
Submitted:
26 September 2023
Posted:
28 September 2023
You are already at the latest version
Abstract
Human disturbances can have severe environmental impacts on freshwater ecosystems. The main aim of this study was to detect the influences of physiochemical variables, land-cover characteristics, and river connectivity on fish assemblages in Lake Chaohu Basin, China. A cluster analysis of the river connectivity variables identified four groups of sites that characterized by significantly different connectivity gradients in local scale. These four groups of sites showed increasing connectivity from the upper reaches to the lower reaches. At the same time, among the four groups, the values of the environmental variables generally increased from the upper reaches with less human activity towards the lower reaches with more human activities. For instance, some main physiochemical variables (e.g., river width, water depth, nitrate, phosphate) significantly increased among the four groups. In contrast, fish taxa richness and diversity indices were not significantly different among the four connectivity groups. However, fish assemblages showed significant variation among the connectivity groups (p=0.026). In addition, the study determined that upper riparian land uses (e.g., woodland and grassland), flow velocity, and elevation were environmental variables regulating the variance in the fish communities, and for the connectivity variables, only river order and number of branches along a path to the left of the main stem affected the variance in the fish communities. Therefore, new practices aimed at maintaining and even increasing the riparian canopy coverage and the flow velocity of rivers should be integrated into local conservation planning for freshwater ecosystems, especially in the upper reaches of the basin.
Keywords:
fish
; river connectivity
; land use
; physiochemical variable
; spatial gradient
; Lake Chaohu Basin
1. Introduction
The species composition, abundance, and distribution of fish assemblages can be affected by both natural conditions and human activities along the spatial gradients of rivers (e.g., along gradients of upper reach-lower reaches) [1,2,3,4]. Many natural factors caused by elevation gradients, such as temperature, precipitation, and flow velocity, co-determine species richness and species distribution patterns [5,6,7,8]. For instance, elevation and several physiochemical variables (e.g., water depth, river width, flow velocity, and conductivity) were the variables that most affected the fish distributions in Ivinhema River Basin [9]. However, some studies have found that water temperature is one of the most important natural factors influencing fish growth and production [10], which also determine how many fish species can live and grow [6]. In addition, elevation, catchment area, and river longitudinal gradients primarily determine the fish assemblages [11,12,13,14,15].
Anthropogenic disturbances (e.g., land use changes, pollutant emissions) can also influence fish assemblages [16,17,18,19,20]. Pollutant emissions and reduced flow velocity stability derived from land use changes (e.g., changes from woodland to cropland and/or built-up land) can reduce the species richness of fish communities [13,21,22,23,24,25,26]. Dam construction is considered one of the major threats to the biodiversity of aquatic organisms because dams can alter the migratory patterns of species, influence the distribution of species, and block the ecological processes of natural rivers [2,4,27,28]. For instance, the construction of Gezhou Dam led to significant losses of several endemic fish species in the Yangtze River because the dam prevented the upstream pathway of fish migration and substantially decreased their breeding grounds [29]. Although a large number of studies have tried to determine the factor with the greatest influence on fish assemblages, the relative importance of isolated and combined effects of natural conditions and anthropogenic disturbances on freshwater fish still needs to be determined for different regions, as these factors are characterized by a range of spatial and temporal scales [16,30,31].
Recently, numerous studies have focused on the influences of loss of connectivity and fragmentation of habitats generally caused by dam construction and land use changes on fish assemblages [2,8,17,25,32,33]. One of the main reasons for focusing on these influences is that many connectivity-related variables are the most important factors affecting freshwater fish assemblages in rivers [1,2,16,18,34,35]. River connectivity (e.g., hydrologic connectivity) can be defined as the water-mediated transport of matter, energy, and/or organisms within or between elements of the hydrologic cycle [18,19,20,36]. Therefore, river connectivity has an important influence on the variance among freshwater fish species. For instance, the connectivity of lakes, which is correlated with lake size, depth, and distance from rivers, is crucial to structuring the fish assemblages in the fluvial lakes of the Mississippi River [37]. Although very important to the contribution of connectivity-related variables to fish distribution, the combined influence of natural conditions and human activities have been proposed to be more important than the isolated influence of losses of connectivity on freshwater fish species distributions [8,13,38].
Generally, fish assemblages are likely to be affected by many factors; however, in this study, we focused on land use characteristics, physiochemical variables and river connectivity in local scale. Therefore, the aims of this study were to test which variables regulate fish assemblages. Therefore, we tried to identify the key influence variables that currently structure the fish assemblages in a lowland basin of China (Lake Chaohu Basin) and determine which factor was the most important.
2. Materials and Methods
2.1. Study area
The Lake Chaohu Basin is located in the lower reaches of the Yangtze River, with an area of 1.41×104 km2 (Figure 1). The average elevation within this area is approximately 65 m and ranges from 0 to 1498 m (higher in the west and lower in the east). More than 70% of the total area is lowland plain, with elevations ranging from 0-50 m [39]. Woodland is the main land use type in the western mountains, and cropland is the main land use type in the eastern plains (Figure 1). Generally, the most prevalent land use type in Lake Chaohu Basin was cropland (60.1% of total area), followed by woodland (15.2%), built-up land (11.8%), water bodies (8.7%) and grassland (4.2%) in 2015.
Figure 1.
Location of the sampling sites and the land use in 2015 in the Lake Chaohu Basin, China.
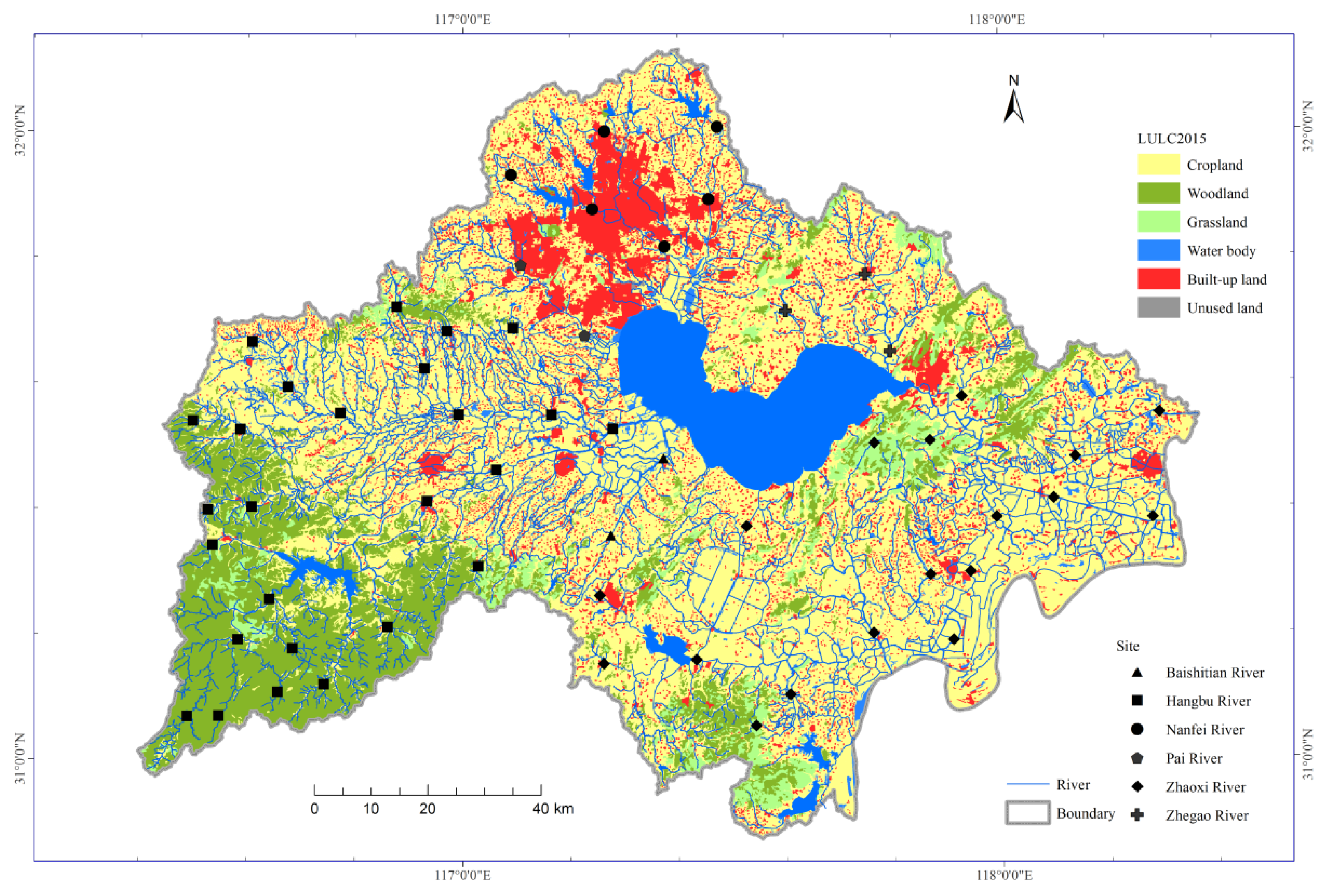
There is a high-density river network with more than 33 rivers centripetally distributed around Lake Chaohu [39]. Most of the inflow comes from the Hangbu River, Baishitian River, and Nanfei River to Lake Chaohu [12]. However, only one river, the Yuxi River, links the lake to the Yangtze River [39]. The Hangbu River has the highest drainage density, followed by the Baishitian River, Zhaoxi River (including the Zhao River and Yuxi River), Pai River, Nanfei River, and Zhegao River [39].
2.2. Fish sampling and diversity indices
During April 2013, fishes were collected using a backpack electro-fishing unit (CWB-2000 p, China; 12-V import, 250-V export) by wading in two passes [40]. The sampling sites were distributed in all six main subcatchments (Hangbu River, Baishitian River, Zhaoxi River, Pai River, Nanfei River, and Zhegao River) in the Lake Chaohu basin. The sampling encompassed complete sets of the characteristic segment forms (e.g., stream, pools and riffles) [41,42]. A total 57 sampling sites were surveyed in this study (Figure 1). A free-flowing segment of 50 m in length was sampled for each sampling site, the entire segment was sampled when the segment was less than 50-m length. When the water depth effect was 1 m, we directly waded into the water for sampling; when the water depth was greater than 1 m, we used a rubber boat to assist sampling. All electro-fishing passes were conducted using a uniform sampling effort, with approximately 30 min of sampling time for each 50-m segment performed by the same three persons [40]. All fishes caught in the 40-m blocked segment were stored in nets, which were placed in the segment to keep the fish as alive as possible. Then fishes were identified to the lowest practical level (usually species), counted, and returned together to the sampling sites if alive. Unidentifiable specimens were fixed in 10% formalin, and transferred to 75% ethanol for identification in the laboratory according to more characterizes [2].
Four diversity indices, Shannon-Wiener index, Berger-Parker dominance index, Margalef's richness index, Pielou’s equitability index, were calculated for each site based on species abundance data [43]. Species with total percentage abundance less than 0.5% were excluded from the analysis to reduce possible biases [44]. The diversity indices can be calculated as follows:
Where H, BP, R and J are Shannon-Wiener index, Berger-Parker dominance index, Margalef's richness index and Pielou’s equitability index, respectively. ni is the number of individuals of the i th species and N is the total number of individuals. Nmax is the number of individuals of dominant species. S is the number of taxa.
2.3. Measured environmental variables
2.3.1. Local-scale physiochemical variables
For each sampling sites, a set of physiochemical variables was surveyed during the sampling period. The river width (width) at each site was recorded in situ using a laser rangefinder (SNDWAY SW-1500A). Flow velocity (Flow) was measured in situ by a portable propeller-type current meter (LS300-A). Water temperature (Temp), pH, dissolved oxygen (DO), electrical conductivity (EC) and turbidity (Turb) were measured in situ using a YSI 6600 V2 multi-parameter water quality sonde (Yellow Springs, USA). Water depth (Depth) was measured in situ by hydrologic sounding rod (Chinese National Standard, GB/T 27992.1-2011). Some physiochemical variables (Flow, Temp, pH, DO, EC, Turb and Depth) were measured along cross-stream transects at 3- to 5-m intervals (river width ≤ 25m, 3-m intervals; river width > 25m, 5-m intervals), with monitoring points spaced 1-m apart [42]. Each sampling sites should have no less than 3 cross-stream transects. The cross-stream transects were set at the place where the water flow was smooth and the water surface had no obvious slope. The monitoring point was located at 0.5 m below the surface of water or 1/2 water depth. For each physiochemical variable above, the average value of all monitoring points of each sampling site was taken as the value of this sampling site.
Besides, an integrated water sample was collected using a 1-L organic glass water sampler. All the water samples were kept at 4°C prior to the analyses. Other physicochemical factors, including total dissolved solid (TDS), dissolved organic carbon (DOC), total nitrogen (TN), ammonium (NH4+-N), nitrate (NO3--N), total phosphorus (TP), and phosphate (PO4 3+-P), were measured in the laboratory according to standard methods (American Public Health Association, 2012). After extraction in 90% hot ethanol of water samples, Chlorophyll a (Chl-a) concentrations was detected by Spectrophotometry [45].
2.3.2. Land use characteristics
Riparian buffer zone was used to identify the land use characteristics for the sampling site. Segments with 1 km upstream and downstream from the sampling site were used as up-segment and down-segment, respectively. Therefore, upstream and downstream riparian buffer zones could be confirmed by a 1-km buffer zone along the up-segment and down-segment, respectively. Land use characteristics for each riparian buffer zone were determined using the 30-m-accuracy land use datasets in Lake Chaohu Basin (Figure 1), with five categories: cropland, woodland, grassland, water body and built-up land. The land use datasets obtained from the Resource and Environment Science and Data Center (https://www.resdc.cn/). The percentage of each category of land use in each buffer zone was calculated using ArcGIS 10.1 (Environmental Systems Research Institute, Redlands, CA, USA). Besides, the percentages of cropland, woodland, grassland, water body and built-up land in the upstream riparian buffer zones are abbreviated to U_Crop, U_Wood, U_Grass, U_Water and U_Built, respectively.
2.3.3. River connectivity variables
In this study, the several common river connectivity variables were selected, which include river order, stream-link magnitude, branch link, confluence link, downstream link, upstream segment length, and downstream segment length. Specifically, river order (ROrder) was determined using the Strahler’s method [46]. Stream-link magnitude (Link) is the number of unbranched source streams upstream of a given segment in a river network [47]. The branch link (BLink) is the number of branches along a path to the right (BLink_R) and left (BLink_L) of the main stem (i.e., BLink=BLink_R+BLink_L) [48]. Confluence link (CLink) is the number of confluences downstream from each stream segment [49]. Downstream link (DLink) is the link magnitude of the next downstream confluence [50]. Upstream-segment length (Up_L) is the distance from the nearest upstream confluence site to the sampling site in the same river segment, and downstream-segment length (Down_L) is the distance from the nearest downstream confluence site. Additionally, the location of the sampling site (LSS) in the segment is defined as the ratio of the upstream length to the total segment length (LSS=Up_L/T_L). A digital 1:50,000-scale hydrology map of Lake Chaohu Basin was used to create the river network (Figure 1) and to calculate these river connectivity variables, which was performed in the software ArcGIS 10.1.
2.4. Statistical analysis
There were three groups of environmental variables (19 variables) in our dataset, but not all of them could be directly used to analyze the effects of the variables on fish diversity indices due to multicollinearity among them; thus, it was necessary to select the available variables. First, to reduce the dimensionality and better compare the variables, all the initial values (except for Land use characteristics) were normalized by the min-max normalization method [51]. Second, a variance inflation factor (VIF) was employed to detect the multicollinearity of the variables. If the VIF was more than 5, multicollinearity was likely present; therefore, a threshold value of 5 was set to decrease the effects of multicollinearity [52,53]. The VIF was calculated using the package vegan (version 2.5-7) [54] in R (R Development Core Team, 2013). Finally, to reduce the complexity of the variation partitioning analysis (VPA), the factors with a VIF < 5 in the three groups were further screened according to their relative importance, which was calculated based on the sum of their Akaike weights (SAW) in a mixed-effects meta-regression model using the package glmulti (Version 1.0.8) in R. If the SAW was more than 0.8, the parameter was considered a key driving variables to explain the deviance in the fish diversity indices [55,56]. Key driving variables were considered good predictors of the response variable without the need to invoke other variables [56].
Besides, VPA was performed to disentangle the influential intensity of the key driving variables (SAW > 0.8) on the fish diversity indices [57]. VPA was performed using the function varpart in the package vegan (version 2.5-7) in R. The proportions of the variation in fish diversity indices explained by the unique and combined influences of the key driving variables were reported based on the adjusted R2 values [57,58]. A negative value in the report was not considered because it indicated that the explained variance may be negligible [56]. Finally, we used multiple regression analysis to detect the effects (strength and direction) of each Key driving variable on the fish diversity indices, which was also performed in R.
The physiochemical variables (except for pH), land use variables, river connectivity and species abundance data were log10 (x+1) transformed before the following analysis to reduce the influence of non-normal distribution. In addition, as variances in the local-scale physiochemical variables and diversity indices were not homogeneous, the nonparametric Kruskal-Wallis test was employed to detect the differences in the local-scale physiochemical variables and diversity indices among the connectivity groups using SPSS 20.0 (IBM Co. Ltd, USA) [12].
Several statistical analysis methods were employed to test which variables regulate fish assemblages. Nonmetric multidimensional scaling (NMDS) was employed to assess the spatial variation in fish assemblages based on Bray-Curtis dissimilarity measures of fish abundance data. One-way analysis of similarity (ANOSIM) was also used to determine whether fish assemblages differed significantly among the connectivity groups [2,12]. Besides, similarity percentage (SIMPER) procedures were employed to identity the species that contributed the most to the dissimilarities among groups [21]. These analyses were performed using PAST 3.21 software [43], accordingly.
Furthermore, to identify the key influence variables that currently structure the fish assemblages and determine which factor was the most important, a canonical correspondence analysis (CCA) was performed based on abundance data [2]. A manual forward selection procedure was employed to identify which environmental variables were significantly related to the fish assemblages. The environmental variables with a significance level of p<0.1 were chosen based on 9,999 Monte Carlo permutation tests (Zhang et al. 2018). The CCA was performed using CANOCO 4.5 software [59].
The fish abundance data were log10(x+1) transformed, before the analyses, to reduce the influence of non-normal distribution. Local-scale physiochemical variables (except TDS, TN, NH4+-N, PO43+-P) and land use characteristics (except proportion of cropland) were included in CCA analyses, because TDS, TN, NH4+-N, PO43+-P and proportion of cropland were multiple collinearities with other variables. Species that occurred at fewer than five sites were excluded from NMDS, ANOSIM, and CCA analyses [2].
3. Results
3.1. Clustering river connectivity variables
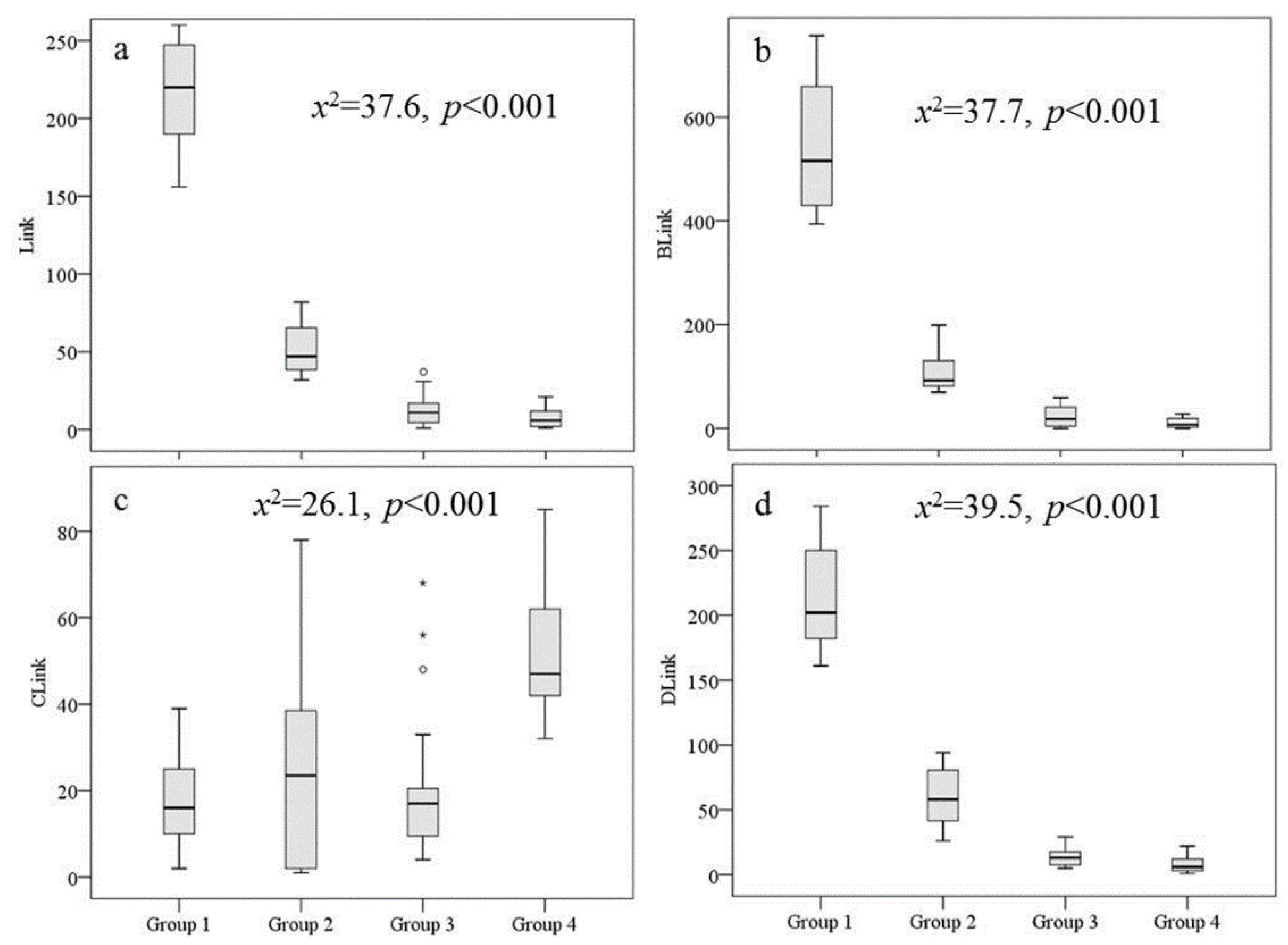
The cluster analysis resulted in four groups of sites with a Euclidean similarity of 200 among sites within a cluster. The sampling sites of the four groups showed an upper-lower reach distribution pattern across the basin (Figure 2). The four groups formed in the cluster analysis had significantly different connectivity according to the Kruskal-Wallis test (Figure 3 and Table A1). For example, sites in Group 1 had the highest Link, BLink, and DLink, followed by the sites in Groups 2, 3 and 4. However, sites in Group 4 had the highest values of CLink, followed by the sites in groups 2, 3 and 1. Generally, these results indicated that sites in Group 1 usually had the highest connectivity from the lower to upper reaches, followed by sites in Groups 2, 3 and 4 (Figures 2 and 3).
Figure 2.
Spatial distribution of the four connectivity groups identified by cluster analysis.
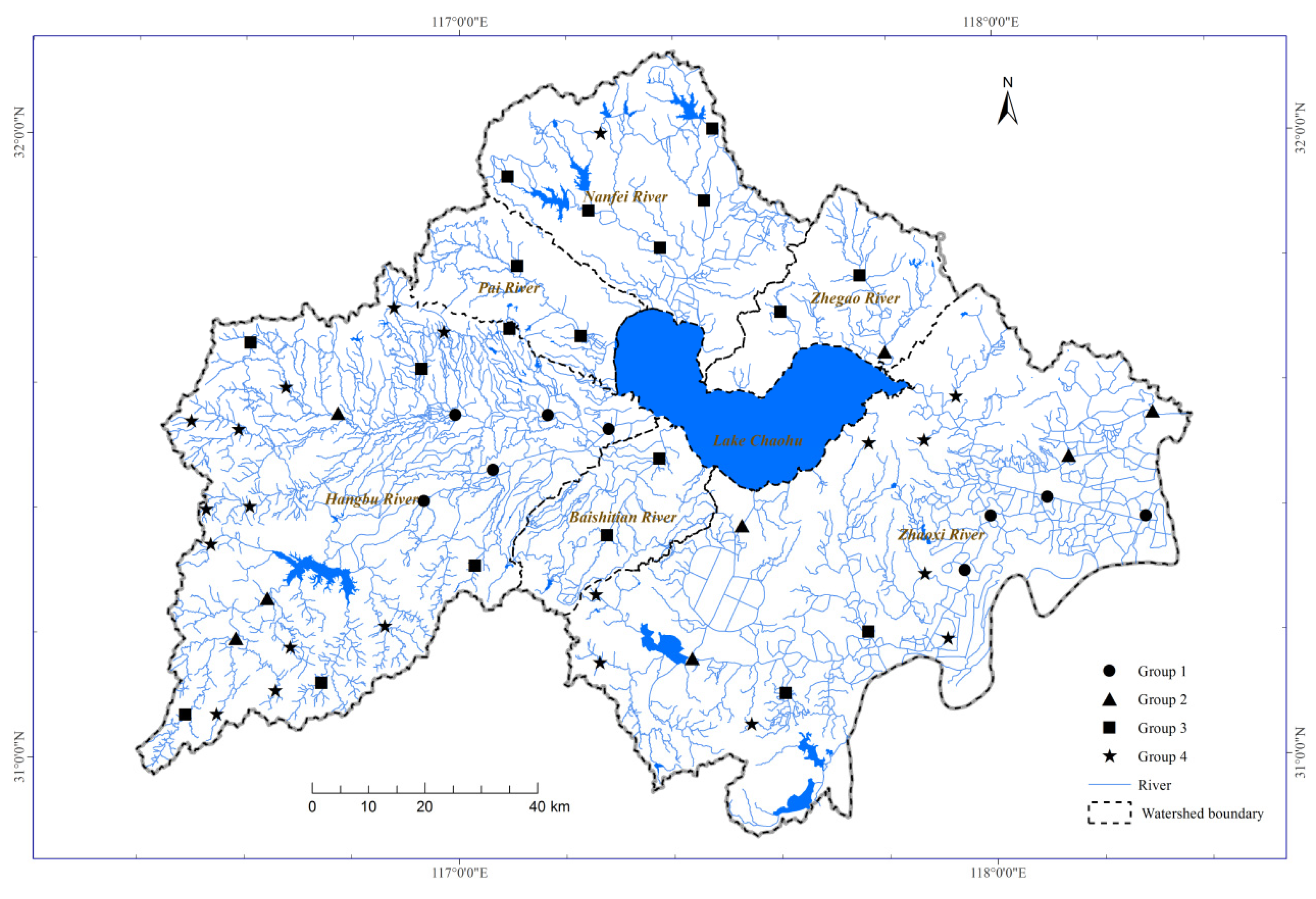
Figure 3.
Box plots of four connectivity variables in the four cluster groups. The Kruskal-Wallis test of the variables among the four groups indicated significant differences between the groups (p<0.05).
Figure 3.
Box plots of four connectivity variables in the four cluster groups. The Kruskal-Wallis test of the variables among the four groups indicated significant differences between the groups (p<0.05).
Therefore, sites from Group 1 were mainly located in the lower reaches with high-connectivity segments (e.g., Hangbu River and Zhaoxi River). Sites from Group 2 were clustered in the upper reaches with moderate-connectivity segments (e.g., Hangbu River and Zhaoxi River). Sites from Group 3 were mainly distributed in the middle reaches with low-connectivity segments (e.g., Nanfei River, Pai River, Zhegao River, and Baishitian River). Sites from Group 4 were situated in the upper reaches with the lowest connectivity segments (e.g., Nanfei River, Pai River, Zhegai River, Baishitian River, Hangbu River, and Zhaoxi River).
Generally, river orders in Lake Chaohu Basin ranged from 1st to 5th (Table A1). Link, BLink_R, BLink_Lf, BLink, CLink, and DLink ranged from 1 to 567, 0 to 970, 0 to 537, 0 to 1423, 1 to 58, and 1 to 499, respectively. Upstream segment length (Up_L), downstream segment length (Down_L) ranged from 0.01 to 5.23 km and 0.01 to 16.50 km, respectively. The location of sampling site (LSS) ranged from 0.01 to 0.99.
3.2. Spatial gradients of physiochemical variables among connectivity groups
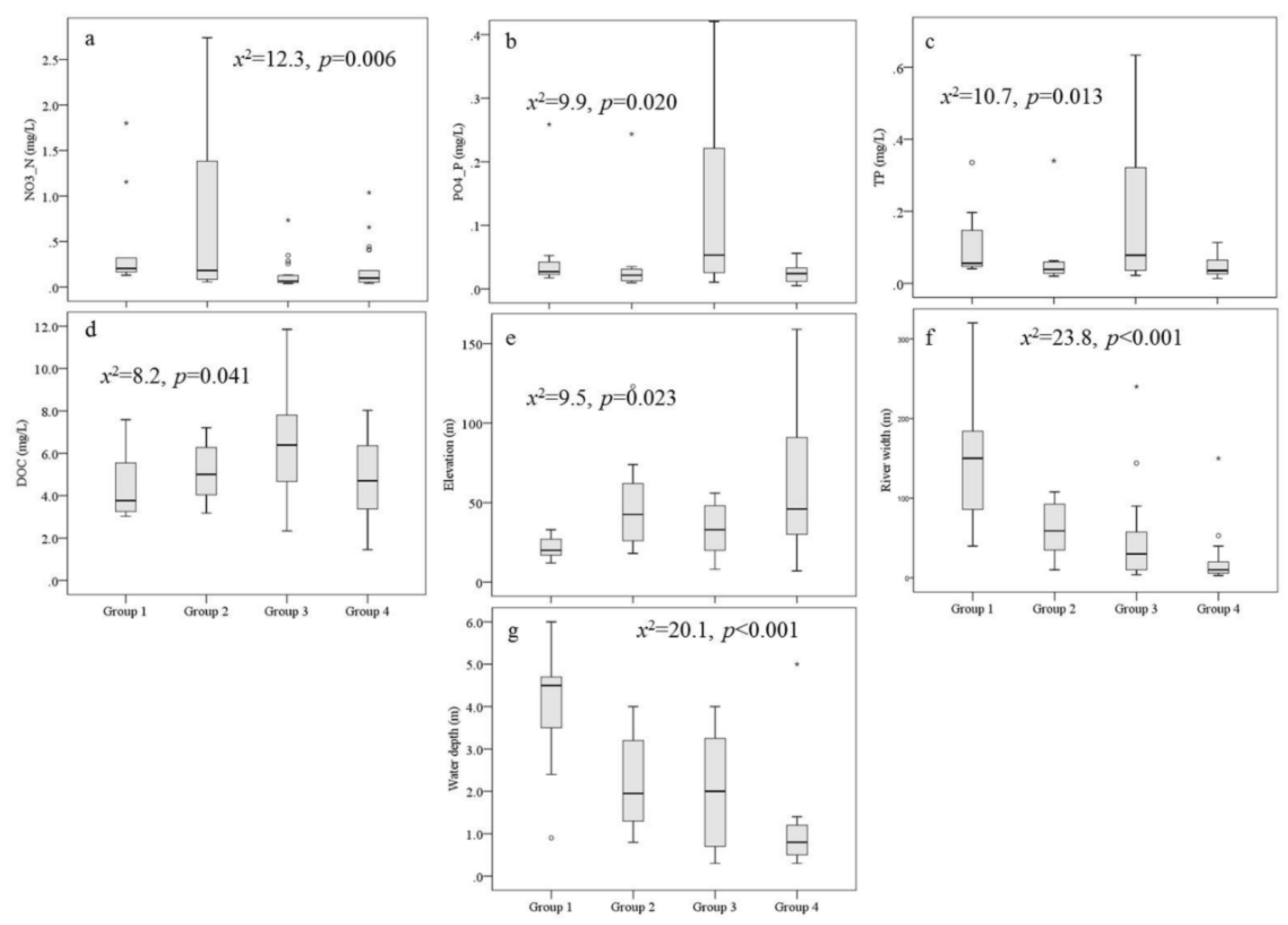
Although many physiochemical variables were not significantly different among the four connectivity groups, several variables (e.g., Width, Depth, NO3--N, TP, DOC, and PO43+-P) were significantly different among the four groups according to the Kruskal-Wallis test (Table A2 and Figure 4). Specifically, in comparison to the other sites and Groups, major sites in Group 3 had the highest pollution (e.g., EC and Turb) and nutrient levels (e.g., TN, NO3--N, TP, PO43+-P, and DOC) and the lowest values for DO, Flow and NH4+-N, whereas sites in Groups 4 and 2 had low nutrient and pollution levels and higher flow velocity. Sites in Group 1 had the highest Width, Depth, and DO and even the highest pH values.
Figure 4.
Box plots of selected physiochemical variables in the four cluster groups. The Kruskal-Wallis test of the variables among the four groups indicated significant differences between the groups (p<0.05).
Figure 4.
Box plots of selected physiochemical variables in the four cluster groups. The Kruskal-Wallis test of the variables among the four groups indicated significant differences between the groups (p<0.05).
There was a significant spatial gradient for the land use characteristics along the river segments (Table A3 and Figure A1). The percentage of grassland in the 1-km buffer along the upstream segments (U_Grass) decreased from the upper reach sites (Group 4) to lower reach sites (Group 1). On the other hand, the percentage of water body in the 1-km buffer along the upstream segments (U_Water) showed the opposite trend, increasing from the upper reach sites (Group 4) to lower reach sites (Group 1). The percentage of woodland in the 1-km buffer along the upstream segments (U_Wood) showed a complex trend along the segments, with the highest values in the upstream segments of sites in Group 4. The next highest U_Wood values were in the upstream segments of sites in Group 2, followed by Group 3 and Group 1. Although the percentage of cropland in the 1-km buffer along the upstream segments (U_Crop) and the percentage of built-up land in the 1-km buffer along the upstream segments (U_Built) were not significantly different among the four connectivity groups, U_Crop increased from the upper reach sites (Group 4) to lower reach sites (Group 1), and the highest U_Built was clustered in Group 3. The same results were found downstream and for all segments of the sites (Table A3 and Figure A1).
3.3. Influence of river connectivity on fish assemblages
A total of 2166 individuals were collected throughout Lake Chaohu Basin at 57 sites, representing 38 species in 35 genera and 13 families. Species richness and number of individuals caught varied from 1 to 14 and 1 to 445, respectively, across sites. Based on the Kruskal-Wallis test, fish taxa richness and diversity indices were not significantly different among the connectivity groups (Table A4).
However, one-way ANOSIM results showed that fish assemblages significantly varied by connectivity groups (global R=0.089, p=0.026). Specifically, fish assemblages could be distinguished between Groups 3 and 4 (R=0.160, p=0.006) and Groups 2 and 4 (R=0.127, p=0.048) (Table 1). The SIMPER analysis revealed that the species that primarily contributed to the dissimilarity between Groups 3 and 4 were Carassius auratus (Linnaeus) (16.72% of contribution), Ctenogobius sp. (13.14%), Hemiculter leucisculus (Basilewsky) (11.45%), and Misgurnus anguillicaudatus (Cantor) (8.53%), while Ctenogobius auratus (18.10%), Ctenogobius sp. (14.24%), H. leucisculus (11.41%), and Acheilognathus barbatulus (Günther) (6.10%) mostly contributed to the difference between Groups 2 and 4. In addition, C. auratus, Ctenogobius sp., and H. leucisculus were the dominant species and occurred at most of the sites throughout Lake Chaohu Basin. Furthermore, the minimum stress value was 0.14 in the NMDS ordination solution for the river connectivity groups (Figure 5a). The NMDS analysis revealed that sampling sites in Group 3 were mainly located on the right of the graph, while the sites in Group 4 were gathered to the left. At the same time, sampling sites in Groups 1 and 2 were mainly clustered to the top of the plot.

Table 1.
One-way ANOSIM showing significance levels of fish community structure among the four groups. The upper triangular matrix showed the p values, and the lower triangular matrix showed the global R statistic.
Table 1.
One-way ANOSIM showing significance levels of fish community structure among the four groups. The upper triangular matrix showed the p values, and the lower triangular matrix showed the global R statistic.
| Group 1 | Group 2 | Group 3 | Group 4 | |
| Group 1 | 0.696 | 0.320 | 0.104 | |
| Group 2 | -0.042 | 0.390 | 0.048* | |
| Group 3 | 0.029 | 0.004 | 0.006** | |
| Group 4 | 0.100 | 0.127 | 0.160 |
* p<0.05, ** p<0.01.
Figure 5.
Nonmetric multidimensional scaling plot based on species abundance data and connectivity variation according to four groups (a) and on stream size according to river order (b) in Lake Chaohu Basin. In the left plot (a), each symbol represents a group (Group 1, blue dot; Group 2, green diamond; Group 3, red square; and Group 4, empty triangle). In the right plot (b), each symbol represents a group (1st-order streams, red square; 2nd-order streams, empty square; 3rd-order streams, empty triangle; 4th-order streams, empty diamond; and 5th-order streams, red dot).
Figure 5.
Nonmetric multidimensional scaling plot based on species abundance data and connectivity variation according to four groups (a) and on stream size according to river order (b) in Lake Chaohu Basin. In the left plot (a), each symbol represents a group (Group 1, blue dot; Group 2, green diamond; Group 3, red square; and Group 4, empty triangle). In the right plot (b), each symbol represents a group (1st-order streams, red square; 2nd-order streams, empty square; 3rd-order streams, empty triangle; 4th-order streams, empty diamond; and 5th-order streams, red dot).
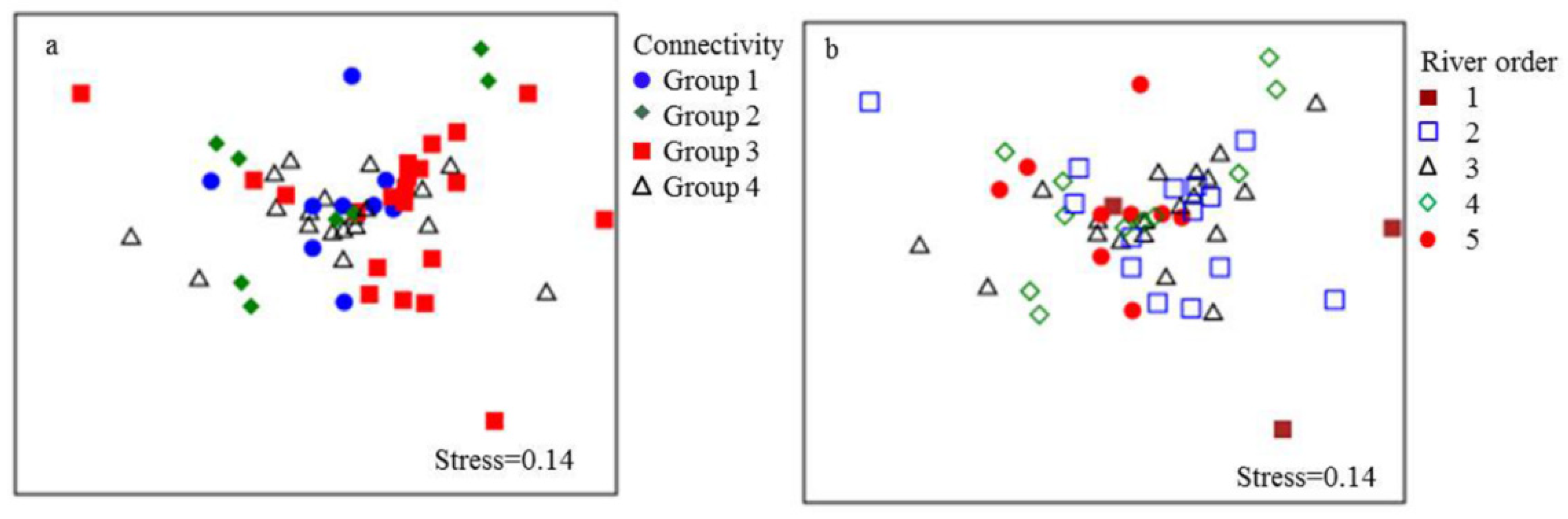
Based on the Kruskal-Wallis test, fish taxa richness and diversity indices were not significantly different among river order groups (Table A5). Similarly, there was also no significant difference in fish assemblages among river orders (global R=0.004, p=0.424). Therefore, river order cannot influence fish assemblages in Lake Chaohu Basin. However, the minimum stress value was 0.14 in the NMDS ordination solution for river orders (Figure 5b). The NMDS analysis revealed that the sampling sites in the 2nd- and 3rd-order streams were mainly located on the right of the graph, while the sampling sites in the 5th-order streams were gathered to the left. At the same time, the sampling sites in the 1st-order streams were mainly clustered to the bottom right, and the sampling sites in the 4th-order streams were mainly located at the top.
3.4. Linking environmental variables to fish assemblages
The forward selection procedure for the CCA identified eight environmental variables (U_Wood, U_Grass, Flow, ROrder, Alka, Elevation, BLink_Lf, and DO) that were highly correlated with the fish communities (Figure 6). U_Wood explained the most variance (15.4%), followed by U_Grass (5.4%), Flow (5.1%), ROrder (3.0%), Alka (2.8%), Elevation (2.7%), BLink_Lf (2.6%), and DO (2.5%). The first and second axes accounted for 16.5 and 6.8%, respectively, of the total variation in fish species abundances. The first axis was highly related to the variables U_Wood (canonical coefficient, r=0.75), Flow (r=0.75), and Elevation (r=0.58), while the second axis was corrected to U_Grass (r=0.42). In addition, the first axis was negatively related to ROrder (r=-0.50). Based on the CCA plot, many sites in the lower reaches of the high-connectivity rivers (Group 1) were clustered around ROrder, while some sites in the upper reaches of the lower-connectivity rivers (Group 4) were gathered around BLink_Lf and Alka. Sites in the lower reaches of high-connectivity rivers (Group 1) were located around BLink_Lf and DO. In addition, sites in the middle reaches of low-connectivity rivers (Group 3) were plotted along Flow, U_Wood, U_Grass, and Elevation.
Figure 6.
Canonical correspondence analysis (CCA) of fish assemblages and environmental variables at the 57 river sites (a: sites and b: species) for which eight environmental variables as significant contributors were examined in Lake Chaohu Basin.
Figure 6.
Canonical correspondence analysis (CCA) of fish assemblages and environmental variables at the 57 river sites (a: sites and b: species) for which eight environmental variables as significant contributors were examined in Lake Chaohu Basin.
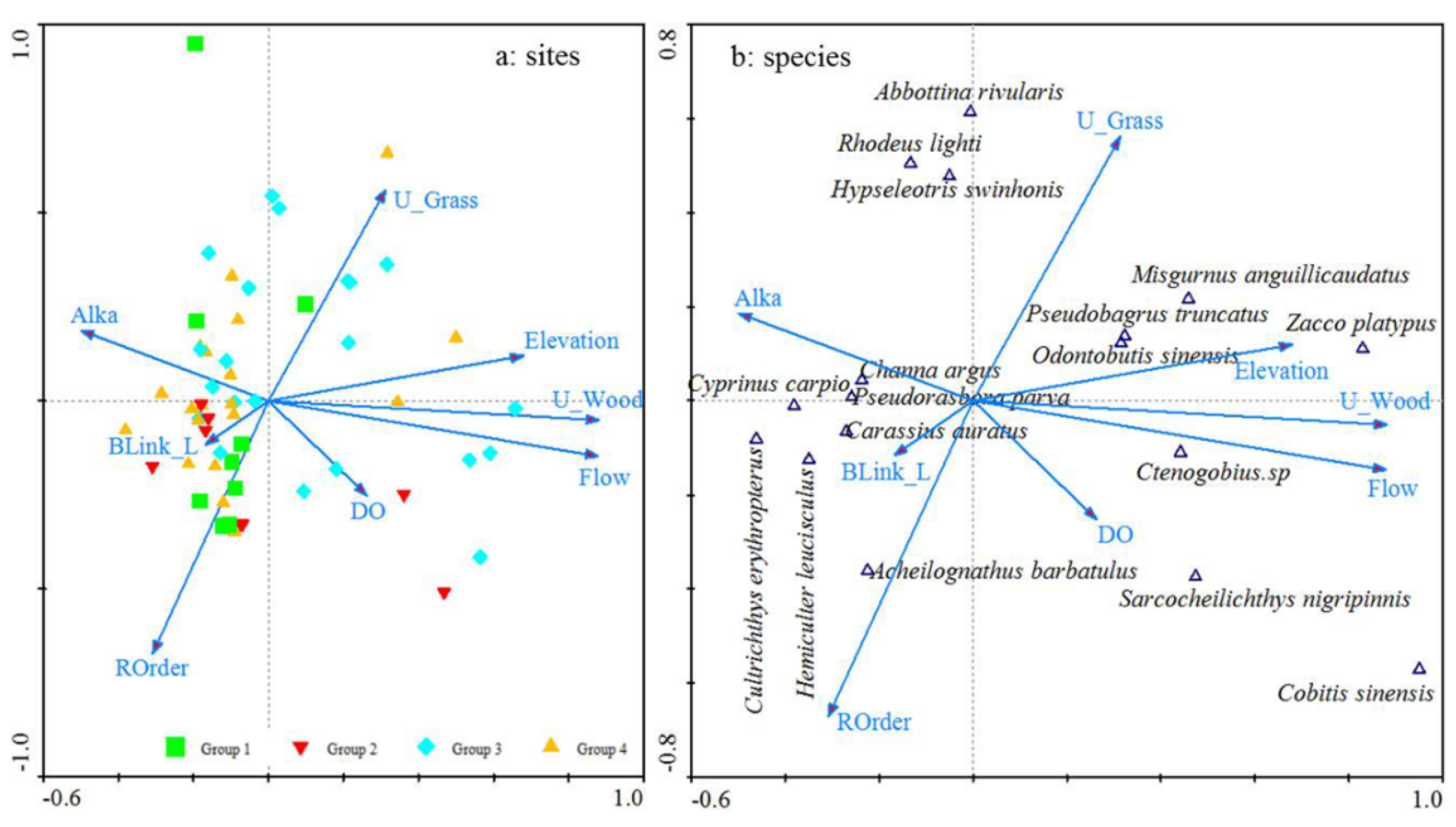
The CCA results also showed the relationships between some common species and the eight environmental variables (Figure 6b). Two dominant species, C. auratus and H. leucisculus, were positively related to BLink_Lf and negatively related to U_Wood, Flow, and Elevation. This result means that these two species would prefer to live in the lower reaches with high BLink_Lf and low Elevation, U_Wood, and Flow. The other dominant species, Ctenogobius sp., were positively correlated with Flow, U_Wood, and Elevation. Similarly, five species belonging to Cypriniformes (Zacco platypus (Temminck and Schlegel) and Misgurnus anguillicaudatus (Cantor)), Perciformes (Ctenogobius sp. and Odontobutis sinensis (Wu, Chen and Chong)), and Siluriformes (Pseudobagrus truncates (Regan)) were positively correlated to Elevation, U_Wood, and Flow. In other words, these six species generally occurred in the upper reaches with high Elevation, U_Wood, and Flow. Moreover, two species of Cypriniformes (Abbottina rivularis (Basilewsky) and Rhodeus lighti (Wu)) and one species of Perciformes (Hypseleotris swinhonis (Günther)) were most frequently found in the upper reaches of high U_Grass and low ROrder. Cobitis sinensis (Sauvage and Dabry de Thiersant) and Sarcocheilichthys nigripinnis (Günther) were found in the upper and middle reaches with high DO.
4. Discussion
4.1. Spatial heterogeneity of environmental variables
Many environmental variables showed significant variation along the longitudinal gradients of the rivers throughout Lake Chaohu Basin. For example, sites in Group 1 had the highest values of the connectivity variables (ROrder, Link, BLink, and DLink), local-size variables (river width, water depth, and DO), and U_Crop. The sites in Group 4 had the highest Elevation, U_Wood, and U_Grass; the lowest river width, water depth and U_Crop; and the lowest nutrient levels (e.g., TP, NO3--N, PO43+-P). Specifically, sites in Group 3 had the highest nutrient levels (e.g., TN, NO3--N, TP, and PO43+-P), which was likely because this area has the highest area of built-up land along the segments of these sites [12,60]. Moreover, the nutrient levels showed an increasing trend from sites in the upper reaches to the lower reaches (Table A2). However, due to anthropogenic disturbances, such as increasing the area of built-up land and cropland, this increasing trend of nutrient variation can be transformed suddenly [12,13,23]. For example, sites in Group 3, which were located in the middle and lower reaches, had the highest nutrient levels. This scenario means that human activities (land use change from woodland and water body to the built-up land and cropland) can strongly influence the water quality in this region [14,23].
Moreover, in this study, a cluster analysis of the river connectivity variables identified four groups of sampling sites that were characterized by significantly different river connectivity and longitudinal gradients. This cluster is significantly different from the results in Yu et al. [61], who simply gathered the sampling sites into two groups, isolated and connected groups. Their study focused on the relationships between the water quality of the isolated and connected wetlands and their surrounding watershed, documenting that the DOC was significantly lower in connected wetlands than in isolated wetlands when the land use did not change significantly, while this difference can be shift completely due to changes in land use. Furthermore, according to varying flood intensities, the sampling sites were grouped into five categories: drought-connected, low flood-connected, moderate flood-connected, high flood-connected, and extreme flood-connected [18,62]. Research has determined the effect of connectivity on water chemistry and shown that lakes with greater connectivity usually have higher DO and NO3--N and lower TN and TP. These different results may vary from those of our study due to the different ways to cluster connectivity and the different subjects, wetlands/lakes vs rivers.
Similarly, some environmental variables did not show significant variation along the rivers, but several main nutrients had remarkably different concentrations among the connectivity groups. This result is also slightly different from the results of Zhang et al. [12]. Their research showed that all the environmental variables had significant differences among their cluster groups. They may have used species abundance data to cluster the sampling sites in their research, and we used the four connectivity variables to cluster the sampling sites in this study due to the aims of our research.
4.2. Connectivity variables slightly influence the variation in fish assemblages
Due to the lack of available data on fish assemblages in Lake Chaohu Basin, temporal variability was not detected in this study. The most significant way in which our study differed from most others is that the sampling sites were grouped according to four river connectivity variables. Thus, we focused on the spatial variance in fish assemblages and tested the hypothesis that fish assemblages are regulated by river connectivity variables in the lowland basin (Lake Chaohu Basin, China). The results of this study cannot support our hypothesis: river connectivity variables regulate fish assemblages. Fish taxa richness and diversity indices were not significantly related to all river connectivity variables (not shown in the text) and no significant difference existed among the connectivity groups (p>0.099, Table A4), fish assemblages significantly varied by connectivity groups (global R=0.089, p=0.026), particularly between Groups 3 and 4 and Groups 2 and 4. In addition, according to the results of the CCA, ROrder and BLink_Lf explain a fraction of the variance (3.0% and 2.6%, respectively). Therefore, the results indicated that the connectivity variables could explain a small part of the variance and slightly influence the variation in fish assemblage in the lowland basin, China. In contrast, the LSS was not significantly different among river order groups (Table A1) and did not explain any variance in fish assemblages (Figure 6), which means that the location of the sampling site in the same segment does not affect the fish assemblages in a river segment; thus, there is no significant difference in fish communities in the same segment. The most likely reason for this result is the high migration ability of fish [1,63].
Our results are slightly different from those of He et al. [2], Osborne and Wiley [50], Smith and Kraft [64], and Yan et al. [1]. These studies demonstrated that CLink, Link, ROrder, or DLink could explain the highest proportion of fish assemblage variance. For instance, Osborne and Wiley [50] found that DLink explained the greatest influence on fish assemblages, while Smith and Kraft [64] showed that CLink and ROrder could explain the highest portion of fish assemblage variance. He et al. [2] found that the downstream, mainstream confluence site and DLink together can explain more than 50% of the total variance in fish assemblages. However, based on our results, the most important environmental variable influencing the variance in fish assemblage was upstream land use, followed by flow velocity, and river connectivity variables were less important. Therefore, our results support the idea that freshwater fish communities are generally determined by environmental varibales in local scale, although river connectivity seems very important to fish assemblages [1,8,9,13,65].
4.3. Upstream land use and flow velocity play more important roles in the variation in fish assemblages
Another aim of this study was to test the relative importance of physiochemical variables, land use characteristics, and river connectivity variables on fish assemblage structure in the lowland basin. This study found that riparian vegetation, such as woodland and grassland areas, could be the most important factor affecting the fish assemblages, followed by the flow velocity of rivers. These two factors can explain 25.9% of the total variation among fish species abundances in Lake Chaohu Basin, although their explanatory power is lower than the factors detected in previous studies [1,2,50,64]. Thus, U_Wood, U_Grass and Flow play a more important role in the variation among fish assemblages than connectivity variables (e.g., ROrder and BLink_Lf, which together accounted for 5.6% of the total variation). These results were highly consistent with results from previous studies, which documented that both local-scale variables (e.g., habitant characteristics) and spatial structure (e.g., connectivity) indices can influence fish assemblages in river networks [8,13,35,50,63].
In addition, some studies have emphasized the role of physiochemical variables [1] and land use characteristics [9] in the variance among fish communities. Land use has been considered a primary driver of environmental impacts on rivers [66,67]. For instance, increasing cropland could lead to high levels of nutrients and sediments and decreased flow stability of rivers [67,68]. Moreover, loss of riparian canopy coverage (e.g., deforestation) could alter river conditions (e.g., river morphology and substrate) and the structure of fish assemblages (e.g., alter the distribution of fish taxa) [67]. Similar results were found in this study, which showed a shift from herbivore (R. lighti), omnivore (Ctenogobius sp. and Z. platypus) and primary carnivore (A. rivularis, H. swinhonis, and O. sinensis) fish assemblages in segments with the highest amount of woodland and grassland along the upper streams to omnivore (H. leucisculus and P. parva) and secondary carnivore (C. argus and C. erythropterus) fish assemblages in segments with the highest cropland and built-up land along the upper streams.
Moreover, other main factors of local-scale physiochemical variables, which can influence the fish assemblages, are flow velocity and elevation (Figure 6). Due to the highest elevations occurring in the southwestern basin, where the mountain area is located, the flow velocity is usually higher in this area than in other regions. Indeed, elevation is unable to affect the fish assemblages directly; it can alter many physiochemical variables, for example flow velocity, temperature, and DO, and then influence the species distribution [7,9]. Therefore, our findings indicate that the factors that most affected the fish assemblages in Lake Chaohu Basin were riparian land use characteristics (U_Wood and U_Grass), followed by local-scale physiochemical variables (Flow and Elevation) and river connectivity variables (ROrder and BLink_Lf).
5. Conclusions
Our results indicated how the fish communities in Lake Chaohu Basin were structured by physicochemical variables, land use, and river connectivity. Sampling sites were clustered into four groups according to river connectivity. Therefore, the influences of the environmental variables on fish assemblages can be detected among the connectivity groups. The results showed that several nutrients (NO3--N, TP, and PO43+-P) were significantly different among the connectivity groups. More importantly, although this study cannot support our hypothesis: river connectivity variables regulate fish assemblages, the results shown that the most important environmental variable influencing the variance in fish assemblages was upstream land use, followed by flow velocity and Elevation, and river connectivity variables were less important, although fish assemblages showed significant variation among the connectivity groups. Therefore, particular attention should be paid to maintaining the riparian canopy coverage upstream areas and the flow velocity of rivers in the upper reaches of the basin because these factors could improve the biodiversity and conservation status of fishes.
Author Contributions
“Conceptualization, Z.Z. and J.G.; methodology, Z.Z. and Y.C.; software, Z.Z. and Y.C.; formal analysis, Z.Z.; investigation, Z.Z. and Y.C.; resources, Z.Z.; writing—original draft preparation, Z.Z.; writing—review and editing, Z.Z., J.G. and Y.C. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Natural Science Foundation of China, China, grant number 41977194.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We would like to send a great thanks to H.B. Yin, T. Xia, and K. Liu for their assistance with fieldwork.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Yan, Y.Z.; Xiang, X.Y.; Chu, L. Zhan, Y.J.; Fu, C.Z. Influences of local habitat and stream spatial position on fish assemblages in a dammed watershed, the Qingyi Stream, China. Ecol. Freshw. Fish. 2011, 20, 199−208. [CrossRef]
- He, D.K.; Kang, Z.J.; Tao, J.; Liu, C.L.; Yang, J.; Chen, Y.F. Hydrologic connectivity driven natural stream fish assemblages in mountain streams in the Yangtze River basin: Implications for stream fish conservation in monsoonal East Asia. Hydrobiologia 2017, 785, 185–206. [CrossRef]
- Deng, X.J.; Xu, Y.P.; Han, L.F. Impacts of human activities on the structural and functional connectivity of a river network in the Taihu Plain. Land Degrad. Dev. 2018, 29, 2575–2588. [CrossRef]
- Yi, Y.J.; Gao, Y.N.; Zhang, S.H. The impact of dams on the river connectivity of the two largest river basins in China. River Res. Appl. 2022, 38(2), 185−193. [CrossRef]
- Bhatt, J.P.; Manish, K.; Pandit, M.K. Elevational Gradients in Fish Diversity in the Himalaya, Water Discharge Is the Key Driver of Distribution Patterns. PLoS ONE 2012, 7, e46237. [CrossRef]
- Carvajal-Quintero, J.D.; Escobar, F.; Alvarado, F.; Villa-Navarro, F.A.; Jaramillo-Villa,Ú.; Maldonado-Ocampo, J.A. Variation in freshwater fish assemblages along a regional elevation gradient in the northern Andes, Colombia. Ecol. Evol. 2015, 5, 2608–2620. [CrossRef]
- Li, Y.H.; Yan, Y.Z.; Zhu, R.; Zhou, K.; Chu, L. Spatial variations in fish assemblages within the headwater streams of the Wanhe watershed, A river network-based approach. Journal of Fishery Sciences of China 2014, 21, 988–999. (in Chinese).
- Valenzuela-Aguayo, F.; McCracken, G.R.; Manosalva, A.; Habit, E.; Ruzzante, D.E. Human-induced habitat fragmentation effects on connectivity, diversity, and population persistence of an endemic fish, Percilia irwini, in the Biobio River basin (Chile). Evol. Appl. 2020, 13, 794–807. [CrossRef]
- Lemke, A.P.; Súarez, Y.R. Influence of local and landscape characteristics on the distribution and diversity of fish assemblages of streams in the Ivinhema River basin, Upper Paraná River. Acta Limnologica Brasiliensia 2013, 25, 451−462. [CrossRef]
- He, H.; Jin, H.; Jeppesen, E.; Li, K.Y.; Liu, Z.W.; Zhang, Y.D. Fish-mediated plankton responses to increased temperature in subtropical aquatic mesocosm ecosystems, Implications for lake management. Water Res. 2018, 144, 304–311. [CrossRef]
- Di Prinzio, C.Y.; Casaux, R.J.; Miserendino, M.L. Effects of land use on fish assemblages in Patagonian low order streams. Ann. Limnol-Int. J. Lim. 2009, 45, 267–277. [CrossRef]
- Zhang, Y.; Cheng, L.; Tolonen, K.E.; Yin, H.B.; Gao, J.F.; Zhang, Z.M.; Li, K.Y. Substrate degradation and nutrient enrichment structuring macroinvertebrate assemblages in agriculturally dominated Lake Chaohu Basins, China. Sci. Total Environ. 2018, 627, 57–66. [CrossRef]
- Shao, X.J.; Fang, Y.; Jawitz, J.W.; Yan, J.G.; Cui, B.S. River network connectivity and fish diversity. Sci. Total Environ. 2019, 689, 21–30. [CrossRef]
- Yu, Z.H.; Wang, Q.; Xu, Y.P.; Lu, M.; Lin, Z.X.; Gao, B. Dynamic impacts of changes in river structure and connectivity on water quality under urbanization in the Yangtze River Delta plain. Ecol. Indic. 2022, 135, 108582. [CrossRef]
- Guan, Q.; Wu, H.T.; Xu, L.;, Kang, Y.J.; Lu, K.L.; Liu, D.D.; Han, D.D.; Xue, Z.S.; Yuan, Y.X.; Wang, W.F.; Zhang, Z.S. Hydrological connectivity shapes multiple diversity facets of snail (Mollusca: Gastropoda) assemblages in freshwater floodplain wetlands. Ecol. Indic. 2023, 153, 110467. [CrossRef]
- Rodeles, A.A.; Galicia, D.; Miranda, R. A new method to include fish biodiversity in river connectivity indices with applications in dam impact assessments. Ecol. Indic. 2020, 117, 106605. [CrossRef]
- González-Ferreras, A.M.; Leal, S.; Barquín, J.; Almodóvar, A. Patterns of genetic diversity of brown trout in a northern Spanish catchment linked to structural connectivity. Aquat. Sci. 2022, 84, 48. [CrossRef]
- Mazur, M.L.C.; Smith, B.; Bird, B.; McMillan, S.; Pyron, M.; Hauswald, C. Hydrologic connectivity and land cover affect floodplain lake water quality, fish abundance, and fish diversity in floodplain lakes of the Wabash-White River basin. River Res. Appl. 2022, 38, 160–172. [CrossRef]
- Nagata, Y.; Ishiyama, N.; Nakamura, F.; Shibata, H.; Fukuzawa, K.; Morimoto, J. Contribution of Hydrological Connectivity in Maintaining Aquatic Plant Communities in Remnant Floodplain Ponds in Agricultural Landscapes. Wetlands 2023, 43(4), 38. [CrossRef]
- Scordo, F.; Seitz, C.; Fiorenza, J.E.; Piccolo, M.C.; Perillo, G.M.E. Human impact changes hydrological connectivity in a Patagonian fluvial basin. J. Hydrol-Reg. Stud. 2023, 45, 101315. [CrossRef]
- Shutes, R.B. Artificial wetlands and water quality improvement. Environ. Int. 2001, 26, 441–447. [CrossRef]
- Gido, K.B.; Dodds, W.K.; Eberle, M.E. Retrospective analysis of fish community change during a half-century of landuse and streamflow changes. J. N. Am. Benthol. Soc. 2010, 29, 970–987. [CrossRef]
- Xu, Z.; Ji, J.; Shi, C. Water geochemistry of the Chaohu Lake Basin rivers, China: Chemical weathering and anthropogenic inputs. Appl. Geochem. 2011, 26, S379–S383. [CrossRef]
- Lane, C.R.; Leibowitz, S.G.; Autrey, B.C.; LeDuc, S.D.; Alexander L.C. Hydrological, physical, and chemical functions and connectivity of non-floodplain wetlands to downstream waters, A review. J. Am. Water Resour. As. 2018, 54, 346–371. [CrossRef]
- Wilkinson, C.L.; Yeo, D.C.J.; Tan, H.H.; Fikri A.H.; Ewers R.M. Land-use change is associated with a significant loss of freshwater fish species and functional richness in Sabah, Malaysia. Biol. Conserv. 2018, 222, 164–171. [CrossRef]
- Xu, N.; Liang, X.S.; Zhang, T.Y.; Dong, J.X.; Wang, Y.; Qu, Y. Spatio-Temporal Evolution Patterns of Hydrological Connectivity of Wetland Biodiversity Hotspots in Sanjiang Plain between 1995 and 2015. Sustainability 2023, 15(6), 4952. [CrossRef]
- Liermann, C.R.; Nilsson, C.; Robertson, J.; Ng, R.Y. Implications of Dam Obstruction for Global Freshwater Fish Diversity. BioScience 2012, 62, 539–548. [CrossRef]
- O'Mara, K.; Venarsky, M.; Stewart-Koster, B.; McGregor, G.B.; Schulz, C.; Kainz, M.; Marshall, J.; Bunn S.E. Connectivity of fish communities in a tropical floodplain river system and predicted impacts of potential new dams. Sci. Total Environ. 2021, 788, 147785. [CrossRef]
- Xie, P.; Three-Gorges Dam, Risk to ancient fish. Science 2003, 302(5648), 1149, 1151.
- Hu, P.; Zeng, Q.H.; Wang, J.H.; Hou, J.M.; Wang, H.; Yang, Z.F.; Liu, H.; Zhao, Y. Identification of hotspots of threatened inland fish species and regions for restoration based on longitudinal river connectivity. J. Environ. Manage. 2021, 290, 112572. [CrossRef]
- Mouchlianitis, F.A.; Bobori, D.; Tsakoumis, E.; Sapounidis, A.; Kritikaki, E.; Ganias K. Does fragmented river connectivity alter the reproductive behavior of the potamodromous fish Alburnus vistonicus? Hydrobiologia 2021, 848, 4029–4044. [CrossRef]
- Sá-Oliveira, J.C.; Isaac, V.J.; Ferrari, S.F. Fish community structure as an indicator of the long-term effects of the damming of an Amazonian river. Environ. Biol. Fish. 2015, 98, 273–286. [CrossRef]
- Larrieu, K.G.; Pasternack, G.B.; Schwindt, S. Automated analysis of lateral river connectivity and fish stranding risks-Part 1: Review, theory and algorithm. Ecohydrology 2021, 14, e2268. [CrossRef]
- Argiroff, W.A.; Zak, D.R.; Lanser, C.M.; Wiley, M.J. Microbial community functional potential and composition are shaped by hydrologic connectivity in riverine floodplain soils. Microb. Ecol. 2017, 73, 630–644. [CrossRef]
- Stoffels, R.J.; Humphries, P.; Bond, N.R.; Price, A.E. Fragmentation of lateral connectivity and fish population dynamics in large rivers. Fish Fish, 2022, 23, 680-696. [CrossRef]
- Pringle, C.M. Hydrologic connectivity and the management of biological reserves, A global perspective. Ecol. Appl. 2001, 11, 981–998. [CrossRef]
- Miranda, L.E. Fish assemblages in oxbow lakes relative to connectivity with the Mississippi River. T. Am. Fish. Soc. 2005, 134, 1480–1489. [CrossRef]
- Branco, P.; Segurado, P.; Santos, J.M.; Pinheiro, P.; Ferreira, M.T. Does longitudinal connectivity loss affect the distribution of freshwater fish? Ecol. Eng. 2012, 48, 70–78. [CrossRef]
- Gao, J.F.; Cai, Y.J.; Xia, T.; Zhang, Z.M.; Yin, H.B.; Huang, Q. Aquatic ecosystem health of Lake Chaohu Basin. Science Press, Beijing, China, 2016. (in Chinese).
- Liu, Y.; Wang, Y.R.; Zhu, Q.; Li, Y.R.; Kang, B.; Chu, L.; Yan, Y.Z. Effects of low-head dams on fish assemblages in subtropical streams: Context dependence on species category and data type. River Res. Applic. 2019, 35, 396-404. [CrossRef]
- Tedesco, P.A.; Ibanez, C.; Moya, N.; Bigorne, R.; Camacho, J.; Goitia, E.; Hugueny, B.; Maldonado, M.; Rivero, M.; Tomanová, S.; Zubieta, J.P.; Oberdorff, T. Local-scale species-energy relationships in fish assemblages of some forested streams of the Bolivian Amazon. C. R. Biol. 2007, 330, 255–264. [CrossRef]
- Datry, T.; Melo, A.S.; Moya, N.; Zubieta, J.; De la Barra, E.; Oberdorff, T. Metacommunity patterns across three Neotropical catchments with varying environmental harshness. Freshwater Biol. 2016, 61, 277–292. [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. (2001) Past: Paleontological statistics software package for education and data analysis. Palaeontol Electron 2001, 4(1), 1–9.
- Paredes del Puerto, J.M.; Garcia, I.D.; Maiztegui, T.; Paracampo, A.H.; Capítulo, L.R.; Garcia de Souza, J.R.; Maroñas, M.E.; Colautti, D.C. Impacts of land use and hydrological alterations on water quality and fish assemblage structure in headwater Pampean streams (Argentina). Aquat. Sci. 2022, 84, 6. [CrossRef]
- Pápista, É.; Ács, É.; Böddi, B. Chlorophyll-a determination with ethanol–a critical test. Hydrobiologia, 2002, 485, 191–198. [CrossRef]
- Strahler, A.N. Quantitative analysis of watershed geomorphology. Transactions American Geophysical Union 1957, 38, 913–920. [CrossRef]
- Shreve, R.L. Statistical law of stream numbers. J. Geol. 1966, 74:17–37. [CrossRef]
- Nieman, D.A. Perspectives on the recent status of redfin pickerel in the Schuylkill River basin of southeast Pennsylvania. In: Warmwater workshop proceedings, esocid management and culture. Soderberg, R., Ed.; American Fisheries Society Northeast Division, Mansfield, Pennsylvania, 1996.
- Fairchild, G.W.; Horwitz, R.J.; Nieman, D.A.; Boyer, M.R.; Knorr, D.F. Spatial variation and historical change in fish assemblages of the Schuylkill River drainage, southeast Pennsylvania. Am. Midl. Nat. 1998, 139, 282–295.
- Osborne, L.L.; Wiley, M.J. Influence of tributary spatial position on the structure of warmwater fish communities. Can. J. Fish. Aquat. Sci. 1992, 49, 671−681. [CrossRef]
- Dou, H.S.; Li, X.B.; Li, S.K.; Dang, D.L.; Li, X.; Lyu, X.; Li, M.Y.; Liu, S.Y. (2020). Mapping ecosystem services bundles for analyzing spatial trade-offs in inner Mongolia, China. J. Clean. Prod. 2020, 256, 120444. [CrossRef]
- Zuur, A.F.; Ieno, E.N.; Smith, G.M. Analysing Ecological Data, Statistics for Biology and Health. Springer, New York, 2007.
- Lemm, J.U.; Venohr, M.; Globevnik, L.; Stefanidis, K.; Panagopoulos, Y.; van Gils, J.; Posthuma, L.; Kristensen, P.; Feld, C.K.; Mahnkopf, J.; Hering, D.; Birk, S. Multiple stressors determine river ecological status at the European scale: Towards an integrated understanding of river status deterioration. Global Change Biol. 2021, 27, 1962–1975. [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O'Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, H.M.H.; Szoecs, E.; Wagner, H. Vegan: Community Ecology Package. Version 2.5-7. https://cran.r-project.org/web/packages/vegan/index.html (accessed 8 May 2023).
- Chen, J.; Luo, Y.; Van Groenigen, K.J.; Hungate, B.A.; Cao, J.; Zhou, X.; Wang, R.W. A keystone microbial enzyme for nitrogen control of soil carbon storage. Sci. Adv. 2018, 4(8), eaaq1689. [CrossRef]
- Li, J.Q.; Bååth, E.; Pei, J.M.; Fang, C.M.; Nie, M. Temperature adaptation of soil microbial respiration in alpine, boreal and tropical soils: An application of the square root (Ratkowsky) model. Global Change Biol. 2021, 27, 1281–1292. [CrossRef]
- Peres-Neto, P.R.; Legendre, P.; Dray, S.; Borcard, D. Variation partitioning of species data matrices: Estimation and comparison of fractions. Ecology 2006, 87(10), 2614-2625. [CrossRef]
- Cai, Y.J.; Xu, J.; Zhang, M.; Wang, J.J.; Heino, J. Different roles for geography, energy and environment in determining three facets of freshwater molluscan beta diversity at broad spatial scales. Sci. Total Environ. 2019, 659, 451-462. [CrossRef]
- ter Braak, C.; Smilauer, P. CANOCO: Reference Manual and CanoDraw for Windows user's Guide, Software for Canonical Community Ordination Version 4.5. Microcomputer Power Ithaca, New York, 2002.
- Zhang, Z.M.; Gao, J.F. Linking landscape structures and ecosystem service value using multivariate regression analysis, a case study of the Chaohu Lake Basin, China. Environ. Earth Sci. 2016, 75, 3. [CrossRef]
- Yu, X.B.; Hawley-Howard, J.; Pitt, A.L.; Wang, J.J.; Baldwin, R.F.; Chow, A.T. Water quality of small seasonal wetlands in the Piedmont ecoregion, South Carolina, USA, Effects of land use and hydrological connectivity. Water Res. 2015, 73, 98–108. [CrossRef]
- MacKinnon, B.D.; Sagin, J.; Baulch, H.M.; Lindenschmidt, K.E.; Jardine T.D. Influence of hydrological connectivity on winter limnology in floodplain lakes of the Saskatchewan River Delta, Saskatchewan. Can. J. Fish. Aquat. Sci. 2016, 73, 140–152. [CrossRef]
- Grenouillet, G.; Pont, D.; Hérissé, C. Within-basin fish assemblage structure, the relative influence of habitat versus stream spatial position on local species richness. Can. J. Fish. Aquat. Sci. 2004, 61, 93–102.
- Smith, T.A., Kraft, C.E. Stream fish assemblages in relation to landscape position and local habitat variables. T. Am. Fish. Soc. 2005, 134, 430–440. [CrossRef]
- Lansac-Tôha, F.M.; Meira, B.R.; Segovia, B.T.; Lansac-Tôha, F.A.; Velho, L.F.M. Hydrological connectivity determining metacommunity structure of planktonic heterotrophic flagellates. Hydrobiologia 2016, 781, 81–94. [CrossRef]
- Matono, P.; Sousa, D.; Ilhéu, M. Effects of land use intensification on fish assemblages in Mediterranean climate streams. Environ. Manage. 2013, 52, 1213–1229. [CrossRef]
- Leitão, R.P.; Zuanon, J.; Mouillot, D.; Leal, C.G.; Hughes, R.M.; Kaufmann, P.R.; Villéger, S.; Pompeu, P.S.; Kasper, D.; de Paula, F.R.; Ferraz, S.F.B.; Gardner, T.A. Disentangling the pathways of land use impacts on the functional structure of fish assemblages in Amazon streams. Ecography 2018, 41, 219–232. [CrossRef]
- Miranda, L.E.; Bies, J.M.; Hann, D.A. Land use structures fish assemblages in reservoirs of the Tennessee River. Mar. Freshwater Res. 2015, 66, 526–534. [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



