
Submitted:
27 September 2023
Posted:
28 September 2023
You are already at the latest version
Abstract
Cellular aging has drawn the attention of researchers, scientists, and biotech businesses for the treatment of a number of medical conditions. Cellular aging is primarily defined by consistent cessation of proliferative development in response to internal and external stressors, including DNA damage, telomere shortening, mitochondrial dysfunction, and regulation of the senescence-associated secretory phenotype (SASP). Disturbances involving these factors may contribute to age-related disease development. Therefore, the current review aims to explore anti-aging factors targeting DNA damage response and SASP regulation and their detailed signaling networks. In addition, it provides an opportunity for researchers to identify not only anti-aging targets, but also anti-aging therapeutic strategies based on identified and non-identified targets.
Keywords:
Anti-aging
; DNA damage response
; senescence-associated secretory phenotype
1. Introduction
Various progeroid syndromes have been linked to genome instability, indicating an important causal role for DNA damage accumulation in the aging process and in the development of age-related pathologies. Aging is a characteristic of practically all biological organisms and is characterized by a steady deterioration in the function of various cells and organs. According to a World Health Organization (WHO) report, by 2030, one in every six people on the planet will be 60 years old or older [1]. Cellular aging is a dynamic and versatile process. This response is initiated by various intrinsic and extrinsic triggers, including oncogenic activation, oxidative and genotoxic stress, telomeres, DNA damage, mitochondrial dysfunction, irradiation, and chemotherapeutic agents. This response is linked to various simple and complicated processes, such as embryogenesis, tumor suppression, organismal aging, and tissue repair [2,3], indicating the importance of DNA damage accumulation during the aging process.
An influential discovery in cellular senescence was that it is a symbol of aging. In 1961, Hayflick and colleagues stated that human fibroblasts demonstrate a finite capacity for cell division before irreversible growth arrest, known as replicative senescence, leading to the hypothesis that aging is caused by cells losing their proliferative ability progressively, which is important to replace damaged or aging cells that naturally accumulate over time. Several studies have been conducted over the last decade to demonstrate the role of excessive or accumulated aging cells in promoting organismal aging and related disabilities [3]. Accumulation of aging cells may disturb the normal cellular environment, ultimately generating abnormal responses and resulting in various aging-related diseases. These diseases include cancer, cardiovascular diseases, and neurodegeneration. Multiple factors are associated with anti-aging, which encourage aging and aging-related diseases [2,3].
However, studying the factors involved in cellular aging, either directly or indirectly, may provide a pathway to develop anti-aging therapies. The theory of this review is based on an in-depth study of the pathological processes involved in identifying targets. When a target is indicated or marked, it is relatively easy to identify substantial therapies for such pathological processes or conditions.
DNA is a complex molecule that contains all the information necessary to develop and maintain an organism. Defects in DNA repair accelerate human aging and, in return, these aging cells produce the senescence-associated secretory phenotype (SASP), which has been considered a focal driver of age-related dysfunction [4]. Therefore, mediating SASP may promote the repair of damaged tissues by correcting the DNA damage response, either to delay aging or to provide protection to aging cells for further damage. Saying good bye to this world or to loved ones is a painful and heart-breaking process. This is the point at which researchers and scientists are unable to bring their loved ones back.
In this review, we explain and highlight natural products and synthetic compounds that are connected either directly or indirectly to the DNA damage response (DDR)or SASP in relation to aging.
Bioactive compounds from natural or synthetic sources have advantages such as diverse structures, affordability, availability, and effectiveness with multiple targeted deliveries to mediate different signaling pathways of aging with the ability to correct the underlying signaling pathway or pathological condition. Therefore, the main aim of this paper is to highlight the hidden factors involved in the prevention, delay, elimination, and replacement of aging cells, or encouraging the production of young cells. Studying these factors and their underlying mechanisms may energize researchers to develop or explore novel anti-aging approaches that can bring more happy moments or add healthier and more valuable years to human life.
2. Materials and Methods
Comprehensive work was conducted to collect data from different authentic electronic resources and databases, including PubMed, Google Scholar, and the Web of Science. We explored natural products and synthetic compounds that specifically target the aging mechanism to enhance a healthy lifespan as an anti-aging therapy.
3. Results
3.1. Signaling pathway involving aging and related aging inducers
Cellular aging is considered a stable and terminal condition of cell growth arrest, in which cells are unable to proliferate despite favorable growth conditions and mitogenic stimuli [4,5]. Aging cells exhibit greater resistance to apoptotic cell death owing to the upregulation of cell survival pathways and exposure to external stress. This prolonged viability may be due to two reasons: either selection of the most death-resistant cells or the involvement of intrinsic properties in the senescence pathway that remain to be elucidated. However, the main factors responsible for the initiation of cell aging and cell fate must be studied in depth. Cell fortune may depend on several factors, such as the type of cell, nature of damage, and duration and intensity of the initial stimulus. This is because cell aging and cell death are interconnected by different signaling pathways, including p53 pathway activation [6]. In addition, it may be possible that cell resistance to apoptotic cell death may depend on p53 activity and levels. However, aging is considered a permanent condition of cell cycle arrest. However, recent trends have shown that cellular aging, which focuses on cancer and tumor cells, may involve epigenetic mechanisms that rearrange cancer cells to a certain level of self-renewal and differentiation in a cell-autonomous manner. Aging is a dynamic process in which several distinct pathways overlap at different stages [7].
3.2. DNA damage response in cellular aging
Various stimuli initiate cellular aging. Among them, nuclear DNA damage is often considered to be the main cause in the form of DNA double strand breaks (DSBs) that are considered to be responsible to activate the DNA damage response pathway [8]. This pathway acts as a barrier to control and block cell cycle progression and prevents the transfer and control of corrupted information to next-generation cells. Several factors involved in DDRs accumulate at DNA damage sites and form a cytologically notable nuclear foci [9]. These nuclear foci contain various events of chromatin modification, such as phosphorylation of histone H2AX and its interlinked proteins, including the activated form of the kinase ataxia telangiectasia, 53BP1, and MDC1. These foci identify each DNA damage site and participate in barrier enforcement and cell cycle arrest. These processes continue until the damage is repaired. If DNA damage continues, it causes extended DDR signaling along with extended proliferative arrest in the form of cellular aging. An early investigation showed that the extended DDR foci detected in cultured aging cells consist of unrepaired DSBs, validating the concept that aging cells are similar to extended barrier activation [10]. Inactivation of DDR signaling kinases, such as CHK2, CHK1, ATR, and ATM, may motivate aging cells to progress into the cell cycle. The tumor suppressor protein p53 stimulates and activates cyclin-dependent kinase inhibitor p21 at the end of the DDR cascade [11]. p16 plays important roles in various aging-related signaling pathways. Both p21 and p16 play roles in cellular aging; p21 plays a role in the early phase of aging [12], while p16 plays a role in the late phase for maintenance of the aging phenotype [13]. Furthermore, along with activation of the DDR cascade, ARF, a tumor suppressor, calms p53, which is involved in the activation of aging. Scientists are attempting to unfold the contributions of two major pathways, the ARF and DDR pathways [14], to p53 and the dependent aging response in cancer. Research indicates that ARF and DDR play antagonistic roles, as in cancer progression, and that ARF activation occurs in a DDR-independent manner [15]. Recently, a regulatory network was identified in a human cancer model, which revealed that ATM suppresses ARF levels [16], and that ARF acts as a barrier to cancer progression when ATM is inactivated. Based on this temporal regulation, DDR leads to ARF engagement, whose activation is noticed in the later phase of tumor development and is less regular than that of DDR [17].
3.3. Telomere shortening and cellular ageing
Over time, aging organisms accumulate cells that participate in body dysfunction. One of the best identified and characterized mechanisms involved in aging and cellular aging is telomere shortening or dysfunction [18]. Therapeutic options for aging may also involve identification of the appropriate cause of telomere shortening below a specific length or dysfunction. Exploring the underlying mechanisms regarding telomerase shortening involves the activation of telomerase activity, either via endogenous TERT expression reactivation [19] or its exogenous transportation. Shortening of telomeres gradually causes activation of DDR and cellular aging, which contributes to aging and aging-related diseases [18]. However, telomere shortening is not essential for predicting lifespan, nor is it responsible for a short lifespan. Telomere shortening may also trigger DDR and promote cellular aging [20]. DDR activation leads to the formation of telomere-stimulated DNA damage foci (TIFs) and interlinked DDR foci (TAFs). Therefore, TIFs and TAFs are considered markers of cellular aging. Because of continuous DDR activation and localization of DSBs within telomeres, cell repair is much less efficient. Persistent localization of DSBs in telomeres maintains cellular aging and stability. Therefore, continuous DDR activation, which triggers cellular aging, can occur in both proliferating and non-proliferating cells [21].
Another study indicated that telomere dysfunction activates DDR to cause cellular aging with the help of SASP, which ultimately leads to the loss of tissue function related to aging [18]. However, telomere dysfunction contribution towards aging needs to be further explored to better understand the association between aging and age-related diseases. Furthermore, another study in a progeria animal model revealed that telomeric also reduces DDR activation, improve lifespan, and work independently of genetic defects [22,23].
3.4. Mitochondrial dysfunction and cellular aging
Abnormal mitochondrial function is associated with increased oxidative stress in aging cells. Aging cell characteristics may depend on mitochondrial structure, mass, and membrane proteins [24]. Mitochondrial sirtuins are responsible for triggering aging across different species by controlling mitochondrial function, owing to their decreased activity or expression with age [25]. In addition, a decline in NAD+ levels with age can reduce sirtuin activity, ultimately contributing to cellular aging. Boosting levels of NAD+ may potentially increase lifespan and is considered a therapeutic strategy for aging by controlling mitochondrial functions [26].
3.5. Oncogene-induced aging
Oncogene activation is considered a potent inducer of cellular aging that activates an early hyperproliferative stage [27]. This stage is further linked to altered DNA replication and eventually initiates DDR pathways, leading to aging. Proliferation arrest can also occur due to the loss of expression of cancer suppressor genes such as PTEN. However, in the beginning, PTEN loss-induced aging is not interlinked with the activation of DDR, but later, via in vivo studies, a linkage between cellular aging, DDR engagement, and hyperproliferation was found [28]. Importantly, unlike the tumor-causing genes BRAF or RAS, P13-AKT pathway activation triggers p53-depedent aging and is frequently triggered in the absence of potential accumulation of DNA damage and noticeable hyperproliferation. All these findings showed a distinct underlying process. Reactive oxygen species accumulate in tumors and cause aging-interlinked DNA damage and induce aging [29].
3.6. Chromatin changes in aging cells
Several aging cells exhibit considerable alterations to their chromatin and epigenetic networks [30]. These alterations are not only associated with aging cells, but also have been found on nearby cells. Ageing-interlinked heterochromatin foci (SAHF) are spatially arranged heterochromatin domains [31]. These domains are identified as dense 4ʹ,6-diamidino-2-phenylindole (DAPI)-positive nuclear structures. These structures are rich in repressive chromatin proteins and markers, including ASFIA, histone co-chaperone HIRA, histone variant macroH2A, high-mobility group protein A, heterochromatin protein 1, and trimethylated histone H3 Lys9. However, SAHF are not considered universal markers of aging. They are usually observed upon tumor activation and exhibit an ATR-dependent manner of DNA replication. Originally, SAHF was thought to repress genes promoting cell cycle arrest [32]. SAHF also promotes a DDR-resistant heterochromatin structure that prevents DDR signaling. In reality, treatment with histone deacetylase inhibitors (HDI) can activate chromatin relaxation and trigger DDR signaling following cell death via apoptosis [33]. This treatment is considered the first senolytic style. This was later supported by the senolytic activity of Panobinostat, an HDI. HDI can trigger cellular aging in normal fibroblasts, which may be due to its effect on DDR signaling [34].
Another study concerning the characteristics of aging cells explored basic heterochromatin domains considered primarily by the distension of pericentromeric satellite sequences that are observed in various species, which result from different mechanisms involving aging triggers [35]. These chromatins structural changes are not only connected to specific repressive histone mark removal, but are also interlinked with alterations involving nuclear structural proteins, such as nuclear lamina loss. This loss releases cytosolic chromatin fragments (CCFs) into the cytoplasm of aging cells [36]. However, oncogene-induced aging, telomere shortening, and DNA damage also contribute to CCfs. However, the formation of CCFs in prolonged aging cells remains to be explored. Notably, CCFs suggest aging-interlinked pancreatic function via cyclic GMP-AMP synthase activation and interferon gene pathway stimulation [37]. Studies have revealed that CCFs can be reduced by using HDI to reduce the gain of the interlinked secretory phenotype.
Recent advances in genomic technology and chromatin modification have led to the elucidation of signaling pathways involved in development and maintenance [38]. Gene-poor regions show DNA hypermethylation during replicative senescence, whereas cancer-suppressor genes display focal hypermethylation. This evidence suggests that aging cells may be epigenetically well-informed regarding cancer alterations. However, later on, this idea was challenged by research showing that oncogene-induced aging exhibits only limited alterations in methylation patterns in comparison to non-oncogene-induced aging cells [39]. This indicates that cancer-interlinked methylome alterations may occur randomly and autonomously during the aging stage. However, a study on DNA methylation alterations revealed that late replicative aging fibroblasts and oncogene-induced aging cells display a noticeable increase in chromatin accessibility at the nucleosome level, along with changes in open chromatin regions towards mediatory elements and repeats [40]. Loosening of chromatin regions leads to an increased expression of transposable elements, which are usually silenced and dormant in unstressed cells [41]. Although transposable elements play a well-defined role in initiating genomic instability through transposition, the reactivation of these transposable elements plays an important role in regulating the non-cell-autonomous functions of aging cells. Numerous studies on genome of H3 Lys27 acetylation, H3 Lys 27 trimethylation, and H3 Lys 4 trimethylation in aging cells have revealed the active activation and deactivation of a wide range of chromatin domains that were considered to be involved in the regulation of aging downstream effectors [42]. Increasing histone levels, HDAC Rpd3 inactivation, decreasing acetylation of H4 K16 and H3 N-terminal, or blockage of signals released from DNA in response to telomere shortening may extend lifespan or lead to a more youthful state [43]. Detailed signaling pathway targeting the DDR is as follows in Figure 1.
3.7. SASP composition and regulation
Another active mechanism through which aging cells exert their pleiotropic biological functions involves SASP program activation and its regulation via growth factors, chemokines, cytokines, and extracellular matrix proteases, which either directly cause aging or influence the nearby environment of aging cells or whole organisms [44]. The activation of the SASP is a self-motivated process that causes aging. SASP was originally defined as an active secretory program involving several bioactive factors, as shown in Figure 2.
The composition of the SASP varies from cell to cell and the type of initial trigger. For example, upon cancer triggering, the amplitude of protein secretion is remarkably high compared to that of other aging-induced factors. Despite the quantitative and qualitative differences among SASP in different tissues and aging models, the main SASP structure consists of monocyte chemoattractant protein 1, CXC chemokine ligand 8, and pro-inflammatory interleukin-6 (IL-6) in all in vitro-generated models [45]. SASP is not only a composite of proinflammatory cytokines but also enzymes involved in ECM (Extra cellular matrix) remodeling, such as tissue metalloproteinase inhibitors, cysteine/serine proteinase inhibitors, and matrix metalloproteinases. A more recent study concerning SASP structural characterization identified specific diverse SASP effectors released with a set of other components that are involved in aging and aging-related diseases [46]. Exosomes are considered the main mediators and controllers of the paracrine aging effect of the SASP and promote tumorigenesis.
Notably, DDR activation promotes SASP in neighboring cells in inflamed ageing [17]. Activation of these two factors may promote the activation of different pro-inflammatory factors, including reactive oxygen species, matrix-degrading enzymes, growth factors, chemokines, and interleukins. Furthermore, NF-KB play a key role to induce a variety of inflammatory SASP contents such as IL-1, IL-6, and TNF-α in an experimental inflammation model. In addition, DDR- and SASP-activated cells release miRNAs into non-senescent cells to promote inflamed aging. However, the exact role of miRNAs is not clear, as miRNAs may protect cells from inflammation; however, they may promote senescence in younger cells via inflammatory pathways. Another recent study identified a correlation with induced cell proliferation [47]. Interestingly, another study revealed that the nuclei of senescent cells abruptly accumulate DNA fragments that control the intrinsic mechanism involved in the production of SASP factors by targeting the activation of the CGAS-sting pathway. This initiative facilitates the type 1 interferon producing pathways with the help of IRF3. Another study support the DDR and SASP relationship indicating that partial sleep may induce gene expression which facilitate the DDA damage accumulation and promote the SASP expression in proinflammatory profile which in turn initiate NF-KB transcriptional regulation [48].
P16 interference can potentially cause senescence but is usually unable to initiate the full SASP cascade [49]. Continuous DDR signaling is required to stimulate inflammatory cytokine secretion. In addition, consistent DDR signaling promotes the paracrine and cell-autonomous functions of aging cells, CHK2 and NBS1, and ATM prime SASP genes for activation [50]. However, hindrance of DDR signaling may reduce the production of cytokines due to genotoxic stress [51]. The opposite effect has been observed in response to p53 inhibition, as it further encourages aging, which in turn generates an inflammatory microenvironment that encourages malignant transformation and anti-aging.
Recently, it has been observed that ATM helps modulate SASP gene expression by moderating the scrapping of modified histone macroH2A1.1 from the SASP genotype to counter oncogenic damage along with DNA disruption [52]. Because DDR activation is a rapid response, whereas SASP establishment is gradual, the SASP must be controlled by additional mechanisms. In fact, even in the absence of DNA disruption, activation of the stress-inducible MAPK p38 was shown to be both essential and sufficient to cause growth arrest and SASP induction via stimulating NF-Kβ activity [53]. Although DDR and p38 activation are separate and work independently, they can activate the SASP.
Aging is one of the variables that continuously causes an elevation in DDR signaling in both proliferating and non-proliferating cells, as well as helping considerably to the build-up of aging cells as people age. Insufficient DNA repair may also play a role in the build-up of DNA damage, activation of DDR, and extensive chromatin alterations [54]. These observations have been made in various cell types and at the organismal level as people age. DDR is a metabolic reprogramming driver that assists with the SASP. As a result, the control of SASP by DDR via numerous pathways could be one of the mechanisms that help DDRs promote aging-associated inflammation [55].
3.8. Transcriptional and epigenetic control of SASP
SASP is primarily controlled at the transcriptional level, and several transcription components and chromatin controllers are involved in its regulation [56]. The transcription factors NF-Kβ and CCAAT/enhancer-binding protein (C/EBP) bind to SASP gene promoters and regulate their activity. Furthermore, the transcription factor GATA4 activates genes that encode IL-6, IL-8, CXCL1 (also known as GRO), granulocyte-macrophage colony-stimulating factor, and ECM proteases and inhibitors, all of which have been implicated in immune responses and inflammation [57]. Because GATA4 levels rise after DDR activation [58] and in senescent cells, it has been proposed that GATA4 serves as a molecular link between DDR signaling and subsequent NF-Kβ activation for full SASP establishment, even though GATA4 regulation of the NF-Kβ pathway occurs indirectly via increased expression and secretion of IL-1, an upstream regulation factor of NF-κβ.
In a model of aging caused by PTEN loss, the JAK2-STAT3 pathway triggers a subclass of SASP components with immunosuppressive characteristics [59]. JAK/STAT inhibitors have been found to successfully rearrange the SASP to improve cancer treatment or remove aging cancer cells controlled by T cells. JAK inhibitors also substantially reduce age-related syndromes in mice. SASP gene expression is time-dependent, and temporal variations in NOTCH1 activity through aging have been shown to influence the SASP structural configuration [60]. Early on, NOTCH1 quantities rise, and NOTCH1 induces growth modifying features and effectors while constraining the proinflammatory arm of the SASP cascade by inhibiting its positive regulator C/EBPβ. In the later phase of aging, the level of NOTCH1 is lowered by the induction of SASP pro-inflammatory cytokines, including IL-1, IL-6, and IL-8 [61]. However, the relationship between ATM inhibition and NOTCHI remains unclear.
In the oncogene induced senescence (OIS), the epigenetic reader bromodomain-carrying polypeptide 4 (BRD4) is drafted as a super-enhancer that is proximally closer to the SASP genes. BRD4 is an acetylated histone-binding protein that is associated with oncogenesis. BRD4 aids in the efficient implementation of cellular aging and, surprisingly, functions as a tumor inhibitor [62]. Undoubtedly, pharmacological and genetic deactivation of BRD4 blunts the SASP, reduces OIS immune-mediated cell removal, and may not be successful in the disposal of damaged cells that are vulnerable to escape from aging. Nonetheless, a small molecule that degrades BRD4 and exhibits senolytic activity was recently discovered by chemical screening. Furthermore, in murine and human cell cultures, BRD4 has been shown to favorably influence telomere elongation [63]. Therefore, BRD4 inhibitors may reduce SASP induction and promote telomere shortening, resulting in increased cell aging. BRD4 competes with polycomb repressor complex 2 (PRC2), which methylates the same histone residue for acetylated histone H3 Lys27 (resulting in trimethylated H3Lys27) [64].
Excessive EZH2 expression prohibits entry into the OIS via various mechanisms, consistent with BRD4 and EZH2 (the catalytic central subunit of PRC2) competing for the same residue and having antagonistic roles [48]. DDR regulation and suppression of SASP gene expression are examples of these mechanisms. MLL1, a transcription-linked histone methyltransferase and oncoprotein, has also been shown to be required for SASP activation; however, this is primarily due to oncogene-linked hyper-replication and DDR interactions, rather than immediate transcriptional regulation of SASP genes.
SASP is also regulated by high mobility group box (HMGB) proteins [65]. In oncogene-based aging cells, HMGB2 directly attaches to and controls SASP gene expression, and its removal reduces SASP without affecting aging proliferation. HMGB1 plays a key role in damage-linked molecular rearrangements, also termed alarmins, which are released to initiate SASP-arbitrated paracrine senescence and activate immunity [66]. In addition, increased nuclear stoma thickness during OIS affects SASP expression by facilitating heterochromatin rearrangement, which is critical for SAHF development.
3.9. SASP and non-specific immunity
cGAS, a DNA sensor, and STING, an accessory protein, have been observed to be important SASP regulators across all species, likely via the activation of NF-Kβ and the interferon response component IRF3. Induction of cGAS-STING happens principally via the identification of ‘self’ double-stranded DNA or in fragments of chromatin present in aging cell cytoplasm [67]. cGAS-STING genetic exhaustion decreases SASP-linked proinflammation and alleviates aging-associated immunosurveillance during aging, as observed in in vivo experiments [68]. In addition to the complex nature of SASP modulation, irregular stimulation of the cGAS-STING mechanism was found to be interconnected with the suppression of DNases, such as DNase 2 and TREX1, which are routinely involved in DNA disruption in the cytoplasm, leading to cytoplasmic interruption of nuclear DNA during the course of aging. Although the phenomena that stimulate the release of cytoplasmic chromatin in proliferation-arrested cells still require in-depth research, these findings reveal that STING suppressors could inherently be applicable for the alleviation of age-based incurable inflammation.
In the stimulation and implementation of the SASP, cGAS-STING is not a single factor that is principally involved. Reportedly inflammasomes and Toll-like receptors increase the maturation and secretion of SASP components. The inflammasome is a combination of several proteins, including caspase 1, which plays an important role in resisting external antigens and is regulated by Toll-like receptors [68].
When enduring longer periods in culture or in vivo, aging cells exhibit intense regulation of interferon type 1 responses and downstream targets. This phenomenon was partially generated by the reinduction of exchangeable segments, resulting in cGAS-STING activation. The aging-linked deleterious implications of the SASP are restricted by its interaction with nucleoside reverse-transcriptase inhibiting factors, mitigating persistent inflammation in mature animals [69].
3.10. Promising and detrimental outcomes of senescence in cells
Aging can be considered a mechanism for coping with any kind of stress to accomplish both crucial and beneficial activities [70]. It also helps in the sustainability of structural configuration and contributes to the recovery of tissue function after injury. For instance, during liver fibrosis, a disorder linked to scraping and scratching of the liver, which reduces its functional ability, aging controls the propagation and expansion of ECM-secreting hepatic stellate cells [71]. This response restricts disease progression following liver impairment. To recover and restore homeostatic equilibrium, the SASP allures natural killer cells from such tissues to abolish aging cells in the diseased part of the liver. The ECM protein CCN1, also called cellular communication network factor 1 (also referred to as CYR61), which mediates the activation of cell aging in hepatic tissues, encourages senescence induction in fibroblasts during wound recovery inside the skin, thereby restraining the scarring of skin tissue [72]. The SASP factor, PDGF-AA, escalates wound closure during recovery. Additionally, the induction of senescence restricts the progression of pancreatic fibrosis. Cell aging is actively involved in tissue repair in several systems. Senescent damage stops fin regeneration in zebrafish after fin amputation [73], whereas cellular senescence is substantially associated with limb regeneration in salamanders. Overall, these observational outcomes indicate that senescence in cells is a mechanism that has been developed to control the tissue impairment response in living organisms and support the restoration and re-architecture to immediately return the tissue to its routine working condition.
Focusing on the detrimental factors of aging reveals that aging is supported by cellular senescence through several pathways [74]. During the aging process, individuals tend to face proinflammatory issues specified by elevated quantities of inflammatory elements, commonly known as inflammation. Inflammation is a detrimental factor in several long-lasting age-related problems, such as cardiovascular issues, certain types of cancer, and neurodegenerative diseases, and may even be linked to early death. Additionally, the existence of inflammatory factors and elements in the blood of old-age people is closely linked to unwanted weight loss, muscle depletion, and general physical lethargy, severe inflammation, and mental stress - declarations of a health issue termed as frailty not long ago [75].
3.11. Senotherapeutic approaches to overcome aging
Senescent cells accumulate several age-linked health issues, as evidenced by the current literature [2]. The finding that the eradication of senescent cells is promising appears to contribute to the absence of persistent poor consequences. Academic and industrial researchers have made it an objective to identify new elements and techniques to diminish senescent cells or their implications in the unavailability of recombinant genetic modifications for human applications. Such therapeutic approaches may be broadly grouped into two different classes: pharmaceutical substances commonly known as senolytic drugs and senomorphic agents that restrain the deadly cell-adventitious complications of senescent cells and involve SASP inhibition factors [76].
The complete removal of aging cells is the core purpose of senomorphic agents. The mode of action mainly relies on alterations to the principal contributing factors to aging, including SASP production and secretion, as well as changes in their capacity to maintain stable growth arrest.This may affect the pro-inflammatory nature of aging cells, which in turn contributes toward delays in aging or aging-related disorders and diseases [77]. Senolytic approaches selectively kill and eliminate senescent cells.
To explore additional senolytic approaches is to explore the intrinsic senolytic system of an organism, called immunosurveillance. In this approach, aging cells are exposed to immunosurveillance through different adaptive and innate immunity mechanisms [78]. The cells used in this approach are macrophages, T cells, and K cells. Based on this approach, it is important to use a combination of mechanisms related to immunosurveillance and the immune system to eliminate aging cells. Aging cells accumulated in diseased and aging tissues lead to a decrease in immunosurveillance. Encouraging the immune system may boost the elimination of aging cells, resulting in an increased lifespan. To explore this connection, an in-depth study was conducted concerning the relationship between aging and NK cells.
Senolytics offer several advantages over senomorphics. For example, senolytics do not require the continuous administration of SASP inhibitors, whereas senomorphics require repeated treatment. In addition, aging-cell removal reduces the possibility of aging, avoiding possible mutations that may promote tumor formation in damaged cells [79]. Furthermore, there is a clear link between the SASP and age-related tissue and organ dysfunction.
The most successful technique for extending health and lifespan is the adoption of caloric restrictions in a variety of species, ranging from yeast to primates. However, the effects of this technique on aging cells must be explored in depth. Recent published literature indicates that cellular aging linked to p16 levels and transcriptional gene expression reports, such as SASP genes, is reduced by caloric restriction techniques [12]. Another study revealed that this technique increased telomere maintenance and decreased DDR levels in mice [80]. Further studies involving postmitotic neurons have shown that this technique reduces the levels of SASP regulators and DDR markers. This connection ultimately reduces the aging load, revealing the connection between aging and caloric restriction.
Over the last few decades, parabiosis has been studied to improve the function of various aging organs, such as the brain, heart, muscles, and liver [81]. The same rejuvenating effects were observed in the brain when younger human plasma was transferred into aging telomerase-deficient mice. Recently it was reported that blood exchange between young and old mice revealed that SASP markers and cellular aging are reduced in old mice exposed to young blood, while increased aging was observed in young mice exposed to old blood [82]. These findings support the idea that blood exchange in humans may be a therapeutic approach for aging-related diseases. Patients with liver damage or acute sepsis have been studied based on this concept.
The following compounds exhibit anti-aging activity by targeting different signaling pathways via different approaches.
3.12. Potential geroprotective and anti-aging compounds
3.12.1. 7-Oxozeanol
(5Z)-7-Oxozeaenol is a naturally occurring anti-protozoan agent originating in fungi. It acts as a strong, irreparable inhibitor of TAK1 and VEGFR2. It has been studied in senescence at early clinical levels in vivo [83]. By activating JNK/p38 MAPKs and NF-KB, TAK1, a member of the mitogen-activated kinase (MAPKKK) family, contributes to pro-inflammatory cellular signaling pathways. Inhibitors of TAK1 catalytic activity has been examined to identify mediators that fight inflammation [83]. A study found that the natural resorcylic lactone 5Z-7-oxozeaenol, which is of fungal origin, is an effective TAK1 inhibitor. The fact that this substance was ineffective at inhibiting the catalytic activity of MEKK1 and ASK1 indicates that 5Z-7-oxozeaenol is a selective TAK1 inhibitor. In cell culture, 5Z-7-oxozeaenol prevents the activation of TAK1, JNK/p38 MAPK, IκB kinases, and NF-κB caused by interleukin-1, which inhibits the synthesis of cyclooxgenase-2. Additionally, 5Z-7-oxozeaenol effectively prevents ear swelling caused by picryl chloride in vivo. Therefore, 5Z-7-oxozeaenol specifically inhibits TAK1 MAPKKK to limit pro-inflammatory signaling [84]. Additionally, 5Z-7-oxozeaenol, which targets the p38 MAPK signaling protein TAK1, reduces the SASP and prevents pro-tumorigenic effects in patients following chemotherapy [85]. The aforementioned results demonstrate the potential advantage of combination anti-cancer therapy using techniques targeting senescence, notwithstanding the possibility of unforeseen adverse side effects from senolytics and senomorphics due to the prevalence of senescent cells and the SASP.
3.12.2. A-1155463
This drug is also effective in treating obesity-associated symptoms. The main mechanism involved in lifespan expansion involves BCL-2 [86]. This is in phase of preclinical studies on senescence, which reveal the senolytic behavior in IMR90 and human umbilical vein endothelial cells (HUVECs) via an in vitro assay. However, in vivo studies are required to evaluate their senolytic activities.
3.12.3. ABT-263
ABT-263, commonly known as navitoclax, is an investigational anti-cancer medication that acts in a manner identical to obatoclax, while inhibiting BCL-2, BCL-XL, and BCL-W, which are pro-survival proteins. In vivo studies indicated the potential capability of ABT-263 in the epidermis to remove aging cells and increase hair follicle stem cell proliferation [87]. However, the main side effect of this drug is severe thrombocytopenia. Recent studies have demonstrated its senolytic effect on aging fibroblasts and found that it decreases photoaging-related secretory phenotype factors, such as SCF, CXCL12, CCL7, CCL5, and IL-6 stimulated MMP-1 in both in vivo and in vitro animal models [88]. Clinical evidence indicates that in patients with osteoarthritis, ABT-263 helps clear senescent cells, which, in turn, strengthens the biological functions of synovial mesenchymal stem cells. However, further SASPs interlinked with UV-induced senescent fibroblasts may be involved and need to be evaluated.
3.12.4. Anakinra
Anakinra is a commercially available biopharmaceutical drug approved for arthritis patients [89]. Anakinra targets the interleukin-1R, a receptor that binds to interleukin 1. There are two types of IL-1R. Type 1 oversees transmission of the inflammatory effects of IL-1, while another type acts as a suppressor of IL-1 [90]. A growing number of senolytic compounds that stimulate apoptosis in cancer cells with senescence have been explored, such as dasatinib and quercetin which treat clinical issues in pulmonary fibrosis, in aging and as well as after radiotherapy, while others include BCL-2 family inhibitors such as novitoclax, HSP90 inhibitors, as well as antibodies directed at SASP-associated elements such anakinra (anti-IL-1β) and infliximab (TNF-α antibody) [91]. Some SASP component-targeting drugs have already been approved for clinical use and may be candidates for medication repositioning. Currently, rheumatoid arthritis is treated with anakinra, an anti-IL-6 receptor antibody, tocilizumab, and tumor necrosis factor inhibitors, such as etanercept and infliximab [92]. These medications may be helpful for specifically suppressing harmful SASP factors because the IL-1 receptor is a major inducer of many other SASP factors and because IL-6 and IL-8 appear to propagate senescence as a common component of the SASP [93,94]. However, because SASP also has certain positive effects, such as wound healing, tissue regeneration, and immune surveillance, and because the majority of SASP components play roles unrelated to SASP, it is probable that medications that target SASP may have unintended side effects.
3.12.5. Azithromycin
Azithromycin is an anti-bacterial drug used to treat several bacterial diseases such as ear infections, throat problems, and diarrhea. It targets senescent human fibroblasts and weakly induces autophagy to drive cell death [95]. Another study reported that it functions as an anti-inflammatory drug that suppresses SASP mediators, including IL-6 and IL-1b [96]. It exhibits high senolytic activity and has a long half-life. However, further studies are needed to determine the molecular mechanisms underlying DNA damage in an aging model.
3.12.6. Canakinumab
Canakinumab, commercially known as Ilaris, is used to treat systemic juvenile idiopathic arthritis [97]. Canakinumab is a monoclonal antibody that efficiently targets interleukin-1 beta [98]. It has been approved for the treatment of cryopyrin-related periodic syndromes. Blocking pro-inflammation pathways, such as NF-Kβ, IL-6, and p38 MAPK signaling, might be used to disrupt senescence in one potential strategy, hence limiting the formation of SASP [99]. Since IL-6 is the "central hub" of variables contributing to localized inflammatory threats, targeting SASP makes sense. In the CANTOS trial, treatment with an IL-1 mAb (canakinumab) decreased the incidence of severe hostile cardiovascular problems by up to 30% in the chronic kidney disease subgroup [100]. While SASP inhibitory elements can lessen the overall implications of senescent cell-induced inflammation with no interference involving anti-cancer pathways, this arbitration may also work through drug-specific procedures that may negatively affect the ability of the immune network to excrete dead cells, raising the possibility of an uncontrolled buildup of senescent cells [101]. Therefore, we must improve our understanding of how senescent cell accumulation, an unintended consequence of SASP therapies, can be prevented.
3.12.7. Cortisol
Cortisol, a glucocorticoid hormone, is a steroid that can be used as a medication under the name hydrocortisone. It elevates blood sugar levels via gluconeogenesis to suppress immunity [102]. In cells, it targets the secretion of interleukin 6 [103], a proinflammatory agent encoded by the IL-6 gene [104].
The effects of corticosterone and cortisol are inhibited by the glucocorticoid antagonist RU-486 [105], which challenges their binding to the glucocorticoid receptors. Additionally, the incorporation of recombinant IL-1 into cells reverses the inhibitory effects of corticosterone and cortisol on NF-Kβ activity and, as a result, IL-6 production, indicating that IL-1 is a crucial target of glucocorticoid activity in senescent cells. Senescence growth arrest cannot be overcome by inhibiting IL-6 and IL-8 secretion caused by glucocorticoids. These outcomes support those of previous studies [106]. Two major elements, X-ray irradiation and RAS expression, cause persistent DNA damage, particularly DNA segments with chromatin alterations reinforcing senescence (DNA-SCARS), which is necessary to sustain the DDR signaling that keeps senescence growth arrest in place [107]. Another study demonstrated how glucocorticoids, working through the GR, hinder the creation of this positive feedback loop by inhibiting IL-1α production and lowering levels of a number of SASP components [103]. However, once established, glucocorticoids do not appear to affect the feedback loop. Consequently, the transcriptional environments that promote the creation and maintenance of the SASP may differ. Glucocorticoids are notable for their capacity to inhibit certain, but not all, contributing elements of the SASP. We cannot rule out the possibility that some of the medications that trickled from the washed cells into the specialized media have independent implications on the invasive ability of cancer cells, despite the fact that the glucocorticoid-treated cells were washed before the collection of specialized media. However, in earlier studies, IL-6 and IL-8 were found to be crucial regulators of the effects of specialized senescent media on the phenotypes of cancer cells [103], and both of these SASP factors are inhibited by glucocorticoids. Corticosterone and cortisol may be examples of medications that can be therapeutically beneficial for circumstances in which the SASP is believed to be damaging, given that the SASP can have positive or detrimental effects depending on the physiological environment. For instance, DNA-damaging radio- and chemotherapy can cause SASP in vivo [93], which may have harmful systemic consequences. They can also result in tumor cells that are not completely destroyed by anti-cancer chemotherapy. Although corticosterone and cortisol individually have negative complications of their own [108], using them concurrently with or soon after radio- or chemotherapy may help mitigate some of these negative systemic effects and even raise the likelihood that patients will survive their treatment without developing cancer, while not interfering with other crucial functions such as tissue repair.
3.12.8. Dasatinib and quercetin
Dasatinib is a targeted therapeutic agent used to treat certain leukemias. Long-term treatment with a combination of dasatinib and quercetin in rats improves age-dependent intervertebral disc degeneration, reduces systemic inflammation, and improves physical condition. Targeted pathways include a significant reduction in aging markers, such as SASP molecules, IL-6, MMP13, p19ARF, and p16INK4a [109]. However, pre-clinical studies should be conducted using larger animal models. Quercetin targets the p13K pathway during aging [110].
3.12.9. EF24
EF24 is a synthetic analog of curcumin, a phytochemical derived from turmeric. It has several biological effects, including anti-oxidant, anti-bacterial, anti-inflammation, and anti-cancer effects. EF24 is also reported to be efficient in decreasing human breast tumors in a mouse model [111]. Enhanced proteasomal degradation of anti-apoptotic BCL-2 proteins expressed by senescent cells displays senolytic efficacy against senescent HUVECs [112]. EF24 is presently used in preclinical models of cellular senescence in the laboratory. The main mechanism of action of this drug for life span extension is by targeting BCL-2, which is an anti-apoptotic protein. It also inhibits Bcl-xL expression in senescent cells. Interestingly, in combination with ABT263, an anti-cancer or Bcl-2 inhibitor, it has been found to be a more effective and safer treatment for age-related diseases [113].
3.12.10. Etanercept
Etanercept is a therapeutic biological product used to treat autoimmune diseases through intervention with TNF, an inflammation-inducing cytokine. It acts as an inhibitor of TNF [114], was accepted for medical use in the United States towards the end of 1998, and is sold under the brand name Enbrel. Targeting individual SASP components that contribute to immune suppression and tumor promotion is a different senomorphic strategy. Various cancers and inflammatory conditions have extensively used monoclonal antibodies and some cytokine signaling-targeting medications, which are also well known in the clinical development process [115]. This includes medications that target TNFα in case of Etanercept and Infliximab, and IL-6/IL-6R by Tocilizumab and Siltuximab [115]. Although simple targeting of IL-6/IL-6R is not sufficient to show therapeutic efficacy in clinical studies for solid tumor cancers, whether it is used individually or in conjunction with chemotherapy [116].
Inhibitors of additional SASP-regulated immune restriction mediators, such as NOTCH and TGFβ (transforming growth factor beta) contenders currently undergoing clinical trials, as well as STING (stimulator of interferon genes) agonists, which are currently being extensively studied [117] in the context of tumor immunotherapy [118] for their capacity to increase anti-tumor resilience proceeding therapy induced senescence, will be tested in the future. Senomorphic-based strategies still lack specificity and centrality, and more detailed research is required to elaborate how their applications in various cancer contexts may affect the SASP background in ways that eventually initiate tumor elimination or development. The senomorphic-associated approach may be innocuous compared to senolytics because senescent and adjacent regular cells are not damaged.
3.12.11. Fisetin
Fisetin belongs to the polyphenol–flavonoid group. It is derived from several plants including vegetables and fruits [119]. Fisetin is known for its anti-cancer effects both in vivo and in vitro, and mainly inhibits the PI3K/AKT/mTOR pathway [120]. It also reduces senescence in human and murine adipose tissues. In vitro analysis has shown that target pathways include NF-KB, IL-6, and TNFα [121]. Another study revealed that it acts as a senolytic agent to increase lifespan. Preclinical studies are underway to investigate these issues in depth.
3.12.12. KU-60019
KU-60019 is an analog of KU-55933 and is ten times more efficient than KU-55933. It inhibits ataxia-telangiectasia mutated (ATM) kinase by specifically targeting it during the cell cycle [122]. Cell cycle arrest and senescence are controlled by the serine/threonine protein kinase ATM kinase, which is activated by DNA double-strand breaks [123,124]. In a chemical screening, the ATM kinase inhibitor KU-60019 was discovered as a treatment for senescence. More precisely, administration of KU-60019 to replicative and stress-induced senescent human dermal fibroblasts (HDFs) leads to higher cumulative population doublings (CPDs), decreased senescence associated beta galactosidase activity, p16 protein expression, reactive oxygen species levels, and abnormal nuclear morphology, and restores the lysosomal/autophagy system, which in turn leads to higher levels of mitophagy and, ultimately, functional recovery of mitochondria and metabolism. Additionally, the administration of KU-60019 to old mice accelerates the healing of cutaneous wounds [125]. This investigation did not reveal the expression of SASP components; however, considering the link between DDR and NF-KB activation [126], it is possible that KU-60019 could operate as a potential senomorphic agent [127]. Accordingly, depending on the stress traits that cause senescence, both ATM activation and inhibition may prevent senescence-associated cellular alterations. In older mice, KU-60019 therapy expedites the healing of cutaneous wounds, and inhibiting ATM activity slows the aging process. Thus, KU-60019 is a viable candidate for treating age-related disorders.
3.12.13. Loperamide
Loperamide is administered to reduce the periodic occurrence of diarrhea. It was initially developed in 1969 and became available in 1976 [128]. It is sold under the brand name Imodium and is commercially available as a generic medication. In cells, it targets heat shock protein 90 (HSP90) to nordihydroguaiaretic acid (NDGA) [129]. Several compounds with senomorphic potential have also been reported. However, the effectiveness of certain compounds is still being investigated, and for other drugs, the targeted route has not been well described. Loperamide and niguldipine, two Ca2+ channel blockers that suppress SASP in primary murine embryonic fibroblasts (MEFs), have been linked to senomorphic activity [129]. Recently, in DNA repair-impaired Ercc1/ mouse embryonic cells, loperamide and NDGA were shown to exhibit senomorphic action [129]. However, as evidently described in the abovementioned sections, cellular senescence is a condition that is also recognized to have a variety of advantageous activities. The function of these activities in senolysis-based therapeutic treatments remains controversial and requires further elucidation. Cumulative evidence strongly suggests that the numerous beneficial and harmful effects of senescence on living creatures are delicately balanced. This factor is crucial for the objective assessment and safe use of any therapeutic intervention based on tiny drug-like compounds that might emerge in the future. The necessity for a deeper understanding of the treatment constraints provided by the innate complexity of aging mechanisms is of the highest significance in this direction.
3.12.14. NDGA
NDGA, generally referred to as nordihydroguaiaretic acid in scientific literature, is a naturally occurring antioxidant found in the creosote bush (Larrea tridentata). Initial reports suggested that NDGA helps to increase the mean life expectancy of rats (AR 1959), D. melanogaster [130], and mosquitos [131]. It generally targets HSP90 in cells [132]. Recently, it was discovered that NDGA, along with loperamide, has senomorphic activity in DNA repair-deficient Ercc1/ murine embryonic fibroblasts [133]. Aspirin, NDGA, and rapamycin have been shown to increase the lifespan in an aging intervention testing program series. These substances also block various inflammatory responses, supporting theories claiming that late-life inflammation is a significant aging mechanism [134]. In a previous study, it was discovered that NDGA lengthened the lives of male but not female UM-HET 3 mice [135]. It is possible that NDGA has positive benefits in both sexes but has negative effects in females that hinder lifespan extension. To ascertain whether NDGA influences the various elements of aging in either or both sexes, further information regarding age-dependent changes in both sexes, particularly those related to general health, is required. More generally, research employing mutant stocks and dietary intervention will complement the work being conducted to define the variables that influence the aging rate in animals and establish links between aging and various illnesses. These medications may help individuals live longer by modulating aging, cancer, or both.
3.12.15. Ouabain
Ouabain, a cardiac glycoside, is a broad-spectrum senolytic agent that triggers the activation of ERK, Akt, p38, and Src in BRAF-senescent cells and prevents cell death [27]. It also inhibits autophagic flux. Using a dual mechanism of action, ouabain, a natural substance from the cardiac glycoside group, has been identified as a senolytic drug that can be used to remove senescent cells and treat cancer [136]. It elevates various pro-apoptotic BCL-2 family proteins in senescent fibroblasts, which triggers apoptosis.
3.12.16. RAD001
RAD001, also known as everolimus and sold under the brand name Afinitor, is an immunosuppressant drug that prevents organ rejection during organ transplantation [137]. Rapamycin and its analogs, such as RAD001, can block mTOR, one of the most important upstream signaling pathways in the processes of cellular senescence and the SASP, and can exhibit a senomorphic function [138]. It functions as an inhibitor of mTOR, similar to sirolimus. The effectiveness of RAD001 as a SASP inhibitor is demonstrated by the fact that RAD001 reduces the activation of mTOR and its direct targets, while the ability of RAD001 to arrest mTOR phosphorylation, cell cycle arrest, and SA—Gal positivity continues, indicating that metabolic processes and cellular senescence are maintained [85].
In many animals, rapalogs, which are mTORC1 (mammalian target of rapamycin complex 1) inhibitors, prolong life and delay age-related traits. However, their molecular pathways are still not completely understood [139]. When 24-month-old rats treated with RAD001 were compared with vehicle-treated rats, the severity of chronic progressive nephropathy lesions was less severe in RAD001-treated kidneys. In addition to highlighting c-Myc regulation as a probable mechanism by which mTORC1 inhibition potentially perturbs age-related phenotypes, this study provides scenarios for the use of mTORC1 inhibitors in the treatment of age-related illnesses [139]. Additionally, to increase the coverage of age-related gene changes that may be counter-regulated by intervention, supplementary research is required to examine other tissues in a similar manner, including immune cells, and to determine whether different pharmacological therapies can be added to a treatment regimen for mTORC1 inhibition.
3.12.17. Rilonacept
Rilonacept, like canakinumab, is a therapeutic drug used to treat cryopyrin-related periodic syndromes, such as familial cold autoinflammatory syndrome and Muckle–Wells syndrome. Rilonacept is a potent inhibitor of both interleukin-1α and β [140,141]. To target cytokine signaling, monoclonal antibodies have been used in synergy with other agents in several cancers and associated inflammation-related settings. This includes agents which target IL-1α/IL-1β/IL1R as in the case of Anakinra, Canakinumab, Rilonacept and TNFα by drugs such as Etanercept, Infliximab [57]. Currently, this could provide an alternative senomorphic strategy while directly targeting particular molecules of the SASP, which guides a compromised immune response and tumor development in aging cells. Owing to its nature of being at an initial stage, more clinical studies are needed before it can be applied in practical real-world situations. Additionally, rilonacept can potently inhibit senescent cells, as it is associated with tumor development, making it a valuable candidate drug for SASP inhibition [142].
3.12.18. Roxithromycin
Roxithromycin, an anti-bacterial drug that belongs to the macrolide class, is used in the treatment of respiratory tract, urinary, and soft tissue infections and selectively destroys aging cells via the induction of apoptosis and SASP factors [95]. It also inhibits TGF-β in in vitro experiments. In a mouse model, it reduces lung injury, fibrosis, and inflammation [96]. Additionally, it attenuates NOX4 expression during lung fibrosis. Collectively, it was found to be a potential anti-aging that should be further tested clinically.
3.12.19. Ruxolitinib
Similar to Jakafi and Jakavi, ruxolitinib is used to treat medium- or high-level myelofibrosis, which adversely affects bone marrow [143]. It is a Janus kinase inhibiting agent with special affinity for JAK1 and JAK2 [144]. After receiving FDA approval, the selective JAK1/2 inhibitor roxolitinib was administered as a therapy for intermediate or high risk patients [145]. When JAK kinase was treated with ruxolitinib, SASP progression was reduced in replicative exhaustion and IR-mediated senescent pre-adipocytes, as well as in senescent HUVECs, indicating that it has senomorphic features, whereas ruxolitinib treatment of old mice substantially increased the ability to control systemic and adipose tissue inflammation and improved physical activity [146]. In contrast, when administered to 22-month-old mice, ruxolitinib treatment resulted in improved adipogenesis, prevented fat loss, decreased lipotoxicity, and increased insulin sensitivity [147]. Finally, senescent cells may be specifically targeted for the delivery of senolytic drugs to reduce their side effects and maximize their benefits. Although numerous senolytic medicines have been studied in vitro and in animal models, clinical trials using focused therapeutic approaches are required to assess their safety and efficacy.
3.12.20. SB203580
SB203580 is a key inhibitor agent of p38 mitogen-activated protein kinase (p38MAPK). Currently, it is observed in the preclinical stages of senescence in scientific laboratories [148]. A wide range of biological stressors, including DNA damage, signal to p38MAPK. Notably, p38MAPK activity in an HCA2 cell model is sufficient to elicit an SASP and that p38MAPK regulates the production of IL-6, IL-8, and GM-CSF through activated NF-KB [148]. Therefore, we investigated whether, and to what extent, NF-KB activity increases during astrocytic senescence. Interestingly, SB203580 dramatically reduced nuclear translocation by inhibiting p38MAPK [149]. In this study, p38MAPK controlled NF-KB expression during senescence. Moreover, it was investigated whether NF-κB is necessary for SASP because p38MAPK controls both SASP and NF-κB activity. Senescent astrocytes were treated with the NF-κB inhibitor BAY 11-7092. Inhibition of NF-κB greatly reduced the release of IL-6 and IL-8. Another report showed that the pyridinyl imidazole p38 MAPK inhibitor SB203580 may target PDK1/PDK2 (3-phosphoinositide-dependent protein kinase), which could at least partially explain the anti-proliferative effect of the medication. The finding that SB203580 can inhibit the PI 3-kinase/PDK1/PKB (protein kinase B) pathway may have significant effects on the interpretation of the results of studies using this drug [150]. The role of SB203580 in the phosphatidylinositol 3-kinase-mediated activation of PKB lends evidence to the alleged function of kinases in cellular survival and proliferation. The discovery that SB203580 specifically targets PDK1 offers a potential mechanism of action [151]. However, as no reports have detected any indication of elevated apoptosis in treated cells, SB203580 appears to have a positive impact on cell cycle progression in the context of IL-2 signaling. It would be intriguing to determine whether SB203580 affects bad phosphorylation, which is regulated by PKB [152]. Lastly, all of these observations point to a function for these kinases and their downstream effectors in retinoblastoma (Rb) regulation, but do not prove that PDK1 or PKB activity is necessary for Rb phosphorylation. Additionally, these findings do not rule out the role of additional SB203580 targets in driving Rb phosphorylation and T cell proliferation.
3.12.21. Simvastatin
Simvastatin is a fungus-derived drug that helps decrease lipid levels in cells [153]. It is administered in combination with exercise, diet, and weight loss to regulate lipid levels. It targets IL-6, IL-8, and MCP1 and is under experimental observation for cellular senescence in vitro at preclinical stages [154]. Simvastatin lowers the SASP by preventing protein prenylation, which in turn inhibits Rac1 and Cdc42 activities. Members of the Rho GTPase family, Rac1 and Cdc42, control actin dynamics and cell division [155]. However, the role of Rac1 in cellular senescence has been the subject of contradictory findings. According to one study, senescent phenotypes require CDK5 to suppress Rac1 activity [156], while Rac1 depletion prevents CCN1-induced senescence, potentially by lowering ROS levels [157]. However, according to another report, constitutive Rac1 activation accelerates aging and mitochondrial oxidative stress [158]. Another study revealed that cellular senescence is caused by either an increase or decrease in Rac1 activity [159]. Therefore, it is probable that Rac1 activity must be maintained within a specific range, and that aberrant Rac1 activity — either an increase in function or a loss of it — can lead to cellular senescence. The authors [154] proved that simvastatin inhibits Rac1 and Cdc42 activation, which in turn inhibits SASP and its cancer-promoting actions, and that SASP induces MEK-ERK1/2 pathway activation, which promotes breast cancer cell proliferation, and that simvastatin inhibits SASP-mediated activation in breast cancer cells. Their data also suggested that breast cancer cells develop endocrine resistance to SASP, which can be reversed by simvastatin.
3.12.22. Siltuximab
Siltuximab (INN; commercial name Sylvant, also labelled CNTO 328) is a globulin fusion protein. It attaches to IL-6 and is being studied for the recovery of neoplastic issues [160]. Siltuximab is also approved for multicentric Castleman disease (MCA) [161]. Antibodies against SASP factors may also have the ability to reduce harmful senescence-related effects. Several oncological settings are testing siltuximab, a neutralizing anti-IL-6 monoclonal antibody authorized to treat MCA [162]. By blocking IL-6 or its receptor, approved medications such as siltuximab may address the issue of senescence. These medications have already been approved to treat cytokine release syndrome (CRS), a condition that can be fatal, as well as rheumatic diseases and cancer triggered by IL-6 [163]. However, beyond their general anti-inflammatory potential, their specific anticancer efficacy as senescence-dependent SASP-suppressing medicines requires in-depth follow-up investigations in suitable model systems and clinical trials.
3.12.23. Tocilizumab
Tocilizumab is an immunosuppressive medication that treats autoimmune diseases and is available under the name of Actemra. It is primarily a humanized monoclonal immunoglobulin that acts against the interleukin-6 receptor (IL-6R) like siltuximab [164]. Several therapeutic medications currently in clinical stages for autoimmune and autoinflammatory diseases, including rheumatoid arthritis, specifically target SASP elements or their receptors [165]. Most pertinent research in this area targets the IL-1 receptor (anakinra), IL-6 receptor (tocilizumab, siltuximab), IL-6 (sirukumab), and TNF-α (adalimumab, etanercept, and infliximab), which have a great potential to be used as precision senostatic agents to prevent the negative effects of the SASP [165,166]. Senolytics can also be used in various forms. Some of these substances are pro-apoptotic medicines that have been used in the treatment of cancer. Apoptosis induces cell death and aids in senescence reversal Senomorphics such as lutikizumab and canakinumab target the IL-1 receptor, tocilizumab targets the IL-6 receptor, etanercept targets TNF, and CL82198 targets MMP13 [167]. In conclusion, the data from these investigations indicate the possibility that senostatic drugs can enhance cancer treatment by regulating or inhibiting pro-inflammatory SASP elements. An effective senostatic intervention may be severely constrained by the context-dependent intricacy and intensity of the SASP network as well as the non-senescence-related functions of the SASP variables. Therefore, care must be taken when using senostatic drugs, and future research should concentrate on resolving these issues.
In summary, agents that restrict the destruction of senescent cells or reduce the harmful complications of the SASP have a variety of benefits that support their use in the treatment of chronic illnesses and age-related conditions. Unfortunately, numerous obstacles, including improved selectivity and decreased off-target effects, still need to be addressed before a drug development strategy can be successful.
4. Discussion
The steady deterioration of the functions of various cells and organs leads to early aging. Reports from the World Health Organization have shown that by 2030, one out of every six people will reach 60 years or older. Cellular aging is a versatile process initiated in response to various intrinsic and extrinsic triggers [168]. These responses are interconnected and involve several simple and complex processes. Reversing aging is impossible; however, the aging process can be delayed or corrected by adopting certain measures. For example, a better understanding of the aging signaling pathway or the identification of aging triggers and how aging can be delayed [39]. The main purpose of this paper is to provide a clear and in-depth review of the possible signaling pathways involved in aging to identify aging targets. Once ageing targets have been explored or studied in depth, researchers can readily develop anti-aging measures.
From the above studies, we now know the most important target, which is the correction of DNA damage in aged cells or modification of the programming of aged cells, such as SASP regulation and modification. Based on this concept, we sought to explore possible means and anti-aging compounds that might overcome the aging process to live a long time along with a healthy lifestyle.
By assessing various recent studies regarding aging signaling networks via this review, we have certain suggestions and opinions for other researchers and scientists, and we hope that our opinions and conclusions will be very helpful for identifying and exploring potential therapeutic anti-aging approaches.
DDR can be controlled or overcome by regulating p53, p16, and ARF [17]. By monitoring these factors via control of corrupted information, transmission of corrupted information into next-generation cells could be avoided, thus a delay in aging can be achieved. This ultimately leads to the production of newly generated healthy cells without any dysfunction and the production of young cells. Therefore, p53, p16, and ARF may act as targeted factors in the development of anti-aging approaches.
To find anti-aging therapeutic approaches at the cellular level by studying signaling networks deeply, DDR can be controlled by modulating telomere shortening via telomerase activation [169]. Therefore, identifying potential agents that act on telomeres shortening, novel anti-aging approaches can be introduced [170]. Introducing agents that can boost NAD+ levels may boost surtuin activity, which functions as an anti-aging gene and novel anti-aging therapy. As an oncogene, PTEN loss induces aging; therefore, by introducing approaches to reduce the loss of, or damage to, PTEN, aging can be delayed or controlled. Another approach to prevent or delay aging is the identification of potential histone deacetylase inhibitors that can act as senolytic agents [171]. The current review reveals that no single compound has been detected targeting the DDR, suggesting that a massive field of aging research targeting the DDR signaling pathway for identifying anti-aging therapeutics in nature needs to be explored.
Activation of the SASP is a self-motivated process that causes aging; therefore, research can be moved towards the exploration of SASP inhibitors [172]. DDR activation is a rapid response, whereas SASP establishment is gradual; therefore, continuous treatment is required to control SASP activation. In addition, SASP and DDR signaling are associated with the promotion of aging. Studies have also indicated that GATA4 interacts with DDR signaling and is involved in aging pathology [17]. Targeting GATA4 has been predicted to deliver novel approaches against aging. This review also reveals that exploring JAK/STAT inhibitors could provide new insights into aging targets. Inhibition of BRD4 reduces SASP activation and suppresses senescence. HMGB1 and HMGB2 are key regulators of SASP expression [173]. Due to the complex nature of SASP modulation, irregular stimulation of the cGAS-STING mechanism has been found to be interconnected during aging.
This review highlights the importance of therapeutic agents from natural and synthetic sources in lifespan extension, as shown in the figures targeting SASP and DDR. Combinations of drugs and use alone may bring some potential effects in delaying the ageing process via different signaling pathways targeting various pharmacological targets. For example, A-1331852, A1155463, and EF24 regulate SASP signaling by targeting BCL-2, BCL-X, BCC-W, and quabain. Azithromycin, SB203580, simvastatin, cortisol, canakinumab, siltuximab, ankinra, rilonacept, and tocilizumab act on SASP signaling pathways via targeting IL-1α,IL-6, IL-8, and IL-1β. RD001 acts on mTOR and mTORCI in the SASP signaling pathway. Another interesting factor highlighted in this review is that no structure-activity relationship has been studied against the aforementioned compounds for anti-aging drugs. Notably, once a potential pharmacological compound is identified against any pathological conditions, scientists always try to find their derivatives either from natural resources or via synthetic pathways. For example, EF24 is a novel derivative of curcumin with significant identified roles such as anti-tumor, anti-bacterial, and anti-inflammatory effects [174]. Thus, curcumin and its derivatives may have greater therapeutic potential. A-1331852 and A1155463, BCL-XL inhibitors, indicate that other already identified BCL-XL inhibitors can be explored and investigated to identify hidden potential compounds for anti-aging. In addition, already identified compounds provide clues and direct methods for searching for new compounds sourced from natural resources or synthetic pathways based on structure-activity relationships.
One major drawback in the development of anti-aging drugs [175] is the complex and continuous signaling pathways in ageing, which cannot be arrested. Also, ageing is not a disease that needs to be cured, but a long, continuous, natural, and irreversible process that is difficult to measure and identify as an anti-aging target. Experimental validation is a major challenge in the development of anti-aging drugs. Negative thoughts and emotions during stress can lead to poor health with rapid ageing that ultimately represent a considerable challenge to developing anti-aging measures. Ethical concerns that could arise from cultural and economic perspectives are another obstacle to the development of anti-aging medicines [175]. To overcome these challenges, in-depth studies are required to identify potential targets for delaying the aging process. As the target is approached, exploring new therapies becomes easy. To eliminate negative thoughts and emotions, one should have a healthy diet and keep engaged in precious moments with family members, which can bring about a happy change in life. Strong social support and good friends’ networks can relieve stress and ultimately delay aging. Economic impacts and social changes bring about drastic changes in delayed aging. Better socioeconomic circumstances delay the aging process, as better living conditions improve health standards and ultimately lead to less spending on healthcare expenditure towards the terminal stages of an individual’s life. Therefore, it is recommended to focus on a healthy lifestyle without stressing on reducing the financial burden during the peak years of life.
The evidence from the above literature review revealed that almost all studies identified were conducted to find anti-aging targets and their detailed signaling networks, such as the DDR and SASP signaling networks, as shown in Figure 3. However, there is also a need to explore bioactive agents from different resources, such as natural and synthetic products, and already recognized potential drugs for different diseases that need to be tested for anti-aging purposes. Recognizing that any potential anti-aging compound will not bring happiness to the world belonging to all age groups, but also boosts the pharmaceutical industries, biotech companies, and investors in terms of revenue. Aging is a very complex mechanism and identifying aging models that match human models is quite challenging for researchers. Therefore, future studies should focus on the identification of anti-aging agents. This highlights the importance of our literature review, as it provides updated information and analysis surrounding the development of anti-aging products.
5. Conclusions
In conclusion, the information, analysis, and suggestions in this article may serve as a valuable resource for the rapid development of anti-aging therapeutic strategies that target the DDR and SASP signaling networks. Incorporating the already identified anti-aging agents and finding their derivatives from different sources can aid and unlock potential compounds in the fight against aging, ultimately improving the quality of life, bringing happiness, and adding more years to spend in this world.
Author Contributions
A.P. designed and directed the project. J.N helped in writing the manuscript. K.S.Y supported the project with A.P
Funding
Currently, this study has no financial support.
Conflicts of Interest
Authors declares no conflict of interest.
References
- Liang, W.; Guan, H. Investigation and analysis on travel and health of the elderly in the old residential areas in Beijing. IRSPSD International; 2023, 11, 85–103. [CrossRef]
- López-Otín, C.; Pietrocola, F.; Roiz-Valle, D.; Galluzzi, L.; Kroemer, G. Meta-hallmarks of aging and cancer. Cell Metab 2023, 35, 12–35. [Google Scholar] [CrossRef] [PubMed]
- Cano, M.; Yeung, Y.T.; Muñoz, M.F.; Ayala, A.; Guerrero-Castilla, A.; Argüelles, S. Basic pathways and targets for antiaging intervention. Anti-Aging Pharmacology; Elsevier; 2023. pp. 13–40.
- Pezone, A.; Olivieri, F.; Napoli, M.V.; Procopio, A.; Avvedimento, E.V.; Gabrielli, A. Inflammation and DNA damage: cause, effect or both. Nat Rev Rheumatol 2023, 19, 200–211. [Google Scholar] [CrossRef]
- Liu, R.M. Aging, cellular senescence, and Alzheimer’s disease. Int J Mol Sci 2022, 23, 23(4):1989. [CrossRef]
- Vaddavalli, P.L.; Schumacher, B. The p53 network: cellular and systemic DNA damage responses in cancer and aging. Trends Genet 2022, 38, 598–612. [Google Scholar] [CrossRef] [PubMed]
- Mi, L.; Hu, J.; Li, N.; Gao, J.; Huo, R.; Peng, X.; Zhang, N.; Liu, Y.; Zhao, H.; Liu, R.; et al. The mechanism of stem cell aging. Stem Cell Rev Rep 2022, 18, 1281–1293. [Google Scholar] [CrossRef]
- Yoon, Y.S.; You, J.S.; Kim, T.K.; Ahn, W.J.; Kim, M.J.; Son, K.H.; Ricarte, D.; Ortiz, D.; Lee, S.J.; Lee, H.J. Senescence and impaired DNA damage responses in alpha-synucleinopathy models. Exp Mol Med 2022, 54, 115–128. [Google Scholar] [CrossRef]
- Wang, F.; Jin, S.; Mayca Pozo, F.M.; Tian, D.; Tang, X.; Dai, Y.; Yao, X.; Tang, J.; Zhang, Y. Chemical screen identifies shikonin as a broad DNA damage response inhibitor that enhances chemotherapy through inhibiting ATM and ATR. Acta Pharm Sin B 2022, 12, 1339–1350. [Google Scholar] [CrossRef] [PubMed]
- González-Bermúdez, L.; Genescà, A.; Terradas, M.; Martín, M. Role of H4K16 acetylation in 53BP1 recruitment to double-strand break sites in in vitro aged cells. Biogerontology 2022, 23, 499–514. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.H.; Ho, T.L.F.; Hariharan, A.; Goh, H.C.; Wong, Y.L.; Verkaik, N.S.; Lee, M.Y.; Tam, W.L.; van Gent, D.C.; Venkitaraman, A.R.; et al. Rapid recruitment of p53 to DNA damage sites directs DNA repair choice and integrity. Proc Natl Acad Sci U S A 2022, 119, e2113233119. [Google Scholar] [CrossRef]
- Duan, J.L.; Ruan, B.; Song, P.; Fang, Z.Q.; Yue, Z.S.; Liu, J.J.; Dou, G.R.; Han, H.; Wang, L. Shear stress–induced cellular senescence blunts liver regeneration through Notch–sirtuin 1–P21/P16 axis. Hepatology 2022, 75, 584–599. [Google Scholar] [CrossRef]
- Lee, S.J.; Lee, D.Y.; O’Connell, J.F.; Egan, J.M.; Kim, Y. Black ginseng ameliorates cellular senescence via p53-p21/p16 pathway in aged mice. Biology 2022, 11, 1108. [Google Scholar] [CrossRef]
- Soto-Palma, C.; Niedernhofer, L.J.; Faulk, C.D.; Dong, X. Epigenetics, DNA damage, and aging. J Clin Invest 2022, 132, e158446. [Google Scholar] [CrossRef] [PubMed]
- Moser, R.; Gurley, K.E.; Nikolova, O.; Qin, G.; Joshi, R.; Mendez, E.; Shmulevich, I.; Ashley, A.; Grandori, C.; Kemp, C.J. Synthetic lethal kinases in Ras/p53 mutant squamous cell carcinoma. Oncogene 2022, 41, 3355–3369. [Google Scholar] [CrossRef] [PubMed]
- Bell, H.N.; Rebernick, R.J.; Goyert, J.; Singhal, R.; Kuljanin, M.; Kerk, S.A.; Huang, W.; Das, N.K.; Andren, A.; Solanki, S.; et al. Reuterin in the healthy gut microbiome suppresses colorectal cancer growth through altering redox balance. Cancer Cell 2022, 40, 185–200.e6. [Google Scholar] [CrossRef] [PubMed]
- Di Micco, R.; Krizhanovsky, V.; Baker, D.; d’Adda di Fagagna, F. Cellular senescence in ageing: from mechanisms to therapeutic opportunities. Nat Rev Mol Cell Biol 2021, 22, 75–95. [Google Scholar] [CrossRef] [PubMed]
- Rossiello, F.; Jurk, D.; Passos, J.F.; d’Adda di Fagagna, F. Telomere dysfunction in ageing and age-related diseases. Nat Cell Biol 2022, 24, 135–147. [Google Scholar] [CrossRef]
- Mao, J.; Zhang, Q.; Wang, Y.; Zhuang, Y.; Xu, L.; Ma, X.; Guan, D.; Zhou, J.; Liu, J.; Wu, X.; et al. Tert activates endogenous retroviruses to promote an immunosuppressive tumour microenvironment. EMBO Rep 2022, 23, e52984. [Google Scholar] [CrossRef]
- Makeeva, V.S. Ionizing radiation effects on telomeres. Biol Bull 2022, 49, 2257–2265. [Google Scholar] [CrossRef]
- Huang, W.; Hickson, L.J.; Eirin, A.; Kirkland, J.L.; Lerman, L.O. Cellular senescence: the good, the bad and the unknown. Nat Rev Nephrol 2022, 18, 611–627. [Google Scholar] [CrossRef]
- Francia, S.; Capozzo, I.; Modafferi, S.; Iannelli, F.; Gioia, U., di Fagagna FdA. DROSHA and DICER RNA products control BMI1-dependent transcriptional repression at DNA damage sites. Europe PMC. 2022.
- Aguado, J.; d’Adda di Fagagna, F.; Wolvetang, E. Telomere transcription in ageing. Ageing Res Rev 2020, 62, 101115. [Google Scholar] [CrossRef]
- Vaena, S.; Chakraborty, P.; Lee, H.G.; Janneh, A.H.; Kassir, M.F.; Beeson, G.; Hedley, Z.; Yalcinkaya, A.; Sofi, M.H.; Li, H.; et al. Aging-dependent mitochondrial dysfunction mediated by ceramide signaling inhibits antitumor T cell response. Cell Rep 2021, 35, 109076. [Google Scholar] [CrossRef]
- Di Emidio, G.; Falone, S.; Artini, P.G.; Amicarelli, F.; D’Alessandro, A.M.; Tatone, C. Mitochondrial sirtuins in reproduction. Antioxidants (Basel) 2021, 10, 1047. [Google Scholar] [CrossRef]
- Covarrubias, A.J.; Perrone, R.; Grozio, A.; Verdin, E. NAD+ metabolism and its roles in cellular processes during ageing. Nat Rev Mol Cell Biol 2021, 22, 119–141. [Google Scholar] [CrossRef] [PubMed]
- l’Hôte v. Senolytic drug discovery and mechanisms of action in BRAF-V600E oncogene-induced senescence: Université Paris-Saclay; 2022.
- Helbling-Leclerc, A.; Garcin, C.; Rosselli, F. Beyond DNA repair and chromosome instability-Fanconi anaemia as a cellular senescence-associated syndrome. Cell Death Differ 2021, 28, 1159–1173. [Google Scholar] [CrossRef] [PubMed]
- Krokidis, M.G.; Prasinou, P.; Efthimiadou, E.K.; Boari, A.; Ferreri, C.; Chatgilialoglu, C. Effects of aging and disease conditions in brain of tumor-bearing mice: evaluation of purine DNA damages and fatty acid pool changes. Biomolecules 2022, 12, 1075. [Google Scholar] [CrossRef]
- Pouikli, A.; Parekh, S.; Maleszewska, M.; Nikopoulou, C.; Baghdadi, M.; Tripodi, I.; Folz-Donahue, K.; Hinze, Y.; Mesaros, A.; Hoey, D.; et al. Chromatin remodeling due to degradation of citrate carrier impairs osteogenesis of aged mesenchymal stem cells. Nat Aging 2021, 1, 810–825. [Google Scholar] [CrossRef] [PubMed]
- Castro-Obregón, S. Lamin B receptor: role on chromatin structure, cellular senescence and possibly aging. Biochem J 2020, 477, 2715–2720. [Google Scholar] [CrossRef]
- Lund, P.J.; Lopes, M.; Sidoli, S.; Coradin, M.; Vitorino, F.N.L.; Da Cunha, J.P.C.; Garcia, B.A. FGF-2 induces a failure of cell cycle progression in cells harboring amplified K-Ras, revealing new insights into oncogene-induced senescence. Mol Omics 2021, 17, 725–739. [Google Scholar] [CrossRef]
- Ashrafian, S.; Zarrineh, M.; Jensen, P.; Nawrocki, A.; Rezadoost, H.; Ansari, A.M.; Farahmand, L.; Ghassempour, A.; Larsen, M.R. Quantitative phosphoproteomics and Acetylomics of safranal anticancer effects in triple-negative breast cancer cells. J Proteome Res 2022, 21, 2566–2585. [Google Scholar] [CrossRef]
- Wu, Y.; Wu, Y.; Yang, Y.; Yu, J.; Wu, J.; Liao, Z.; Guo, A.; Sun, Y.; Zhao, Y.; Chen, J.; et al. Lysyl oxidase-like 2 inhibitor rescues D-galactose-induced skeletal muscle fibrosis. Aging Cell 2022, 21, e13659. [Google Scholar] [CrossRef]
- Zhang, Y.; Amaral, M.L.; Zhu, C.; Grieco, S.F.; Hou, X.; Lin, L.; Buchanan, J.; Tong, L.; Preissl, S.; Xu, X.; et al. Single-cell epigenome analysis reveals age-associated decay of heterochromatin domains in excitatory neurons in the mouse brain. Cell Res 2022, 32, 1008–1021. [Google Scholar] [CrossRef]
- Zhang, N.; Shang, M.; Li, H.; Wu, L.; Dong, M.; Huang, B.; Lu, J.; Zhang, Y. Dual inhibition of H3K9me2 and H3K27me3 promotes tumor cell senescence without triggering the secretion of SASP. Int J Mol Sci 2022, 23, 3911. [Google Scholar] [CrossRef] [PubMed]
- Duan, D.; Shang, M.; Han, Y.; Liu, J.; Liu, J.; Kong, S.H.; Hou, J.; Huang, B.; Lu, J.; Zhang, Y. EZH2–CCF–cGAS axis promotes breast cancer metastasis. Int J Mol Sci 2022, 23, 1788. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Allis, C.D.; Wang, G.G. The language of chromatin modification in human cancers. Nat Rev Cancer 2021, 21, 413–430. [Google Scholar] [CrossRef]
- Zhu, X.; Chen, Z.; Shen, W.; Huang, G.; Sedivy, J.M.; Wang, H.; Ju, Z. Inflammation, epigenetics, and metabolism converge to cell senescence and ageing: the regulation and intervention. Signal Transduct Target Ther 2021, 6, 245. [Google Scholar] [CrossRef] [PubMed]
- Endicott, J.L.; Nolte, P.A.; Shen, H.; Laird, P.W. Cell division drives DNA methylation loss in late-replicating domains in primary human cells. Nat Commun 2022, 13, 6659. [Google Scholar] [CrossRef]
- Yu, Y.C.-Y.; Hui, T.Z.; Kao, T.H.; Liao, H.F.; Yang, C.Y.; Hou, C.C.; Hsieh, H.T.; Chang, J.Y.; Tsai, Y.T.; Pinskaya, M.; et al. Transient DNMT3L expression reinforces chromatin surveillance to halt senescence progression in mouse embryonic fibroblast. Front Cell Dev Biol 2020, 8, 103. [Google Scholar] [CrossRef]
- Chalmers, T.J.; Wu, L.E. Transposable elements cross kingdom boundaries and contribute to inflammation and ageing: somatic acquisition of foreign transposable elements as a catalyst of genome instability, epigenetic dysregulation, inflammation, senescence, and ageing. BioEssays 2020, 42, e1900197. [Google Scholar] [CrossRef]
- Metaxakis, A.; Gkikas, I.; Tavernarakis, N. The epigenetics of aging. Aging; Elsevier 2023, 333–358.
- Liu, H.; Zhao, H.; Sun, Y., Eds. Tumor microenvironment and cellular senescence: understanding therapeutic resistance and harnessing strategies. Semin Cancer Biol; Elsevier 2022, 86, 769–781. [CrossRef]
- Van Petten de Vasconcelos Azevedo, F.; Lopes, D.S.; Zóia, M.A.P.; Correia, L.I.V.; Saito, N.; Fonseca, B.B.; Polloni, L.; Teixeira, S.C.; Goulart, L.R.; de Melo Rodrigues Ávila, V. A New Approach to Inhibiting Triple-Negative Breast Cancer: in vitro, ex vivo and in vivo antiangiogenic Effect of BthTx-II, a PLA2-Asp-49 from Bothrops jararacussu Venom. Biomolecules 2022, 12, 258. [Google Scholar] [CrossRef]
- Tan, H.; Xu, J.; Liu, Y. Ageing, cellular senescence and chronic kidney disease: experimental evidence. Curr Opin Nephrol Hypertens 2022, 31, 235–243. [Google Scholar] [CrossRef]
- Liang, L.; Chai, Y.; Chai, F.; Liu, H.; Ma, N.; Zhang, H.; Zhang, S.; Nong, L.; Li, T.; Zhang, B. Expression of SASP, DNA damage response, and cell proliferation factors in early gastric neoplastic lesions: correlations and clinical significance. Pathol Oncol Res 2022, 28, 1610401. [Google Scholar] [CrossRef]
- Turley, J. Chitin-Derived STING Activators as Adjuvants and Therapeutics; Trinity College Dublin, 2021.
- Salech, F.; SanMartín, C.D.; Concha-Cerda, J.; Romero-Hernández, E.; Ponce, D.P.; Liabeuf, G.; Rogers, N.K.; Murgas, P.; Bruna, B.; More, J.; et al. Senescence markers in peripheral blood mononuclear cells in amnestic mild cognitive impairment and Alzheimer’s disease. Int J Mol Sci 2022, 23, 9387. [Google Scholar] [CrossRef]
- Morales-Valencia, J. From DNA Damage Repair to Cellular Plasticity: How Cancer Cells Respond to Genotoxic Stress; New York University, 2022.
- Morrugares Carmona, R. Determination of New DYRK2 Functions in Response to Genotoxic Stress, 2022.
- Josipovic, N.; Ebbesen, K.K.; Zirkel, A.; Danieli-Mackay, A.; Dieterich, C.; Kurian, L.; Hansen, T.B.; Papantonis, A. circRAB3IP modulates cell proliferation by reorganizing gene expression and mRNA processing in a paracrine manner. RNA 2022, 28, 1481–1495. [Google Scholar] [CrossRef] [PubMed]
- Ohtani, N. The roles and mechanisms of senescence-associated secretory phenotype (SASP): can it be controlled by senolysis? Inflamm Regen 2022, 42, 11. [Google Scholar] [CrossRef]
- Durinikova, E.; Reilly, N.M.; Buzo, K.; Mariella, E.; Chilà, R.; Lorenzato, A.; Dias, J.M.L.; Grasso, G.; Pisati, F.; Lamba, S.; et al. Targeting the DNA damage response pathways and replication stress in colorectal cancer. Clin Cancer Res 2022, 28, 3874–3889. [Google Scholar] [CrossRef] [PubMed]
- Kong, M.; Guo, L.; Xu, W.; He, C.; Jia, X.; Zhao, Z.; Gu, Z. Aging-associated accumulation of mitochondrial DNA mutations in tumor origin. Life Med 2022, 1, 149–167. [Google Scholar] [CrossRef]
- Luzhin, A.; Rajan, P.; Safina, A.; Leonova, K.; Stablewski, A.; Wang, J.; Pal, M.; Kantidze, O.; Gurova, K. Comparison of cell response to chromatin and DNA damage. bioRxiv 2023, 01, 17.524424. [CrossRef]
- Chibaya, L.; Snyder, J.; Ruscetti, M., Eds. Senescence and the tumor-immune landscape: implications for cancer immunotherapy. Semin Cancer Biol; Elsevier 2022, 86, 827–845. [CrossRef]
- Ma, Y.; Li, S.; Ye, S.; Hu, D.; Wei, L.; Xiao, F. Hexavalent chromium triggers hepatocytes premature senescence via the GATA4/NF-κB signaling pathway mediated by the DNA damage response. Ecotoxicol Environ Saf 2022, 239, 113645. [Google Scholar] [CrossRef]
- Purohit, M.; Gupta, G.; Afzal, O.; Altamimi, A.S.A.; Alzarea, S.I.; Kazmi, I.; Almalki, W.H.; Gulati, M.; Kaur, I.P.; Singh, S.K.; et al. Janus kinase/signal transducers and activator of transcription (JAK/STAT) and its role in Lung inflammatory disease. Chem Biol Interact 2023, 371, 110334. [Google Scholar] [CrossRef]
- Molnár, A.Á.; Pásztor, D.; Merkely, B. Cellular senescence, aging and non-aging processes in calcified aortic valve stenosis: from Bench-Side to bedside. Cells 2022, 11, 3389. [Google Scholar] [CrossRef]
- Laubach, K.N.; Yan, W.; Kong, X.; Sun, W.; Chen, M.; Zhang, J.; Chen, X. p73α1, a p73 C-terminal isoform, regulates tumor suppression and the inflammatory response via Notch1. Proc Natl Acad Sci U S A 2022, 119, e2123202119. [Google Scholar] [CrossRef]
- Barrows, J.K.; Lin, B.; Quaas, C.E.; Fullbright, G.; Wallace, E.N.; Long, D.T. BRD4 promotes resection and homology-directed repair of DNA double-strand breaks. Nat Commun 2022, 13, 3016. [Google Scholar] [CrossRef]
- Markouli, M.; Strepkos, D.; Piperi, C. Impact of histone modifications and their therapeutic targeting in hematological malignancies. Int J Mol Sci 2022, 23, 13657. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Wang, Y.; Jaganathan, A.; Sun, Y.; Ma, N.; Li, N.; Han, X.; Sun, X.; Yi, H.; Fu, S.; et al. BRD4-PRC2 represses transcription of T-helper 2-specific negative regulators during T-cell differentiation. EMBO J 2023, 42, e111473. [Google Scholar] [CrossRef]
- He, J.; Liu, M.W.; Wang, Z.Y.; Shi, R.J. Protective effects of the notoginsenoside R1 on acute lung injury by regulating the miR-128-2-5p/Tollip signaling pathway in rats with severe acute pancreatitis. Innate Immun 2022, 28, 19–36. [Google Scholar] [CrossRef]
- Yao, J.; Miao, Y.; Zhang, Y.; Zhu, L.; Chen, H.; Wu, X.; Yang, Y.; Dai, X.; Hu, Q.; Wan, M.; et al. Dao-Chi Powder ameliorates pancreatitis-induced intestinal and cardiac injuries via regulating the Nrf2-HO-1-HMGB1 signaling pathway in rats. Front Pharmacol 2022, 13, 922130. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Simon, M.; Seluanov, A.; Gorbunova, V. DNA damage and repair in age-related inflammation. Nat Rev Immunol 2023, 23, 75–89. [Google Scholar] [CrossRef] [PubMed]
- Frediani, E.; Scavone, F.; Laurenzana, A.; Chillà, A.; Tortora, K.; Cimmino, I.; Leri, M.; Bucciantini, M.; Mangoni, M.; Fibbi, G.; et al. Olive phenols preserve lamin B1 expression reducing cGAS/STING/NFκB-mediated SASP in ionizing radiation-induced senescence. J Cell Mol Med 2022, 26, 2337–2350. [Google Scholar] [CrossRef]
- Kirkland, J.L. Tumor dormancy and disease recurrence. Cancer Metastasis Rev 2023, 42, 9–12. [Google Scholar] [CrossRef]
- Choudhary, S.; Sharma, K.; Silakari, O. The interplay between inflammatory pathways and COVID-19: A critical review on pathogenesis and therapeutic options. Microb Pathog 2021, 150, 104673. [Google Scholar] [CrossRef]
- Li, Y.; Adeniji, N.T.; Fan, W.; Kunimoto, K.; Török, N.J. Non-alcoholic fatty liver disease and liver fibrosis during aging. Aging Dis 2022, 13, 1239–1251. [Google Scholar] [CrossRef]
- Kim, K.H.; Cheng, N.; Lau, L.F. Cellular communication network factor 1-stimulated liver macrophage efferocytosis drives hepatic stellate cell activation and liver fibrosis. Hepatol Commun 2022, 6, 2798–2811. [Google Scholar] [CrossRef]
- Sun, G. The Role of BMP Signaling in DNA-Damage Responses During Zebrafish Heart Regeneration; Universität Ulm, 2022.
- Chaib, S.; Tchkonia, T.; Kirkland, J.L. Cellular senescence and senolytics: the path to the clinic. Nat Med 2022, 28, 1556–1568. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, C.A.; Wang, B.; Demaria, M. Senescence and cancer—role and therapeutic opportunities. Nat Rev Clin Oncol 2022, 19, 619–636. [Google Scholar] [CrossRef]
- Secomandi, L.; Borghesan, M.; Velarde, M.; Demaria, M. The role of cellular senescence in female reproductive aging and the potential for senotherapeutic interventions. Hum Reprod Update 2022, 28, 172–189. [Google Scholar] [CrossRef] [PubMed]
- Morsli, S.; Doherty, G.J.; Muñoz-Espín, D. Activatable senoprobes and senolytics: novel strategies to detect and target senescent cells. Mech Ageing Dev 2022, 202, 111618. [Google Scholar] [CrossRef]
- Mittal, D.; Gubin, M.M.; Schreiber, R.D.; Smyth, M.J. New insights into cancer immunoediting and its three component phases—elimination, equilibrium and escape. Curr Opin Immunol 2014, 27, 16–25. [Google Scholar] [CrossRef]
- Naylor, R.M.; Baker, D.J.; Van Deursen, J.M. Senescent cells: a novel therapeutic target for aging and age-related diseases. Clin Pharmacol Ther 2013, 93, 105–116. [Google Scholar] [CrossRef] [PubMed]
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. Hallmarks of aging: an expanding universe. Cell 2023, 186, 243–278. [Google Scholar] [CrossRef]
- Yousefzadeh, M.J.; Robbins, P.D.; Huffman, D.M. Heterochronic parabiosis: a valuable tool to investigate cellular senescence and other hallmarks of aging. Aging (Albany, NY) 2022, 14, 3325–3328. [Google Scholar] [CrossRef]
- Jeon, O.H.; Mehdipour, M.; Gil, T.H.; Kang, M.; Aguirre, N.W.; Robinson, Z.R.; Kato, C.; Etienne, J.; Lee, H.G.; Alimirah, F.; et al. Systemic induction of senescence in young mice after single heterochronic blood exchange. Nat Metab 2022, 4, 995–1006. [Google Scholar] [CrossRef]
- Yang, J.Y.; Deng, X.Y.; Li, Y.S.; Ma, X.C.; Feng, J.X.; Yu, B.; Chen, Y.; Luo, Y.L.; Wang, X.; Chen, M.L.; et al. Structure of Schlafen13 reveals a new class of tRNA/rRNA-targeting RNase engaged in translational control. Nat Commun 2018, 9, 1165. [Google Scholar] [CrossRef]
- Ninomiya-Tsuji, J.; Kajino, T.; Ono, K.; Ohtomo, T.; Matsumoto, M.; Shiina, M.; Mihara, M.; Tsuchiya, M.; Matsumoto, K. A resorcylic acid lactone, 5Z-7-oxozeaenol, prevents inflammation by inhibiting the catalytic activity of TAK1 MAPK kinase kinase. J Biol Chem 2003, 278, 18485–18490. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Fu, D.; Xu, Q.; Cong, X.; Wu, C.; Zhong, X.; Ma, Y.; Lv, Z.; Chen, F.; Han, L.; et al. The senescence-associated secretory phenotype is potentiated by feedforward regulatory mechanisms involving Zscan4 and TAK1. Nat Commun 2018, 9, 1723. [Google Scholar] [CrossRef] [PubMed]
- Tao, Z.F.; Hasvold, L.; Wang, L.; Wang, X.; Petros, A.M.; Park, C.H.; Boghaert, E.R.; Catron, N.D.; Chen, J.; Colman, P.M.; et al. Discovery of a potent and selective BCL-XL inhibitor with in vivo activity. ACS Med Chem Lett 2014, 5, 1088–1093. [Google Scholar] [CrossRef]
- Sharma, A.K.; Roberts, R.L.; Benson Jr., R.D.; Pierce, J.L.; Yu, K.; Hamrick, M.W.; McGee-Lawrence, M.E. The senolytic drug navitoclax (ABT-263) causes trabecular bone loss and impaired osteoprogenitor function in aged mice. Front Cell Dev Biol 2020, 8, 354. [CrossRef]
- Kim, H.; Jang, J.; Song, M.J.; Park, C.H.; Lee, D.H.; Lee, S.H.; Chung, J.H. Inhibition of matrix metalloproteinase expression by selective clearing of senescent dermal fibroblasts attenuates ultraviolet-induced photoaging. Biomed Pharmacother 2022, 150, 113034. [Google Scholar] [CrossRef] [PubMed]
- Cohen, S.; Hurd, E.; Cush, J.; Schiff, M.; Weinblatt, M.E.; Moreland, L.W.; Kremer, J.; Bear, M.B.; Rich, W.J.; McCabe, D. Treatment of rheumatoid arthritis with anakinra, a recombinant human interleukin-1 receptor antagonist, in combination with methotrexate: results of a twenty-four–week, multicenter, randomized, double-blind, placebo-controlled trial. Arthritis Rheum 2002, 46, 614–624. [Google Scholar] [CrossRef]
- Kuno, K.; Matsushima, K. The IL-1 receptor signaling pathway. J Leukoc Biol 1994, 56, 542–547. [Google Scholar] [CrossRef]
- Faget, D.V.; Ren, Q.; Stewart, S.A. Unmasking senescence: context-dependent effects of SASP in cancer. Nat Rev Cancer 2019, 19, 439–453. [Google Scholar] [CrossRef]
- Schett, G.; Dayer, J.M.; Manger, B. Interleukin-1 function and role in rheumatic disease. Nat Rev Rheumatol 2016, 12, 14–24. [Google Scholar] [CrossRef]
- Coppé, J.P.; Patil, C.K.; Rodier, F.; Sun, Y.; Muñoz, D.P.; Goldstein, J.; Nelson, P.S.; Desprez, P.Y.; Campisi, J. Senescence-associated secretory phenotypes reveal cell-nonautonomous functions of oncogenic RAS and the p53 tumor suppressor. PLOS Biol 2008, 6, 2853–2868. [Google Scholar] [CrossRef] [PubMed]
- Kuilman, T.; Michaloglou, C.; Vredeveld, L.C.; Douma, S.; van Doorn, R.; Desmet, C.J.; Aarden, L.A.; Mooi, W.J.; Peeper, D.S. Oncogene-induced senescence relayed by an interleukin-dependent inflammatory network. Cell 2008, 133, 1019–1031. [Google Scholar] [CrossRef]
- Ozsvari, B.; Nuttall, J.R.; Sotgia, F.; Lisanti, M.P. Azithromycin and Roxithromycin define a new family of “senolytic” drugs that target senescent human fibroblasts. Aging (Albany, NY) 2018, 10, 3294–3307. [Google Scholar] [CrossRef]
- Zhang, X.; Dong, Y.; Li, W.C.; Tang, B.X.; Li, J.; Zang, Y. Roxithromycin attenuates bleomycin-induced pulmonary fibrosis by targeting senescent cells. Acta Pharmacol Sin 2021, 42, 2058–2068. [Google Scholar] [CrossRef]
- Dhimolea, E., Ed. Canakinumab. mAbs; Taylor & Francis 2010, 2, 3–13. [CrossRef]
- Kuemmerle-Deschner, J.B.; Ramos, E.; Blank, N.; Roesler, J.; Felix, S.D.; Jung, T.; Stricker, K.; Chakraborty, A.; Tannenbaum, S.; Wright, A.M.; et al. Canakinumab (ACZ885, a fully human IgG1 anti-IL-1β mAb) induces sustained remission in pediatric patients with cryopyrin-associated periodic syndrome (CAPS). Arthritis Res Ther 2011, 13, R34. [Google Scholar] [CrossRef] [PubMed]
- Chien, Y.; Scuoppo, C.; Wang, X.; Fang, X.; Balgley, B.; Bolden, J.E.; Premsrirut, P.; Luo, W.; Chicas, A.; Lee, C.S.; et al. Control of the senescence-associated secretory phenotype by NF-κB promotes senescence and enhances chemosensitivity. Genes Dev 2011, 25, 2125–2136. [Google Scholar] [CrossRef] [PubMed]
- Ridker, P.M.; MacFadyen, J.G.; Glynn, R.J.; Koenig, W.; Libby, P.; Everett, B.M.; Lefkowitz, M.; Thuren, T.; Cornel, J.H. Inhibition of interleukin-1β by canakinumab and cardiovascular outcomes in patients with chronic kidney disease. J Am Coll Cardiol 2018, 71, 2405–2414. [Google Scholar] [CrossRef]
- Tasdemir, N.; Banito, A.; Roe, J.S.; Alonso-Curbelo, D.; Camiolo, M.; Tschaharganeh, D.F.; Huang, C.H.; Aksoy, O.; Bolden, J.E.; Chen, C.C.; et al. BRD4 connects enhancer remodeling to senescence immune SurveillanceBRD4. Cancer Discov 2016, 6, 612–629. [Google Scholar] [CrossRef] [PubMed]
- Cummings, B. Human Anatomy and Physiology; Auflage: San Francisco, 2001. [Google Scholar]
- Laberge, R.M.; Zhou, L.; Sarantos, M.R.; Rodier, F.; Freund, A.; de Keizer, P.L.; Liu, S.; Demaria, M.; Cong, Y.S.; Kapahi, P.; et al. Glucocorticoids suppress selected components of the senescence-associated secretory phenotype. Aging Cell 2012, 11, 569–578. [Google Scholar] [CrossRef]
- Ferguson-Smith, A.C.; Chen, Y.F.; Newman, M.S.; May, L.T.; Sehgal, P.B.; Ruddle, F.H. Regional localization of the interferon-β2B-cell stimulatory factor 2/hepatocyte stimulating factor gene to human chromosome 7p15-p21. Genomics 1988, 2, 203–208. [Google Scholar] [CrossRef]
- Chauntry, A.J.; Bishop, N.C.; Hamer, M.; Kingsnorth, A.P.; Chen, Y.L.; Paine, N.J. Sedentary behaviour is associated with heightened cardiovascular, inflammatory and cortisol reactivity to acute psychological stress. Psychoneuroendocrinology 2022, 141, 105756. [Google Scholar] [CrossRef]
- Han, X.; Lei, Q.; Xie, J.; Liu, H.; Li, J.; Zhang, X.; Zhang, T.; Gou, X. Potential regulators of the senescence-associated secretory phenotype during senescence and aging. J Gerontol A Biol Sci Med Sci 2022, 77, 2207–2218. [Google Scholar] [CrossRef]
- Rodier, F.; Muñoz, D.P.; Teachenor, R.; Chu, V.; Le, O.; Bhaumik, D.; Coppé, J.P.; Campeau, E.; Beauséjour, C.M.; Kim, S.H.; et al. DNA-SCARS: distinct nuclear structures that sustain damage-induced senescence growth arrest and inflammatory cytokine secretion. J Cell Sci 2011, 124, 68–81. [Google Scholar] [CrossRef] [PubMed]
- Moghadam-Kia, S.; Werth, V.P. Prevention and treatment of systemic glucocorticoid side effects. Int J Dermatol 2010, 49, 239–248. [Google Scholar] [CrossRef]
- Novais, E.J.; Tran, V.A.; Johnston, S.N.; Darris, K.R.; Roupas, A.J.; Sessions, G.A.; Shapiro, I.M.; Diekman, B.O.; Risbud, M.V. Long-term treatment with senolytic drugs dasatinib and quercetin ameliorates age-dependent intervertebral disc degeneration in mice. Nat Commun 2021, 12, 5213. [Google Scholar] [CrossRef]
- Cui, Z.; Zhao, X.; Amevor, F.K.; Du, X.; Wang, Y.; Li, D.; Shu, G.; Tian, Y.; Zhao, X. Therapeutic application of quercetin in aging-related diseases: SIRT1 as a potential mechanism. Front Immunol 2022, 13, 943321. [Google Scholar] [CrossRef]
- Chu, J.J.; Ji, W.B.; Zhuang, J.H.; Gong, B.F.; Chen, X.H.; Cheng, W.B.; Liang, W.D.; Li, G.R.; Gao, J.; Yin, Y. Nanoparticles-based anti-aging treatment of Alzheimer’s disease. Drug Deliv 2022, 29, 2100–2116. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; He, Y.; Zhang, R.; Zheng, G.; Zhou, D. The curcumin analog EF24 is a novel senolytic agent. Aging (Albany, NY) 2019, 11, 771–782. [Google Scholar] [CrossRef]
- Zhang, L.; Pitcher, L.E.; Prahalad, V.; Niedernhofer, L.J.; Robbins, P.D. Targeting cellular senescence with senotherapeutics: senolytics and senomorphics. FEBS Journal 2023, 290, 1362–1383. [Google Scholar] [CrossRef]
- Klareskog, L.; van der Heijde, D.; de Jager, J.P.; Gough, A.; Kalden, J.; Malaise, M.; Martín Mola, E.; Pavelka, K.; Sany, J.; Settas, L.; et al. Therapeutic effect of the combination of etanercept and methotrexate compared with each treatment alone in patients with rheumatoid arthritis: double-blind randomized controlled trial. Lancet 2004, 363, 675–681. [Google Scholar] [CrossRef]
- Rider, P.; Carmi, Y.; Cohen, I. Biologics for targeting inflammatory cytokines, clinical uses, and limitations. Int J Cell Biol 2016, 2016, 9259646. [Google Scholar] [CrossRef] [PubMed]
- Fizazi, K.; De Bono, J.S.; Flechon, A.; Heidenreich, A.; Voog, E.; Davis, N.B.; Qi, M.; Bandekar, R.; Vermeulen, J.T.; Cornfeld, M.; et al. Randomised phase II study of siltuximab (CNTO 328), an anti-IL-6 monoclonal antibody, in combination with mitoxantrone/prednisone versus mitoxantrone/prednisone alone in metastatic castration-resistant prostate cancer. Eur J Cancer 2012, 48, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Moore, G.; Annett, S.; McClements, L.; Robson, T. Top notch targeting strategies in cancer: a detailed overview of recent insights and current perspectives. Cells 2020, 9, 1503. [Google Scholar] [CrossRef] [PubMed]
- Le Naour, J.; Zitvogel, L.; Galluzzi, L.; Vacchelli, E.; Kroemer, G. Trial watch: STING agonists in cancer therapy. Oncoimmunology 2020, 9, 1777624. [Google Scholar] [CrossRef]
- Sahu, B.D.; Kalvala, A.K.; Koneru, M.; Mahesh Kumar, J.; Kuncha, M.; Rachamalla, S.S.; Sistla, R. Ameliorative effect of fisetin on cisplatin-induced nephrotoxicity in rats via modulation of NF-κB activation and antioxidant defence. PLOS ONE 2014, 9, e105070. [Google Scholar] [CrossRef]
- Zhu, Y.; Doornebal, E.J.; Pirtskhalava, T.; Giorgadze, N.; Wentworth, M.; Fuhrmann-Stroissnigg, H.; Niedernhofer, L.J.; Robbins, P.D.; Tchkonia, T.; Kirkland, J.L. New agents that target senescent cells: the flavone, fisetin, and the BCL-XL inhibitors, A1331852 and A1155463. Aging (Albany, NY) 2017, 9, 955–963. [Google Scholar] [CrossRef] [PubMed]
- Molagoda, I.M.N.; Jayasingha, J.A.C.C.; Choi, Y.H.; Jayasooriya, R.G.P.T.; Kang, C.H.; Kim, G.Y. Fisetin inhibits lipopolysaccharide-induced inflammatory response by activating β-catenin, leading to a decrease in endotoxic shock. Sci Rep 2021, 11, 8377. [Google Scholar] [CrossRef]
- Huang, C.; Filippone, N.R.; Reiner, T.; Roberts, S. Sensors and Inhibitors for the detection of ataxia telangiectasia mutated (ATM) protein kinase. Mol Pharm 2021, 18, 2470–2481. [Google Scholar] [CrossRef] [PubMed]
- Abuetabh, Y.; Wu, H.H.; Chai, C.; Al Yousef, H.; Persad, S.; Sergi, C.M.; Leng, R. DNA damage response revisited: the p53 family and its regulators provide endless cancer therapy opportunities. Exp Mol Med 2022, 54, 1658–1669. [Google Scholar] [CrossRef]
- Aghali, A.; Koloko Ngassie, M.L.; Pabelick, C.M.; Prakash, Y.S. Cellular senescence in aging lungs and diseases. Cells 2022, 11, 1781. [Google Scholar] [CrossRef]
- Park, J.Y.; Lee, H.; Song, E.S.; Lee, Y.H.; Kuk, M.U.; Ko, G.; Kwon, H.W.; Byun, Y.; Park, J.T. Restoration of lysosomal and mitochondrial function through p38 mitogen-activated protein kinase inhibition ameliorates senescence. Rejuvenation Res 2022, 25, 291–299. [Google Scholar] [CrossRef]
- Kang, H.T.; Park, J.T.; Choi, K.; Kim, Y.; Choi, H.J.C.; Jung, C.W.; Lee, Y.S.; Park, S.C. Chemical screening identifies ATM as a target for alleviating senescence. Nat Chem Biol 2017, 13, 616–623. [Google Scholar] [CrossRef]
- Özeş, A.R.; Miller, D.F.; Özeş, O.N.; Fang, F.; Liu, Y.; Matei, D.; Huang, T.; Nephew, K.P. NF-κB-HOTAIR axis links DNA damage response, chemoresistance and cellular senescence in ovarian cancer. Oncogene 2016, 35, 5350–5361. [Google Scholar] [CrossRef]
- Rebbeck, T.R.; Devesa, S.S.; Chang, B.L.; Bunker, C.H.; Cheng, I.; Cooney, K.; Eeles, R.; Fernandez, P.; Giri, V.N.; Gueye, S.M.; et al. Global patterns of prostate cancer incidence, aggressiveness, and mortality in men of african descent. Prostate Cancer 2013, 2013, 560857. [Google Scholar] [CrossRef] [PubMed]
- Fuhrmann-Stroissnigg, H.; Ling, Y.Y.; Zhao, J.; McGowan, S.J.; Zhu, Y.; Brooks, R.W.; Grassi, D.; Gregg, S.Q.; Stripay, J.L.; Dorronsoro, A.; et al. Identification of HSP90 inhibitors as a novel class of senolytics. Nat Commun 2017, 8, 422. [Google Scholar] [CrossRef]
- Fleming, J.E.; Miquel, J.; Cottrell, S.F.; Yengoyan, L.S.; Economos, A.C. Is cell aging caused by respiration-dependent injury to the mitochondrial genome? Gerontology 1982, 28, 44–53. [Google Scholar] [CrossRef]
- Richie Jr., J.P.; Mills, B.J.; Lang, C.A. Dietary nordihydroguaiaretic acid increases the life span of the mosquito. Proc Soc Exp Biol Med 1986, 183, 81–85. [CrossRef]
- Harrison, D.E.; Strong, R.; Allison, D.B.; Ames, B.N.; Astle, C.M.; Atamna, H.; Fernandez, E.; Flurkey, K.; Javors, M.A.; Nadon, N.L.; et al. Acarbose, 17-α-estradiol, and nordihydroguaiaretic acid extend mouse lifespan preferentially in males. Aging Cell 2014, 13, 273–282. [Google Scholar] [CrossRef]
- Lozano-Torres, B.; Estepa-Fernández, A.; Rovira, M.; Orzáez, M.; Serrano, M.; Martínez-Máñez, R.; Sancenón, F. The chemistry of senescence. Nat Rev Chem 2019, 3, 426–441. [Google Scholar] [CrossRef]
- Jenny, N.S. Inflammation in aging: cause, effect, or both? Discov Med 2012, 13, 451–460. [Google Scholar] [PubMed]
- Strong, R.; Miller, R.A.; Astle, C.M.; Floyd, R.A.; Flurkey, K.; Hensley, K.L.; Javors, M.A.; Leeuwenburgh, C.; Nelson, J.F.; Ongini, E.; et al. Nordihydroguaiaretic acid and aspirin increase lifespan of genetically heterogeneous male mice. Aging Cell 2008, 7, 641–650. [Google Scholar] [CrossRef]
- Guerrero, A.; Herranz, N.; Sun, B.; Wagner, V.; Gallage, S.; Guiho, R.; Wolter, K.; Pombo, J.; Irvine, E.E.; Innes, A.J.; et al. Cardiac glycosides are broad-spectrum senolytics. Nat Metab 2019, 1, 1074–1088. [Google Scholar] [CrossRef]
- Tedesco-Silva, H.; Saliba, F.; Barten, M.J.; De Simone, P.; Potena, L.; Gottlieb, J.; Gawai, A.; Bernhardt, P.; Pascual, J. An overview of the efficacy and safety of everolimus in adult solid organ transplant recipients. Transplant Rev (Orlando) 2022, 36, 100655. [Google Scholar] [CrossRef]
- Laberge, R.M.; Sun, Y.; Orjalo, A.V.; Patil, C.K.; Freund, A.; Zhou, L.; Curran, S.C.; Davalos, A.R.; Wilson-Edell, K.A.; Liu, S.; et al. MTOR regulates the pro-tumorigenic senescence-associated secretory phenotype by promoting IL1A translation. Nat Cell Biol 2015, 17, 1049–1061. [Google Scholar] [CrossRef] [PubMed]
- Shavlakadze, T.; Zhu, J.; Wang, S.; Zhou, W.; Morin, B.; Egerman, M.A.; Fan, L.; Wang, Y.; Iartchouk, O.; Meyer, A.; et al. Short-term low-dose mTORC1 inhibition in aged rats counter-regulates age-related gene changes and blocks age-related kidney pathology. J Gerontol A Biol Sci Med Sci 2018, 73, 845–852. [Google Scholar] [CrossRef] [PubMed]
- Terkeltaub, R.; Sundy, J.S.; Schumacher, H.R.; Murphy, F.; Bookbinder, S.; Biedermann, S.; Wu, R.; Mellis, S.; Radin, A. The interleukin 1 inhibitor Rilonacept in treatment of chronic gouty arthritis: results of a placebo-controlled, monosequence crossover, non-randomised, single-blind pilot study. Ann Rheum Dis 2009, 68, 1613–1617. [Google Scholar] [CrossRef]
- Hoffman, H.M.; Throne, M.L.; Amar, N.J.; Sebai, M.; Kivitz, A.J.; Kavanaugh, A.; Weinstein, S.P.; Belomestnov, P.; Yancopoulos, G.D.; Stahl, N.; et al. Efficacy and safety of Rilonacept (interleukin-1 Trap) in patients with cryopyrin-associated periodic syndromes: results from two sequential placebo-controlled studies. Arthritis Rheum 2008, 58, 2443–2452. [Google Scholar] [CrossRef]
- Liao, C.; Xiao, Y.; Liu, L. The dynamic process and its dual effects on tumors of therapy-induced senescence. Cancer Manag Res 2020, 12, 13553–13566. [Google Scholar] [CrossRef]
- Mesa, R.A.; Yasothan, U.; Kirkpatrick, P. Ruxolitinib. Nat Rev Drug Discov 2012, 11, 103–104. [Google Scholar] [CrossRef]
- Wang, L.; Wang, B.; Gasek, N.S.; Zhou, Y.; Cohn, R.L.; Martin, D.E.; Zuo, W.; Flynn, W.F.; Guo, C.; Jellison, E.R.; et al. Targeting p21Cip1 highly expressing cells in adipose tissue alleviates insulin resistance in obesity. Cell Metab 2022, 34, 75–89. [Google Scholar] [CrossRef] [PubMed]
- Harrison, C.N.; Talpaz, M.; Mead, A.J. Ruxolitinib is effective in patients with intermediate-1 risk myelofibrosis: a summary of recent evidence. Leuk Lymphoma 2016, 57, 2259–2267. [Google Scholar] [CrossRef]
- Xu, M.; Tchkonia, T.; Ding, H.; Ogrodnik, M.; Lubbers, E.R.; Pirtskhalava, T.; White, T.A.; Johnson, K.O.; Stout, M.B.; Mezera, V.; et al. JAK inhibition alleviates the cellular senescence-associated secretory phenotype and frailty in old age. Proc Natl Acad Sci U S A 2015, 112, E6301–E6310. [Google Scholar] [CrossRef]
- Xu, M.; Palmer, A.K.; Ding, H.; Weivoda, M.M.; Pirtskhalava, T.; White, T.A.; Sepe, A.; Johnson, K.O.; Stout, M.B.; Giorgadze, N.; et al. Targeting senescent cells enhances adipogenesis and metabolic function in old age. eLife 2015, 4, e12997. [Google Scholar] [CrossRef]
- Freund, A.; Patil, C.K.; Campisi, J. P38MAPK is a novel DNA damage response-independent regulator of the senescence-associated secretory phenotype. EMBO J 2011, 30, 1536–1548. [Google Scholar] [CrossRef]
- Hou, J.; Cui, C.; Kim, S.; Sung, C.; Choi, C. Ginsenoside F1 suppresses astrocytic senescence-associated secretory phenotype. Chem Biol Interact 2018, 283, 75–83. [Google Scholar] [CrossRef]
- Lali, F.V.; Hunt, A.E.; Turner, S.J.; Foxwell, B.M. The pyridinyl imidazole inhibitor SB203580 blocks phosphoinositide-dependent protein kinase activity, protein kinase B phosphorylation, and retinoblastoma hyperphosphorylation in interleukin-2-stimulated T cells independently of p38 mitogen-activated protein kinase. J Biol Chem 2000, 275, 7395–7402. [Google Scholar] [CrossRef]
- Gordon, G.M.; LaGier, A.J.; Ponchel, C.; Bauskar, A.; Itakura, T.; Jeong, S.; Patel, N.; Fini, M.E. A cell-based screening assay to identify pharmaceutical compounds that enhance the regenerative quality of corneal repair. Wound Repair Regen 2016, 24, 89–99. [Google Scholar] [CrossRef] [PubMed]
- del Peso, Ld.; González-Garcıa, M.; Page, C.; Herrera, R.; Nuñez, G. Interleukin-3-induced phosphorylation of BAD through the protein kinase Akt. Science 1997, 278, 687–689. [Google Scholar] [CrossRef] [PubMed]
- Cesca, T.G.; Faqueti, L.G.; Rocha, L.W.; Meira, N.A.; Meyre-Silva, C.; De Souza, M.M.; Quintão, N.L.; Silva, R.M.; Filho, V.C.; Bresolin, T.M. Antinociceptive, anti-inflammatory and wound healing features in animal models treated with a semisolid herbal medicine based on Aleurites moluccana L. Willd. Euforbiaceae standardized leaf extract: semisolid herbal. J Ethnopharmacol 2012, 143, 355–362. [Google Scholar] [CrossRef]
- Liu, S.; Uppal, H.; Demaria, M.; Desprez, P.Y.; Campisi, J.; Kapahi, P. Simvastatin suppresses breast cancer cell proliferation induced by senescent cells. Sci Rep 2015, 5, 17895. [Google Scholar] [CrossRef] [PubMed]
- Jaffe, A.B.; Hall, A. Rho GTPases: biochemistry and biology. Annu Rev Cell Dev Biol 2005, 21, 247–269. [Google Scholar] [CrossRef] [PubMed]
- Alexander, K.; Yang, H.S.; Hinds, P.W. Cellular senescence requires CDK5 repression of Rac1 activity. Mol Cell Biol 2004, 24, 2808–2819. [Google Scholar] [CrossRef]
- Kim, K.H.; Chen, C.C.; Monzon, R.I.; Lau, L.F. Matricellular protein CCN1 promotes regression of liver fibrosis through induction of cellular senescence in hepatic myofibroblasts. Mol Cell Biol 2013, 33, 2078–2090. [Google Scholar] [CrossRef]
- Deshpande, S.S.; Qi, B.; Park, Y.C.; Irani, K. Constitutive activation of rac1 results in mitochondrial oxidative stress and induces premature endothelial cell senescence. Arterioscler Thromb Vasc Biol 2003, 23, e1–e6. [Google Scholar] [CrossRef]
- Wang, L.; Yang, L.; Debidda, M.; Witte, D.; Zheng, Y. Cdc42 GTPase-activating protein deficiency promotes genomic instability and premature aging-like phenotypes. Proc Natl Acad Sci U S A 2007, 104, 1248–1253. [Google Scholar] [CrossRef]
- Korneev, K.V.; Atretkhany, K.-S.N.; Drutskaya, M.S.; Grivennikov, S.I.; Kuprash, D.V.; Nedospasov, S.A. TLR-signaling and proinflammatory cytokines as drivers of tumorigenesis. Cytokine 2017, 89, 127–135. [Google Scholar] [CrossRef]
- Van Rhee, F.; Wong, R.S.; Munshi, N.; Rossi, J.F.; Ke, X.Y.; Fosså, A.; Simpson, D.; Capra, M.; Liu, T.; Hsieh, R.K.; et al. Siltuximab for multicentric Castleman’s disease: a randomised, double-blind, placebo-controlled trial. Lancet Oncol 2014, 15, 966–974. [Google Scholar] [CrossRef]
- Chen, R.; Chen, B. Siltuximab (CNTO 328): a promising option for human malignancies. Drug Des Devel Ther 2015, 9, 3455–3458. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Teachey, D.T.; Pequignot, E.; Frey, N.; Porter, D.; Maude, S.L.; Grupp, S.A.; June, C.H.; Melenhorst, J.J.; Lacey, S.F. Measuring IL-6 and sIL-6R in serum from patients treated with tocilizumab and/or siltuximab following CAR T cell therapy. J Immunol Methods 2016, 434, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Emery, P.; Keystone, E.; Tony, H.P.; Cantagrel, A.; Van Vollenhoven, R.; Sanchez, A.; Alecock, E.; Lee, J.; Kremer, J. IL-6 receptor inhibition with tocilizumab improves treatment outcomes in patients with rheumatoid arthritis refractory to anti-tumour necrosis factor BioLogicals: results from a 24-week multicentre randomised placebo-controlled trial. Ann Rheum Dis 2008, 67, 1516–1523. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, S.; Kawamoto, S.; Ohtani, N.; Hara, E. Impact of senescence-associated secretory phenotype and its potential as a therapeutic target for senescence-associated diseases. Cancer Sci 2017, 108, 563–569. [Google Scholar] [CrossRef]
- Lau, L.; Porciuncula, A.; Yu, A.; Iwakura, Y.; David, G. Uncoupling the senescence-associated secretory phenotype from cell cycle exit via interleukin-1 inactivation unveils its protumorigenic role. Mol Cell Biol 2019, 39, e00586-18. [Google Scholar] [CrossRef]
- Coryell, P.R.; Diekman, B.O.; Loeser, R.F. Mechanisms and therapeutic implications of cellular senescence in osteoarthritis. Nat Rev Rheumatol 2021, 17, 47–57. [Google Scholar] [CrossRef]
- Colich, N.L.; Rosen, M.L.; Williams, E.S.; McLaughlin, K.A. Biological aging in childhood and adolescence following experiences of threat and deprivation: A systematic review and meta-analysis. Psychol Bull 2020, 146, 721–764. [Google Scholar] [CrossRef] [PubMed]
- Moulahoum, H.; Ghorbanizamani, F.; Khiari, Z.; Toumi, M.; Benazzoug, Y.; Tok, K.; Timur, S.; Zihnioglu, F. Artemisia alleviates AGE-induced liver complications via MAPK and RAGE signaling pathways modulation: a combinatorial study. Mol Cell Biochem 2022, 477, 2345–2357. [Google Scholar] [CrossRef]
- Furrer, R.; Handschin, C. Drugs, clocks and exercise in ageing: hype and hope, fact and fiction. J Physiol 2023, 601, 2057–2068. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Lankhorst, L.; Bernards, R. Exploiting senescence for the treatment of cancer. Nat Rev Cancer 2022, 22, 340–355. [Google Scholar] [CrossRef] [PubMed]
- Jin, P.; Li, X.; Xia, Y.; Li, H.; Li, X.; Yang, Z.Y.; Wang, Z.; Xu, C.; Fang, T.; Zhou, D.; et al. Bepotastine sensitizes ovarian cancer to PARP inhibitors through suppressing NF-κB-triggered SASP in cancer-associated fibroblasts. Mol Cancer Ther 2023, 22, 447–458. [Google Scholar] [CrossRef] [PubMed]
- Sofiadis, K.; Josipovic, N.; Nikolic, M.; Kargapolova, Y.; Übelmesser, N.; Varamogianni-Mamatsi, V.; Zirkel, A.; Papadionysiou, I.; Loughran, G.; Keane, J.; et al. HMGB1 coordinates SASP-related chromatin folding and RNA homeostasis on the path to senescence. Mol Syst Biol 2021, 17, e9760. [Google Scholar] [CrossRef]
- Rodrigues, F.C.; Anil Kumar, N.V.; Thakur, G. Developments in the anticancer activity of structurally modified curcumin: an up-to-date review. Eur J Med Chem 2019, 177, 76–104. [Google Scholar] [CrossRef]
- Zhao, R.C.; Stambler, I. The urgent need for international action for anti-aging and disease prevention. Aging Dis 2020, 11, 212–215. [Google Scholar] [CrossRef]
Figure 1.
Detailed signaling pathway targeting DDRs.
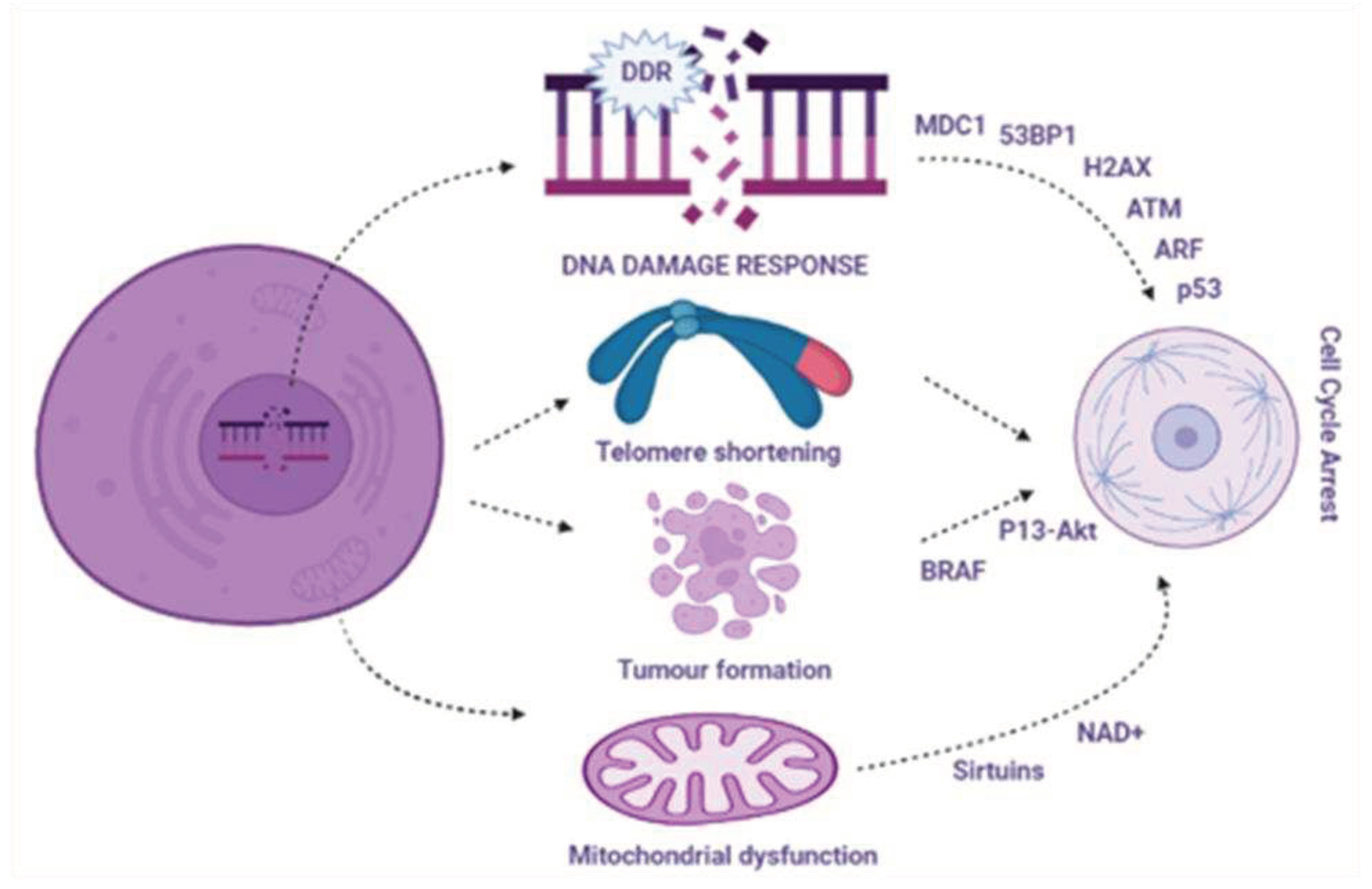
Figure 2.
Detailed signaling pathway targeting the SASP.
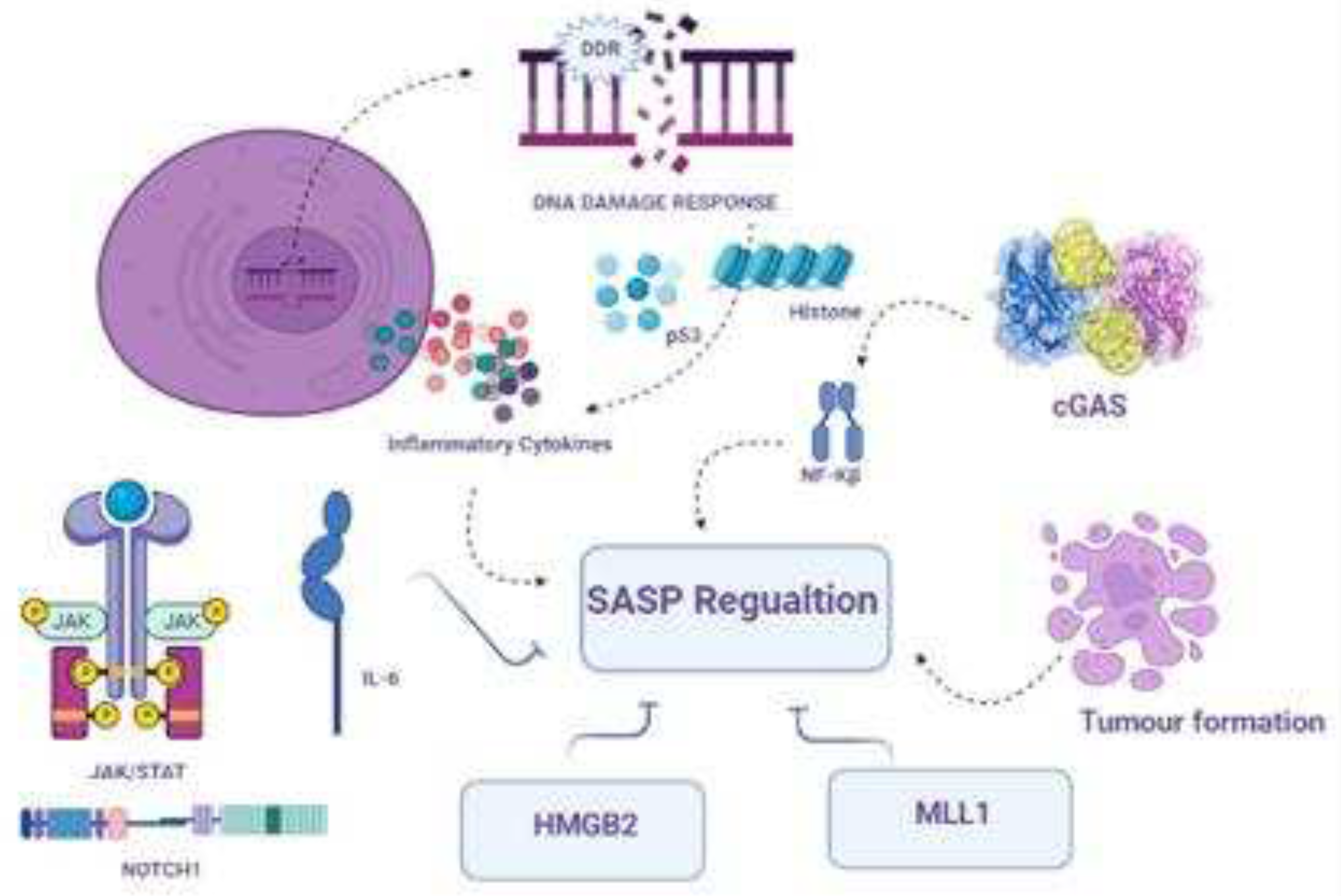
Figure 3.
Links among potential geroprotective and anti-aging compounds with senescence via DDR and SASP signaling pathways.
Figure 3.
Links among potential geroprotective and anti-aging compounds with senescence via DDR and SASP signaling pathways.
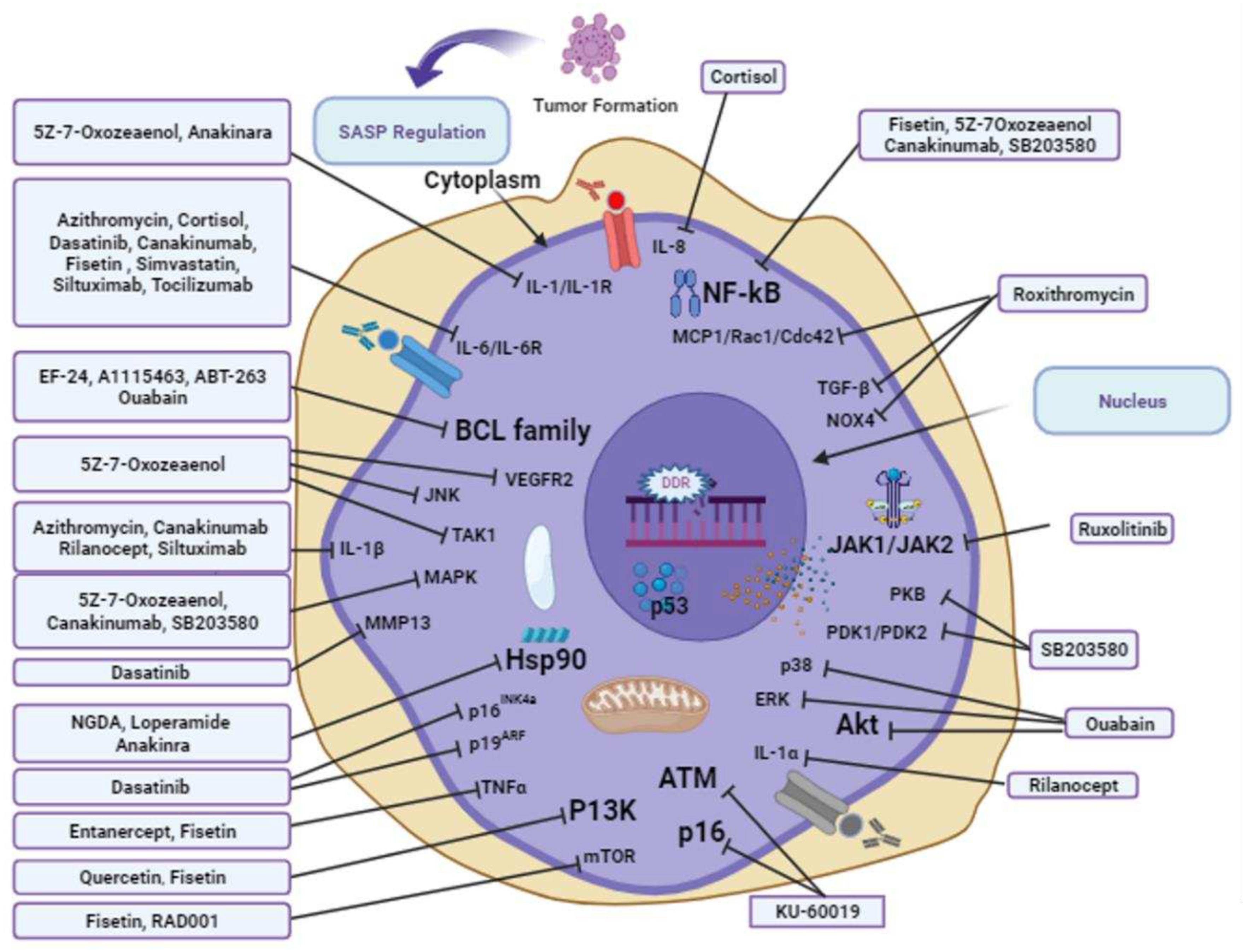
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



