
Submitted:
28 September 2023
Posted:
28 September 2023
You are already at the latest version
Abstract
Rhabdomyosarcoma is a cancer arising from arrested myogenic differentiation, seen mainly in children or adolescents. Metastatic rhabdomyosarcoma is often fatal even with aggressive cytotoxic chemotherapies, surgery, and irradiation. SALIQ is an acronym for a multidrug augmentation regimen designed as an adjunct to current rhabdomyosarcoma chemotherapies. SALIQ uses five common non-oncology drugs, repurposed from general medicine use, to promote malignant clone maturation and inhibit rhabdomyosarcoma growth. The five drugs are: the cholesterol lowering drug simvastatin, the acne medicine tretinoin (ATRA), the psychiatric drug lithium carbonate, the antifungal drug itraconazole, and the food supplement quercetin. All five drugs are in common use for non-cancer conditions, are cheap, have an eminently safe side effect profile, and all five have preclinical evidence and good rationale for inhibiting rhabdomyosarcoma growth.
Keywords:
chemotherapy
; repurposed drugs
; rhabdomyosarcoma
1. Introduction
This paper introduces SALIQ, a new adjunctive multidrug regimen for treating rhabdomyosarcoma that is projected to impose a low side effect burden. SALIQ is an acronym for the regimen that uses five already marketed non-oncology drugs with well-established data showing that they can inhibit one or another of the known growth drivers active in rhabdomyosarcoma.
Rhabdomyosarcoma is mainly a childhood cancer expressing myogenic markers MyoD, myogenin, and muscle related proteins, myosin heavy chain, skeletal α-actin, and desmin [1,2,3,4]. Four basic categories are recognized - embryonal, alveolar, spindle cell/sclerosing, and pleomorphic [5]. Embryonal (e-rhabdomyosarcoma) and alveolar (a-rhabdomyosarcoma) predominate. Children presenting with metastatic rhabdomyosarcoma have a 3-year event-free survival of 30% and overall survival of 49% after standard current cytotoxic treatments [6]. Regimens from intensive chemotherapy up to high doses with stem-cell support and radiotherapy have been tested over the years to little avail [6,7,8]. Studies like these in widely metastatic rhabdomyosarcoma using intensive chemotherapy imposed significant side effect burdens. Therefore we are pursuing regimens that do not lower quality of life for these children. Examples of lower side effect burden regimens: low-dose continuous cytotoxic chemotherapy (daily without interruption), or non-genotoxic drugs active against specific molecular targets, or repurposed multidrug regimens like SALIQ.
Because rhabdomyosarcoma is a cancer of defective myocyte maturation, an overarching theme of SALIQ development was to find drugs to remove blocks to myoblast maturation in rhabdomyosarcoma cells.
Rhabdomyosarcomas are histologically similar, immunohistochemical stain positive for desmin, myogenin, and MyoD but are otherwise biologically diverse [1,3,4,6]. All forms of rhabdomyosarcoma are diseases of impaired myogenic differentiation [7,8].
The approach of SALIQ was to identify the known abnormalities blocking differentiation in rhabdomyosarcoma then looking at currently marketed drugs that have data that they might be repurposed to inhibit these identified differentiation blocks. Many different forces are active in impeding differentiation of the rhabdomyosarcoma cell. Only some of these are discussed here.
Rhabdomyosarcoma’s molecular classification is evolving [9]. a-rhabdomyosarcoma is more common in adolescents and carries a worse prognosis. Many variables influence 5 year survival after diagnosis - most prominently a worse prognosis if metastases are present. Many different treatment regimens have been reported over that last 2 decades using various combinations of cyclophosphamide, dactinomycin, doxorubicin, ifosfamide, radiotherapy, surgical resection, temozolomide, topotecan, vincristine, vinorelbine, and others. Yet a significant number of children still succumb to the disease.
The SALIQ drugs are: 1) the cholesterol lowering drug simvastatin, 2) all trans retinoic acid (ATRA), a vitamin A metabolite used topically in the treatment of acne, 3) the mood stabilizing drug lithium carbonate, 4) the antifungal drug itraconazole, and 5) the dietary supplement quercetin. Since quercetin is found in high amounts in onions, increased dietary intake of onions and dried onion powder are an additional option. How these drugs might impede rhabdomyosarcoma growth will be presented in section 3 below. Before that, in section 2, three core elements of rhabdomyosarcoma growth relevant to the SALIQ drugs´ actions are reviewed. See Table 1 for quick reference to these.
2. Some elements of rhabdomyosarcoma relevant to SALIQ
2. A. PAX and FOXO1
A fusion product of two transcription factors, PAX (Pax7 or Pax3) and FOXO1 is the result of a common chromosomal translocation found in 80% of a-rhabdomyosarcoma. Degree of PAX-FOXO1 expression is correlated with shorter survival [10]. a-rhabdomyosarcoma without this fusion protein clinically resembles e-rhabdomyosarcoma [11]. Both PAX and FOXO1 have an N-terminal DNA binding and a C-terminal transactivation domain. The PAX-FOXO1 fusion protein is more stable than PAX transcription factor alone. Both PAX3/PAX7 DNA binding and transcriptional activation of the fusion protein is greater than wild-type PAX proteins.The FOXO1 transactivation domain is responsible for interacting with basal transcriptional machinery in this fusion product, abnormally and excessively promoting transcription of PAX binding DNA sites (genes). The PAX fragment is selecting which genes are going to be transcriptionally upregulated by the FOXO1 transactivation domain [1,2]. An unhappy union.
PAX-FOXO1 is resistant to proteolytic degradation, is gene amplified, has increased transcription activity, and increased translation in a-rhabdomyosarcoma. Individually PAX and FOXO1 are retained in cytosol - and hence inactive - after phosphorylation by Akt. The fusion protein PAX-FOXO1 is resistant to Akt phosphorylation, hence transported to nucleus more readily than the individual components would be [2]. Pax-FOXO1 fusion proteins are resistant to Akt activity and therefore predominantly reside in the nucleus. Pax3-FOXO1 and Pax7-FOXO1 exhibit up to 100 fold greater transcriptional activity compared to wild type Pax3 and Pax7 proteins.
2. B. Glycogen synthase kinase 3b
PAX3-FOXO1 enhances glycogen synthase kinase 3beta (GSK3) transcription/translation. GSK3 is a serine-threonine kinase that is activated by its tyrosine-279/216 phosphorylation and inhibited by its phosphorylation on serine 9 [12]. The phosphatase PP2A removes GSK3’s inhibiting serine 9 phosphorylation, reactivating GSK3. PP2A activity is inversely correlated with the level of phosphorylated GSK3 in kainic acid excitotoxic mouse brain [13].
Parenthetical note: Inactive PP2A becomes activated in the presence of several common dopamine receptor 2 inhibiting antipsychotic medicines (haloperidol, paliperidone, penfluridol, perphenazine, thioridazine) [14]. Since both lithium [15,16] and phenothiazine class antipsychotic medicines [17,18] have extensive preclinical databases showing potential cancer growth inhibition, it seems they may work against each other in that role vis a vis GSK3 function. The combination might be best avoided until more is known.
GSK3 inhibition allows non phosphorylated beta-catenin’s nuclear maturation functions to proceed. Beta-catenin phosphorylated by GSK3 is degraded in the proteasome. This process is depicted in Figure 1. See section 5. below on lithium for details.
GSK3 overactivity contributes to the undifferentiated, proliferative phenotype in rhabdomyosarcoma [19].
2. C. MyoD, myogenin, and myogenesis
Myogenin is a gene derepressed by the functioning transcription factor MyoD. Myogenin’s protein product, myogenin, is essential for myogenic differentiation [2,20,21]. Any event blocking or hindering myogenin transcription would be predicted to contribute to maintaining rhabdomyosarcoma [22].
It is probable that e-rhabdomyosarcoma results from an earlier differentiation stage than a-rhabdomyosarcoma because a-rhabdomyosarcomas express three times more myogenin than does e-rhabdomyosarcoma.
MyoD is a muscle specific transcription factor. It can induce a differentiated nonmuscle cell to redifferentiate to a myocyte without going through a pluripotent stem cell stage but cannot do this without myogenin [21]. MyoD and myogenin exist in an amplifying feedback loop maintaining each other’s expression in normal skeletal muscle development and maturation [2,20,21]. Many MyoD cofactors (peptides, proteins, miRNAs, long noncoding RNAs, histone deacetylases, etc) are recognized, and potentially required, that allow or inhibit MyoD to act as a myocyte differentiation promoting transcription factor [23].
Rhabdomyosarcomas express MyoD but preliminary data indicate that these cofactors are preventing MyoD from derepressing maturation programs in rhabdomyosarcoma [2].
3. Simvastatin
Simvastatin is a common cholesterol lowering medicine, widely prescribed worldwide. Simvastatin and other cholesterol lowering drugs of this class, the “statins”, are 3-hydroxy-3-methyl-glutaryl-CoA (HMG-CoA) reductase inhibitors. They have a considerable preclinical database showing potential for cancer growth inhibition [25,26,27,28,29,30]. Clear clinical benefit from statin use has not yet been shown.
Currently marketed examples of statins are atorvastatin, fluvastatin, lovastatin, pitavastatin, pravastatin, rosuvastatin, and simvastatin. Pravastatin and rosuvastatin are hydrophilic, simvastatin and atorvastatin are lipophilic [31,32]. It has been the more lipophilic statins that have shown greater preclinical potential to inhibit cancer growth [25,30].
In adults, high doses of simvastatin (~80 mg/day) carry a higher risk of muscle pain or rhabdomyolysis compared with other statins [33]. This might be a plus as well as a minus when treating rhabdomyosarcoma. This higher muscle pain with simvastatin compared to that seen with other statins may indicate that simvastatin, as a lipophilic statin, attains higher muscle tissue levels than do other statins [34].
HMG-CoA reductase is the rate-limiting step in cholesterol synthesis, converting HMG-CoA to mevalonic acid. Simvastatin is highly lipophilic, metabolized by CYP3A4, and has a T½ of 2 hours although clinical suppression of HMG-CoA reductase is continuous [35].
Oddly, i) despite these preclinical observations of potential benefit of simvastatin, and 2) the generally benign nature of simvastatin, and 3) the serious threat to life from rhabdomyosarcoma, yet as of 2023 there has been no published clinical trial of simvastatin or any other HMG-CoA reductase inhibitor as treatment adjunct in rhabdomyosarcoma.
Proliferation and invasion of mature muscle cells, including those within the venous wall, are inhibited by lipophilic statins atorvastatin or simvastatin but not by hydrophilic statins like pravastatin [36].
HMG-CoA reductase statins also inhibit trans-prenyltransferases including geranylgeranyl pyrophosphate synthase resulting in depletion of geranylgeranyl diphosphate stores in muscle cells and rhabdomyosarcoma growth inhibition [37,38]. Simvastatin inhibits rhabdomyosarcoma cell line growth in vitro by depletion of geranylgeranyl diphosphate [38]. Lovastatin also inhibited in vitro proliferation of rhabdomyosarcoma cells [39].
It remains unproven which of the seven approved and marketed statins would be most effective as rhabdomyosarcoma treatment adjunct. Preclinical evidence would indicate simvastatin.
4. ATRA
ATRA (tretinoin is synonym) is a normal and essential metabolic product of retinoic acid forms (vitamin A) present in our food. ATRA is probably the primary biologically active form of the retinoids. It activates all three main retinoid receptors (RAR): RAR-alpha, RAR-gamma, and RXR. Vitamin A is an essential nutrient as animals cannot synthesize it [40]. ATRA drives differentiation in many different cell lineages and settings [41]. It is also used as a pharmaceutical driver of differentiation in pediatric neuroblastoma and pediatric acute promyelocytic leukemia [42,43,44]. ATRA has been used successfully as a differentiation inducing agent in treating selected pediatric cancers for decades [42,45,46].
Sine oculis homeobox homolog 1 (SIX1) is a transcription factor crucial in embryogenesis. SIX1 is highly expressed in rhabdomyosarcoma. SIX1 is one of the several transcription factors active in maintaining a muscle progenitor-like state in rhabdomyosarcoma. SIX1 loss induces partial differentiation of rhabdomyosarcoma cells and impedes those cells’ growth in vitro [47]. SIX1 is highly overexpressed in rhabdomyosarcoma where it forms part of the myocyte maturation block [47,48,49]. SIX1 is upregulated, forming part of the drive to pathologic growth, invasion, and metastasis across a variety of cancers [50,51,52,53]. ATRA reduced SIX1 expression in a myoblast cell line [54].
mRNA and protein of SIX1 and its transcriptional coactivators EYA1 and EYA2 are elevated in pulmonary fibrosis. The increased SIX1 here is a reparative reactivation on a basis of chronic alveolar damage - a wound repair triggered developmental program reactivation [55].
ATRA slowed proliferation, and resulted in a more differentiated myocyte phenotype in rhabdomyosarcoma cell lines but did not effect survival in a xenograft murine rhabdomyosarcoma model [56,57]. nanoM ATRA in vitro promoted nontransformed myoblast maturation as reflected by increased cell content of myogenin, myosin heavy chain, and myocyte fusion while inhibiting proliferation [58]. ATRA promoted in vitro differentiation of a patient derived a-rhabdomyosarcoma cell line [59]. Many authors using many different rhabdomyosarcoma cell lines have shown that ATRA increased differentiation signs - myotube-like giant cell formation, increased creatinine kinase, reduced proliferation, etc [60,61,62,63]. However not all rhabdomyosarcoma cell lines are equally sensitive to ATRA mediated differentiation or proliferation inhibition [64,65,66].
ATRA increases rhabdomyosarcoma cell lines’ synthesis and secretion of myogenin [67,68]. As discussed in section 2.C. above, myogenin is the sine qua non for muscle cell differentiation.
Rhabdomyosarcoma uses many autocrine growth signaling systems, synthesizing VEGF and expressing VEGF receptor 1 among them. Anti-VEGF monoclonal antibody slows rhabdomyosarcoma growth and forced expression of the PAX-FOXO1 fusion protein into non-malignant muscle cells, induced expression of VEGF receptor 1 [69,70,71]. ATRA decreased rhabdomyosarcoma cell growth and synthesis of VEGF. Adding exogenous VEGF rescued proliferation [71].
In vitro, 3 microM ATRA inhibited the hepatocyte growth factor (HGF) secretion from a glioblastoma cell line [72]. HGF as agonist for its cognate receptor c-Met is discussed below as a growth drive element in rhabdomyosarcoma.
A search for published research showed no clinical trial results of ATRA (tretinoin), isotretinoin, or any other retinoid in rhabdomyosarcoma.
5. Lithium
Lithium, as lithium carbonate, is commonly used in psychiatry to reduce wide mood swings where its primary locus of action is inhibition of GSK3 [73]. Lithium inhibits GSK-3 in a variety of clinical settings and diseases [74,75,76]. Lithium also has an extensive database on its adjunctive use in treating various cancers [77,78,79].
Experience with Li+ use in children is limited. Lithium has a narrow therapeutic index when treating psychiatric problems. In children weighing 20 kg to 30 kg, 600 mg to 1500 mg/day in divided doses would be a typical dose. Lithium plasma levels should be kept around 1 mEq/L Li+ until further experience might dictate other levels best. Severe side effects are seen as plasma levels approach or exceed 2 mEq/L necessitating regular Li+ blood level monitoring.
GSK3 is the central enzyme inhibited by Li+ that results in some inhibition of malignant growth [15,79,80]. As briefly presented in section 2.B. above, GSK3 is a serine/threonine kinase expressed in all mammalian cells. GSK3 is constitutively active and is negatively regulated by phosphorylation on serine-21 and/or serine-9, primarily by Akt, but also through growth factor stimulation of mitogen activated protein kinases (MAPKs), mTOR, protein kinase A (PKA), and protein kinase C (PKC).
Normal human plasma Li+ levels during psychiatric treatment are typically ~ 1mEq. In vitro cytotoxic LD50 to rhabdomyosarcoma cell lines was between 20 and 50 mM LiCl but in vitro colony suppression was 10% of controls at 10 mM LiCl [81]. Since 10 mM LiCl = about 2 mEq/L Li+ we might get only slight Li+ mediated rhabdomyosarcoma growth suppression.
Li+ stabilization of beta-catenin resulting in its enhanced nuclear signaling, induced embryonic stem cell differentiation in hemangioblast-like cells [82]. It is the intent of Li+ use in SALIQ to do the same in rhabdomyosarcoma cells. Post-natal myogenesis and myocyte maturation processes are accelerated by reduction of GSK3 activity [83,84]. A low-therapeutic (0.5 mEq) dose of Li promoted in vitro myoblast fusion and myogenic differentiation in a myoblast cell line [85].
PAX3-FOXO1 enhances GSK3 activity which in turn represses myogenesis activity. GSK3 repressed muscle creatine kinase promoter activity and hyperphosphorylated myogenin, rendering it nonfunctional in driving muscle differentiation in rhabdomyosarcoma cells [19]. Pharmacological inhibition of GSK3 rescues PAX3-FOXO1 repression of myogenesis in rhabdomyosarcoma cells [19]. This chain of events is schematically depicted in Figure 1.
GSK3 action is integral to inhibiting the WNT/β-catenin pathway. as depicted in Figure 1. Wnt signaling activity is a pro-differentiating force in normal muscle [86,87] as well as in rhabdomyosarcoma [7,88,89]. Wnt agonists decreased proliferation and promoted differentiation in a-rhabdomyosarcoma cells [90].
Again, despite the preclinical data showing Li+ induction of rhabdomyosarcoma maturation and growth suppression plus the wealth of clinical experience and ease of Li use in treating psychiatric illnesses, still as of 2023 there have been no published clinical trials of Li+ in rhabdomyosarcoma.
6. Itraconazole, and Hh
Itraconazole is a widely prescribed antifungal drug. Itraconazole is used in the treatment of glioblastoma as part of the CUSP9v3 protocol and is seeing a resurgence of interest in treating other cancers [91,92,93]. Itraconazole has poor solubility and erratic absorption, necessitating taking itraconazole with Coke™, lemon juice, or other low pH drink. Itraconazole is among the strongest CYP3A4 inhibitors in common use. Itraconazole 5 mg/kg/day in divided doses of an oral solution would be a common dose for fungal prophylaxis in children aged 2–18 years undergoing immunosuppressive cytotoxic cancer treatments [94].
Hedgehog signaling (Hh) is one of the crucial signaling systems directing embryogenesis, wound healing, bone marrow homeostasis and other physiological functions [95,96,97]. Itraconazole inhibits Hh function across a wide range of clinical and experimental conditions [92,93,98,99,100,101].
One could reasonably deduce that normal cells either have work around from an Hh signaling block or that itraconazole in human clinical use provides only weak Hh inhibition since even high doses of itraconazole do not result in major organ system failures.
Hh signaling engagement participates in facilitating growth and treatment resistance across a wide variety of cancers [102,103,104,105,106,107]. This is also the case in rhabdomyosarcoma [88,108,109,110,111,112,113]. Multiple interventions are being explored to add Hh signaling inhibition to current rhabdomyosarcoma treatments [110,114].
Hepatocyte growth factor (HGF) signals via its cognate receptor c-Met to stimulate growth, invasion in mature cells and is crucial during morphogenesis. HGF added to cultured rhabdomyosarcoma cells resulted in increased Hh signaling and migration while a phytochemical from ginger plants, zerumbone, decreased that HGF-Hh provoked nondirected motility [115].
7. Quercetin and onions
Quercetin is widely available as a food supplement. It is found in high amounts in a variety of vegetables, the highest amounts in onions [116]. Problems of poor solubility and strong first-pass catabolism have made human studies of quercetin’s anticancer potential difficult [116]. Formulation to improve solubility and absorption are available but are largely of untested effectiveness in achieving these goals. Quercetin is FDA certified as Generally Regarded as Safe (GRAS) and as such unregulated, making pharmacological attributes of quercetin products uncertain. Although multiple quercetin cancer growth inhibiting effects have been shown in preclinical study, none have yet been shown to do so in human disease [117,118,119,120].
A quercetin with lecithin proprietary product as Quercetin Phytosome 500 mg, gave 150 ng/ml plasma level at 6 hours in healthy humans [121]. A second study in healthy humans showed ~200 ng/ml plasma Cmax [122]. If this is typical for quercetin use in humans, blood levels are unlikely to be sufficient for significant rhabdomyosarcoma inhibition. However, worth a try given quercetin’s benignity.
Pannexin 1 expression is decreased in rhabdomyosarcoma, while increasing its levels decreased in vitro growth [123]. Quercetin exposure increased pannexin 1 transcription in rhabdomyosarcoma cells [124].
HGF and its receptor, c-Met’s tyrosine kinase overexpression contributes to suppressing differentiation in rhabdomyosarcoma [125,126,127,128,129]. Quercetin binds to the ATP binding site of c-Met preventing its in vitro activation by HGF [130]. Quercetin suppressed in vitro melanoma migration by preventing HGF activation of c-Met [131].
Quercetin inhibited HGF stimulation of c-Met in a medulloblastoma cell line with an IC50 of 12 microM [132]. Prostate cancer cells exposed to quercetin had reduced c-Met and Akt phosphorylation and a lower IC50 of doxorubicin [133]. In vitro 25 microM quercetin inhibited glioblastoma cell spheroid formation and migration but c-Met inhibition might not have mediated this [134]. Quercetin had an IC50 of 35 microM to HepG2 hepatoma cells [135]. It is questionable whether this microM concentration is clinically achievable.
c-Met receptor is expressed in myogenic precursor cells, and upon signaling stimulation by its ligand, HGF induces normal myogenic precursor stem cells’ proliferation and migration [136]. c-Met mRNA and protein expression was elevated in rhabdomyosarcoma, exogenous HGF increasing motility but not proliferation [137]. HGF/c-Met signaling contributes to rhabdomyosarcoma’s radioresistance [138]. Rhabdomyosarcoma cells can move toward differentiation with inhibition of c-Met signaling [139].
Abnormally increased c-Met is widely expressed in rhabdomyosarcoma [140,141,142]. Upregulation of c-Met activity contributes to the ability to establish rhabdomyosarcoma metastases. c-Met becomes unrepressed by the rhabdomyosarcoma fusion protein PAX-FOXO1, schematically shown in Figure 1 [143,144,145]. Stronger c-Met expression correlates to greater rhabdomyosarcoma tissue invasion [146]. Histologically, tumors with lower c-Met expression were characterized as more mature and differentiated, suggesting that c-Met expression in rhabdomyosarcoma functions as a suppressor of differentiation.
Clinical study NCT03245151 is of a multi-tyrosine kinase inhibitor, lenvatinib, in pediatric cancers including rhabdomyosarcoma. It is underway but results as of this writing in mid-2023 are unknown. Higher HGF/c-Met signaling activity is associated with lenvatinib resistance in adult cancers [147]. This would potentially make quercetin plus lenvatinib a particularly attractive combination for rhabdomyosarcoma. Lenvatinib blood levels, and hence side effects, can be expected to increase in people on strong CYP3A4 inhibitors like itraconazole [148]. These side effects can often be treated by simple dose reduction [148].
Studies in humans show few side effects from up to 2000 mg quercetin/day. Based on that lack, clinical utility might be doubtful but the preclinical studies showing potential quercetin benefits plus the rarity of side effects make trying quercetin reasonable.
Onions contain high levels of quercetin but absorption from that source is uncertain. Increasing onion intake per individual taste would be a harmless intervention.
8. Pharmacological considerations
The estimated rank order of side effect risk of the SALIQ drugs is, lowest to highest, quercetin, simvastatin, itraconazole, lithium carbonate, ATRA. As a conservative approach, treatment initiation should be in that order and at lower doses than their target doses. Quercetin and simvastatin can be started together on day one. ATRA, Li+ and itraconazole are best started one at a time at weekly intervals. Thus after four weeks a patient will be on all five SALIQ drugs at a low starting dose, suggested doses given in Table 3. Then bring doses up to target dose in the same order. As always in these kinds of multidrug regimens, requirement for dose reductions per individual tolerance can be expected.
8. A. Simvastatin
Studies of pediatric simvastatin use in treating hyperlipidemia showed a side effect profile no different from that of placebo [149]. Being a CYP3A4 substrate, simvastatin is technically contraindicated with itraconazole. But given the likely outcome of metastatic rhabdomyosarcoma after several failed chemotherapies, a careful try of the combination with itraconazole could be considered. There are no published reports of simvastatin overdose in children or adults.
8. B. ATRA
Many potential ATRA side effects have been reported, some serious, but fatalities are rare. Alanine aminotransferase or aspartate aminotransferase elevation or headache would be the most common side effects from ATRA [150]. ATRA is strongly teratogenic so birth control must be in place as applicable.
8. C. Lithium carbonate
Li+ is usually given as lithium carbonate. Blood Li+ levels are universally available and must be done regularly during Li+ treatment. Lithium carbonate treatment of children is not common because pediatric onset manic-depression is not common. But we do have experience in pediatric lithium carbonate use that shows good tolerability in pediatric populations [151]. Thyroid function must be monitored due to Li+ potential to inhibit thyroid function.
8. D. Itraconazole
Since itraconazole is only absorbed in low pH conditions, it must be given with an acidic beverage such as lemon juice, Coke™, or similar. Proton pump inhibitors must be avoided [152,153]. Blood levels are readily available as a standard lab test. About 4% of people on chronic itraconazole will develop a reversible elevation of hepatic function transaminases, and 1 in 9000 develop structural liver damage.
Many drugs are formally contraindicated with itraconazole due to itraconazole’s strong CYP3A4 inhibition. Specifically ATRA and simvastatin are metabolized by CYP3A4 so use is formally contraindicated with itraconazole due to potential for increased blood levels. Many of the drugs listed as contraindicated can be safely used at lower doses than usual with appropriate monitoring and dose adjustment as needed. As with most of our current cytotoxic drugs the risk of serious or fatal side effects must be weighed against the potential benefits and the likelihood of death from the cancer. Greatly complicating this decision process is the unknown degree of benefit - if any - from the SALIQ adjuvant regimen. In clinical practice the combination itraconazole together with statins, including simvastatin, is used with appropriate monitoring and dose reductions [153].
8. E. Quercetin
This is predicted to be the most benign of the SALIQ drugs. There are no known side effects in human studies.
8. Discussion and Conclusions
As amply argued elsewhere, many in the oncology community believe that adding one or two drugs will not potently inhibit a currently deadly metastatic tumor like rhabdomyosarcoma [154,155,156,157,158]. We can not expect any one or two drugs added to current cytotoxic chemotherapies to have strong disease inhibiting effects. Aggressively growing tumors have many workarounds to any one or two growth drive blocks. For now, multidrug regimens will be required. We therefore turn to multidrug combination cancer treatments to achieve fractional cell killing or fractional growth inhibition through drug additivity, as in Palmer et al [156]. And as in Kilmister et al “...effective treatment of cancer may require a multi-target strategy with multi-step inhibition of signaling pathways...in lieu of the long-standing pursuit of a ‘silver-bullet’ single-target approach” [158]. And as Lindsey et al say specifically for osteosarcoma, it “continues rapidly modifying its genotype, thus making potential targeted molecular therapeutics increasingly impractical” [159].
SALIQ also follows a military principle of breaking a crucial enemy supply chain in more than one place. Both ATRA and itraconazole inhibit Hh signaling but at two different points along the Hh signaling chain [160].
There exists more growth driving systems in rhabdomyosarcoma than the ones discussed in this paper. Dozens of cofactors interact with the growth driving systems discussed here. Some of these interacting cofactors enhance, others inhibit, function of those systems. SALIQ takes aim at inhibiting some of the more important physiological growth promoting systems. There are, however, others.
The in vitro SALIQ drug levels showing rhabdomyosarcoma growth inhibition or differentiation promotion might be higher than those attainable in humans. This potential problem might be overcome as in the works of Chow et al and of Hu et al. These authors showed that using four different antihypertensive medicines each with different mechanism of action at doses too low to be effective individually, became effective when combined all four in a single tablet [161,162]. Table 4 lists the quarter doses, usual doses, and primary mechanism of action of the four antihypertension medicines combined in the quadpill of Chow et al [161]. Should the same principle hold in rhabdomyosarcoma SALIQ could be effective even if the predicted human tumor tissue levels would be lower than in vitro effective drug levels.
A caveat: when embarking on any untested treatment regimen like SALIQ, meticulous following by the treating physician is essential.
- Frequent meetings in person with the treating physician for a standard review of systems and targeted physical exam. Weekly is a minimum during the uptitration establishment phase unless experience with a given regimen allows longer evaluation intervals.
- Frequent blood test monitoring for bone marrow, liver and kidney function.
- Starting with low doses, adding one drug at a time, reevaluating after several days then increasing dose as tolerated every few days until target dose met or side effects dictate a lower dose.
- Patients must have 24/7 ready direct access to the treating physician by phone.
- Careful attention required to assure patient is taking medicines as directed. People often have trouble keeping these multidrug regimens and the frequent dose changes straight.
- Once stability of dose of all drugs and tolerability established, monthly meetings will suffice.
- When these conditions cannot be met, use of multidrug regimens like SALIQ or CUSP9v3 is not recommended until wider experiences with them can determine more suited monitoring recommendations.
These seven recommendations all stem from a basic principle of combat, whether it is fencing, chess, go, or actual warfare. General and warfare theorist Carl von Clausewitz (1780-1831) expressed it as “Every plan which enters too much into the detail of the course of the combat is therefore faulty and ruinous, for detail does not depend merely on general grounds, but on other particulars which it is impossible to know beforehand.” General and later President of the USA Dwight D. Eisenhower (1890-1969) said it this way: “In preparing for battle I have always found that plans are useless, but planning is indispensable.”. These military aphorisms express rules for effective practice of medicine too and are particularly applicable for SALIQ use.
The five drugs of SALIQ might not be the ideal drugs to inhibit the relevant growth and treatment resistance pathways active in rhabdomyosarcoma, but these are the drugs we have today in 2023. Given the likely unfavorable outcome once a rhabdomyosarcoma has widely metastasized plus the expected benign nature of adding the SALIQ drugs to standard treatments, the risk/potential benefit is well worth pilot study.
Figure 1.
Schematic showing relationship between pathologic fusion protein PAX-FOXO1 and several intracellular physiological changes consequent to that. See text for details and references. Omitted are many cofactors that interact with the processes shown here, some of which enhance, some of which inhibit a given reaction. Intermediate steps in these reactions are also omitted from the schematic. Dotted arrows indicate processes inhibited by the indicated drug.
Figure 1.
Schematic showing relationship between pathologic fusion protein PAX-FOXO1 and several intracellular physiological changes consequent to that. See text for details and references. Omitted are many cofactors that interact with the processes shown here, some of which enhance, some of which inhibit a given reaction. Intermediate steps in these reactions are also omitted from the schematic. Dotted arrows indicate processes inhibited by the indicated drug.
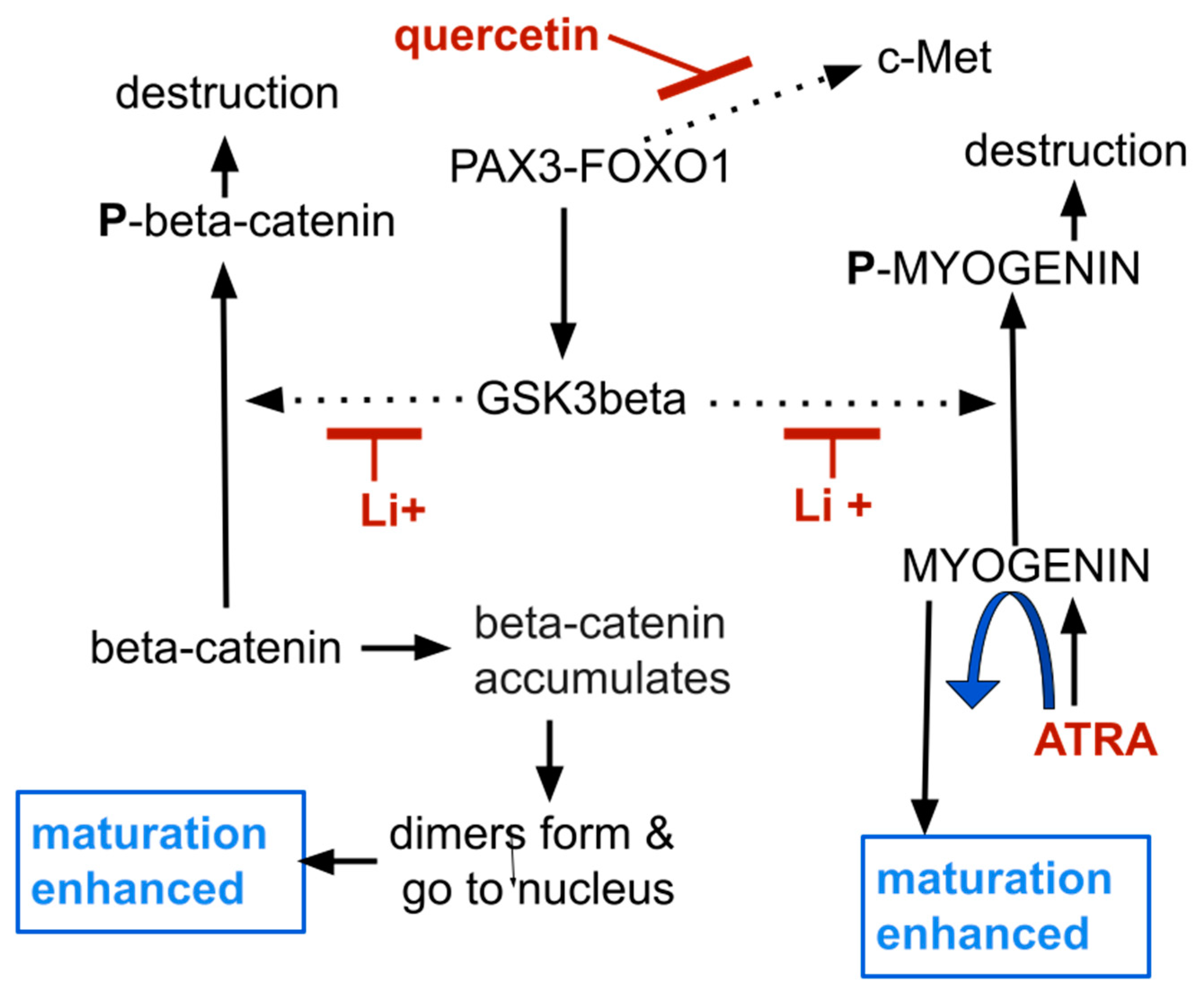

Table 1.
List of some rhabdomyosarcoma cells’ physiological elements relevant to the SALIQ regimen.
| c-MET | a tyrosine kinase stimulated by HGF, its transcription enhanced by Pax3-FOXO1 |
| FOXO1 | transcription factor directing transcription of target genes |
| GGD | geranylgeranyl diphosphate |
| GSK3 | glycogen synthase kinase 1 beta, a major inhibitor of myogenesis function |
| HGF | hepatocyte growth factor, the cognate ligand of c-Met |
| Hh | Hedgehog signaling |
| HMG-CoA reductase | 3-hydroxy-3-methyl-glutaryl-coenzyme A reductase, a statin, the target enzyme simvastatin inhibits |
| MyoD | transcription factor for myocyte differentiation |
| myogenesis | transcription factor necessary for myocyte maturation. |
| Pannexin 1 | an ion channel with lower function in rhabdomyosarcoma |
| PAX | paired box transcription factor, PAX3 or PAX7, protein transcription factor |
| PAX-FOXO1 | PAX3-FOXO1 or PAX7-FOXO1, common abnormal fusion protein driver in rhabdomyosarcoma |
Table 2.
Overview of the SALIQ drugs. APL, acute promyelocytic leukemia; BSA, body surface area; ped, pediatric.
Table 2.
Overview of the SALIQ drugs. APL, acute promyelocytic leukemia; BSA, body surface area; ped, pediatric.
| drug | ped dose | comments |
|---|---|---|
| simvastatin | 10 to 40 mg/d | 0.1% rhabdomyolysis risk over 4 years, CYP3A4 metabolism |
| ATRA | 7 mg/m2/BSA bid | is 30% of standard ATRA ped APL dose, CYP3A4 metabolism |
| lithium carbonate | 300-600 mg/d | little experience with Li+ in children, renal elimination |
| itraconazole | 5 mg/kg/day | strong CYP3A4 inhibition |
| quercetin | 1 to 2 g/d | potentially ineffective |
Table 3.
Suggested starting order, doses, and uptitration schedule. * This cannot be predicted due to strong CYP3A4 inhibition by itraconazole. Doses can be titrated by careful monitoring. ** Blood monitoring required, dose adjusted to target blood level of 1 mEq/L. Recommended starting order - quercetin, simvastatin, itraconazole, lithium carbonate, ATRA. Note that simvastatin with itraconazole is contraindicated due to potential for increased simvastatin levels. These doses are for older children and teenagers. Dose reductions to tolerability can be expected.
Table 3.
Suggested starting order, doses, and uptitration schedule. * This cannot be predicted due to strong CYP3A4 inhibition by itraconazole. Doses can be titrated by careful monitoring. ** Blood monitoring required, dose adjusted to target blood level of 1 mEq/L. Recommended starting order - quercetin, simvastatin, itraconazole, lithium carbonate, ATRA. Note that simvastatin with itraconazole is contraindicated due to potential for increased simvastatin levels. These doses are for older children and teenagers. Dose reductions to tolerability can be expected.
| drug | starting dose | target dose |
|---|---|---|
| simvastatin | 10 mg/day | 20 to 40 mg/day |
| ATRA | 7 mg/m2/BSA bid | cannot be predicted * |
| lithium carbonate | 300 mg/day | 600 mg/day ** |
| itraconazole | 2.5 mg/kg/day | 5 mg/kg/day |
| quercetin | O.5 g/day | 2 g/day |
Table 4.
Data from Chow et al, ref. 161 showing quadpill dose with percent of common human dose in treating hypertension. ARB, angiotensin receptor blocker. MOA = primary mechanism of blood pressure lowering action.
Table 4.
Data from Chow et al, ref. 161 showing quadpill dose with percent of common human dose in treating hypertension. ARB, angiotensin receptor blocker. MOA = primary mechanism of blood pressure lowering action.
| drug | quadpill dose/d (%) | Common dose/d | MOA |
|---|---|---|---|
| irbesartan | 37·5 mg (25%) | 150 mg | ARB |
| amlodipine | 1·25 mg (25%) | 5 mg | Ca++ channel blocker |
| indapamide | 0·625 mg (25%) | 2.5 mg | thiazide diuretic |
| bisoprolol | 2·5 mg (25%) | 10 mg | beta blocker |
Abbreviations
All trans retinoic acid (ATRA); glycogen synthase kinase 3beta (GSK3); hepatocyte growth factor (HGF); 3-hydroxy-3-methyl-glutaryl-CoA (HMG-CoA); retinoid receptors (RAR); Sine oculis homeobox homolog 1 (SIX1)
References
- Hettmer, S.; Linardic, C.M.; Kelsey, A.; Rudzinski, E.R.; Vokuhl, C.; Selfe, J.; Ruhen, O.; Shern, J.F.; Khan, J.; Kovach, A.R.; et al. Molecular testing of rhabdomyosarcoma in clinical trials to improve risk stratification and outcome: A consensus view from European paediatric Soft tissue sarcoma Study Group, Children’s Oncology Group and Cooperative Weichteilsarkom-Studiengruppe. Eur. J. Cancer 2022, 172, 367–386. [Google Scholar] [CrossRef] [PubMed]
- Keller, C.; Guttridge, D.C. Mechanisms of impaired differentiation in rhabdomyosarcoma. FEBS J. 2013, 280, 4323–4334. [Google Scholar] [CrossRef] [PubMed]
- Melcón, S.G.; Codina, J.S.d.T. Molecular biology of rhabdomyosarcoma. Clin. Transl. Oncol. 2007, 9, 415–419. [Google Scholar] [CrossRef] [PubMed]
- Xia, S.J.; Pressey, J.G.; Barr, F.G. Molecular pathogenesis of rhabdomyosarcoma. Cancer Biol. Ther. 2002, 1, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Leiner, J.; Le Loarer, F. The current landscape of rhabdomyosarcomas: an update. Virchows Arch. 2019, 476, 97–108. [Google Scholar] [CrossRef] [PubMed]
- Parham, D.M.; Giannikopoulos, P. Rhabdomyosarcoma: From Obscurity to Clarity in Diagnosis … But With Ongoing Challenges in Management: The Farber-Landing Lecture of 2020. Pediatr. Dev. Pathol. 2021, 24, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Yu, P.Y.; Guttridge, D.C. Dysregulated Myogenesis in Rhabdomyosarcoma. Curr. Top. Dev. Biol. 2017, 126, 285–297. [Google Scholar] [CrossRef]
- Pomella, S.; Danielli, S.G.; Alaggio, R.; Breunis, W.B.; Hamed, E.; Selfe, J.; Wachtel, M.; Walters, Z.S.; Schäfer, B.W.; Rota, R.; et al. Genomic and Epigenetic Changes Drive Aberrant Skeletal Muscle Differentiation in Rhabdomyosarcoma. Cancers 2023, 15, 2823. [Google Scholar] [CrossRef]
- Agaram, N.P. Evolving classification of rhabdomyosarcoma. Histopathology 2021, 80, 98–108. [Google Scholar] [CrossRef]
- Skapek, S.X.; Anderson, J.; Barr, F.G.; Bridge, J.A.; Gastier-Foster, J.M.; Parham, D.M.; Rudzinski, E.R.; Triche, T.; Hawkins, D.S. PAX-FOXO1 fusion status drives unfavorable outcome for children with rhabdomyosarcoma: A children’s oncology group report. Pediatr. Blood Cancer 2013, 60, 1411–1417. [Google Scholar] [CrossRef]
- Williamson, D.; Missiaglia, E.; Pritchard-Jones, K.; Oberlin, O.; Shipley, J.M.; Delattre, O.; De Reyniès, A.; Pierron, G.; Thuille, B.; Palenzuela, G.; et al. Fusion Gene-Negative Alveolar Rhabdomyosarcoma is Clinically and Molecularly Indistinguishable from Embryonal Rhabdomyosarcoma. J. Clin. Oncol. 2010, 28, 2151–2158. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yang, R.; Gu, J.; Yin, X.; Jin, N.; Xie, S.; Wang, Y.; Chang, H.; Qian, W.; Shi, J.; et al. Cross talk between PI3K-AKT-GSK-3β and PP2A pathways determines tau hyperphosphorylation. Neurobiol. Aging 2015, 36, 188–200. [Google Scholar] [CrossRef]
- Chu, D.; Tan, J.; Xie, S.; Jin, N.; Yin, X.; Gong, C.-X.; Iqbal, K.; Liu, F. GSK-3β is Dephosphorylated by PP2A in a Leu309 Methylation-Independent Manner. J. Alzheimer’s Dis. 2015, 49, 365–375. [Google Scholar] [CrossRef]
- O’connor, C.M.; Perl, A.; Leonard, D.; Sangodkar, J.; Narla, G. Therapeutic targeting of PP2A. Int. J. Biochem. Cell Biol. 2018, 96, 182–193. [Google Scholar] [CrossRef] [PubMed]
- Snitow, M.E.; Bhansali, R.S.; Klein, P.S. Lithium and Therapeutic Targeting of GSK-3. Cells 2021, 10, 255. [Google Scholar] [CrossRef] [PubMed]
- Villegas-Vázquez, E.Y.; Quintas-Granados, L.I.; Cortés, H.; Carmen, M.G.-D.; Leyva-Gómez, G.; Rodríguez-Morales, M.; Bustamante-Montes, L.P.; Silva-Adaya, D.; Pérez-Plasencia, C.; Jacobo-Herrera, N.; et al. Lithium: A Promising Anticancer Agent. Life 2023, 13, 537. [Google Scholar] [CrossRef] [PubMed]
- Manoharan, G.B.; Okutachi, S.; Abankwa, D. Potential of phenothiazines to synergistically block calmodulin and reactivate PP2A in cancer cells. PLOS ONE 2022, 17, e0268635. [Google Scholar] [CrossRef]
- Chien, W.; Sun, Q.-Y.; Lee, K.L.; Ding, L.-W.; Wuensche, P.; Torres-Fernandez, L.A.; Tan, S.Z.; Tokatly, I.; Zaiden, N.; Poellinger, L.; et al. Activation of protein phosphatase 2A tumor suppressor as potential treatment of pancreatic cancer. Mol. Oncol. 2015, 9, 889–905. [Google Scholar] [CrossRef] [PubMed]
- Dionyssiou, M.G.; Ehyai, S.; Avrutin, E.; Connor, M.K.; McDermott, J.C. Glycogen synthase kinase 3β represses MYOGENIN function in alveolar rhabdomyosarcoma. Cell Death Dis. 2014, 5, e1094. [Google Scholar] [CrossRef]
- Myer, A.; Olson, E.N.; Klein, W.H. MyoD Cannot Compensate for the Absence of Myogenin during Skeletal Muscle Differentiation in Murine Embryonic Stem Cells. Dev. Biol. 2001, 229, 340–350. [Google Scholar] [CrossRef]
- Adhikari, A.; Kim, W.; Davie, J. Myogenin is required for assembly of the transcription machinery on muscle genes during skeletal muscle differentiation. PLOS ONE 2021, 16, e0245618. [Google Scholar] [CrossRef] [PubMed]
- Sebire, N.J.; Malone, M. Myogenin and MyoD1 expression in paediatric rhabdomyosarcomas. J. Clin. Pathol. 2003, 56, 412–416. [Google Scholar] [CrossRef] [PubMed]
- Battistelli, C.; Garbo, S.; Maione, R. MyoD-Induced Trans-Differentiation: A Paradigm for Dissecting the Molecular Mechanisms of Cell Commitment, Differentiation and Reprogramming. Cells 2022, 11, 3435. [Google Scholar] [CrossRef] [PubMed]
- Calhabeu, F.; Hayashi, S.; E Morgan, J.; Relaix, F.; Zammit, P.S. Alveolar rhabdomyosarcoma-associated proteins PAX3/FOXO1A and PAX7/FOXO1A suppress the transcriptional activity of MyoD-target genes in muscle stem cells. Oncogene 2012, 32, 651–662. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Hu, J.-W.; He, X.-R.; Jin, W.-L.; He, X.-Y. Statins: a repurposed drug to fight cancer. J. Exp. Clin. Cancer Res. 2021, 40, 241. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.-H.; Liu, C.-H.; Ding, D.-C. Statins as Repurposed Drugs in Gynecological Cancer: A Review. Int. J. Mol. Sci. 2022, 23, 13937. [Google Scholar] [CrossRef] [PubMed]
- Zhu, P.-F.; Wang, M.-X.; Chen, Z.-L.; Yang, L. Targeting the Tumor Microenvironment: A Literature Review of the Novel Anti-Tumor Mechanism of Statins. Front. Oncol. 2021, 11. [Google Scholar] [CrossRef]
- Xie, L.; Zhu, G.; Shang, J.; Chen, X.; Zhang, C.; Ji, X.; Zhang, Q.; Wei, Y. An overview on the biological activity and anti-cancer mechanism of lovastatin. Cell. Signal. 2021, 87, 110122. [Google Scholar] [CrossRef]
- Duarte, J.A.; de Barros, A.L.B.; Leite, E.A. The potential use of simvastatin for cancer treatment: A review. Biomed. Pharmacother. 2021, 141, 111858. [Google Scholar] [CrossRef]
- Joharatnam-Hogan, N.; Alexandre, L.; Yarmolinsky, J.; Lake, B.; Capps, N.; Martin, R.M.; Ring, A.; Cafferty, F.; E Langley, R. Statins as Potential Chemoprevention or Therapeutic Agents in Cancer: a Model for Evaluating Repurposed Drugs. Curr. Oncol. Rep. 2021, 23, 29. [Google Scholar] [CrossRef]
- Hamelin, B.A.; Turgeon, J. Hydrophilicity/lipophilicity: relevance for the pharmacology and clinical effects of HMG-CoA reductase inhibitors. Trends Pharmacol. Sci. 1998, 19, 26–37. [Google Scholar] [CrossRef] [PubMed]
- Vuu, Y.M.; Shahib, A.K.; Rastegar, M. The Potential Therapeutic Application of Simvastatin for Brain Complications and Mechanisms of Action. Pharmaceuticals 2023, 16, 914. [Google Scholar] [CrossRef] [PubMed]
- Backes, J.M.; A Howard, P.; Ruisinger, J.F.; Moriarty, P.M. Does Simvastatin Cause More Myotoxicity Compared with Other Statins? Ann. Pharmacother. 2009, 43, 2012–2020. [Google Scholar] [CrossRef] [PubMed]
- Jamal, S.M.; Eisenberg, M.J.; Christopoulos, S. Rhabdomyolysis associated with hydroxymethylglutaryl-coenzyme A reductase inhibitors. Am. Hear. J. 2004, 147, 956–965. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, T.R.; A Tobert, J. Simvastatin: a review. Expert Opin. Pharmacother. 2004, 5, 2583–2596. [Google Scholar] [CrossRef] [PubMed]
- Turner, N.A.; Midgley, L.; O’Regan, D.J.; E Porter, K. Comparison of the Efficacies of Five Different Statins on Inhibition of Human Saphenous Vein Smooth Muscle Cell Proliferation and Invasion. J. Cardiovasc. Pharmacol. 2007, 50, 458–461. [Google Scholar] [CrossRef]
- Araki, M.; Motojima, K. Hydrophobic statins induce autophagy in cultured human rhabdomyosarcoma cells. Biochem. Biophys. Res. Commun. 2008, 367, 462–467. [Google Scholar] [CrossRef] [PubMed]
- Araki, M.; Maeda, M.; Motojima, K. Hydrophobic statins induce autophagy and cell death in human rhabdomyosarcoma cells by depleting geranylgeranyl diphosphate. Eur. J. Pharmacol. 2012, 674, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Codenotti, S.; Zizioli, D.; Mignani, L.; Rezzola, S.; Tabellini, G.; Parolini, S.; Giacomini, A.; Asperti, M.; Poli, M.; Mandracchia, D.; et al. Hyperactive Akt1 Signaling Increases Tumor Progression and DNA Repair in Embryonal Rhabdomyosarcoma RD Line and Confers Susceptibility to Glycolysis and Mevalonate Pathway Inhibitors. Cells 2022, 11, 2859. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.; Qiao, G.; Liu, Y.; Tian, L.; Hui, N.; Li, J.; Ma, Y.; Li, H.; Zhao, Q.; Cao, W.; et al. Overview of all-trans-retinoic acid (ATRA) and its analogues: Structures, activities, and mechanisms in acute promyelocytic leukaemia. Eur. J. Med. Chem. 2021, 220, 113451. [Google Scholar] [CrossRef]
- Brown, G. Retinoic acid receptor regulation of decision-making for cell differentiation. Front. Cell Dev. Biol. 2023, 11, 1182204. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, C.; Matthay, K.K.; Villablanca, J.G.; Maurer, B.J. Retinoid therapy of high-risk neuroblastoma. Cancer Lett. 2003, 197, 185–192. [Google Scholar] [CrossRef]
- Giuli, M.V.; Hanieh, P.N.; Giuliani, E.; Rinaldi, F.; Marianecci, C.; Screpanti, I.; Checquolo, S.; Carafa, M. Current Trends in ATRA Delivery for Cancer Therapy. Pharmaceutics 2020, 12, 707. [Google Scholar] [CrossRef] [PubMed]
- Masetti, R.; Biagi, C.; Zama, D.; Vendemini, F.; Martoni, A.; Morello, W.; Gasperini, P.; Pession, A. Retinoids in Pediatric Onco-Hematology: the Model of Acute Promyelocytic Leukemia and Neuroblastoma. Adv. Ther. 2012, 29, 747–762. [Google Scholar] [CrossRef] [PubMed]
- Ni, X.; Hu, G.; Cai, X. The success and the challenge of all trans retinoic acid in the treatment of cancer. Crit Rev Food Sci Nutr. 2019, 59, S71–S80. [Google Scholar] [CrossRef]
- Calleja, E.M.; Warrell, R.P. Differentiating agents in pediatric malignancies: All-trans-retinoic acid and arsenic in acute promyelocytic leukemia. Curr. Oncol. Rep. 2000, 2, 519–523. [Google Scholar] [CrossRef]
- Hsu, J.Y.; Danis, E.P.; Nance, S.; O’Brien, J.H.; Gustafson, A.L.; Wessells, V.M.; Goodspeed, A.E.; Talbot, J.C.; Amacher, S.L.; Jedlicka, P.; et al. SIX1 reprograms myogenic transcription factors to maintain the rhabdomyosarcoma undifferentiated state. Cell Rep. 2022, 38, 110323–110323. [Google Scholar] [CrossRef]
- Yu, Y.; Khan, J.; Khanna, C.; Helman, L.; Meltzer, P.S.; Merlino, G. Expression profiling identifies the cytoskeletal organizer ezrin and the developmental homeoprotein Six-1 as key metastatic regulators. Nat. Med. 2004, 10, 175–181. [Google Scholar] [CrossRef]
- Ehinger, D.; Frostberg, H.; Larsson, S.; Gisselsson, D. SIX1 as a Novel Immunohistochemical Marker in the Differential Diagnosis of Rhabdomyosarcoma. Fetal Pediatr. Pathol. 2023, 42, 1–12. [Google Scholar] [CrossRef]
- Zhu, G.; Liu, Y.; Zhao, L.; Lin, Z.; Piao, Y. The Significance of SIX1 as a Prognostic Biomarker for Survival Outcome in Various Cancer Patients: A Systematic Review and Meta-Analysis. Front. Oncol. 2021, 11, 622331. [Google Scholar] [CrossRef]
- Huang, S.; Lin, W.; Wang, L.; Gao, Y.; Yuan, X.; Zhang, P.; Chen, Y.; Chu, Q. SIX1 Predicts Poor Prognosis and Facilitates the Progression of Non-small Lung Cancer via Activating the Notch Signaling Pathway. J. Cancer 2022, 13, 527–540. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Zhao, Q.; Yang, X.; Wang, T.; Yuan, S.; Meng, Q. SIX1: A Prognostic Biomarker in Uterine Corpus Endometrial Carcinoma. Comb. Chem. High Throughput Screen. 2023, 26, 330–338. [Google Scholar] [CrossRef] [PubMed]
- Zou, J.; Huang, Y.; Chen, Y.; Wu, Z.; Xie, H.; Zhou, H.; Xing, C. FOXC2-induced circCASK aggravates colorectal cancer progression by upregulating SIX1 expression. IUBMB Life 2023, 75, 659–672. [Google Scholar] [CrossRef] [PubMed]
- Adachi, Y.; Masuda, M.; Sakakibara, I.; Uchida, T.; Niida, Y.; Mori, Y.; Kamei, Y.; Okumura, Y.; Ohminami, H.; Ohnishi, K.; et al. All-trans retinoic acid changes muscle fiber type via increasing GADD34 dependent on MAPK signal. Life Sci. Alliance 2022, 5, e202101345. [Google Scholar] [CrossRef] [PubMed]
- Wilson, C.; Mertens, T.C.; Shivshankar, P.; Bi, W.; Collum, S.D.; Wareing, N.; Ko, J.; Weng, T.; Naikawadi, R.P.; Wolters, P.J.; et al. Sine oculis homeobox homolog 1 plays a critical role in pulmonary fibrosis. J. Clin. Investig. 2022, 7, e142984. [Google Scholar] [CrossRef] [PubMed]
- Ohi, S. Characterization, anticancer drug susceptibility and atRA-induced growth inhibition of a novel cell line (HUMEMS) established from pleural effusion of alveolar rhabdomyosarcoma of breast tissue. Hum. Cell 2007, 20, 39–51. [Google Scholar] [CrossRef]
- Al-Tahan, A.; Sarkis, O.; Harajly, M.; Bs, O.K.B.; Zibara, K.; Boulos, F.; Dighe, D.; Kregel, S.; Bazarbachi, A.; El-Sabban, M.; et al. Retinoic acid fails to induce cell cycle arrest with myogenic differentiation in rhabdomyosarcoma. Pediatr. Blood Cancer 2011, 58, 877–884. [Google Scholar] [CrossRef]
- Li, Q.; Zhang, T.; Zhang, R.; Qin, X.; Zhao, J. All-trans retinoic acid regulates sheep primary myoblast proliferation and differentiation in vitro. Domest. Anim. Endocrinol. 2019, 71, 106394. [Google Scholar] [CrossRef]
- Ogose, A.; Motoyama, T.; Watanabe, H.; Hotta, T. In vitro differentiation and proliferation in a newly established human rhabdomyosarcoma cell line. Virchows Arch. 1995, 426, 385–391. [Google Scholar] [CrossRef]
- Ramp, U.; Gerharz, C.D.; Doehmer, J.; Oster, O.; Gabbert, H.E. Uniform response of c-raf expression to differentiation induction and inhibition of proliferation in a rat rhabdomyosarcoma cell line. Virchows Arch. B Cell Pathol. Incl. Mol. Pathol. 1990, 59, 271–280. [Google Scholar] [CrossRef]
- Gerharz, C.D.; Bracke, M.E.; Mareel, M.M.; Gabbert, H.E. Modulation of invasive potential in different clonal subpopulations of a rat rhabdomyosarcoma cell line (BA-HAN-1) by differentiation induction. Clin. Exp. Metastasis 1993, 11, 55–67. [Google Scholar] [CrossRef] [PubMed]
- Górski, G.K.; Donaldson, M.H.; McMorrow, L.E. Synergistic inhibition of human rhabdomyosarcoma cells by sodium phenylacetate and tretinoin. Vitr. Cell. Dev. Biol. - Anim. 1993, 29, 189–191. [Google Scholar] [CrossRef] [PubMed]
- Gabbert, H.E.; Gerharz, C.D.; Biesalski, H.K.; Engers, R.; Luley, C. Terminal differentiation and growth inhibition of a rat rhabdomyosarcoma cell line (BA-HAN-1C) in vitro after exposure to retinoic acid. Cancer Res. 1988, 48, 5264–5269. [Google Scholar] [PubMed]
- Ricaud, S.; Vernus, B.; Bonnieu, A. Response of human rhabdomyosarcoma cell lines to retinoic acid: Relationship with induction of differentiation and retinoic acid sensitivity. Exp. Cell Res. 2005, 311, 192–204. [Google Scholar] [CrossRef] [PubMed]
- Barlow, J.W.; Wiley, J.C.; Mous, M.; Narendran, A.; Gee, M.F.; Goldberg, M.; Sexsmith, E.; Malkin, D. Differentiation of rhabdomyosarcoma cell lines using retinoic acid. Pediatr. Blood Cancer 2005, 47, 773–784. [Google Scholar] [CrossRef] [PubMed]
- Palomares, T.; Castro, B.; del Olmo, M.; Iglesias, A.; Bilbao, P.; Alonso-Varona, A. Influence of the level of γ-glutamyltranspeptidase activity on the response of poorly and moderately differentiated rhabdomyosarcoma cell lines to all-trans-retinoic acid. Anti-Cancer Drugs 2006, 17, 1127–1139. [Google Scholar] [CrossRef] [PubMed]
- Arnold, H.H.; Gerharz, C.D.; E Gabbert, H.; Salminen, A. Retinoic acid induces myogenin synthesis and myogenic differentiation in the rat rhabdomyosarcoma cell line BA-Han-1C. J. Cell Biol. 1992, 118, 877–887. [Google Scholar] [CrossRef] [PubMed]
- Arnold, H.H.; Braun, T.; Bober, E.; Buchberger, A.; Winter, B.; Salminen, A. Regulation of myogenin expression in normal and transformed myogenic cell lines. Symp Soc Exp Biol. 1992, 46, 37–51. [Google Scholar]
- Crouch, G.D.; Helman, L.J. All-trans-retinoic acid inhibits the growth of human rhabdomyosarcoma cell lines. Cancer Res. 1991, 51, 4882–4887. [Google Scholar]
- Miyoshi, K.; Kohashi, K.; Fushimi, F.; Yamamoto, H.; Kishimoto, J.; Taguchi, T.; Iwamoto, Y.; Oda, Y. Close correlation between CXCR4 and VEGF expression and frequent CXCR7 expression in rhabdomyosarcoma. Hum. Pathol. 2014, 45, 1900–1909. [Google Scholar] [CrossRef]
- Gee, M.F.W.; Tsuchida, R.; Eichler-Jonsson, C.; Das, B.; Baruchel, S.; Malkin, D. Vascular endothelial growth factor acts in an autocrine manner in rhabdomyosarcoma cell lines and can be inhibited with all-trans-retinoic acid. Oncogene 2005, 24, 8025–8037. [Google Scholar] [CrossRef] [PubMed]
- Chattopadhyay, N.; Butters, R.R.; Brown, E.M. Agonists of the retinoic acid- and retinoid X-receptors inhibit hepatocyte growth factor secretion and expression in U87 human astrocytoma cells. Mol. Brain Res. 2001, 87, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Dandekar, M.P.; Valvassori, S.S.; Dal-Pont, G.C.; Quevedo, J. Glycogen Synthase Kinase-3β as a Putative Therapeutic Target for Bipolar Disorder. Curr. Drug Metab. 2018, 19, 663–673. [Google Scholar] [CrossRef] [PubMed]
- Malhi, G.S.; Outhred, T. Therapeutic Mechanisms of Lithium in Bipolar Disorder: Recent Advances and Current Understanding. CNS Drugs 2016, 30, 931–949. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Singh, A.; Kumar, T.; Kumar, T.; Velagala, V.R.; Velagala, V.R.; Thakre, S.; Thakre, S.; Joshi, A.; Joshi, A. The Actions of Lithium on Glaucoma and Other Senile Neurodegenerative Diseases Through GSK-3 Inhibition: A Narrative Review. Cureus 2022, 14, e28265. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, D.; Beaulieu, J.M. Inhibition of glycogen synthase kinase 3 by lithium, a mechanism in search of specificity. Front. Mol. Neurosci. 2022, 15, 1028963. [Google Scholar] [CrossRef]
- Bilir, A.; Aynacioglu, A.S.; Tuna, M.Y. The Possible Interactions and Therapeutic Roles of Lithium Chloride and Midkine on Cancer Treatment. Crit. Rev. Oncog. 2019, 24, 35–45. [Google Scholar] [CrossRef]
- Yang, C.; Zhu, B.; Zhan, M.; Hua, Z.-C. Lithium in Cancer Therapy: Friend or Foe? Cancers 2023, 15, 1095. [Google Scholar] [CrossRef]
- Neofytou, C.; Backlund, A.; Blomgren, K.; Hermanson, O. Irradiation and lithium treatment alter the global DNA methylation pattern and gene expression underlying a shift from gliogenesis towards neurogenesis in human neural progenitors. Transl. Psychiatry 2023, 13, 258. [Google Scholar] [CrossRef]
- Natale, G.; Fini, E.; Calabrò, P.F.; Carli, M.; Scarselli, M.; Bocci, G. Valproate and lithium: Old drugs for new pharmacological approaches in brain tumors? Cancer Lett. 2023, 560, 216125. [Google Scholar] [CrossRef]
- Schleicher, S.B.; Zaborski, J.J.; Riester, R.; Zenkner, N.; Handgretinger, R.; Kluba, T.; Traub, F.; Boehme, K.A. Combined application of arsenic trioxide and lithium chloride augments viability reduction and apoptosis induction in human rhabdomyosarcoma cell lines. PLOS ONE 2017, 12, e0178857. [Google Scholar] [CrossRef] [PubMed]
- Mnatsakanyan, H.; Salmeron-Sanchez, M.; Rico, P. Lithium Directs Embryonic Stem Cell Differentiation Into Hemangioblast-Like Cells. Adv. Biol. 2021, 5, 2000569. [Google Scholar] [CrossRef] [PubMed]
- Pansters, N.A.; Schols, A.M.; Verhees, K.J.; de Theije, C.C.; Snepvangers, F.J.; Kelders, M.C.; Ubags, N.D.; Haegens, A.; Langen, R.C. Muscle-specific GSK-3β ablation accelerates regeneration of disuse-atrophied skeletal muscle. Biochim. et Biophys. Acta (BBA) - Mol. Basis Dis. 2015, 1852, 490–506. [Google Scholar] [CrossRef] [PubMed]
- Theeuwes, W.; Gosker, H.; Langen, R.; Pansters, N.; Schols, A.; Remels, A. Inactivation of glycogen synthase kinase 3β (GSK-3β) enhances mitochondrial biogenesis during myogenesis. Biochim. et Biophys. Acta (BBA) - Mol. Basis Dis. 2018, 1864, 2913–2926. [Google Scholar] [CrossRef]
- Kurgan, N.; Whitley, K.C.; Maddalena, L.A.; Moradi, F.; Stoikos, J.; Hamstra, S.I.; Rubie, E.A.; Kumar, M.; Roy, B.D.; Woodgett, J.R.; et al. A Low-Therapeutic Dose of Lithium Inhibits GSK3 and Enhances Myoblast Fusion in C2C12 Cells. Cells 2019, 8, 1340. [Google Scholar] [CrossRef]
- Girardi, F.; Le Grand, F. Wnt Signaling in Skeletal Muscle Development and Regeneration. Prog Mol Biol Transl Sci. 2018, 153, 157–179. [Google Scholar] [CrossRef]
- De-Paula, V.J.; dos Santos, C.C.C.; Luque, M.C.A.; Ali, T.M.; Kalil, J.E.; Forlenza, O.V.; Cunha-Neto, E. Acute and chronic lithium treatment increases Wnt/β-catenin transcripts in cortical and hippocampal tissue at therapeutic concentrations in mice. Metab. Brain Dis. 2020, 36, 193–197. [Google Scholar] [CrossRef]
- Ramadan, F.; Fahs, A.; Ghayad, S.E.; Saab, R. Signaling pathways in Rhabdomyosarcoma invasion and metastasis. Cancer Metastasis Rev. 2020, 39, 287–301. [Google Scholar] [CrossRef] [PubMed]
- Chen, E. Wnt Signaling in Rhabdomyosarcoma - A Potential Targeted Therapy Option. Curr. Drug Targets 2016, 17, 1245–1251. [Google Scholar] [CrossRef]
- Annavarapu, S.R.; Cialfi, S.; Dominici, C.; Kokai, G.K.; Uccini, S.; Ceccarelli, S.; McDowell, H.P.; Helliwell, T.R. Characterization of Wnt/β-catenin signaling in rhabdomyosarcoma. Lab. Investig. 2013, 93, 1090–1099. [Google Scholar] [CrossRef]
- Halatsch, M.-E.; Kast, R.E.; Karpel-Massler, G.; Mayer, B.; Zolk, O.; Schmitz, B.; Scheuerle, A.; Maier, L.; Bullinger, L.; Mayer-Steinacker, R.; et al. A phase Ib/IIa trial of 9 repurposed drugs combined with temozolomide for the treatment of recurrent glioblastoma: CUSP9v3. Neuro-Oncol. Adv. 2021, 3, vdab075. [Google Scholar] [CrossRef] [PubMed]
- Li, C.-L.; Fang, Z.-X.; Wu, Z.; Hou, Y.-Y.; Wu, H.-T.; Liu, J. Repurposed itraconazole for use in the treatment of malignancies as a promising therapeutic strategy. Biomed. Pharmacother. 2022, 154, 113616. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Liu, W.; Wang, J.Q.; Tang, Z. “Hedgehog pathway”: a potential target of itraconazole in the treatment of cancer. J. Cancer Res. Clin. Oncol. 2020, 146, 297–304. [Google Scholar] [CrossRef] [PubMed]
- Bury, D.; Tissing, W.J.E.; Muilwijk, E.W.; Wolfs, T.F.W.; Brüggemann, R.J. Clinical Pharmacokinetics of Triazoles in Pediatric Patients. Clin. Pharmacokinet. 2021, 60, 1103–1147. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.T.H.; Zhao, Z.; Ingham, P.W. Hedgehog signalling. Development 2016, 143, 367–372. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Cui, B.; Li, X.; Zhao, X.; Huang, T.; Ding, X. The emerging roles of Hedgehog signaling in tumor immune microenvironment. Front. Oncol. 2023, 13, 1171418. [Google Scholar] [CrossRef]
- Zhang, Y.; Beachy, P.A. Cellular and molecular mechanisms of Hedgehog signalling. Nat. Rev. Mol. Cell Biol. 2023, 24, 668–687. [Google Scholar] [CrossRef]
- Ban, L.; Mei, T.; Su, Q.; Li, W.; Huang, Z.; Liu, L.; Wu, Y.; Lv, S.; Wang, A.; Li, S. Anti-fungal drug itraconazole exerts anti-cancer effects in oral squamous cell carcinoma via suppressing Hedgehog pathway. Life Sci. 2020, 254, 117695. [Google Scholar] [CrossRef]
- Deng, H.; Huang, L.; Liao, Z.; Liu, M.; Li, Q.; Xu, R. Itraconazole inhibits the Hedgehog signaling pathway thereby inducing autophagy-mediated apoptosis of colon cancer cells. Cell Death Dis. 2020, 11, 539. [Google Scholar] [CrossRef]
- Gerber, D.E.; Putnam, W.C.; Fattah, F.J.; Kernstine, K.H.; Brekken, R.A.; Pedrosa, I.; Skelton, R.; Saltarski, J.M.; Lenkinski, R.E.; Leff, R.D.; et al. Concentration-dependent Early Antivascular and Antitumor Effects of Itraconazole in Non–Small Cell Lung Cancer. Clin. Cancer Res. 2020, 26, 6017–6027. [Google Scholar] [CrossRef]
- Freitas, R.D.; Dias, R.B.; Vidal, M.T.A.; Valverde, L.d.F.; Costa, R.G.A.; Damasceno, A.K.A.; Sales, C.B.S.; Rocha, L.d.O.S.d.; dos Reis, M.G.; Soares, M.B.P.; et al. Inhibition of CAL27 Oral Squamous Carcinoma Cell by Targeting Hedgehog Pathway With Vismodegib or Itraconazole. Front. Oncol. 2020, 10, 563838. [Google Scholar] [CrossRef] [PubMed]
- Xia, R.; Xu, M.; Yang, J.; Ma, X. The role of Hedgehog and Notch signaling pathway in cancer. Mol. Biomed. 2022, 3, 44. [Google Scholar] [CrossRef] [PubMed]
- Martelli, A.M.; Paganelli, F.; Truocchio, S.; Palumbo, C.; Chiarini, F.; McCubrey, J.A. Understanding the Roles of the Hedgehog Signaling Pathway during T-Cell Lymphopoiesis and in T-Cell Acute Lymphoblastic Leukemia (T-ALL). Int. J. Mol. Sci. 2023, 24, 2962. [Google Scholar] [CrossRef] [PubMed]
- Suchors, C.; Kim, J. Canonical Hedgehog Pathway and Noncanonical GLI Transcription Factor Activation in Cancer. Cells 2022, 11, 2523. [Google Scholar] [CrossRef]
- Tesanovic, S.; Krenn, P.W.; Aberger, F. Hedgehog/GLI signaling in hematopoietic development and acute myeloid leukemia—From bench to bedside. Front. Cell Dev. Biol. 2022, 10, 944760. [Google Scholar] [CrossRef] [PubMed]
- Meister, M.T.; Boedicker, C.; Linder, B.; Kögel, D.; Klingebiel, T.; Fulda, S. Concomitant targeting of Hedgehog signaling and MCL-1 synergistically induces cell death in Hedgehog-driven cancer cells. Cancer Lett. 2019, 465, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Lai, Q.; Wang, D.; Pei, J.; Tian, B.; Gao, Y.; Gao, Z.; Xu, X. Hedgehog signaling regulates the development and treatment of glioblastoma (Review). Oncol. Lett. 2022, 24, 294. [Google Scholar] [CrossRef]
- Yoon, J.W.; Lamm, M.; Chandler, C.; Iannaccone, P.; Walterhouse, D. Up-regulation of GLI1 in vincristine-resistant rhabdomyosarcoma and Ewing sarcoma. BMC Cancer 2020, 20, 511. [Google Scholar] [CrossRef]
- Zarzosa, P.; Garcia-Gilabert, L.; Hladun, R.; Guillén, G.; Gallo-Oller, G.; Pons, G.; Sansa-Girona, J.; Segura, M.F.; de Toledo, J.S.; Moreno, L.; et al. Targeting the Hedgehog Pathway in Rhabdomyosarcoma. Cancers 2023, 15, 727. [Google Scholar] [CrossRef]
- Manzella, G.; Schäfer, B.W. Interfering with Hedgehog Pathway: New Avenues for Targeted Therapy in Rhabdomyosarcoma. Curr. Drug Targets 2016, 17, 1228–1234. [Google Scholar] [CrossRef]
- Chelsky, Z.L.; Paulson, V.A.; Chen, E.Y. Molecular analysis of 10 pleomorphic rhabdomyosarcomas reveals potential prognostic markers and druggable targets. Genes, Chromosom. Cancer 2021, 61, 138–147. [Google Scholar] [CrossRef] [PubMed]
- Drummond, C.J.; Hanna, J.A.; Garcia, M.R.; Devine, D.J.; Heyrana, A.J.; Finkelstein, D.; Rehg, J.E.; Hatley, M.E. Hedgehog Pathway Drives Fusion-Negative Rhabdomyosarcoma Initiated From Non-myogenic Endothelial Progenitors. Cancer Cell 2018, 33, 108–124. [Google Scholar] [CrossRef] [PubMed]
- Almazán-Moga, A.; Zarzosa, P.; Molist, C.; Velasco, P.; Pyczek, J.; Simon-Keller, K.; Giralt, I.; Vidal, I.; Navarro, N.; Segura, M.F.; et al. Ligand-dependent Hedgehog pathway activation in Rhabdomyosarcoma: the oncogenic role of the ligands. Br. J. Cancer 2017, 117, 1314–1325. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Lin, W.; Li, C.; Ueki, H.; Xue, R.; Sadahira, T.; Hu, H.; Wada, K.; Li, N.; Liu, C.; et al. Repurposing of posaconazole as a hedgehog/SMO signaling inhibitor for embryonal rhabdomyosarcoma therapy. 2021, 11, 4528–4540.
- Urla, C.; Stagno, M.J.; Fuchs, J.; Warmann, S.W.; Schmid, E. Anticancer bioactivity of zerumbone on pediatric rhabdomyosarcoma cells. J. Cancer Res. Clin. Oncol. 2022, 149, 3313–3323. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Fu, K.; Wang, C.; Ma, C.; Gong, L.; Zhou, H.; Xue, X.; Peng, C.; Li, Y. Protective effects of dietary quercetin on cerebral ischemic injury: pharmacology, pharmacokinetics and bioavailability-enhancing nanoformulations. Food Funct. 2023, 14, 4470–4489. [Google Scholar] [CrossRef] [PubMed]
- Lotfi, N.; Yousefi, Z.; Golabi, M.; Khalilian, P.; Ghezelbash, B.; Montazeri, M.; Shams, M.H.; Baghbadorani, P.Z.; Eskandari, N. The potential anti-cancer effects of quercetin on blood, prostate and lung cancers: An update. Front. Immunol. 2023, 14, 1077531. [Google Scholar] [CrossRef]
- Shabir, I.; Pandey, V.K.; Shams, R.; Dar, A.H.; Dash, K.K.; Khan, S.A.; Bashir, I.; Jeevarathinam, G.; Rusu, A.V.; Esatbeyoglu, T.; et al. Promising bioactive properties of quercetin for potential food applications and health benefits: A review. Front. Nutr. 2022, 9, 999752. [Google Scholar] [CrossRef]
- Sethi, G.; Rath, P.; Chauhan, A.; Ranjan, A.; Choudhary, R.; Ramniwas, S.; Sak, K.; Aggarwal, D.; Rani, I.; Tuli, H.S. Apoptotic Mechanisms of Quercetin in Liver Cancer: Recent Trends and Advancements. Pharmaceutics 2023, 15, 712. [Google Scholar] [CrossRef]
- Maugeri, A.; Calderaro, A.; Patanè, G.T.; Navarra, M.; Barreca, D.; Cirmi, S.; Felice, M.R. Targets Involved in the Anti-Cancer Activity of Quercetin in Breast, Colorectal and Liver Neoplasms. Int. J. Mol. Sci. 2023, 24, 2952. [Google Scholar] [CrossRef]
- Riva, A.; Ronchi, M.; Petrangolini, G.; Bosisio, S.; Allegrini, P. Improved Oral Absorption of Quercetin from Quercetin Phytosome®, a New Delivery System Based on Food Grade Lecithin. Eur. J. Drug Metab. Pharmacokinet. 2019, 44, 169–177. [Google Scholar] [CrossRef]
- Solnier, J.; Chang, C.; Roh, K.; Du, M.; Kuo, Y.C.; Hardy, M.; Lyon, M.; Gahler, R. Quercetin LipoMicel—A Novel Delivery System to Enhance Bioavailability of Quercetin. J Nat Health Prod Res. 2021, 3, 1–8. [Google Scholar] [CrossRef]
- Xiang, X.; Langlois, S.; St-Pierre, M.-E.; Barré, J.F.; Grynspan, D.; Purgina, B.; Cowan, K.N. Pannexin 1 inhibits rhabdomyosarcoma progression through a mechanism independent of its canonical channel function. Oncogenesis 2018, 7, 89. [Google Scholar] [CrossRef] [PubMed]
- Xiang, X.; Hoang, H.-D.; Gilchrist, V.H.; Langlois, S.; Alain, T.; Cowan, K.N. Quercetin induces pannexin 1 expression via an alternative transcript with a translationally active 5′ leader in rhabdomyosarcoma. Oncogenesis 2022, 11, 9. [Google Scholar] [CrossRef] [PubMed]
- Skrzypek, K.; Kusienicka, A.; Szewczyk, B.; Adamus, T.; Lukasiewicz, E.; Miekus, K.; Majka, M. Constitutive activation of MET signaling impairs myogenic differentiation of rhabdomyosarcoma and promotes its development and progression. Oncotarget 2015, 6, 31378–31398. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Wang, Y.; Meng, L.; Liu, Y.; Pang, Y.; Cui, W.; Zhang, L.; Li, Z.; Liu, Q.; Shang, H.; et al. c-MET expression potentially contributes to the poor prognosis of rhabdomyosarcoma. Int J Clin Exp Pathol. 2018, 11, 4083–4092. [Google Scholar]
- Taulli, R.; Scuoppo, C.; Bersani, F.; Accornero, P.; Forni, P.E.; Miretti, S.; Grinza, A.; Allegra, P.; Schmitt-Ney, M.; Crepaldi, T.; et al. Validation of Met as a Therapeutic Target in Alveolar and Embryonal Rhabdomyosarcoma. Cancer Res 2006, 66, 4742–4749. [Google Scholar] [CrossRef] [PubMed]
- Rees, H.; Williamson, D.; Papanastasiou, A.; Jina, N.; Nabarro, S.; Shipley, J.; Anderson, J. The MET receptor tyrosine kinase contributes to invasive tumour growth in rhabdomyosarcomas. Growth Factors 2006, 24, 197–208. [Google Scholar] [CrossRef]
- Perrone, C.; Pomella, S.; Cassandri, M.; Pezzella, M.; Milano, G.M.; Colletti, M.; Cossetti, C.; Pericoli, G.; Di Giannatale, A.; de Billy, E.; et al. MET Inhibition Sensitizes Rhabdomyosarcoma Cells to NOTCH Signaling Suppression. Front. Oncol. 2022, 12, 835642. [Google Scholar] [CrossRef] [PubMed]
- Baby, B.; Antony, P.; Vijayan, R. Interactions of quercetin with receptor tyrosine kinases associated with human lung carcinoma. Nat. Prod. Res. 2017, 32, 2928–2931. [Google Scholar] [CrossRef]
- Cao, H.-H.; Cheng, C.-Y.; Su, T.; Fu, X.-Q.; Guo, H.; Li, T.; Tse, A.K.-W.; Kwan, H.-Y.; Yu, H.; Yu, Z.-L. Quercetin inhibits HGF/c-Met signaling and HGF-stimulated melanoma cell migration and invasion. Mol. Cancer 2015, 14, 103. [Google Scholar] [CrossRef]
- Labbé, D.; Provençal, M.; Lamy, S.; Boivin, D.; Gingras, D.; Béliveau, R. The Flavonols Quercetin, Kaempferol, and Myricetin Inhibit Hepatocyte Growth Factor-Induced Medulloblastoma Cell Migration. J. Nutr. 2009, 139, 646–652. [Google Scholar] [CrossRef]
- Shu, Y.; Xie, B.; Liang, Z.; Chen, J. Quercetin reverses the doxorubicin resistance of prostate cancer cells by downregulating the expression of c-met. Oncol. Lett. 2017, 15, 2252–2258. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.; Jung, N.; Lee, S.; Sohng, J.K.; Jung, H.J. Apigenin Inhibits Cancer Stem Cell-Like Phenotypes in Human Glioblastoma Cells via Suppression of c-Met Signaling. Phytotherapy Res. 2016, 30, 1833–1840. [Google Scholar] [CrossRef] [PubMed]
- AbouAitah, K.; Swiderska-Sroda, A.; Farghali, A.A.; Wojnarowicz, J.; Stefanek, A.; Gierlotka, S.; Opalinska, A.; Allayeh, A.K.; Ciach, T.; Lojkowski, W. Folic acid-conjugated mesoporous silica particles as nanocarriers of natural prodrugs for cancer targeting and antioxidant action. Oncotarget 2018, 9, 26466–26490. [Google Scholar] [CrossRef] [PubMed]
- Allen, R.E.; Sheehan, S.M.; Taylor, R.G.; Kendall, T.L.; Rice, G.M. Hepatocyte growth factor activates quiescent skeletal muscle satellite cells in vitro. J. Cell. Physiol. 1995, 165, 307–312. [Google Scholar] [CrossRef] [PubMed]
- Otabe, O.; Kikuchi, K.; Tsuchiya, K.; Katsumi, Y.; Yagyu, S.; Miyachi, M.; Iehara, T.; Hosoi, H. MET/ERK2 pathway regulates the motility of human alveolar rhabdomyosarcoma cells. Oncol. Rep. 2016, 37, 98–104. [Google Scholar] [CrossRef] [PubMed]
- Jankowski, K.; Kucia, M.; Wysoczynski, M.; Reca, R.; Zhao, D.; Trzyna, E.; Trent, J.; Peiper, S.; Zembala, M.; Ratajczak, J.; et al. Both hepatocyte growth factor (HGF) and stromal-derived factor-1 regulate the metastatic behavior of human rhabdomyosarcoma cells, but only HGF enhances their resistance to radiochemotherapy. Cancer Res. 2003, 63, 7926–7935. [Google Scholar] [PubMed]
- Miekus, K.; Lukasiewicz, E.; Jarocha, D.; Sekula, M.; Drabik, G.; Majka, M. The decreased metastatic potential of rhabdomyosarcoma cells obtained through MET receptor downregulation and the induction of differentiation. Cell Death Dis. 2013, 4, e459. [Google Scholar] [CrossRef]
- Chen, Y.; Takita, J.; Mizuguchi, M.; Tanaka, K.; Ida, K.; Koh, K.; Igarashi, T.; Hanada, R.; Tanaka, Y.; Park, M.-J.; et al. Mutation and expression analyses of theMET andCDKN2A genes in rhabdomyosarcoma with emphasis onMET overexpression. Genes, Chromosom. Cancer 2007, 46, 348–358. [Google Scholar] [CrossRef]
- Diomedi-Camassei, F.; McDowell, H.P.; De Ioris, M.A.; Uccini, S.; Altavista, P.; Raschellà, G.; Vitali, R.; Mannarino, O.; De Sio, L.; Cozzi, D.A.; et al. Clinical Significance of CXC Chemokine Receptor-4 and c-Met in Childhood Rhabdomyosarcoma. Clin. Cancer Res. 2008, 14, 4119–4127. [Google Scholar] [CrossRef]
- Yan, D.; Da Dong, X.; Chen, X.; Wang, L.; Lu, C.; Wang, J.; Qu, J.; Tu, L. MicroRNA-1/206 Targets c-Met and Inhibits Rhabdomyosarcoma Development. J. Biol. Chem. 2009, 284, 29596–29604. [Google Scholar] [CrossRef] [PubMed]
- Lukasiewicz, E.; Miekus, K.; Kijowski, J.; Drabik, G.; Wilusz, M.; Bobis-Wozowicz, S.; Majka, M. Inhibition of rhabdomyosarcoma’s metastatic behavior through downregulation of MET receptor signaling. Folia Histochem. et Cytobiol. 2010, 47, 485–489. [Google Scholar] [CrossRef] [PubMed]
- Saini, M.; Verma, A.; Mathew, S.J. SPRY2 is a novel MET interactor that regulates metastatic potential and differentiation in rhabdomyosarcoma. Cell Death Dis. 2018, 9, 237. [Google Scholar] [CrossRef] [PubMed]
- Ginsberg, J.P.; Davis, R.J.; Bennicelli, J.L.; Nauta, L.E.; Barr, F.G. Up-regulation of MET but not neural cell adhesion molecule expression by the PAX3-FKHR fusion protein in alveolar rhabdomyosarcoma. Cancer Res. 1998, 58, 3542–3546. [Google Scholar]
- Ferracini, R.; Olivero, M.; DI Renzo, M.F.; Martano, M.; De Giovanni, C.; Nanni, P.; Basso, G.; Scotlandi, K.; Lollini, P.L.; Comoglio, P. Retrogenic expression of the MET proto-oncogene correlates with the invasive phenotype of human rhabdomyosarcomas. Oncogene 1996, 12, 1697–1705. [Google Scholar] [PubMed]
- Fu, R.; Jiang, S.; Li, J.; Chen, H.; Zhang, X. Activation of the HGF/c-MET axis promotes lenvatinib resistance in hepatocellular carcinoma cells with high c-MET expression. Med Oncol. 2020, 37, 24. [Google Scholar] [CrossRef] [PubMed]
- Hao, Z.; Wang, P. Lenvatinib in Management of Solid Tumors. Oncologist 2020, 25, e30. [Google Scholar] [CrossRef]
- Wagner, J.; Abdel-Rahman, S.M. Jonathan Wagner, DO1,2,3 and Susan M. Abdel-Rahman, PharmD2,3 1Ward Family Heart Center and 2Division of Clinical Pharmacology, Toxicology and Therapeutic Innovation, Children’s Mercy Hospital, Kansas City, Missouri 3Department of Pediatrics, University o; Do; D, P. Pediatric Statin Administration: Navigating a Frontier with Limited Data. J. Pediatr. Pharmacol. Ther. 2016, 21, 380–403. [Google Scholar] [CrossRef]
- Voorberg, A.N.; Kamphuis, E.; Christoffers, W.A.; Romeijn, G.L.E.; Oosterhaven, J.A.F.; Schuttelaar, M.L.A. Efficacy and safety of oral alitretinoin versus oral azathioprine in patients with severe chronic hand eczema: Results from a prematurely discontinued randomized controlled trial. Contact Dermat. 2022, 87, 366–368. [Google Scholar] [CrossRef]
- Findling, R.L.; McNamara, N.K.; Pavuluri, M.; Frazier, J.A.; Rynn, M.; Scheffer, R.; Kafantaris, V.; Robb, A.; DelBello, M.; Kowatch, R.A.; et al. Lithium for the Maintenance Treatment of Bipolar I Disorder: A Double-Blind, Placebo-Controlled Discontinuation Study. J. Am. Acad. Child Adolesc. Psychiatry 2018, 58, 287–296. [Google Scholar] [CrossRef]
- Piérard, G.; Arrese, J.; Piérard-Franchimont, C. Itraconazole. Expert Opin. Pharmacother. 2000, 1, 287–304. [Google Scholar] [CrossRef] [PubMed]
- Lestner, J.; Hope, W.W. Itraconazole: an update on pharmacology and clinical use for treatment of invasive and allergic fungal infections. Expert Opin. Drug Metab. Toxicol. 2013, 9, 911–926. [Google Scholar] [CrossRef] [PubMed]
- Kast, R.E.; Alfieri, A.; Assi, H.I.; Burns, T.C.; Elyamany, A.M.; Gonzalez-Cao, M.; Karpel-Massler, G.; Marosi, C.; Salacz, M.E.; Sardi, I.; et al. MDACT: A New Principle of Adjunctive Cancer Treatment Using Combinations of Multiple Repurposed Drugs, with an Example Regimen. Cancers 2022, 14, 2563. [Google Scholar] [CrossRef] [PubMed]
- Halatsch, M.-E.; Dwucet, A.; Schmidt, C.J.; Mühlnickel, J.; Heiland, T.; Zeiler, K.; Siegelin, M.D.; Kast, R.E.; Karpel-Massler, G. In Vitro and Clinical Compassionate Use Experiences with the Drug-Repurposing Approach CUSP9v3 in Glioblastoma. Pharmaceuticals 2021, 14, 1241. [Google Scholar] [CrossRef] [PubMed]
- Palmer, A.C.; Chidley, C.; Sorger, P.K. A curative combination cancer therapy achieves high fractional cell killing through low cross-resistance and drug additivity. eLife 2019, 8, e50036. [Google Scholar] [CrossRef] [PubMed]
- Nogales, C.; Mamdouh, Z.M.; List, M.; Kiel, C.; Casas, A.I.; Schmidt, H.H. Network pharmacology: curing causal mechanisms instead of treating symptoms. Trends Pharmacol. Sci. 2021, 43, 136–150. [Google Scholar] [CrossRef] [PubMed]
- Kilmister, E.J.; Koh, S.P.; Weth, F.R.; Gray, C.; Tan, S.T. Cancer Metastasis and Treatment Resistance: Mechanistic Insights and Therapeutic Targeting of Cancer Stem Cells and the Tumor Microenvironment. Biomedicines 2022, 10, 2988. [Google Scholar] [CrossRef]
- Lindsey, B.A.; Markel, J.E.; Kleinerman, E.S. Osteosarcoma Overview. Rheumatol. Ther. 2016, 4, 25–43. [Google Scholar] [CrossRef]
- Cosio, T.; Di Prete, M.; Campione, E. Arsenic Trioxide, Itraconazole, All-Trans Retinoic Acid and Nicotinamide: A Proof of Concept for Combined Treatments with Hedgehog Inhibitors in Advanced Basal Cell Carcinoma. Biomedicines 2020, 8, 156. [Google Scholar] [CrossRef]
- Chow, C.K.; Atkins, E.R.; Hillis, G.S.; Nelson, M.R.; Reid, C.M.; Schlaich, M.P.; Hay, P.; Rogers, K.; Billot, L.; Burke, M.; et al. Initial treatment with a single pill containing quadruple combination of quarter doses of blood pressure medicines versus standard dose monotherapy in patients with hypertension (QUARTET): a phase 3, randomised, double-blind, active-controlled trial. Lancet 2021, 398, 1043–1052. [Google Scholar] [CrossRef]
- Hu, L.; Wang, D.; Liu, H.; Zhang, Q.; Sun, D.; Zhang, L.; Chen, X.; Chang, G.; Wang, J. A double-blind, placebo-controlled trial on the antihypertensive treatment effect of a quadruple single-pill combination. J. Clin. Hypertens. 2021, 23, 815–822. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



