Submitted:
26 September 2023
Posted:
28 September 2023
You are already at the latest version
Abstract
Radiotherapy and cisplatin-based chemotherapy belong to the main treatment modalities for head and neck squamous cell carcinoma (HNSCC), and induce cancer cell death by generating DNA damage, including the most severe double strand breaks (DSBs). Alterations in DSB response and repair genes may affect individual DNA repair capacity and treatment sensitivity, contributing to therapy resistance and poor prognosis often observed in HNSCC. In this study, we investigated the association of a panel of single nucleotide polymorphisms (SNPs) in 20 DSB signaling and repair genes with therapy results and prognosis in 505 HNSCC patients treated non-surgically with DNA damage-inducing therapies. In the multivariate analysis, there were a total of 14 variants associated with overall, locoregional recurrence-free or metastasis-free survival. Moreover, we identified 10 of these SNPs as independent predictors of therapy failure and unfavorable prognosis in the whole group or in two treatment subgroups. They were MRE11A rs2155209, XRCC5 rs828907, RAD51 rs1801321, rs12593359, LIG4 rs1805388, CHEK1 rs558351, TP53 rs1042522, ATM rs1801516, XRCC6 rs2267437 and NBS1 rs2735383. Only CHEK1 rs558351 remained statistically significant after correction for multiple testing. These results suggest that specific germline variants related to DSB response and repair may be potential genetic modifiers of therapy effects and disease progression in HNSCC treated with radiotherapy and cisplatin-based chemoradiation.
Keywords:
DNA repair
; genetic polymorphism
; head and neck cancer
; radiotherapy
; survival
; CHEK1
; MRE11
; XRCC5
; XRCC6
; RAD51
; LIG4
; ATM
; TP53
; NBS1
1. Introduction
In recent years, an increase in the incidence of head and neck cancer (HNC) has been observed both in Poland and in the world [1,2]. HNCs, the majority of which are squamous cell carcinomas (HNSCCs), represent a significant clinical problem as patients often present with advanced disease, and current treatments are associated with high toxicity and resistance, adversely affecting patients’ quality of life and survival rates. Radiation therapy (RT) and cisplatin-based chemotherapy (CT) are essential therapeutic strategies in HNC [2]. Their mechanism of action is based on the ability to induce various types of DNA damage, both directly and indirectly, the most harmful of which are double strand breaks (DSBs) and interstrand crosslinks (ICLs). Generated by ionizing radiation (IR) and free radicals, DSBs are substrates for DNA repair proteins belonging to two major pathways – fast, more error-prone non-homologous end joining (NHEJ), and accurate but slower homologous recombination repair (HR). In addition to the repair of DSBs, HR also participates in ICL and single-strand gaps repair [3,4].
DSBs trigger DNA damage response (DDR) signaling cascade that involves a network of multiple proteins acting as sensors, transducers/mediators or effectors, and comprises damage recognition, cell cycle arrest, DNA repair and cell death [5]. The MRE11-RAD50-NBS1 (MRN) protein complex is responsible for initial DSBs sensing, signal transduction and response to lesions, and functions in both HR and NHEJ [6]. Activation of the ATM-CHEK2 and ATR-CHEK1 checkpoint kinase pathways leads to phosphorylation of the tumor suppressor protein p53 resulting in the transcription of many downstream target genes that regulate cell cycle, apoptosis, senescence and repair [7]. The ATM together with DNA-dependent protein kinase (DNA-PK) promote NHEJ, which represents a dominant pathway for DSBs repair in mammalian cells. Active DNA-PK complex consists of a catalytic subunit (DNA-PKcs, also known as PRKDC or XRCC7) and Ku70/Ku80 (alias XRCC6/XRCC5) heterodimer necessary for damage detection. Then, other proteins such as XRCC4 and DNA ligase IV (LIG4) are recruited, which allows direct ligation of the DNA ends [4]. In turn, in the HR pathway, that requires a template strand to repair the break, RAD51 recombinase plays a central role in homology search and strand exchange events. RAD51 paralogs, such as e.g. XRCC2 and XRCC3, as well as many other proteins, including BRCA2, RAD52 and RAD54, participate in RAD51 recruitment and the formation and stabilization of the presynaptic filament [3]. Interacting with multiple proteins, BRCA1 is implicated in DDR, including cell cycle checkpoint activation, transcription regulation and promoting HR repair [8].
Impaired DDR mechanisms may lead to genomic instability and malignant transformation, as well as may be important for anticancer therapy. Rare germline mutations in certain key genes involved in DSB signaling and repair are associated with cancer-prone syndromes and increased sensitivity to IR or chemotherapeutic agents. [3,9]. However, numerous data indicate that also common inherited variation, such as single nucleotide polymorphisms (SNPs), in these genes may modulate DSB repair capacity, susceptibility to cancer and treatment effects [10,11]. Similarly to other solid tumors, HNC is characterized by radio- and chemoresistance, which contribute to poor therapy outcomes and survival. Enhanced DSB repair may be an important mechanism of therapeutic resistance and, consequently, disease progression in many solid cancers [12]. In HNSCC, for example, NBS1 and XRCC5 overexpression was associated with metastasis and locoregional failure after RT [13,14]. In oral cancer, high MRE11 and RAD51 levels correlated with radiation resistance and poor prognosis [15,16]. Significant inter-individual differences in response to RT and CT are observed in HNC patients, which may be due to, among others, host genetic factors such as SNPs. In HNC, SNPs in HR and NHEJ genes have been rarely investigated in the context of treatment efficiency, cancer progression and patient survival. Therefore, the objective of our study was to assess the influence of common SNPs in 20 core genes involved in DSB response and repair on therapy results and prognosis in patients with unresected HNSCC receiving DNA damaging treatment.
2. Materials and Methods
2.1. Study group
The study group comprised 505 Caucasian patients with primary HNSCC, qualified to curative-intent therapy at the Maria Skłodowska-Curie National Research Institute of Oncology in Gliwice. The inclusion criteria were: tumor located in the larynx (LSCC), oropharynx (OPSCC) or hypopharynx (HPSCC), clinical stage T1-4N0-3M0, WHO 0–1 performance status, treatment with radical RT, no surgery for HNC and no previous treatment for other malignancy. Patients received RT alone (n = 244, 48%) or combined with cisplatin-based CT (n = 261, 52%). The median total radiation dose was 70 Gy (range 50–72 Gy). In the combination treatment subgroup (RT+CT), there were 127 (25%) individuals given induction CT (docetaxel 75 mg/m2, cisplatin 75 mg/m2 and 5-fluorouracil 750 mg/m2, or cisplatin 100 mg/m2 and 5-fluorouracil 1000 mg/m2) and 211 (42%) patients given concurrent radiochemotherapy (based on cisplatin 100 mg/m2). Details of treatment and follow-up of the patients have been described previously [17]. The mean age at diagnosis was 59.7 years (range 30–87 years), with a median age of 59 years. Out of the patients, 362 (72%) had clinical stage III or IV, 398 (79%) were males and 393 (78%) had cigarette smoking history. Clinicopathological data were obtained from the medical records. The demographic and clinical parameters are presented in Table 1.
2.2. SNP genotyping
Genomic DNA was extracted from frozen peripheral blood using Genomic Maxi AX kit (A&A Biotechnology, Gdynia, Poland). SNP identification was performed using commercially available Taqman SNP Genotyping Assays (Applied Biosystems, Foster City, CA, USA), following the manufacturer’s standard protocol. Genotyping was repeated in 10% of randomly selected samples and resulted in 100% concordance.
There were 31 SNPs in 20 candidate genes investigated in the study (Table S1). We focused on known variants in key genes involved in the DSB response and repair that have been previously reported in the literature. Variant selection included SNPs that were functional or possibly functional, and/or located in regions likely influencing gene expression or protein levels/function, and/or related to cancer, and had a minor allele frequency (MAF) ≥ 10% in the European population [18]. One SNP (i.e. XRCC2 rs3218384), that failed to be genotyped using the appropriate Taqman assay, was excluded from further analysis.
2.3. Statistical analysis
The study endpoints were overall survival (OS), locoregional recurrence-free survival (LRFS) and metastasis-free survival (MFS). OS was calculated from the date of diagnosis to the date of death from any cause or the last known date alive. LRFS and MFS were calculated from the last day of treatment to the date of clinically detectable relapse (local and/or regional for LRFS, and distant for MFS) or the last examination without evidence of disease. The Kaplan-Meier plots and log-rank test were used to compare survival curves. The association between each SNP and survival was tested under additive, dominant and recessive genetic models, and the best model (i.e. with the lowest p value) was selected for the analysis. Univariate and multivariate Cox proportional hazards regression was used to estimate the hazard ratios (HRs) and 95% confidence intervals (CIs). All multivariate models were adjusted for potential confounders including median age at diagnosis, sex, T stage, N stage, tumor subsite, chemotherapy use, smoking, alcohol use, local and regional relapse (for OS and MFS), and metastasis or second primary cancer (SPC) diagnosed during follow-up (for OS only). A backward stepwise regression was also performed in order to identify independent risk factors for each endpoint. Pearson’s chi-square test and Spearman’s correlation were applied to examine the associations between variables. The Bonferroni correction was used to account for multiple testing (with the level of significance set at ≤ 0.002). However, due to the exploratory nature of this study, uncorrected p values were reported and p ≤ 0.05 was considered the threshold for statistical significance. All tests were two-sided and analyses were carried out using STATISTICA 13.1 (TIBCO Software Inc., Palo Alto, CA, USA).
3. Results
The genotype distribution was shown in Table S1 and the observed MAFs in the group were in line with the data reported for European populations [18]. The median OS was 71.4 months (range 4–161), while the median LRFS and MFS were not reached. The 5-year OS rate was 54.8%, the 2-year LRFS rate was 73.0% and the 5-year MFS rate was 83.7%. During the follow-up time (median 81.3 months), there were 251 (50%) deaths and 152 (30%) locoregional recurrences, 60 (12%) patients developed distant metastasis and SPC was diagnosed in 59 (12%) patients.
In total, there were six SNPs significantly associated with survival endpoints in the univariate analysis. In the whole group, patients with one or two XRCC5 rs1051677 C alleles had shorter MFS than TT homozygotes (p log-rank 0.048, HR 1.81, 95% CI 1.02–3.20; Figure 1A). The TP53 rs1042522 CC homozygotes showed reduced LRFS compared to G variant carriers overall (p log-rank 0.008, HR 1.88, 95% CI 1.15–3.08; Figure 1B) and in the RT alone subgroup (p log-rank 0.031, HR 2.09, 95% CI 1.04–4.21; Figure 1CLIG4 rs10131 CC genotype was associated with an unfavorable OS (p log-rank 0.020, HR 1.83, 95% CI 1.07–3.14; Figure 1D) and LRFS (p log-rank 0.038, HR 1.92, 95% CI 0.96–3.83; Figure 1E). Also, the DNA-PKcs rs7003908 A variant conferred a decreased LRFS in this subset (p log-rank 0.040, HR 1.96, 95% CI 0.94–4.05; Figure 1FNBS1 rs2735383 GG and RAD51 rs12593359 GG homozygotes treated with RT+CT showed reduced MFS (p log-rank 0.005, HR 2.85, 95% CI 1.34–6.06, and p log-rank 0.033, HR 2.15, 95% CI 1.07–4.32, respectively; Figure 1G, H) compared to other rs2735383 and rs12593359 genotype carriers. None of the associations were statistically significant after multiple testing correction.
Multivariate models adjusted for clinicopathological parameters revealed an association between 14 SNPs in 10 genes and the studied endpoints (Table 2). Only four SNPs previously identified in the univariate models (i.e. rs1042522, rs7003908, rs2735383 and rs12593359) were confirmed in this analysis, and RAD51 rs12593359 was associated with two endpoints (OS and MFS). When all patients were considered, LIG4 rs1805388 GG, MRE11A rs2155209 TT, XRCC5 rs828907 T and RAD51 rs1801321 GG carriers showed an increased risk of death (HR 1.35, p = 0.028, HR 1.36, p = 0.019, HR 1.41, p = 0.022 and HR 1.37, p = 0.016, respectively). The TP53 rs1042522 CC genotype and ATM rs1801516 A allele were associated with an elevated risk of locoregional relapse (HR 1.89, p = 0.013 and HR 1.48, p = 0.029, respectively), whereas individuals with ATM rs189037 A, XRCC6 rs2267437 CC, NBS1 rs1805787 CC and rs1805794 G were at higher risk of distant relapse (HR 2.14, p = 0.049, HR 1.89, p = 0.023, HR 1.81, p = 0.036 and HR 2.00, p = 0.020, respectively). In the subgroup given combination treatment (Table 2), the MRE11A rs2155209 TT, XRCC5 rs828907 T, RAD51 rs1801321 GG and rs12593359 GG showed an association with an increased risk of death (HR 1.54, p = 0.024, HR 1.76, p = 0.012, HR 1.58, p = 0.018 and HR 1.56, p = 0.041, respectively), while there was only one variant, DNA-PKcs rs7003908 A, that conferred a higher risk of locoregional failure (HR 2.14, p = 0.045). The XRCC6 rs2267437 CC, RAD51 rs12593359 GG, NBS1 rs1805794 GG and rs2735383 GG genotypes were associated with an elevated risk of distant relapse after RT+CT (HR 2.44, p = 0.022, HR 2.88, p = 0.004, HR 3.12, p = 0.017 and HR 3.22, p = 0.005, respectively). In patients treated with RT alone, two SNPs showed an association with the studied outcomes. The CHEK1 rs558351 TT homozygotes were at a 2.5-fold higher risk of death compared to variant C carriers (HR 2.54, p = 2x10-5), and individuals with the TP53 rs1042522 CC genotype had a more than two-fold increased risk of locoregional recurrence (HR 2.16, p = 0.047; Table 2). After the adjustment for multiple comparisons, only the effect of CHEK1 rs558351 on OS remained statistically significant.
By performing a stepwise selection procedure, 10 of the above SNPs were identified as independent predictors of OS, LRFS or MFS (Table 3). In the whole group, MRE11A rs2155209 TT, XRCC5 rs828907 T, RAD51 rs1801321 GG, LIG4 rs1805388 GG, N > 0, local and regional relapse, as well as metastasis or SPC were independent risk factors for unfavorable OS. The TP53 rs1042522 CC and ATM rs1801516 A, together with T3–4, N > 0 and non-oropharyngeal tumor localization, were independent indicators of poor LRFS, while the XRCC6 rs2267437 CC genotype, HPSCC and regional recurrence after treatment had an independent negative effect on MFS. In patients who received the combination therapy (RT+CT), MRE11A rs2155209 TT, XRCC5 rs828907 T, RAD51 rs1801321 GG, alcohol consumption, local and regional failure, and metastasis or SPC were independent indicators of poor OS, whereas RAD51 rs12593359 GG, NBS1 rs2735383 GG, non-OPSCC and regional relapse independently predicted shorter MFS. There were no independent predictors of LRFS in the RT+CT subgroup. In the RT alone subgroup, CHEK1 rs558351 TT, N > 0, local recurrence after treatment and metastasis or SPC were found to be independent risk factors for inferior OS. The TP53 rs1042522 CC genotype, T3–4, N > 0 and non-OPSCC were independently associated with reduced LRFS in these patients.
4. Discussion
There are many indications that common germline variants in genes involved in DSB response and repair may modify the efficiency of these processes and thus be of great importance for cancer susceptibility as well as the success of anticancer treatments based on DNA damage induction, such as DSBs. However, there are little data regarding the role of SNPs in these genes as predictors of therapeutic response and patient survival in HNC since previous reports have explored mainly radiation-induced toxicity or cancer risk. In this study, we hypothesized that these SNPs, by altering the levels and activity of the encoded proteins, could modulate individual sensitivity to radiation and chemoradiation, resulting in variable treatment efficacy and prognosis in HNSCC. Using multivariate analysis, we identified 14 SNPs associated with survival outcomes in HNSCC patients treated non-surgically with RT alone or with combination therapy, and 10 of which were independent indicators of unfavorable OS, LRFS or MFS in the final models. One variant, i.e. CHEK1 rs558351, survived adjusting for multiple comparisons.
In our report, MRE11A rs2155209 TT, XRCC5 rs828907 T, LIG4 rs1805388 GG and RAD51 rs1801321 GG independently predicted poor OS in the entire cohort, and rs2155209 TT, rs828907 T and rs1801321 GG were also independent risk factors for OS in the combination treatment subgroup. Possessing endonuclease and exonuclease activities, MRE11 is the core of the MRN complex, essential in early DSB recognition and signaling, cell cycle checkpoint regulation, telomere maintenance, DNA recombination, meiosis, and immune response to viral infections [6]. The data show that MRE11 may be a biomarker of response to RT and its altered expression correlates with radiosensitivity in cancer. In oral cancer, high MRE11 levels were associated with advanced stage, progression and metastasis, radio- and chemoresistance, as well as reduced OS [15]. The rs2155209 T>C SNP in the 3’ untranslated region (UTR) may influence microRNA (miRNA) binding and gene expression. It was predicted to be within miR-584, miR-744, miR-1296 and miR-296–5p binding site, and the C allele showed reduced activity in the reporter gene assay [19,20]. The C variant was also shown to increase the risk of bladder and breast cancers [21,22], while reducing colorectal cancer risk [20]. To date, no study has examined this SNP in HNC, and its prognostic significance has only been found in colorectal cancer [20]. In our HNSCC patients, it was the TT genotype that conferred an increased risk of death, which would be in line with the observation by Naccarati et al. [20] that the T variant resulted in higher MRE11A expression presumably due to less effective interactions with miRNAs.
Similarly to MRE11, XRCC5 (Ku80) functions in a protein complex and, together with XRCC6 (Ku70), participates in DSB recognition in NHEJ. Besides, XRCC6/XRCC5 dimer is involved in DDR, V(D)J recombination and telomere maintenance. In HNC, XRCC5 overexpression correlated with radioresistance, locoregional failure and mortality [14,23]. The XRCC5 rs828907 -1401G>T is located in the promoter region, what may affect the gene expression, and has been linked to increased cancer susceptibility in the Asian population. In HNC, the T allele has been identified as a risk factor for oral cancer [24] and for RT-induced subcutaneous fibrosis in OPSCC [25]. In our HNSCC patients, the T variant contributed to a shorter OS, however, to date, no studies have addressed the functional significance of this SNP or the role in cancer prognosis. Thus, our work is the first to draw attention to its possible impact on cancer survival, which may be of additional clinical relevance in HNC as the data indicate a potential predictive role of XRCC5 in immunotherapy combined with RT [26].
The ATP-dependent DNA ligase 4 is required for NHEJ and V(D)J recombination, and the LIG4 deficiency syndrome, caused by rare LIG4 gene mutations, is manifested by increased radiosensitivity, neurological abnormalities, immunodeficiency and cancer predisposition [9]. In oral cancer, high LIG4 expression has been found to predict unfavorable outcome [27]. In our HNSCC group, the LIG4 rs1805388 GG genotype was independently associated with shorter OS. The rs1805388 G>A causes a Thr to Ile exchange at codon 9 (T9I), and the Ile (A) variant has been shown to impair the ligation and adenylation activity of the protein [28]. The Ile variant was also found to be associated with high chromosomal instability in lung cancer [29] and increased DNA damage levels after occupational exposure to lead [30]. This would suggest better repair in the G (Thr) carriers, thus supporting our findings and the hypothesis that more efficient DNA damage removal may lead to worse therapy outcomes. However, other authors have reported an increased radioresistance in healthy individuals [31] and higher mRNA expression in bone marrow [32] in A allele carriers. Similarly, data on rs1805388 in relation to cancer risk and prognosis are inconsistent. In small HNC studies, it was not associated with RT toxicity, while A allele was protective against cancer [33,34]. The effect on survival has only been studied in lung cancer and, contrary to our observations, the A allele was a risk factor for progression and poor response to platinum-based CT [35].
The RAD51 recombinase is a critical player in HR pathway, that interacts with many other key proteins, including BRCA1, BRCA2, CHEK1 and p53. It is upregulated in various cancers, which has been linked to resistance to RT and anticancer drugs [36]. RAD51 overexpression has also been shown to lead to transcriptional activation of pro-metastatic genes and promoting cancer progression [37]. Elevated RAD51 levels were found in oral cancer, especially in patients with lymph node metastases, and indicated a poor prognosis [16]. The RAD51 rs1801321 -61G>T (alias -172G>T) in the 5’UTR has been demonstrated to enhance promoter activity [38]. Similarly, in silico analysis has shown that this SNP is located in the P300/CBP transcription factor binding site and the T variant may increase RAD51 expression [39]. In addition, healthy TT homozygotes had lower levels of IR-induced chromatid breaks [39], while in the GG carriers higher levels of DNA damage detected with comet assay were observed [40]. With regard to cancer risk, the T allele showed a protective effect in HNC but had no impact in other cancers [39,41]. The above data could suggest that this frequently studied functional SNP confers increased DNA damage repair capacity in T variant carriers, which may result in a poorer prognosis. However, in our report, the GG genotype was an independent predictor of reduced OS after the combination treatment and overall. Considering therapy outcomes and prognosis, no association was found between this SNP and RT toxic effects in HNC and lung cancer [25,33,42], as well as response to platinum-based CT and OS in gastric cancer [43]. Whereas, contrary to our findings, in the only study evaluating the effect of rs1801321 on HNC prognosis, the T allele was associated with poor survival in OPSCC, however, the study group was very small, limited to one subsite and the result was not confirmed in multivariate analysis [44]. Thus, further research is warranted to assess the role of RAD51 rs1801321 in HNC progression and prognosis.
Notably, the strongest association in our study, that remained statistically significant after multiple testing correction, was observed between CHEK1 rs558351 and OS. We found that the TT genotype was an independent risk factor for poor OS after RT. CHEK1 kinase is an essential molecule for DDR coordination, cell cycle regulation, cell proliferation and survival [7]. The rs558351 -664C>T is located in the 5’UTR and, according to HaploReg v4.2 [45,46], may disrupt regulatory motifs and protein binding. It has also been predicted to markedly alter RNA secondary structure [47]. This may cause a phenotypic effect as such SNPs are able to change binding affinity of proteins and miRNAs [48]. Very little is known about the role of this variant in cancer and other human diseases. It has only been examined in a large case-control study for colorectal cancer risk in the Chinese population, but no effect has been found [47]. Thus, our study is the first to indicate the potential prognostic role of this common SNP in cancer, in particular in RT treated HNSCC patients. Increased CHEK1 expression has been reported in various cancers, including HNC, and found to correlate with radioresistance and poor outcome, whereas CHEK1 inhibition has been demonstrated to have radiosensitizing effects [7,49]. One could speculate that the T variant, possibly conferring a higher CHEK1 activity, may result in limited sensitivity to RT in TT homozygotes and thus reduced OS. Our data suggest that this SNP deserves further attention and efforts are needed to explore its functional relevance, role in solid cancer progression and treatment response. Our observation also takes on more importance, since CHEK1 is considered a promising therapeutic target in HNC [7,50].
This study identified two SNPs independently related to a high risk of locoregional failure. We demonstrated that the TP53 rs1042522 CC homozygotes did not benefit from treatment with either RT alone or overall. This well-known, non-synonymous SNP (nsSNP) occurring in the proline-rich domain and resulting in G>C transversion at codon 72 (R72P, Arg72Pro) in exon 4 has been extensively functionally investigated and significant biochemical and biological differences between the variants have been found. The 72Arg variant was shown to induce apoptosis more efficiently, while the 72Pro was more effective in G1 cell cycle arrest and activating DNA repair [51,52]. The rs1042522 has been linked to the risk of various cancers and other diseases, as well as to aging. Data on HNC do not clearly confirm the relationship between rs1042522 and the development of this cancer, but some of them suggest that the 72Pro may be a susceptibility allele [53,54]. In HNC, studies on the predictive and prognostic value of this SNP are very scarce. Consistent with our results, the 72Pro allele was also associated with poor prognosis in early-stage HNSCC treated with RT [55] and in breast cancer [56]. The higher DNA repair efficiency in the 72Pro cell lines compared to the 72Arg cells [57] and lower apoptotic activity of the variant may result in worse therapy response, although it has been reported that the impact of rs1042522 on clinical outcome may depend on p53 mutation status of the tumor [58]. The second SNP linked to locoregional recurrence in our study was ATM rs1801516 causing D1835N substitution in exon 37. This nsSNP may alter the exonic splicing enhancer and regulatory motifs, but its functional consequences are unclear [59]. It has been primarily examined in the context of radiosensitivity. The 1835Asn variant was found to be related to increased RT-induced normal tissue toxicity in a meta-analysis including breast and prostate cancers [60], as well as in a small nasopharyngeal cancer study [61]. There is very little data on the prognostic impact of this SNP in cancer. In our HNSCC cohort, the A (1835Asn) allele was an independent indicator of poor LRFS, confirming previous observations showing the association of AA genotype with progression in colorectal cancer [62].
In this report, the XRCC6 rs2267437 CC, NBS1 rs2735383 GG and RAD51 rs12593359 GG genotypes independently predicted increased risk of distant failure. The rs2267437 -61C>G (alias -1310C>G) functional SNP in the XRCC6 promoter region affects the binding of transcription factors and gene expression [63,64]. In line with our results, healthy carriers of the C allele had lower DSBs induction, which may suggest lower radiosensitivity [31]. The C variant was also shown to cause higher promoter activity and mRNA levels [64]. Although no correlation was found in HNC between this SNP and cancer susceptibility or RT toxicity, meta-analyses demonstrated that the G allele may be a risk factor in other cancers [33,34,65]. Two previous studies on HNC and renal cancer also showed no association with prognosis [66,64]. Therefore, our work is probably the first to indicate that rs2267437 contributes to HNSCC progression. Furthermore, we reported that rs2735383 GG and rs12593359 GG homozygotes were more than two times as likely to develop distant relapse after combination therapy. Both SNPs are located in 3’UTRs thus may affect NBS1 and RAD51 post-transcriptional regulation by miRNAs. The rs2735383 541C>G was predicted to be within miR-499-5p, miR-508-3p, miR-629 and miR-509-5p binding sites [19,67]. The C allele was shown to decrease the gene expression and alter miR-509-5p binding in colorectal cancer [67], as well as miR-629 binding in lung cancer cells [68]. Also, the mRNA levels in CC genotype carriers were found to be lower in colorectal, lung and laryngeal cancer tissues [67 -69 ]. Thus, it could be assumed that C variant is associated with less efficient DNA repair, especially that CC homozygotes had higher cancer risk, including LSCC [69,70], and more chromatid breaks in lymphocytes [68]. Although a few previous reports on the NBS1 rs2735383 impact on survival demonstrated no correlation in bladder, breast and oropharyngeal cancers [19,71], the existing data together with our findings may suggest that GG genotype is a marker for effective DSB early response and repair, resulting in lower sensitivity to DNA damaging treatment. In turn, the functional RAD51 rs12593359 T>G was proposed to affect miR-129-3p binding, and the GG genotype correlated with lower mRNA levels in lymphoblastoid cell lines and fibroblasts, as well as with better OS in hepatocellular carcinoma [72,73]. Conversely, in our RT+CT subgroup, the GG carriers were at higher risk of metastasis. In HNC, no prior research addressed the prognostic role of rs12593359, and the only case-control study showed no association with HNSCC risk [74]. Given the correlation of RAD51 overexpression with treatment resistance and aggressive phenotype in many cancers, as well as the therapeutic potential of RAD51 inhibitors [36], subsequent studies on RAD51 genetic variants may yield clinically valuable results.
To summarize, our data show that MRE11A rs2155209 TT, XRCC5 rs828907 T, RAD51 rs1801321 GG, rs12593359 GG, LIG4 rs1805388 GG, CHEK1 rs558351 TT, TP53 rs1042522 CC, ATM rs1801516 A, XRCC6 rs2267437 CC and NBS1 rs2735383 GG independently predict unfavorable clinical outcome in HNSCC patients receiving DNA damaging therapy. In the stratified analysis, these SNPs were specific risk modifiers with effects that varied by treatment. This is also the first report demonstrating the association of MRE11A rs2155209, XRCC5 rs828907, LIG4 rs1805388, ATM rs1801516 and RAD51 rs12593359 with survival in HNSCC, as well as the first to indicate that CHEK1 rs558351 may play a role in cancer disease. Although the present work has several limitations, including a moderate sample size and the number of SNPs analyzed, our results highlight variants worth validation in larger populations and other cancer types, as well as more thorough functional studies.
5. Conclusions
One way to improve the survival of HNC patients is to better understand the host genetic factors underlying the mechanisms that modulate treatment sensitivity. Our observations suggest that common germline variants in crucial DSB response and repair genes have the potential to constitute predictive and prognostic biomarkers for use alongside conventional clinical factors in HNSCC treated with DNA damage-inducing therapies. They may also be of broader importance for anticancer therapy, as DDR inhibition or deficiency increases the efficacy of immunotherapy.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: The genotype frequency in the group, location and functionality of SNPs selected for the study.
Author Contributions
Conceptualization, D.B.; methodology, D.B., A.G.-K. and M.K.; investigation, M.K., A.G.-K., D.B. and T.R.; validation, M.K., A.G.-K. and D.B.; formal analysis and data interpretation, D.B.; resources, D.B., T.R. and K.S.; data curation, D.B. and T.R.; writing—original draft preparation, D.B.; writing—review and editing, D.B. and M.K.; visualization, D.B.; supervision and project administration, D.B.; funding acquisition, D.B. All authors have read and agreed to the published version of the manuscript.
Funding
This study was funded by the National Science Centre (NCN), Poland, grant number 2016/23/B/NZ5/03470 to D.B.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki, and approved by the Institutional Ethics Committee of Maria Skłodowska-Curie National Research Institute of Oncology, Gliwice Branch (protocol code KB/430-37/18).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
The data presented in this study are available on reasonable request from the corresponding author.
Acknowledgments
We thank Iwona Domińczyk, Zofia Kołosza and Marcin Motnyk (The Provincial Office for Cancer Registration) for help in clinical and epidemiological data acquisition, as well as Iwona Matuszczyk, Teresa Stępień and Małgorzata Żmuda for their excellent technical assistance. We are also grateful to Monika Pietrowska and Piotr Widłak for their valuable contribution to the collection of research material.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Pinkas, W.; Jankowski, M.; Wierzba, W. Awareness of Head and Neck Cancers: A 2021 Nationwide Cross-Sectional Survey in Poland. J. Clin. Med. 2022, 11, 538. [Google Scholar] [CrossRef]
- Johnson, D.E.; Burtness, B.; Leemans, C.R.; Lui, V.W.Y.; Bauman, J.E.; Grandis, J.R. Head and neck squamous cell carcinoma. Nat. Rev. Dis. Primers 2020, 6, 92. [Google Scholar] [CrossRef] [PubMed]
- Krejci, L.; Altmannova, V.; Spirek, M.; Zhao, X. Homologous recombination and its regulation. Nucleic Acids Res. 2012, 40, 5795–5818. [Google Scholar] [CrossRef] [PubMed]
- Davis, A.J.; Chen, D.J. DNA double strand break repair via non-homologous end-joining. Transl. Cancer Res. 2013, 2, 130–143. [Google Scholar] [CrossRef]
- Papalouka, C.; Adamaki, M.; Batsaki, P.; Zoumpourlis, P.; Tsintarakis, A.; Goulielmaki, M.; Fortis, S.P.; Baxevanis, C.N.; Zoumpourlis, V. DNA Damage Response Mechanisms in Head and Neck Cancer: Significant Implications for Therapy and Survival. Int. J. Mol. Sci. 2023, 24, 2760. [Google Scholar] [CrossRef] [PubMed]
- Syed, A.; Tainer, J.A. The MRE11-RAD50-NBS1 Complex Conducts the Orchestration of Damage Signaling and Outcomes to Stress in DNA Replication and Repair. Annu. Rev. Biochem. 2018, 87, 263–294. [Google Scholar] [CrossRef]
- Smith, H.L.; Southgate, H.; Tweddle, D.A.; Curtin, N.J. DNA damage checkpoint kinases in cancer. Expert Rev. Mol. Med. 2020, 22, e2. [Google Scholar] [CrossRef]
- Huen, M.S.; Sy, S.M.; Chen, J. BRCA1 and its toolbox for the maintenance of genome integrity. Nat. Rev. Mol. Cell Biol. 2010, 11, 138–148. [Google Scholar] [CrossRef]
- El Nachef, L.; Berthel, E.; Ferlazzo, M.L.; Le Reun, E.; Al-Choboq, J.; Restier-Verlet, J.; Granzotto, A.; Sonzogni, L.; Bourguignon, M.; Foray, N. Cancer and Radiosensitivity Syndromes: Is Impaired Nuclear ATM Kinase Activity the Primum Movens? Cancers 2022, 14, 6141. [Google Scholar] [CrossRef]
- Parliament, M.B.; Murray, D. Single nucleotide polymorphisms of DNA repair genes as predictors of radioresponse. Semin. Radiat. Oncol. 2010, 20, 232–240. [Google Scholar] [CrossRef]
- Wang, M.; Chu, H.; Zhang, Z.; Wei, Q. Molecular epidemiology of DNA repair gene polymorphisms and head and neck cancer. J. Biomed. Res. 2013, 27, 179–192. [Google Scholar] [CrossRef]
- Liu, Y.P.; Zheng, C.C.; Huang, Y.N.; He, M.L.; Xu, W.W.; Li, B. Molecular mechanisms of chemo- and radiotherapy resistance and the potential implications for cancer treatment. MedComm. (2020) 2021, 2, 315–340. [Google Scholar] [CrossRef]
- Yang, M.H.; Chang, S.Y.; Chiou, S.H.; Liu, C.J.; Chi, C.W.; Chen, P.M.; Teng, S.C.; Wu, K.J. Overexpression of NBS1 induces epithelial-mesenchymal transition and co-expression of NBS1 and Snail predicts metastasis of head and neck cancer. Oncogene 2007, 26, 1459–1467. [Google Scholar] [CrossRef] [PubMed]
- Moeller, B.J.; Yordy, J.S.; Williams, M.D.; Giri, U.; Raju, U.; Molkentine, D.P.; Byers, L.A.; Heymach, J.V.; Story, M.D.; Lee, J.J.; Sturgis, E.M.; Weber, R.S.; Garden, A.S.; Ang, K.K.; Schwartz, D.L. DNA repair biomarker profiling of head and neck cancer: Ku80 expression predicts locoregional failure and death following radiotherapy. Clin. Cancer Res. 2011, 17, 2035–2043. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.Y.; Chen, Y.K.; Lo, S.; Chi, T.C.; Chen, Y.H.; Hu, S.C.; Chen, Y.W.; Jiang, S.S.; Tsai, F.Y.; Liu, W.; Li, R.N.; Hsieh, Y.C.; Huang, C.J.; Yuan, S.F. MRE11 promotes oral cancer progression through RUNX2/CXCR4/AKT/FOXA2 signaling in a nuclease-independent manner. Oncogene 2021, 40, 3510–3532. [Google Scholar] [CrossRef]
- Li, Y.; Li, J.; Sun, J.; Liu, Y.; Liu, D.; Du, L.; Wang, B.; Liu, W. Expression of RAD51 and Its Clinical Impact in Oral Squamous Cell Carcinoma. Anal. Cell. Pathol. (Amst). 2020, 2020, 1827676. [Google Scholar] [CrossRef] [PubMed]
- Butkiewicz, D.; Gdowicz-Kłosok, A.; Krześniak, M.; Rutkowski, T.; Krzywon, A.; Cortez, A.J.; Domińczyk, I.; Składowski, K. Association of Genetic Variants in ANGPT/TEK and VEGF/VEGFR with Progression and Survival in Head and Neck Squamous Cell Carcinoma Treated with Radiotherapy or Radiochemotherapy. Cancers 2020, 12, 1506. [Google Scholar] [CrossRef] [PubMed]
- Ensembl Database 103. Available online: http://www.ensembl.org/ (accessed on 25 February 2021).
- Teo, M.T.; Landi, D.; Taylor, C.F.; Elliott, F.; Vaslin, L.; Cox, D.G.; Hall, J.; Landi, S.; Bishop, D.T.; Kiltie, A.E. The role of microRNA-binding site polymorphisms in DNA repair genes as risk factors for bladder cancer and breast cancer and their impact on radiotherapy outcomes. Carcinogenesis 2012, 33, 581–586. [Google Scholar] [CrossRef] [PubMed]
- Naccarati, A.; Rosa, F.; Vymetalkova, V.; Barone, E.; Jiraskova, K.; Di Gaetano, C.; Novotny, J.; Levy, M.; Vodickova, L.; Gemignani, F.; Buchler, T.; Landi, S.; Vodicka, P.; Pardini, B. Double-strand break repair and colorectal cancer: gene variants within 3’ UTRs and microRNAs binding as modulators of cancer risk and clinical outcome. Oncotarget 2016, 7, 23156–23169. [Google Scholar] [CrossRef]
- Choudhury, A.; Elliott, F.; Iles, M.M.; Churchman, M.; Bristow, R.G.; Bishop, D.T.; Kiltie, A.E. Analysis of variants in DNA damage signalling genes in bladder cancer. BMC Med. Genet. 2008, 9, 69. [Google Scholar] [CrossRef]
- Wu, Z.; Wang, P.; Song, C.; Wang, K.; Yan, R.; Li, J.; Dai, L. Evaluation of miRNA-binding-site SNPs of MRE11A, NBS1, RAD51 and RAD52 involved in HRR pathway genes and risk of breast cancer in China. Mol. Genet. Genomics 2015, 290, 1141–1153. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.W.; Kim, S.Y.; Yi, S.L.; Son, S.H.; Song, D.Y.; Moon, S.Y.; Kim, J.H.; Choi, E.K.; Ahn, S.D.; Shin, S.S.; Lee, K.K.; Lee, S.W. Expression of Ku80 correlates with sensitivities to radiation in cancer cell lines of the head and neck. Oral Oncol. 2006, 42, 979–986. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.F.; Tseng, H.C.; Chiu, C.F.; Liang, S.Y.; Tsai, C.W.; Tsai, M.H.; Bau, D.T. Association between DNA double strand break gene Ku80 polymorphisms and oral cancer susceptibility. Oral Oncol. 2009, 45, 789–793. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Mathew, D.; Bhat, S.A.; Ghoshal, S.; Pal, A. Genetic Variants of DNA Repair Genes as Predictors of Radiation-Induced Subcutaneous Fibrosis in Oropharyngeal Carcinoma. Front. Oncol. 2021, 11, 652049. [Google Scholar] [CrossRef] [PubMed]
- Kumazawa, T.; Mori, Y.; Sato, H.; Permata, T.B.M.; Uchihara, Y.; Noda, S.E.; Okada, K.; Kakoti, S.; Suzuki, K.; Ikota, H.; Yokoo, H.; Gondhowiardjo, S.; Nakano, T.; Ohno, T.; Shibata, A. Expression of non-homologous end joining factor, Ku80, is negatively correlated with PD-L1 expression in cancer cells after X-ray irradiation. Oncol. Lett. 2022, 23, 29. [Google Scholar] [CrossRef]
- Joshi, J.S.; Vora, H.H.; Ghosh, N.R.; Tankshali, R.N.; Jetly, D.H.; Trivedi, T.I. Nonhomologous end joining repair pathway molecules as predictive biomarkers for patients with oral squamous cell carcinoma. J. Cancer Res. Ther. 2021, 17, 1031–1038. [Google Scholar] [CrossRef] [PubMed]
- Girard, P.M.; Kysela, B.; Härer, C.J.; Doherty, A.J.; Jeggo, P.A. Analysis of DNA ligase IV mutations found in LIG4 syndrome patients: the impact of two linked polymorphisms. Hum. Mol. Genet. 2004, 13, 2369–2376. [Google Scholar] [CrossRef]
- Tseng, R.C.; Hsieh, F.J.; Shih, C.M.; Hsu, H.S.; Chen, C.Y.; Wang, Y.C. Lung cancer susceptibility and prognosis associated with polymorphisms in the nonhomologous end-joining pathway genes: a multiple genotype-phenotype study. Cancer 2009, 115, 2939–2948. [Google Scholar] [CrossRef]
- García-Lestón, J.; Roma-Torres, J.; Vilares, M.; Pinto, R.; Prista, J.; Teixeira, J.P.; Mayan, O.; Conde, J.; Pingarilho, M.; Gaspar, J.F.; Pásaro, E.; Méndez, J.; Laffon, B. Genotoxic effects of occupational exposure to lead and influence of polymorphisms in genes involved in lead toxicokinetics and in DNA repair. Environ. Int. 2013, 43, 29–36. [Google Scholar] [CrossRef]
- Mumbrekar, K.D.; Goutham, H.V.; Vadhiraja, B.M.; Bola Sadashiva, S.R. Polymorphisms in double strand break repair related genes influence radiosensitivity phenotype in lymphocytes from healthy individuals. DNA repair 2016, 40, 27–34. [Google Scholar] [CrossRef]
- Ribeiro, H.L., Junior; Soares Maia, A.R.; Costa, M.B.; Farias, I.R.; de Paula Borges, D.; de Oliveira, R.T.; de Sousa, J.C.; Magalhães, S.M.; Pinheiro, R.F. Influence of functional polymorphisms in DNA repair genes of myelodysplastic syndrome. Leuk. Res. 2016, 48, 62–72. [Google Scholar] [CrossRef] [PubMed]
- Venkatesh, G.H.; Manjunath, V.B.; Mumbrekar, K.D.; Negi, H.; Fernandes, D.J.; Sharan, K.; Banerjee, S.; Bola Sadashiva, S.R. Polymorphisms in radio-responsive genes and its association with acute toxicity among head and neck cancer patients. PloS One 2014, 9, e89079. [Google Scholar] [CrossRef]
- Werbrouck, J.; De Ruyck, K.; Duprez, F.; Van Eijkeren, M.; Rietzschel, E.; Bekaert, S.; Vral, A.; De Neve, W.; Thierens, H. Single-nucleotide polymorphisms in DNA double-strand break repair genes: association with head and neck cancer and interaction with tobacco use and alcohol consumption. Mutat. Res. 2008, 656, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.H.; Xu, X.L.; Ruan, H.H.; Xu, W.Z.; Li, D.; Feng, J.G.; Han, Q.B.; Mao, W.M. The impact of functional LIG4 polymorphism on platinum-based chemotherapy response and survival in non-small cell lung cancer. Med Oncol. 2014, 31, 959. [Google Scholar] [CrossRef] [PubMed]
- Demeyer, A.; Benhelli-Mokrani, H.; Chénais, B.; Weigel, P.; Fleury, F. Inhibiting homologous recombination by targeting RAD51 protein. Biochim. Biophys Acta Rev. Cancer 2021, 1876, 188597. [Google Scholar] [CrossRef]
- Wiegmans, A.P.; Al-Ejeh, F.; Chee, N.; Yap, P.Y.; Gorski, J.J.; Da Silva, L.; Bolderson, E.; Chenevix-Trench, G.; Anderson, R.; Simpson, P.T.; Lakhani, S.R.; Khanna, K.K. Rad51 supports triple negative breast cancer metastasis. Oncotarget 2014, 5, 3261–3272. [Google Scholar] [CrossRef]
- Hasselbach, L.; Haase, S.; Fischer, D.; Kolberg, H.C.; Stürzbecher, H.W. Characterisation of the promoter region of the human DNA-repair gene Rad51. Eur. J. Gynaecol. Oncol. 2005, 26, 589–598. [Google Scholar]
- Lu, J.; Wang, L.E.; Xiong, P.; Sturgis, E.M.; Spitz, M.R.; Wei, Q. 172G>T variant in the 5’ untranslated region of DNA repair gene RAD51 reduces risk of squamous cell carcinoma of the head and neck and interacts with a P53 codon 72 variant. Carcinogenesis 2007, 28, 988–994. [Google Scholar] [CrossRef]
- Goricar, K.; Erculj, N.; Zadel, M.; Dolzan, V. Genetic polymorphisms in homologous recombination repair genes in healthy Slovenian population and their influence on DNA damage. Radiol. Oncol. 2012, 46, 46–53. [Google Scholar] [CrossRef]
- Zhao, M.; Chen, P.; Dong, Y.; Zhu, X.; Zhang, X. Relationship between Rad51 G135C and G172T variants and the susceptibility to cancer: a meta-analysis involving 54 case-control studies. PloS One 2014, 9, e87259. [Google Scholar] [CrossRef]
- Yin, M.; Liao, Z.; Huang, Y.J.; Liu, Z.; Yuan, X.; Gomez, D.; Wang, L.E.; Wei, Q. Polymorphisms of homologous recombination genes and clinical outcomes of non-small cell lung cancer patients treated with definitive radiotherapy. PloS One 2011, 6, e20055. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Liu, Z.Y.; Li, C.B.; Gao, S.; Ding, L.H.; Wu, X.L.; Wang, Z.Y. Genetic polymorphisms of DNA repair pathways influence the response to chemotherapy and overall survival of gastric cancer. Tumour Biol. 2015, 36, 3017–3023. [Google Scholar] [CrossRef] [PubMed]
- Santos, E.M.; Santos, H.B.P.; de Matos, F.R.; Machado, R.A.; Coletta, R.D.; Galvão, H.C.; Freitas, R.A. Clinicopathological significance of SNPs in RAD51 and XRCC3 in oral and oropharyngeal carcinomas. Oral Dis. 2019, 25, 54–63. [Google Scholar] [CrossRef]
- HaploReg v4.2. database. Available online: https://pubs.broadinstitute.org/mammals/haploreg/haploreg.php (accessed on 02 May 2023).
- Ward, L.D.; Kellis, M. HaploReg: a resource for exploring chromatin states, conservation, and regulatory motif alterations within sets of genetically linked variants. Nucleic Acids Res. 2012, 40, D930–D934. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zhang, Y.; Chen, M.; Wang, Y.; Feng, Y.; Xu, Z.; Zhang, D.; Sun, Y.; Fu, Z. Association of genetic variants in ATR-CHEK1 and ATM-CHEK2 pathway genes with risk of colorectal cancer in a Chinese population. Oncotarget 2018, 9, 26616–26624. [Google Scholar] [CrossRef] [PubMed]
- Shatoff, E.; Bundschuh, R. Single nucleotide polymorphisms affect RNA-protein interactions at a distance through modulation of RNA secondary structures. PLoS Comput. Biol. 2020, 16, e1007852. [Google Scholar] [CrossRef] [PubMed]
- Barker, H.E.; Patel, R.; McLaughlin, M.; Schick, U.; Zaidi, S.; Nutting, C.M.; Newbold, K.L.; Bhide, S.; Harrington, K.J. CHK1 Inhibition Radiosensitizes Head and Neck Cancers to Paclitaxel-Based Chemoradiotherapy. Mol. Cancer Ther. 2016, 15, 2042–2054. [Google Scholar] [CrossRef] [PubMed]
- van Harten, A.M.; Buijze, M.; van der Mast, R.; Rooimans, M.A.; Martens-de Kemp, S.R.; Bachas, C.; Brink, A.; Stigter-van Walsum, M.; Wolthuis, R.M.F.; Brakenhoff, R.H. Targeting the cell cycle in head and neck cancer by Chk1 inhibition: a novel concept of bimodal cell death. Oncogenesis 2019, 8, 38. [Google Scholar] [CrossRef]
- Pim, D.; Banks, L. p53 polymorphic variants at codon 72 exert different effects on cell cycle progression. Int. J. Cancer 2004, 108, 196–199. [Google Scholar] [CrossRef]
- Jeong, B.S.; Hu, W.; Belyi, V.; Rabadan, R.; Levine, A.J. Differential levels of transcription of p53-regulated genes by the arginine/proline polymorphism: p53 with arginine at codon 72 favors apoptosis. FASEB J. 2010, 24, 1347–1353. [Google Scholar] [CrossRef]
- Ji, X.; Neumann, A.S.; Sturgis, E.M.; Adler-Storthz, K.; Dahlstrom, K.R.; Schiller, J.T.; Wei, Q.; Li, G. p53 codon 72 polymorphism associated with risk of human papillomavirus-associated squamous cell carcinoma of the oropharynx in never-smokers. Carcinogenesis 2008, 29, 875–879. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.J.; Niu, J.; Wei, S.; Yin, M.; Liu, Z.; Wang, L.E.; Sturgis, E.M.; Wei, Q. A novel functional DEC1 promoter polymorphism -249T>C reduces risk of squamous cell carcinoma of the head and neck. Carcinogenesis 2010, 31, 2082–2090. [Google Scholar] [CrossRef] [PubMed]
- Azad, A.K.; Bairati, I.; Samson, E.; Cheng, D.; Mirshams, M.; Qiu, X.; Savas, S.; Waldron, J.; Wang, C.; Goldstein, D.; Xu, W.; Meyer, F.; Liu, G. Validation of genetic sequence variants as prognostic factors in early-stage head and neck squamous cell cancer survival. Clin. Cancer Res. 2012, 18, 196–206. [Google Scholar] [CrossRef] [PubMed]
- Tommiska, J.; Eerola, H.; Heinonen, M.; Salonen, L.; Kaare, M.; Tallila, J.; Ristimäki, A.; von Smitten, K.; Aittomäki, K.; Heikkilä, P.; Blomqvist, C.; Nevanlinna, H. Breast cancer patients with p53 Pro72 homozygous genotype have a poorer survival. Clin. Cancer Res. 2005, 11, 5098–5103. [Google Scholar] [CrossRef]
- Siddique, M.; Sabapathy, K. Trp53-dependent DNA-repair is affected by the codon 72 polymorphism. Oncogene 2006, 25, 3489–3500. [Google Scholar] [CrossRef]
- Vivenza, D.; Monteverde, M.; Lattanzio, L.; Tonissi, F.; Astesana, V.; Denaro, N.; Comino, A.; Russi, E.; Lo Nigro, C.; Merlano, M. Correlation of TP53 and MDM2 genotypes and clinical outcome in platinum-treated head and neck cancer patients with more than 10 years’ follow-up. Int. J. Biol. Markers 2016, 31, e183–e192. [Google Scholar] [CrossRef]
- Heikkinen, K.; Rapakko, K.; Karppinen, S.M.; Erkko, H.; Nieminen, P.; Winqvist, R. Association of common ATM polymorphism with bilateral breast cancer. Int. J. Cancer 2005, 116, 69–72. [Google Scholar] [CrossRef]
- Andreassen, C.N.; Rosenstein, B.S.; Kerns, S.L.; Ostrer, H.; De Ruysscher, D.; Cesaretti, J.A.; Barnett, G.C.; Dunning, A.M.; Dorling, L.; West, C.M.L.; Burnet, N.G.; Elliott, R.; Coles, C.; Hall, E.; Fachal, L.; Vega, A.; Gómez-Caamaño, A.; Talbot, C.J.; Symonds, R.P.; De Ruyck, K.; International Radiogenomics Consortium (RgC). Individual patient data meta-analysis shows a significant association between the ATM rs1801516 SNP and toxicity after radiotherapy in 5456 breast and prostate cancer patients. Radiother. Oncol. 2016, 121, 431–439. [Google Scholar] [CrossRef]
- Alsbeih, G.; El-Sebaie, M.; Al-Rajhi, N.; Al-Harbi, N.; Al-Hadyan, K.; Al-Qahtani, S.; Alsubael, M.; Al-Shabanah, M.; Moftah, B. Among 45 variants in 11 genes, HDM2 promoter polymorphisms emerge as new candidate biomarker associated with radiation toxicity. 3 Biotech. 2014, 4, 137–148. [Google Scholar] [CrossRef]
- Kweekel, D.M.; Antonini, N.F.; Nortier, J.W.; Punt, C.J.; Gelderblom, H.; Guchelaar, H.J. Explorative study to identify novel candidate genes related to oxaliplatin efficacy and toxicity using a DNA repair array. Br. J. Cancer 2009, 101, 357–362. [Google Scholar] [CrossRef]
- Ouimet, M.; Cassart, P.; Larivière, M.; Kritikou, E.A.; Simard, J.; Sinnett, D. Functional analysis of promoter variants in KU70 and their role in cancer susceptibility. Genes Chromosomes Cancer 2012, 51, 1007–1013. [Google Scholar] [CrossRef]
- Wang, W.; Pan, X.; Huo, X.; Yan, F.; Wang, M.; Wang, D.; Gao, Y.; Cao, Q.; Luo, D.; Qin, C.; Yin, C.; Zhang, Z. A functional polymorphism C-1310G in the promoter region of Ku70/XRCC6 is associated with risk of renal cell carcinoma. Mol. Carcinog. 2012, 51 Suppl 1, E183–E190. [Google Scholar] [CrossRef]
- Jia, J.; Ren, J.; Yan, D.; Xiao, L.; Sun, R. Association between the XRCC6 polymorphisms and cancer risks: a systematic review and meta-analysis. Medicine (Baltimore) 2015, 94, e283. [Google Scholar] [CrossRef] [PubMed]
- Guberina, M.; Sak, A.; Pöttgen, C.; Tinhofer-Keilholz, I.; Budach, V.; Balermpas, P.; Von der Grün, J.; Rödel, C.M.; Gkika, E.; Grosu, A.L.; Abdollahi, A.; Debus, J.; Belka, C.; Pigorsch, S.; Combs, S. E.; Mönnich, D.; Zips, D.; De-Colle, C.; Welz, S.; Linge, A. ERCC2 gene single-nucleotide polymorphism as a prognostic factor for locally advanced head and neck carcinomas after definitive cisplatin-based radiochemotherapy. Pharmacogenomics J. 2021, 21, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Li, J.T.; Zhong, B.Y.; Xu, H.H.; Qiao, S.Y.; Wang, G.; Huang, J.; Fan, H.Z.; Zhao, H.C. Associations between NBS1 Polymorphisms and Colorectal Cancer in Chinese Population. PloS One 2015, 10, e0132332. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Li, Y.; Cheng, M.; Huang, D.; Zheng, J.; Liu, B.; Ling, X.; Li, Q.; Zhang, X.; Ji, W.; Zhou, Y.; Lu, J. A functional polymorphism at microRNA-629-binding site in the 3’-untranslated region of NBS1 gene confers an increased risk of lung cancer in Southern and Eastern Chinese population. Carcinogenesis 2012, 33, 338–347. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Liao, J.; Zhao, H.; Chen, F.; Zhu, X.; Li, J.; Nong, Q. NBS1 rs2735383 polymorphism is associated with an increased risk of laryngeal carcinoma. BMC Cancer 2018, 18, 175. [Google Scholar] [CrossRef] [PubMed]
- Gao, P.; Ma, N.; Li, M.; Tian, Q.B.; Liu, D.W. Functional variants in NBS1 and cancer risk: evidence from a meta-analysis of 60 publications with 111 individual studies. Mutagenesis 2013, 28, 683–697. [Google Scholar] [CrossRef]
- Zhu, L.; Sturgis, E.M.; Zhang, H.; Lu, Z.; Tao, Y.; Wei, Q.; Li, G. Genetic variants in microRNA-binding sites of DNA repair genes as predictors of recurrence in patients with squamous cell carcinoma of the oropharynx. Int. J. Cancer 2017, 141, 1355–1364. [Google Scholar] [CrossRef]
- Chen, F.; Zhang, H.; Pu, F. Association between a functional variant in RAD51 gene’s 3’ untranslated region and its mRNA expression in lymphoblastoid cell lines. Springerplus 2016, 5, 1688. [Google Scholar] [CrossRef] [PubMed]
- Qiu, M.; Liu, Y.; Lin, Q.; Jiang, Y.; Zhou, Z.; Wen, Q.; Liang, X.; Zhou, X.; Yu, H. A functional variant in the RAD51 3’ UTR is associated with survival of hepatocellular carcinoma patients. Gene 2013, 851, 146964. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Gao, F.; Dahlstrom, K.R.; Li, G.; Sturgis, E.M.; Zevallos, J.P.; Wei, Q.; Liu, Z. A variant at a potentially functional microRNA-binding site in BRIP1 was associated with risk of squamous cell carcinoma of the head and neck. Tumour Biol. 2016, 37, 8057–8066. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The Kaplan-Meier analysis for the studied SNPs (with p ≤ 0.05 only): (A) XRCC5 rs1051677 in relation to metastasis-free survival (MFS) and (B) TP53 rs1042522 in relation to locoregional recurrence-free survival (LRFS) in the whole group; (C) TP53 rs1042522 in relation to LRFS in the RT alone subgroup; (D) LIG4 rs10131 in relation to overall survival (OS); (E) LIG4 rs10131 and (F) DNA-PKcs rs7003908 in relation to LRFS, and (G) NBS1 rs2735383 and (H) RAD51 rs12593359 in relation to MFS in the combination treatment (RT+CT) subgroup. Number of events and n are shown in the brackets.
Figure 1.
The Kaplan-Meier analysis for the studied SNPs (with p ≤ 0.05 only): (A) XRCC5 rs1051677 in relation to metastasis-free survival (MFS) and (B) TP53 rs1042522 in relation to locoregional recurrence-free survival (LRFS) in the whole group; (C) TP53 rs1042522 in relation to LRFS in the RT alone subgroup; (D) LIG4 rs10131 in relation to overall survival (OS); (E) LIG4 rs10131 and (F) DNA-PKcs rs7003908 in relation to LRFS, and (G) NBS1 rs2735383 and (H) RAD51 rs12593359 in relation to MFS in the combination treatment (RT+CT) subgroup. Number of events and n are shown in the brackets.
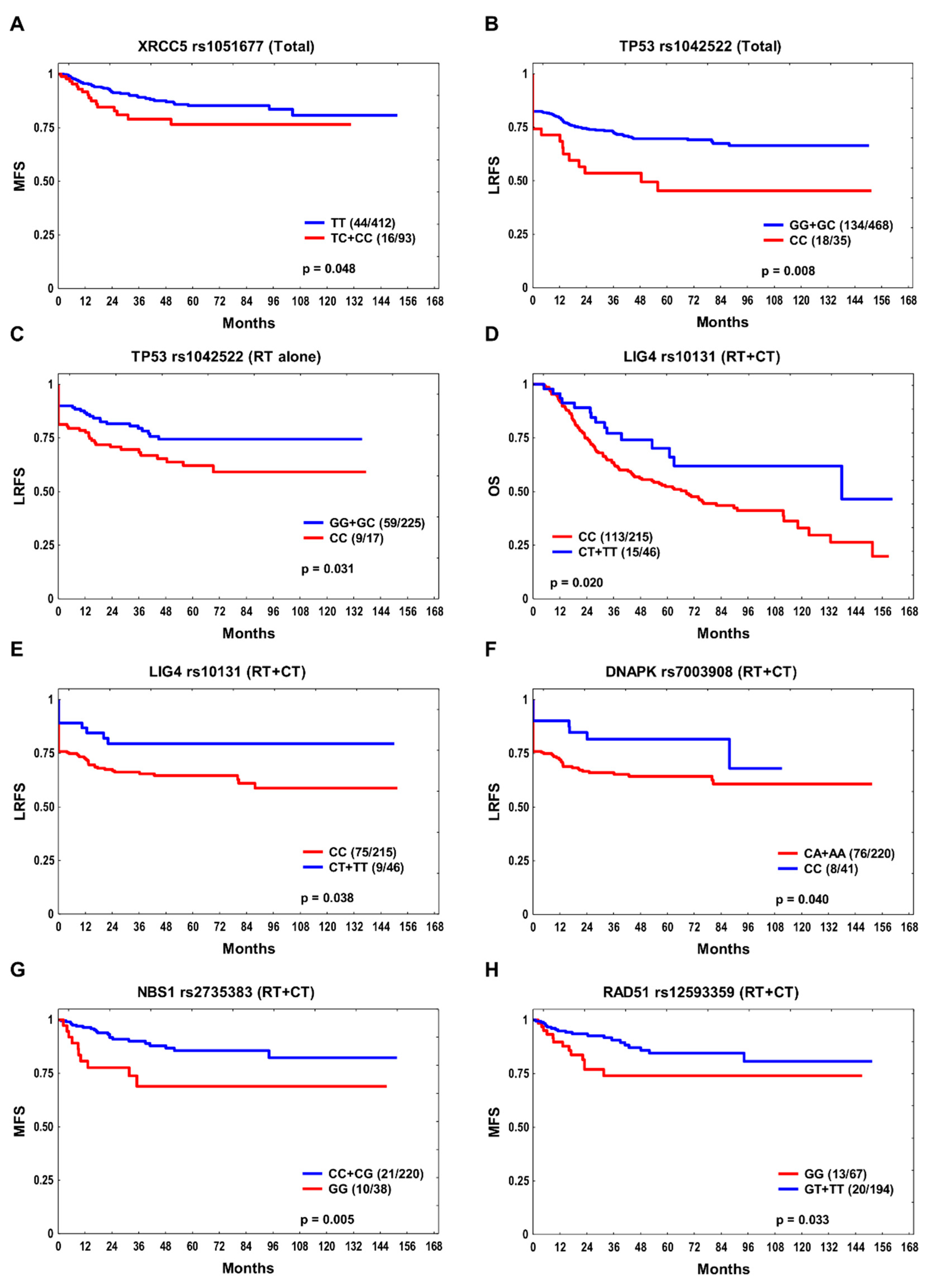

Table 1.
Study population characteristics.
| Parameter | Total n = 505 |
RT+CT n = 261 |
RT alone n = 244 |
|---|---|---|---|
| Age at diagnosis (median) < 59 years ≥ 59 years |
235 (47%) 270 (53%) |
153 (59%) 108 (41%) |
82 (34%) 162 (66%) |
| Sex Male Female |
398 (79%) 107 (21%) |
53 (20%) 208 (80%) |
54 (22%) 190 (78%) |
| Tumor site Oropharynx Hypopharynx Larynx |
212 (42%) 63 (12%) 230 (46%) |
147 (56%) 47 (18%) 67 (26%) |
65 (27%) 16 (6%) 163 (67%) |
| T stage 1–2 3–4 |
252 (50%) 253 (50%) |
81 (31%) 180 (69%) |
171 (70%) 73 (30%) |
| N stage 0 1–3 |
207 (41%) 298 (59%) |
40 (15%) 221 (85%) |
167 (68%) 77 (32%) |
| Smoking status Never Ever |
112 (22%) 393 (78%) |
61 (23%) 200 (77%) |
51 (21%) 193 (79%) |
| Alcohol consumption a Never Ever |
124 (25%) 378 (75%) |
65 (25%) 194 (75%) |
59 (24%) 184 (76%) |
RT, radiotherapy; CT, chemotherapy; RT+CT, combination treatment; a No data for three patients.
Table 2.
The association between SNPs and the studied endpoints—multivariate analysis (only SNPs with p ≤ 0.05 are shown).
Table 2.
The association between SNPs and the studied endpoints—multivariate analysis (only SNPs with p ≤ 0.05 are shown).
| Gene | SNP | Genotype | Total | RT+CT | RT alone | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Events/n | HR (95% CI) | p | Events/n | HR (95% CI) | p | Events/n | HR (95% CI) | p | |||
| OS | |||||||||||
| LIG4 | rs1805388 | GG | 165/322 | 1.35 (1.03–1.77) | 0.028 | - | - | - | - | - | - |
| MRE11A | rs2155209 | TT | 131/245 | 1.36 (1.05–1.75) | 0.019 | 72/134 | 1.54 (1.06–2.23) | 0.024 | - | - | - |
| XRCC5 | rs828907 | GT+TT | 182/354 | 1.41 (1.05–1.88) | 0.022 | 98/189 | 1.76 (1.13–2.72) | 0.012 | - | - | - |
| RAD51 | rs1801321 | GG | 105/203 | 1.37 (1.06–1.77) | 0.016 | 55/109 | 1.58 (1.08–2.30) | 0.018 | - | - | - |
| RAD51 | rs12593359 | GG | - | - | - | 33/67 | 1.56 (1.02–2.38) | 0.041 | - | - | - |
| CHEK1 | rs558351 | TT | - | - | - | - | - | - | 38/67 | 2.54 (1.66–3.90) | 2x10-5 |
| LRFS | |||||||||||
| TP53 | rs1042522 | CC | 18/35 | 1.89 (1.14–3.12) | 0.013 | - | - | - | 9/17 | 2.16 (1.01–4.62) | 0.047 |
| ATM | rs1801516 | GA+AA | 47/133 | 1.48 (1.04–2.12) | 0.029 | - | - | - | - | - | - |
| DNA-PKcs | rs7003908 | CA+AA | - | - | - | 76/220 | 2.14 (1.02–4.50) | 0.045 | - | - | - |
| MFS | |||||||||||
| ATM | rs189037 | GA+AA | 51/396 | 2.14 (1.00–4.57) | 0.049 | - | - | - | - | - | - |
| NBS1 | rs1805787 | CC | 38/281 | 1.81 (1.04–3.16) | 0.036 | - | - | - | - | - | - |
| XRCC6 | rs2267437 | CC | 22/128 | 1.89 (1.09–3.26) | 0.023 | 13/68 | 2.44 (1.14–5.26) | 0.022 | - | - | - |
| NBS1 | rs1805794 | CG+GG | 41/306 | 2.00 (1.12–3.58) | 0.020 | - | - | - | - | - | - |
| NBS1 | rs1805794 | GG | - | - | - | 7/32 | 3.12 (1.22–7.95) | 0.017 | - | - | - |
| NBS1 | rs2735383 | GG | - | - | - | 10/38 | 3.22 (1.42–7.32) | 0.005 | - | - | - |
| RAD51 | rs12593359 | GG | - | - | - | 13/67 | 2.88 (1.39–5.96) | 0.004 | - | - | - |
RT, radiotherapy; RT+CT, combination treatment; HR, hazard ratio; CI, confidence interval; OS, overall survival; LRFS, locoregional recurrence-free survival; MFS, metastasis-free survival.
Table 3.
The final models for OS, LRFS and MFS (stepwise multiple regression analysis).
| Endpoint | Variables | HR (95% CI) | p |
|---|---|---|---|
| Total | |||
| OS | N > 0 Local recurrence Regional recurrence Metastasis/SPC MRE11A rs2155209 TT XRCC5 rs828907 GT+TT LIG4 rs1805388 GG RAD51 rs1801321 GG |
1.34 (1.02–1.76) 4.43 (3.34–5.89) 1.85 (1.32–2.58) 1.97 (1.50–2.59) 1.29 (1.00–1.65) 1.36 (1.02–1.81) 1.33 (1.01–1.74) 1.32 (1.02–1.70) |
0.038 <1x10-6 0.0003 1x10-6 0.048 0.038 0.040 0.037 |
| LRFS | T3–4 N > 0 Non-OPSCC TP53 rs1042522 CC ATM rs1801516 GA+AA |
1.78 (1.25–2.54) 1.65 (1.12–2.41) 1.71 (1.20–2.44) 1.90 (1.16–3.12) 1.47 (1.04–2.09) |
0.001 0.011 0.003 0.011 0.030 |
| MFS | HPSCC Regional recurrence XRCC6 rs2267437 CC |
3.06 (1.64–5.70) 5.14 (2.94–9.02) 1.78 (1.05–3.03) |
0.0004 <1x10-6 0.032 |
| RT+CT | |||
| OS | Alcohol: ever Local recurrence Regional recurrence Metastasis/SPC MRE11A rs2155209 TT XRCC5 rs828907 GT+TT RAD51 rs1801321 GG |
2.12 (1.31–3.43) 5.36 (3.51–8.20) 1.85 (1.22–2.81) 2.37 (1.59–3.52) 1.51 (1.05–2.18) 1.67 (1.08–2.56) 1.49 (1.03–2.16) |
0.002 <1x10-6 0.004 2x10-5 0.026 0.020 0.034 |
| MFS | Non-OPSCC Regional recurrence NBS1 rs2735383 GG RAD51 rs12593359 GG |
2.13 (1.03–4.42) 5.43 (2.51–11.75) 2.74 (1.28–5.87) 2.31 (1.10–4.86) |
0.042 2x10-5 0.010 0.027 |
| RT alone | |||
| OS | N > 0 Local recurrence Metastasis/SPC CHEK1 rs558351 TT |
2.27 (1.55–3.33) 4.27 (2.88–6.31) 2.45 (1.66–3.60) 2.47 (1.63–3.77) |
3x10-5 <1x10-6 6x10-6 2x10-5 |
| LRFS | T3–4 N > 0 Non-OPSCC TP53 rs1042522 CC |
3.12 (1.87–5.21) 1.90 (1.11–3.26) 2.01 (1.09–3.68) 2.15 (1.05–4.41) |
1x10-5 0.020 0.025 0.036 |
| MFS | Regional recurrence | 6.54 (2.81–15.24) | 1x10-5 |
RT, radiotherapy; RT+CT, combination treatment; HR, hazard ratio; CI, confidence interval; OS, overall survival; LRFS, locoregional recurrence-free survival; MFS, metastasis-free survival; HPSCC, hypopharyngeal squamous cell carcinoma; Non-OPSCC, non-oropharyngeal squamous cell carcinoma; SPC, second primary cancer.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



