
Submitted:
25 September 2023
Posted:
26 September 2023
You are already at the latest version
Abstract
Objective: There are currently no known treatment methods for ensuring complete recovery from facial nerve damage. Thus, various basic and clinical studies continue to be conducted to achieve this end. As part of this research, autophagy is being investigated for its role in nerve regeneration. Here, we review the existing literature regarding the involvement and signifi-cance of autophagy in facial nerve injury and regeneration. Methods: A comprehensive litera-ture review was conducted to assess the induction and role of autophagy in facial nerve injury and subsequent regeneration. Results: Studies conducted on rats and mice have demonstrated activation of autophagy and expression of related factors in facial and sciatic nerves. Among the most studied of these factors in relation to degeneration and regeneration of facial and sciatic nerves are LC3II/I, PI3K, mTOR, Beclin-1, ATG3, ATG5, ATG7, ATG9, and ATG12. Conclusion: This analysis indicates that autophagy is involved in the process of nerve regeneration follow-ing facial and sciatic nerve damage. Inadequate autophagy induction or failure of autophagy re-sponses can result in regeneration issues after facial or sciatic nerve damage. Animal studies suggest that autophagy plays an important role in facial and sciatic nerve degeneration and re-generation.
Keywords:
facial nerve
; degeneration
; regeneration
; autophagy
1. Introduction
1-1. Facial nerve and facial palsy
The facial nerve is a mixed nerve containing both motor and sensory functions. Motor nerves are responsible for movement of the muscles of the face and neck, and contain parasympathetic components that are responsible for the secretory actions of lacrimal and salivary glands. It also contains specialized sensory nerves in the anterior two-thirds of the tongue that detect taste and general sensory nerves that are responsible for deep perception of the auricle, posterior wall of the ear canal, ear lobe, and soft tissue of the face. Thus, the facial nerve consists of two efferent nerves and two afferent nerves and collectively serves four functions. The facial nerve can be classified into upper and lower segments based on its motor nucleus. The upper segment of the motor nucleus receives inputs from both sides of the brain – cross fibers and non-cross fibers from the pons – governing facial expressions of both sides of the face. In contrast, nerve fibers in the lower segment of the motor nucleus are exclusively innervated by the contralateral cerebral cortex. Thus, in cases where upper lesions occur on one side of the motor nucleus, because the upper face is controlled by both hemispheres, paralysis mainly manifests in the facial expression muscles at the bottom of the face around the lower region of the mouth. This paralysis does not affect the expression muscles of the forehead or eye circumference or impair taste, salivation, or tear secretion. In contrast, in the case of a lesion below the unilateral motor nucleus, paralysis affects both upper and lower facial muscles on the lesion side as a whole [1,2].
Although facial nerve palsy is not a life-threatening disease, it has a devastating effect on those afflicted, impacting their emotional state and social lives. Thus, effective treatment and positive prognosis are of paramount importance. When damage to the cell body of a neuron is sufficient to destroy it, the neuron can no longer survive. However, if the axon is only partially cut and the cell body is not damaged, the neuron can regenerate the axon. Moreover, under appropriate conditions, this cell may re-form synapses to cells with which it had previously synapsed, allowing for the possibility of full restoration of function. Changes in nerve fibers after nerve damage vary depending on the degree of damage. In the case of mild damage such as neuropraxia, nerve fibers go through the process of local demyelination and remyelination; but in case of the severe damage, axonal degeneration and regeneration occur [3,4].
1-2. Definition and types of autophagy
The term ‘autophagy’ is derived from the Greek words ‘auto’ meaning self and ‘phagy’ meaning to eat. There are three types of autophagy: (1) microautophagy, in which intracellular substances directly enter the lysosome through invagination of the lysosomal membrane; (2) macroautophagy, where an autophagosome composed of a double lipid membrane surrounds a substance and fuses with lysosomes; and (3) chaperone-mediated autophagy, which degrades proteins with specific target motifs through a process mediated by a chaperone complex and lysosomal-associated membrane protein type 2A. Where not otherwise specified, ‘autophagy’ generally refers to macroautophagy. Autophagy is further classified into aggrephagy, lipophagy, mitophagy, plexophagy, ribophagy and xenophagy, depending on the nature of the material that is loaded and digested. Autophagy plays an important role not only in the degradation of protein aggregates, but also in the removal of damaged intracellular organelles, including mitochondria, endoplasmic reticulum and peroxisomes; it also is involved in removing extracellular pathogens, such as bacteria, viruses, and parasites. In various organisms, including mice, whole-body or tissue-specific deletion of autophagy-related genes (Atg) causes serious disorders and death [5,6], supporting the hypothesis that autophagy is an important process in maintaining health.
1-3. Molecular mechanisms controlling autophagy
Since the first identification of autophagy-related genes in yeast by Ohsumi et al. in 1993, more than 35 Atg genes have been discovered [7,8]. In addition to being intricately regulated by the proteins it produces, the autophagy process is under negative regulation by mTOR (mammalian target of rapamycin) and positive regulation by AMPK (adenosine monophosphate-activated protein kinase). The process itself can be divided into four major steps: 1) initiation and vesicle nucleation, 2) vesicle elongation, 3) fusion and degradation, and 4) termination. The initiation step is regulated by the ULK1 protein, with the ULK1-containing complex, ULK1-Atg13-Atg101-FIP200, dissociating from the mTORC1 complex as a result of nutrient deficiency-dependent dephosphorylation of ULK1. This leads to increased activation of the beclin 1/Vps34 complex (beclin 1-Atg14L-Vps15-Vps34 complex), inducing the production of PI3P (phosphatidylinositol-3-phosphate) and initiating nucleation of vesicles with a double-membrane structure, facilitated by the DFCP1 and WIPI proteins that gather at the site. Vesicles subsequently elongate through the actions of proteins that constitute the two ubiquitin-like conjugation systems, ATG12-ATG5-ATG16 and LC3-phosphatidylethanolamine, resulting in the formation of an autophagosome. The resulting mature autophagosome then fuses with a lysosome to form an autolysosome, within which isolated intracellular substances or organelles are degraded by lysosomal hydrolytic enzymes. As a consequence, nutrients (e.g., amino acids) released through autolysosomal digestion increase the activation of mTOR protein, a major negative regulator of the mTORC1 complex, ultimately leading to termination of the autophagy process [9,10].
2. Induction of autophagy in various diseases
The autophagy process, involving the transport of intracellular substances surrounded by a double membrane structure to a lysosome and subsequent autolysomal degradation, acts together with the ubiquitin proteasome system to play an important role in maintaining intracellular protein quality control. Autophagy also plays an important role in the degradation of protein aggregates and the removal of damaged intracellular organelles and extracellular pathogens. Thus, not surprisingly, dysregulation of autophagy can act as a major cause of various diseases. In the absence of sufficient nutrients to generate adequate ATP, autophagy also breaks down proteins into amino acids, lipid droplets into fatty acids, and glycogen into glucose, thereby serving a regulatory function that maintains intracellular energy balance. Conversely, under energy-replete conditions, autophagic degradation of energy sources (proteins, fat particles, and sugar sources) is suppressed and their storage increases. However, disruptions in the regulation of autophagy result in an energy imbalance [11,12].
Ongoing studies have investigated potential positive or negative effects of autophagy on human diseases, but the underlying mechanisms have proven more complex than expected. Consequently, whether and how autophagy is involved in maintaining human health or the development of disease remains unclear. Activation of autophagy in cancer acts as a tumor suppressor, whereas inactivation of autophagy allows survivals of cancer cells under nutrient-poor conditions. In muscular disorders, autophagy may increase as a compensatory response to defects in lysosome function, whereas inactivation of autophagy can lead to the accumulation of autophagosomes, potentially impairing cell function. In neurodegeneration, autophagy activation contributes to the timely removal of protein aggregates before they become toxic, whereas autophagy inactivation might trigger cell death in neurons burdened with aggregated proteins. In the case of pathogen infection, autophagy activation serves as a cellular defense against invasion of bacteria and viruses; conversely, inactivation of autophagy allows pathogens to establish a replicative niche [11,12,13,14].
While the involvement of autophagy has been studied in various diseases, its induction and contribution to nerve degeneration and regeneration after facial nerve injury (FNI) has not yet been established. To bridge this knowledge gap and shed light on the clearance of damaged nerve debris and innate immune induced by facial nerve damage, we conducted a review of the relevant literature. To this end, we analyzed and summarized the results of previous studies on the involvement of autophagy in nerve regeneration. Our search encompassed literature databases, focusing on studies published in English. Studies were included if they 1) were prospective or retrospective investigations of autophagy and facial or peripheral nerves; 2) considered facial nerve degeneration and regeneration; and 3) included human patients and/or animal studies on autophagy and facial or peripheral nerves. From 1993 to 2023, we identified 188 studies based on search terms from PubMed electronic database. No autophagy, no facial or peripheral nerve, review article, off topic, not written in English were excluded. A total of 14 peripheral nerve studies that met these criteria, including 11 involving sciatic nerves, 2 involving facial nerves, and 1 involving the inferior alveolar nerve, were included in this review (Figure 1).
3. Expression of autophagy-related factors, with or without autophagy inducers, after peripheral nerve injury
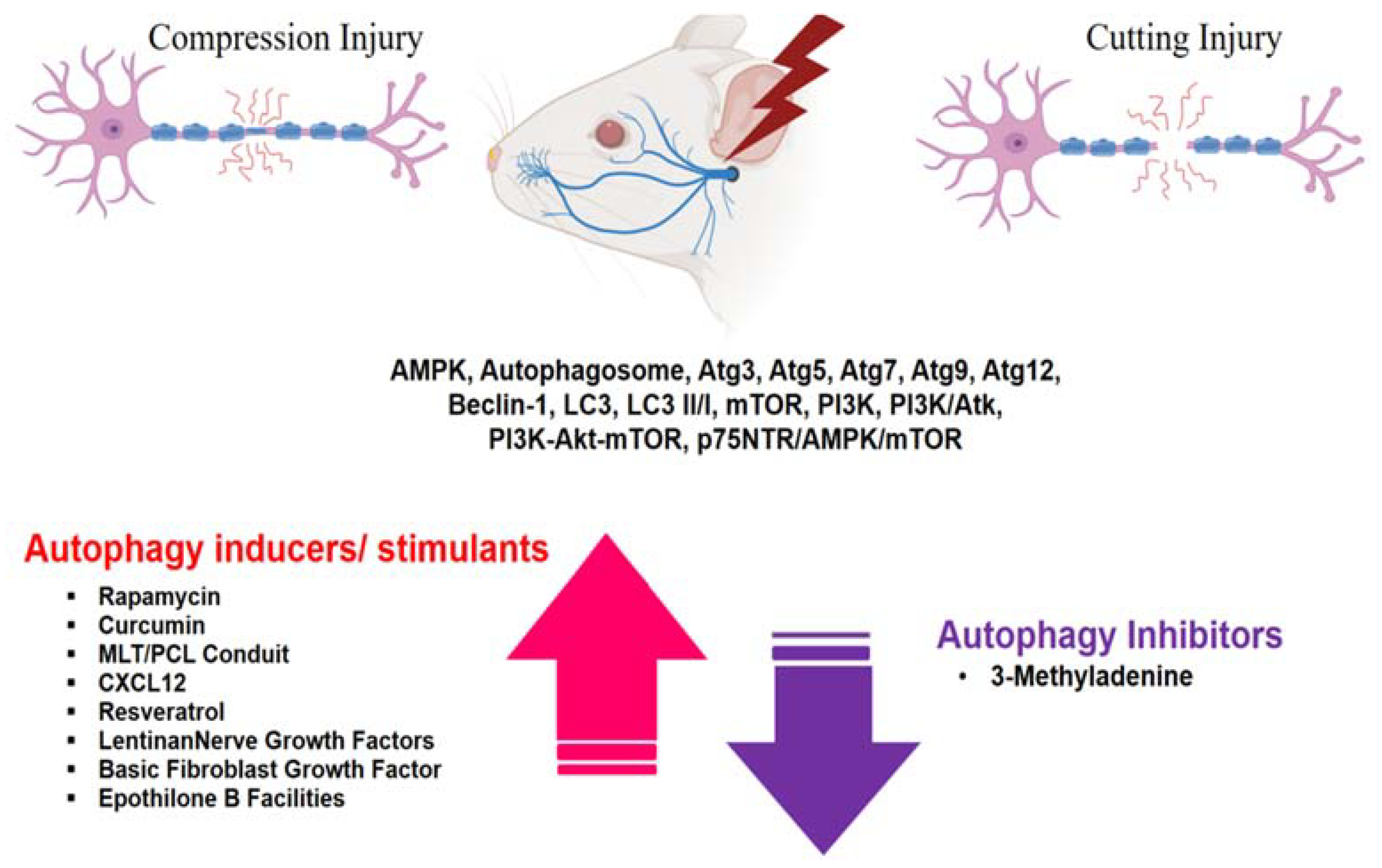
In explorations of the potential role of autophagy in the context of peripheral nerve injury, the expression of autophagy-related factors and associated pathways looms large. This section delves into dynamic changes in these factors and processes in the context of peripheral nerve injury, with or without stimulation of autophagy-inducing factors, particularly as it relates to nerve degeneration/regeneration and recovery (Figure 2).
3-1. Upregulation of autophagy-related factors without stimulation of autophagy-inducing factors after sciatic nerve injury (Table 1)
One previous study on nerve degeneration and regeneration following sciatic nerve injury in genetically engineered mouse models investigated modulation of autophagy-related factors after injury [15], reporting distinct upregulation of these factors post-nerve injury. In this study, using Atg7fl/fl, Mpz-Cre, Mpz-Cre, and Mpz-Cre+;Atg7loxP/loxP mice, the sciatic nerve was exposed and transected, and a temporal analysis of autophagy-related genes and proteins was conducted. During demyelination, several autophagy-related genes, especially those for the ULK complex, ATG9 cycling system and ATG7, which are essential for autophagosome formation, were strongly induced. Several ATG proteins, including ATG7, ATG16L complex, Wipi2 and beclin-1 (also known as ATG6), were also increased in injured nerves compared with uninured nerves, concomitant with a decrease in the degeneration of myelin protein zero (MPZ) and myelin basic protein (MBP). Furthermore, a substantial increase in the level of LC3 II, indicative of autophagosome initiation, was observed in response to injury in vivo. An examination of nerve homogenates from uninjured and injured Atg7-cKO mice revealed a significant reduction in ATG7, ATG5-12 and LC3 II in intact and severed nerves, suggesting impaired autophagy. These findings underscore the important role of inductive autophagy in Wallerian degeneration.
Given its role in cellular self-cleaning and repair mechanisms, ATG5 (autophagy related gene 5), a key player in the autophagy process, holds particular significance in the context of nerve regeneration. In one study investigating whether autophagy induction is important for motor nerve regeneration [16], researchers subjected 12-week-old Sprague-Dawley rats to a nerve crush injury (three times for 30 seconds with fine forceps) or cutting injury of the right sciatic nerve. After surgery, ATG5 (AAVrh10-ATG5) or control protein, in this case green fluorescence protein (GFP; AAVrh10-GFP), was injected into spinal motor neurons and nerve conduction tests were performed. At 48 and 60 days after nerve injury, rats in the AAVrh10-ATG5 group showed higher nerve conduction amplitudes in gastrocnemius and tibialis anterior muscles than those in the AAVrh10-GFP group (p < 0.05). Motor performance of hindlimb movements was also better in the AAVrh10-ATG5 group than in the AAVrh10-GFP group. Collectively, these findings indicate that overexpression of ATG5 increases motor axon regeneration, establishing a connection between ATG5 expression and the process of motor neuron regeneration in the context of neuronal damage.

Table 1.
Expression of autophagy-related factors in sciatic nerves without stimulation of autophagy-inducing factors after sciatic nerve injury.
Table 1.
Expression of autophagy-related factors in sciatic nerves without stimulation of autophagy-inducing factors after sciatic nerve injury.
| Author/Year/ Reference | Study design | Species and/or sample | Nerve/Injury methods | Detection Method | Target substance(s) associated with autophagy | Results : Conclusions |
| Gomez-Sanchez JA, et al. 2015 [15] | Animal study |
Atg7flfl mice, Mpz-Cre mice, Mpz-Cre+, Atg7loxP/loxP mice |
Sciatic nerve transection |
Immunohistochemistry Electron microscopy qRT-PCR Western blotting Lipid analysis Proteomic analysis Behavioral tests |
ATG5, ATG7, ATG9, ATG12, LC3-II |
Several autophagy-related genes were expressed during demyelination; in particular, the ULK complex, ATG9 cycling system and ATG7, which are essential for autophagosome formation, were strongly induced. ATG7, ATG5, ATG12 and LC3 II were significantly reduced in homogenates of uninjured and injured nerves from Atg7-cKO mice, suggesting impaired autophagy. : These results reveal an important role for inductive autophagy during Wallerian degeneration, and point to potential mechanistic targets for accelerating myelin clearance and improving demyelinating disease. |
| Romeo-Guitart D, et al. 2019 [16] | Animal study | Sprague-Dawley rats | Sciatic nerve/crush and transection injury | Immunohistochemistry, Western blotting Cell culture Electrophysiological and functional assessments |
Atg5 |
Nerve conduction amplitudes were higher in gastrocnemius and tibialis anterior muscles in the AAVrh10-ATG5 group than in the AAVrh10-GFP group at 48 days and 60 days post injury. : ATG5 overexpression in spinal motoneurons stimulates mTOR-independent autophagy and facilitates a growth-competent state after nerve transection, improving motor axonal regeneration and electromyographic responses. |
AAV, adeno-associated virus; ATG, autophagy-related genes; GFP, green fluorescent protein; LC 3-II, microtubule-associated protein-1 light chain 3-II protein; mTOR, mammalian target of the rapamycin; qRT-PCR; quantitative reverse transcription-polymerase chain reaction.
3-2. Activation of autophagy and expression of related factors in facial and sciatic nerves with stimulation of autophagy-inducing factors after peripheral nerve injury (Table 2)
3-2-1. Autophagy responses to rapamycin and 3-methyladenine
The influence of rapamycin, an allosteric inhibitor of mTOR – and thus an activator of autophagy – and 3-methyladenine (3-MA), an autophagy inhibitor, on the response to nerve injury was tested in adult female Sprague–Dawley rats. In these studies [17], the sciatic nerve was clamped with forceps three times for 10 seconds each at 10-second intervals; for sham controls, the nerve was exposed without performing a crush injury. Five days after surgery, rats were intraperitoneally injected with rapamycin (1 mg/kg; crush + Rapa group), 3-MA (50 mg/kg; crush + 3-MA group), or vehicle (DMSO) control (crush + Veh group). A fourth group received sham surgery and vehicle injection (sham + Veh group). Motor function was assessed by evaluating standing time and footprint intensity of the operated limbs at 1, 2, 3, and 6 weeks after injury. Mean standing time in the rapamycin-treated group was 2-fold longer than that in the crush + Veh group at 1 and 2 weeks after injury (p < 0.05), whereas mean footprint strength was similar between the two groups at 2 weeks. These results indicate that animals treated with rapamycin are better able to support body weight on the injured limb from 2 weeks after injury, probably as a result of enhanced autophagy in the acute phase. In contrast, average standing time in the 3-MA group was significantly lower compared with sham-operated animals at 1 and 2 weeks post-injury (45% and 15%, respectively), and mean footprint strength was approximately 20–30% lower (p < 0.05). These results provide evidence supporting the role of autophagy in nerve regeneration and motor function recovery in the peripheral nervous system, and may serve as a basis for the development of therapies that improve the outcome of peripheral nerve injury through pharmacological induction of autophagy.
In a related set of experiments [18], sciatic nerves of 8-week-old male Sprague-Dawley rats (n = 42) were dissected instead of clamped and autophagy was studied. In this study, rats were randomly divided into four groups: sham neurorrhaphy (control group), transected + DMSO group (sciatic nerve transected, repaired, and then injected with DMSO), Rapa group (sciatic nerve transected, repaired, and then injected with rapamycin), and a 3-MA group (sciatic nerve transected, repaired, and then injected with 3-MA). The sciatic nerve function index (SFI), which ranges from -10 (normal) to -100 (severe damage), was used as a recovery metric. There was no change in the sham-treated control group, whereas rats injected with rapamycin to induce autophagy showed improved hind-limb function (SFI: -44.56 ± 5.86) compared with rats injected with DMSO (-59.74 ± 5.11; p < 0.05). In contrast, functional recovery in rats injected with 3-MA was significantly inhibited (SFI: -72.15 ± 15.84; p < 0.05). Immunostaining for S-100 and LC3 increased in the Rapa group; merged staining also increased in the Rapa group (115.5 ± 11.56) compared with DMSO (28.5 ± 4.203 pixels/cm2; p < 0.001), but decreased in the 3-MA group (10 ± 4.546 pixels per cm2; p < 0.05). In addition, neurofilament immunostaining and merged staining with LC3 tended to increase in the Rapa group, suggesting improved neurofilament continuity and nerve function. Collectively, these findings reinforce the importance of autophagy in peripheral nerve regeneration and suggest its role in scar reduction.
Another related study [19] focused on the inferior alveolar nerve (IAN) instead of the sciatic nerve. In this study, the IAN of Sprague-Dawley rats was transected, and rapamycin, 3-MA or vehicle was administered to the IAN transection (IANX) site once a day for 10 days after surgery. All rats that underwent surgery failed to show a motor response in the lower lip for 4 days after IANX. The elevated head-withdrawal threshold (HWT) gradually decreased from day 6 after IANX; in contrast, the head-withdrawal threshold (HWT) of IANX rats was significantly higher than that of control rats throughout the experimental period. In sections proximal and distal to the severed IAN, rapamycin administration to the injured area led to a significant reduction in the expression level of phosphorylated p70S6K (p-p70s6k), a downstream target of mTOR. In contrast, beclin-1 and microtubule-associated protein-1 light chain 3-II (MAP1LC3) protein levels in the proximal and distal stumps of the severed IAN were increased in the Rapa group compared with the control group. Thus, recovery from IANX-induced sensory disturbance of the lower lip was markedly improved by rapamycin. These results suggested that rapamycin administration is a promising treatment for IANX-induced sensory impairment.
Table 2.
Expression of autophagy-related factors in facial and sciatic nerves with stimulation of autophagy-inducing/inhibiting factors after peripheral nerve injury.
Table 2.
Expression of autophagy-related factors in facial and sciatic nerves with stimulation of autophagy-inducing/inhibiting factors after peripheral nerve injury.
| Author/Year/ Reference | Study design | Species and/or sample | Nerve/Injury methods | Autophagy stimulants/ inhibitors Detection Method |
Target substance(s) associated with autophagy | Results : Conclusions |
| Huang H, et al. 2016 [17] | Animal study | Sprague-Dawley rats | Sciatic nerve crush injury | Rapamycin and 3-MA Electron microscopy Western blotting Immunocytochemistry CatWalk gait analysis |
Autophagosome LC3-II |
The mean standing time in the Rapa group was twofold higher than in the crush + Veh group at 1 and 2 weeks post injury (p < 0.05). In contrast, the mean standing time was significantly lower in the 3-MA group at 1 and 2 weeks post-injury (45% and 15%, respectively), whereas the mean footprint intensity was ~20–30% lower at 1, 2, and 3 weeks post-injury compared with sham-operated animals (p < 0.05). Expression of the autophagy marker LC3-II was higher in the Rapa group and lower in the 3-MA group (p < 0.05) compared with controls. : Modulation of autophagy in PNI could be an effective pharmacological approach for promoting nerve regeneration and reestablishing motor function. |
| Ko PY, et al. 2017 [18] | Animal study | Sprague-Dawley rats | Sciatic nerve transection | Rapamycin or 3-MA Behavioral test Immunohistochemistry |
LC3 | Induction of autophagy by injection (i.p.) of rats with rapamycin significantly improved hind-limb function compared with injection of DMSO (-44.56 ± 5.86 vs. -59.74 ± 5.11; p < 0.05). Neurofilament, LC3, and merged staining trended higher in the Rapa group. : A deficiency of Schwann cell autophagic activity might be an early event in nerve scar formation, and modulating autophagy might be a powerful pharmacological approach for improving functional outcomes. |
| Zhao Z, et al. 2017 [20] | Animal study | Sprague-Dawley rats | Sciatic nerve compression (mechanical clamping) | Curcumin Western blotting qRT-PCR Flow cytometry |
LC3-II Beclin-1 |
Curcumin treatment increased the levels of LC3-II and beclin-1 in the negative control group (p < 0.05). : Curcumin accelerated repair of injured sciatic nerves in rats by reducing Schwann cell apoptosis and promoting myelinization. |
| Yun Qian et al. 2018 [23] | Animal study | Sprague-Dawley rats | Sciatic nerve/Dissection |
Melatonin and polymeric nerve conduits Walking tract analysis Morphological evaluation Immunofluorescence evaluation Western blotting |
Beclin-1 LC3A/B, Atg3, Atg5, Atg7 |
LC3A/B expression was significantly higher in MLT/PCL and autograft groups than the PCL group (p < 0.05). Beclin-1 and LC3-I expression were 2.6- and 4.1-fold higher, respectively, in the MLT/PCL group than in the PCL group (p < 0.05). ATG3, ATG5 and ATG7 expression were 1.5-, 1.8- and 2.2- fold higher in the MLT/PCL group than the PCL group, but were significantly lower in both groups compared with the autograft group (p < 0.05). : 3D melatonin nerve scaffolds increase autophagy in peripheral nerve regeneration. |
| Gao D, et al. 2019 [25] | In vivo | Sprague-Dawley rats | Facial nerve compression injury (mechanical clamping) | CXCL12 Cell proliferation assay TUNEL staining ELISA Immunofluorescence Western blotting TEM |
LC3II/I PI3K-Akt-mTOR |
CXCL12 induced a time-dependent increase in LC3B II and reduction in P62 – two markers of autophagy – in Schwann cells. Treatment with CXCL12 decreased PI3K, AKT and mTOR phosphorylation, but increased expression of the autophagy marker LC3II/I. : CXCL12 promotes the migration of Schwann cells and is potentially a key molecule in the repair of FNI. |
| Zhang J, et al. 2020 [26] |
Animal study | Sprague-Dawley rats | Sciatic nerve crush injury | Resveratrol SFI TEM Immunohistochemistry Western blot analysis |
Resveratrol LC3-II/IBeclin1 |
SFI values on days 7 and 14 post injury were significantly higher in the crush+Res group than the control group (p < 0.01). Levels of LC3-II/I and beclin-1 were highest in the crush+Res group and lowest in the sham group. : Resveratrol facilitated peripheral nerve repair by improving autophagy in Schwann cells, at least partially accelerating the Wallerian degeneration process after SNCI. |
| Li Rui, et al. 2020 [31] | Animal study | Wistar rats | Sciatic nerve crush injury | NGF Electron microscopy Immunofluorescence Western blotting qRT-PCR Myelin phagocytosis assays Detection of autophagic flow |
LC3 p75NTR/AMPK/mTOR axis |
NGF treatment resulted in an increase in the fluorescence intensity of the autophagic marker protein LC3 in the PNI+NGF group compared with the PNI group. Compared with the sham group, the p-AMPK/AMPK ratio increased whereas both p-mTOR/mTOR and p-p70s6k/p70s6k ratios decreased after PNI. These effects were further enhanced by NGF treatment in the PNI+NGF group. : The effect of NGF on promoting early nerve regeneration is closely associated with its acceleration of autophagic clearance of myelin debris in Schwann cells. |
| Inada T, et al. 2021 [18] | Animal study | Sprague-Dawley rats | Inferior alveolar nerve transection | Rapamycin, 3-MA Western blotting Immunohistochemistry Mechanical sensitivity assay |
Beclin-1, LC3 |
The head-withdrawal threshold (HWT) in IANX rats was significantly higher than that in sham-operated rats throughout the experimental period. Rapamycin increased both LC3-II and beclin-1 expression in the injured site of the inferior alveolar nerve on day 6 after nerve transection. : Rapamycin-induced facilitation of recovery from the sensory disturbance following IANX was mediated through the clearance of myelin debris by Schwann cells. |
| Xiao H, et al. 2022 [32] | Animal study | Sprague-Dawley rats | Sciatic nerve compression injury |
Lentinan Gait analysis Network pharmacology Histological analysis Immunofluorescence Cell culture Western blotting |
AMPK/mTOR LC3-II/LC3-I ratio |
Lentinan promoted autophagic flux in vivo via the AMPK/mTOR signaling pathway, accelerated the clearance of myelin debris by SCs, and inhibited neuronal apoptosis, thereby promoting neurological recovery. Lentinan treatment increased the LC3-II/LC3-I ratio. : Lentinan promotes nerve regeneration primarily by accelerating the autophagic clearance of myelin debris in Schwann cells, a process likely regulated by the AMPK/mTOR signaling pathway. |
| Zhang Z, et al. 2022 [33] | Animal study | Sprague-Dawley rats | Sciatic nerve crush injury | ADSCs+Mel SFI Western blotting Immunofluorescence |
LC3-II/LC3-I Beclin-1 |
The number of autophagosomes and lysosomes and the expression of LC3-II/LC3-I and beclin-1 proteins were prominently increased in the ADSCs + Mel group compared with the Model group, and significantly decreased in the 3-MA group. : ADSCs combined with melatonin promotes sciatic nerve regeneration in rats by altering the early autophagic activity of the injured sciatic nerve. |
| Hu B, et al. 2022 [34] | Sprague-Dawley rats | Facial nerve compression injury (mechanical clamping) | bFGF Fluorescence labeling and imaging Western blotting Facial nerve functional scoring |
LC3B, Beclin-1 ATG5 |
Abnormal nerve morphologies were improved in the bFGF group and P-bFGF group. Nerve fibers in the P-bFGF group showed significantly greater and more regular regeneration compared with those in the bFGF group. Immunofluorescence staining showed that the fluorescence intensity of LC3B was slightly increased by FNI and was further enhanced by bFGF or P-bFGF treatment, with P-bFGF treatment producing significantly greater fluorescence intensity than bFGF. Western blotting showed that changes in the expression of LC3B-II, beclin-1, and ATG5 paralleled changes in the fluorescence intensity of LC3B. : P-bFGF effectively promotes cell proliferation, myelination and functional recovery, and also reduces apoptosis of nerve cells after FNI. |
|
| Zhou J, et al. 2020 [35] | Animal study | Sprague-Dawley rats | Sciatic nerve crush injury | EpoB Walking track Assessment Western blotting Von Frey filament test TEM |
PI3K/Akt | SFI values for EpoB-treatment groups gradually decreased over 4 weeks, indicating that EpoB treatment facilitated the recovery of sensory function after nerve crush injury. EpoB treatment significantly increased LC3B II levels and inhibited PI3K and Akt phosphorylation in Schwann cells. : EpoB is capable of promoting axonal regeneration and remyelination as well as functional recovery in a rat model of nerve crush injury. |
AAV, adeno-associated virus; ADSC, adipose-derived stem cells; AMPK, activated protein kinase; Akt, protein kinase B; ATG, autophagy-related genes; bFGF, basic fibroblast growth factor; ELISA, enzyme-linked immunoassay; EpoB, epothilone B; IANX, inferior alveolar nerve transection; IGF, insulin-like growth factor; LC 3-II, microtubule-associated protein-1 light chain 3-II protein; Mel, melatonin; 3-MA, 3-methyladenine; mTOR, mammalian target of the rapamycin; Mel, melatonin; MLT/PCL, melatonin and polymeric nerve conduits; MBP, myelin basic protein; MPZ, myelin protein zero; NGF, nerve growth factor; PNI, peripheral nerve injury; p75NTR, 75 kD neurotrophin receptor; PI3K; phosphatidylinositol 3-kinase; qRT-PCR; quantitative reverse transcription-polymerase chain reaction; RAPA, rapamycin; SNCI, sciatic nerve compression injury; SFI, sciatic nerve function index; TEM, transmission electron microscopy; ULK1, Unc51-like autophagy activating kinase 1.
3-2-2. Curcumin
Given its intricate interactions with diverse signaling pathways and potential to modulate an array of cellular processes, curcumin – a natural product with broad health benefits and low toxicity – is an attractive candidate for investigating in the context of nerve injury. In one such investigation [20] employing a rat model of sciatic nerve injury, the sciatic nerve of male Sprague-Dawley rats was clamped with a 2-mm-wide needle holder, after which nerve-damaged rats were randomly divided into four groups (n = 10 rats/group): negative control (NC), curcumin, PD98059 (MEK inhibitor) + curcumin, and insulin-like growth factor 1 (IGF 1) + curcumin. In groups treated with curcumin, rats were intraperitoneally administered curcumin (100 mg/kg/d, dissolved in DMSO at 20 mg/ml) for 60 days. Following surgery, LC3-II and beclin-1 levels significantly increased, whereas the levels of p62 significantly decreased, in all curcumin-containing groups compared with the NC group, suggesting initiation of autophagy in the damaged sciatic nerve. The greatest increase in LC3-II and beclin 1 was detected in the group given curcumin alone, suggesting that injury-induced autophagy in the sciatic nerve is enhanced by curcumin treatment. Curcumin administration also significantly increased the levels of phosphorylated-ERK1/2 (p-ERK1/2) and decreased levels of p-Akt, indicating that curcumin promotes ERK1/2 pathway activation and inhibits Akt/mTOR pathway activation in the injured sciatic nerve. Taken together, these observations indicate that curcumin promotes injury-induced cell autophagy in the rat sciatic nerve by targeting Akt/mTOR and ERK1/2 pathways.
3-2-3. Three-dimensional melatonin nerve scaffolds
Damage to peripheral nerves gives rise to various disorders in patients. In the case of nerve transection, autogenous nerve transplantation is a good standard treatment for severe peripheral nerve defects. However, a number of factors, including post-traumatic inflammation, oxidative stress and misdirected nerve regeneration, can inhibit functional recovery of peripheral nerves. One widely used and effective method for bolstering peripheral nerve regeneration is to position a nerve conduit between two transected nerves to support nerve regrowth. Although this method is not sufficient for long nerve defects, it is effective for short defects, with traditional polymeric nerve conduits such as polyglycolic acid and polycaprolactone (PCL) seeing wide use in clinical practice [21,22,23]. In one preclinical study [24], 30 male Sprague-Dawley rats (weight, 200-250 g) were randomly assigned to three groups: Mel (melatonin)/PCL, PCL, and autologous transplant groups. After the sciatic nerve segment was cut, the two nerve endings were ligated using a 15-mm nerve guide conduit. SFI, used as a metric to assess recovery, was significantly higher in the Mel/PCL group than the PCL group at both 6 and 12 weeks postoperatively (p < 0.05). At 6 weeks, SFI remained lower in Mel/PCL than in autograft group (p < 0.05), showing good results for nerve regeneration and nerve recovery. The function and role of autophagy in the regenerated sciatic nerve was confirmed by assessing LC3A/B expression by immunohistochemical analysis. Beclin-1 and LC3-I expression were 2.6- and 4.1-fold higher, respectively, in the Mel/PCL group than in the PCL group (p < 0.05). ATG3, ATG5 and ATG7 expression were also higher in the Mel/PCL group than the PCL group, although this increase was more modest (1.5-, 1.8- and 2.2-fold, respectively). These results show that the Mel/PCL conduit can clear nerve debris and trigger autophagy, fostering nerve regeneration.
3-2-4. CXCL12
CXCL12 has garnered interest as a potential therapeutic target for enhancing nerve regeneration, and emerging evidence suggests that it might play a role in regulating autophagy. In one study investigating this connection [25], male Sprague-Dawley rats were separated into surgery and sham groups. Rats in the surgery group were subjected to nerve-crush injury of the main trunk of the right extracranial facial nerve (compressed once using mosquito forceps for 50 seconds at a point 0.5 cm from the stylomastoid foramen), with and without treatment with recombinant rat CXCL12, injected near the posterior mastoid of the rat ear at a dose of 4 μg/kg/d. The CXCL12 group was further subdivided into 3- and 28-day subgroups. Rats were sacrificed on days 0, 1, 3, 7, 17 and 28, and CXCL12 levels and expression of various markers of autophagy were assessed at each time points.
Expression of endogenous CXCL12 protein was largely unchanged in sham-operated rats. However, in the nerve-injured experimental group, CXCL12 expression transiently increased 1 day after FNI, decreased at 14 days, and returned to normal by 28 days. These dynamic changes in the expression of CXCL12 increasing after FNI and later decreasing – suggest that CXCL12 may be involved in repair of FNI. Administration of CXCL12 after FNI increased levels of LC3B II and decreased p62, two markers of autophagy, in a time-dependent manner in Schwann cells, producing a maximum effect at 24 hours. Treatment with CXCL12 also markedly reduced the expression of p-PI3K, p-AKT, p-mTOR and p-S6, and significantly increased the number of autophagosomes after 24 hours compared with the control group, suggesting that CXCL12 increases autophagy in Schwann cells. Taken together, these observations demonstrate that CXCL12 exerts a therapeutic effect on FNI. The fact that CXCL12 plays a pivotal role in the regulation of Schwann cell migration through enhancement of autophagy by the PI3K-AKT-mTOR signaling pathway further reinforces the possible importance of CXCL12 in FNI.
3-2-5. Resveratrol
Resveratrol, which has been investigated for potential therapeutic interventions by virtue of its multifarious effects, has been shown to modulate autophagy by influencing the key autophagy-regulating signaling pathways, mTOR and AMPK. In one study investigating the potential of resveratrol to promote recovery after nerve injury [26], resveratrol (100 mg/kg/d) was administered intraperitoneally for 7 days to rats subjected to sciatic nerve crush injury (SNCI). In addition to the resveratrol treatment group (crush+Res) were a 3-MA treatment group (crush+3-MA), a vehicle control group (crush+Veh) and a sham-operated group. SFI, measured as an index of functional recovery, was determined for each rat on days 7 and 14 after SNCI. SFI was significantly higher in the crush+Res group on days 7 (p < 0.01) and 14 (p < 0.05) after nerve injury compared with the control group. On day 28, the SFI for the crush+Res group was higher than that for crush+Veh and crush+3-MA groups (p < 0.05). The SFI for the crush+3-MA group was lower than that for the crush+Veh group (p < 0.01). Levels of the myelin-specific protein, MPZ, and autophagy-related proteins, LC3B, p62 and beclin 1, in sciatic nerve slices were confirmed by Western blotting. On day 4 post SNCI, the expression of MPZ in the distal end of the injured nerve was significantly lower than that in the control group. Levels of LC3-II/I and beclin 1 were highest in the crush+Res group and lowest in the sham group. Collectively, these results show that resveratrol can promote the recovery of damaged peripheral nerves, consistent with previous reports that resveratrol improves autophagy in Schwann cells, helps peripheral nerve repair, and partially accelerates the process of Wallerian degeneration after SNCI.
3-2-6. Nerve growth factor
Nerve growth factor (NGF) has an established role in nerve development and repair and is known to modulate the AMPK/mTOR pathway; as such, it has the potential to modulate autophagy and contribute to nerve damage and recovery mechanisms. The role of NGF in modulating autophagy was investigated [31] in a rat sciatic nerve crush-injury model in which 8-week-old male Wistar rats subjected to nerve injury were administered 0.2 mL of an NGF solution (20 μg/ml), designated the peripheral nerve injury plus NGF (PNI+NGF) group, or the same amount of saline (PNI group), injected intramuscularly once a day for 5 days. NGF treatment (PNI+NGF group) increased the fluorescence intensity of the autophagy marker protein, LC3, compared with saline (PNI group). The regulation of NGF-mediated autophagy in Schwann cells during Wallerian degeneration by p75NTR/AMPK/mTOR signaling was confirmed by examining the levels of p-AMPK, p-p70s6k, and p-mTOR in the sciatic nerve of injured mice by Western blotting 5 days after PNI. Compared with results obtained in the control group, the p-AMPK/AMPK ratio increased after PNI whereas p-mTOR/mTOR and p-p70s6k/p70s6k ratios decreased after PNI. p75NTR is highly expressed in Schwann cells [27], and NGF stimulates downstream signaling pathways by activating p75NTR [28]. AMPK/mTOR signaling is also known to exert neuroprotective effects against focal cerebral ischemia through activation of autophagy [29], and AMPK upregulates autophagy by inhibiting mTOR activation [30]. Collectively, these studies show that administration of NGF not only activates autophagy in dedifferentiated Schwann cells and accelerates myelin debris removal and phagocytosis, it also promotes axon and myelin regeneration in the early stage after PNI.
3-2-7. Lentinan
Lentinan, a natural product with broad pharmacological activity, can modulate the autophagy process by which Schwann cells remove myelin fragments after PNI. The potential to harness this activity the promote recovery after nerve injury was investigated in rats subjected to sciatic nerve compression injury (SNCI) [32]. After injury, the experimental group received 20 mg/kg of lentinan by intraperitoneal injection every day for 14 days, and the control group was injected with normal saline, after which neurological recovery was evaluated in each group. Interestingly, although both groups showed improvement in sciatic nerve function over time, a difference between the control group and the lentinan group appeared after 7 days of treatment and became more pronounced after 2 weeks, at which point the SFI for the lentinan group was significantly higher than that for the PNI (saline-treated) group. In addition, the lentinan group showed delayed muscle atrophy and a much higher muscle-wet-weight ratio than the PNI group. These results suggest that lentinan can significantly promote early functional recovery after SCNI. Lentinan promoted neurological recovery by stimulating autophagic flux in vivo through the AMPK/mTOR signaling pathway, accelerating the removal of myelin debris by Schwann cells and inhibiting neuronal cell death. It was further found that the LC3-II/LC3-I ratio, an important marker of autophagic flux that reflects the involvement of LC3 in the formation of autophagosomes during autophagy induction, increased in the PNI group after nerve compression injury. Lentinan also promoted phagocytosis of myelin fragments by Schwann cells in vitro. Collectively, these results suggest that lentinan accelerates the autophagic clearance of myelin debris in Schwann cells, a process likely regulated by the AMPK/mTOR signaling pathway that mainly promotes nerve regeneration.
3-2-8. Adipose-derived stem cells
Adipose-derived stem cells (ADSCs) have the capacity to bolster nerve regeneration through modulation of autophagy activity. The contribution of ADSCs in harnessing autophagy-related mechanisms for effective nerve repair was investigated [33] in a rat PNI model in which sciatic nerves of Sprague-Dawley rats (n = 12) were crushed with hemostatic forceps (three times for 10 seconds each at 10-second intervals) and then divided into the following groups: Control group (sham+PBS), Model group (crush+PBS), ADSCs+Mel group (crush+ADSCs+melatonin), and 3-MA group (crush+3-MA). Compared with the control group, the SFI for the Model group decreased 21.1-fold (p < 0.01), indicating limb dysfunction after nerve injury. SFI values exhibited time-dependent increases in treatment group rats, measured 1, 2, 3 and 4 weeks after surgery, indicating improved recovery of neurological function in treated rats compared with crush+PBS rats (p < 0.01). Recovery of regenerated nerves 4 weeks after treatment, based on SFI values, was best for the ADSCs+Mel group compared with Mel and ADSC groups (p < 0.01). The ADSCs+Mel group showed significantly increased myelin regeneration and number of motor neurons as well as reduced gastrocnemius atrophy. An analysis performed 1 week after PNI showed a significant increase in the number of autophagosomes and lysosomes and elevated expression of LC3-II/LC3-I and beclin-1 protein in the ADSCs+Mel group compared with the Model group; by comparison, these indicators were significantly decreased in the 3-MA group. After 4 weeks, sciatic nerve function in the ADSCs+Mel group was similar to that of the Control group. A quantitative analysis revealed an increase in the LC3-II/LC3-I ratio and protein levels of beclin-1 in the Model group compared with the control group (p < 0.01). Beclin-1 levels were also increased in ADSCs, Mel, and ADSCs+Mel groups compared with controls (p < 0.01). In addition, the LC3-II/LC3-I ratio and beclin-1 protein levels were higher in the ADSCs+Mel group (p < 0.01) compared with ADSC- and Mel-only groups. The effects of 3-MA on LC3-II/LC3-I ratio and beclin-1 protein levels were opposite those of the ADSCs+Mel group (p < 0.01). This study showed that various indices of nerve regeneration were significantly reduced after treatment with the autophagy inhibitor 3-MA. Taken together with the improved outcome observed in the ADSCs+Mel group, these observations suggest that ADSCs combined with Mel can promote sciatic nerve regeneration in rats by altering autophagy activity in the early stages of sciatic nerve injury.
3-2-9. Basic fibroblast growth factor
Although studies have shown that basic fibroblast growth factor (bFGF) can activate autophagy and promote peripheral nerve repair, the role of bFGF in the facial nerve and its molecular mechanism of action are not clear. In a study designed to address this question [34], male Sprague Dawley (SD) rats were randomly divided into five groups (n = 10/group): sham group, FNI group (FNI+saline), poloxamer group (FNI+poloxamer), bFGF group (FNI+bFGF) and P-bFGF group (FNI+P-bFGF). The FNI model was created by clamping the facial nerve for 60 seconds with hemostatic forceps. Normal saline (control) or bFGF (0.1 mg/ml) was administered on days 0, 3 and 7, and rats were sacrificed on day 7. Facial nerve morphological abnormalities induced by FNI were improved in bFGF and P-bFGF groups compared with the FNI group. Nerve fibers in the P-bFGF group displayed significantly more regular regeneration than those in the bFGF group. The fluorescence intensity of LC3B was slightly increased in the FNI group, but was enhanced to a greater degree in bFGF and P-bFGF treatment groups, with the P-bFGF group showing significantly greater LC3B intensity than the bFGF group. Western blot analyses showed that LC3B-II, beclin 1 and ATG5 expression paralleled changes in the fluorescence intensity of LC3B. Taken together, the results of this study showed that P-bFGF effectively promotes cell proliferation, myelination and functional recovery, and reduces neuronal apoptosis after FNI.
3-2-10. Facilitation by epothilone B
Epothilone B (EpoB) is an FDA-approved antineoplastic agent known for its ability to induce α-tubulin polymerization and improve microtubule stability. It has also recently gained attention for its regenerative effects on the central nervous system. However, the potential therapeutic effects of EpoB on peripheral nerve regeneration has only begun to be investigated. In one such study [35], a sciatic nerve injury model was created by compressing the sciatic nerve of adult male Sprague–Dawley rats with a vascular clamp for 50 seconds. Rats in the EpoB group (n = 10 rats/group) received an intraperitoneal injection of 1 ml of 150 µg/ml EpoB every day for 7 days. SFI values were not significantly different between control and EpoB groups within the first 2 weeks. However, the SFI of the EpoB group was significantly increased at both 3 weeks (-27.50 ± 1.58) and 4 weeks (-17.32 ± 2.17) compared with the control group, where the 4-week value was -29.73 ± 1.97 (p < 0.05 at both time points), suggesting that EpoB treatment achieved superior recovery of motor function. At 24 hours after EpoB treatment, LC3B II levels were significantly increased, and PI3K and Akt phosphorylation were significantly inhibited, in Schwann cells (p < 0.05). Transmission electron microscopy further showed that EpoB significantly increased the number of autophagosomes (5 ± 1) in Schwann cells at this time point compared with controls (2 ± 1; p < 0.05). To further investigate the role of autophagy in EpoB-enhanced SC migration, this study also tested the effect of 3-MA on cell migration, demonstrating that this autophagy inhibitor significantly inhibited EpoB-induced autophagosome production in Schwann cells (p < 0.05) and abolished EpoB-enhanced migration of these cells (p < 0.05). Collectively, these results suggest that EpoB induces autophagy and that this autophagy is involved in EpoB-enhanced migration of Schwann cells.
4. Conclusions
Although research on the facial nerve remains limited, studies of autophagy conducted on peripheral nerves, including the sciatic nerve, have yielded noteworthy findings. These investigations have revealed the expression of a number of autophagy-related substances in compression and transection nerve injury models, including AMPK, Atg3, Atg5, Atg7, Atg9, Atg12, beclin-1, LC3, LC3 II/I, mTOR, PI3K, PI3K/Atk, PI3K-Akt-mTOR and p75NTR/AMPK/mTOR, as well as changes in autophagosome dynamics. In the case of PNI, administration of autophagy inducers or inhibitors increased or decreased autophagy-related substances, suggesting that autophagy is closely related to peripheral nerve degeneration and regeneration. It is imperative that future studies delve in other unexplored autophagy-related factors post nerve damage and further explore the intricate interplay between autophagy, inflammation- and immunity-related factors, and nerve regeneration-related factors.
Author Contributions
Conceptualization, S.G.Y.; Data curation, Y.J.K., D.K.Y., S.Y.J., S.S.K., J.L., and J.H.Y.; Formal analysis, J.L., J.H.Y., and J.M.L.; Funding acquisition: S.S.K. and S.G.Y.; Methodology, D.C.P., Y.J.K., D.K.Y., and J.M.L.; validation, Project administration, D.C.P. and S.G.Y.; Visualization, D.K., D.C.P., and S.S.K.; Writing - original draft, D.K.Y., Y.J.K., D.C.P., and S.G.Y.; Writing - review & editing, D.C.P. and S.G.Y. All authors have read and agreed to the published version of the manuscript.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgement
This work was supported by a National Research Foundation of Korea (NRF) grant funded by the Korea government (MSIT) (NRF 2018R1A6A1A03025124)(NRF 2019R1A2C1086807)(NRF 2022R1A2C1091779). This research was supported by a grant of the Korea Health Technology R&D Project through the Korea Health Industry Development Institute (KHIDI), funded by the Ministry of Health & Welfare, Republic of Korea (grant number: HV22C0233). The funders had no role in study design, data collection, data analysis, data interpretation, or writing of the report.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kim, K.H. The Role of Autophagy in the Pathogenesis and Treatment of Metabolic Diseases. Molecular and Cellular Biology Newsletter. 2004, 1–9. [Google Scholar] [CrossRef]
- Dobie, R.A. Test of facial nerve function. In Otolaryngology Head and Neck Surgery, St. Louis Mosby, 3rd ed.; Cummings, C.W., Fredrickson, J.M., Harker, L.A., et al., Eds.; 1998; pp. 2757–2766. [Google Scholar]
- Byun, J.Y. Facial paralysis disorders. Anatomy and evaluation of facial nerve. In Korean Society of Otorhinolaryngology−Head and Neck Surgery, 3rd ed.; Seoul. KoonJa, 2018; pp. 913–932. [Google Scholar]
- Hu, Y. Axon injury induced endoplasmic reticulum stress and neurodegeneration. Neural Regen Res. 2016, 11, 1557–1559. [Google Scholar] [CrossRef]
- Kim, K.H.; Lee, M.S. Autophagy–a key player in cellular and body metabolism. Nat Rev Endocrinol. 2014, 10, 322–37. [Google Scholar] [CrossRef]
- Ichimura, Y.; Komatsu, M. Pathophysiological role of autophagy: lesson from autophagy-deficient mouse models. Exp Anim. 2011, 60, 329–345. [Google Scholar] [CrossRef]
- Tsukada, M.; Ohsumi, Y. Isolation and characterization of autophagy-defective mutants of Saccharomyces cerevisiae. FEBS Lett. 1993, 333, 169–174. [Google Scholar] [CrossRef]
- Mizushima, N.; Yoshimori, T.; Ohsumi, Y. The role of Atg proteins in autophagosome formation. Annu. Rev. Cell Dev. Biol. 2011, 27, 107–132. [Google Scholar] [CrossRef]
- Russell, R.C.; Yuan, H.X.; Guan, K.L. Autophagy regulation by nutrient signaling. Cell Res. 2013, 24, 42–57. [Google Scholar] [CrossRef]
- Nixon, R.A. The role of autophagy in neurodegenerative disease. Nature medicine. 2013, 19, 983–997. [Google Scholar] [CrossRef]
- Shntani, T.; Klionsky, D.J. Autophagy in health and disease; A double-edged sword. Science. 2004, 306, 990–995. [Google Scholar] [CrossRef]
- Cuervo, A.M. Autophagy; in sickness and in health. Trends in Cell Biol. 2004, 14, 70–77. [Google Scholar] [CrossRef]
- Mehrpour, M.; Esclatine, A.; Beau, I.; Codogno, P. Autophagy in health and disease. 1. Regulation and significance of autophagy: an overview. Am J Physiol Cell Physiol. 2010, 298, C776–85. [Google Scholar] [CrossRef] [PubMed]
- Choi, A.M.; Ryter, S.W.; Levine, B. Autophagy in human health and disease. N. Engl. J. Med. 2013, 368, 651–662. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Sanchez, J.A.; Carty, L.; Iruarrizaga-Lejarreta, M.; Palomo-Irigoyen, M.; Varela-Rey, M.; Jessen, K.R. Schwann cell autophagy, myelinophagy, initiates myelin clearance from injured nerves. J Cell Biol. 2015, 210, 153–68. [Google Scholar] [CrossRef] [PubMed]
- Romeo-Guitart, D.; Leiva-Rodriguez, T.; Forés, J.; Casas, C. Improved Motor Nerve Regeneration by SIRT1/Hif1a-Mediated Autophagy. Cells. 2019, 30, 1354. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.C.; Chen, L.; Zhang, H.X.; Li, S.F.; Li, C.X. Autophagy Promotes Peripheral Nerve Regeneration and Motor Recovery Following Sciatic Nerve Crush Injury in Rats. J Mol Neurosci. 2016, 58, 416–23. [Google Scholar] [CrossRef] [PubMed]
- Ko, P.Y.; Yang, C.C.; Kuo, Y.L.; Su, F.C.; Jou, I.M. Schwann- Cell Autophagy, Functional Recovery, and Scar Reduction After Peripheral Nerve Repair. J Mol Neurosci. 2018, 64, 601–610. [Google Scholar] [CrossRef] [PubMed]
- Inada, T.; Sato, H.; Hayashi, Y.; Hitomi, S.; Ando, M.; Shinoda, M. Rapamycin Accelerates Axon Regeneration Through Schwann Cell-mediated Autophagy Following Inferior Alveolar Nerve Transection in Rats. Neuroscience. 2021, 468, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Li, X.; Li, Q. Curcumin accelerates the repair of sciatic nerve injury in rats through reducing Schwann cells apoptosis and promoting myelinization. 2017, 92, 1103–1110. [Google Scholar] [CrossRef]
- Shintani, K.; Uemura, T.; Takamatsu, K. Protective effect of biodegradable nerve conduit against peripheral nerve adhesion after neurolysis. J Neurosurg. 2017, 20, 1–10. [Google Scholar] [CrossRef]
- Chang, Y.C.; Chen, M.H.; Liao, S.Y. Multichanneled nerve guidance conduit with spatial gradients of neurotrophic factors and oriented nanotopography for repairing the peripheral nervous system. ACS Appl Mater Interfaces. 2017, 9, 37623–37636. [Google Scholar] [CrossRef]
- Salehi, M.; Naseri-Nosar, M.; Ebrahimi-Barough, S. Polyurethane/gelatin nanofibrils neural guidance conduit containing platelet-rich plasma and melatonin for transplantation of schwann cells. Cell Mol Neurobiol. 2018, 38, 703–713. [Google Scholar] [CrossRef]
- Qian, Y.; Han, Q.; Zhao, X.; Song, J.; Cheng, Y.; Fang, Z.; Qian, Y. 3D melatonin nerve scaffold reduces oxidative stress and inflammation and increases autophagy in peripheral nerve regeneration. J Pineal Res. 2018, 65, e12516. [Google Scholar] [CrossRef] [PubMed]
- Gao, D.; Tang, T.; Zhu, J.; Tang, Y.; Sun, H.; Li, S. CXCL12 has therapeutic value in facial nerve injury and promotes Schwann cells autophagy and migration via PI3K-AKT-mTOR signal pathway. International journal of biological macromolecules 2019, 124, 460–468. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Ren, J.; Liu, Y.; Huang, D.; Lu, L. Resveratrol regulates the recovery of rat sciatic nerve crush injury by promoting the autophagy of Schwann cells. Life Sci. 2020, 256, 117959. [Google Scholar] [CrossRef] [PubMed]
- Bothwell, M. Functional interactions of neurotrophins and neurotrophin receptors. Annu Rev Neurosci. 1995, 18, 223–53. [Google Scholar] [CrossRef]
- Zampieri, N.; Chao, M. Structural biology. The p75 NGF receptor exposed. Science. 2004, 304, 833–4. [Google Scholar] [CrossRef]
- Jiang, T.; Yu, J.T.; Zhu, X.C.; Wang, H.F.; Tan, M.S.; Cao, L. Acute metformin preconditioning confers neuroprotection against focal cerebral ischaemia by pre-activation of AMPK-dependent autophagy. Br J Pharmacol. 2014, 171, 3146–57. [Google Scholar] [CrossRef]
- Matsui, Y.; Takagi, H.; Qu, X.; Abdellatif, M.; Sakoda, H.; Asano, T. Distinct roles of autophagy in the heart during ischemia and reperfusion: roles of AMP-activated protein kinase and Beclin 1 in mediating autophagy. Circ Res. 2007, 100, 914–22. [Google Scholar] [CrossRef]
- Li, R.; Li, D.; Wu, C.; Ye, L.; Wu, Y. , Yuan, Y.; Xiao, J. Nerve growth factor activates autophagy in Schwann cells to enhance myelin debris clearance and to expedite nerve regeneration. Theranostics. 2020, 10, 1649–1677. [Google Scholar] [CrossRef]
- Xiao, H.; Wei, C.; Liu, H.; Li, Z.; Zheng, C.; Luo, J. Lentinan alleviates sciatic nerve injury by promoting autophagy to remove myelin fragments. Phytother Res. 2023, 10. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, M.; Zhang, Z.; Sun, Y.; Wang, J.; Liu, Y. ADSCs Combined with Melatonin Promote Peripheral Nerve Regeneration through Autophagy. Int J Endocrinol. 2022, 5861553. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Zhang, H.; Xu, M.; Li, L.; Wu, M.; Zhang, S.; Ni, L. Delivery of Basic Fibroblast Growth Factor Through an In Situ Forming Smart Hydrogel Activates Autophagy in Schwann Cells and Improves Facial Nerves Generation via the PAK-1 Signaling Pathway. Front Pharmacol. 2022, 13, 778680. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Li, S.; Gao, J.; Hu, Y.; Chen, S.; Luo, X.; Zhang, H.; Luo, Z.; Huang, J. Epothilone B Facilitates Peripheral Nerve Regeneration by Promoting Autophagy and Migration in Schwann Cells. Frontiers in Cellular Neuroscience. 2020, 14, 143. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Review flow diagram.
Figure 2.
Expression of target substances associated with autophagy in the context of facial and sciatic nerve degeneration and regeneration after nerve injury.
Figure 2.
Expression of target substances associated with autophagy in the context of facial and sciatic nerve degeneration and regeneration after nerve injury.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



