconclusive increase of 1.10% (0.029 nM cm-2) was recorded for variant D1. A similar trend was also observed in genotype ´29-17´, with a statistically significant decrease in variant D2 (0.191 nM cm-2; 0.60%) and an inconclusive decrease in variant D3 (0.048 nM cm-2; 2.66%). A significant increase in carotenoid content was determined in variant D1, with an increase of 0.551 nM cm-2 or 30.58%.
3.2. Gas Exchange Parameters
Stomatal conductance (gs) was affected not only by the experimental variant but also by the genotype used. The highest average stomatal conductance was observed in plants of genotype ´29-17´ (0.102 M m-2 s-1) and the lowest in the control (0.073 M m-2 s-1), while no significant differences were found between genotype´284-17´ and cv. ´Bohemia´, see Table 3. The highest stomatal conductance within the variants was exhibited by plants growing on variant D2, which had a gs value of 0.090 M m-2 s-1. Plants from variants C and D1 showed similar values. In contrast, plants from variant D3 had statistically significant lowest gs values, 26.67% lower than those of variant D2.
Stomatal conductance (gs) was affected not only by the experimental variant but also by the genotype used. The highest average stomatal conductance was observed in plants of genotype ´29-17´ (0.102 M m-2 s-1) and the lowest in the control (0.073 M m-2 s-1), while no significant differences were found between genotype ´284-17´ and cv. ´Bohemia´, see Table 3. The highest stomatal conductance within the variants was exhibited by plants growing on variant D2, which had a gs value of 0.090 M m-2 s-1. Plants from variants C and D1 showed similar values. In contrast, plants from variant D3 had statistically significant lowest gs values, 26.67% lower than those of variant D2.
The effect of differential response to water deficit within the genotypes studied was demonstrated, with genotype ´29-17´, conclusively reducing stomatal conductance by 37.26% compared to the control (0.133 M m-2 s-1). A significant decrease in gs would be observed in this genotype for all stressed variants. On the other hand, the genotype ´284-17´ increased the gs value by 37.29 % compared to the control (0.069 M m-2 s-1) and a similar trend was observed for the ´Bohemia´ (0.092 M m-2 s-1, increase by 29.52 %). On the other hand, the lowest reduction in genotype ´284-17´ was in the D3 variant (20.76 %) and in the ´Bohemia´ in the D1 variant (6.00 %).
Rate of photosynthesis (Pn) was significantly affected at the α=0.05 significance level by the variant experiment and genotype, as documented in Figure 1A, C, E. They show that the lowest Pn during the experiment was exhibited by genotype ´284-17´, which had Pn of 7.842 M m-2 s-1. On the other hand, the highest rate of photosynthesis was recorded by the control plants of genotype ´29-17´ (12.213 M m-2 s-1). Photosynthesis decreased as a function of the duration of exposure drought. After rehydration, photosynthesis increased compared to the water deficit, but did not reach the values of the control plants. The highest decrease in photosynthesis due to water deficit was found in the ´Bohemia´, where rate of photosynthetic decreased by 3.243 M m-2 s-1 or 26.21% and 2.825 M m-2 s-1 or 22.83%, respectively. Similarly, the Pn decreased in these variants in the case of the genotype ´284-17´. On the other hand, in case of genotype ´29-17´, the reduction in photosynthesis rate was conclusively the lowest. In the case of variant D2 photosynthetic rate decreased by 1.060 M m-2 s-1 (3.9%) and 1.126 M m-2 s-1 (8.74%) for variant D1.
M m-2 s-1. On the other hand, the highest rate of photosynthesis was recorded by the control plants of genotype ´29-17´ (12.213 M m-2 s-1). Photosynthesis decreased as a function of the duration of exposure drought. After rehydration, photosynthesis increased compared to the water deficit, but did not reach the values of the control plants. The highest decrease in photosynthesis due to water deficit was found in the ´Bohemia´, where rate of photosynthetic decreased by 3.243 M m-2 s-1 or 26.21% and 2.825 M m-2 s-1 or 22.83%, respectively. Similarly, the Pn decreased in these variants in the case of the genotype ´284-17´. On the other hand, in case of genotype ´29-17´, the reduction in photosynthesis rate was conclusively the lowest. In the case of variant D2 photosynthetic rate decreased by 1.060 M m-2 s-1 (3.9%) and 1.126 M m-2 s-1 (8.74%) for variant D1.The following were found statistically significant differences of transpiration rates (E) between the treatments wheat genotypes in plants growing under control conditions, where the lowest rate of transpiration was in genotype ´Bohemia´ (1.656 mM m-2 s-1) and the highest by genotype ´284-17´ (2.097 mM m-2 s-1), see Figure 1B, D, F. Under drought stress, the transpiration decreased depending on the duration of drought stress and genotype.
The highest decrease in rate of transpiration was found for the genotype ´284-17´, variant D3 a decrease of 27.17% (1.308 mM m-2 s-1) and for variant D1 a decrease in transpiration of 0.358 mM m-2 s-1 or 26.93% was observed. In genotype ´29-17´, the drought caused a decrease in rate of transpiration for variant D1 by 0.542 mM m-2 s-1 or 25.86 %. On the other hand, the lowest decrease in transpiration rate was found in genotype ´29-17´, variant D1 (0.554 mM m-2 s-1; 19.95 D3 (decrease by 0.139 mM m-2 s-1; 6.64%) increased for variants D2 and D3, with increases of 0.137 mM m-2 s-1 or 8.24% and 0.243 mM m-2 s-1 or 14,66 %, respectively.
3.3. Parameters of Fluorescence
The results presented in Figure 2A, C, E demonstrate that water deficit affects fluorescence parameters (Fv/Fm) depending on the experimental variant. The results show that the Fv/Fm values obtained were lower than the generally reported value of about 0.820 for all control plants of the genotypes studied. However, this reduction was not statistically significant. In stressed plants, a non-significant decrease of 0.001 in the Fv/Fm ratio value was observed compared to the control.
There were also no statistically significant differences in the value of the Fv/Fm ratio between genotypes. Nevertheless, it can be stated that the lower fluorescence value was observed in the ´Bohemia´ (0.797) and the higher in genotype ´29-17´ (0.800). The control plants of cv. ´Bohemia´ and genotype ´284-17´ had the lowest fluorescence at the beginning of the experiment (0.761, 0.774 and 0.803) and the highest fluorescence at the end of the experiment (0.816). On the other hand, for genotype ´29-17´, the lowest value of Fv/Fm ratio was detected at the end of the experiment (0.781) and on the contrary, the highest value was detected on the 18th day of the experiment (0.811). A similar trend was also found for the stressed variants of all genotypes studied, with variant D2 showing the lowest Fv/Fm ratio values compared to the other stressed variants.
Since no conclusive differences between genotypes and variants were found, another fluorescence parameter was used, which is the Fv/F0 ratio. The values obtained for this ratio are shown in Figure 2B, D, F. Significant differences were found between the genotypes studied, with the lowest value of the Fv/F0 parameter being found in the cv. ´Bohemia´ (3.963) and the highest in genotype ´29-17´ (4.083). A decrease in Fv/F0 values was observed in plants growing under water deficit conditions compared to control plants.
The exception is the plants from variant D3, which had a higher ratio compared to the control by 0.006. Plants from variant D2 had the lowest Fv/F0 ratio, which decreased by 0.049 to 3.991 compared to the control plants. No conclusive differences in Fv/F0 were found among the control plants of the genotypes studied within each measurement date. Differences were found only between day 1 (3.328 and 3.578) and day 18 (4.329 and 4.512) for the cv. ´Bohemia´and genotype ´284-17´.
In the case of genotype ´29-17´, the lowest Fv/F0 value was determined on day the 5th (3.835) and the highest on day 18 (4.353). Consistent with the Fv/Fm fluorescence results, it can be concluded that the Fv/F0 ratio also decreased in the stressed variants of all genotypes studied, with the lowest values being achieved by the D2 variant compared to the other stressed variants. No conclusive differences were found within genotypes between measurement dates, except on day the 18th, when conclusive differences were found within measurement dates.
3.4. Water potential (ψw)
Effect of water deficit on wheat plants was monitored based on water potential values (ψw), see Figure 3A-C. The measured values show a conclusive effect of genotype on this characteristic. Among the genotypes studied, the cv. ´Bohemia´ showed the highest water potential (-1.44 MPa) and the genotype ´284-17´ the lowest (-1.54 MPa).
The water potential decreased with water deficit; thus, the effect of the variant was confirmed. At a significance level of =0.05, the lowest water potential was the lowest in the D2 variant (-1.87 MPa) and the highest in the control plants (-1.14 MPa). Lower water potential was observed for variant D1 compared to variant D3.
=0.05, the lowest water potential was the lowest in the D2 variant (-1.87 MPa) and the highest in the control plants (-1.14 MPa). Lower water potential was observed for variant D1 compared to variant D3. For all genotypes studied, a clear reduction in water potential due to water deficit was found in D2 variant. For this variant, the most significant decrease was observed in the genotype ´284-17´ (reduction of 75.58%, -2.02 MPa), followed by genotype ´29-17́´ (60.04%, -1.84 MPa) and the least decrease in water potential was observed in the cv. ´Bohemia´ (54.42 %, -1.74 MPa). On the other hand, the lowest decrease in water potential was again the same for all genotypes. A statistically significant difference between the control and stressed variant was not found in genotype ´284-17´. In this case, the decrease in water potential compared to compared to the control was 0.75 % and its value was -1.14 MPa. On the other hand, in the ´Bohemia´ as the highest, with a decrease of 25.60 % (-1.41 MPa), and for genotype ´284-17´ - 23,59 % (-1.42 MPa).
3.5. Statistical analysis
From Table 4 shows that when changing the variant to D1, the total chlorophyll content is on average 1,55 nM cm-2 lower than in the control plants. Similarly, a reduction in total chlorophyll content can be found when changing the variant to D2. In this case, the chlorophyll content decreased on average 1,61 nM cm-2. When assessing the effect of genotype, it can be noted that in the case of genotype change under control conditions to genotypes ´284-17´ and 29-17´, the content of of total chlorophyll by an average of 8,03 nM cm-2 and 4.14 nM cm-2 higher than in in the case of the variety ´Bohemia´. Statistical analysis shows that water deficit and genotype have a statistically significant effect on total chlorophyll content.

Table 4.
Di, d1 + β2Di, d2 + β3Di, d3 + γ1Di, v1 + γ2Di, v2 + εi) using dummy variables (coefficient significant at α = 0.05 significance level is indicated in bold)..
Table 4.
Di, d1 + β2Di, d2 + β3Di, d3 + γ1Di, v1 + γ2Di, v2 + εi) using dummy variables (coefficient significant at α = 0.05 significance level is indicated in bold)..
| coefficient | transpiration | photosynthesis | Fv/Fm | Fv/F0 | total chlorophylls | carotenoids | water potential |
|---|---|---|---|---|---|---|---|
| constant | 1.705 | 11.005 | 0.796 | 3.98 | 7.811 | 1.256 | 1,089 |
| D1 (d1) | -0.244 | -1.678 | 0.0002 | 0.0017 | -1.553 | 0.143 | -0,416 |
| D2 (d2) | -0.086 | -0.785 | -0.0017 | -0.0540 | -1.608 | -0.072 | -0,725 |
| D3 (d3) | -0.017 | -1.655 | 0.0007 | 0.0050 | -0.462 | -0.109 | -0,282 |
| Genotype ́´284-17́ (v1) |
0.021 | -2.095 | 0.0025 | 0.0720 | 8.030 | 1.242 | -0,099 |
| genotype ́´29-17́ (v2) |
0.183 | 1.499 | 0.0039 | 0.0201 | 4.137 | 0.638 | -0,063 |
The carotenoid content, a statistically significant effect of genotype was found, with genotype under control conditions to genotypes ´284-17´ and ´29-17´, the content of carotenoids was on average 1.24 nM cm-2 and 0.64 nM cm-2 higher than in the case of the variety ´Bohemia´. Rate of photosynthesis was significantly affected by water deficit, when changing the variant to D1, D2 and D3, the value of photosynthesis rate was on average 1.68 M m-2 s-1, 0.79M m-2 s-1 and 1.65 M m-2 s-1 lower than the control, respectively. Similarly, it was shown effect of genotype on changes in photosynthetic rate. Differences were found between genotypes ´284-17´ and ´29-17´ compared to the cv. ´Bohemia´, where both new cultivars the rate of photosynthetic was on average 2.09 M m-2 s-1 and 1.5 M m-2 s-1 lower.
M m-2 s-1, 0.79M m-2 s-1 and 1.65 M m-2 s-1 lower than the control, respectively. Similarly, it was shown effect of genotype on changes in photosynthetic rate. Differences were found between genotypes ´284-17´ and ´29-17´ compared to the cv. ´Bohemia´, where both new cultivars the rate of photosynthetic was on average 2.09 M m-2 s-1 and 1.5 M m-2 s-1 lower. If the transpiration rate is changed to D1, its value is on average 0.24 mM m-2 s-1 lower than the control. If we change the genotype to genotype ´284-17´, the rate of transpiration is on average 0.18 mM m-2 s-1 higher than in control cv. ´Bohemia´. It follows that the water deficit and genotype have a statistically significant effect on transpiration rate.
Evaluation of the effect of genotype and water deficit on fluorescence parameters there was no statistically significant effect.
4. Discusion
The obtained results of the content of photosynthetically active pigments confirm the conclusions of Radzikowska et al. [40] by spelt, Sayed [41] in cereal crops and Zhang et al. [42] by carrot, because the cv. ΄Bohemia΄ showed the lowest chlorophyll content compared to the other monitored genotypes. At the same time, it was confirmed that the observed genotypes of wheat responded differently to the influence of water deficit by reducing the chlorophyll content, see the work of spelt and maize [43,44], when among the genotypes observed, it decreased the least the content of chlorophyll a and b in the leaves due to the drought in the cv. ΄Bohemia΄ and, on the other hand, the most genotype ´284-17´. As a result of the water deficit, the chlorophyll content in the leaves and the ratio of a to b decrease. This reduction can be caused not only by the degradation of chlorophyll, the formation of chlorosis, but above all by the inhibition of chlorophyll synthesis, but also by the formation of reactive oxygen species, which is also confirmed in their work on legumes [45,46] and chives [47]. For all monitored genotypes, the content of total chlorophylls increased after subsequent rehydration, but the values of the control plants were not reached. This conclusion is confirmed by, for example, [48,49]. Similarly, to the chlorophyll content in the leaves, the carotenoid content was influenced by genotype, with genotype ´284-17´ showing the highest carotenoid content and the cv. ΄Bohemia΄ the lowest. Genotypic differences in pigment content are reported, for example, in Catharanthus roseus by Ababaf et al. [50]. Carotenoids, as plant pigments, are involved in reducing the concentration of reactive oxygen species and free radicals, as evidenced by Ramel et al. [51] and Hou et al. [52]. Changes in the concentration of carotenoids due to water deficit are confirmed by works, for example, in maize [53] and soybeans [54]. As a result of the water deficit, the content of carotenoids increased in variant D2, when it was the variant with the most pronounced stress, but this is a genotypic characteristic. Only the cv. ΄Bohemia΄ responded to the most pronounced drought stress by increasing carotenoid concentrations. This conclusion is confirmed by Ababaf et al. [50] in Catharanthus roseus and Taheri et al. [55] in Anchusa italica. The increase in carotenoid concentration in stressed plants may be due to the late accumulation and degradation of carotenoids.
The reduced content of photosynthetically active pigments in leaves (non-stomatal inhibition of photosynthesis) due to water deficit contributed to the reduced CO2 assimilation. However, photosynthesis is not only limited by the pigment content, but also by stomatal conductance, which decreases due to stomatal closure. This conclusion is confirmed, e.g., by [56]. According to [57], the decrease in photosynthesis is due to stomatal closure, a relatively fast process that also reduces water loss. The decrease in stomatal conductance due to water deficit was observed in all genotypes studied within the first few days after induction of stress. The above conclusion is also supported by Figure 2 and Figure 3, which conclude that stomatal closure occurs within 4 days of stress. Furthermore, the effect of genotype on stomatal conductance was found, with genotype ́29-17 ́ having the highest. Water deficit resulted in a decrease in photosynthetic rate in all genotypes studied, with the cv. ΄Bohemia΄ and genotype ́284-17 ́ showing the lowest photosynthesis.
This agrees with the findings of e.g., [40] who compared sown and spelt varieties and also [58] for durum wheat. Reduction of CO2 assimilation due to stomatal closure (stomatal inhibition) leads to limited growth of aboveground biomass, disruption of sink-source relationship and distribution of ATP, NADPH. Photosynthesis is therefore one of the metabolic processes that is affected by water deficit [56,59,60]. At the onset of water deficit, it is likely that the stomatal inhibition of photosynthesis was due to a reduction in stomatal conductance, which was subsequently supplemented by non-stomatal inhibition caused by reduced pigment content, altered electron transport within photosystem II (PSII), and possibly altered enzyme activity, especially Rubisco [59,60]. The photosynthetic rate increased after the re-watering, as also confirmed in maize [61] and Lupinus albus by Pinheriro et al. [62]. The increase in photosynthesis and transpiration rates is due not only to turgor recovery but also to cell growth and new tissue. Similar to photosynthesis rate, transpiration rate was limited by water deficit. The above reduction is not only related to the progressive loss of turgor, but subsequently to the closure of stomata [11,15,20], reducing nutrient uptake. The relationship between stomatal conductance and gas exchange (photosynthesis, transpiration) that was confirmed in our research is also in agreement with the results in maize [63,64], wheat [65,66,67] and soyabean [68]. The reduction in transpiration due to drought was further confirmed by the work Further, the effect of genotype on this characteristic was confirmed in agreement with the work of Wasaya et al. [49], Bakhshandeh et al. [69] and Poudel et al. [70].
One of the important parameters that can be used to determine the level of resistance/sensitivity to stresses are fluorescence parameters. The reported characteristics of Fv/Fm - the maximum quantum yield of PSII [71] and Fv/F0 (non-QA- electron transfer) [72] are the main indicators of the degree of damage to PSII by stressors [73]. Fv/Fm is a measure of the maximum photochemical efficiency of PSII when all reaction centers are open, when its value is in the range of 0.83. Consistent with work [74] in bean, [75,76] in wheat a decrease in Fv/Fm fluorescence ratio values has been demonstrated in plants stressed by water deficit. The decrease in fluorescence may be a way of defense mechanism of the plant due to increased heat and energy dissipation, reduction in carboxylation activity. Thus, the effect of drought is to reduce electron transfer during photosynthesis. Furthermore, according to the results of [77], photoinhibition occurs, which reduces fluorescence. A decrease in Fm has been shown, as also stated by e.g. Gilmore and Björkman [78], Zlatev [79]. However, the effect of genotype on fluorescence changes was not confirmed, the above is in contradiction with the results of spelt who detected changes in Fv/Fm ratio in the context of comparison between sown wheat and spelt varieties. Nevertheless, lower fluorescence was detected in the cv. ΄Bohemia΄ and higher in the genotype '29-17'.
The Fv/Fo ratio is a useful parameter for determining the degree of sensitivity or resistance of a given genotype to a stressor. It has been shown that the cv. ΄Bohemia΄ is more sensitive to drought stress and the genotype '29-17' is tolerant. This confirms the results of [80,81]. This parameter was adversely affected by stress, with partial chlorophyll degradation and reduced electron transfer rate. According to [82,83], the above parameter is influenced by increased Fo, which is related to the inactivation of PSII and the change in the acceptor of reduced plastoquinone. The increase in F0 is accompanied by a decrease in Fm at high stress levels, indicating degradation of the PS II light-harvesting complex, as reported by [83]. The above was confirmed for the D3 variant.
The water deficit effect on juvenile wheat plants resulted in a decrease in leaf water potential. This reduction was due to the gradual dehydration of the tissues and the gradual loss of turgor and closure of stomata. The reduction of leaf water potential due to drought in wheat has been confirmed by, for example [84] and, in the case of osmotic stress [85]. Furthermore, the effect of genotype was confirmed, with the highest decrease in water potential due to drought observed in the cv. ΄Bohemia΄ and the lowest in the genotype ́284-17 ́. Genotypic differences in stress response have been confirmed in wheat by, for example [84,86]. In the case of genotype ́284-17 ́, osmotic adaptation to water deficit may be involved, where osmotic adaptation is one of the important physiological adaptations of plants to stress. Osmotic adjustment leads to maintenance of cellular turgor tension and increased water transport based on changes in the concentrations of substances inside the cell and external solution. The influence of osmotic adjustment as an adaptation to water deficit has also been reported, e.g., by [87,88,89].
5. Conclusions
The obtained results indicate differences in drought response of the studied sown wheat genotypes. The content of photosynthetically active pigments (chlorophyll a and b, carotenoids) was influenced by genotype, with genotype ´284-17´ having a higher content of chlorophylls and carotenoids and cv. ´Bohemia´ the lowest. The chlorophyll content decreased under the effect of water deficit, i.e., most significantly after the irrigation period (D2). The carotenoid content increased only in the ´Bohemia´ variety due to water deficit (D2). In contrast, the carotenoid content of the newly bred genotypes (genotypes ´284-17´ and ´29-17´) decreased due to the water deficit. Drought and subsequent rehydration affected the primary metabolism of juvenile wheat plants. Wheat plants reduced photosynthetic rates due to stomatal closure and reduced substomal CO2 levels caused by water deficit. The above effect was observed in genotype ´29-17´ and cv 'Bohemia'. The rate of transpiration in genotype ´284-17´ decreased because of water deficit. On the contrary, in genotype ´29-17´ and cv. ´Bohemia´, rate of transpiration increased under water deficit. This effect is inappropriate with respect to drought tolerance. Among the genotypes studied, genotype ´29-17´ showed higher photosynthetic and transpiration rates compared to genotype ´284-17´ and the cv. ´Bohemia´. In all genotypes studied, the there was a clear reduction in water potential due to water deficit, most markedly in genotype ´284-17´, followed by genotype ´29-17´ and least markedly in water potential decreased in the cv. ´Bohemia´. Genotype ´29-17´ appears to be promising with regard to drought resistance.
Author Contributions
S.L. and T.R. designed and supervised the project. F.H., S.L. analyzed the data and wrote the manuscript. F.H., H.H. and S.L. participated in the determination of physiological parameters. J.P., T.R. participated in the material preparation. All authors discussed the results and commented on the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Ministry of Agriculture of the Czech Republic, Project No. QK 1910343.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author. The data are not publicly available due to the involvement of other unpublished papers.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the result.
References
- Lipiec, J.; Doussan, C.; Nosalewicz, A.; Kondracka, K. Effect of drought and heat stresses on plant growth and yield: a review. Int. Agrophys. 2013, 27(4), 463–477. [Google Scholar] [CrossRef]
- Khan, M.A.; Iqbal, M.; Akram, M.; Ahmad, M.; Hassan, M.W.; Jamil, M. Recent advances in molecular tool development for drought tolerance breeding in cereal crops: A review. Zemdirb. Agric. 2013, 100, 325–334. [Google Scholar] [CrossRef]
- Asfaw, A.; Blair, M.W. Quantification of Drought Tolerance in Ethiopian Common Bean Varieties. Agric. Sci. 2014, 5, 124–139. [Google Scholar] [CrossRef]
- Sekhon, H.S., Singh, G., Sharma, P., Bains, T.S. Water use efficiency under stress environments. In.: Yadav, S.S.; Redden, R. (eds.): Climate change and management of cool season grain legume crops. Springer. 2010, 207-227.
- Tian, L.; Li, J.; Bi, W.; Zuo, S.; Li, L.; Li, W.; Sun, L. Effects of waterlogging stress at different growth stages on the photosynthetic characteristics and grain yield of spring maize (Zea mays L.) under field conditions. Agric. Water Manag. 2019, 218, 250–258. [Google Scholar] [CrossRef]
- Bettaieb, I.; Zakhama, N.; Wannes, W.A.; Kchouk, M. , Marzouk; B. Water deficit effects on Salvia officinalis fatty acids and essential oils composition. Sci. Hortic. 2009, 120, 271–275. [Google Scholar] [CrossRef]
- Mladenov, P.; Aziz, S.; Topalova, E.; Renaut, J.; Planchon, S.; Raina, A.; Tomlekova, N. Physiological responses of common bean genotypes to drought stress. Agron. 2023, 13(4), 1022. [Google Scholar] [CrossRef]
- Khaleghi, A.; Naderi, R.; Brunetti, C.; Maserti, B.E.; Salami, S.A.; Babalar, M. Morphological, physiochemical and antioxidant responses of Maclura pomifera to drought stress. Sci. Rep. 2019, 9(1), e19250. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Lu, M.; Wang, Y.; Wang, Y.; Liu, Z.; Chen, S. Response mechanism of plants to drought stress. Horticulturae. 2021, 7(3), 1–36. [Google Scholar] [CrossRef]
- Keerthi Sree, Y.; Lakra, N.; Manorama, K.; Ahlawat, Y.; Zaid, A.; Elansary, H.O.; Saved, S.R.M.; Mahmoud, E.A. Drought-Induced Morpho-Physiological, Biochemical, Metabolite Responses and Protein Profiling of Chickpea (Cicer arietinum L.). Agron. 2023, 13(7), 1814.
- Majeed, S.; Nawaz, F.; Naeem, M.; Ashraf, M.Y.; Ejaz, S.; Ahmad, K.S.; Saba, T.; Farid, G.; Khalid, I.; Mehmood, K. Nitric oxide regulates water status and associated enzymatic pathways to inhibit nutrients imbalance in maize (Zea mays L.) under drought stress. Plant Physiol. Biochem. 2020, 155, 147–160. [Google Scholar] [CrossRef]
- Ding, Y.; Nie, Y.; Chen, H.; Wang, K.; Querejeta, J.I. Water uptake depth is coordinated with leaf water potential, water-use efficiency and drought vulnerability in karst vegetation. New Phytol. 2021, 229(3), 1339–1353. [Google Scholar] [CrossRef] [PubMed]
- Hura, T.; Hura, K.; Ostrowska, A.; Urban, K. Non-rolling flag leaves use an effective mechanism to reduce water loss and light-induced damage under drought stress. Ann. Bot. 2022, 130(3), 393–408. [Google Scholar] [CrossRef] [PubMed]
- Sewore, B.M.; Abe, A.; Nigussie, M. Evaluation of bread wheat (Triticum aestivum L.) genotypes for drought tolerance using morpho-physiological traits under drought-stressed and well-watered conditions. Plos one 2023, 18(5), e0283347. [Google Scholar] [CrossRef]
- Liu, H.; Song, S.; Zhang, H.; Li, Y.; Niu, L.; Zhang, J.; Wang, W. Signaling transduction of ABA, ROS, and Ca2+ in plant stomatal closure in response to drought. Int. J. Mol. Sci. 2022, 23(23), 14824. [Google Scholar] [CrossRef]
- Onyemaobi, O.; Sangma, H.; Garg, G.; Wallace, X.; Kleven, S.; Suwanchaikasem, P.; Roessner, U.; Dolferus, R. Reproductive stage drought tolerance in wheat: Importance of stomatal conductance and plant growth regulators. Genes 2021, 12(11), 1742. [Google Scholar] [CrossRef] [PubMed]
- Khalvandi, M.; Siosemardeh, A.; Roohi, E.; Keramati, S. Salicylic acid alleviated the effect of drought stress on photosynthetic characteristics and leaf protein pattern in winter wheat. Heliyon. 2021, 7(1), e05908. [Google Scholar] [CrossRef]
- Liang, G.; Liu, J.; Zhang, J.; Guo, J. Effects of drought stress on photosynthetic and physiological parameters of tomato. J. Am. Soc. Hortic. Sci. 2020, 145(1), 12–17. [Google Scholar] [CrossRef]
- Parry, M.A.; Andralojc, P.J.; Khan, S.; Lea, P.J.; Keys, A.J. Rubisco activity: effects of drought stress. Ann. Bot. 2002, 89(7), 833–839. [Google Scholar] [CrossRef]
- Grzesiak, M.T.; Rzepka, A.; Hura, T.; Hura, K.; Skoczowski, A. Changes in response to drought stress of triticale and maize genotypes differing in drought tolerance. Photosynthetica, 2007, 45, 280–287. [Google Scholar] [CrossRef]
- Sabagh, A.E.; Hossain, A.; Islam, M.S.; Barutcular, C.; Hussain, S.; Hasanuzzaman, M.; Akram, T.; Mubeen, M.; Nasim, W.; Fahad, S.; Kumar, N.; Meena, R.S.; Ferhat, K.; Fwerhat, K.; Mehmet, Y.; Ratnasekera, D.; Saneoka, H. Drought and salinity stresses in barley: consequences and mitigation strategies. Aust. J. Crop Sci. 2019, 13(6), 810–820. [Google Scholar] [CrossRef]
- Rahim, F.P.; María Alejandra, T.T.; Víctor Manuel, Z.V.; José Elías, T.R.; Maginot, N.H. Stomatal traits and barley (Hordeum vulgare L.) forage yield in drought conditions of Northeastern Mexico. Plants 2021, 10(7), 1318. [Google Scholar] [CrossRef]
- Farooq, M.; Rizwan, M.; Nawaz, A.; Rehman, A.; Ahmad, R. Application of natural plant extracts improves the tolerance against combined terminal heat and drought stresses in bread wheat. J. Agron. Crop Sci. 2017, 203, 528–538. [Google Scholar] [CrossRef]
- Popko, M.; Michalak, I.; Wilk, R.; Gramza, M.; Chojnacka, K.; Górecki, H. Effect of the new plant growth biostimulants based on amino acids on yield and grain quality of winter wheat. Molecules. 2018, 23(2), 470. [Google Scholar] [CrossRef]
- Moustakas, M.; Sperdouli, I.; Moustaka, J. Early drought stress warning in plants: Color pictures of photosystem II photochemistry. Climate. 2022, 10(11), 179. [Google Scholar] [CrossRef]
- Chaves, M.M.; Flexas, J.; Pinheiro, C. Photosynthesis under drought and salt stress: regulation mechanisms from whole plant to cell. Ann. Bot. 2009, 103, 551–560. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Al-Suhaibani, N.; Ali, N.; Akmal, M.; Alotaibi, M.; Refay, Y.; Dindaroglu, Y.; Abdul-Wajid, H.H.; Battaglia, M.L. Drought stress impacts on plants and different approaches to alleviate its adverse effects. Plants. 2021, 10(2), 259. [Google Scholar] [CrossRef]
- Aliyeva, D.R.; Aydinli, L.M.; Pashayeva, A.N.; Zulfugarov, I.S.; Huseynova, I.M. Photosynthetic machinery and antioxidant status of wheat genotypes under drought stress followed by rewatering. Photosynthetica. 2020, 58(5), 1217–1225. [Google Scholar] [CrossRef]
- Mwadzingeni, L.; Shimelis, H.; Dube, E.; Laing, M. D.; Tsilo, T.J. Breeding wheat for drought tolerance: Progress and technologies. J. Integr. Agric. 2016, 15(5), 935–943. [Google Scholar] [CrossRef]
- Rizwan, M.; Mahboob, W.; Faheem, M.; Shimelis, H.; Hameed, A.; Sial, M.A.; Shokat, S. Can we exploit supernumerary spikelet and spike branching traits to boost bread wheat (Triticum aestivum L.) yield? Appl. Ecol. Environ. Res. 2020, 18(5), 6243–6258. [Google Scholar] [CrossRef]
- Wang, Y.; Du, F.; Wang, J.; Wang, K.; Tian, C.; Qi, X.; Lu, F.; Liz, X.; Jiao, Y. (2022). Improving bread wheat yield through modulating an unselected AP2/ERF gene. Nat. Plants. 2022, 8(8), 930–939. [Google Scholar] [CrossRef]
- Zhang, X.; Qiao, L.; Li, X.; Yang, Z.; Liu, C.; Guo, H.; Chang, Z.; Zheng, J.; Zhangm, S.; Chang, S.; Chang, L.; Chen, F.; Jia, J.; Yan, L.; Chang, Z. Genetic incorporation of the favorable alleles for three genes associated with spikelet development in wheat. Front. Plant Sci., 2022, 13, e892642. [Google Scholar] [CrossRef]
- Li, Y.; Li, L.; Zhao, M.; Guo, L.; Guo, X.; Zhao, D.; Batool, A.; Dong, B.; Xu, H.; Cui, S.; Zhang, A.; Fu, X.; Jing, R.; Liu, X. Wheat FRIZZY PANICLE activates VERNALIZATION1-A and HOMEOBOX4-A to regulate spike development in wheat. Plant Biotechnol. J. 2021, 19(6), 1141–1154. [Google Scholar] [CrossRef] [PubMed]
- Du, D.; Zhang, D.; Yuan, J.; Feng, M.; Li, Z.; Wang, Z.; Zhang, Z.; Li, X.; Ke, W.; Li, R.; Chen, Z.; Chai, L.; Hu, Z.; Guo, W.; Ni, Z. FRIZZY PANICLE defines a regulatory hub for simultaneously controlling spikelet formation and awn elongation in bread wheat. New Phytol. 2021, 231(2), 814–833. [Google Scholar] [CrossRef]
- Bienkowska, T.; Suchowilska, E.; Wiwart, M. Triticum polonicum L. as promising source material for breeding new wheat cultivars. J. Elem. 2020, 25(1), 237–248. [Google Scholar]
- Romanov, B.V.; Pimonov, K.I.; Lipskij, D.D. Produkcionnye ocobennosti pšenicy Triticum petropavlovskyi. Izvestiya Niznevolzskogo agrouniversitetskogo kompleksa: nauka i vyssee professionalnoe obrazovanie. 2020, 4(60), 172–182. [only Russian].
- Porra, R.; Thompson, W.; Kriedemann, P. Determination of accurate extinction coefficients and simultaneous equations for assaying chlorophylls a and b extracted with four different solvents: Verification of the concentration of chlorophyll standards by atomic absorption spectroscopy. Biochim. Biophys. Acta (BBA)-Bioenerg. 1989, 975, 384–394. [Google Scholar] [CrossRef]
- Kuklova, M.; Hnilickova, H.; Kukla, J.; Hnilicka, F. Environmental impact of the Al smelter on physiology and macronutrient contents in plants and Cambisols. Plant Soil Environ. 2015, 61, 72–78. [Google Scholar] [CrossRef]
- Banks, J.M.; Hirons, A.D. Alternative methods of estimating the water potential at turgor loss point in Acer genotypes. Plant Methods. 2019, 15, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Radzikowska, D.; Sulewska, H.; Bandurska, H.; Ratajczak, K.; Szymańska, G.; Kowalczewski, P.Ł.; Głowicka-Wołoszyn, R. Analysis of physiological status in response to water deficit of spelt (Triticum aestivum ssp. spelta) cultivars in reference to common wheat (Triticum aestivum ssp. vulgare). Agron. 2022, 12(8), 1822. [Google Scholar] [CrossRef]
- Sayed, O.H. Chlorophyll fluorescence as a tool in cereal crop research. Photosynthetica. 2003, 41, 321–330. [Google Scholar] [CrossRef]
- Zhang, R.R.; Wang, Y.H.; Li, T.; Tan, G.F.; Tao, J.P.; Su, X.J.; Xu, S.H.; Tian, Y.S.; Xiong, A.S. Effects of simulated drought stress on carotenoid contents and expression of related genes in carrot taproots. Protoplasma, 2021, 258, 379–390. [Google Scholar] [CrossRef]
- Hussain, I.; Rasheed, R.; Ashraf, M.; Mohsin, M.; Ali, S.; Rashid, A.; Akram, M.; Nisar, J.; Riaz, M. Foliar Applied acetylsalicylic acid induced growth and key-biochemical changes in chickpea (Cicer arietinum L.) under drought stress. Dose-Response 2020, 18(4), 1–13. [Google Scholar] [CrossRef]
- Shafiq, S.; Akram, N.A.; Ashraf, M.; García-Caparrós, P.; Ali, O.M.; Latef, A.A.H.A. Influence of glycine betaine (natural and synthetic) on growth, metabolism and yield production of drought-stressed maize (Zea mays L.) plants. Plants, 2021, 10, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Talebi, R.; Ensafi, M.H.; Baghebani, N.; Karami, E.; Mohammadi, K. Physiological responses of chickpea (Cicer arietinum) genotypes to drought stress. Environ. Exp. Biol. 2013, 11, 9–15. [Google Scholar]
- Mahdavi Mashaki, K.; Garg, V.; Nasrollahnezhad Ghomi, A.A.; Kudapa, H.; Chitikineni, A.; Zaynali Nezhad, K.; Yamchi, A.; Soltanloo, H.; Varshney, R.K.; Thudi, M. RNA-Seq analysis revealed genes associated with drought stress response in kabuli chickpea (Cicer arietinum L.). PLoS ONE. 2018, 13, e0199774. [Google Scholar] [CrossRef]
- Egert, M.; Tevini, M. Influence of drought on some physiological parameters symptomatic for oxidative stress in leaves of chives (Allium schoenoprasum). Environ. Exp. Bot. 2002, 48, 43–49. [Google Scholar] [CrossRef]
- Barboričová, M.; Filaček, A.; Vysoká, D.M.; Gašparovič, K.; Živčák, M.; Brestic, M. Sensitivity of fast chlorophyll fluorescence parameters to combined heat and drought stress in wheat genotypes. Plant, Soil and Environ. 2022, 68(7), 309–316. [Google Scholar] [CrossRef]
- Wasaya, A.; Manzoor, S.; Yasir, T.A.; Sarwar, N.; Mubeen, K.; Ismail, I.A.; Raza, A.; Rehman, A.; Hossain, A.; EL Sabagh, A. Evaluation of fourteen bread wheat (Triticum aestivum L.) genotypes by observing gas exchange parameters, relative water and chlorophyll content, and yield attributes under drought stress. Sustainability 2021, 13(9), 4799. [Google Scholar] [CrossRef]
- Ababaf, M.; Omidi, H.; Bakhshandeh, A. Changes in antioxidant enzymes activities and alkaloid amount of Catharanthus roseus in response to plant growth regulators under drought condition. Ind. Crops Prod. 2021, 167, 113505. [Google Scholar] [CrossRef]
- Ramel, F.; Mialoundama, A.S.; Havaux, M. Nonenzymic carotenoid oxidation and photooxidative stress signalling in plants. J. Exp. Bot. 2013, 64(3), 799–805. [Google Scholar] [CrossRef]
- Khalilzadeh, R.; Seyed Sharifi, R.; Jalilian, J. Antioxidant status and physiological responses of wheat (Triticum aestivum L.) to cycocel application and bio fertilizers under water limitation condition. J. Plant Interact. 2016, 11(1), 130–137. [Google Scholar] [CrossRef]
- Mohammadkhani, N.; Heidari, R. Effects of water stress on respiration, photosynthetic pigments and water. Pak. J. Biol. Sci. 2007, 10(22), 4022–4028. [Google Scholar] [CrossRef]
- Rys, M.; Szaleniec, M.; Skoczowski, A.; Stawoska, I.; Janeczko, A. FT-Raman spectroscopy as a tool in evaluation the response of plants to drought stress. Open Chem. 2015, 13(1), 1091–1100. [Google Scholar] [CrossRef]
- Taheri, Z.; Vatankhah, E.; Jafarian, V. Methyl jasmonate improves physiologicalical responses of Anchusa italica under salinity stress. South Afr. J. Bot. 2020, 0, 375–382. [Google Scholar] [CrossRef]
- Kollist, H.; Zandalinas, S.I.; Sengupta, S.; Nuhkat, M.; Kangasjärvi, J.; Mittler, R. Rapid responses to abiotic stress: priming the landscape for the signal transduction network. Trends Plant Sci. 2019, 24, 25–37. [Google Scholar] [CrossRef]
- Onyemaobi, O.; Sangma, H.; Garg, G.; Wallace, X.; Kleven, S.; Suwanchaikasem, P.; Roessner, U.; Dolferus, R. Reproductive stage drought tolerance in wheat: Importance of stomatal conductance and plant growth regulators. Genes. 2021, 12(11), 1742. [Google Scholar] [CrossRef] [PubMed]
- Dib, T.A.; Monneveux, P.; Acevedo, E.; Nachit, M.M. Evaluation of proline analysis and chlorophyll fluorescence quenching measurements as drought tolerance indicators in durum wheat (Triticum turgidum L. var. durum). Euphytica. 1994, 79, 65–73. [Google Scholar] [CrossRef]
- Kandel, S. Wheat responses, defence mechanisms and tolerance to drought stress: a review article. Int. J. Appl. Sci. Biotechnol. 2021, 8, 99–109. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, X.; Han, Z.; Feng, H.; Wang, Y.; Kang, J.; Han, X.; Wang, L.; Wang, C.; Li, H.; Ma, G. Analysis of physiological indicators associated with drought tolerance in wheat under drought and re-watering conditions. Antioxidants. 2022, 11(11), 2266. [Google Scholar] [CrossRef] [PubMed]
- Badr, A.; Brüggemann, W. Comparative analysis of drought stress response of maize genotypes using chlorophyll fluorescence measurements and leaf relative water content. Photosynthetica. 2020, 58(2), 38–645. [Google Scholar] [CrossRef]
- Pinheriro, C.; Passarinho, J.A.; Ricardo, C.P. Effect of drought and rewatering on the metabolism of Lupinus albus organs. J. Plant Physiol. 2004, 161, 1203–1210. [Google Scholar] [CrossRef] [PubMed]
- Abid, M.; Ali, S.; Qi, L.K.; Zahoor, R.; Tian, Z.; Jiang, D.; Snider, J.L.; Dai, T. Physiological and biochemical changes during drought and recovery periods at tillering and jointing stages in wheat (Triticum aestivum L.). Sci. Rep. 2018, 8, 4615. [Google Scholar] [CrossRef]
- Qi, M.; Liu, X.; Li, Y.; Song, H.; Yin, Z.; Zhang, F.; He, Q.; Xu, Z.; Zhou, G. Photosynthetic resistance and resilience under drought, fooding and rewatering in maize plants. Photosynth. Res. 2021, 148, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Nisa, W.; Nisa, V.; Nagoo, S.A.; Dar, Z.A. Drought tolerance mechanism in wheat: a review. Pharma Innovation, 2019, 2, 714–724. [Google Scholar]
- Qayyum, A.; Al Ayoubi, S.; Sher, A.; Bibi, Y.; Ahmad, S.; Shen, Z.; Jenks, M. Improvement in drought tolerance in bread wheat is related to an improvement in osmolyte production, antioxidant enzyme activities, and gaseous exchange. Saudi J. Biol. Sci. 2021, 28(9), 5238–5249. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Cernusak, L.A.; Song, X. Dynamic responses of gas exchange and photochemistry to heat interference during drought in wheat and sorghum. Funct. Plant Biol. 2020, 47(7), 611–627. [Google Scholar] [CrossRef]
- Katam, R.; Shokri, S.; Murthy, N.; Singh, S.K.; Suravajhala, P.; Khan, M.N.; Bahmani, M.; Sakata, K.; Reddy, K.R. Proteomics, physiological, and biochemical analysis of cross tolerance mechanisms in response to heat and water stresses in soybean. PLoS ONE. 2020, 15, e0233905. [Google Scholar] [CrossRef]
- Bakhshandeh, S.; Corneo, P.E.; Yin, L.; Dijkstra, F.A. Drought and heat stress reduce yield and alter carbon rhizodeposition of different wheat genotypes. J. Agron. Crop Sci. 2019, 205(2), 157–167. [Google Scholar] [CrossRef]
- Poudel, M.R.; Ghimire, S.; Pandey, M.P.; Dhakal, K.H.; Thapa, D.B.; Poudel, H.K. Evaluation of wheat genotypes under irrigated, heat stress and drought conditions. J Biol Today's World. 2020, 9(1), 1–12.
- Papageorgiou, G.C., Stamatakis, K.2004). Water and solute transport in cyanobacteria as probed by chlorophyll fluorescence. In Chlorophyll a fluorescence: a signature of photosynthesis (pp. 663–678). Dordrecht: Springer Netherlands.
- Kramer, D.M.; Johnson, G.; Kiirats, O.; Edwards, G.E. New fluorescence parameters for the determination of QA redox state and excitation energy fluxes. Photosynth. Res. 2004, 79, 209–218. [Google Scholar] [CrossRef]
- Peterson, R. , Havir, E. Photosynthetic properties of an Arabidopsis thaliana mutant possessing a defective PsbS gene. Planta. 2001, 214, 142–152. [Google Scholar] [CrossRef]
- Afshar Mohamadian, M.; Omidipour, M.; Jamal Omidi, F. Effect of different drought stress levels on chlorophyll fluorescence indices of two bean cultivars. J. Plant Res. (Iran. J. Biol.). 2018, 31, 511–525. [Google Scholar]
- Wu, X.; Tang, Y.; Li, C.; Wu, C.; Huang, G. Chlorophyll fluorescence and yield responses of winter wheat to waterlogging at different growth stages. Plant Prod. Sci. 2015, 18, 284–294. [Google Scholar] [CrossRef]
- Sun, Z.W.; Ren, L.K.; Fan, J.W.; Li, Q.; Wang, K.J.; Guo, M.M.; Li, J.; Zhang, G.X.; Yang, Z.Y.; Chen, F.; Li, X.N. Salt response of photosynthetic electron transport system in wheat cultivars with contrasting tolerance. Plant, Soil and Environ. 2016, 62(11), 515–521. [Google Scholar] [CrossRef]
- Lu, C.; Zhang, J. Effects of water stress on photosynthesis, chlorophyll fluorescence and photoinhibition in wheat plants. Funct. Plant Biol. 1998, 25, 883–892. [Google Scholar] [CrossRef]
- Gilmore, A.; Björkman, O. Temperature-sensitive coupling and uncoupling of ATPase-mediated, nonradiative energy dissipation: Similarities between chloroplasts and leaves. Planta. 1995, 197, 646–654. [Google Scholar] [CrossRef]
- Zlatev, Z. Drought-induced changes in chlorophyll fluorescence of young wheat plants. Biotechnol. Biotechnol. Equip. 2009, 23, 438–441. [Google Scholar] [CrossRef]
- Simova-Stoilova, L.; Pecheva, D.; Kirova, E. Drought stress response in winter wheat varieties –changes in leaf proteins and proteolytic activities. Acta Bot. Croat. 2020, 79, 121–130. [Google Scholar] [CrossRef]
- Larouk, C.; Gabon, F.; Kehel, Z.; Djekoun, A.; Nachit, M.; Amri, A. Chlorophyll fluorescence and drought tolerance in a mapping population of durum wheat. Curr. Agric. Res. J 2021, 70(3-4), 123–134. [Google Scholar] [CrossRef]
- Zlatev, Z.S. Drought-induced changes and recovery of photosynthesis in two bean cultivars (Phaseolus vulgaris L.). Emir. J. Food Agric. 2013, 25, 1014–1023. [Google Scholar] [CrossRef]
Figure 1.
Effect of experimental variation on leaf gas exchange rate of juvenile wheat plants, rate of photosynthesis (Pn) - M m-2 s-1 and rate of transpiration (E) - mM m-2 s-1, depending on wheat genotype, with photosynthetic rates shown in graphs A (´Bohemia´), C (´284-17´), and E (´ 29-17´). Transpiration rates are shown in graphs B (´Bohemia´), D (´284-17´), and F (´29-17´). The lines in the graphs indicate the standard error (S.E.) values at the = 0.05 significance level.
M m-2 s-1 and rate of transpiration (E) - mM m-2 s-1, depending on wheat genotype, with photosynthetic rates shown in graphs A (´Bohemia´), C (´284-17´), and E (´ 29-17´). Transpiration rates are shown in graphs B (´Bohemia´), D (´284-17´), and F (´29-17´). The lines in the graphs indicate the standard error (S.E.) values at the = 0.05 significance level.
Figure 1.
Effect of experimental variation on leaf gas exchange rate of juvenile wheat plants, rate of photosynthesis (Pn) - M m-2 s-1 and rate of transpiration (E) - mM m-2 s-1, depending on wheat genotype, with photosynthetic rates shown in graphs A (´Bohemia´), C (´284-17´), and E (´ 29-17´). Transpiration rates are shown in graphs B (´Bohemia´), D (´284-17´), and F (´29-17´). The lines in the graphs indicate the standard error (S.E.) values at the = 0.05 significance level.
M m-2 s-1 and rate of transpiration (E) - mM m-2 s-1, depending on wheat genotype, with photosynthetic rates shown in graphs A (´Bohemia´), C (´284-17´), and E (´ 29-17´). Transpiration rates are shown in graphs B (´Bohemia´), D (´284-17´), and F (´29-17´). The lines in the graphs indicate the standard error (S.E.) values at the = 0.05 significance level.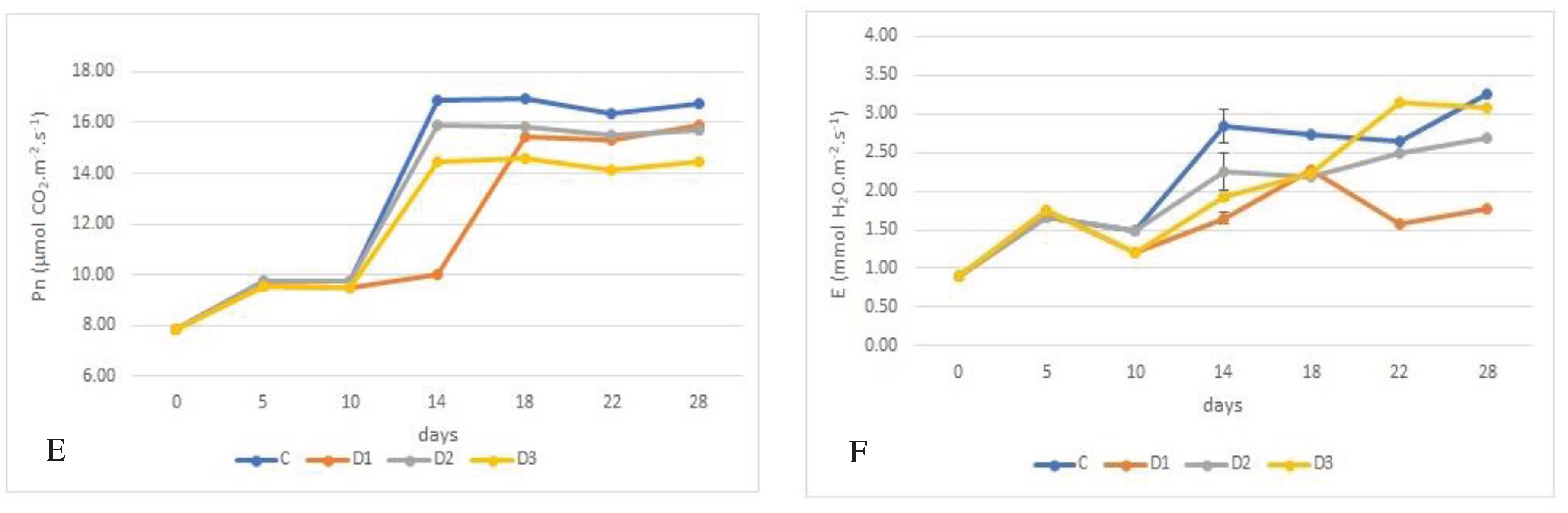
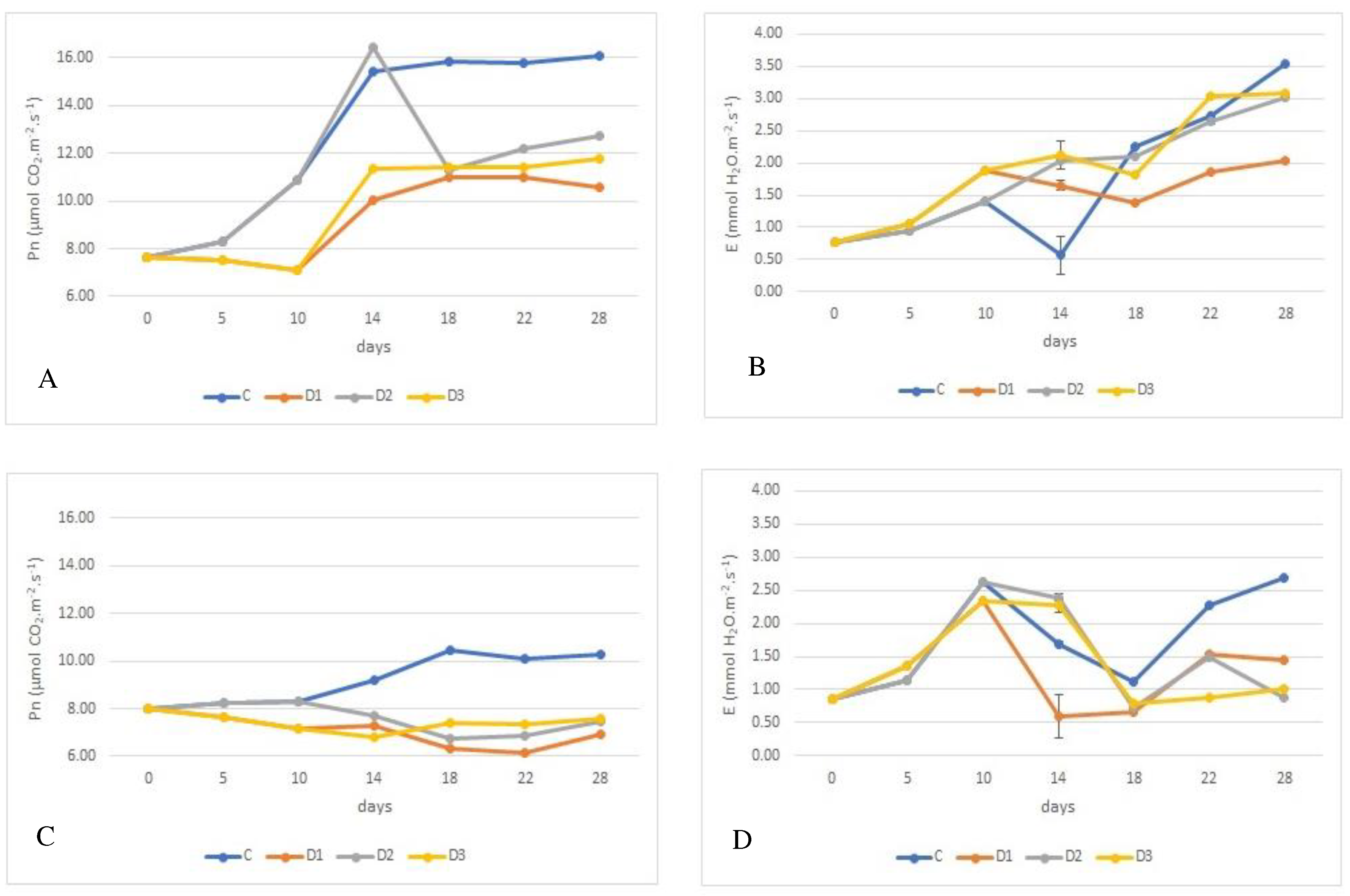
Figure 2.
Effect of experimental variation on parameters of fluorescence by juvenile wheat plants, FV/Fm and Fv/F0, depending on wheat genotype, with Fv/Fm shown in graphs A (´Bohemia´), C (´284-1), and E (´29-17´) and Fv/F0 are shown in graphs B (´Bohemia´), D (´284-17´), and F (´29-17´). The lines in the graphs indicate the standard error (S.E.) values at the = 0.05 significance level.
= 0.05 significance level.
Figure 2.
Effect of experimental variation on parameters of fluorescence by juvenile wheat plants, FV/Fm and Fv/F0, depending on wheat genotype, with Fv/Fm shown in graphs A (´Bohemia´), C (´284-1), and E (´29-17´) and Fv/F0 are shown in graphs B (´Bohemia´), D (´284-17´), and F (´29-17´). The lines in the graphs indicate the standard error (S.E.) values at the = 0.05 significance level.
= 0.05 significance level.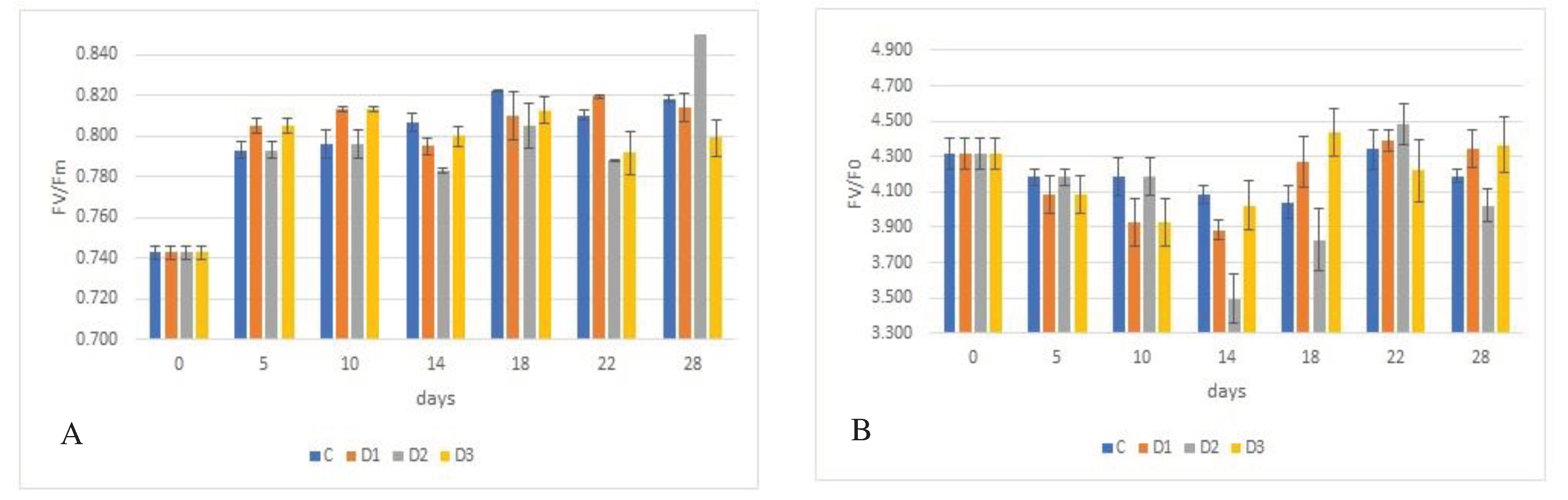
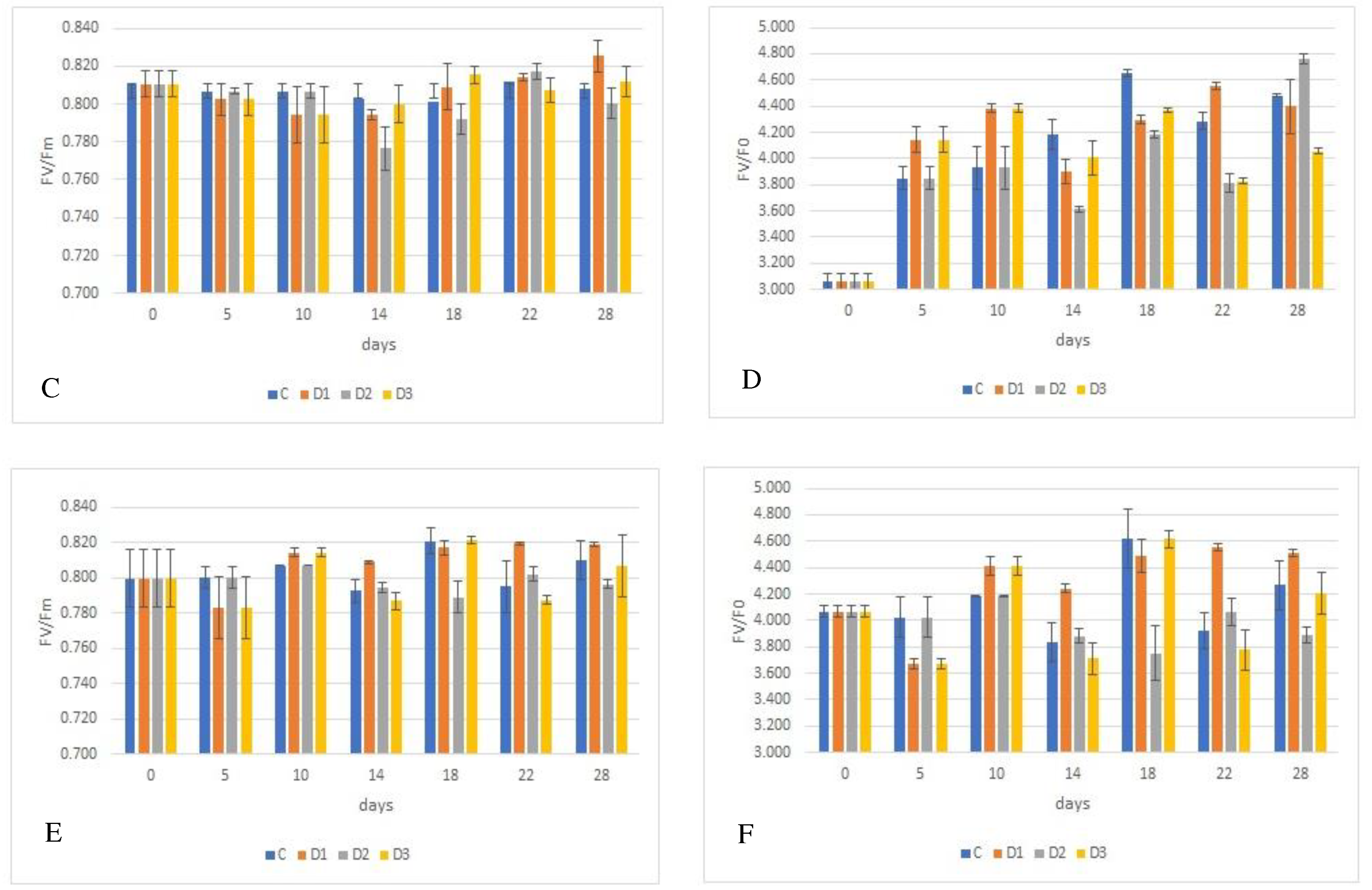
Figure 3.
Effect of experimental variant and time on plant water regime, as determined by water potential (MPa) of leaves of juvenile wheat plants, as a function of wheat genotype, where water potential values (ψw) is shown in graphs A (´Bohemia´), B (´284-17´) and C (´29-17´). The lines in the graphs indicate the standard error (S.E.) values at a significance level of = 0.05.
= 0.05.
Figure 3.
Effect of experimental variant and time on plant water regime, as determined by water potential (MPa) of leaves of juvenile wheat plants, as a function of wheat genotype, where water potential values (ψw) is shown in graphs A (´Bohemia´), B (´284-17´) and C (´29-17´). The lines in the graphs indicate the standard error (S.E.) values at a significance level of = 0.05.
= 0.05.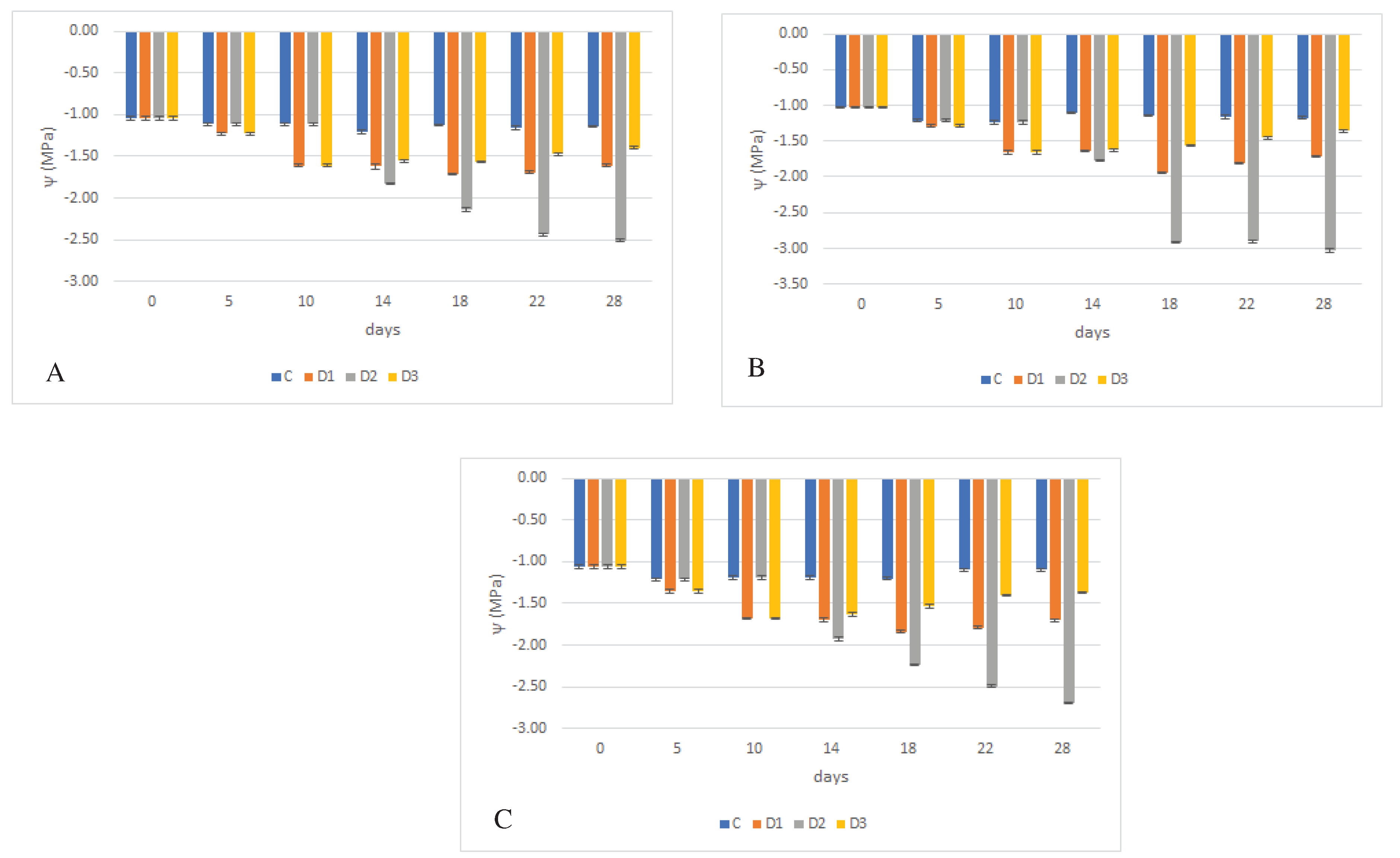
Table 1.
Scheme of the experiment.
| Variant | Day of experiment | |||||
|---|---|---|---|---|---|---|
| 0 – 5th | 6th – 10th | 10th – 14th | 14th – 19th | 19th – 24th | 24th – 28th | |
| Control (C) | irrigation | irrigation | irrigation | irrigation | irrigation | irrigation |
| Drought 1 (D1) | drought | drought | irrigation | drought | drought | irrigation |
| Drought 2 (D2) | irrigation | irrigation | drought | drought | drought | drought |
| Drought 3 (D3) | drought | drought | irrigation | irrigation | irrigation | irrigation |
Table 2.
Effect of variation on the content of photosynthetically active pigments (nM cm-2), total chlorophylls (Chltot) and carotenoids (Car), depending on wheat genotypes. Statistically significant differences at the α = 0.05 significance level are indicated by letters.
Table 2.
Effect of variation on the content of photosynthetically active pigments (nM cm-2), total chlorophylls (Chltot) and carotenoids (Car), depending on wheat genotypes. Statistically significant differences at the α = 0.05 significance level are indicated by letters.
|
Variant |
Days |
´Bohemia´ | ´287-17´ | ´29-17´ | |||
| Chltot | Car | Chltot | Car | Chltot | Car | ||
|
C |
0 | 5.368 ± 0.133 | 0.889 ± 0.030 | 15.118 ± 0.059 | 2.271 ± 0.031 | 9.027 ± 0.067 | 1.330 ± 0.005 |
| 5 | 5.860 ± 0.480 | 1.001 ± 0.084 | 15.509 ± 0.926 | 2.102 ± 0.130 | 11.277 ± 0.837 | 1.558 ± 0.153 a, b | |
| 10 | 7.027 ± 0.064 | 1.201 ± 0.014 | 15.279 ± 1.541 | 2.636 ± 0.223 | 11.801 ± 0.325 | 1.934 ± 0.041 | |
| 14 | 7.596 ± 0.738 | 1.223 ± 0.124 | 16.698 ± 5.123 | 2.571 ±0.637 d | 11.944 ± 1.376 | 1.912 ± 0.268 a, b, c | |
| 18 | 7.962 ± 0.246 | 1.343 ± 0.042 | 17.232 ± 0.596 | 2.789 ± 0.102 | 12.511 ± 0.549 | 1.923 ± 0.072 | |
| 22 | 8.881 ± 0.483 c, d | 1.471 ± 0.057 | 17.490 ± 0.795 | 2.862 ± 0.344 | 13.635 ± 0.317 | 2.085 ± 0.079 | |
| 28 | 9.468 ± 0.434 a | 1.524 ± 0.077 a, b, c | 19.948 ± 6.642 f | 3.288 ± 0.207 f | 13.703 ± 0.618 | 2.230 ± 0.021 b, c, d | |
|
D1 |
0 | 5.368 ± 0.133 | 0.889 ± 0.030 | 15.118 ± 0.059 c | 2.271 ± 0.031 | 9.027 ± 0.067 d | 1.330 ± 0.005 a |
| 5 | 5.266 ± 0.448 | 0.890 ± 0.048 | 14.496 ± 0.295 | 2.355 ± 0.076 | 11.012 ± 0.081 | 1.791 ± 0.034 | |
| 10 | 5.086 ± 0.377 a | 0.979 ± 0.080 b | 14.065 ± 1.361 | 2.181 ± 0.226 | 10.995 ± 1.589 | 1.797 ± 0.239 | |
| 14 | 7.599 ± 0.740 | 1.181 ± 0.102 | 15.400 ± 0.026 | 3.306 ± 0.077 | 11.674 ± 0.021 | 3.214 ± 0.044 | |
| 18 | 7.487 ± 0.180 | 1.148 ± 0.036 | 13.141 ± 0.001 | 3.190 ± 0.162 | 11.666 ± 0.027 | 3.213 ± 0.042 e | |
| 22 | 7.293 ± 0.786 a, b, c | 1.182 ± .0132 | 12.351 ± 0.015 | 3.130 ± 0.126 | 10.388 ± 0.004 | 2.902 ± 0.017 d, e | |
| 28 | 8.638 ± 0.378 b | 1.425 ± 0.076 | 11.597 ± 0.011 | 2.580 ± 1.002 | 9.683 ± 0.012 | 2.745 ± 052 c, d, e | |
|
D2 |
0 | 5.369 ± 0.133 | 0.889 ± 0.030 | 15.118 ± 0.059 | 2.271 ± 0.031 | 9.027 ± 0.067 | 1.330 ± 0.005 |
| 5 | 5.860 ± 0.480 a, b, c | 1.001 ± 0.084 c | 15.509 ± 0.926 | 2.102 ± 0.130 | 11.277 ± 0.837 | 1.556 ± 0.153 | |
| 10 | 7.027 ± 0.064 | 1.201 ± 0.014 | 15.279 ± 1.541 | 2.636 ± 0.223 | 11.801 ± 0.325 | 1.934 ± 0.041 | |
| 14 | 8.090 ± 1.618 a | 1.279 ± 0.318 | 13.923 ± 0.863 b | 2.201 ± 0.132 | 11.232 ± 0.481 | 1.792 ± 0.078 | |
| 18 | 6.960 ± 0.010 | 2.472 ± 0.206 a, d | 13.366 ± 0.032 | 2.191 ± 0.398 | 10.693 ± 1.018 e | 1.646 ± 0.160 | |
| 22 | 6.898 ± 0.031 a, b, c, d | 2.576 ± 0.335 a | 10.915 ± 0.004 | 2.962 ± 0.221 e | 10.307 ± 0.532 | 1.591 ± 0.114 | |
| 28 | 6.061 ± 0.173 | 1.097 ± 0.007 | 10.570 ± 1.227 a | 1.803 ± 0.031 a | 9.676 ± 1.158 | 1.597 ± 0.118 | |
|
D3 |
0 | 5.368 ± 0.133 a, b | 0.889 ± 0.030 | 15.118 ± 0.059 | 2.271 ± 0.076 | 9.027 ± 0.067 | 1.330 ± 0.005 |
| 5 | 5.266 ± 0.448 | 0.890 ± 0.048 | 14.496 ± 0.295 | 2.355 ± 0.226 | 11.012 ± 0.081 f | 1.791 ± 0.034 | |
| 10 | 5.086 ± 0.377 | 0.979 ± 0.080 b | 14.065 ± 1.361 | 2.181 ± 0.180 b | 10.995 ± 1.589 | 1.797 ± 0.239 | |
| 14 | 8.605 ± 0.986 b, c, d | 1.465 ± 0.124 | 14.380 ± 1.005 | 2.256 ± 0.015 | 11.224 ± 2.353 | 1.796 ± 0.446 | |
| 18 | 8.711 ± 1.313 c, d | 1.456 ± 0.149 | 16.456 ± 2.241 e | 2.352 ± 0.042 c | 11.860 ± 0.929 | 1.714 ± 0.101 | |
| 22 | 9.151 ± 0.356 a | 1.493 ± 0.052 a, b, c | 16.959 ± 0.012 | 2.360 ± 0.184 | 12.916 ± 1.963 b | 2.021 ± 0.310 | |
| 28 | 9.788 ± 1.784 a | 1.617 ± 0.257 a, c | 17.361 ± 2.708 d | 2.568 ± 0.327 | 13.443 ± 2.366 a | 2.046 ± 0.320 | |
Table 3.
Effect of variation on the stomatal conductance (gs) - M m-2 s-1, depending on wheat genotypes. Statistically significant differences at the α = 0.05 significance level are indicated by letters.
Table 3.
Effect of variation on the stomatal conductance (gs) - M m-2 s-1, depending on wheat genotypes. Statistically significant differences at the α = 0.05 significance level are indicated by letters.
| variant | days | ´Bohemia´ | ´284-17´ | ´29-17´ |
|---|---|---|---|---|
| 0 | 0.010 ± 0 a | 0.010 ± 0 a | 0.010 ± 0 a | |
| 5 | 0.138 ± 0.004 f, g | 0.142 ± 0.003 c, d | 0.220 ± 0.005 d | |
| 10 | 0.140 ± 0.003 f, g | 0.146 ± 0.006 c, d | 0.282 ± 0.005 f | |
| C | 14 | 0.097 ± 0.008 d | 0.152 ± 0.010 c, d | 0.258 ± 0.015 e. f |
| 18 | 0.097 ± 0.005 d, e | 0.126 ± 0.009 b, c, d | 0.131 ± 0.007 c | |
| 22 | 0.050 ± 0.004 a, b, c | 0.031 ± 0.002 a, b | 0.041 ± 0.002 a. b | |
| 28 | 0.087 ± 0.003 c, d, e | 0.019 ± 0.002 a | 0.070 ± 0.004 a. b | |
| 0 | 0.010 ± 0 a | 0.010 ± 0 a | 0.010 ± 0 a | |
| 5 | 0.123 ± 0.006 f, g | 0.164 ± 0.030 d | 0.231 ± 0.005 d. e | |
| 10 | 0.147 ± 0.009 e, f | 0.158 ± 0.005 c, d | 0.270 ± 0.006 f | |
| D1 | 14 | 0.015 ± 0.001 a | 0.015 ± 0.001 a | 0.015 ± 0.001 a |
| 18 | 0.160 ± 0.009 g | 0.119 ± 0.006 b, c, d | 0.217 ± 0.009 d | |
| 22 | 0.025 ± 0.002 a | 0.071 ± 0.004 a, b, c | 0.028 ± 0.002 a. b | |
| 28 | 0.042 ± 0.002 a, b, c | 0.050 ± 0.003 a, b | 0.035 ± 0.003 a. b | |
| 0 | 0.010 ± 0 a | 0.010 ± 0 a | 0.010 ± 0 a | |
| 5 | 0.138 ± 0.004 f, g | 0.142 ± 0.003 c, d | 0.220 ± 0.005 d | |
| 10 | 0.140 ± 0.003 f, g | 0.146 ± 0.006 c, d | 0.282 ± 0.005 f | |
| D2 | 14 | 0.020 ± 0 a | 0.028 ± 0.001 a | 0.024 ± 0.001 a |
| 18 | 0.251 ± 0.005 h | 0.202 ± 0.035 | 0.026 ± 0.001 a | |
| 22 | 0.036 ± 0.002 a, b | 0.031 ± 0.003 a, b | 0.026 ± 0.003 a. b | |
| 28 | 0.047 ± 0.003 a, b, c | 0.047 ± 0.003 a, b | 0.033 ± 0.003 a. b | |
| 0 | 0.010 ± 0 a | 0.010 ± 0 a | 0.010 ± 0 a | |
| 5 | 0.123 ± 0.006 e, f | 0.164 ± 0.030 d | 0.231 ± 0.005 f | |
| 10 | 0.147 ± 0.009 f, g | 0.158 ± 0.005 d | 0.270 ± 0.006 d. e | |
| D3 | 14 | 0.026 ± 0.001 a | 0.026 ± 0.001 a | 0.020 ± 0 a |
| 18 | 0.023 ± 0.002 a | 0.018 ± 0.001 a | 0.020 ± 0 a | |
| 22 | 0.042 ± 0.003 a, b, c | 0.031 ± 0.002 a, b | 0.040 ± 0.004 a. b | |
| 28 | 0.075 ± 0.003 b, c, d | 0.020 ± 0.002 a | 0.050 ± 0.003 a. b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



