
Submitted:
22 September 2023
Posted:
25 September 2023
You are already at the latest version
Abstract
According to the International Agency for Research on Cancer (IARC) of the World Health Organization (data of year 2020), Breast Cancer (BC) is one of the most common cancer types worldwide, with large geographical variations occurring between countries and world regions and highest incidence rates in countries that have undergone economic transition. The risk factors for BC include women ageing, genetic mutations, reproductive history, dense breast tissues, personal history of BC or specific non-cancerous breast diseases, family history of breast or ovarian cancer, previous treatment using radiation therapy, and exposure to hormone-like drugs such as diethylstilbestrol (DES). Additional risk factors include being overweight or having obesity after menopause, and taking hormones. MicroRNAs (miRNAs or miRs) are small non-coding RNA molecules that have an essential role in gene expression and are significantly linked to the tumor development and angiogenesis process in different types of cancer, including BC. In the last years, the knowledge about miRNAs role in BC has significantly improved, and complex interactions among coding and non-coding RNA has been elucidated. In this context, an increasing number of papers had been published regarding the role of miR-125 in BC. In this review, we summarize the state-of-the-art about this research topic in addition to elaborating on the need to set novel ethical and legal standards for the governance of such innovations in healthcare.
Keywords:
Breast Cancer (BC)
; MicroRNAs (miRNAs)
; non-coding RNA
; competing endogenous RNA (ceRNA)
; personalized medicine
; ethical and legal challenges
1. Introduction
Breast Cancer (BC) is the most commonly diagnosed cancer type, accounting for 1 in 8 cancer diagnoses worldwide. Available data suggest that incidence and mortality in high-resource countries has been declining whereas incidence and mortality in low-resource countries has been increasing [1]. In 2020, there were about 2.3 million new cases of breast cancer globally and about 685,000 deaths from this disease, with large geographical variations observed between countries and world regions. Breast cancer incidence rates are highest in countries that have undergone economic transition, but transitioning countries carry a disproportionate share of breast cancer deaths [2]. In the United States, breast cancer alone is expected to account for 29% of all new cancers in women [3]. The 2018 GLOBOCAN (Global Cancer Data) data shows that age-standardized incidence rates (ASIR) of BC are strongly and positively associated with the Human Development Index (HDI) [4].
2. Clinical features of BC
2.1. Risk factors of BC
A variety of risk factors for breast cancer have been well-established by epidemiologic studies including race, ethnicity, family history of cancer, and genetic traits, as well as modifiable exposures such as increased alcohol consumption, physical inactivity, exogenous hormones, and certain female reproductive factors. Younger age at menarche, low parity, late menopause and older age at first full-term pregnancy may influence breast cancer risk through long-term effects on sex hormone levels or by other biological mechanisms, although recent studies suggest that triple negative breast cancers may have a distinct etiology [5]. Nearly one fourth of all BC cases are related to family history: women, whose mother or sister has a BC, are more prone to this disease [6]. In the past decades the incidence of pregnancy-associated BC (PABC) increased [7]. Possible explanations are the trend to postpone childbearing and the general increase in the incidence of BC. Young patients with BC and not completed family planning should be referred for counseling about fertility preservation options before the initiation of adjuvant treatment. Both endogenous and exogenous estrogens are associated with the risk of breast cancer. The endogenous estrogen is usually produced by the ovary in premenopausal women and ovariectomy can reduce the risk of breast cancer. The main sources of exogenous estrogen are the oral contraceptives and the hormone replacement therapy (HRT) [8]. Modern lifestyles such as excessive alcohol consumption and too much dietary fat intake can increase the risk of BC as well. Alcohol consumption can elevate the level of estrogen-related hormones in the blood and trigger the estrogen receptor pathways [9].
Women that having extremely dense breast tissue is significantly associated with increased breast cancer risk compared to having scattered dense breast tissue [10]. In this scenario many patients in repoductive age and that have diagnosis of malignancies need to perform a fertility sparing surgery treatment (FST) [11,12,13,14] preserving their fertility by oocyte freezing before gonadotoxic treatments [15,16]. As known, a pregnancy following breast cancer does not have a negative impact on prognosis [17]. Women older than forty y.o. planning a family often need to recure to in vitro fertilization program (IVF) with not few psycological implication in their infertile pathway to get pregnancy [18].
Currently available data disagree on the linkage between combined oral contraceptives (COCs) and other cancers, e.g. liver malignancy risk assessments. Hormonal contraceptives have been reported to increase the risk of not only hepatocellular carcinoma but also intrahepatic cholangiocarcinoma. Still, COCs safety and effectiveness has been recognized, and their effects are reversible. While patients who choose to rely on COCs for birth control ought to be warned of the possibility of carcinogenic side-effects, it should also be pointed out to them that COCs may also decrease the risks of endometrial, colorectal, and ovarian cancers [19,20].
The inherited susceptibility to BC is partially attributed to the mutations of BC-related genes such as BRCA1 and BRCA2 [21]. Additional genes that, during the years, had been implicated in BC pathogenesis include PTEN [22], TP53 [23], CDH1 [24], STK11 [25], CHEK2 [26], PALB2 [27], ATM [28], RAD51C [29], RAD51D [30], BARD1 [31], NF1 [32], BRIP1 [33], CASP8, CTLA4, NBN and, possibly, CYP19A1, TERT and XRCC3 [34].
2.2. BC diagnosis
BC is diagnosed via ultrasonography, which is a standard imaging procedure. In recent years, it has advanced to the point that it can now be used for breast imaging. Ultrasound is a technology used as a follow-up examination to clarify equivocal findings [35,36]. Mammography screening for malignancy is commonly used to detect the disease [37]. Breast MRI (magnetic resonance imaging) is used in conjunction with mammography as a support tool [38]; in this case, MRI can be helpful in deciding whether to have a breast-conserving mastectomy or surgery [39,40]. A biopsy is done when mammograms, other imaging tests, or a physical exam shows a breast change that may be cancer. A biopsy is presently the most affordable way to diagnose cancer. Surely, CT (computerized tomography) scans, MRIs, ultrasound, and PET (positron emission tomography) scans may also give information about extension and position of cancer in the body. Laboratory tests of cancer cells (from biopsy or surgery) and blood tests can also be used to help staging some types of cancer [41]. A recently developed technique is the Digital Breast Tomosynthesis (DBT), that is a subset of the mammography procedure, with the difference that the X-ray tube rotates around a narrow angle (15–60°) from the compressed breast tissue and produces 3D breast imaging information [42]. In the same way, the Contrast-enhanced mammography (CEDM) represents the angiogenic pattern of the masses, which depicts anatomical information of the tissue. The sensitivity of this procedure is high, about 85–90%, but it has a low specificity compared to conventional mammography; also, it has limitations in the diagnosis of ductal carcinoma of the breast [43].
Tumor vascularization depends on the size and severity of the pathology. There are disorders of vascular distribution in malignant breast lesions. Contrast-enhanced ultrasound (CEUS) is used in clinical research to observe different vascular structures. CEUS uses vein injection of gas microbubbles to improve the return of the waves [44]. In ultrasonography, the color Doppler estimates the local blood flow using mean Doppler shift coding in the region of interest, and colors it. Color Doppler imaging heavily depends on technical factors. Vascular flow sensitivity can be optimized through scale reduction by power Doppler imaging (with no flow direction) and by heightening color rate. Since breast lesions are relatively superficial, the vessels can be easily closed when squeezed between the transducer and chest wall [45].
Although MRI is not commonly used for BC imaging, it is a suitable method for screening patients with a high risk of BC (20–25%) [43]. According to the American Cancer Society (ACS), high-risk patients are recommended to be screened annually using MRI imaging. The measure of high risk is for those individuals with mutations in the BRCA1 and BRCA2 genes who have a history of chest radiotherapy at ages 10 to 30 years [46]. Each of these methods has different subdivisions and their advantages and disadvantages have been discussed. Although there are ways to improve these methods, it should be kept in mind that with the simultaneous use of the capabilities of each imaging technique and their combination, early detection of BC can be significantly improved [47].
2.3. BC staging, grading and classification
The stage of a cancer gives information about its extension and spreading. In the TNM staging system, the acronym stands for tumor, node, metastasis and represents the standard classification of any cancer. The TNM staging system is the most common way to stage BC, too. Staging for breast cancer is very complex. Many different factors are considered before doctors can confirm the cancer stage, and on this basis an appropriate therapy is chosen. The TNMEIO system was suggested by the European Institute of Oncology (EIO) in an effort to include tumor characteristics affecting treatment decisions in the TNM system [48] (Table 1).
Perou and Sorlie proposed the “Molecular Classification” terminology in BC for the first time in year 2000 with a comprehensive study showing the differences in gene expression; in this study, BC was divided into different sub-groups according to various gene expression [49,50].
All breast cancers these days are tested for expression, or detectable effect, of the estrogen receptor (ER), progesterone receptor (PR) and HER2/neu proteins. These tests utilize principles of immunohistochemistry and once the status of these proteins it known, prognosis can be predicted, and appropriate novel therapies may be chosen for treatment. BC is usually, but not always, primarily classified by its histological appearance [51]. Some of the histological types include:
- Ductal Carcinoma in situ (DCIS). This is a very early form of cancer that has not spread. DCIS is a type of early BC inside of the ductal system that has not attacked the nearby tissue. It is one of the common types of non-invasive cancer.
- Infiltrating or invasive ductal carcinoma (IDC), i.e. the most widespread kind of BC. It begins in the milk ducts and can spread to surrounding tissues and eventually to other parts of the body.
- Medullary carcinoma. It constitutes roughly 15% of all BCs. Middle aged women are more likely to be affected. Cellular histology shows a resemblance to the brain medulla (gray matter).
- Lobular Carcinoma in situ (LCIS), recently renamed “lobular neoplasia”, i.e. a non-invasive and less common form of tumor. It is unlikely to turn into invasive cancer. LCIS is considered more akin to a “marker” that BC may develop.
- Infiltrating Lobular Carcinoma (ILC), i.e. the second most common type of BC after invasive ductal carcinoma. The cancer starts in the lobules or lobes and then spreads. Initial apparent thickening in the upper-outer section of the breast is usually reported. Such carcinomas are usually positive for estrogen and progesterone receptors, and therefore hormone therapy can be a valid therapeutic option.
- Tubular carcinoma. The cancer cells come to resemble tiny tubules. Women aged 50 and older are typically affected. Adjuvant systemic therapy is an effective therapeutic option.
- Mucinous carcinoma or colloid. It is a somewhat rare and invasive BC that seldom spreads to the lymph nodes. The cancer cells are known to produce mucus. Jelly-like tumors result from the combination of mucous and cancer cells.
- Paget’s disease. This leads to an eczema-like change in the skin of the nipple. Itchiness, scaling and oozing discharge from the nipple have been reported. An underlying BC is reported in 90% of patients with such symptoms have. Women in their 50s are most likely to be affected, although Paget’s disease can happen at any age.
- Inflammatory breast cancer, i.e. a rare, yet rather aggressive type of BC, which can cause the blockage of the lymph vessels in breast skin. The cancer manifests itself over a rather large area blanketing the breast, rather than a lump, and causes swelling and inflammation.
- Triple negative breast cancer. Such an BC type is known to be negative for estrogen receptor (ER), progesterone receptor (PR) and HER2/neu proteins. The acronym HER stands for “human epidermal growth factor receptor”)
- Metastatic breast cancer, i.e. a later BC stage which has metastasized to other organs (typically liver, brain, bones among others) [52].
Recently, the eighth edition of the American Joint Commission of Cancer (AJCC) staging system for BC approved major changes in the classification system, adopting an anatomy-based and histology-based subdivision built on the original TNM staging system and adding various biomarkers, to refine the prognostic information for better selection of therapy with improved outcome [53].
2.4. BC treatment
BC therapy involves a multidisciplinary approach comprising surgery, radiotherapy, neoadjuvant and adjuvant therapy. Effective therapy of BC requires maximum therapeutic efficacy [54]. There is increasing recognition that the care of a BC patient depends on highly individualized clinical features, including the stage at presentation, the biological subset of BC, the genetic factors that may underlie BC risk, the genomic signatures that advise treatment recommendations, the extent of response before surgery in patients who receive neoadjuvant therapy, and patient preferences. This customized approach to treatment requires integration of clinical care between patients and radiology, pathology, genetics, and surgical, medical and radiation oncology providers with relatively minor adverse effects and therefore better quality of life [55]. The main treatments for BC are:
- surgery
- radiotherapy
- chemotherapy
- hormone therapy
- targeted therapy
- Immunotherapy.
- Neoadjuvant systemic therapy for non-metastatic BC.
Chemotherapy, which combines DNA targeting drugs and microtubule poisons, is administrated to early, locally advanced, and metastatic BC. In the adjuvant setting, chemotherapy is used to kill remaining or spread cancer cells. In the neoadjuvant setting, chemotherapy is used to reduce tumor size and allow breast conservation.
Radiotherapy is an adjuvant treatment given to BC patients after surgery. Nearly half of all BCs are treated with radiotherapy alone or in combination with other treatments.
Neoadjuvant chemotherapy is broadly administered in early-stage breast cancer (EBC) and locally advanced BC. Such a therapeutic option can in fact result in better chances for breast-conserving surgery (BCS) by downstaging the disease and to determine tumor response to therapy [56]. Moreover, radiotherapy along with BCS has been shown to lead to higher long term overall survival rates and better local regional control, lowering local recurrence risks [57]. Therefore, patients with early stage BC, hypofractionated whole breast irradiation following breast conserving surgery now represent the standard of care based on randomized data with long-term efficacy and toxicity outcomes and postmastectomy radiation therapy represents a standard approach for patients with locally advanced BC [58].
As for surgery treatments, mastectomy remains a valid surgical alternative in selected cases and is usually associated with immediate reconstructive procedures. New surgical procedures called "conservative mastectomies" are emerging as techniques that combine oncological safety and cosmesis by entirely removing the breast parenchyma sparing the breast skin and nipple-areola complex. Unfortunately, axillary lymph node status still constitute a critical point in the planning of adjuvant treatments, and a rather reliable predictor of local disease free and overall survival. Moreover, currently no preoperative diagnostic tool can conclusively determine whether cancer cells have spread to the axillary lymphatic basins, hence there is no clarifying that point until after surgery completion [59]. Definitely, targeted therapy is the closest step to precision medicine that combines a companion biomarker with a specific treatment to select the right patient to treat. For example recently PARP (poly-ADP ribose polymerase) inhibitors are FDA-approved for people who were born with a mutation, or change, on the BRCA genes. The BC can be early-stage or metastatic and must be HER2-negative [60], immunotherapy is an emerging BC treatment. As for most treatments, immunotherapy is not beneficial to all patients, and its efficacy needs to be optimized through patient stratification and accurate predictive biomarkers.
The search for predictive biomarkers is an active field of research that includes genomic, proteomic and/or machine learning approaches. Endocrine therapy is the prevalent option for estrogen Receptors (ER)-positive BCs, since it is effective at inhibiting either estrogen production or ER signaling. Despite the high success rate of endocrine therapy, 20 to 50% of patients are reportedly resistant or develop resistance, particularly in metastatic disease. Documented resistance to endocrine therapy among ER-positive tumors seems to support the conclusion that evaluation of ER expression is not enough to reliably foresee treatment response [61]. Surely new predictive markers of sensitivity to selected therapies and innovative evaluation criteria to assess long-term treatment efficacy will be the essential prerequisites to address the biological complexity and heterogeneity of metastatic BC (MBC) [62].
3. Epigenetics of BC and the role of miR-125
3.1. microRNA: biogenesis and classification
MicroRNA (miRNA or miR) are short (20-25 nucleotides), single-stranded, non-coding RNA molecules whose main function is gene expression control, mainly silencing. They exert this downregulation by binding the 3’ end of target mRNA(s) through sequence homology and promoting either their degradation or impairing their translation [63]. It is estimated that more than 2500 miRs are encoded in the human genome, regulating over 60% of human genes [64]; in addition, thanks to imperfect sequence pairing, they also can bind multiple targets, thus amplifying their intracellular action. They map in several genomic locations, either intra- or intergenic, and in the former case they may reside inside introns, exons or regulator sequences of other genes [65], either in the sense or in the antisense strand, making their classification a complex topic [66]. Their nomenclature is complex as well and mirrors their evolution and biogenesis [67]. miRNAs are usually transcribed by RNA polymerase II (Pol II) as independent transcription units or together with their hosting genes [68]. In the first case (canonical pathway, the most common), the transcript forms a hairpin structure by folding on itself, creating what is called a pri-miRNA which may contain one or more miR precursor(s), in this case forming a cluster. Once transcribed, the pri-miRNA is processed by the microprocessor complex, containing the ribonuclease Drosha and the RNA binding protein DiGeorge Syndrome Critical Region 8 (DGCR8), which cleaves it to produce the precursor-miRNA (pre-miRNA). The pre-miRNA is then exported to the cytoplasm trough the action of Exportin5(XPO5)/RanGTP complex and further processed by the RNase III endonuclease Dicer to produce the mature miRNA duplex. Finally, either the 5p (at the 5’ end) or 3p (at the 3’ end) strands of the mature miRNA duplex is loaded into the Argonaute (AGO) family of proteins to form a miRNA-induced silencing complex (miRISC); notably, sometimes both strands – separately – can be used in this process, thus the proportion of AGO-loaded 5p or 3p strand varies greatly depending on the cell type, the cellular environment and on the biochemical characteristics of the sequence [69]. Multiple, non-canonical pathways of miRNA biogenesis exist as well [67]. These pathways may act of pri-miRNA either transcribed independently, or on those contained inside the introns/exons of other genes and generated during the splicing process (these splicing-derived miRNAs being called mirtrons) and can be broadly subdivided in Drosha/DGCR8-independent and Dicer-independent pathways; sometimes, importin 1 is used for their export in the cytoplasm [67]. However, in the end they all are loaded into the miRISC complex to proceed to their final maturation and exert their function.
Beyond the number identifier (usually higher for miRNA described chronologically later) additional nomenclature rules are established to identify unequivocally each miR [70,71]. miRNAs with almost identical sequence are identified by a progressive lowercase letter after the identification (ID) number (miR-XXXa, miR-XXXb, etc) while miR with identical sequence but mapping in different genomic locations are indicated by a progressive number separated from the ID number by a dash (i.e., miR-XXX-1, miR-XXX-2, etc). To further distinguish miRNAs of species, an additional three-letter code and a dash may be added at the beginning of the miR name (i.e., hsa-miR-XXX indicates a human -Homo sapiens – miR). Finally, the 5p and 3p notation described above is used when the two miRNAs are roughly equivalent in their intracellular amount; if one is significantly more abundant than the other, then the rarer has an asterisk at the end of its name (for example, miR-XXX-5p*).
3.2. The miR-125 family: molecular organization and roles in human pathology
miR-125 is a highly conserved family of microRNAs whose members had been found also in nematodes (named lin-4 in 1993, the first miR described ever) [72]. The miR-125 family in H. sapiens includes three members, namely miR-125a, miR-125b-1 and miR-125b-2. MIR125A gene maps to chromosome 19q13.41 [73] and miR-125a is part of a cluster together with miR-99b and let-7e [74]. MIR125B1 gene maps to chromosome 11q24.1 and in this locus, it is part of a cluster including LET7A2 and MIR100 genes [74,75]; these miRNAs are inside the third intron of the MIR100HG gene [76]. Finally, MIR125B2 gene maps to chromosome 21q21.1 where it is included in a cluster together with the MIR99A and LET7C genes [74,75], inside intron 6 of MIR99AHG gene [76]. miR-125a and miR-125b differ only by a central diuridine insertion and a U-to-C change in miR-125a [77]. All members of the family show both 5p and 3p forms (Figure 1).
The miR-125 family is involved in several cell metabolic pathways controlling differentiation, proliferation, apoptosis, metastasis formation, drug resistance and immune system function because of targeting of messenger RNAs related to these cellular processes [80]. As described for miRNAs in general (see previous section), also miR-125 molecules have a complex behavior inside the cell, which mirrors their expression pattern in different tissues/cell types [78,79], their ample variety of targets [78,79], the intracellular role of their targets, and the way miR and mRNAs are either up- or down-regulated upon expression.
In the muscle, miR-125b action on insulin-like growth factor II (IGF-II) negatively regulates myoblast differentiation in vitro and muscle regeneration in vivo [81], while overexpression of miR-125b-5p protects skeletal muscle from atrophy in vitro and in vivo by targeting TRAF6 through inactivation of several ubiquitin-proteasome system (UPS)- and autophagy-lysosome system (ALS)-related proteins [82]; interestingly, the same interaction between miR-125b-5p and TRAF6 plays a critical role also in allergic asthma by modulating macrophage polarization [83] and in the myocardium, where it protects from ischaemia/reperfusion injury by preventing p53-mediated apoptotic signaling and suppressing TRAF6-mediated NF-κB activation [84]. The same miR is also involved in smooth muscle phenotype [85] and in the proliferation and migration of vascular smooth muscle cells induced by platelet-derived growth factor BB by regulating the expression levels of AAMP (angio-associated migratory cell) and SRF (serum response factor) proteins [86]. Also, their role in cardiac muscle is well described in the literature. Beyond the above mentioned work [84], miR-125 family action is related to cardiomyocytes and the development of heart in embryonic mammals (reviewed in [87]); it regulates muscle-enriched transcription factors in cardiac and skeletal myocytes [88]; it can modulate cardiac progenitor cells proliferation and migration potential through the MALAT1/JMJD6 axis in hypoxia [89] while in the HOTAIR/MMP2 axis miR-125 regulates cardiomyocytes proliferation and apoptosis under oxidative stress conditions [90]; it has been recently shown that cardiac-specific miR-125b deficiency induces perinatal death and cardiac hypertrophy [91]. Also miR-125a plays roles in cardiac muscle, by improving cardiac function and fibrosis and reducing inflammatory responses in DCM (diabetic cardiomyopathy) mice, although these data need further validation in man [92].
miR-125 is one of the most abundant microRNAs in the central nervous system (CNS) in mice and men [93]. In man, miR-125b promotes neuronal differentiation in human cells by repressing at least ten target mRNAs involved in those pathways – although in the same work the authors report a list of 164 deregulated genes, affecting several pathways [94] – and plays a key role for cocaine-induced PARP-1 expression (miR125 downregulation, inducing PARP-1 upregulation) [95]. It also regulates dendritic spine morphology and synaptic maturation [96]; it is implicated in synaptic plasticity [97]; it regulates neural stem/progenitor cell proliferation by targeting Musashi1 [98]; it promotes astrogliogenesis [99] and is involved in astrogliosis and glial cell proliferation [100]. miR-125b deregulation is linked to several brain disorders including Down syndrome [101], acute ischemic stroke [102,103] and Alzheimer’s disease [104,105]. Recently, it has been shown that miR-125b-2 is imprinted in human but not mouse brain, where it regulates hippocampal circuit and function, and affects the hippocampal expression of Grin2a and NMDAR-mediated currents [106]. Its deregulation has been linked also to CNS tumor formation and growth, such as pediatric low-grade glioma [107], where it regulates cell growth and invasion; it regulates cell growth arrest and apoptosis of human neuroblastoma- and medulloblastoma-derived cell lines [108,109]; it inhibits cell apoptosis through p53 and p38MAPK-independent pathways in glioblastoma cells [110] and in glioma it targets BMF [111]. Finally, miR-125a-5p is up-regulated in myasthenia gravis (an autoimmune disease) thymuses [112].
In the immune system, miR-125 members regulate hematopoiesis, inflammation, and immune cell function. miR-125a controls stem cell homeostasis during hematopoiesis [113,114,115]; miR125 plays a role in immune cell identity [116]; miR-125b potentiates macrophage activation [117] and regulates their differentiation and polarization [118]; it is downregulated during bacterial infections (reviewed in [119]). It is involved in periodontitis as well [120] and some authors report that miR-125a-5p modulates macrophage polarization by targeting E26 transformation-specific variant 6 gene during orthodontic tooth movement [121]. miR-125-5p targeting IL-6 R regulates macrophage inflammatory response and intestinal epithelial cell apoptosis in ulcerative colitis through JAK1/STAT3 and NF-κB pathway [122]. miR-125b-1-3p is expressed in hMSCs-Ad exosomes and can promote T lymphocyte apoptosis and alleviate atherosclerosis (AS) by down-regulating BCL11B expression, thus providing potential molecular targets for the clinical treatment of AS [123].
3.3. miR-125 and cancer
One of the most studied topics is the role of miR-125 in cancer and beyond the above-mentioned tumors of the CNS, it is deregulated in several other tumors [124]. miR-125 is involved in neointimal hyperplasia [125]. In ovarian cancer cells Dahl and collaborators showed that overexpression of miR-125a induces conversion of highly invasive ovarian cancer cells from a mesenchymal to an epithelial morphology, suggesting miR-125a is a negative regulator of epithelial-to-mesenchymal transition (EMT) [126], while another study showed that ectopic expression of miR-125b in ovarian cancer cells induced cell cycle arrest and led to reduction in proliferation and clonal formation through the inhibition of the translation of BCL3 proto-oncogene mRNA [127]. Recently, a study on the same tumor cells revealed that serum miR-125b levels are a useful diagnostic biomarker to predict the responses to chemotherapy in patients, its expression being associated with FIGO stage, lymph node metastasis, and chemoresistance [128]. Huang and colleagues showed that miR-125b may act as a tumor suppressor in bladder urothelium by targeting E2F3, and its downregulation may contribute to tumorigenesis [129]. Use of miR-125b as a biomarker of bladder cancer in urine supernatant has been reported as well [130]. In the same tumor, hypoxia-regulated miRNAs include miR-125-3p [131]. miR-125 is also included in a panel of 8 miRNAs highly significant and consistently dysregulated for predicting survival in bladder cancer [132]. In hepatocellular carcinoma (HCC) it has been shown that the ectopic expression of miR-125a inhibits proliferation and metastasis by targeting MMP11 and VEGF [133], while others showed that ectopic expression of miR-125b reduces the cellular proliferation and cell cycle progression of HCC cells by targeting Mcl-1 and IL6R [134]. In another study the authors showed that miR-125b suppresses human liver cancer cell proliferation and metastasis by directly targeting oncogene LIN28B2 [135]. Additional miR-125 interactors in HCC had been identified in the years, including Pokemon [136], TRAF6 [137], hexokinase II [138] and FOXM1 [139]. As for the skin tumors, in melanoma, miR-125 controls the expression of NCAM [140] and c-Jun [141]; in cutaneous squamous cell carcinoma miR-125b controls the expression of MMP13 [142] and STAT3 [143]; in basal cell carcinoma, it is one of the most deregulated miRNAs [144]. The same altered pathway (miR-125b/STAT3) is also involved in osteosarcoma development [145,146] , and in the same tumor additional pathways affected by miR-125 deregulation include ErbB2 [147] and BAP1 [148]. In lung cancer, miR-125 mutations significantly affect the survival of non-small cell lung cancer (NSCLC) patients [149] and its interactors include EGFR [150], HER2 [151] and MMP13 [152]. In pancreatic cancer miR-125 is deregulated as well [153,154] and NEDD9 is one of the recognized targets [155]. Also in prostate cancer miR-125 is deregulated [156,157,158] and its targets comprise BAK1 [159,160], p53 and PUMA [160]. A role of these miRNAs has been ascribed also in thyroid cancer, where it has been shown that miR-125b could directly act on Foxp3 expression by binding to its 3' UTR mRNA and inhibiting its expression; overexpression of miR-125b markedly sensitizes thyroid cancer cells to cisplatin treatment by inducing autophagy through an Atg7 pathway in vitro and in vivo [161]. In gastric cancer it has been reported that miR-125b promotes cell migration and invasion by targeting PPP1CA-Rb signal pathways, resulting in a poor prognosis [162]. In colon cancer miR-125a-5p inhibits cell proliferation and induces apoptosis via targeting BCL2, BCL2L12 and MCL1 [163]. The use of miR-125b as a marker in clear-cell renal cell carcinoma has been explored as well, with good results [164]. Hematological malignancies are also a possible byproduct of miR-125 dysregulation; due to the ample literature available, we redirect the reader to specific and comprehensive reviews [74,80,165,166].
3.4. Role of miR-125 in BC
A relatively large amount of research findings are available regarding the role of miR-125 in BC. The reports showing altered expression of miRNAs in BC started to be published more than 20 years ago and the research is still running in search of an affordable diagnostic panel [167,168,169,170,171,172], and among the first targets recognized, we recall here ERBB2 and ERBB3 [173]. In 2011 Zhang and colleagues demonstrated the action of miR-125b on the regulation of the ETS1 proto-oncogene in human invasive breast cancer [174]. Rajabi et al. found that miR-125b, downregulated in BC, can reduce the expression of MUC1 (an oncoprotein), whose silencing causes DNA damage-induced apoptosis in cancer cells [175]. Tang and collaborators studied the effects of miR-125 deregulation on metastasis formation, finding that miR-125b induces metastasis by targeting STARD13 in MCF-7 and MDA-MB-231 breast cancer cells [176], in contrast with the tumor suppressive action described before. Using the same BC cell lines, Metheetrairut and collaborators showed that forced expression of miR-125b results in radiosensitivity, as seen by reduced clonogenic survival, enhanced apoptotic activity and enhanced senescence post ionizing radiation treatment. Moreover, re-expression of c-JUN in MDA-MB-231 cells promoted radioresistance and abrogated miR-125-mediated radiosensitization, suggesting that overexpression of miR-125b causes sensibilization to γ-irradiation, indicating this miRNA as a possible target for adjuvant therapy [177].In contrast, Wang and co-worker found an association between miR-125b expression and chemoresistance [178], again indicating an oncogenic role for this miRNA. In line with these last results, Zhou and collaborators found that miR-125b confers the resistance of BC cells to paclitaxel through suppression of pro-apoptotic Bcl-2 antagonist killer 1 (Bak1) expression [179]. He and collaborators studied the expression of miR-125a-5p/3p and miR-125b in 143 pairs of BC and normal adjacent tissues finding that miR-125a-5p and miR-125b were significantly down-regulated in BC tissue samples and that the expression level of miR-125a-5p was significantly higher in younger patients (<35 years) than in older ones, with a gradual reduction in miR-125a-5p expression was observed in BC tissue samples correlated to increasing age [180]. Recently, a paper showed the oncosuppressor role of miR-125b via the inhibition of proliferation, migration, and invasion of BC cells through targeting MMP11 protein [181].
3.5. Further mining miR-125 function in BC: competing endogenous RNA networks (ceRNETs).
A fundamental way to control gene expression through miRNAs has been elucidated in recent years, consisting in the so called ceRNETs. It has been shown that miRNAs are controllers of target mRNAs by altering their half-life or translation. However, they are also controlled, in many cases, by other long non-coding RNAs (lncRNA), or even other mRNAs, which “sponge” miRNAs through sequence homology, avoiding their interaction with mRNA targets [66]. In other words, lncRNA and mRNA compete for binding miRNAs; these two molecules form a competing endogenous RNA (ceRNA) couple. If the lncRNA efficiently sponges the miRNA, then miRNA inhibitory action is not accomplished, and the mRNA is regularly translated. In this case, the lncRNA (inhibitor of an inhibitor) has a function resembling that of an enhancer of gene expression. Hence, if the mRNA encodes an oncoprotein, the lncRNA has an oncogenic effect, while the miRNA has an oncosuppressive role. The same, with opposite effects, occurs in case of the mRNA coding an oncosuppressor. The three molecules, taken together, form what is currently known as a regulatory axis, and the sum of many axes creates the ceRNET. In the ceRNETs, lncRNAs and mRNAs constitute the nodes of the network, while the miRNAs represent their connections. An increasing number of papers has been published in recent years depicting the increasing structure and complexity of the ceRNET in BC (see [182] and references therein), including the action of pseudogenes in this phenomenon. In fact, Welch and collaborators found that 309 pseudogenes exhibit significant differential expression among breast cancer subtypes and their expression pattern allows recognizing tumor samples from normal samples and discriminates the basal subtype from the luminal and Her2 subtypes; of them, 177 transcribed pseudogenes possess binding sites for co-expressed miRNAs that are also predicted to target their parent genes [183]. Recently, in a work by Zhu and collaborators the authors took advantage of the data available in exoRbase database and derived from exosomes of human BC samples [184]. Their study allowed to identify a ceRNA network including 19 mRNA nodes, 2 lncRNA nodes, 8 circular RNA nodes, and 41 miRNA connections. KEGG enrichment analysis showed that differentially expressed mRNA in the regulatory network are mainly enriched in the p53 signaling pathway.
Literature describing portions of a miR-125-centered ceRNET is constantly growing. It is likely that the miR-125 interactions with mRNAs described in the previous section will become axes of the growing BC ceRNET as well, as soon as the appropriate lncRNA is identified in the pathway. However, some axes have been already described and some of them, being interconnected, can be used to build a basic version of this network (Figure 2).
In 2004 Rieger and collaborators discovered a new human cytochrome P450 (CYP), termed CYP4Z1, which is specifically expressed in mammary gland and breast carcinoma [185]. They also found a transcribed pseudogene, named CYP4Z2P, that codes for a truncated CYP protein (340 amino acids vs. 505) with 96% identity to CYP4Z1. Both CYPs are highly expressed in breast cancer, although the expression level of CYP4Z2P is approximately 20 times lower than that of CYP4Z1 in mammary tissues, and barely expressed elsewhere. Later it was shown that increased expression of CYP4Z1 promotes tumor angiogenesis and growth in human breast cancer [186] and that CYP4Z2P 3’-UTR is involved in promoting breast cancer angiogenesis through the VEGF/VEGFR2 pathway [187]. In 2015 Zheng and collaborators showed that the action of CYP4Z2P 3’-UTR is sponging several miRNAs, including miR-125a-3p, and that this pseudogene acts as a ceRNA in respect to CYP4Z1, enhancing its expression levels [188,189]. They also showed that tumor angiogenesis-is promoted by inducing the phosphorylation of ERK1/2 and PI3K/Akt pathways. The same group also showed later that the deregulation of these ceRNA also confer tamoxifen resistance in BC through the enhancement of the transcriptional activity of ERα via its phosphorylation dependent on cyclin-dependent kinase 3 (CDK3) [190]; that downregulation of CYP4Z1 or the CYP4Z2P 3'-UTR promotes cell apoptosis, mirroring the functions and modulating the expression of human telomerase reverse transcriptase (hTERT) [191]; and that the transcriptional factor six2 activates these CYPs ceRNET by directly binding to their promoters, thus activating the downstream PI3K/Akt and ERK1/2 pathways and consequently being involved not only in chemoresistance, but also regulating the stemness of BC cells [192,193].
STARD13 (StAR-related lipid transfer domain protein 13, also known as deleted in liver cancer 2 protein (DLC-2)) is a Rho GTPase-activating protein (GAP) that selectively activates RhoA and CDC42 and suppresses cell growth by inhibiting actin stress fiber assembly in hepatocellular carcinoma (HCC) [194]; this protein is ubiquitously expressed in normal tissues and downregulated in HCC. In mice, STARD13, promotes angiogenesis through the actions of RhoA [195]. Its role is well established in BC as well, where it act as a tumor suppressor gene [196], where it regulates cell motility and invasion [197], endothelial differentiation [198], metastasis formation [199,200], cell migration [201], and apoptosis [202]. It has also been shown that STARD13 exerts its function in BC through its participation to ceRNETs, such as the one involving positive TGF-β/miR-9 regulatory loop mediated by the STARD13/YAP axis [203], or the one involving hsa-miR-21-3p [204], or even the more complex network which involves 5 different miRNAs and that controls YAP/TAZ nuclear accumulation and transcriptional activity via modulation of Hippo and Rho-GTPase/F-actin signaling pathways [205]. A direct link between miR-125 and STARD13 has been described, too. Li and coworkers showed that CDH5, HOXD1, and HOXD10 are putative STARD13 ceRNAs and display concordant patterns with STARD13 in different metastatic potential breast cancer cell lines and tissues; in addition, they also show that the 3’ UTR of STARD13 mRNA can bind miR-125b (and also miR-9 and miR-10b, indicating that this mRNA may target multiple pathways simultaneously) [206], thus confirming their previous study about this interaction [207] and showing that the transcripts of the tumor suppressors genes CDH5, HOXD1, and HOXD10 inhibit BC metastasis in vitro and in vivo by competing with STARD13 mRNA for these three miRs. Interestingly, as mentioned before [205], CDH5, HOXD1 and HOXD10, together with STARD13, are BC players also in a different ceRNET, competing for a different set of miRNAs, indicating that STARD13 role in BC is very complex. In 2017 Hu and collaborators discovered another ceRNET axis in which STARD13 and miR-125b control the CCR2 (cysteine–cysteine chemokine receptor 2) expression levels [208]. In this case, they found that the CCR2 3' UTR harbors three miR-125 binding sites that both inhibits MDA-MB-231 and MCF-7 cell metastasis by repressing epithelial-mesenchymal transition (EMT) in vitro, and suppresses breast cancer metastasis in vivo, through the competition with STARD13 in a miR-125b-dependent and protein coding-independent manner. Another component of the same ceRNET is TP53INP1 (tumor protein p53-inducible nuclear protein 1). TP53INP1 is an antiproliferative and proapoptotic protein involved in cell stress response which acts as a dual regulator of transcription and autophagy and is modulated by p53 in response to stress; it also interacts with kinases HIPK2 and PKCδ, which phosphorylate p53, creating a positive feedback loop between p53 and TP53INP1 [209]. TP53INP1 is also involved in SPARC (secreted protein acidic and rich in cysteine) mediated-promotive effects on cancer cell migration and metastasis [210]. In 2018 Zheng et al. found a ceRNA interaction between STARD13 and TP53INP1 mediated by competitively binding to miR-125b in BC [211]. In this case STARD13 promotes upregulation of TP53INP1, causing the inhibition of BC cell metastasis through competitively binding to miR-125b thanks to the inhibition of SPARC gene expression. Later, Guo and co-workers also found a ceRNET axis in BC involving miR-125b, STARD13 and BMF (Bcl-2-modifying factor) [212]. BMF is a member of the BCL2 protein family and controls apoptosis in several cell types [213]. The authors [212] found that miR-125b directly binds the 3’ UTR, and thus downregulates, BMF expression, and that STARD13, sponging miR-125b, upregulates BMF in BC both in vitro and in vivo. All together, these results may suggest novel therapies for BC treatment and aiding in selecting adequate drugs, depending on the molecular biology of the tumor, in perspective aiming at the goal of personalized medicine. Indeed, a recent work showed that tanshinone IIA (an effective component extracted from Salvia miltiorrhiza which regulates the stemness of tumor cells) attenuates this phenotype in BC cells, by downregulating miR-125b levels and upregulating its target gene STARD13, while miR-125b overexpression or STARD13 knockdown impairs the inhibitory effects of tanshinone IIA on the stemness of BC cells. [214].
3.6. Beyond clinical factors: new standards for a legally and ethically tenable implementation of personalized medicine
The broad-ranging and potentially revolutionary scope of personalized medicine has been well established and acknowledged universally. The notion of personalized medicine can be summarized as the medical model relying on characterization of individual patients’ phenotypes and genotypes (e.g. the centerpiece of this article applied to breast cancer, molecular profiling, medical imaging, lifestyle data) for the ultimate purpose of configuring and modulating the most effective therapeutic pathways for any given individual patient with the right timing; personalized medicine as a prognostic avenue can identify and assess the predisposition to disease in order to delineate timely and individually outlined prevention strategies [215]. Such practices are extremely promising as far as the development of novel therapeutic avenues options is concerned, tailored to each patient’s genetic make-up for maximum effectiveness. Such a breakthrough however is bound to give rise to a host of challenges from the legal, ethical, social and policy-making perspectives in order to guarantee the necessary standards of fairness, equitable access and non-discrimination among others [65,216]. New criteria need to be developed for the effective and equitable governance of personalized medicine, which must take into account key aspects such as individual privacy, the risk of stratification and discrimination of sub-populations segments on the basis of ethnicity, equal access opportunities, and how to ensure oversight for the fair allocation of funding [217]. As personalized medicine practices expand and further develop, they are likely to become increasingly mainstream, which will make ethical, legal and social implications all the more urgent to deal with, in order to guarantee te ethically sound implementation of such a major scientific breakthrough, which in conjunction with other technologies such as big data analysis and artificial intelligence has the potential to change healthcare at its very core [218]. Major European institutions have proven fully aware of such looming changes. The potential of personalized medicine has in fact been recognized in two highly relevant policy papers, focused on oncological diagnostics or therapeutics, advocating for personalized medicine to be fully harnessed and gradually implemented on a large scale for the sake of patients [219,220], despite the lingering difficulties linked to the still high level of uncertainty regarding outcomes [221]. The EU papers outline the key factors that can be instrumental in fulfilling the potential of personalized medicine, primarily a thoroughly structured, synergistic conjunction of health data and new technologies, in order to lay the groundwork for the comprehensive and well-balanced development of personalized medicine techniques [222]. The considerable potential of personalized and precision medicine to offer valuable oncological treatment options will be fulfilled through the elaboration of prevention and treatment pathways which will allow patients and doctors to identify the best-suited and most effective therapeutic course. Overall, such novel tailored approaches are likely to benefit healthcare systems to a substantial degree too, especially by reducing wasteful spending on trials and ineffective treatments and streamlining resource allocation more efficiently. In 2022, the EU plans took it upon itself to put in place new measures meant to make the most out of new innovative technologies based on AI, big data, and genomics. It is worth mentioning in that regard the so-called European Cancer Imaging Initiative, designed to implement and optimize the delivery of personalized medicine and innovative approaches through the application of new computer-aided tools [223,224]. Also deserving attention is the new Partnership on Personalized Medicine (also known as EP PerMed), phased in 2023 and funded by the EU’s chief funding program aimed at fostering research and development, Horizon Europe, which can rely on a budget of €95.5 billion. Among its priorities, EP PerMed is set to find common ground projects for public and private sector stakeholders, in order to define key objectives to direct research funding, harmonize the current broad range of personalized medicine approaches and strategies, develop new effective training blueprints, and support policy-makers. Just as importantly, it is considered a priority to provide support for longer life expectancy by increasing healthy life years and to contribute to guarantee the sustainability of each member state’s healthcare system [225].Through a multidisciplinary concerted effort, EP PerMed will ultimately outline fundamental priorities to further research and education in personalized medicine, in addition to fostering cancer prevention, diagnosis, and therapeutic research endeavors. Rationalizing the implementation of personalized medicine by defining the roles of all parties involved in research, policy-making, industry and society at large will be vital for the purpose of ensuring such novel techniques and processes do not lose sight of the interests and rights of everyone, thus guaranteeing an ethically viable use for maximum benefit.
Table 2 describes the stakeholders at play and the expected outcomes for the harmonized development of personalized medicine initiatives in the European Union
It will be ultimately essential to be able to rely on a clean-cut a set of criteria for the definition, delivery and oversight of personalized medicine-based approaches to be transposed in clinical and medical research settings. Other than the EU initiative, it is noteworthy how such a set of goals had already been acknowledged by the International Consortium for Personalized Medicine (ICPerMed), started in November 2016 [226,227]. The ICPerMed has laid out and described quite thoroughly the distinctive traits and features of personalized medicine and what it will finally come down to: the natural evolution of healthcare in the dawning age of biotechnology and big data. Such a broad-ranging breakthrough however certainly requires comprehensive adjustments and restructuring in the very modalities and processes through which healthcare is delivered. To that end, large-scale implementation calls for emphasizing training and new sets of skills for healthcare professionals, which will be able to rely on novel means and instruments to deliver care to their patients [228]. The path undertaken by the ICPerMed has been aided and supervised by European and international expert and specialist consultants in key research areas, who have greatly contributed to defining and weighing the opportunities and pitfalls posed by personalized medicine and identifying any particular concerns and management challenges [229]. To that end, as a preliminary step towards launching the partnership, the European Commission is set to outline a road map laying out in detail tailored preventive strategies and approaches [230]. The very real possibility that data may in all likelihood change healthcare at its core has been recognized and assessed by established European policies, both at the individual patient level and insofar as healthcare systems are concerned (the European Alliance for Personalized Medicine has issued in that respect a rather compelling report titled “Cooperating on Data: The Missing Element in Bringing Real Innovation to Europe’s Healthcare System” [231]). It is in fact worth considering that clinical studies, diagnostic results, medical records and patient information are but a part of the data sources available in healthcare. Completing the full transition towards the digitization of patient records is poised to be a major contributing factors in this highly consequential breakthrough. Comprehensive digital archives from which to collect and elaborate big data will be of utmost importance to that end. Data drawn from digital applications, wearable devices, and social media, will also be instrumental at providing information on environment- and lifestyle-related factors, socio-demographics, genomics, metabolomics, proteomics, radiomics, standardized electronic health records, or precision medicine platforms [232,233]. In order to outline an ethically and legally sound path towards the large-scale implementation of personalized medicine, it is necessary to streamline and optimize key aspects suchas the management of biobanking and informed consent, confidentiality [234], access to treatment, clinical translation, and direct-to-consumer genetic testing. In addition, the stratification and genetic discrimination of sub-populations based on ethnicity will have to be prevented through the introduction of targeted measures [235,236]. Other hurdles which have been pointed out and need to be addressed have to do with the insufficient level of genetic literacy and substandard comprehension of personal and familial implications of germline and somatic genomic testing among patients, which may negatively affect the informed consent process [237]. An element of inequality may arise from each patient’s socioeconomic status, healthcare insurance policy (or level of coverage by the national healthcare system), and cancer care facilities [238]. Even though patients residing in countries with publicly funded universal healthcare are less likely to be affected by access inequalities, such systems often limit public coverage to treatments and techniques of already established efficacy [239]. In addition, seeking the highest possible degree of harmonization will prevent “healthcare tourism” practices, i.e. patients with adequate financial means traveling to countries where any given procedure is legally available [240,241]. Personalized medicine and its wide-ranging clinical applications will have to rely on full validation in terms of their efficacy and safety, and such a process will likely need larger study samples compared to more conventional procedures whose clinical value has already been acknowledged. That in turn may mean that personalized medicine techniques and therapeutic approaches may take longer to be acknowledged as fully evidence-based and approved for large-scale use [242]. Such implications can negatively affect the fair and equitable allocation of healthcare funding. The already mentioned European Union Communication issued in 2022 [219] in fact points to legal and ethical criteria as potentially difficult barriers to overcome, for the purpose of harnessing personalized medicine and the European Digital Strategy to their full capabilities. That principles holds true for many different innovations which open up new prospects, for instance, for transplant patients through regenerative medicine [243,244]. Lawsuits arising from negligence and malpractice charges [245] will probably become increasingly common as personalized medicine techniques do as well [246]. That however entails that medical procedures grow more complex, and as they do, the risk of litigation resulting from alleged medical malpractice will be ever more substantial [246]. The parties that could be held liable in such instances are not only the healthcare professionals implementing diagnostic and therapeutic actions, but range from manufacturing firms of genome sequencers and medical devices, laboratories, pharmaceutical companies, and healthcare facilities. The legal concept at the core of “genetic malpractice” is in fact the doctors’ failure to identify or accurately interpret genetic testing. Such error-prone dynamics can be complicated even more by the lingering differences in the medical research community about the values, scope and timing of genetic testing when defining prognostic and therapeutic pathways, or whether such testing should be considered and carried out at all, and under what circumstances [227,246,247]. Research findings and case-law [245,246,247] are still inconclusive at this stage as to how and to what extent these new litigation profiles linked to clinical genomics-based personalized medicine will impact healthcare professionals and facilities. A major factor in determining how such legal actions end is surely the specific set of conditions, environmental and contributing factors specific to each case, in addition to the legal strategies chosen by plaintiffs, counselors, expert witnesses, and magistrates [248]. It appears therefore quite plausible that judicial decisions at first will have a major impact on the future incidence, sustainability, and frequency of such actions. Medical and legal analysts will in fact weigh such rulings and adjust their management and use of such techniques accordingly, by putting in place all necessary safeguards [249,250]. Current uncertainty makes it all the more urgent to rely on harmonized and widely agreed upon legislative, regulatory, and policy criteria, to be incapsulated in evidence-based guidelines and best practices specifying when and where genetic testing may be advisable, or even necessary, and when it is not (at least with the current level of knowledge) [251]. Such a degree of objectivity will provide valuable guidance based on sound scientific and clinical elements which will inform court decisions, thus protecting physicians and healthcare professionals from malpractice lawsuits which are often frivolous and harm the quality of care for everyone [252].
5. Conclusions
BC is a heterogeneous disease, thus patients that are histologically diagnosed with the same cancer type, might have different molecular characteristics, genetic mutations or tumor microenvironments that can deeply influence prognosis or treatment response. Consequently, the challenge in personalized medicine is to distinguish these diverse molecular characteristics, separate patients accordingly, and treat them using a tailored approach which considers all these features. Personalized medicine might profoundly improve patient outcomes thanks to diagnostic tests that can identify specific biomarkers, allowing doctors to select the most effective treatment for each patient, reducing the risk of adverse reactions and increasing the likelihood of a successful outcome. In this context, miR-125 and its targets are emerging as promising biomarkers in BC classification.
It is interesting to remark that, depending on the study, miR-125 has been described having either oncogenic or oncosuppressive role. However, this should not be surprising because of at least three reasons: cell changes in space and time, and miR regulation. First, different cancers – and sometimes also different stages of the same cancer, or even different populations of the same tumoral mass – have different metabolic needs [253,254]. So, in presence of both the same miRNA and corresponding mRNA target, the effect on the tumoral cell metabolism may significantly vary, allowing different spatial effects of miRNAs on the transcriptome. Second, the evolution of cancer during time is associated with alterations in the cancer cell proteome; new genes are activated, other genes are suppressed or lost (for example through aneuploidy [255,256] or copy number variations)[257,258,259,260] also as a response to the internal and external microenvironmental interactions [258,259,260]. This is obviously true also for every single cancer subpopulation (the space variable described before). Third, we recall here the organization of the miR-125b genes: this miRNA is transcribed from two different loci in the genome, which are under two different promoters[74], thus it is quite straightforward to hypothesize that the two copies are not fully interchangeable in their function despite their sequence identity, and they may be transcribed under different cellular conditions; consequently, different targets may be present at the time of miR-125 expression. In addition, the two clusters miR-125b is embedded in, are different; the transcription of the cluster as a single pri-miRNA using the same promoter suggests that these diverse miRNAs need to act in concert, thus the two different clusters, upon expression, likely modify the host cell proteome in different ways. As a consequence, it is possible to theorize that inconsistencies in the role of miR-125 (oncogene vs. oncosuppressor) might just reflect differences in the analyzed cells or in the used analytical protocols, more than real contradictions, thus further pushing researchers and physicians to characterize each patient at the molecular level. Indeed, some clues for this possibility have been showed for BC cell lines used to verify miR-125 function, where it has been shown that miR-125 is expressed in MCF-7 spheroids but not in MDA-MB-231 spheroids and that the unique cluster of miRNAs found in each cell type are associated with their chemoresistance properties and cancer progression, and most likely influencing the maintenance of the spheroid-enriched cancer stem cells properties [258]. Similar differences in different BC cell lines have been reported also by Ahram et al. who compared MDA-MB-453, MCF-7 and T47D cells finding that miR-125b is highly expressed in T47D cells and slightly downregulated in MDA-MB-453 cells, with all the predictable consequences related to their targets fate [261]. All together, these data underline the importance and complexity of the expression of miR-125 family members in the etiopathogenesis of BC, and the need to further and better characterize this biomarker in BC patients. Personalized medicine undoubtedly constitutes a broad-ranging breakthrough with huge potential to change healthcare to its very core. At the same time, such a potential constitutes such a breakthrough that the current sets of ethical, legal and policy-making standards which provide us guidance today may be outpaced and ultimately inadequate. New criteria therefore need to be devised if we are to rely on equitable, effective healthcare for all.
Author Contributions
Conceptualization, R.P., G.G. and S.Z.; methodology, R.P., G.G., G.C., A.L.; validation, A.L., G.N. and S.Z.; formal analysis, R.P., G.G., G.C., S.Z.; investigation, R.P., G.G., G.C., A.L. and E.M.; resources, R.P., G.G., G.C., G.N. and S.Z.; data curation, R.P., G.G., G.C., S.Z.; writing—original draft preparation, R.P., G.G., G.C., G.N., E.M. and S.Z.; writing—review and editing, R.P., G.G., G.C., A.L. and S.Z.; visualization, R.P., G.G., G.C., G.N., E.M. and S.Z.; supervision, R.P., G.G., E.M. and S.Z.; project administration, R.P., G.G., G.C., G.N., E.M. and S.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
None.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Winters, S.; Martin, C.; Murphy, D.; Shokar, N.K. Breast Cancer Epidemiology, Prevention, and Screening. In Progress in Molecular Biology and Translational Science; 2017; Vol. 151.
- Arnold, M.; Morgan, E.; Rumgay, H.; Mafra, A.; Singh, D.; Laversanne, M.; Vignat, J.; Gralow, J.R.; Cardoso, F.; Siesling, S.; et al. Current and Future Burden of Breast Cancer: Global Statistics for 2020 and 2040. Breast 2022, 66. [Google Scholar] [CrossRef] [PubMed]
- DeSantis, C.E.; Fedewa, S.A.; Goding Sauer, A.; Kramer, J.L.; Smith, R.A.; Jemal, A. Breast Cancer Statistics, 2015: Convergence of Incidence Rates between Black and White Women. CA Cancer J Clin 2016, 66. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R. Global, Regional, National Burden of Breast Cancer in 185 Countries: Evidence from GLOBOCAN 2018. Breast Cancer Res Treat 2021, 187. [Google Scholar] [CrossRef] [PubMed]
- Coughlin, S.S. Epidemiology of Breast Cancer in Women. In Advances in Experimental Medicine and Biology; 2019; Vol. 1152.
- Majeed, W.; Aslam, B.; Javed, I.; Khaliq, T.; Muhammad, F.; Ali, A.; Raza, A. Breast Cancer: Major Risk Factors and Recent Developments in Treatment. Asian Pacific Journal of Cancer Prevention 2014, 15. [Google Scholar] [CrossRef] [PubMed]
- Galati, F.; Magri, V.; Arias-Cadena, P.A.; Moffa, G.; Rizzo, V.; Pasculli, M.; Botticelli, A.; Pediconi, F. Pregnancy-Associated Breast Cancer: A Diagnostic and Therapeutic Challenge. Diagnostics 2023, 13. [Google Scholar] [CrossRef] [PubMed]
- Key, T. Sex Hormones and Risk of Breast Cancer in Premenopausal Women: A Collaborative Reanalysis of Individual Participant Data from Seven Prospective Studies. Lancet Oncol 2013, 14. [Google Scholar] [CrossRef]
- Jung, S.; Wang, M.; Anderson, K.; Baglietto, L.; Bergkvist, L.; Bernstein, L.; van den Brandt, P.A.; Brinton, L.; Buring, J.E.; Heather Eliassen, A.; et al. Alcohol Consumption and Breast Cancer Risk by Estrogen Receptor Status: In a Pooled Analysis of 20 Studies. Int J Epidemiol 2016, 45. [Google Scholar] [CrossRef]
- Bodewes, F.T.H.; van Asselt, A.A.; Dorrius, M.D.; Greuter, M.J.W.; de Bock, G.H. Mammographic Breast Density and the Risk of Breast Cancer: A Systematic Review and Meta-Analysis. Breast 2022, 66. [Google Scholar] [CrossRef]
- Giampaolino, P.; Cafasso, V.; Boccia, D.; Ascione, M.; Mercorio, A.; Viciglione, F.; Palumbo, M.; Serafino, P.; Buonfantino, C.; De Angelis, M.C.; et al. Fertility-Sparing Approach in Patients with Endometrioid Endometrial Cancer Grade 2 Stage IA (FIGO): A Qualitative Systematic Review. Biomed Res Int 2022, 2022. [Google Scholar] [CrossRef]
- Mutlu, L.; Manavella, D.D.; Gullo, G.; McNamara, B.; Santin, A.D.; Patrizio, P. Endometrial Cancer in Reproductive Age: Fertility-Sparing Approach and Reproductive Outcomes. Cancers (Basel) 2022, 14. [Google Scholar] [CrossRef]
- Gullo, G.; Cucinella, G.; Chiantera, V.; Dellino, M.; Cascardi, E.; Török, P.; Herman, T.; Garzon, S.; Uccella, S.; Laganà, A.S. Fertility-Sparing Strategies for Early-Stage Endometrial Cancer: Stepping towards Precision Medicine Based on the Molecular Fingerprint. Int J Mol Sci 2023, 24. [Google Scholar] [CrossRef] [PubMed]
- Piergentili, R.; Gullo, G.; Basile, G.; Gulia, C.; Porrello, A.; Cucinella, G.; Marinelli, E.; Zaami, S. Circulating MiRNAs as a Tool for Early Diagnosis of Endometrial Cancer—Implications for the Fertility-Sparing Process: Clinical, Biological, and Legal Aspects. Int J Mol Sci 2023, 24, 11356. [Google Scholar] [CrossRef] [PubMed]
- Gullo, G.; Perino, A.; Cucinella, G. Open vs. Closed Vitrification System: Which One Is Safer? Eur Rev Med Pharmacol Sci 2022, 26. [Google Scholar]
- Zaami, S.; Stark, M.; Signore, F.; Gullo, G.; Marinelli, E. Fertility Preservation in Female Cancer Sufferers: (Only) a Moral Obligation? European Journal of Contraception and Reproductive Health Care 2022, 27. [Google Scholar] [CrossRef] [PubMed]
- Knabben, L.; Mueller, M.D. Breast Cancer and Pregnancy. In Proceedings of the Hormone Molecular Biology and Clinical Investigation; 2017; Vol. 32.
- Burgio, S.; Polizzi, C.; Buzzaccarini, G.; Laganà, A.S.; Gullo, G.; Perricone, G.; Perino, A.; Cucinella, G.; Alesi, M. Psychological Variables in Medically Assisted Reproduction: A Systematic Review. Przeglad Menopauzalny 2022, 21. [Google Scholar] [CrossRef] [PubMed]
- Smrekar, K.; Lodise, N.M. Combined Oral Contraceptive Use and Breast Cancer Risk: Select Considerations for Clinicians. Nurs Womens Health 2022, 26. [Google Scholar] [CrossRef] [PubMed]
- Kamani, M.O.; Akgor, U.; Gültekin, M. Review of the Literature on Combined Oral Contraceptives and Cancer. Ecancermedicalscience 2022, 16. [Google Scholar] [CrossRef]
- Petrucelli, N.; Daly, M.B.; Pal, T. BRCA1- and BRCA2-Associated Hereditary Breast and Ovarian Cancer; 1993.
- Carbognin, L.; Miglietta, F.; Paris, I.; Dieci, M.V. Prognostic and Predictive Implications of PTEN in Breast Cancer: Unfulfilled Promises but Intriguing Perspectives. Cancers (Basel) 2019, 11. [Google Scholar] [CrossRef]
- Shahbandi, A.; Nguyen, H.D.; Jackson, J.G. TP53 Mutations and Outcomes in Breast Cancer: Reading beyond the Headlines. Trends Cancer 2020, 6. [Google Scholar] [CrossRef]
- Corso, G.; Veronesi, P.; Sacchini, V.; Galimberti, V. Prognosis and Outcome in CDH1-Mutant Lobular Breast Cancer. European Journal of Cancer Prevention 2018, 27. [Google Scholar] [CrossRef]
- Beggs, A.D.; Latchford, A.R.; Vasen, H.F.A.; Moslein, G.; Alonso, A.; Aretz, S.; Bertario, L.; Blanco, I.; Bülow, S.; Burn, J.; et al. Peutz - Jeghers Syndrome: A Systematic Review and Recommendations for Management. Gut 2010, 59. [Google Scholar] [CrossRef] [PubMed]
- Apostolou, P.; Papasotiriou, I. Current Perspectives on CHEK2 Mutations in Breast Cancer. Breast Cancer : Targets and Therapy 2017, 9, 331. [Google Scholar] [CrossRef] [PubMed]
- Nepomuceno, T.C.; Carvalho, M.A.; Rodrigue, A.; Simard, J.; Masson, J.Y.; Monteiro, A.N.A. PALB2 Variants: Protein Domains and Cancer Susceptibility. Trends Cancer 2021, 7, 188–197. [Google Scholar] [CrossRef] [PubMed]
- Stucci, L.S.; Internò, V.; Tucci, M.; Perrone, M.; Mannavola, F.; Palmirotta, R.; Porta, C. The ATM Gene in Breast Cancer: Its Relevance in Clinical Practice. Genes (Basel) 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; McInerny, S.; Zethoven, M.; Cheasley, D.; Lim, B.W.X.; Rowley, S.M.; Devereux, L.; Grewal, N.; Ahmadloo, S.; Byrne, D.; et al. Combined Tumor Sequencing and Case-Control Analyses of RAD51C in Breast Cancer. J Natl Cancer Inst 2019, 111, 1332–1338. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Li, Y.; Ouyang, T.; Li, J.; Wang, T.; Fan, Z.; Fan, T.; Lin, B.; Xie, Y. Associations between RAD51D Germline Mutations and Breast Cancer Risk and Survival in BRCA1/2-Negative Breast Cancers. Annals of Oncology 2018, 29, 2046–2051. [Google Scholar] [CrossRef] [PubMed]
- Śniadecki, M.; Brzeziński, M.; Darecka, K.; Klasa-Mazurkiewicz, D.; Poniewierza, P.; Krzeszowiec, M.; Kmieć, N.; Wydra, D. BARD1 and Breast Cancer: The Possibility of Creating Screening Tests and New Preventive and Therapeutic Pathways for Predisposed Women. Genes (Basel) 2020, 11, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Suarez-Kelly, L.P.; Yu, L.; Kline, D.; Schneider, E.B.; Agnese, D.M.; Carson, W.E. Increased Breast Cancer Risk in Women with Neurofibromatosis Type 1: A Meta-Analysis and Systematic Review of the Literature. Hered Cancer Clin Pract 2019, 17, 1–13. [Google Scholar] [CrossRef]
- Khan, U.; Khan, M.S. Prognostic Value Estimation of BRIP1 in Breast Cancer by Exploiting Transcriptomics Data Through Bioinformatics Approaches. Bioinform Biol Insights 2021, 15. [Google Scholar] [CrossRef]
- Zhang, B.; Beeghly-Fadiel, A.; Long, J.; Zheng, W. Genetic Variants Associated with Breast-Cancer Risk: Comprehensive Research Synopsis, Meta-Analysis, and Epidemiological Evidence. Lancet Oncol 2011, 12, 477–488. [Google Scholar] [CrossRef]
- Niell, B.L.; Freer, P.E.; Weinfurtner, R.J.; Arleo, E.K.; Drukteinis, J.S. Screening for Breast Cancer. Radiol Clin North Am 2017, 55, 1145–1162. [Google Scholar] [CrossRef] [PubMed]
- Andreea, G.I.; Pegza, R.; Lascu, L.; Bondari, S.; Stoica, Z.; Bondari, A. The Role of Imaging Techniques in Diagnosis of Breast Cancer. 2012.
- Albert, U.S.; Altland, H.; Duda, V.; Engel, J.; Geraedts, M.; Heywang-Köbrunner, S.; Hölzel, D.; Kalbheim, E.; Koller, M.; König, K.; et al. 2008 Update of the Guideline: Early Detection of Breast Cancer in Germany. J Cancer Res Clin Oncol 2009, 135, 339–354. [Google Scholar] [CrossRef] [PubMed]
- Lima, Z.S.; Ebadi, M.R.; Amjad, G.; Younesi, L. Application of Imaging Technologies in Breast Cancer Detection: A Review Article. Open Access Maced J Med Sci 2019, 7, 838–848. [Google Scholar] [CrossRef] [PubMed]
- Gerami, R.; Joni, S.S.; Akhondi, N.; Etemadi, A.; Fosouli, M.; Eghbal, A.F.; Souri, Z. A Literature Review on the Imaging Methods for Breast Cancer. Int J Physiol Pathophysiol Pharmacol 2022, 14, 171. [Google Scholar] [PubMed]
- Zeng, Z.; Amin, A.; Roy, A.; Pulliam, N.E.; Karavites, L.C.; Espino, S.; Helenowski, I.; Li, X.; Luo, Y.; Khan, S.A. Preoperative Magnetic Resonance Imaging Use and Oncologic Outcomes in Premenopausal Breast Cancer Patients. NPJ Breast Cancer 2020, 6. [Google Scholar] [CrossRef] [PubMed]
- El Bairi, K.; Haynes, H.R.; Blackley, E.; Fineberg, S.; Shear, J.; Turner, S.; de Freitas, J.R.; Sur, D.; Amendola, L.C.; Gharib, M.; et al. The Tale of TILs in Breast Cancer: A Report from The International Immuno-Oncology Biomarker Working Group. NPJ Breast Cancer 2021, 7. [Google Scholar] [CrossRef] [PubMed]
- Dromain, C.; Balleyguier, C. Contrast-Enhanced Digital Mammography. 2010, 187–198. [CrossRef]
- Mortezazadeh, T.; Gholibegloo, E.; Riyahi, A.N.; Haghgoo, S.; Musa, A.E.; Khoobi, M. Glucosamine Conjugated Gadolinium (III) Oxide Nanoparticles as a Novel Targeted Contrast Agent for Cancer Diagnosis in MRI. J Biomed Phys Eng 2020, 10, 25–38. [Google Scholar] [CrossRef]
- Guo, R.; Lu, G.; Qin, B.; Fei, B. Ultrasound Imaging Technologies for Breast Cancer Detection and Management: A Review. Ultrasound Med Biol 2018, 44, 37–70. [Google Scholar] [CrossRef]
- Carpentier, B.; Hayward, J.; Strachowski, L. Enhancing Your Acoustics: Ultrasound Image Optimization of Breast Lesions. J Ultrasound Med 2017, 36, 1479–1485. [Google Scholar] [CrossRef]
- Drukteinis, J.S.; Mooney, B.P.; Flowers, C.I.; Gatenby, R.A. Beyond Mammography: New Frontiers in Breast Cancer Screening. Am J Med 2013, 126, 472–479. [Google Scholar] [CrossRef]
- Iranmakani, S.; Mortezazadeh, T.; Sajadian, F.; Ghaziani, M.F.; Ghafari, A.; Khezerloo, D.; Musa, A.E. A Review of Various Modalities in Breast Imaging: Technical Aspects and Clinical Outcomes. Egyptian Journal of Radiology and Nuclear Medicine 2020, 51. [Google Scholar] [CrossRef]
- Veronesi, U.; Viale, G.; Rotmensz, N.; Goldhirsch, A. Rethinking TNM: Breast Cancer TNM Classification for Treatment Decision-Making and Research. Breast 2006, 15, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Eliyatkin, N.; Yalcin, E.; Zengel, B.; Aktaş, S.; Vardar, E. Molecular Classification of Breast Carcinoma: From Traditional, Old-Fashioned Way to A New Age, and A New Way. J Breast Health 2015, 11, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Perou, C.M.; Sørile, T.; Eisen, M.B.; Van De Rijn, M.; Jeffrey, S.S.; Ress, C.A.; Pollack, J.R.; Ross, D.T.; Johnsen, H.; Akslen, L.A.; et al. Molecular Portraits of Human Breast Tumours. Nature 2000, 406, 747–752. [Google Scholar] [CrossRef]
- Sinn, H.P.; Kreipe, H. A Brief Overview of the WHO Classification of Breast Tumors, 4th Edition, Focusing on Issues and Updates from the 3rd Edition. Breast Care (Basel) 2013, 8, 149–154. [Google Scholar] [CrossRef]
- Hortobagyi, G.N.; Edge, S.B.; Giuliano, A. New and Important Changes in the TNM Staging System for Breast Cancer. Am Soc Clin Oncol Educ Book 2018, 38, 457–467. [Google Scholar] [CrossRef] [PubMed]
- Giuliano, A.E.; Connolly, J.L.; Edge, S.B.; Mittendorf, E.A.; Rugo, H.S.; Solin, L.J.; Weaver, D.L.; Winchester, D.J.; Hortobagyi, G.N. Breast Cancer-Major Changes in the American Joint Committee on Cancer Eighth Edition Cancer Staging Manual. CA Cancer J Clin 2017, 67, 290–303. [Google Scholar] [CrossRef]
- Fisusi, F.A.; Akala, E.O. Drug Combinations in Breast Cancer Therapy. Pharm Nanotechnol 2019, 7, 3. [Google Scholar] [CrossRef]
- Burstein, H.J.; Curigliano, G.; Thürlimann, B.; Weber, W.P.; Poortmans, P.; Regan, M.M.; Senn, H.J.; Winer, E.P.; Gnant, M.; Aebi, S.; et al. Customizing Local and Systemic Therapies for Women with Early Breast Cancer: The St. Gallen International Consensus Guidelines for Treatment of Early Breast Cancer 2021. Ann Oncol 2021, 32, 1216–1235. [Google Scholar] [CrossRef]
- Tanaka, S.; Iwamoto, M.; Kimura, K.; Matsunami, N.; Morishima, H.; Yoshidome, K.; Nomura, T.; Morimoto, T.; Yamamoto, D.; Tsubota, Y.; et al. Phase II Study of Neoadjuvant Anthracycline-Based Regimens Combined With Nanoparticle Albumin-Bound Paclitaxel and Trastuzumab for Human Epidermal Growth Factor Receptor 2-Positive Operable Breast Cancer. Clin Breast Cancer 2015, 15, 191–196. [Google Scholar] [CrossRef]
- Darby, S.; McGale, P.; Correa, C.; Taylor, C.; Arriagada, R.; Clarke, M.; Cutter, D.; Davies, C.; Ewertz, M.; Godwin, J.; et al. Effect of Radiotherapy after Breast-Conserving Surgery on 10-Year Recurrence and 15-Year Breast Cancer Death: Meta-Analysis of Individual Patient Data for 10,801 Women in 17 Randomised Trials. Lancet 2011, 378, 1707–1716. [Google Scholar] [CrossRef] [PubMed]
- Sayan, M.; Yehia, Z.A.; Ohri, N.; Haffty, B.G. Hypofractionated Postmastectomy Radiation Therapy. Adv Radiat Oncol 2020, 6. [Google Scholar] [CrossRef] [PubMed]
- Franceschini, G.; Sanchez, A.M.; Di Leone, A.; Magno, S.; Moschella, F.; Accetta, C.; Masetti, R. New Trends in Breast Cancer Surgery: A Therapeutic Approach Increasingly Efficacy and Respectful of the Patient. G Chir 2015, 36, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Tung, N.; Garber, J.E. PARP Inhibition in Breast Cancer: Progress Made and Future Hopes. NPJ Breast Cancer 2022, 8. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues-Ferreira, S.; Nahmias, C. Predictive Biomarkers for Personalized Medicine in Breast Cancer. Cancer Lett 2022, 545. [Google Scholar] [CrossRef] [PubMed]
- Naeli, P.; Winter, T.; Hackett, A.P.; Alboushi, L.; Jafarnejad, S.M. The Intricate Balance between MicroRNA-Induced MRNA Decay and Translational Repression. FEBS J 2023, 290. [Google Scholar] [CrossRef] [PubMed]
- Friedman, R.C.; Farh, K.K.H.; Burge, C.B.; Bartel, D.P. Most Mammalian MRNAs Are Conserved Targets of MicroRNAs. Genome Res 2009, 19, 92. [Google Scholar] [CrossRef]
- Rodriguez, A.; Griffiths-Jones, S.; Ashurst, J.L.; Bradley, A. Identification of Mammalian MicroRNA Host Genes and Transcription Units. Genome Res 2004, 14, 1902–1910. [Google Scholar] [CrossRef]
- Piergentili, R.; Basile, G.; Nocella, C.; Carnevale, R.; Marinelli, E.; Patrone, R.; Zaami, S. Using NcRNAs as Tools in Cancer Diagnosis and Treatment—The Way towards Personalized Medicine to Improve Patients’ Health. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front Endocrinol (Lausanne) 2018, 9, 388354. [Google Scholar] [CrossRef]
- Lee, Y.; Kim, M.; Han, J.; Yeom, K.H.; Lee, S.; Baek, S.H.; Kim, V.N. MicroRNA Genes Are Transcribed by RNA Polymerase II. EMBO J 2004, 23, 4051–4060. [Google Scholar] [CrossRef] [PubMed]
- Meijer, H.A.; Smith, E.M.; Bushell, M. Regulation of MiRNA Strand Selection: Follow the Leader? Biochem Soc Trans 2014, 42, 1135–1140. [Google Scholar] [CrossRef] [PubMed]
- Ambros, V.; Bartel, B.; Bartel, D.P.; Burge, C.B.; Carrington, J.C.; Chen, X.; Dreyfuss, G.; Eddy, S.R.; Griffiths-Jones, S.; Marshall, M.; et al. A Uniform System for MicroRNA Annotation. RNA 2003, 9, 277–279. [Google Scholar] [CrossRef] [PubMed]
- Griffiths-Jones, S.; Grocock, R.J.; van Dongen, S.; Bateman, A.; Enright, A.J. MiRBase: MicroRNA Sequences, Targets and Gene Nomenclature. Nucleic Acids Res 2006, 34, D140. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.C.; Feinbaum, R.L.; Ambros, V. The C. Elegans Heterochronic Gene Lin-4 Encodes Small RNAs with Antisense Complementarity to Lin-14. Cell 1993, 75, 843–854. [Google Scholar] [CrossRef] [PubMed]
- Duan, R.; Pak, C.H.; Jin, P. Single Nucleotide Polymorphism Associated with Mature MiR-125a Alters the Processing of Pri-MiRNA. Hum Mol Genet 2007, 16, 1124–1131. [Google Scholar] [CrossRef] [PubMed]
- Shaham, L.; Binder, V.; Gefen, N.; Borkhardt, A.; Izraeli, S. MiR-125 in Normal and Malignant Hematopoiesis. Leukemia 2012 26:9 2012, 26, 2011–2018. [Google Scholar] [CrossRef]
- Ciafrè, S.A.; Galardi, S.; Mangiola, A.; Ferracin, M.; Liu, C.G.; Sabatino, G.; Negrini, M.; Maira, G.; Croce, C.M.; Farace, M.G. Extensive Modulation of a Set of MicroRNAs in Primary Glioblastoma. Biochem Biophys Res Commun 2005, 334, 1351–1358. [Google Scholar] [CrossRef]
- Emmrich, S.; Streltsov, A.; Schmidt, F.; Thangapandi, V.R.; Reinhardt, D.; Klusmann, J.H. LincRNAs MONC and MIR100HG Act as Oncogenes in Acute Megakaryoblastic Leukemia. Mol Cancer 2014, 13. [Google Scholar] [CrossRef]
- Lagos-Quintana, M.; Rauhut, R.; Yalcin, A.; Meyer, J.; Lendeckel, W.; Tuschl, T. Identification of Tissue-Specific MicroRNAs from Mouse. Curr Biol 2002, 12, 735–739. [Google Scholar] [CrossRef]
- Huang, H.Y.; Lin, Y.C.D.; Cui, S.; Huang, Y.; Tang, Y.; Xu, J.; Bao, J.; Li, Y.; Wen, J.; Zuo, H.; et al. MiRTarBase Update 2022: An Informative Resource for Experimentally Validated MiRNA-Target Interactions. Nucleic Acids Res 2022, 50, D222–D230. [Google Scholar] [CrossRef]
- MiRTarBase: The Experimentally Validated MicroRNA-Target Interactions Database Available online:. Available online: https://mirtarbase.cuhk.edu.cn/~miRTarBase/miRTarBase_2022/php/index.php (accessed on 24 August 2023).
- Sun, Y.M.; Lin, K.Y.; Chen, Y.Q. Diverse Functions of MiR-125 Family in Different Cell Contexts. J Hematol Oncol 2013, 6, 6. [Google Scholar] [CrossRef] [PubMed]
- Ge, Y.; Sun, Y.; Chen, J. IGF-II Is Regulated by MicroRNA-125b in Skeletal Myogenesis. J Cell Biol 2011, 192, 69–81. [Google Scholar] [CrossRef] [PubMed]
- Qiu, J.; Zhu, J.; Zhang, R.; Liang, W.; Ma, W.; Zhang, Q.; Huang, Z.; Ding, F.; Sun, H. MiR-125b-5p Targeting TRAF6 Relieves Skeletal Muscle Atrophy Induced by Fasting or Denervation. Ann Transl Med 2019, 7, 456–456. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; He, L.; Li, X.; Pei, W.; Yang, H.; Zhong, M.; Zhang, M.; Lv, K.; Zhang, Y. LncRNA AK089514/MiR-125b-5p/TRAF6 Axis Mediates Macrophage Polarization in Allergic Asthma. BMC Pulm Med 2023, 23. [Google Scholar] [CrossRef]
- Wang, X.; Ha, T.; Zou, J.; Ren, D.; Liu, L.; Zhang, X.; Kalbfleisch, J.; Gao, X.; Williams, D.; Li, C. MicroRNA-125b Protects against Myocardial Ischaemia/Reperfusion Injury via Targeting P53-Mediated Apoptotic Signalling and TRAF6. Cardiovasc Res 2014, 102, 385–395. [Google Scholar] [CrossRef] [PubMed]
- Medrano, S.; Monteagudo, M.C.; Sequeira-Lopez, M.L.S.; Pentz, E.S.; Ariel Gomez, R. Two MicroRNAs, MiR-330 and MiR-125b-5p, Mark the Juxtaglomerular Cell and Balance Its Smooth Muscle Phenotype. Am J Physiol Renal Physiol 2012, 302, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Chen, S.; Gao, Y.; Yu, C.; Nie, Z.; Lu, R.; Sun, Y.; Guan, Z. MicroRNA-125b Inhibits the Proliferation of Vascular Smooth Muscle Cells Induced by Platelet-Derived Growth Factor BB. Exp Ther Med 2021, 22. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tan, J.; Wang, L.; Pei, G.; Cheng, H.; Zhang, Q.; Wang, S.; He, C.; Fu, C.; Wei, Q. MiR-125 Family in Cardiovascular and Cerebrovascular Diseases. Front Cell Dev Biol 2021, 9. [Google Scholar] [CrossRef]
- Lozano-Velasco, E.; Galiano-Torres, J.; Jodar-Garcia, A.; Aranega, A.E.; Franco, D. MiR-27 and MiR-125 Distinctly Regulate Muscle-Enriched Transcription Factors in Cardiac and Skeletal Myocytes. Biomed Res Int 2015, 2015. [Google Scholar] [CrossRef]
- Li, L.; Wang, Q.; Yuan, Z.; Chen, A.; Liu, Z.; Wang, Z.; Li, H. LncRNA-MALAT1 Promotes CPC Proliferation and Migration in Hypoxia by up-Regulation of JMJD6 via Sponging MiR-125. Biochem Biophys Res Commun 2018, 499, 711–718. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Zhang, M.; Chen, W.; Wang, R.; Ye, Z.; Wang, Y.; Li, X.; Cai, C. LncRNA-HOTAIR Inhibition Aggravates Oxidative Stress-Induced H9c2 Cells Injury through Suppression of MMP2 by MiR-125. Acta Biochim Biophys Sin (Shanghai) 2018, 50, 996–1006. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.Y.; Lee, D.S.; Choong, O.K.; Chang, S.K.; Hsu, T.; Nicholson, M.W.; Liu, L.W.; Lin, P.J.; Ruan, S.C.; Lin, S.W.; et al. Cardiac-Specific MicroRNA-125b Deficiency Induces Perinatal Death and Cardiac Hypertrophy. Sci Rep 2021, 11. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhu, Y.; Liu, S.; Liu, J.; Li, X. NORAD Lentivirus ShRNA Mitigates Fibrosis and Inflammatory Responses in Diabetic Cardiomyopathy via the CeRNA Network of NORAD/MiR-125a-3p/Fyn. Inflamm Res 2021, 70, 1113–1127. [Google Scholar] [CrossRef] [PubMed]
- Sempere, L.F.; Freemantle, S.; Pitha-Rowe, I.; Moss, E.; Dmitrovsky, E.; Ambros, V. Expression Profiling of Mammalian MicroRNAs Uncovers a Subset of Brain-Expressed MicroRNAs with Possible Roles in Murine and Human Neuronal Differentiation. Genome Biol 2004, 5. [Google Scholar] [CrossRef] [PubMed]
- Le, M.T.N.; Xie, H.; Zhou, B.; Chia, P.H.; Rizk, P.; Um, M.; Udolph, G.; Yang, H.; Lim, B.; Lodish, H.F. MicroRNA-125b Promotes Neuronal Differentiation in Human Cells by Repressing Multiple Targets. Mol Cell Biol 2009, 29, 5290–5305. [Google Scholar] [CrossRef] [PubMed]
- Dash, S.; Balasubramaniam, M.; Rana, T.; Godino, A.; Peck, E.G.; Goodwin, J.S.; Villalta, F.; Calipari, E.S.; Nestler, E.J.; Dash, C.; et al. Poly (ADP-Ribose) Polymerase-1 (PARP-1) Induction by Cocaine Is Post-Transcriptionally Regulated by MiR-125b. eNeuro 2017, 4. [Google Scholar] [CrossRef] [PubMed]
- Edbauer, D.; Neilson, J.R.; Foster, K.A.; Wang, C.F.; Seeburg, D.P.; Batterton, M.N.; Tada, T.; Dolan, B.M.; Sharp, P.A.; Sheng, M. Regulation of Synaptic Structure and Function by FMRP-Associated MicroRNAs MiR-125b and MiR-132. Neuron 2010, 65, 373–384. [Google Scholar] [CrossRef]
- Åkerblom, M.; Petri, R.; Sachdeva, R.; Klussendorf, T.; Mattsson, B.; Gentner, B.; Jakobsson, J. MicroRNA-125 Distinguishes Developmentally Generated and Adult-Born Olfactory Bulb Interneurons. Development 2014, 141, 1580–1588. [Google Scholar] [CrossRef]
- Gioia, U.; Di Carlo, V.; Caramanica, P.; Toselli, C.; Cinquino, A.; Marchioni, M.; Laneve, P.; Biagioni, S.; Bozzoni, I.; Cacci, E.; et al. Mir-23a and Mir-125b Regulate Neural Stem/Progenitor Cell Proliferation by Targeting Musashi1. RNA Biol 2014, 11, 1105–1112. [Google Scholar] [CrossRef]
- Shenoy, A.; Danial, M.; Blelloch, R.H. Let-7 and MiR-125 Cooperate to Prime Progenitors for Astrogliogenesis. EMBO J 2015, 34, 1180–1194. [Google Scholar] [CrossRef] [PubMed]
- Pogue, A.I.; Cui, J.G.; Li, Y.Y.; Zhao, Y.; Culicchia, F.; Lukiw, W.J. Micro RNA-125b (MiRNA-125b) Function in Astrogliosis and Glial Cell Proliferation. Neurosci Lett 2010, 476, 18–22. [Google Scholar] [CrossRef] [PubMed]
- Klusmann, J.H.; Li, Z.; Böhmer, K.; Maroz, A.; Koch, M.L.; Emmrich, S.; Godinho, F.J.; Orkin, S.H.; Reinhardt, D. MiR-125b-2 Is a Potential OncomiR on Human Chromosome 21 in Megakaryoblastic Leukemia. Genes Dev 2010, 24, 478–490. [Google Scholar] [CrossRef] [PubMed]
- Tiedt, S.; Prestel, M.; Malik, R.; Schieferdecker, N.; Duering, M.; Kautzky, V.; Stoycheva, I.; Böck, J.; Northoff, B.H.; Klein, M.; et al. RNA-Seq Identifies Circulating MiR-125a-5p, MiR-125b-5p, and MiR-143-3p as Potential Biomarkers for Acute Ischemic Stroke. Circ Res 2017, 121, 970–980. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Li, L.; Dong, L.; Cheng, J.; Zhao, C.; Bao, C.; Wang, H. Circulating MRNA and MicroRNA Profiling Analysis in Patients with Ischemic Stroke. Mol Med Rep 2020, 22, 792–802. [Google Scholar] [CrossRef] [PubMed]
- Banzhaf-Strathmann, J.; Benito, E.; May, S.; Arzberger, T.; Tahirovic, S.; Kretzschmar, H.; Fischer, A.; Edbauer, D. MicroRNA-125b Induces Tau Hyperphosphorylation and Cognitive Deficits in Alzheimer’s Disease. EMBO J 2014, 33, 1667–1680. [Google Scholar] [CrossRef] [PubMed]
- Alexandrov, P.N.; Dua, P.; Hill, J.M.; Bhattacharjee, S.; Zhao, Y.; Lukiw, W.J. MicroRNA (MiRNA) Speciation in Alzheimer’s Disease (AD) Cerebrospinal Fluid (CSF) and Extracellular Fluid (ECF). Int J Biochem Mol Biol 2012, 3, 365. [Google Scholar]
- Chou, M.Y.; Cao, X.; Hou, K.C.; Tsai, M.H.; Lee, C.Y.; Kuo, M.F.; Wu, V.C.; Huang, H.Y.; Akbarian, S.; Chang, S.K.; et al. Mir125b-2 Imprinted in Human but Not Mouse Brain Regulates Hippocampal Function and Circuit in Mice. Commun Biol 2023, 6. [Google Scholar] [CrossRef]
- Yuan, M.; Da Silva, A.C.A.L.; Arnold, A.; Okeke, L.; Ames, H.; Correa-Cerro, L.S.; Vizcaino, M.A.; Ho, C.Y.; Eberhart, C.G.; Rodriguez, F.J. MicroRNA (MiR) 125b Regulates Cell Growth and Invasion in Pediatric Low Grade Glioma. Sci Rep 2018, 8. [Google Scholar] [CrossRef]
- Ferretti, E.; De Smaele, E.; Po, A.; Marcotullio, L. Di; Tosi, E.; Espinola, M.S.B.; Rocco, C. Di; Riccardi, R.; Giangaspero, F.; Farcomeni, A.; et al. MicroRNA Profiling in Human Medulloblastoma. Int J Cancer 2009, 124, 568–577. [Google Scholar] [CrossRef]
- Laneve, P.; Di Marcotullio, L.; Gioia, U.; Fiori, M.E.; Ferretti, E.; Gulino, A.; Bozzoni, I.; Caffarelli, E. The Interplay between MicroRNAs and the Neurotrophin Receptor Tropomyosin-Related Kinase C Controls Proliferation of Human Neuroblastoma Cells. Proc Natl Acad Sci U S A 2007, 104, 7957–7962. [Google Scholar] [CrossRef] [PubMed]
- Wu, N.; Lin, X.; Zhao, X.; Zheng, L.; Xiao, L.; Liu, J.; Ge, L.; Cao, S. MiR-125b Acts as an Oncogene in Glioblastoma Cells and Inhibits Cell Apoptosis through P53 and P38MAPK-Independent Pathways. Br J Cancer 2013, 109, 2853–2863. [Google Scholar] [CrossRef] [PubMed]
- Xia, H.F.; He, T.Z.; Liu, C.M.; Cui, Y.; Song, P.P.; Jin, X.H.; Ma, X. MiR-125b Expression Affects the Proliferation and Apoptosis of Human Glioma Cells by Targeting Bmf. Cell Physiol Biochem 2009, 23, 347–358. [Google Scholar] [CrossRef] [PubMed]
- Cron, M.A.; Maillard, S.; Delisle, F.; Samson, N.; Truffault, F.; Foti, M.; Fadel, E.; Guihaire, J.; Berrih-Aknin, S.; Le Panse, R. Analysis of MicroRNA Expression in the Thymus of Myasthenia Gravis Patients Opens New Research Avenues. Autoimmun Rev 2018, 17, 588–600. [Google Scholar] [CrossRef] [PubMed]
- Wojtowicz, E.E.; Lechman, E.R.; Hermans, K.G.; Schoof, E.M.; Wienholds, E.; Isserlin, R.; van Veelen, P.A.; Broekhuis, M.J.C.; Janssen, G.M.C.; Trotman-Grant, A.; et al. Ectopic MiR-125a Expression Induces Long-Term Repopulating Stem Cell Capacity in Mouse and Human Hematopoietic Progenitors. Cell Stem Cell 2016, 19, 383–396. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Lu, J.; Schlanger, R.; Zhang, H.; Wang, J.Y.; Fox, M.C.; Purton, L.E.; Fleming, H.H.; Cobb, B.; Merkenschlager, M.; et al. MicroRNA MiR-125a Controls Hematopoietic Stem Cell Number. Proc Natl Acad Sci U S A 2010, 107, 14229–14234. [Google Scholar] [CrossRef] [PubMed]
- Emmrich, S.; Rasche, M.; Schöning, J.; Reimer, C.; Keihani, S.; Maroz, A.; Xie, Y.; Li, Z.; Schambach, A.; Reinhardt, D.; et al. MiR-99a/100~125b Tricistrons Regulate Hematopoietic Stem and Progenitor Cell Homeostasis by Shifting the Balance between TGFβ and Wnt Signaling. Genes Dev 2014, 28, 858–874. [Google Scholar] [CrossRef]
- Allantaz, F.; Cheng, D.T.; Bergauer, T.; Ravindran, P.; Rossier, M.F.; Ebeling, M.; Badi, L.; Reis, B.; Bitter, H.; D’Asaro, M.; et al. Expression Profiling of Human Immune Cell Subsets Identifies MiRNA-MRNA Regulatory Relationships Correlated with Cell Type Specific Expression. PLoS One 2012, 7. [Google Scholar] [CrossRef]
- Chaudhuri, A.A.; So, A.Y.-L.; Sinha, N.; Gibson, W.S.J.; Taganov, K.D.; O’Connell, R.M.; Baltimore, D. MicroRNA-125b Potentiates Macrophage Activation. J Immunol 2011, 187, 5062–5068. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, M.; Zhong, M.; Suo, Q.; Lv, K. Expression Profiles of MiRNAs in Polarized Macrophages. Int J Mol Med 2013, 31, 797–802. [Google Scholar] [CrossRef]
- Staedel, C.; Darfeuille, F. MicroRNAs and Bacterial Infection. Cell Microbiol 2013, 15, 1496–1507. [Google Scholar] [CrossRef]
- Taheri, M.; Khoshbakht, T.; Hussen, B.M.; Abdullah, S.T.; Ghafouri-Fard, S.; Sayad, A. The Emerging Role of MicroRNA in Periodontitis: Pathophysiology, Clinical Potential and Future Molecular Perspectives. Curr Stem Cell Res Ther 2022, 17. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Zhang, N.; Lin, Z. MicroRNA-125a-5p Modulates Macrophage Polarization by Targeting E26 Transformation-Specific Variant 6 Gene during Orthodontic Tooth Movement. Arch Oral Biol 2021, 124. [Google Scholar] [CrossRef] [PubMed]
- Yao, D.; Zhou, Z.; Wang, P.; Zheng, L.; Huang, Y.; Duan, Y.; Liu, B.; Li, Y. MiR-125-5p/IL-6R Axis Regulates Macrophage Inflammatory Response and Intestinal Epithelial Cell Apoptosis in Ulcerative Colitis through JAK1/STAT3 and NF-ΚB Pathway. Cell Cycle 2021, 20, 2547–2564. [Google Scholar] [CrossRef]
- Yu, C.; Tang, W.; Lu, R.; Tao, Y.; Ren, T.; Gao, Y. Human Adipose-Derived Mesenchymal Stem Cells Promote Lymphocyte Apoptosis and Alleviate Atherosclerosis via MiR-125b-1-3p/BCL11B Signal Axis. Ann Palliat Med 2021, 10, 2123–2133. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Zhang, S.; Ma, X. Prognostic Value of MicroRNA-125 in Various Human Malignant Neoplasms: A Meta-Analysis. Clin Lab 2015, 61, 1667–1674. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Gao, B.; Yue, J.; Liu, F.; Liu, Y.; Fu, W.; Si, Y. Exosomes from Mesenchymal Stem Cells Expressing MiR-125b Inhibit Neointimal Hyperplasia via Myosin IE. J Cell Mol Med 2019, 23, 1528–1540. [Google Scholar] [CrossRef] [PubMed]
- Cowden Dahl, K.D.; Dahl, R.; Kruichak, J.N.; Hudson, L.G. The Epidermal Growth Factor Receptor Responsive MiR-125a Represses Mesenchymal Morphology in Ovarian Cancer Cells. Neoplasia 2009, 11, 1208–1215. [Google Scholar] [CrossRef]
- Guan, Y.; Yao, H.; Zheng, Z.; Qiu, G.; Sun, K. MiR-125b Targets BCL3 and Suppresses Ovarian Cancer Proliferation. Int J Cancer 2011, 128, 2274–2283. [Google Scholar] [CrossRef]
- Chen, Z.; Guo, X.; Sun, S.; Lu, C.; Wang, L. Serum MiR-125b Levels Associated with Epithelial Ovarian Cancer (EOC) Development and Treatment Responses. Bioengineered 2020, 11, 311–317. [Google Scholar] [CrossRef]
- Huang, L.; Luo, J.; Cai, Q.; Pan, Q.; Zeng, H.; Guo, Z.; Dong, W.; Huang, J.; Lin, T. MicroRNA-125b Suppresses the Development of Bladder Cancer by Targeting E2F3. Int J Cancer 2011, 128, 1758–1769. [Google Scholar] [CrossRef] [PubMed]
- Pospisilova, S.; Pazzourkova, E.; Horinek, A.; Brisuda, A.; Svobodova, I.; Soukup, V.; Hrbacek, J.; Capoun, O.; Hanus, T.; Mares, J.; et al. MicroRNAs in Urine Supernatant as Potential Non-Invasive Markers for Bladder Cancer Detection. Neoplasma 2016, 63, 799–808. [Google Scholar] [CrossRef] [PubMed]
- Blick, C.; Ramachandran, A.; Mccormick, R.; Wigfield, S.; Cranston, D.; Catto, J.; Harris, A.L. Identification of a Hypoxia-Regulated MiRNA Signature in Bladder Cancer and a Role for MiR-145 in Hypoxia-Dependent Apoptosis. Br J Cancer 2015, 113, 634–644. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Tang, K.; Xiao, H.; Zeng, J.; Guan, W.; Guo, X.; Xu, H.; Ye, Z. A Panel of Eight-MiRNA Signature as a Potential Biomarker for Predicting Survival in Bladder Cancer. J Exp Clin Cancer Res 2015, 34. [Google Scholar] [CrossRef] [PubMed]
- Bi, Q.; Tang, S.; Xia, L.; Du, R.; Fan, R.; Gao, L.; Jin, J.; Liang, S.; Chen, Z.; Xu, G.; et al. Ectopic Expression of MiR-125a Inhibits the Proliferation and Metastasis of Hepatocellular Carcinoma by Targeting MMP11 and VEGF. PLoS One 2012, 7. [Google Scholar] [CrossRef]
- Jia, H.Y.; Wang, Y.X.; Yan, W.T.; Li, H.Y.; Tian, Y.Z.; Wang, S.M.; Zhao, H.L. MicroRNA-125b Functions as a Tumor Suppressor in Hepatocellular Carcinoma Cells. Int J Mol Sci 2012, 13, 8762–8774. [Google Scholar] [CrossRef] [PubMed]
- Liang, L.; Wong, C.M.; Ying, Q.; Fan, D.N.Y.; Huang, S.; Ding, J.; Yao, J.; Yan, M.; Li, J.; Yao, M.; et al. MicroRNA-125b Suppressesed Human Liver Cancer Cell Proliferation and Metastasis by Directly Targeting Oncogene LIN28B2. Hepatology 2010, 52, 1731–1740. [Google Scholar] [CrossRef] [PubMed]
- Kong, J.; Liu, X.; Li, X.; Wu, J.; Wu, N.; Chen, J.; Fang, F. MiR-125/Pokemon Auto-Circuit Contributes to the Progression of Hepatocellular Carcinoma. Tumour Biol 2016, 37, 511–519. [Google Scholar] [CrossRef]
- Xie, C.; Zhang, L.Z.; Chen, Z.L.; Zhong, W.J.; Fang, J.H.; Zhu, Y.; Xiao, M.H.; Guo, Z.W.; Zhao, N.; He, X.; et al. A HMTR4-PDIA3P1-MiR-125/124-TRAF6 Regulatory Axis and Its Function in NF Kappa B Signaling and Chemoresistance. Hepatology 2020, 71, 1660–1677. [Google Scholar] [CrossRef]
- Jiang, J.X.; Gao, S.; Pan, Y.Z.; Yu, C.; Sun, C.Y. Overexpression of MicroRNA-125b Sensitizes Human Hepatocellular Carcinoma Cells to 5-Fluorouracil through Inhibition of Glycolysis by Targeting Hexokinase II. Mol Med Rep 2014, 10, 995–1002. [Google Scholar] [CrossRef]
- Xu, Z.; Pei, C.; Cheng, H.; Song, K.; Yang, J.; Li, Y.; He, Y.; Liang, W.; Liu, B.; Tan, W.; et al. Comprehensive Analysis of FOXM1 Immune Infiltrates, M6a, Glycolysis and CeRNA Network in Human Hepatocellular Carcinoma. Front Immunol 2023, 14. [Google Scholar] [CrossRef]
- Chen, W.; Wang, T.; Li, W.; Yin, S. MiR-125b Acts as a Tumor Suppressor of Melanoma by Targeting NCAM. JBUON 2021, 26, 182–188. [Google Scholar]
- Kappelmann, M.; Kuphal, S.; Meister, G.; Vardimon, L.; Bosserhoff, A.K. MicroRNA MiR-125b Controls Melanoma Progression by Direct Regulation of c-Jun Protein Expression. Oncogene 2013, 32, 2984–2991. [Google Scholar] [CrossRef] [PubMed]
- Xu, N.; Zhang, L.; Meisgen, F.; Harada, M.; Heilborn, J.; Homey, B.; Grandér, D.; Ståhle, M.; Sonkoly, E.; Pivarcsi, A. MicroRNA-125b down-Regulates Matrix Metallopeptidase 13 and Inhibits Cutaneous Squamous Cell Carcinoma Cell Proliferation, Migration, and Invasion. J Biol Chem 2012, 287, 29899–29908. [Google Scholar] [CrossRef] [PubMed]
- Tian, K.; Liu, W.; Zhang, J.; Fan, X.; Liu, J.; Zhao, N.; Yao, C.; Miao, G. MicroRNA-125b Exerts Antitumor Functions in Cutaneous Squamous Cell Carcinoma by Targeting the STAT3 Pathway. Cell Mol Biol Lett 2020, 25. [Google Scholar] [CrossRef] [PubMed]
- Sand, M.; Skrygan, M.; Sand, D.; Georgas, D.; Hahn, S.A.; Gambichler, T.; Altmeyer, P.; Bechara, F.G. Expression of MicroRNAs in Basal Cell Carcinoma. Br J Dermatol 2012, 167, 847–855. [Google Scholar] [CrossRef] [PubMed]
- Liu, L. hong; Li, H.; Li, J. ping; Zhong, H.; Zhang, H. chon; Chen, J.; Xiao, T. MiR-125b Suppresses the Proliferation and Migration of Osteosarcoma Cells through down-Regulation of STAT3. Biochem Biophys Res Commun 2011, 416, 31–38. [Google Scholar] [CrossRef]
- Yang, Y.; Chen, Y.; Liu, J.; Zhang, B.; Yang, L.; Xue, J.; Zhang, Z.; Qin, L.; Bian, R. MiR-125b-5p/STAT3 Axis Regulates Drug Resistance in Osteosarcoma Cells by Acting on ABC Transporters. Stem Cells Int 2023, 2023. [Google Scholar] [CrossRef]
- Tang, X.Y.; Zheng, W.; Ding, M.; Guo, K.J.; Yuan, F.; Feng, H.; Deng, B.; Sun, W.; Hou, Y.; Gao, L. MiR-125b Acts as a Tumor Suppressor in Chondrosarcoma Cells by the Sensitization to Doxorubicin through Direct Targeting the ErbB2-Regulated Glucose Metabolism. Drug Des Devel Ther 2016, 10, 571–583. [Google Scholar] [CrossRef]
- Gao, S.; Sun, H.; Cheng, C.; Wang, G. BRCA1-Associated Protein-1 Suppresses Osteosarcoma Cell Proliferation and Migration Through Regulation PI3K/Akt Pathway. DNA Cell Biol 2017, 36, 386–393. [Google Scholar] [CrossRef]
- Wu, S.; Shen, W.; Yang, L.; Zhu, M.; Zhang, M.; Zong, F.; Geng, L.; Wang, Y.; Huang, T.; Pan, Y.; et al. Genetic Variations in MiR-125 Family and the Survival of Non-Small Cell Lung Cancer in Chinese Population. Cancer Med 2019, 8, 2636–2645. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Mao, W.; Zheng, S.; Ye, J. Epidermal Growth Factor Receptor-Regulated MiR-125a-5p--a Metastatic Inhibitor of Lung Cancer. FEBS J 2009, 276, 5571–5578. [Google Scholar] [CrossRef] [PubMed]
- Yagishita, S.; Fujita, Y.; Kitazono, S.; Ko, R.; Nakadate, Y.; Sawada, T.; Kitamura, Y.; Shimoyama, T.; Maeda, Y.; Takahashi, F.; et al. Chemotherapy-Regulated MicroRNA-125-HER2 Pathway as a Novel Therapeutic Target for Trastuzumab-Mediated Cellular Cytotoxicity in Small Cell Lung Cancer. Mol Cancer Ther 2015, 14, 1414–1423. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Wei, F.; Yu, J.; Zhao, H.; Jia, L.; Ye, Y.; Du, R.; Ren, X.; Li, H. Matrix Metalloproteinase 13: A Potential Intermediate between Low Expression of MicroRNA-125b and Increasing Metastatic Potential of Non-Small Cell Lung Cancer. Cancer Genet 2015, 208, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Bloomston, M.; Frankel, W.L.; Petrocca, F.; Volinia, S.; Alder, H.; Hagan, J.P.; Liu, C.G.; Bhatt, D.; Taccioli, C.; Croce, C.M. MicroRNA Expression Patterns to Differentiate Pancreatic Adenocarcinoma from Normal Pancreas and Chronic Pancreatitis. JAMA 2007, 297, 1901–1908. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Paris, P.L.; Chen, J.; Ngo, V.; Yao, H.; Frazier, M.L.; Killary, A.M.; Liu, C.G.; Liang, H.; Mathy, C.; et al. Next Generation Sequencing of Pancreatic Cyst Fluid MicroRNAs from Low Grade-Benign and High Grade-Invasive Lesions. Cancer Lett 2015, 356, 404–409. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.; Zhong, Y.; Wu, T.; Sheng, Y.; Dai, Y.; Xu, L.; Bao, C. Anti-Proliferative and Apoptosis-Promoting Effect of MicroRNA-125b on Pancreatic Cancer by Targeting NEDD9 via PI3K/AKT Signaling. Cancer Manag Res 2020, 12, 7363–7373. [Google Scholar] [CrossRef]
- Walter, B.A.; Valera, V.A.; Pinto, P.A.; Merino, M.J. Comprehensive MicroRNA Profiling of Prostate Cancer. J Cancer 2013, 4, 350–357. [Google Scholar] [CrossRef]
- Li, W.; Dong, Y.; Wang, K.J.; Deng, Z.; Zhang, W.; Shen, H.F. Plasma Exosomal MiR-125a-5p and MiR-141-5p as Non-Invasive Biomarkers for Prostate Cancer. Neoplasma 2020, 67, 1314–1318. [Google Scholar] [CrossRef]
- Konoshenko, M.Y.; Lekchnov, E.A.; Bryzgunova, O.E.; Zaporozhchenko, I.A.; Yarmoschuk, S.V.; Pashkovskaya, O.A.; Pak, S.V.; Laktionov, P.P. The Panel of 12 Cell-Free MicroRNAs as Potential Biomarkers in Prostate Neoplasms. Diagnostics (Basel) 2020, 10. [Google Scholar] [CrossRef]
- Shi, X.B.; Xue, L.; Yang, J.; Ma, A.H.; Zhao, J.; Xu, M.; Tepper, C.G.; Evans, C.P.; Kung, H.J.; White, R.W.D.V. An Androgen-Regulated MiRNA Suppresses Bak1 Expression and Induces Androgen-Independent Growth of Prostate Cancer Cells. Proc Natl Acad Sci U S A 2007, 104, 19983–19988. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.B.; Xue, L.; Ma, A.H.; Tepper, C.G.; Kung, H.J.; White, R.W.D. MiR-125b Promotes Growth of Prostate Cancer Xenograft Tumor through Targeting pro-Apoptotic Genes. Prostate 2011, 71, 538–549. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Wu, J.; Ren, J.; Vlantis, A.C.; Li, M. yue; Liu, S.Y.W.; Ng, E.K.W.; Chan, A.B.W.; Luo, D.C.; Liu, Z.; et al. MicroRNA-125b Interacts with Foxp3 to Induce Autophagy in Thyroid Cancer. Mol Ther 2018, 26, 2295–2303. [Google Scholar] [CrossRef]
- Wu, J.G.; Wang, J.J.; Jiang, X.; Lan, J.P.; He, X.J.; Wang, H.J.; Ma, Y.Y.; Xia, Y.J.; Ru, G.Q.; Ma, J.; et al. MiR-125b Promotes Cell Migration and Invasion by Targeting PPP1CA-Rb Signal Pathways in Gastric Cancer, Resulting in a Poor Prognosis. Gastric Cancer 2015, 18, 729–739. [Google Scholar] [CrossRef] [PubMed]
- Tong, Z.; Liu, N.; Lin, L.; Guo, X.; Yang, D.; Zhang, Q. MiR-125a-5p Inhibits Cell Proliferation and Induces Apoptosis in Colon Cancer via Targeting BCL2, BCL2L12 and MCL1. Biomed Pharmacother 2015, 75, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Fu, Q.; Liu, Z.; Pan, D.; Zhang, W.; Xu, L.; Zhu, Y.; Liu, H.; Xu, J. Tumor MiR-125b Predicts Recurrence and Survival of Patients with Clear-Cell Renal Cell Carcinoma after Surgical Resection. Cancer Sci 2014, 105, 1427–1434. [Google Scholar] [CrossRef] [PubMed]
- Testa, U.; Pelosi, E. MicroRNAs Expressed in Hematopoietic Stem/Progenitor Cells Are Deregulated in Acute Myeloid Leukemias. Leuk Lymphoma 2015, 56, 1466–1474. [Google Scholar] [CrossRef] [PubMed]
- Alemdehy, M.F.; Erkeland, S.J. MicroRNAs: Key Players of Normal and Malignant Myelopoiesis. Curr Opin Hematol 2012, 19, 261–267. [Google Scholar] [CrossRef]
- Mattie, M.D.; Benz, C.C.; Bowers, J.; Sensinger, K.; Wong, L.; Scott, G.K.; Fedele, V.; Ginzinger, D.; Getts, R.; Haqq, C. Optimized High-Throughput MicroRNA Expression Profiling Provides Novel Biomarker Assessment of Clinical Prostate and Breast Cancer Biopsies. Mol Cancer 2006, 5. [Google Scholar] [CrossRef]
- Iorio, M.V.; Ferracin, M.; Liu, C.G.; Veronese, A.; Spizzo, R.; Sabbioni, S.; Magri, E.; Pedriali, M.; Fabbri, M.; Campiglio, M.; et al. MicroRNA Gene Expression Deregulation in Human Breast Cancer. Cancer Res 2005, 65, 7065–7070. [Google Scholar] [CrossRef]
- Mar-Aguilar, F.; Luna-Aguirre, C.M.; Moreno-Rocha, J.C.; Araiza-Chávez, J.; Trevino, V.; Rodríguez-Padilla, C.; Reséndez-Pérez, D. Differential Expression of MiR-21, MiR-125b and MiR-191 in Breast Cancer Tissue. Asia Pac J Clin Oncol 2013, 9, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Liang, F.; Yang, M.; Tong, N.; Fang, J.; Pan, Y.; Li, J.; Zhang, X. Identification of Six Key MiRNAs Associated with Breast Cancer through Screening Large-Scale Microarray Data. Oncol Lett 2018, 16, 4159–4168. [Google Scholar] [CrossRef] [PubMed]
- Braicu, C.; Raduly, L.; Morar-Bolba, G.; Cojocneanu, R.; Jurj, A.; Pop, L.A.; Pileczki, V.; Ciocan, C.; Moldovan, A.; Irimie, A.; et al. Aberrant MiRNAs Expressed in HER-2 Negative Breast Cancers Patient. J Exp Clin Cancer Res 2018, 37. [Google Scholar] [CrossRef] [PubMed]
- Incoronato, M.; Grimaldi, A.M.; Mirabelli, P.; Cavaliere, C.; Parente, C.A.; Franzese, M.; Staibano, S.; Ilardi, G.; Russo, D.; Soricelli, A.; et al. Circulating MiRNAs in Untreated Breast Cancer: An Exploratory Multimodality Morpho-Functional Study. Cancers (Basel) 2019, 11. [Google Scholar] [CrossRef] [PubMed]
- Scott, G.K.; Goga, A.; Bhaumik, D.; Berger, C.E.; Sullivan, C.S.; Benz, C.C. Coordinate Suppression of ERBB2 and ERBB3 by Enforced Expression of Micro-RNA MiR-125a or MiR-125b. J Biol Chem 2007, 282, 1479–1486. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yan, L.X.; Wu, Q.N.; Du, Z.M.; Chen, J.; Liao, D.Z.; Huang, M.Y.; Hou, J.H.; Wu, Q.L.; Zeng, M.S.; et al. MiR-125b Is Methylated and Functions as a Tumor Suppressor by Regulating the ETS1 Proto-Oncogene in Human Invasive Breast Cancer. Cancer Res 2011, 71, 3552–3562. [Google Scholar] [CrossRef] [PubMed]
- Rajabi, H.; Jin, C.; Ahmad, R.; McClary, A.C.; Joshi, M.D.; Kufe, D. MUCIN 1 ONCOPROTEIN EXPRESSION IS SUPPRESSED BY THE MiR-125b ONCOMIR. Genes Cancer 2010, 1, 62–68. [Google Scholar] [CrossRef]
- Tang, F.; Zhang, R.; He, Y.; Zou, M.; Guo, L.; Xi, T. MicroRNA-125b Induces Metastasis by Targeting STARD13 in MCF-7 and MDA-MB-231 Breast Cancer Cells. PLoS One 2012, 7. [Google Scholar] [CrossRef]
- Metheetrairut, C.; Adams, B.D.; Nallur, S.; Weidhaas, J.B.; Slack, F.J. Cel-Mir-237 and Its Homologue, Hsa-MiR-125b, Modulate the Cellular Response to Ionizing Radiation. Oncogene 2017, 36, 512–524. [Google Scholar] [CrossRef]
- Wang, H.; Tan, G.; Dong, L.; Cheng, L.; Li, K.; Wang, Z.; Luo, H. Circulating MiR-125b as a Marker Predicting Chemoresistance in Breast Cancer. PLoS One 2012, 7. [Google Scholar] [CrossRef]
- Zhou, M.; Liu, Z.; Zhao, Y.; Ding, Y.; Liu, H.; Xi, Y.; Xiong, W.; Li, G.; Lu, J.; Fodstad, O.; et al. MicroRNA-125b Confers the Resistance of Breast Cancer Cells to Paclitaxel through Suppression of pro-Apoptotic Bcl-2 Antagonist Killer 1 (Bak1) Expression. J Biol Chem 2010, 285, 21496–21507. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Xu, F.; Huang, W.; Luo, S.Y.; Lin, Y.T.; Zhang, G.H.; Du, Q.; Duan, R.H. MiR-125a-5p Expression Is Associated with the Age of Breast Cancer Patients. Genet Mol Res 2015, 14, 17927–17933. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wei, Y.; Fan, X.; Zhang, P.; Wang, P.; Cheng, S.; Zhang, J. MicroRNA-125b as a Tumor Suppressor by Targeting MMP11 in Breast Cancer. Thorac Cancer 2020, 11, 1613–1620. [Google Scholar] [CrossRef] [PubMed]
- Abdollahzadeh, R.; Daraei, A.; Mansoori, Y.; Sepahvand, M.; Amoli, M.M.; Tavakkoly-Bazzaz, J. Competing Endogenous RNA (CeRNA) Cross Talk and Language in CeRNA Regulatory Networks: A New Look at Hallmarks of Breast Cancer. J Cell Physiol 2019, 234, 10080–10100. [Google Scholar] [CrossRef] [PubMed]
- Welch, J.D.; Baran-Gale, J.; Perou, C.M.; Sethupathy, P.; Prins, J.F. Pseudogenes Transcribed in Breast Invasive Carcinoma Show Subtype-Specific Expression and CeRNA Potential. BMC Genomics 2015, 16, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Zhu, K.; Wang, Q.; Wang, L. Analysis of Competitive Endogenous RNA Regulatory Network of Exosomal Breast Cancer Based on ExoRBase. Evolutionary Bioinformatics 2022, 18. [Google Scholar] [CrossRef] [PubMed]
- Rieger, M.A.; Ebner, R.; Bell, D.R.; Kiessling, A.; Rohayem, J.; Schmitz, M.; Temme, A.; Rieber, E.P.; Weigle, B. Identification of a Novel Mammary-Restricted Cytochrome P450, CYP4Z1, with Overexpression in Breast Carcinoma. Cancer Res 2004, 64, 2357–2364. [Google Scholar] [CrossRef]
- Yu, W.; Chai, H.; Li, Y.; Zhao, H.; Xie, X.; Zheng, H.; Wang, C.; Wang, X.; Yang, G.; Cai, X.; et al. Increased Expression of CYP4Z1 Promotes Tumor Angiogenesis and Growth in Human Breast Cancer. Toxicol Appl Pharmacol 2012, 264, 73–83. [Google Scholar] [CrossRef]
- Zheng, L.; Li, X.; Gu, Y.; Ma, Y.; Xi, T. Pseudogene CYP4Z2P 3’UTR Promotes Angiogenesis in Breast Cancer. Biochem Biophys Res Commun 2014, 453, 545–551. [Google Scholar] [CrossRef]
- Zheng, L.; Li, X.; Gu, Y.; Lv, X.; Xi, T. The 3’UTR of the Pseudogene CYP4Z2P Promotes Tumor Angiogenesis in Breast Cancer by Acting as a CeRNA for CYP4Z1. Breast Cancer Res Treat 2015, 150, 105–118. [Google Scholar] [CrossRef]
- Zheng, L.; Li, X.; Gu, Y.; Lv, X.; Xi, T. Correction to: The 3′UTR of the Pseudogene CYP4Z2P Promotes Tumor Angiogenesis in Breast Cancer by Acting as a CeRNA for CYP4Z1 (Breast Cancer Research and Treatment, (2015), 150, 1, (105-118), 10.1007/S10549-015-3298-2). Breast Cancer Res Treat 2020, 179, 521–522. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Li, X.; Meng, X.; Chou, J.; Hu, J.; Zhang, F.; Zhang, Z.; Xing, Y.; Liu, Y.; Xi, T. Competing Endogenous RNA Networks of CYP4Z1 and Pseudogene CYP4Z2P Confer Tamoxifen Resistance in Breast Cancer. Mol Cell Endocrinol 2016, 427, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Zheng, L.; Xin, Y.; Tan, Z.; Zhang, Y.; Meng, X.; Wang, Z.; Xi, T. The Competing Endogenous RNA Network of CYP4Z1 and Pseudogene CYP4Z2P Exerts an Anti-Apoptotic Function in Breast Cancer. FEBS Lett 2017, 591, 991–1000. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Guo, Q.; Xiang, C.; Liu, S.; Jiang, Y.; Gao, L.; Ni, H.; Wang, T.; Zhao, Q.; Liu, H.; et al. Transcriptional Factor Six2 Promotes the Competitive Endogenous RNA Network between CYP4Z1 and Pseudogene CYP4Z2P Responsible for Maintaining the Stemness of Breast Cancer Cells. J Hematol Oncol 2019, 12. [Google Scholar] [CrossRef]
- Zheng, L.; Guo, Q.; Xiang, C.; Liu, S.; Jiang, Y.; Gao, L.; Ni, H.; Wang, T.; Zhao, Q.; Liu, H.; et al. Correction to: Transcriptional Factor Six2 Promotes the Competitive Endogenous RNA Network between CYP4Z1 and Pseudogene CYP4Z2P Responsible for Maintaining the Stemness of Breast Cancer Cells (Journal of Hematology and Oncology (2019) 12 (23). https://doi.org/10.1186/S13045-019-0697-6). [CrossRef]
- Ching, Y.P.; Wong, C.M.; Chan, S.F.; Leung, T.H.Y.; Ng, D.C.H.; Jin, D.Y.; Ng, I.O.L. Deleted in Liver Cancer (DLC) 2 Encodes a RhoGAP Protein with Growth Suppressor Function and Is Underexpressed in Hepatocellular Carcinoma. J Biol Chem 2003, 278, 10824–10830. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Chen, N.T.; Shih, Y.P.; Liao, Y.C.; Xue, L.; Lo, S.H. DLC2 Modulates Angiogenic Responses in Vascular Endothelial Cells by Regulating Cell Attachment and Migration. Oncogene 2010, 29, 3010–3016. [Google Scholar] [CrossRef] [PubMed]
- Ullmannova, V.; Popescu, N.C. Expression Profile of the Tumor Suppressor Genes DLC-1 and DLC-2 in Solid Tumors. Int J Oncol 2006, 29, 1127–1132. [Google Scholar] [CrossRef]
- Hanna, S.; Khalil, B.; Nasrallah, A.; Saykali, B.A.; Sobh, R.; Nasser, S.; El-Sibai, M. StarD13 Is a Tumor Suppressor in Breast Cancer That Regulates Cell Motility and Invasion. Int J Oncol 2014, 44, 1499–1511. [Google Scholar] [CrossRef]
- D’Ippolito, E.; Plantamura, I.; Bongiovanni, L.; Casalini, P.; Baroni, S.; Piovan, C.; Orlandi, R.; Gualeni, A.V.; Gloghini, A.; Rossini, A.; et al. MiR-9 and MiR-200 Regulate PDGFRβ-Mediated Endothelial Differentiation of Tumor Cells in Triple-Negative Breast Cancer. Cancer Res 2016, 76, 5562–5572. [Google Scholar] [CrossRef]
- Hu, J.; Li, X.; Guo, X.; Guo, Q.; Xiang, C.; Zhang, Z.; Xing, Y.; Xi, T.; Zheng, L. The CCR2 3’UTR Functions as a Competing Endogenous RNA to Inhibit Breast Cancer Metastasis. J Cell Sci 2017, 130, 3399–3413. [Google Scholar] [CrossRef]
- Basak, P.; Leslie, H.; Dillon, R.L.; Muller, W.J.; Raouf, A.; Mowat, M.R.A. In Vivo Evidence Supporting a Metastasis Suppressor Role for Stard13 (Dlc2) in ErbB2 (Neu) Oncogene Induced Mouse Mammary Tumors. Genes Chromosomes Cancer 2018, 57, 182–191. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.; Liu, X.; Xiong, B.; Sun, Y. Homeobox B4 Inhibits Breast Cancer Cell Migration by Directly Binding to StAR-Related Lipid Transfer Domain Protein 13. Oncol Lett 2017, 14, 4625–4632. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Xiang, C.; Zhang, Z.; Zhang, F.; Xi, T.; Zheng, L. Displacement of Bax by BMF Mediates STARD13 3’UTR-Induced Breast Cancer Cells Apoptosis in an MiRNA-Depedent Manner. Mol Pharm 2018, 15, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Chen, Y.; Zhao, Q.; Xie, T.; Xiang, C.; Guo, Q.; Zhang, W.; Zhou, Y.; Yuan, Y.; Zhang, Y.; et al. A Positive TGF-β/MiR-9 Regulatory Loop Promotes the Expansion and Activity of Tumour-Initiating Cells in Breast Cancer. Br J Pharmacol 2023, 180. [Google Scholar] [CrossRef] [PubMed]
- Amirfallah, A.; Knutsdottir, H.; Arason, A.; Hilmarsdottir, B.; Johannsson, O.T.; Agnarsson, B.A.; Barkardottir, R.B.; Reynisdottir, I. Hsa-MiR-21-3p Associates with Breast Cancer Patient Survival and Targets Genes in Tumor Suppressive Pathways. PLoS One 2021, 16. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Xiang, C.; Li, X.; Guo, Q.; Gao, L.; Ni, H.; Xia, Y.; Xi, T. STARD13-Correlated CeRNA Network-Directed Inhibition on YAP/TAZ Activity Suppresses Stemness of Breast Cancer via Co-Regulating Hippo and Rho-GTPase/F-Actin Signaling. J Hematol Oncol 2018, 11. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zheng, L.; Zhang, F.; Hu, J.; Chou, J.; Liu, Y.; Xing, Y.; Xi, T. STARD13-Correlated CeRNA Network Inhibits EMT and Metastasis of Breast Cancer. Oncotarget 2016, 7, 23197. [Google Scholar] [CrossRef] [PubMed]
- Tang, F.; Zhang, R.; He, Y.; Zou, M.; Guo, L.; Xi, T. MicroRNA-125b Induces Metastasis by Targeting STARD13 in MCF-7 and MDA-MB-231 Breast Cancer Cells. PLoS One 2012, 7. [Google Scholar] [CrossRef]
- Hu, J.; Li, X.; Guo, X.; Guo, Q.; Xiang, C.; Zhang, Z.; Xing, Y.; Xi, T.; Zheng, L. The CCR2 3’UTR Functions as a Competing Endogenous RNA to Inhibit Breast Cancer Metastasis. J Cell Sci 2017, 130, 3399–3413. [Google Scholar] [CrossRef]
- Seillier, M.; Peuget, S.; Gayet, O.; Gauthier, C.; N’Guessan, P.; Monte, M.; Carrier, A.; Iovanna, J.L.; Dusetti, N.J. TP53INP1, a Tumor Suppressor, Interacts with LC3 and ATG8-Family Proteins through the LC3-Interacting Region (LIR) and Promotes Autophagy-Dependent Cell Death. Cell Death Differ 2012, 19, 1525–1535. [Google Scholar] [CrossRef]
- Seux, M.; Peuget, S.; Montero, M.P.; Siret, C.; Rigot, V.; Clerc, P.; Gigoux, V.; Pellegrino, E.; Pouyet, L.; N’Guessan, P.; et al. TP53INP1 Decreases Pancreatic Cancer Cell Migration by Regulating SPARC Expression. Oncogene 2011 30:27 2011, 30, 3049–3061. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Li, X.; Chou, J.; Xiang, C.; Guo, Q.; Zhang, Z.; Guo, X.; Gao, L.; Xing, Y.; Xi, T. StarD13 3’-Untranslated Region Functions as a CeRNA for TP53INP1 in Prohibiting Migration and Invasion of Breast Cancer Cells by Regulating MiR-125b Activity. Eur J Cell Biol 2018, 97, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Xiang, C.; Zhang, Z.; Zhang, F.; Xi, T.; Zheng, L. Displacement of Bax by BMF Mediates STARD13 3’UTR-Induced Breast Cancer Cells Apoptosis in an MiRNA-Depedent Manner. Mol Pharm 2018, 15, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Puthalakath, H.; Villunger, A.; O’Reilly, L.A.; Beaumont, J.G.; Coultas, L.; Cheney, R.E.; Huang, D.C.S.; Strasser, A. Bmf: A Proapoptotic BH3-Only Protein Regulated by Interaction with the Myosin V Actin Motor Complex, Activated by Anoikis. Science 2001, 293, 1829–1832. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Jia, Q.; Zhou, Y.; Jiang, X.; Song, L.; Wu, Y.; Wang, A.; Chen, W.; Wang, S.; Lu, Y. Tanshinone IIA Attenuates the Stemness of Breast Cancer Cells via Targeting the MiR-125b/STARD13 Axis. Exp Hematol Oncol 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Barkley, D.; Rao, A.; Pour, M.; França, G.S.; Yanai, I. Cancer Cell States and Emergent Properties of the Dynamic Tumor System. Genome Res 2021, 31, 1719. [Google Scholar] [CrossRef] [PubMed]
- Official Journal of the European Union. Notices From European Union Institutions, Bodies, Offices And Agencies. Council conclusions on personalised medicine for patients (2015/C 421/03). Issued on 17th December 2015. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/PDF/?uri=CELEX:52015XG1217 (accessed on 20 August 2023).
- Salari, P.; Larijani, B. Ethical Issues Surrounding Personalized Medicine: A Literature Review. Acta Med. Iran. 2017, 55, 209–217. [Google Scholar]
- Fiore, R.N.; Goodman, K.W. Precision Medicine Ethics: Selected Issues and Developments in next-Generation Sequencing, Clinical Oncology, and Ethics. Curr. Opin. Oncol. 2016, 28, 83–87. [Google Scholar] [CrossRef]
- Ramaswami, R.; Bayer, R.; Galea, S. Precision Medicine from a Public Health Perspective. Annu Rev Public Health 2018, 39, 153–168. [Google Scholar] [CrossRef]
- European Commission. Europe’s Beating Cancer Plan Communication from the Commission to the European Parliament and the Council. Issued in February 2022. Available online: https://health.ec.europa.eu/system/files/2022-02/eu_cancer-plan_en_0.pdf (accessed on 20 August 2023).
- European Commission. Directorate General for Research and Innovation. Conquering Cancer: Mission Possible; Publications Office: Luxembourg, 2020. Available online: https://op.europa.eu/en/publication-detail/-/publication/b389aad3-fd56-11ea-b44f-01aa75ed71a1/https://op.europa.eu/en/publication-detail/-/publication/b389aad3-fd56-11ea-b44f-01aa75ed71a1 (accessed on 29 August 2023).
- Hickman, J.A.; Tannock, I.F.; Meheus, L.; Hutchinson, L. The European Union and personalised cancer medicine. Eur. J. Cancer 2021, 150, 95–98. [Google Scholar] [CrossRef]
- Vos, S.; van Delden, J.J.M.; van Diest, P.J.; Bredenoord, A.L. Moral Duties of Genomics Researchers: Why Personalized Medicine Requires a Collective Approach. Trends Genet 2017, 33, 118–128. [Google Scholar] [CrossRef] [PubMed]
- Morsella, A.; Cadeddu, C.; Castagna, C.; Hoxhaj, I.; Sassano, M.; Wang, C.M.; Wang, L.; Klessova, S.; de Belvis, A.G.; Boccia, S.; et al. “Integrating China in the International Consortium for Personalized Medicine”: The Coordination and Support Action to Foster Collaboration in Personalized Medicine Development between Europe and China. Public Health Genom. 2021, 24, 310–314. [Google Scholar] [CrossRef] [PubMed]
- McGowan, M.L.; Settersten, R.A.; Juengst, E.T.; Fishman, J.R. Integrating genomics into clinical oncology: Ethical and social challenges from proponents of personalized medicine. Urol. Oncol. Semin. Orig. Investig. 2014, 32, 187–192. [Google Scholar] [CrossRef] [PubMed]
- European Commission. Smart Specialisation Platform. European Partnership on Personalised Medicine. TOPIC ID: HORIZON-HLTH-2023-CARE-08-01. Last Updated on Aug 9, 2023. Available online: https://ec.europa.eu/info/funding-tenders/opportunities/portal/screen/opportunities/topic-details/horizon-hlth-2023-care-08-01 (accessed on 30 August 2023).
- Venne, J.; Busshoff, U.; Poschadel, S.; Menschel, R.; Evangelatos, N.; Vysyaraju, K.; Brand, A. International Consortium for Personalized Medicine: An International Survey about the Future of Personalized Medicine. Per. Med. 2020, 17, 89–100. [Google Scholar] [CrossRef] [PubMed]
- Brothers, K.B.; Rothstein, M.A. Ethical, legal and social implications of incorporating personalized medicine into healthcare. Per. Med. 2015, 12, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Vicente, A.M.; Ballensiefen, W.; Jönsson, J.-I. How Personalised Medicine Will Transform Healthcare by 2030: The ICPerMed Vision. J. Transl. Med. 2020, 18, 180. [Google Scholar] [CrossRef] [PubMed]
- Sisodiya, S.M. Precision Medicine and Therapies of the Future. Epilepsia 2021, 62 Suppl 2, S90–S105. [Google Scholar] [CrossRef]
- Nimmesgern, E.; Norstedt, I.; Draghia-Akli, R. Enabling Personalized Medicine in Europe by the European Commission’s Funding Activities. Per. Med. 2017, 14, 355–365. [Google Scholar] [CrossRef]
- Horgan, D.; Bernini, C.; Thomas, P.P.M.; Morre, S.A. Cooperating on Data: The Missing Element in Bringing Real Innovation to Europe’s Healthcare Systems. Public Health Genom. 2019, 22, 77–101. [Google Scholar] [CrossRef]
- He, K.Y.; Ge, D.; He, M.M. Big Data Analytics for Genomic Medicine. Int. J. Mol. Sci. 2017, 18, 412. [Google Scholar] [CrossRef]
- Medenica, S.; Zivanovic, D.; Batkoska, L.; Marinelli, S.; Basile, G.; Perino, A.; Cucinella, G.; Gullo, G.; Zaami, S. The Future Is Coming: Artificial Intelligence in the Treatment of Infertility Could Improve Assisted Reproduction Outcomes-The Value of Regulatory Frameworks. Diagnostics (Basel) 2022, 12, 2979. [Google Scholar] [CrossRef]
- Sharrer, G.T. Personalized Medicine: Ethical Aspects. Methods Mol Biol 2017, 1606, 37–50. [Google Scholar] [CrossRef] [PubMed]
- Brittain, H.K.; Scott, R.; Thomas, E. The Rise of the Genome and Personalised Medicine. Clin Med (Lond) 2017, 17, 545–551. [Google Scholar] [CrossRef] [PubMed]
- Borry, P.; Bentzen, H.B.; Budin-Ljøsne, I.; Cornel, M.C.; Howard, H.C.; Feeney, O.; Jackson, L.; Mascalzoni, D.; Mendes, Á.; Peterlin, B.; et al. The Challenges of the Expanded Availability of Genomic Information: An Agenda-Setting Paper. J. Community Genet. 2018, 9, 103–116. [Google Scholar] [CrossRef] [PubMed]
- Hassan, M.; Awan, F.M.; Naz, A.; deAndrés-Galiana, E.J.; Alvarez, O.; Cernea, A.; Fernández-Brillet, L.; Fernández-Martínez, J.L.; Kloczkowski, A. Innovations in Genomics and Big Data Analytics for Personalized Medicine and Health Care: A Review. Int J Mol Sci 2022, 23, 4645. [Google Scholar] [CrossRef] [PubMed]
- Klein, M.E.; Parvez, M.M.; Shin, J.-G. Clinical Implementation of Pharmacogenomics for Personalized Precision Medicine: Barriers and Solutions. J Pharm Sci 2017, 106, 2368–2379. [Google Scholar] [CrossRef]
- Gavan, S.P.; Thompson, A.J.; Payne, K. The Economic Case for Precision Medicine. Expert Rev. Precis. Med. Drug Dev. 2018, 3, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Piersanti, V.; Consalvo, F.; Signore, F.; Del Rio, A.; Zaami, S. Surrogacy and “Procreative Tourism”. What Does the Future Hold from the Ethical and Legal Perspectives? Medicina (Kaunas) 2021, 57, 47. [Google Scholar] [CrossRef]
- Zhong, L.; Deng, B.; Morrison, A.M.; Coca-Stefaniak, J.A.; Yang, L. Medical, Health and Wellness Tourism Research-A Review of the Literature (1970-2020) and Research Agenda. Int J Environ Res Public Health 2021, 18, 10875. [Google Scholar] [CrossRef]
- Callier, S.L.; Abudu, R.; Mehlman, M.J.; Singer, M.E.; Neuhauser, D.; Caga-Anan, C.; Wiesner, G.L. Ethical, Legal, and Social Implications of Personalized Genomic Medicine Research: Current Literature and Suggestions for the Future. Bioethics 2016, 30, 698–705. [Google Scholar] [CrossRef]
- Varone, M.C.; Napoletano, G.; Negro, F. Decellularization and Tissue Engineering: Viable Therapeutic Prospects for Transplant Patients and Infertility? Eur Rev Med Pharmacol Sci 2021, 25, 6164–6166. [Google Scholar] [CrossRef] [PubMed]
- Baker, H.B.; McQuilling, J.P.; King, N.M.P. Ethical Considerations in Tissue Engineering Research: Case Studies in Translation. Methods 2016, 99, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Marchant, G.E.; Lindor, R.A. Personalized medicine and genetic malpractice. Genet. Med. 2013, 15, 921–922. [Google Scholar] [CrossRef] [PubMed]
- Ferrara, S.D.; Viel, G. [Personalized medicine and medical malpractice.]. G Ital Med Lav Ergon 2017, 39, 186–189. [Google Scholar] [PubMed]
- Marchant, G.; Barnes, M.; Evans, J.P.; LeRoy, B.; Wolf, S.M. ; LawSeq Liability Task Force From Genetics to Genomics: Facing the Liability Implications in Clinical Care. J. Law Med. Ethics 2020, 48, 11–43. [Google Scholar] [CrossRef] [PubMed]
- Ledford, H. US Personalized-Medicine Industry Takes Hit from Supreme Court. Nature 2016, 536, 382. [Google Scholar] [CrossRef] [PubMed]
- Donohue, K.E.; Gooch, C.; Katz, A.; Wakelee, J.; Slavotinek, A.; Korf, B.R. Pitfalls and Challenges in Genetic Test Interpretation: An Exploration of Genetic Professionals Experience with Interpretation of Results. Clin Genet 2021, 99, 638–649. [Google Scholar] [CrossRef] [PubMed]
- Stevens, Y.A.; Senner, G.D.; Marchant, G.E. Physicians’ Duty to Recontact and Update Genetic Advice. Per Med 2017, 14, 367–374. [Google Scholar] [CrossRef]
- Otten, E.; Plantinga, M.; Birnie, E.; Verkerk, M.A.; Lucassen, A.M.; Ranchor, A.V.; Van Langen, I.M. Is There a Duty to Recontact in Light of New Genetic Technologies? A Systematic Review of the Literature. Genet Med 2015, 17, 668–678. [Google Scholar] [CrossRef]
- Roberts, J.L.; Foulkes, A.L. GENETIC DUTIES. William Mary Law Rev 2020, 62, 143–211. [Google Scholar]
- Niida, A.; Iwasaki, W.M.; Innan, H. Neutral Theory in Cancer Cell Population Genetics. Mol Biol Evol 2018, 35, 1316–1321. [Google Scholar] [CrossRef] [PubMed]
- Ben-David, U.; Amon, A. Context Is Everything: Aneuploidy in Cancer. Nat Rev Genet 2020, 21, 44–62. [Google Scholar] [CrossRef] [PubMed]
- Newburger, D.E.; Kashef-Haghighi, D.; Weng, Z.; Salari, R.; Sweeney, R.T.; Brunner, A.L.; Zhu, S.X.; Guo, X.; Varma, S.; Troxell, M.L.; et al. Genome Evolution during Progression to Breast Cancer. Genome Res 2013, 23, 1097–1108. [Google Scholar] [CrossRef] [PubMed]
- Murakami, F.; Tsuboi, Y.; Takahashi, Y.; Horimoto, Y.; Mogushi, K.; Ito, T.; Emi, M.; Matsubara, D.; Shibata, T.; Saito, M.; et al. Short Somatic Alterations at the Site of Copy Number Variation in Breast Cancer. Cancer Sci 2021, 112, 444–453. [Google Scholar] [CrossRef] [PubMed]
- Soysal, S.D.; Tzankov, A.; Muenst, S.E. Role of the Tumor Microenvironment in Breast Cancer. Pathobiology 2015, 82, 142–152. [Google Scholar] [CrossRef] [PubMed]
- Hinshaw, D.C.; Shevde, L.A. The Tumor Microenvironment Innately Modulates Cancer Progression. Cancer Res 2019, 79, 4557–4567. [Google Scholar] [CrossRef] [PubMed]
- Tan, Z.; Kan, C.; Sun, M.; Yang, F.; Wong, M.; Wang, S.; Zheng, H. Mapping Breast Cancer Microenvironment Through Single-Cell Omics. Front Immunol 2022, 13. [Google Scholar] [CrossRef] [PubMed]
- Boo, L.; Ho, W.Y.; Ali, N.M.; Yeap, S.K.; Ky, H.; Chan, K.G.; Yin, W.F.; Satharasinghe, D.A.; Liew, W.C.; Tan, S.W.; et al. Phenotypic and MicroRNA Transcriptomic Profiling of the MDA-MB-231 Spheroid-Enriched CSCs with Comparison of MCF-7 MicroRNA Profiling Dataset. PeerJ 2017, 5. [Google Scholar] [CrossRef]
- Ahram, M.; Mustafa, E.; Zaza, R.; Abu Hammad, S.; Alhudhud, M.; Bawadi, R.; Zihlif, M. Differential Expression and Androgen Regulation of MicroRNAs and Metalloprotease 13 in Breast Cancer Cells. Cell Biol Int 2017, 41, 1345–1355. [Google Scholar] [CrossRef]
Figure 1.
schematic representation of human miR-125 illustrating the structure of the pre-miRNA of both miR-125a and miR-125b, and their sequence. Color codes: red highlight for the 5p form, blue highlight for the 3p form. Numbers before and after the sequences indicate the number of nucleotides trimmed away from the mature miRNA. Data re-trieved and partially modified from miRTarBase v9 update 2022 [78,79]
Figure 1.
schematic representation of human miR-125 illustrating the structure of the pre-miRNA of both miR-125a and miR-125b, and their sequence. Color codes: red highlight for the 5p form, blue highlight for the 3p form. Numbers before and after the sequences indicate the number of nucleotides trimmed away from the mature miRNA. Data re-trieved and partially modified from miRTarBase v9 update 2022 [78,79]
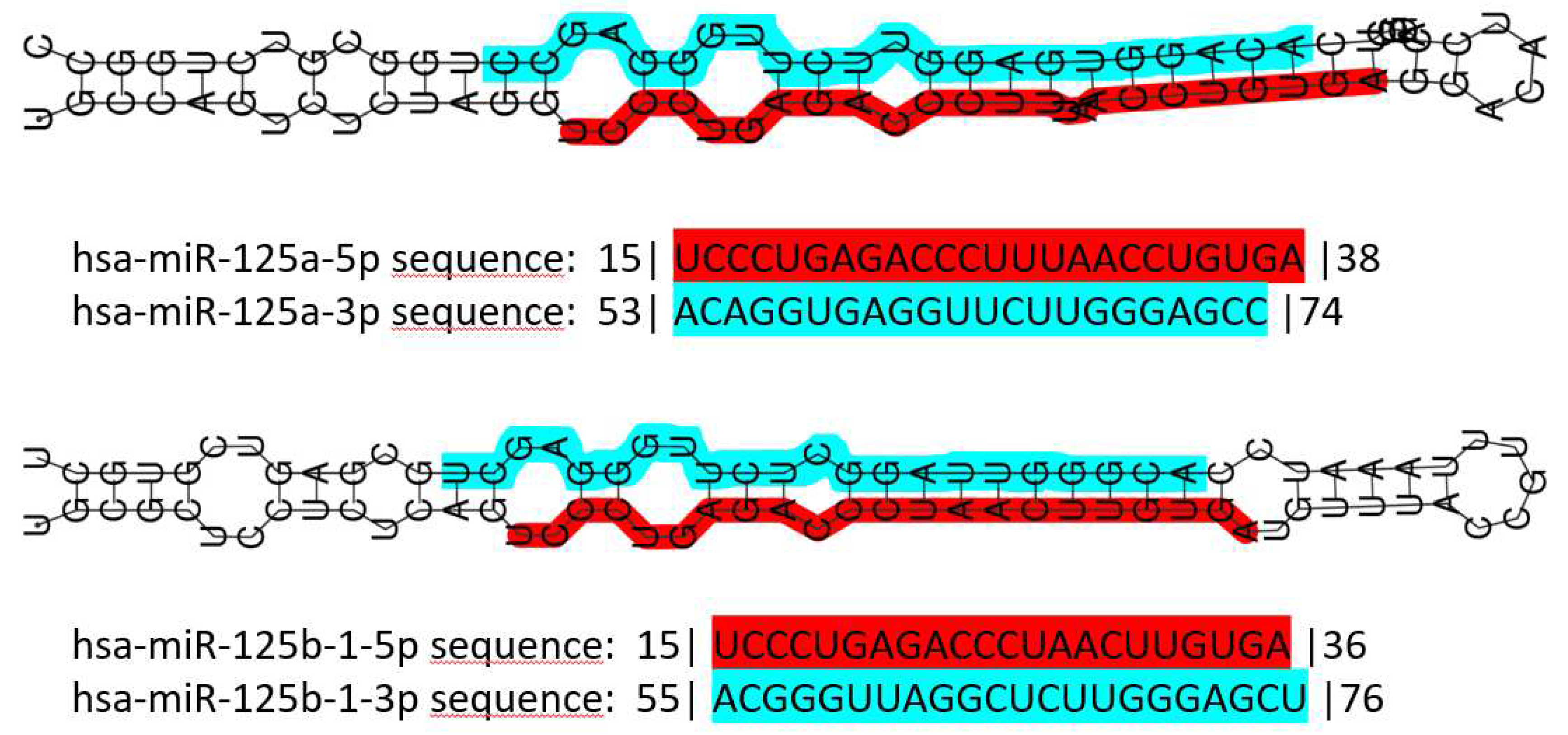
Figure 2.
schematic representation of two miR-125-centered ceRNETs in BC. A: a simple ceRNET having only one axis, where a pseudogene mRNA (CYP4Z2P) inhibits miR-125a-3p action on the target mRNA (CYP4Z1) by sponging it, thus enhancing CYP4Z1 expression. B: a more complex ceRNET in which the interaction between miR-125b and STARD13 mRNA control the expression of multiple target mRNAs. See text for references and further explanations.
Figure 2.
schematic representation of two miR-125-centered ceRNETs in BC. A: a simple ceRNET having only one axis, where a pseudogene mRNA (CYP4Z2P) inhibits miR-125a-3p action on the target mRNA (CYP4Z1) by sponging it, thus enhancing CYP4Z1 expression. B: a more complex ceRNET in which the interaction between miR-125b and STARD13 mRNA control the expression of multiple target mRNAs. See text for references and further explanations.
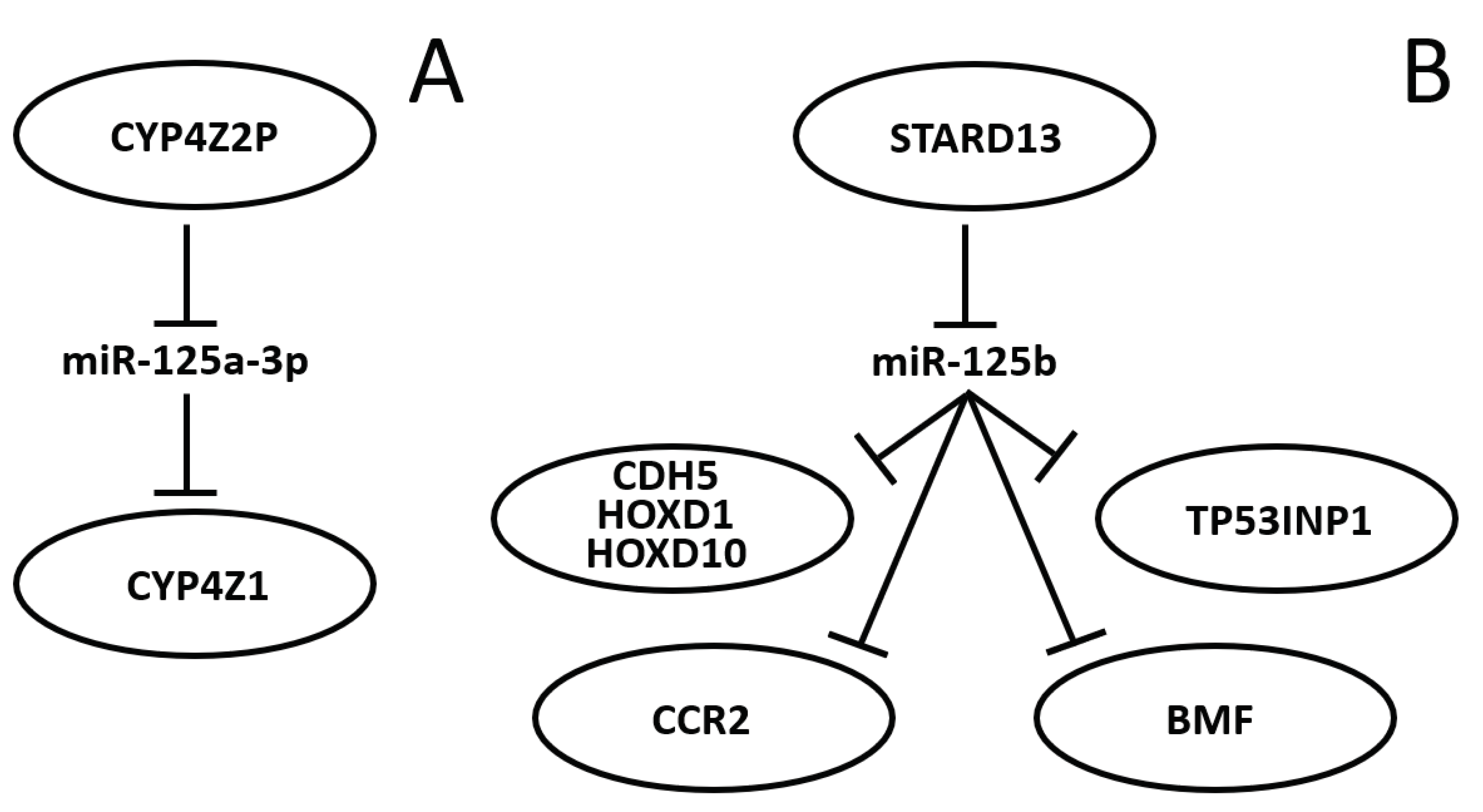

Table 1.
TNM staging for BC.
| Tumor | Node | Metastasis | |||
|---|---|---|---|---|---|
| Tx | no primary tumor information | Nx | not assessable | Mx | not assessed |
| T0 | no primary tumor evidence | N0 | no clinically positive nodes | M0 | no evidence |
| TIS | carcinoma in-situ (primary sites) | N1 | single, ipsilateral, size <3 cm | M1 | metastasis present at distance |
| T1 | size <2 cm | N2a | single, ipsilateral, size 3-6 cm | ||
| T2 | size 2 to 4cm | N2b | multiple, ipsilateral, size <6 cm | ||
| T3 | size >4 cm | N3 | massive/ipsilateral/bilateral/controlateral | ||
| T4 | size >4 cm, pterygoid muscle, base of tongue or skin involved | N3a | ipsilateral node(s), one more than 6 cm | ||
| N3b | bilateral | ||||
| N4 | controlateral | ||||
Table 2.
Summation of stakeholders’ roles and outcomes to be pursued through targeted measures aimed at personalized medicine development, uptake and awareness as defined by the European Partnership on Personalised Medicine.
Table 2.
Summation of stakeholders’ roles and outcomes to be pursued through targeted measures aimed at personalized medicine development, uptake and awareness as defined by the European Partnership on Personalised Medicine.
| Stakeholder(s) involved | Outcome(s) to be pursued |
|---|---|
| European countries/regions in cooperation with international partners | Broad-ranging collaborative research efforts for the development of innovative personalized medicine approaches regarding prevention, diagnosis and treatment |
| Healthcare authorities, policymakers |
|
| Healthcare industries and manufacturers, policymakers |
|
| Healthcare providers and professionals | Improvement of health outcomes, prevention of diseases and foster population health through the equitable and cost-effective implementation of personalized medicine |
| Connected local/regional ecosystems of stakeholders | Fostering the uptake of successful innovations in personalized medicine, in order to improve healthcare outcomes and strengthening European competitiveness |
| Citizens, patients and healthcare professionals | Broader knowledge of personalized medicine and resulting better involvement in its implementation through feedback |
| All stakeholders | Better cooperation and establishment of networks of national and regional knowledge and awareness-building hubs for personalized medicine. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



