
Submitted:
23 September 2023
Posted:
26 September 2023
You are already at the latest version
Abstract
Abstract: Sarcopenia poses a significant challenge to public health and can severely impact the quality of life of aging populations. Despite extensive efforts to study muscle degeneration using traditional animal models, there is still a lack of effective diagnostic tools, precise biomarkers, and treatments for sarcopenia. Zebrafish models have emerged as powerful tools in biomedical research, providing unique insights into age-related muscle disorders like sarcopenia. The advantages of using zebrafish models include their rapid growth outside of the embryo, optical transparency during early developmental stages, high reproductive potential, ease of husbandry, compact size, and genetic tractability. By deepening our understanding of the molecular processes underlying sarcopenia, we may develop novel diagnostic tools and effective treatments that can improve the lives of aging individuals affected by this condition. This review aims to explore the unique advantages of zebrafish as a model for sarcopenia research, highlight recent breakthroughs, outline potential avenues for future investigations, and emphasize the distinctive contributions that zebrafish models offer. Our research endeavors to contribute significantly to addressing the urgent need for practical solutions to the impact of sarcopenia on aging populations, ultimately striving to enhance the quality of life for individuals affected by this condition.
Keywords:
Sarcopenia
; Zebrafish
; Age-related muscle deficits
; Aging
; Biomarkers
; Electrical Impedance Myography
1. Content
Zebrafish (Danio rerio) has emerged as a highly promising animal model in the realm of biomedical research, offering invaluable insights into a diverse array of human diseases, including but not limited to skeletal muscle atrophy, aging, and sarcopenia. This diminutive aquatic organism has earned its place as an indispensable model organism for investigating crucial pathways and therapeutics associated with various facets of human health, such as neurodegenerative disorders, skeletomuscular ailments, and cardiovascular conditions [1,2,3,4]. Genetically, the zebrafish genome shares an astonishing 70% similarity with the human genome, with approximately 82% of genes associated with human ailments having identifiable orthologues in zebrafish [5,6].
Primary sarcopenia, a condition characterized by age-related loss of muscle mass and function, that significantly compromises the quality of life in affected individuals, has been extensively studied utilizing model organisms, including mice, rats, flies, and worms. These models have facilitated the identification of therapeutic strategies and genetic modifications aimed at ameliorating muscle-related degenerative processes in aging individuals [6]. However, practical diagnostic modalities, accurate biomarkers for risk stratification or follow-up, and effective treatments for primary Sarcopenia are still elusive. Zebrafish models offer a variety of muscle growth mechanisms, bridging the evolutionary gap between fly and mouse models. Despite the apparent senescence and fixed lifetime of mammalian skeletal muscle, fish tissues maintain significant plasticity even at the adult stage. This allows researchers to examine the metabolic functions such as peroxisome-resident proteins using morpholino-mediated knockdown or transcription activator-like effector nucleases [6,7,8].
Zebrafish, in particular, has assumed a significant role in biochemical, physiological, genetic, and developmental research due to several inherent advantages: (i) rapid ex utero growth, with adulthood attained within a mere three months, (ii) optical transparency of embryos and early larvae, (iii) high reproductive potential, (iv) compact size at approximately 1.5 inches when fully matured, and a short generation time, (v) facile husbandry and reproduction, and (vi) the ability for genetic manipulation [9,10,11,12] (Figure 1).
The primary objective of this study is to elucidate the manifold advantages of employing zebrafish as a model for sarcopenia, shedding light on its resemblances and disparities relative to the human skeletal muscle. This manuscript will also critically evaluate recent breakthroughs in the field, delineate potential avenues for future research, and underscore the distinctive contributions that the zebrafish model offers in comparison to other fish models.
This literature review employs a comprehensive method involving systematic database searches, including PubMed, Scopus, and Web of Science, followed by assessment of the relevance and quality of identified research articles.
2. Zebrafish as an Animal Model of Sarcopenia
Within the context of skeletal muscle research, Zebrafish exhibits striking parallels with the human skeletal muscle system, including the presence of satellite-like cells responsible for muscle repair and regeneration, and the anatomical distinction between fast and slow muscles. The fast muscles of zebrafish are situated around the vertebrae, while the slow muscles reside beneath the body’s surface [9,12]. Moreover, zebrafish harbors a collection of orthologous genes analogous to human Myogenic Regulatory Factors (MRFs) [4]. Skeletal muscle, constituting a substantial portion (35-50%) of the human body’s volume, plays an integral role in physical mobility and critical metabolic processes, such as basal metabolic rate, glucose uptake, and lipid metabolism [3,4]. The maintenance of muscle mass within myofibers hinges on a delicate balance between protein synthesis and breakdown, with disruptions in this equilibrium resulting in reduced muscle mass, myofiber size, and subsequently, metabolic disorders and muscular atrophy [13].
In light of the remarkable progress achieved in recent years, zebrafish has solidified its status as a potential model for elucidating the molecular underpinnings of sarcopenia and conducting rigorous therapeutic screening. Recent research exemplifies the incorporation of cutting-edge methodologies, such as Electrical Impedance Myography, showcasing substantial advancements within this field.
3. Current challenges and opportunities
3.1. Aging and Sarcopenia
As we age, our bodies endure a variety of changes. If any of these alterations continue for an extended period of time, they may contribute to the development of various illnesses [14]. Despite the possible participation of various molecular pathways in the aging process, we still do not fully comprehend its complexities [15]. Many nutritional (such as vitamins and minerals) and hormonal deficits have been identified during natural aging, and these shortcomings are suspected to be contributing factors to chronic diseases that are particularly prevalent among the elderly (such as sarcopenia and neurodegenerative disease) [16,17]. Among nutritional factors are magnesium, zinc, vitamin D, vitamin B, and so forth; however, the specific link between these deficits and illnesses in elderly individuals is in doubt [17,18,19,20,21]. In the context of aging, several investigations in various Zebrafish tissues (such as cardiac, neural, and skeletomuscular tissues) have been implemented in order to find the fundamental mechanisms of aging due to its advantages over other animal models and strong similarities to humans [3,22,23,24].
Sarcopenia is defined as chronic loss of skeletal muscle mass and efficiency throughout the body, and can be categorized into two forms: primary (age-associated) and secondary (illness-associated) [25,26]. Primary sarcopenia is caused by aging alone, whereas secondary sarcopenia occurs in the presence of other disorders along with aging [27,28]. This age-related muscle atrophy may cause falls and subsequent fractures, osteoporosis, frailty, metabolic shifts, increased heart disease, immunodeficiency, and various other disorders among elderly individuals [29,30].
Aging results in alterations to muscle mass and function in virtually everyone, but the rapidity at which these changes occur varies from person to person for a multitude of factors, the majority of which are still unknown (Figure 2) [31,32]. (Figure 1) Its prevalence in the elderly population can vary from 10% to 27% [33], and with the increasing elderly population, it is predicted to become a significant health issue by 2030 [34]. Revealing the economic facet, approximated $40.4 billion USD as the potential hospitalization cost for sarcopenia patients [35]. In this context, Zebrafish presents a distinct path to hope; their potential utility in unraveling the complexity of sarcopenia and its underlying processes offers a viable path for cutting-edge treatments and therapeutic approaches.
3.2. Pathogenesis
Sarcopenia has been associated with several processes, including oxidative stress, mitochondrial dysfunction, senescent satellite cell accumulation, protein homeostasis imbalance, inappropriate diet, NAD+ deterioration, increased myostatin and GDF-15, immunosenescence, gut microbiota alteration, and hormonal deviations [25,27,36,37,38,39,40] (Figure 3). Despite extensive investigations in these areas, more detailed research efforts are still needed. As Zebrafish gains increasing momentum in the field of aging, more different Zebrafish models and experiments will be performed in these areas to validate and further expand the animal model with different strains and high-output platforms. Other possible biomarkers for cancer, disorders of neurological development, hepatotoxicity, and other conditions also have been identified in zebrafish, providing novel insights into the identification of molecular pathways and the early detection of these pathologies [41,42,43].
Sustained oxidative stress may contribute to the progression of sarcopenia by influencing synaptic junctions (reducing acetylcholine and synaptic effectiveness), protein balance (increased protein degradation and decreased protein synthesis), and most importantly, mitochondria, which are the main source of ROS [44,45]. Dysfunctioning mitochondria are proposed to be caused by the inactivation of the SIRT1-PGC1α axis, crucial for mitochondrial biogenesis [46]. Subsequently, the accumulation of persistent oxidative damage and the resulting mitochondrial malfunction collide with the potential of telomere breakage, creating an environment permissive to cellular senescence [37].
Satellite cells (SCs) in skeletal muscle have regenerative capabilities but this ability may diminish with age due to various factors (such as telomere shrinkage, defective mitochondria, and so on), leading to cellular senescence [47,48]. Different regulatory pathways, including p53-p21CIP1-CDK2 and p16Ink4a-CDK4/6, play a role in the initiation of senescence [49,50]. The contribution of p53 to sarcopenia is still debated [51]. The total count of SCs during aging remains uncertain due to conflicting findings [52]. As previously noted, hyperplastic development persists in zebrafish skeletal muscle throughout life (unlike humans) due to specific cells that generate Pax7, similar to human satellite cells. These Pax7+ cells were additionally identified to be situated similarly to human SCs between the basement membrane and the sarcolemma [53]. Further investigations uncovered a zebrafish ortholog to mammalian FOXM1 that may have comparable functions in muscle tissue maintenance such as cellular senescence regulation [54]. These fish also can be employed to investigate the association between telomere shrinkage and sarcopenia as zebrafish and human telomeres are remarkably comparable [55]. Zebrafish models have been developed to simulate age-related disorders, including telomere malfunction, lipofuscin buildup, vitamin E depletion, and obesity-associated sarcopenia [56,57,58].
Protein homeostasis (production and breakdown) in skeletal muscle includes numerous mechanisms, and its disruption contributes to the occurrence of sarcopenia [59]. Myostatin, synthesized by various tissues, including skeletal muscle, suppresses protein synthesis by inhibiting the AKT-mTOR axis after binding to the activin type IIB receptor [60]. Increased myostatin has been associated with aging and is considered a possible mechanism for sarcopenia development [61]. Deactivation of the AKT-mTOR axis not only suppresses protein synthesis but also accelerates protein degradation through the elimination of AKT’s inhibitory impact on Forkhead box O (FoxO) [60]. Notably, myostatin depletion in mutant zebrafish resulted in a growth in muscle tissue as well as a metabolic shift toward lipid utilization. Although the particular mechanism of myostatin on skeletal muscle differs across humans and zebrafish in some domains, the study provides insight into the general molecular interplay throughout metabolic alterations and muscle development [62].
In conclusion, several processes, such as long-term low-level inflammation, hormonal shifts, cellular senescence, mitochondrial failure, altered myokines, and others, may contribute to sarcopenia, but the precise root cause remains unknown. Although a substantial number of experiments are required to unravel these intricate phenomena, using zebrafish as a high-output potent model has the capacity to elucidate the intricate in vivo mechanisms underlying the development of Sarcopenia, facilitating the identification of novel pathways and mediators essential for an in-depth understanding of these intricate mechanisms.
3.3. Diagnosis:
Sarcopenia is diagnosed by assessing muscle quantity (mass) and quality (strength and performance). Various tests are used for strength analysis, physical performance evaluation, and assessing muscle mass through imaging technologies [26,63]. Despite the available assessments, there are limitations in specific diagnostic domains. Certain exercise or nutrition programs may show a beneficial impact on some evaluations but not others, reducing their reliability [64,65]. Thus, improving diagnostic tools and procedures is urgent.
The decline in skeletal muscle size and strength is not proportional. Studies show that muscle mass decreases by 6% every ten years after mid-age, while strength declines faster, and the rate is twice as high in men compared to women [66,67]. These findings indicate measuring mass alone may not reliably detect sarcopenia in its early stages. Previous investigations suggested that reduced muscle mass accounts for a small percentage of the decline in muscular strength, and other areas require additional research since, in particular cases, conserving or improving muscle mass did not result in the anticipated rise in muscle strength [66].
Henderson et. al. used the sarcopenic Zebrafish animal model to address the complex interplay between muscle degeneration due to aging, and oxidative damage resulting from vitamin E deficiency, particularly in the context of skeletal muscle metabolism. The study employs metabolomics to analyze the skeletal muscle of aging zebrafish subjected to long-term vitamin E deficiency. It reveals distinct metabolic changes associated with aging and vitamin E deficiency, indicating overlapping but unique alterations in metabolic pathways, underscoring the need for further comprehensive investigation using confirmatory approaches to understand their combined impact on muscle health [57]. Several biomarkers have been proposed to diagnose sarcopenia early, including inflammation-related biomarkers (TNF-α and CRP), nutritional-related biomarkers (insulin-like growth factor-1 and vitamin D), and genetic biomarkers (Rho GTPase activating protein 36 and family sequence similarity 171, member protein A1) [68,69,70]. However, there is no gold standard biomarker for its etiology, requiring further research. Carbonic anhydrase 2 (ca2), selenium-binding protein 1 (selenbp1), and myosin regulatory light chain 2 a and b (myl2a/b) have been proposed as age markers in zebrafish due to their striking resemblance in function and structure in their mammalian orthologs. It should be emphasized that age biomarkers may have altered in opposite ways in various tissues, for example, cardiac myl2b has been elevated till middle age in contrast to muscle myl2b in zebrafish [71].
3.4. Treatment
Resistance training (RT) with a protein-rich diet is considered an effective starting point for managing age-associated skeletal muscle degradation [72,73]. Different exercise variables and the patient’s general health condition complicate implementing research findings in clinical practice [74,75], requiring researchers to develop ideal exercise instructions for sarcopenia therapy [76]. Additionally, exercise training in zebrafish has been found to lead to white muscle fiber hypertrophy and increased expression of genes involved in muscle synthesis and energy metabolism [77]. Aged zebrafish exhibit deficits in swimming performance due to weaker and shorter movements, indicating impairments in motor function [78].
Nutritional supplementation also plays a crucial role in sarcopenia management [73]. For example, vitamin D supplementation has shown conflicting outcomes in different studies, necessitating further investigation [19]. Whey protein peptide intake has been advantageous in both the prevention and management of sarcopenia [79,80]. Similarly, essential amino acid supplementation might trigger myocyte renewal and specialization [81,82].
Combining exercise with a healthy diet can induce mitochondrial biogenesis, increase satellite cell activity, and inhibit inflammatory cytokines resulting in enhanced protein synthesis and decreased protein breakdown [83].
Pharmacological therapies are also considered for patients unable to undergo exercise interventions [84]. Although several therapies have been suggested, there are currently no FDA-approved medications for sarcopenia [85]. In an interesting study, the protective properties of Rhus coriaria extracts on cells subjected to oxidative damage were explored. It was shown that both human myoblast and zebrafish embryos demonstrated a slowing or halting of muscle atrophy [86]. This highlights the significance of using these fish in the development of pharmaceuticals.
In addition to the techniques described above, stem cell and mitochondrial transplantation have demonstrated an improvement in muscle mass and strength [52,87]. However, these approaches face challenges, such as the difficulty of isolation and transferring procedures, which may result in transferring malfunctioning mitochondria and be detrimental to the tissue. Moreover, the specific molecular effects of the transferring paths are not well-known [88]. Therefore, further exploration and improvement are needed to successfully treat this disorder using these approaches.
In summary, given the existing lack of sufficient sarcopenia management and prevention measures, the need to discover and deliver effective approaches to address this issue continues unabated.
4. Latest Zebrafish Advances in Sarcopenia
Recent evidence suggests Electrical Impedance Myography (EIM) as a feasible screening tool for skeletal muscle age-associated illnesses due to its affordability, simplicity of use, and ease of transport over other imaging tools [89]. EIM relies on the composition and subsequent conductivity and electrical activity of the targeted muscle since it has been demonstrated that muscle conductivity and derived charge can alter in the presence of additional variables such as muscular atrophy and edema [90]. EIM has been performed on zebrafish using two separate techniques (surface and needle) to examine muscle tissue characteristics. Although the core principles of both approaches are the same, they differ in terms of accuracy and invasiveness. In general, when compared to alternative methods such as histological analysis and physical performance assessments in the study of zebrafish muscle tissue, EIM has proven to be less invasive, providing faster results, and greater ease of application, establishing its significance for sarcopenia research [78,91].
Rutkove et. al. emphasize the lack of pharmacological treatments for sarcopenia and the impracticality of high-throughput chemical screens in mammalian models. (Table 1) A novel surface electrical impedance myography (sEIM) platform has been lately introduced to quantitatively and noninvasively assess skeletal muscle health in adult zebrafish, effectively detecting muscle atrophy in aged zebrafish and correlating these measurements with established morphometric parameters. This demonstrates the utility of sEIM in studying the genetic factors contributing to muscle atrophy, offering potential applications as a noninvasive “virtual biopsy” in zebrafish for muscle and gerontology research. The study also investigated the impact of genetic deletion of gpr27, an orphan G-protein coupled receptor (GPCR), on skeletal muscle atrophy in aged zebrafish. Surface electrical impedance myography (sEIM) and histological analysis revealed that the absence of gpr27 exacerbated muscle atrophy, highlighting its role in modulating muscle health in aging animals [81].
Exercise has been the cornerstone of treatment to prevent the onset/progression and treat sarcopenia. This has also been evaluated in Zebrafish. Exercise is known to effectively delay or prevent the onset and progression of sarcopenia, yet the underlying molecular mechanisms remain poorly understood. Sun et al. have utilized aged Zebrafish as a valuable model to elucidate the pathological changes associated with sarcopenia, including reduced muscle fiber cross-sectional area, imbalanced protein synthesis and degradation, increased oxidative stress, and mitochondrial dysfunction. After subjecting these zebrafish to eight weeks of exercise intervention, attenuation of these pathological changes was observed, particularly in relation to the activation of the AMPK/SIRT1/PGC-1α axis and downregulation of 15-PGDH, providing potential therapeutic targets for the treatment of age-related sarcopenia through exercise intervention. Thus, confirming in the ZF model the benefit of exercise in terms of mitochondrial function [92].
5. Future perspectives:
5.1. Novel pathways and molecules for improving mitochondrial function.
Sarcopenia and aging have been associated with progressive mitochondrial dysfunction that is characterized by deficiencies in mitochondrial content within the muscle in addition to decreased mitochondrial biogenesis, impaired mitochondrial permeability transition pore function and increased mtDNA mutations [92,93]. Zebrafish is an excellent model to study muscle-related degenerations and sarcopenia due to their short lifespan, and gradual aging like humans while having conserved aging markers as in humans [94]. For instance, the zebrafish myosepta mimics the human tendon, which both decline with age therefore, the zebrafish model can be a perfect tool in studying mitochondrial tendon biomechanics and aging [94,95]. Due to these similarities, exercise intervention regimens related to mitochondria can be applied in the zebrafish. For example, it has been demonstrated that exercise significantly improved the expression of mitochondrial function biogenesis-related proteins while decreasing the expression of mitochondrial fission-related proteins in the zebrafish model [92]. In another study, aerobic exercise-induced in zebrafish, repressed miR-128, has been reported to impair mitochondrial biogenesis by inhibiting PGC-1α [58,96]. Another study corroborated the importance of zebrafish as a model in identifying potential novel pathways for mitochondrial function. Here, the findings revealed that a high fat diet reduced exercise capacity in a zebrafish model of obesity through pathways of decreased mitochondrial function such as decreases in the expression of PGC-1α, AMPK and SIRT-1.
Taken together, these examples of studies from the zebrafish model highlight the potential of zebrafish in pioneering novel pathways and molecules for improving mitochondrial function with the aim of developing therapies. In addition, while a lot of interventions such as supplementation with NAD as well as other dietary improvements have been targeted to larger rodent models like mice, none has been tested on zebrafish for the improvement on aged muscle, despite the many advantages of the zebrafish model highlighted in this review as well as in many others. Therefore, focus should now be placed on the zebrafish model in understanding pathways related to mitochondrial function.
5.2. Identification of novel genes and candidates in population-wide studies
Genome-wide associations (GWAS) identifies genes and variants associated with or causal of disease, yet we need animal models to identify and dissect the function in vivo. Regarding sarcopenia, there is a strong and substantial heritability estimated to be at 50% in the transmission of defective genes through the genetic component [97]. Because there are no reliable markers to identify sarcopenia, GWAS studies are very much justified such that they can be used to identify single nucleotide polymorphisms (SNPs) related to sarcopenia. An additional advantage relating to sarcopenia and GWAS studies is that as a teleost model species, its genome has already been sequenced [3]. A key strength in the zebrafish genome is the existence of gene ohnologs or gene duplicates [3]. The zebrafish ohnologs have been found to be advantageous in the study of muscle diseases such as sarcopenia because they result in mild phenotypes when deleted as opposed to embryonic lethality as observed in mammalian paralogs [3]. Although not related to sarcopenia, much recently, GWAS studies are now being employed in zebrafish models [98], and already there is the identification of novel muscle pathways such as MPP7 that affect bone mineralization [99]. With the advent of new tools such as CRISPR gene editing and zebrafish’s smaller genome size and ohnologs, the zebrafish is an ideal candidate to be utilized in the identification of novel genes and candidates highlighted in population-wide studies.
5.3. Complimentary with other animal models
The short-lived multifaceted Turquoise Killifish, has continued to rapidly emerge as a more practical experimental candidate for studying aging and diapause [100,101,102]. The use of this animal model will increase in the next decades as complimentary to current well-established vertebrate models like zebrafish and mice, as the Turquoise Killfish offers a compressed lifespan (4-6 months vs 2-3 years) and a long diapause state [102,103]. It was demonstrated that short-lived Turquoise Killifish upregulated H3K27 methylation complexes leading to chromatin hoarding and downregulated H3K27 acetylation complexes in the aging process [104]. Recently, a comparative study between short-lived Turquoise killifish and the longer-lived zebrafish demonstrated initial central regulation of the NPY neuropeptide in aging models with peripheral localization in both aged models elucidating new mechanisms of further study for both fish species [105].
6. Conclusion:
Recent advances in the Zebrafish animal model have highlighted the potential role of this organism in elucidating the pathogenesis, diagnosis, monitoring, and treatment of Sarcopenia. As deterioration of muscle function is expected with aging, a better understanding of the impact of aging in muscle can help not only in Sarcopenia management, but also in reducing the trends of muscle decay with age. The gradual implementation of advanced molecular techniques in Zebrafish and the recent validation of numerous models of Sarcopenia, will build the foundations for a better understanding and improved management of Sarcopenia in the upcoming decades.
Author Contributions
Conceptualization, S.C.; Methodology, S.C., B.O., F.M. .; Investigation, S.C., F.M., L.A. S.S., B.K., D.M., B.O.; Resources, S.C., F.M., L.A., S.S., B.K., D.M., B.O.; Data Curation, B.O., F.M., D.M.; Writing – Original Draft Preparation, F.M., L.A., S.S. , B.K.; Writing – Review & Editing, S.C. B.K. S.S. D.M. & B.O.; Visualization, S.C. F.M.; Supervision, B.O. D.M. S.C.
Funding
No external funding was obtained from any source to support the execution of this review.
Conflicts of Interest
The authors declare that they have no conflicts of interest to disclose.
References
- Bowley, G.; Kugler, E.; Wilkinson, R.; Lawrie, A.; van Eeden, F.; Chico, T.J.A.; Evans, P.C.; Noël, E.S.; Serbanovic-Canic, J. Zebrafish as a tractable model of human cardiovascular disease. Br. J. Pharmacol. 2021, 179, 900–917. [Google Scholar] [CrossRef] [PubMed]
- Bashirzade, A.A.; Zabegalov, K.N.; Volgin, A.D.; Belova, A.S.; Demin, K.A.; de Abreu, M.S.; Babchenko, V.Y.; Bashirzade, K.A.; Yenkoyan, K.B.; Tikhonova, M.A.; et al. Modeling neurodegenerative disorders in zebrafish. Neurosci. Biobehav. Rev. 2022, 138, 104679. [Google Scholar] [CrossRef] [PubMed]
- Daya, A.; Donaka, R.; Karasik, D. Zebrafish models of sarcopenia. Dis Model Mech [Internet] 2020, 13, dmm042689. [Google Scholar] [CrossRef]
- Ichii, S.; Matsuoka, I.; Okazaki, F.; Shimada, Y. Zebrafish Models for Skeletal Muscle Senescence: Lessons from Cell Cultures and Rodent Models. Molecules 2022, 27, 8625. [Google Scholar] [CrossRef] [PubMed]
- Adhish, M.; Manjubala, I. Effectiveness of zebrafish models in understanding human diseases—A review of models. Heliyon 2023, 9, e14557. [Google Scholar] [CrossRef]
- Kijima, Y.; Wantong, W.; Igarashi, Y.; Yoshitake, K.; Asakawa, S.; Suzuki, Y.; Watabe, S.; Kinoshita, S. Age-Associated Different Transcriptome Profiling in Zebrafish and Rats: an Insight into the Diversity of Vertebrate Aging. Mar. Biotechnol. 2022, 24, 1–16. [Google Scholar] [CrossRef]
- Johnston, I.A.; Bower, N.I.; Macqueen, D.J. Growth and the regulation of myotomal muscle mass in teleost fish. J. Exp. Biol. 2011, 214, 1617–1628. [Google Scholar] [CrossRef]
- Bhandari, S.; Kim, Y.-I.; Nam, I.-K.; Hong, K.; Jo, Y.; Yoo, K.-W.; Liao, W.; Lim, J.-Y.; Kim, S.-J.; Um, J.-Y.; et al. Loss of pex5 sensitizes zebrafish to fasting due to deregulated mitochondria, mTOR, and autophagy. Cell. Mol. Life Sci. 2023, 80, 1–20. [Google Scholar] [CrossRef]
- Guyon, J.R.; Steffen, L.S.; Howell, M.H.; Pusack, T.J.; Lawrence, C.; Kunkel, L.M. Modeling human muscle disease in zebrafish. Biochim Biophys Acta Mol Basis Dis [Internet] 2007, 1772, 205–15. [Google Scholar] [CrossRef]
- Tesoriero, C.; Greco, F.; Cannone, E.; Ghirotto, F.; Facchinello, N.; Schiavone, M.; Vettori, A. Modeling Human Muscular Dystrophies in Zebrafish: Mutant Lines, Transgenic Fluorescent Biosensors, and Phenotyping Assays. Int. J. Mol. Sci. 2023, 24, 8314. [Google Scholar] [CrossRef]
- Teame, T.; Zhang, Z.; Ran, C.; Zhang, H.; Yang, Y.; Ding, Q.; Xie, M.; Gao, C.; Ye, Y.; Duan, M.; et al. The use of zebrafish (Danio rerio) as biomedical models. Anim. Front. 2019, 9, 68–77. [Google Scholar] [CrossRef] [PubMed]
- Christian, C.J.; Benian, G.M. Animal models of sarcopenia. Aging Cell [Internet] 2020, 19. [Google Scholar] [CrossRef] [PubMed]
- Ryu, B.; Je, J.-G.; Jeon, Y.-J.; Yang, H.-W. Zebrafish Model for Studying Dexamethasone-Induced Muscle Atrophy and Preventive Effect of Maca (Lepidium meyenii). Cells 2021, 10, 2879. [Google Scholar] [CrossRef] [PubMed]
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. Hallmarks of aging: An expanding universe. Cell 2023, 186, 243–278. [Google Scholar] [CrossRef]
- Schmeer, C.; Kretz, A.; Wengerodt, D.; Stojiljkovic, M.; Witte, O. W. Dissecting aging and senescence—current concepts and open lessons. Cells 2019, 8, 1446. [Google Scholar] [CrossRef]
- Hill, M.; Třískala, Z.; Honců, P.; Krejčí, M.; Kajzar, J.; Bičíková, M.; et al. Aging, hormones and receptors. Physiol Res 2020, 69 (Suppl 2), S255–72. [Google Scholar] [CrossRef]
- Ames, B.N. Prolonging healthy aging: Longevity vitamins and proteins. Proc. Natl. Acad. Sci. 2018, 115, 10836–10844. [Google Scholar] [CrossRef]
- Barbagallo, M.; Veronese, N.; Dominguez, L.J. Magnesium in aging, health and diseases. Nutrients 2021, 13, 463. [Google Scholar] [CrossRef]
- Remelli, F.; Vitali, A.; Zurlo, A.; Volpato, S. Vitamin D Deficiency and Sarcopenia in Older Persons. Nutrients 2019, 11, 2861. [Google Scholar] [CrossRef]
- Wang, Z.; Zhu, W.; Xing, Y.; Jia, J.; Tang, Y. B vitamins and prevention of cognitive decline and incident dementia: a systematic review and meta-analysis. Nutr Rev 2022, 80, 931–49. [Google Scholar] [CrossRef]
- Sun, R.; Wang, J.; Feng, J.; Cao, B. Zinc in Cognitive Impairment and Aging. Biomolecules 2022, 12, 1000. [Google Scholar] [CrossRef]
- Reuter, H.; Perner, B.; Wahl, F.; Rohde, L.; Koch, P.; Groth, M.; Buder, K.; Englert, C. Aging Activates the Immune System and Alters the Regenerative Capacity in the Zebrafish Heart. Cells 2022, 11, 345. [Google Scholar] [CrossRef] [PubMed]
- Monma, Y.; Shimada, Y.; Nakayama, H.; Zang, L.; Nishimura, N.; Tanaka, T. Aging-associated microstructural deterioration of vertebra in zebrafish. Bone Rep. 2019, 11, 100215. [Google Scholar] [CrossRef] [PubMed]
- Zambusi, A.; Burhan. P.; Di Giaimo, R.; Schmid, B.; Ninkovic, J. Granulins Regulate Aging Kinetics in the Adult Zebrafish Telencephalon. Cells 2020, 9, 350. [Google Scholar] [CrossRef] [PubMed]
- Nishikawa, H.; Fukunishi, S.; Asai, A.; Yokohama, K.; Nishiguchi, S.; Higuchi, K. Pathophysiology and mechanisms of primary sarcopenia (Review). Int. J. Mol. Med. 2021, 48, 1–8. [Google Scholar] [CrossRef]
- Cruz-Jentoft, AJ.; Bahat, G.; Bauer, J.; Boirie, Y.; Bruyère, O.; Cederholm, T.; et al. Sarcopenia: revised European consensus on definition and diagnosis. Age Ageing 2019, 48, 16–31. [Google Scholar] [CrossRef]
- Bauer, J.; Morley, J.E.; Schols, A.M.W.J.; Ferrucci, L.; Cruz-Jentoft, A.J.; Dent, E.; et al. Sarcopenia: A time for action. An SCWD position paper. J Cachexia Sarcopenia Muscle 2019, 10, 956–61. [Google Scholar] [CrossRef]
- Supriya, R.; Singh, K.P.; Gao, Y.; Gu, Y.; Baker, J.S. Effect of Exercise on Secondary Sarcopenia: A Comprehensive Literature Review. Biology 2021, 11, 51. [Google Scholar] [CrossRef]
- Sayer, A.A.; Cruz-Jentoft, A. Sarcopenia definition, diagnosis and treatment: consensus is growing. Age Ageing 2022, 51. [Google Scholar] [CrossRef]
- Tournadre, A.; Vial, G.; Capel, F.; Soubrier, M.; Boirie, Y. Sarcopenia. Joint Bone Spine 2019, 86, 309–14. [Google Scholar] [CrossRef]
- Sartori, R.; Romanello, V.; Sandri, M. Mechanisms of muscle atrophy and hypertrophy: implications in health and disease. Nat. Commun. 2021, 12, 1–12. [Google Scholar] [CrossRef]
- Larsson, L.; Degens, H.; Li, M.; Salviati, L.; Lee, Y.I.; Thompson, W.; Kirkland, J.L.; Sandri, M. Sarcopenia: Aging-Related Loss of Muscle Mass and Function. Physiol. Rev. 2019, 99, 427–511. [Google Scholar] [CrossRef] [PubMed]
- Petermann-Rocha, F.; Balntzi, V.; Gray, S.R.; Lara, J.; Ho, F.K.; Pell, J.P.; et al. Global prevalence of sarcopenia and severe sarcopenia: a systematic review and meta-analysis. J Cachexia Sarcopenia Muscle 2022, 13, 86–99. [Google Scholar] [CrossRef] [PubMed]
- Colleluori, G.; Villareal, D.T. Aging, obesity, sarcopenia and the effect of diet and exercise intervention. Exp. Gerontol. 2021, 155, 111561–111561. [Google Scholar] [CrossRef] [PubMed]
- Goates, S.; Du, K.; Arensberg, M.B.; Gaillard, T.; Guralnik, J.; Pereira, S.L. ECONOMIC IMPACT OF HOSPITALIZATIONS IN US ADULTS WITH SARCOPENIA. J. Frailty Aging 2019, 8, 1–7. [Google Scholar] [CrossRef]
- Migliavacca, E.; Tay, S.K.H.; Patel, H.P.; Sonntag, T.; Civiletto, G.; McFarlane, C.; Forrester, T.; Barton, S.J.; Leow, M.K.; Antoun, E.; et al. Mitochondrial oxidative capacity and NAD+ biosynthesis are reduced in human sarcopenia across ethnicities. Nat. Commun. 2019, 10, 1–14. [Google Scholar] [CrossRef]
- He, Y.; Xie, W.; Li, H.; Jin, H.; Zhang, Y.; Li, Y. Cellular Senescence in Sarcopenia: Possible Mechanisms and Therapeutic Potential. Front. Cell Dev. Biol. 2022, 9, 793088. [Google Scholar] [CrossRef]
- Ticinesi, A.; Nouvenne, A.; Cerundolo, N.; Catania, P.; Prati, B.; Tana, C.; Meschi, T. Gut Microbiota, Muscle Mass and Function in Aging: A Focus on Physical Frailty and Sarcopenia. Nutrients 2019, 11, 1633. [Google Scholar] [CrossRef]
- Dao, T.; Green, A.E.; A Kim, Y.; Bae, S.-J.; Ha, K.-T.; Gariani, K.; Lee, M.-R.; Menzies, K.J.; Ryu, D. Sarcopenia and Muscle Aging: A Brief Overview. Endocrinol. Metab. 2020, 35, 716–732. [Google Scholar] [CrossRef]
- Abdelrahman, Z.; Wang, X.; Wang, D.; Zhang, T.; Zhang, Y.; Wang, X.; Chen, Z. Identification of novel pathways and immune profiles related to sarcopenia. Front. Med. 2023, 10. [Google Scholar] [CrossRef]
- Ayobahan, S.U.; Eilebrecht, S.; Baumann, L.; Teigeler, M.; Hollert, H.; Kalkhof, S.; Eilebrecht, E.; Schäfers, C. Detection of biomarkers to differentiate endocrine disruption from hepatotoxicity in zebrafish (Danio rerio) using proteomics. Chemosphere 2019, 240, 124970. [Google Scholar] [CrossRef] [PubMed]
- Juvale, I.I.A.; Che Has, A.T. The potential role of miRNAs as predictive biomarkers in neurodevelopmental disorders. J Mol Neurosci 2021, 71, 1338–55, Available from: https://pubmed.ncbi.nlm.nih.gov/33774758/. [Google Scholar] [CrossRef]
- Wawruszak, A.; Okoń, E.; Dudziak, K. A Review of the Role of the Zebrafish (Danio reiro) in Preclinical and Clinical Models of Biomarker Identification and Drug Targeting, and Developments in Personalized Medicine in Breast Cancer. Experiment 2023, 29, e940550–1. [Google Scholar] [CrossRef] [PubMed]
- Liguori, I.; Russo, G.; Curcio, F.; Bulli, G.; Aran, L.; Della-Morte, D.; et al. Oxidative stress, aging, and diseases. Clin Interv Aging 2018, 13, 757–72, Available from: https://pubmedncbinlmnihgov/29731617/. [Google Scholar] [CrossRef]
- Kadoguchi, T.; Shimada, K.; Miyazaki, T.; Kitamura, K.; Kunimoto, M.; Aikawa, T.; Sugita, Y.; Ouchi, S.; Shiozawa, T.; Yokoyama-Nishitani, M.; et al. Promotion of oxidative stress is associated with mitochondrial dysfunction and muscle atrophy in aging mice. Geriatr. Gerontol. Int. 2019, 20, 78–84. [Google Scholar] [CrossRef]
- Daussin, F.N.; Boulanger, E.; Lancel, S. From mitochondria to sarcopenia: Role of inflammaging and RAGE-ligand axis implication. Exp. Gerontol. 2021, 146, 111247. [Google Scholar] [CrossRef] [PubMed]
- Hong, X.; Campanario, S.; Ramírez-Pardo, I.; Grima-Terrén, M.; Isern, J.; Muñoz-Cánoves, P. Stem cell aging in the skeletal muscle: The importance of communication. Ageing Res. Rev. 2021, 73, 101528. [Google Scholar] [CrossRef]
- Mankhong, S.; Kim, S.; Moon, S.; Kwak, H.-B.; Park, D.-H.; Kang, J.-H. Experimental Models of Sarcopenia: Bridging Molecular Mechanism and Therapeutic Strategy. Cells 2020, 9, 1385. [Google Scholar] [CrossRef]
- Pacifici, F.; Della-Morte, D.; Piermarini, F.; Arriga, R.; Scioli, M.G.; Capuani, B.; Pastore, D.; Coppola, A.; Rea, S.; Donadel, G.; et al. Prdx6 Plays a Main Role in the Crosstalk between Aging and Metabolic Sarcopenia. Antioxidants 2020, 9, 329. [Google Scholar] [CrossRef]
- Zhang, X.; Habiballa, L.; Aversa, Z.; Ng, Y.E.; Sakamoto, A.E.; Englund, D.A.; Pearsall, V.M.; White, T.A.; Robinson, M.M.; Rivas, D.A.; et al. Characterization of cellular senescence in aging skeletal muscle. Nat. Aging 2022, 2, 601–615. [Google Scholar] [CrossRef]
- Ebert, S.M.; Dierdorff, J.M.; Meyerholz, D.K.; Bullard, S.A.; Al-Zougbi, A.; DeLau, A.D.; Tomcheck, K.C.; Skopec, Z.P.; Marcotte, G.R.; Bodine, S.C.; et al. An investigation of p53 in skeletal muscle aging. J. Appl. Physiol. 2019, 127, 1075–1084. [Google Scholar] [CrossRef] [PubMed]
- Cai, Z.; Liu, D.; Yang, Y.; Xie, W.; He, M.; Yu, D.; Wu, Y.; Wang, X.; Xiao, W.; Li, Y. The role and therapeutic potential of stem cells in skeletal muscle in sarcopenia. Stem Cell Res. Ther. 2022, 13, 1–12. [Google Scholar] [CrossRef]
- Berberoglu, M.A.; Gallagher, T.L.; Morrow, Z.T.; Talbot, J.C.; Hromowyk, K.J.; Tenente, I.M.; Langenau, D.M.; Amacher, S.L. Satellite-like cells contribute to pax7-dependent skeletal muscle repair in adult zebrafish. Dev. Biol. 2017, 424, 162–180. [Google Scholar] [CrossRef]
- Ferreira, F.J.; Carvalho, L.; Logarinho, E.; Bessa, J. foxm1 Modulates Cell Non-Autonomous Response in Zebrafish Skeletal Muscle Homeostasis. Cells 2021, 10, 1241. [Google Scholar] [CrossRef] [PubMed]
- Carneiro, M.C.; de Castro, I.P.; Ferreira, M.G. Telomeres in aging and disease: lessons from zebrafish. Dis. Model. Mech. 2016, 9, 737–748. [Google Scholar] [CrossRef] [PubMed]
- Kishi, S.; Bayliss, P.E.; Uchiyama, J.; Koshimizu, E.; Qi, J.; Nanjappa, P.; Imamura, S.; Islam, A.; Neuberg, D.; Amsterdam, A.; et al. The Identification of Zebrafish Mutants Showing Alterations in Senescence-Associated Biomarkers. PLOS Genet. 2008, 4, e1000152–e1000152. [Google Scholar] [CrossRef]
- Henderson, T.D.; Choi, J.; Leonard, S.W.; Head, B.; Tanguay, R.L.; Barton, C.L.; Traber, M.G. Chronic Vitamin E Deficiency Dysregulates Purine, Phospholipid, and Amino Acid Metabolism in Aging Zebrafish Skeletal Muscle. Antioxidants 2023, 12, 1160. [Google Scholar] [CrossRef]
- Zou, Y.-Y.; Chen, Z.-L.; Sun, C.-C.; Yang, D.; Zhou, Z.-Q.; Xiao, Q.; Peng, X.-Y.; Tang, C.-F. A High-Fat Diet Induces Muscle Mitochondrial Dysfunction and Impairs Swimming Capacity in Zebrafish: A New Model of Sarcopenic Obesity. Nutrients 2022, 14, 1975. [Google Scholar] [CrossRef]
- Breen, L.; Phillips, S.M. Skeletal muscle protein metabolism in the elderly: Interventions to counteract the 'anabolic resistance' of ageing. Nutr. Metab. 2011, 8, 68–68. [Google Scholar] [CrossRef]
- Bilski, J.; Pierzchalski, P.; Szczepanik, M.; Bonior, J.; Zoladz, J.A. Multifactorial Mechanism of Sarcopenia and Sarcopenic Obesity. Role of Physical Exercise, Microbiota and Myokines. Cells 2022, 11, 160. [Google Scholar] [CrossRef]
- E Yarasheski, K.; Bhasin, S.; Sinha-Hikim, I.; Pak-Loduca, J.; Gonzalez-Cadavid, N.F. Serum myostatin-immunoreactive protein is increased in 60-92 year old women and men with muscle wasting. Journal of nutrition health and aging 2002, 6, 343–348. [Google Scholar]
- Gao, Y.; Dai, Z.; Shi, C.; Zhai, G.; Jin, X.; He, J.; et al. Depletion of myostatin b promotes somatic growth and lipid metabolism in zebrafish. Front Endocrinol (Lausanne) 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Evans, W.J.; Hellerstein, M.; Orwoll, E.; Cummings, S.; Cawthon, P.M. D3-Creatine dilution and the importance of accuracy in the assessment of skeletal muscle mass. J. Cachex- Sarcopenia Muscle 2019, 10, 14–21. [Google Scholar] [CrossRef]
- Lu, L.; Mao, L.; Feng, Y.; Ainsworth, B.E.; Liu, Y.; Chen, N. Effects of different exercise training modes on muscle strength and physical performance in older people with sarcopenia: a systematic review and meta-analysis. BMC Geriatr. 2021, 21, 1–30. [Google Scholar] [CrossRef]
- Uchitomi, R.; Oyabu, M.; Kamei, Y. Vitamin D and Sarcopenia: Potential of Vitamin D Supplementation in Sarcopenia Prevention and Treatment. Nutrients 2020, 12, 3189. [Google Scholar] [CrossRef] [PubMed]
- Goodpaster, B.H.; Park, S.W.; Harris, T.B.; Kritchevsky, S.B.; Nevitt, M.; Schwartz, A.V.; et al. The loss of skeletal muscle strength, mass, and quality in older adults: The health, aging and body composition study. J Gerontol A Biol Sci Med Sci 2006, 61, 1059–64. [Google Scholar] [CrossRef] [PubMed]
- Janssen, I. Evolution of sarcopenia research. Appl Physiol Nutr Metab 2010, 35, 707–12. [Google Scholar] [CrossRef]
- Papadopoulou, S.K.; Voulgaridou, G.; Kondyli, F.S.; Drakaki, M.; Sianidou, K.; Andrianopoulou, R.; Rodopaios, N.; Pritsa, A. Nutritional and Nutrition-Related Biomarkers as Prognostic Factors of Sarcopenia, and Their Role in Disease Progression. Diseases 2022, 10, 42. [Google Scholar] [CrossRef]
- Lin, S.; Ling, M.; Chen, C.; Cai, X.; Yang, F.; Fan, Y. Screening Potential Diagnostic Biomarkers for Age-Related Sarcopenia in the Elderly Population by WGCNA and LASSO. BioMed Res. Int. 2022, 2022, 1–14. [Google Scholar] [CrossRef]
- Picca, A.; Calvani, R.; Cesari, M.; Landi, F.; Bernabei, R.; Coelho-Júnior, H.J.; et al. Biomarkers of physical frailty and sarcopenia: Coming up to the place? Int J Mol Sci 2020, 21, 5635. [Google Scholar] [CrossRef]
- Feng, S.; Wang, S.; Wang, Y.; Yang, Q.; Wang, D.; Li, H. Identification and expression of carbonic anhydrase 2, myosin regulatory light chain 2 and selenium-binding protein 1 in zebrafish Danio rerio: Implication for age-related biomarkers. Gene Expr. Patterns 2018, 29, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Dent, E.; Morley, J.E.; Cruz-Jentoft, A.J.; Arai, H.; Kritchevsky, S.B.; Guralnik, J.; Bauer, J.M.; Pahor, M.; Clark, B.C.; Cesari, M.; et al. International Clinical Practice Guidelines for Sarcopenia (ICFSR): Screening, Diagnosis and Management. J. Nutr. Health Aging 2018, 22, 1148–1161. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.-K.; Woo, J.; Assantachai, P.; Auyeung, T.-W.; Chou, M.-Y.; Iijima, K.; Jang, H.C.; Kang, L.; Kim, M.; Kim, S.; et al. Asian Working Group for Sarcopenia: 2019 Consensus Update on Sarcopenia Diagnosis and Treatment. J. Am. Med. Dir. Assoc. 2020, 21, 300–307. [Google Scholar] [CrossRef] [PubMed]
- Barton, C.; King, M.; Dascombe, B.; Taylor, N.; Silva, D.d.O.; Holden, S.; Goff, A.; Takarangi, K.; Shields, N. Many physiotherapists lack preparedness to prescribe physical activity and exercise to people with musculoskeletal pain: A multi-national survey. Phys. Ther. Sport 2021, 49, 98–105. [Google Scholar] [CrossRef]
- Hurst, C.; Robinson, S.M.; Witham, M.D.; Dodds, R.M.; Granic, A.; Buckland, C.; De Biase, S.; Finnegan, S.; Rochester, L.; A Skelton, D.; et al. Resistance exercise as a treatment for sarcopenia: prescription and delivery. Age Ageing 2022, 51. [Google Scholar] [CrossRef]
- Tsekoura, M.; Billis, E.; Kastrinis, A.; Katsoulaki, M.; Fousekis, K.; Tsepis, E.; et al. The effects of exercise in patients with sarcopenia. In GeNeDis 2020; Springer International Publishing: Cham, 2021; pp. 281–90. [Google Scholar]
- Hasumura, T.; Meguro, S. Exercise quantity-dependent muscle hypertrophy in adult zebrafish (Danio rerio). J. Comp. Physiol. B 2016, 186, 603–614. [Google Scholar] [CrossRef]
- Rutkove, S.B.; Callegari, S.; Concepcion, H.; Mourey, T.; Widrick, J.; Nagy, J.A.; Nath, A.K. Electrical impedance myography detects age-related skeletal muscle atrophy in adult zebrafish. Sci. Rep. 2023, 13, 1–14. [Google Scholar] [CrossRef]
- Wu, X.; Yu, X.; Zhu, N.; Xu, M.; Li, Y. Beneficial effects of whey protein peptides on muscle loss in aging mice models. Front. Nutr. 2022, 9, 897821. [Google Scholar] [CrossRef]
- Kerasioti, E.; Stagos, D.; Priftis, A.; Aivazidis, S.; Tsatsakis, A.M.; Hayes, A.W.; Kouretas, D. Antioxidant effects of whey protein on muscle C2C12 cells. Food Chem. 2014, 155, 271–278. [Google Scholar] [CrossRef]
- Drummond, M.J.; Glynn, E.L.; Fry, C.S.; Dhanani, S.; Volpi, E.; Rasmussen, B.B. Essential amino acids increase MicroRNA-499, −208b, and −23a and downregulate myostatin and myocyte enhancer factor 2C mRNA expression in human skeletal muscle. J Nutr 2009, 139, 2279–84. [Google Scholar] [CrossRef]
- Barbiera, A.; Pelosi, L.; Sica, G.; Scicchitano, B.M. Nutrition and microRNAs: Novel Insights to Fight Sarcopenia. Antioxidants 2020, 9, 951. [Google Scholar] [CrossRef]
- Papadopoulou, S.K.; Papadimitriou, K.; Voulgaridou, G.; Georgaki, E.; Tsotidou, E.; Zantidou, O.; et al. Exercise and nutrition impact on osteoporosis and sarcopenia—the incidence of osteosarcopenia: A narrative review. Nutrients 2021, 13, 4499. [Google Scholar] [CrossRef]
- Bellanti, F.; Lo Buglio, A.; Vendemiale, G. Muscle delivery of mitochondria-targeted drugs for the treatment of sarcopenia: Rationale and perspectives. Pharmaceutics 2022, 14, 2588. [Google Scholar] [CrossRef]
- Hardee, J.P.; Lynch, G.S. Current pharmacotherapies for sarcopenia. Expert Opin Pharmacother 2019, 20, 1645–57. [Google Scholar] [CrossRef]
- Najjar, F.; Rizk, F.; Carnac, G.; Nassar, R.; Jabak, S.; Sobolev, AP.; et al. Protective effect ofRhus coriariafruit extracts against hydrogen peroxide-induced oxidative stress in muscle progenitors and zebrafish embryos. PeerJ 2017, 5, e4144. [Google Scholar] [CrossRef] [PubMed]
- Turkel, I.; Ozerklig, B.; Yılmaz, M.; Ulger, O.; Kubat, G.B.; Tuncer, M. Mitochondrial transplantation as a possible therapeutic option for sarcopenia. J Mol Med 2023, 101, 645–69. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Pan, M.; Zhou, M.; Tang, Q.; Chen, M.; Hong, W.; et al. Mitochondria transplantation from stem cell for mitigating sarcopenia. Aging Dis 2023. Available from: https://pubmed.ncbi.nlm.nih.gov/37196123. [Google Scholar] [CrossRef] [PubMed]
- Clark, B.C.; Rutkove, S.; Lupton, E.C.; Padilla, C.J.; Arnold, W.D. Potential utility of electrical impedance myography in evaluating age-related skeletal muscle function deficits. Front Physiol 2021, 12. [Google Scholar] [CrossRef]
- Cebrián-Ponce, Á.; Irurtia, A.; Carrasco-Marginet, M.; Saco-Ledo, G.; Girabent-Farrés, M.; Castizo-Olier, J. Electrical impedance myography in health and physical exercise: A systematic review and future perspectives. Front Physiol 2021, 12. [Google Scholar] [CrossRef]
- Rutkove, S.B.; Chen, Z.-Z.; Pandeya, S.; Callegari, S.; Mourey, T.; Nagy, J.A.; et al. Surface electrical impedance myography detects skeletal muscle atrophy in aged wildtype zebrafish and aged gpr27 knockout zebrafish. Biomedicines 2023, 11, 1938, Available from: https://pubmed.ncbi.nlm.nih.gov/37509577/. [Google Scholar] [CrossRef]
- Sun, C.-C.; Yang, D.; Chen, Z.-L.; Xiao, J.-L.; Xiao, Q.; Li, C.-L.; et al. Exercise intervention mitigates zebrafish age-related sarcopenia via alleviating mitochondrial dysfunction. FEBS J 2023, 290, 1519–30. [Google Scholar] [CrossRef]
- Rogeri, P.S.; Zanella, R. , Jr, Martins, G.L.; Garcia, M.D.A.; Leite, G.; Lugaresi, R.; et al. Strategies to prevent sarcopenia in the aging process: Role of protein intake and exercise. Nutrients 2021, 14, 52. [Google Scholar] [CrossRef] [PubMed]
- Van houcke, J.; De Groef, L.; Dekeyster, E.; Moons, L. The zebrafish as a gerontology model in nervous system aging, disease, and repair. Ageing Res Rev 2015, 24 Pt B, 358–68, Available from: https://pubmed.ncbi.nlm.nih.gov/26538520/. [Google Scholar] [CrossRef]
- Shah, R.R.; Nerurkar, N.L.; Wang, C.C.; Galloway, J.L. Tensile properties of craniofacial tendons in the mature and aged zebrafish. J Orthop Res 2015, 33, 867–73. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.-L.; Guo, C.; Zou, Y.-Y.; Feng, C.; Yang, D.-X.; Sun, C.-C.; et al. Aerobic exercise enhances mitochondrial homeostasis to counteract D-galactose-induced sarcopenia in zebrafish. Exp Gerontol 2023, 180, 112265. [Google Scholar] [CrossRef]
- Willems, S.M.; Wright, D.J.; Day, F.R.; Trajanoska, K.; Joshi, P.K.; Morris, J.A.; et al. Large-scale GWAS identifies multiple loci for hand grip strength providing biological insights into muscular fitness. Nat Commun 2017, 8. [Google Scholar] [CrossRef]
- Kwon, R.Y.; Watson, C.J.; Karasik, D. Using zebrafish to study skeletal genomics. Bone [Internet] 2019, 126, 37–50. [Google Scholar] [CrossRef]
- Xiao, S.-M.; Kung, A.W.C.; Gao, Y.; Lau, K.-S.; Ma, A.; Zhang, Z.-L.; et al. Post-genome wide association studies and functional analyses identify association of MPP7 gene variants with site-specific bone mineral density. Hum Mol Genet 2012, 21, 1648–57. [Google Scholar] [CrossRef]
- Polačik, M.; Blažek, R.; Reichard, M. Laboratory breeding of the short-lived annual killifish Nothobranchius furzeri. Nat Protoc 2016, 11, 1396–413. [Google Scholar] [CrossRef]
- Kim, Y.; Nam, HG.; Valenzano, DR. The short-lived African turquoise killifish: an emerging experimental model for ageing. Dis Model Mech 2016, 9, 115–29. [Google Scholar] [CrossRef]
- Hu, C.-K.; Brunet, A. The African turquoise killifish: A research organism to study vertebrate aging and diapause. Aging Cell 2018, 17. [Google Scholar] [CrossRef] [PubMed]
- Królak, M. The epizootiological situation with regard to animal brucellosis in Poland 1985. Przegl Epidemiol 1987, 41. [Google Scholar]
- Cencioni, C.; Heid, J.; Krepelova, A.; Rasa, S.M.M.; Kuenne, C.; Guenther, S.; et al. Aging triggers H3K27 trimethylation hoarding in the chromatin of Nothobranchius furzeri skeletal muscle. Cells 2019, 8, 1169. [Google Scholar] [CrossRef] [PubMed]
- Giaquinto, D.; De Felice, E.; Attanasio, C.; Palladino, A.; Schiano, V.; Mollo, E.; et al. Central and peripheral NPY age-related regulation: A comparative analysis in fish translational models. Int J Mol Sci 2022, 23, 3839. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Summary of the numerous advantages in Zebrafish as a model for unraveling the mechanistic pathways and potential therapies as a model for primary Sarcopenia.
Figure 1.
Summary of the numerous advantages in Zebrafish as a model for unraveling the mechanistic pathways and potential therapies as a model for primary Sarcopenia.
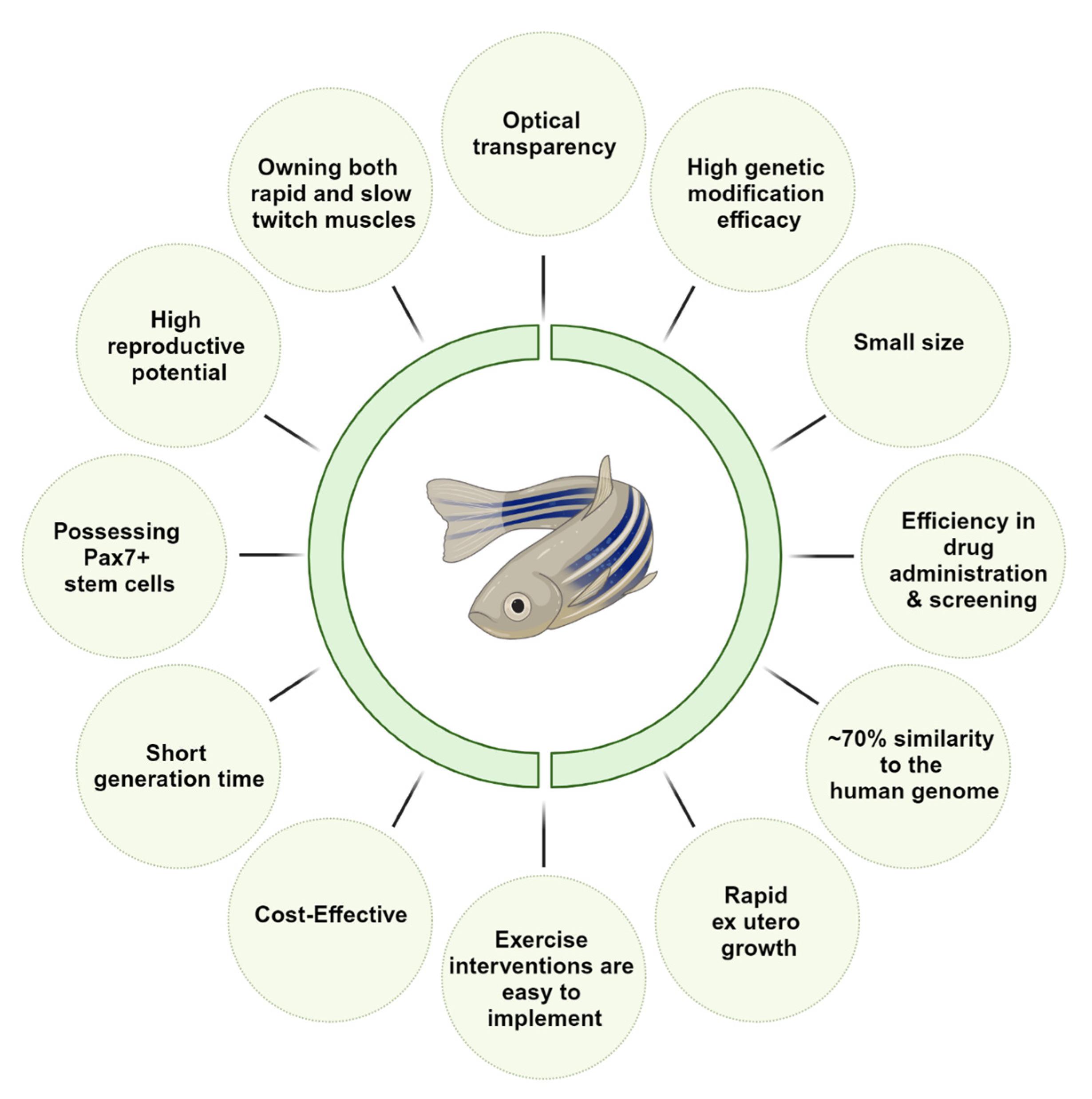
Figure 2.
Deterioration of muscle aging and function through the lifespan of an individual. Zebrafish as a Sarcopenia model would not only be beneficial to those with the disease but also to understand the mechanisms leading to muscle degradation in all individuals through lifespan.
Figure 2.
Deterioration of muscle aging and function through the lifespan of an individual. Zebrafish as a Sarcopenia model would not only be beneficial to those with the disease but also to understand the mechanisms leading to muscle degradation in all individuals through lifespan.
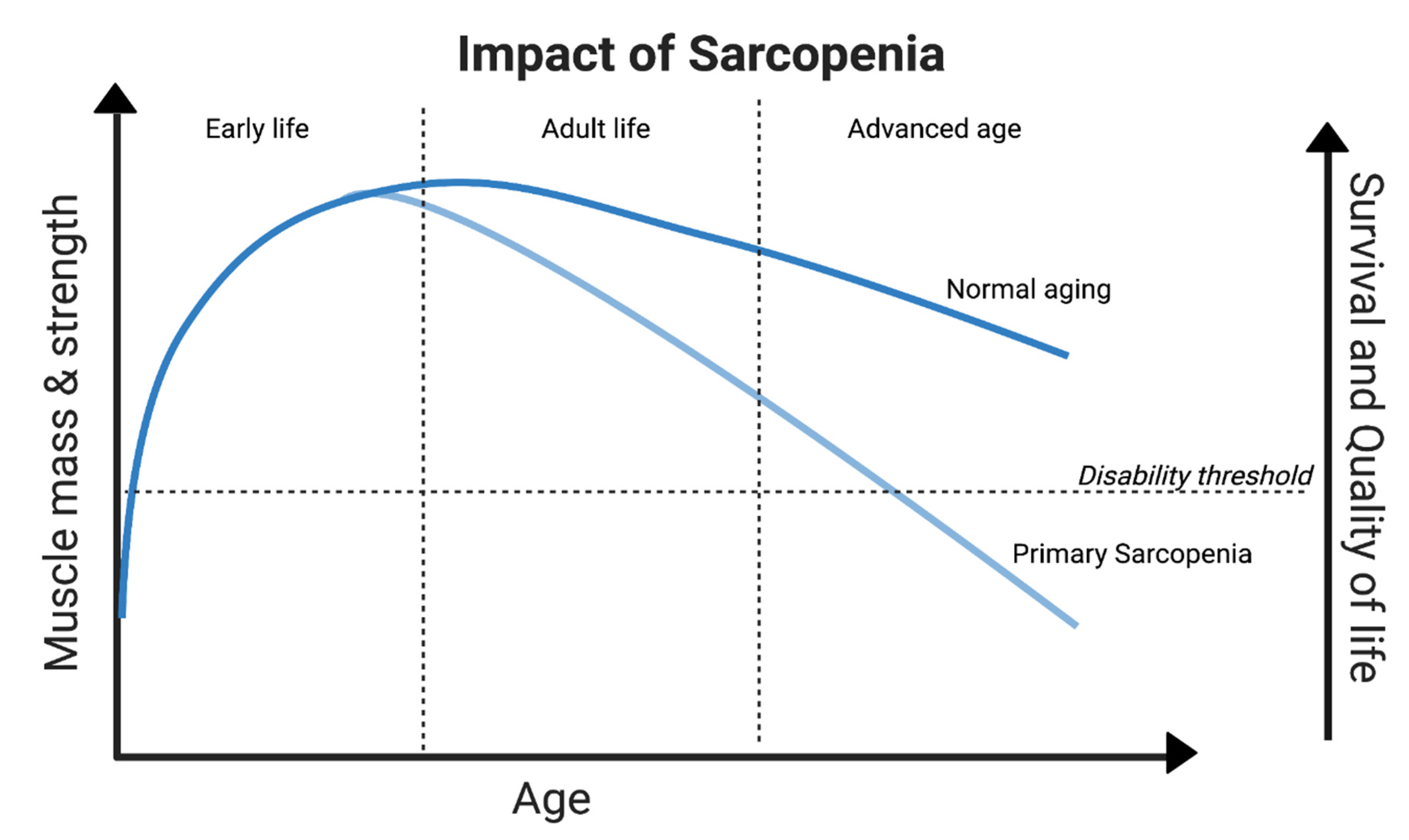
Figure 3.
Multiple pathways lead to the deterioration of organ systems and ultimately lead to worsening of muscle mass and function impacting the functionality of the individual, leading to primary sarcopenia.
Figure 3.
Multiple pathways lead to the deterioration of organ systems and ultimately lead to worsening of muscle mass and function impacting the functionality of the individual, leading to primary sarcopenia.
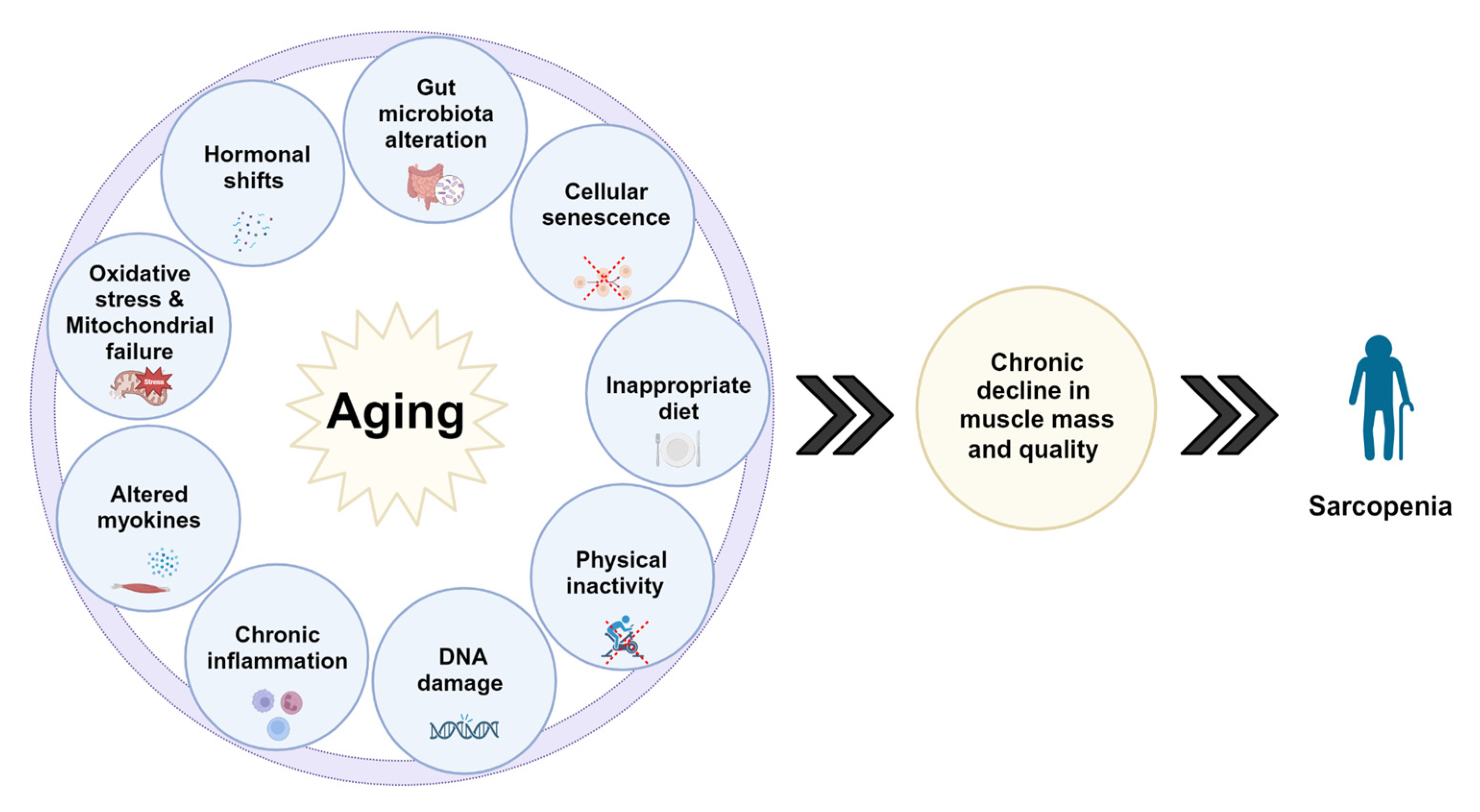

Table 1.
Latest Experimental Studies in Zebrafish As Sarcopenia Model.
| Title | Authors | Zebrafish animals used | Intervention and/or exposure | Results |
|---|---|---|---|---|
| Electrical impedance myography detects age-related skeletal muscle atrophy in adult zebrafish | Rutkove SB, Callegari S, Concepcion H, Mourey T, Widrick J, Nagy JA, Nath AK. | Wildtype casper (6 & 33 months) and Tübingen (4 & 24 months) zebrafish | Relationship between swimming efficacy, age, and EIM measures | EIM effectively correlates with age-related muscle atrophy in adult zebrafish. |
| Exercise intervention mitigates zebrafish age-related sarcopenia via alleviating mitochondrial dysfunction | Sun CC, Yang D, Chen ZL, Xiao JL, Xiao Q, Li CL, Zhou ZQ, Peng XY, Tang CF, Zheng L | AB strain male zebrafish (21 &6 months) | swimming efficacy on mitochondrial homeostasis and protein regulation in sarcopenia | Exercise reduced age-related muscle atrophy by improving muscle structure, decreasing protein breakdown, and restoring mitochondrial activity. |
| A High-Fat Diet Induces Muscle Mitochondrial Dysfunction and Impairs Swimming Capacity in Zebrafish: A New Model of Sarcopenic Obesity | Zou YY, Chen ZL, Sun CC, Yang D, Zhou ZQ, Xiao Q, Peng XY, Tang CF. | Adult male AB strain zebrafish (4 months) | High-fat diet (16 weeks) induced sarcopenic obesity models and its effect on swimming capacity and muscle atrophy | High-fat diet for an extended period of time resulted in muscular atrophy, reduced swimming ability, increased body weight, higher muscle triglycerides, fatty liver characteristics, and downregulation of mitochondrial and fatty acid metabolism genes. |
| Surface Electrical Impedance Myography Detects Skeletal Muscle Atrophy in Aged Wildtype Zebrafish and Aged gpr27 Knockout Zebrafish | Rutkove SB, Chen ZZ, Pandeya S, Callegari S, Mourey T, Nagy JA, Nath AK. | Tübingen-strain wild type (8,12 & 36 months) and gpr27 knockout zebrafish | Muscle atrophy detection in young, aged and mutant zebrafish by surface EIM(sEIM) | sEIM effectively recognized muscle structural changes. |
| Aerobic exercise enhances mitochondrial homeostasis to counteract D-galactose-induced sarcopenia in zebrafish | Chen ZL, Guo C, Zou YY, Feng C, Yang DX, Sun CC, Wen W, Jian ZJ, Zhao Z, Xiao Q, Zheng L, Peng XY, Zhou ZQ, Tang CF. | Wild-type male AB strain zebrafish (7 months) | Correlation between D-galactose induced sarcopenia models and aerobic activity | Aerobic exercise enhances muscle function and quality by regulating miR-128/IGF-1 pathway and improves mitochondrial homeostasis in aging muscle. |
| Chronic Vitamin E Deficiency Dysregulates Purine, Phospholipid, and Amino Acid Metabolism in Aging Zebrafish Skeletal Muscle | Henderson TD, Choi J, Leonard SW, Head B, Tanguay RL, Barton CL, Traber MG. | Zebrafish (55 dpf) supplemented for 12 or 18 months | Vitamin E inadequate and adequate diet for 12 or 18 months | The metabolic pathway alterations in skeletal muscle observed in aging and vitamin E deprivation exhibit some similarities but also demonstrate distinct modifications. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



