
Submitted:
27 September 2023
Posted:
28 September 2023
You are already at the latest version
Abstract
Cotton leaf curl Kokhran virus (CLCuKoV) (genus, Begomovirus; family, Geminiviridae) is highly infectious, widespread and the most dangerous pathogen of cotton (Gossypium hirsutum L.) that is responsible to a serious disorder, cotton leaf curl disease (CLCuD). Begomoviruses are spread very efficiently by the whitefly Bemisia tabaci cryptic species, causing economic losses to cotton crop, all over the world. The ‘Lucknow’ strain of CLCuKoV has emerged as a divergent isolate that could cause CLCuD. The monopartite ssDNA genome of CLCuKoV-Lu (2.7 Kb) contains six open reading frames (ORFs) that was shown to encode four major proteins. RNA interference (RNAi)-based antiviral innate immunity is a sequence-specific biological phenomenon and a powerful tool to control plant viruses. The present study aims to determine cotton locus-derived microRNAs (ghr-miRNAs) that are identified for targeting the CLCuKoV-Lu ss-DNA-encoded mRNAs using a predictive approach that involves four computational algorithms, miRanda, RNA22, psRNATarget and RNAhybrid. Mature ghr-miRNA sequences (n=80) from allotetraploid upland cotton (2n = 4x = 52) were selected from the miRBase and were tested for alignment with the CLCuKoV-Lu genome. Among the 80 cotton locus-derived ghr-miRNAs evaluated, only one consensus cotton locus-derived ghr-miRNA (ghr-miR2950) was concluded to have effective ghr-miRNA target site at common nucleotide position 82 in the CLCuKoV-Lu genome respectively, using a stringent criterion, identified by all the algorithms used. The miRNA targeting is reliant on base pairing of miRNA-mRNA target pairings. Conservation of the hybridization binding site of the predicted ghr-miR2950 was validated using multiple sequence alignment within all the strains of CLCuKoV. We constructed a regulatory interaction network of miRNA–mRNA to identify novel targets. The efficacy of the predicted miRNAs against CLCuKoV-Lu was evaluated by RNAi-mediated targeted mRNA cleavage. The current investigated miRNA targets provide evidence for the development of CLCuD-resistant cotton plants.
Keywords:
Cotton leaf curl Kokhran virus
; microRNAs
; RNA interference
; prediction
; computational algorithms
; target binding sites
; host-virus interaction
1. Introduction
The allotetraploid upland cotton (Gossypium hirsutum L.) is an important essential fiber-producing industrial cash crop which is grown on several continents of the world attributed 40% world’s agriculture production. Cotton is an important natural textile fiber made up of 90% cellulose and is a renewable natural source for global textiles industry. The quality parameters of cotton fibers have a direct influence on agro-based textile industry [1,2,3]. Being Allotetraploid nature, upland cotton genome contains 52 chromosomes (2n = 4x = 52)[4,5]. The first draft genome sequences of model polyploidy upland allotetraploid cotton species was accomplished and physical map was released in 2015 [6]. Cotton leaf curl disease (CLCuD) is a major biotic and economic constraint of cotton and is caused by at least four species in the genus Begomovirus (Geminiviridae) [7,8]. CLCuD complex is caused by monopartite begomoviruses (BGVs) [9,10]. CLCuD outbreaks caused by CLCuKoV significantly constrain cotton production in Pakistan [11,12,13]. BGVs are known to be spread efficiently by polyphagous whitelyfly vector, i.e., Bemisia tabaci [14,15,16,17].
Cotton leaf curl Kokhran virus-Lucknow (CLCuKoV-Lu) has recently emerged, and has been reported as new ‘strain’ of CLCuKoV in the begomovirus genus of the Geminiviridae family [18,19]. The architecture of the CLCuKoV genome inside virion is made up of a single copy of circular, single-stranded (ss) DNA molecule. The monopartite genome of a CLCuKoV encodes six proteins. The transcription and regulation of these proteins are governed from a large intergenic region (LIR) using bidirectional mode of transcription [12,20,21]. The plus (+) virion-sense (VS) and negative (-) complementary-sense (CS) strands encode ORFs V1 and V2, and ORFs C1, C2, C3 and C4, respectively [15,18,22]. CLCuKoV-Lu was first noticed in Lucknow in 2010 on guar (Cyamopsis tetragonoloba) plant, exhibiting leaf curl symptoms [19]. Rolling circle amplification (RCA) is widely used molecular technique to identify and isolate BGVs. Loop-mediated isothermal amplification (LAMP) assay, quantitative real-time polymerase chain reaction (qPCR) assay, multiplex PCR and immunofluorescence assay are also optimized and standardized as molecular diagnostics methods for CLCuD [23,24,25,26,27,28,29].
Although efforts have been redirected to control CLCuD, there are no resistant cultivars available yet in Pakistan. Considering the DNA nature of CLCuKoV, RNA interference (RNAi) has emerged as a robust and efficient tool for targeting microRNA-induced silencing complex (miRISC)-mediated gene silencing in eukaryotes [30,31,32,33]. RNAi-based silencing is a sequence-specific double-stranded (ds) RNA-mediated gene regulatory innate antiviral, natural mechanism to inhibit viral gene replication and transcription. The RNAi machinery is composed of two core central components, Dicer and Argonaute. These are involved for loading of 20-30 nucleotides long RNA molecule for processing of miRNA/miRNA* duplex and further was incorporated into RISCs. The dsRNA was processed and are broken down to short 21–24 nucleotides siRNA that was degraded. [34,35,36,37,38]. The plant microRNAs (miRNAs) are the smallest, widespread, highly conserved, non-coding, single stranded (ss) RNA molecules ranging from 18-24 nt encoded by MIR genes. The endogenous miRNAs play important role in regulating plant gene expression and key biological process by recruiting RISC complex [39,40,41].
Upland Cotton plant has been investigated for potential diverse molecular mechanism and was explored mature miRNAs that are critical for normal growth [42,43,44]. Cotton miRNAs are good source of immunity to control biotic and abiotic stress response networks [45,46,47,48,49]. The artificial microRNA (amiRNA)-based gene silencing impart resistance against invading viruses [50]. The amiRNA construct was transformed to induce gene silencing of target plant virus in 2006 [51]. Experimentally verified locus-derived mature miRNAs in the cotton genome was predicted to regulate gene expression. A subset of high-confidence mature miRNAs in cotton is designed to have predicted target sites in the CLCuKoV-Lu genome.
Our present an algorithmic in silico approach relied on prediction of upland allotetraploid cotton genome-encoded miRNAs targeting CLCuKoV-Lu. The research work summarized key strategies for analyzing most effective target sites of ghr-miRNAs in the CLCuKoV-Lu genome. To explore host-virus complex mechanism, miRNA-mRNA target site interactions were also explored. The study aims to elucidate the predicted ghr-miRNAs for the generation of CLCuKoV-Lu-resistant cotton plants in future.
2. Materials and Methods
2.1. Biological Data Retrieval
There are currently 80 mature cotton locus-derived ghr-microRNAs (commonly called Gossypium hirsutum -microRNAs (ghr-miR156-ghr-miR7514) (Accession IDs: MIMAT0005806-MIMAT0029164) (Table S1 (supplementary Materials)) identified. In a previous study, 78 stem-loop (precursor) cotton locus-derived ghr-miRNAs (ghr-MIR156-ghr-MIR7514) (Accession IDs: MI0005638-MI0024206) have been identified (Table S2). The cotton locus-derived ghr-miRNAs were retrieved from the miRBase latest version (v22.1) database (http://mirbase.org/) (accessed on 26 December 2018). miRBase offers biological information and annotation on miRNAs [52]. The full-length genome (2761 nucleotides) of CLCuKoV-Lu (accession number GU385879) was downloaded from the NCBI GenBank database (http://ncbi.nlm.nih.gov) (accessed on 19 December 2018) [53].
2.2. Target Prediction
The current in silico approach was based on the most widely used, publicly available miRNA prediction algorithms such as miRanda, RNA22, psRNATarget, and RNAhybrid. These algorithms were used to identify the CLCuKoV-Lu genome predicting the ‘most effective’ miRNA binding sites of the cotton miRNAs (Table 1). The G.hirsutum locus-derived ghr-miRNA sequences and the CLCuKoV-Lu genome-predicted transcripts (in FASTA format) were analyzed.
2.3. miRanda
miRanda is considered seed-matching, scoring scheme, dynamic programming and conservation consisting of three major steps to identify target sites since its release in 2003 [54]. miRanda has been used to predict the candidate target sites (CTSs) based on minimum free energy hypothesis. It works on RNA duplex dimerization and target sequence complementarity score [55]. The miRanda algorithm (written in C programming language) is a user-friendly web tool. The miRanda software was downloaded using the online source website (http://www.microrna.org/) (accessed on 23 March 2019). Prediction analysis was performed at default parameters. The miRanda program was set at MFE threshold: −20 Kcal/mol, score threshold: 140.00, gap open penalty: −9.000, gap extend penalty: −4.000 and scaling parameter: 4.00.
2.4. RNA22
The RNA22 algorithm is considered patter-recognition approach based on non-seed interaction for the prediction of target binding sites.[56]. The RNA22 is non-seed based algorithm and can be accessed web server (http://cm.jefferson.edu/rna22v1.0/) (accessed on January 26, 2019). Non-seed-based biological significant miRNA-mRNA interaction was calculated using target patterns and maximum folding energy (MFE)[57]. Site complementarity is also unique feature of RNA22 algorithms. Default parameters were set after loading cotton miRNA sequences and CLCuKoV-Lu genomic template: (63%), specificity (61%) and MFE (−15.00 Kcal/mol).
2.5. psRNATarget
The psRNATarget algorithm is considered small RNA target prediction using plant specific features coupled with seed region matching and scoring scheme. The web server is used to predict binding sites of the plant miRNAs listed in the web server on the basis of complementary scoring schema [58]. The psRNATarget web server has been updated and developed to report the inhibition pattern [59]. The fasta sequence of CLCuKoV-Lu genome was set in the server after selecting G. hirsutum miRNAs in the psRNATarget web server (http://plantgrn.noble.org/psRNATarget) (accessed on 26 October 2020). Target sites of cotton miRNAs were identified using default criteria of prediction: expectation cut-off value 6.5 and mode of inhibition ‘Cleavage’ was selected.
2.6. RNAhybrid
RNAhybrid is a seed-based flexible online web-based computational algorithm that is based on intermolecular hybridization predicting accurate miRNA binding sites in the target sequence. The RNAhybrid algorithm is MFE-based hybridization model. Site complementarity, MFE and seed match key features [60]. The fasta sequence of CLCuKoV-Lu genome and miRNAs were loaded after setting defaults parameter in the RNAhybrid web server http://bibiserv.techfak.uni-bielefeld.de/rnahybrid (accessed on 2 19 December 2020). MFE is a model feature and set at (−20.00 Kcal/mol).
2.7. RNAfold and RNAcofold
RNAfold is a web-based algorithm to identify accurate secondary structures of the target single-stranded (ss) miRNA precursor [61]. Precursor sequences was uploaded in the web server (http://rna.tbi.univie.ac.at/cgi-bin/RNAWebSuite/RNAfold.cgi) (accessed on September 6, 2022) under user-defined default settings. RNAcofold is a web-based algorithm and specifically intended to estimate the cofolding free energy (ΔG) of RNA duplex sequences. MFE and base-pairing pattern of miRNA-mRNA target duplex were estimated to evaluate miRNA-mRNA interaction [62]. FASTA sequences of unique duplex pair were set in the RNAcofold web server (http://rna.tbi.univie.ac.at/cgi-bin/RNAWebSuite/RNAcofold.cgi) (accessed May 6, 2022) under user-defined default settings.
2.8. Mapping of Network-based miRNA-Target Interactions
A Circos plot was generated using CIRCOS algorithm. We used the circos package v0.69-9R-Language [63].
2.9. Identification of miRNA Binding Site Distribution
The genomic transcript of monopartite CLCuKoV strains were downloaded from NCBI Genbank nucleotide database (accessed on 26 March 2018). The accession numbers of CLCuKoV strains under study are classified: GU385879.2, AM421522.1, AJ496286.1, HF549182.1, and FN5520001.1[18]. MEGA X (version 10.0.5) was used to align conservation of binding affinity of the predicted consensus miRNAs [64]. MUSCLE, a multiple sequence alignment algorithm was used to align a set of CLCuKoV genomic sequences [65]. The miRNA binding site sequences were mapped CLUSTALW algorithm[66].
2.9. Statistical Analysis
miRNA target prediction data were further processed for graphical representations using R-language (version 3.1.1, software version 3.5.1 [67].
2.10. Genome Annotation
pDRAW32 DNA analysis (AcaClone 1.1.147) is DNA annotation tool and was downloaded from the source web. It is used for ssDNA genomic annotation of CLCuKoV-Lu and further editing. The genome analysis and production of graphical output of CLCuKoV-Lu ORFs were generated.
3. Results
3.1. Cotton miRNAs-mRNA Interaction Pairs in CLCuKoV-Lu Genome
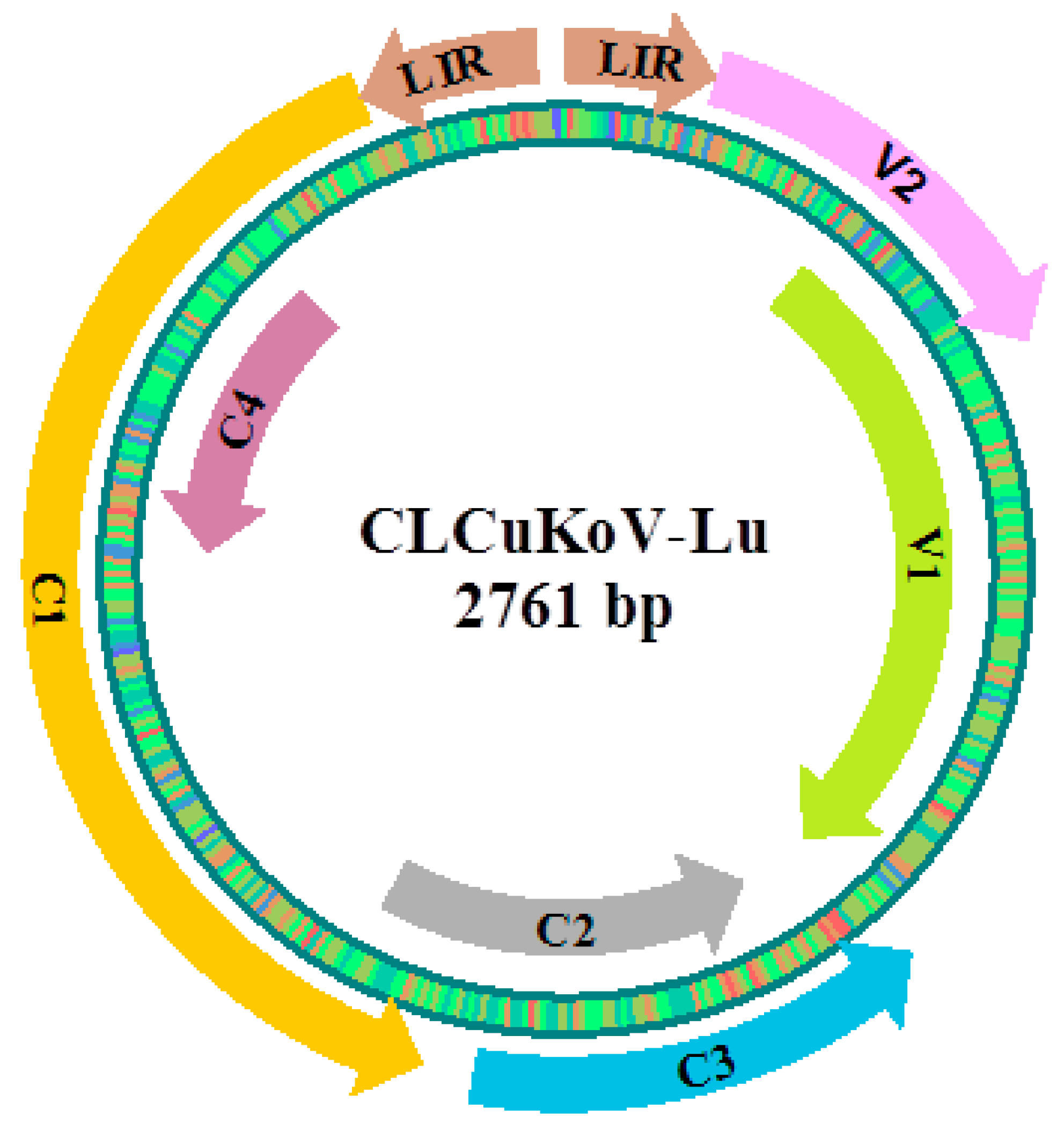
The genetic map of CLCuKoV-Lu has a single copy of circular ssDNA biomolecule that is composed of 2750 nucleotides (Figure 1).
Figure 1.
The genetic makeup of CLCuKoV-Lu is composed of six ORFs. The colored arrows are showing ORFs. The plus (+)-strand comprises virion-sense (encapsidated strand) ORFs (V1 and V2). The negative (−)-strand is composed of complementary-sense strand ORFs (C1, C2, C3 and C4). Replication of viral genome and transcription of the viral coding strands are governed by a non-coding large intergenic region (LIR).
Figure 1.
The genetic makeup of CLCuKoV-Lu is composed of six ORFs. The colored arrows are showing ORFs. The plus (+)-strand comprises virion-sense (encapsidated strand) ORFs (V1 and V2). The negative (−)-strand is composed of complementary-sense strand ORFs (C1, C2, C3 and C4). Replication of viral genome and transcription of the viral coding strands are governed by a non-coding large intergenic region (LIR).
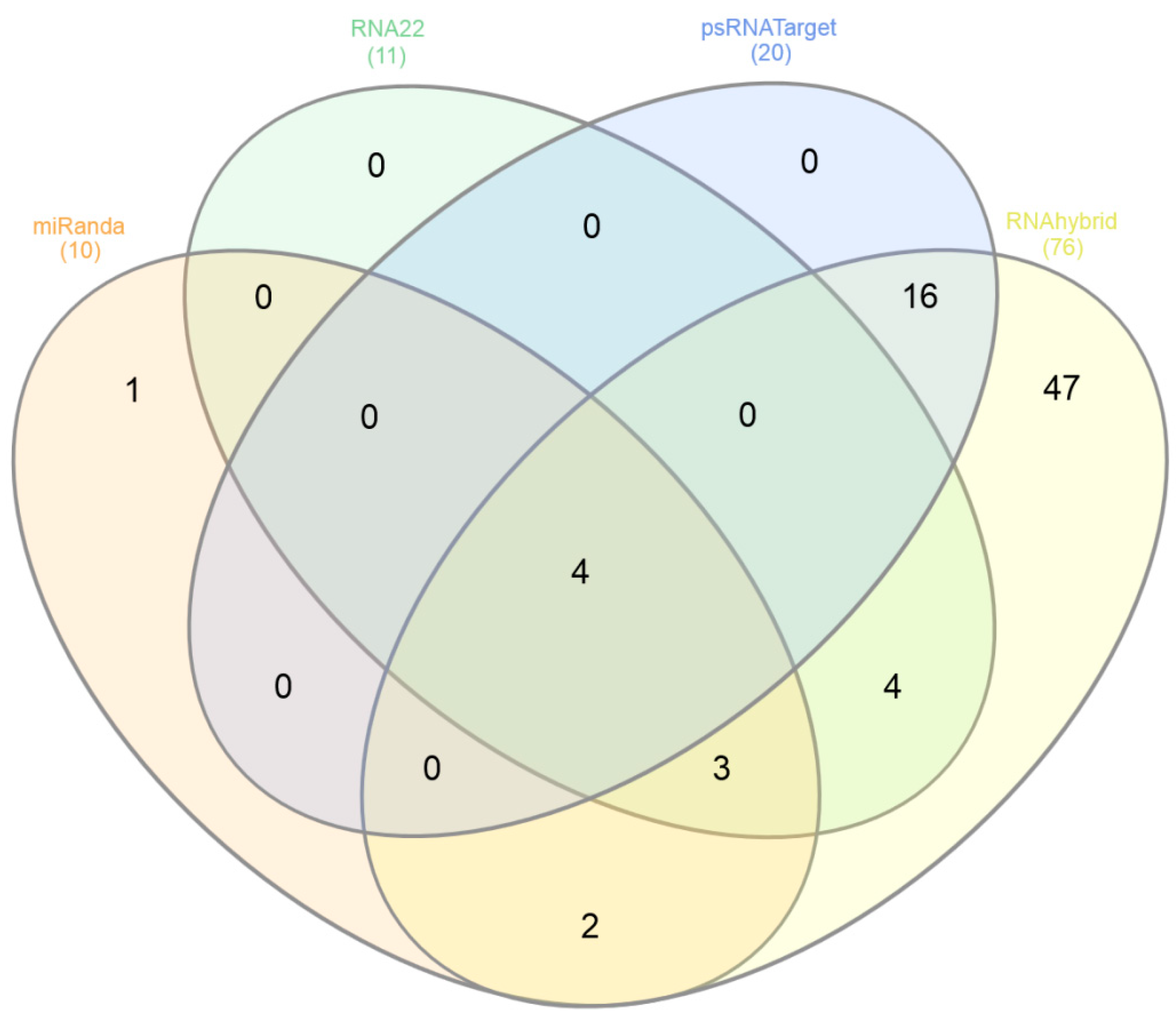
The CLCuKoV-Lu genome possesses six overlapping open reading frames (ORFs) that are shown to encode four major proteins. Using the miRBase web-based tool for miRNA genomics, and 3 distinct prediction algorithms (miRanda, RNA22 and psRNATarget), we search out cotton genome encoded miRNAs possess the capacity to target the CLCuKoV-Lu genome. For this we first accessed the newly emerged novel CLCuKoV-Lu genome from GenBank and the experimentally verified mature cotton locus-derived ghr-miRNAs were retrieved from the miRBase (version 22) to evaluate the binding strength of miRNA-mRNA target interactions. CLCuKoV-Lu genomic sequence potentially targeted by cotton miRNAs was searched by the miRanda algorithm which predicted 11 miRNA-mRNA target pairings. RNA22 predicted effective target binding sites of 11 cotton miRNAs at 11 genomic loci in the CLCuKoV-Lu genome. psRNATarget predicted cleavable target candidates: 20 cotton miRNAs and 26 loci. RNAhybrid 76 miRNA-mRNA target pairs (Figure 2) (Table S3-S4 and File S1).
Figure 2.
The Venn diagram showing miRNA-mRNA target pairs in the CLCuKoV-Lu genome. Four widely used computational databases (miRanda, RNA22, psRNATarget and RNAhybrid) were mined to estimate host-virus interaction pairs. Binding sites are key feature to determine degree of overlap exhibited by in silico tools used in this study. The intersection of four computational tools’ graph concludes four common ghr-miRNAs.
Figure 2.
The Venn diagram showing miRNA-mRNA target pairs in the CLCuKoV-Lu genome. Four widely used computational databases (miRanda, RNA22, psRNATarget and RNAhybrid) were mined to estimate host-virus interaction pairs. Binding sites are key feature to determine degree of overlap exhibited by in silico tools used in this study. The intersection of four computational tools’ graph concludes four common ghr-miRNAs.
3.2. V1 encoding coat protein (CP)
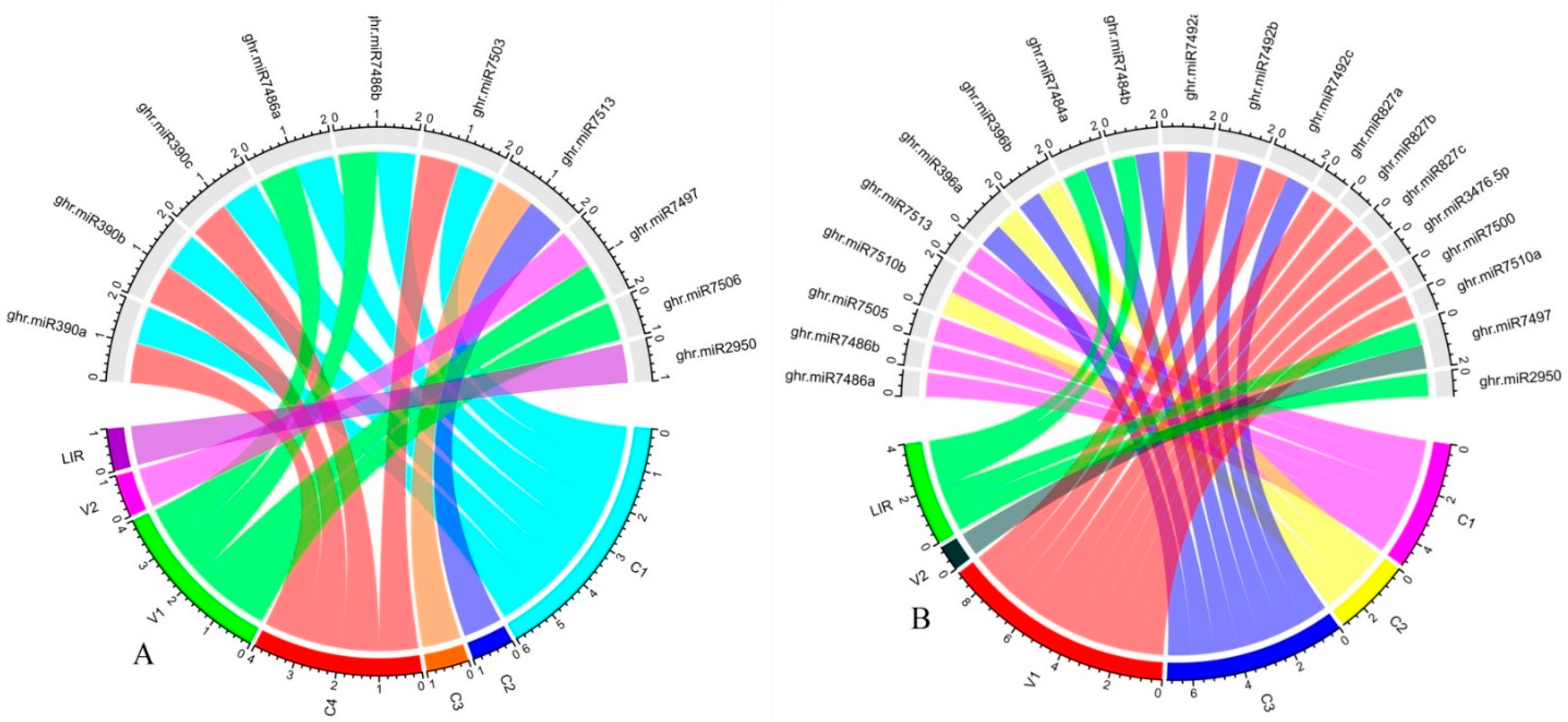
The begomoviral V1 ORF (291-1061 (770 nt) encodes the coat protein (CP), which is required for encapsidation of the begomoviral ssDNA genome into virions, vector-mediated transmission, and virus movement [68,69,70]. V1 was targeted by four cotton locus-derived ghr-miRNAs: ghr-miR7486 (a, b) (locus 846), ghr-miR7497 (locus 349) and ghr-miR7506 (locus 509), as indicated by miRanda (Figure 3A). RNA22 identified two miRNAs: ghr-miR169a (locus 691) and ghr-miR7512 (locus 917) (Figure 3B). The psRNATarget algorithm predicted ten miRNAs: ghr-miR827 (a, b and c) (locus 740), ghr-miR3476-5p (locus 765), ghr-miR7492 (a, b and c) (locus 901), ghr-miR7497 (locus 459 ghr-miR7500 (locus 674) and ghr-miR7510a (locus 805) (Figure 3C). RNAhybrid identified nine ghr-miRNAs: ghr-miR393, ghr-miR482 (a, b), ghr-miR7486 (a, b), ghr-miR7490, ghr-miR7504a, ghr-miR7510a and ghr-miR7512 at nucleotide positions 611, 581, 849, 670, 694 and 917, respectively (Figure 3D and Table 2) and (Table S3-S4 and File S1).
3.3. V2 encoding pre-coat protein
V2 ORF (131-487 nt) is composed of 356 nucleotides encoding a pre-coat protein. The V2 protein of the viral genome is involved in symptom development, movement, transmission and regulation [70,71,72,73]. The ghr-miR7497 targeted the V2 gene at (locus 349) and (loci 155, 459), as identified by the miRanda and psRNATarget algorithms, respectively (Figure 3A and 3C). RNAhybrid identified six ghr-miRNAs to target overlapping region of V1 and V2 ORF: ghr-miR164, ghr-miR479, ghr-miR3476-5p, ghr-miR7497, ghr-miR7498 and ghr-miR7507 at nucleotide positions 375, 371, 421, 362, 370 and 364, respectively (Figure 3D and Table 2) and (Table S3-S4 and File S1).
3.4. C1 encoding replication-associated protein (Rep)
The C1 ORF (1505-2581 (1076 bases) encodes a replication-associated protein (Rep), that is essential for ssDNA replication, transcription, regulation [18,74,75,76]. miRanda predicted six miRNAs: ghr-miR390 (a, b and c) (locus 2278), ghr-miR7486 (a, b) (locus 2488) and ghr-miR7503 (locus 2214) (Figure 3A). C1 was also targeted by six miRNAs: ghr-miR390 (a, b and c) (locus 2281), ghr-miR393 (locus 1735) and ghr-miR7486 (a, b) (locus 2488) by RNA22 (Figure 3B). Five potential miRNA candidates were detected for silencing the C1 gene by psRNATarget: ghr-miR7486 (a, b) (locus 2488), ghr-miR7505 (locus 2049), ghr-miR7510b (locus 1581) and ghr-miR7513 (locus 2540) (Figure 3C). The RNAhybrid algorithm predicted twenty one ghr-miRNAs: ghr-miR156 (a, b, c, d), ghr-miR162a, ghr-miR166b, ghr-miR169a, ghr-miR398, ghr-miR827 (a, b, c), ghr-miR2949-3p, ghr-miR3476-3p, ghr-miR7491, ghr-miR7492 (a, b, c), ghr-miR7500, ghr-miR7501, ghr-miR7505, and ghr-miR7506 at nucleotide positions 2500, 1665, 2038, 1820, 2469, 2473, 1655, 2470, 1743, 2566, 2467, 2428, 2036 and 1795, respectively (Figure 3D and Table 2) and (Table S3-S4 and File S1).
3.5. C2 encoding transcription activator protein (TrAP)
The C2 ORF of begomoviruses (1153-1599) (446 nt) encoded transcriptional activator protein (TrAP) that is essential for symptom development in infected plants [15,77,78,79]. Among the targeted genes of CLCuKoV-Lu, C2 has few binding sites of cotton miRNAs. miRanda predicted hybridization of ghr-miR7513 at locus 1350 in the overlapping region of C2 and C3 ORFs (Figure 3A). RNA22 predicted two miRNAs: ghr-miR7489 (locus 1408) and ghr-miR7513 (2540) in the C2 and C3 ORFs (Figure 3B). Three miRNA were predicted to have binding affinity with C2 by psRNATarget: ghr-miR396 (a, b) at locus 1250 in the C2 and C3 overlapping ORFs and ghr-miR7510b at locus 1581 in the C2 and C1 overlapping ORFs (Figure 3C). RNAhybrid predicted cotton ghr-miRNAs: ghr-miR394 (a, b), ghr-miR7504b in the C2 ORF (Figure 3D) and (Table 2, Table S3-S4 and File S1).
3.6. C3 encoding replication enhancer protein (REn)
The C3 ORF of begomoviruses (1058-1459) (401nt) encode a replication enhancer protein (REn) that is involved in replication of the viral ssDNA genome[80,81]. The psRNATarget algorithm identified five miRNAs in the C3 ORF: (locus 1250), ghr-miR7484 (a, b) (1081) and ghr-miR7492 (a, b and c) (1094). In addition psRNATarget predicted in the overlapping region of C2 and C3: ghr-miR396 (a, b) (Figure 3C). RNAhybrid predicted two cotton miRNAs targeting C3 ORF: ghr-miR7484 (a, b) at nucleotide position 1077. Further, RNAhybrid identified cotton miRNAs in the C2 and C3 overlapping region: ghr-miR167 (a, b), ghr-miR396 (a, b), ghr-miR2949 (a-5p, b, c), ghr-miR7489, ghr-miR7493, ghr-miR7494, ghr-miR7511 and ghr-miR7513 at genomic positions 1400, 1226, 1242, 1447, 1352, 1399 and 1350 respectively (Figure 4D) and (Table 2, Table S3-S4 and File S1).
3.7. C4 encoding transcription regulator protein
The C4 ORF of begomoviruses (2091-2429) (338 bases) encodes a transcription regulator protein that work as viral effector [76,82,83]. miRanda and RNA22 predicted binding of ghr-miR390 (a, b and c) at consensus genomic locus 2281. The ghr-miR7503 targeted the C1 gene to have binding affinity at genomic locus 2214, as indicated by miRanda (Figure 3A-3B). RNAhybrid predicted cotton miRNAs in the C4 and C1 overlapping region: ghr-miR160, ghr-miR172, ghr-miR390 (a, b and c), ghr-miR399d, ghr-miR7488, ghr-miR7495 (a, b), ghr-miR7503, ghr-miR7508, ghr-miR7509 and ghr-miR7510b at nucleotide positions 2044, 2177, 2196, 2047, 2258, 2276, 2214, 2282, 2162 and 2175, respectively (Figure 3D) and (Table 2, Table S3-S4 and File S1).
3.8. Large Intergenic Region
Large intergenic region (LIR) drives the transcriptional regulation of the V1 and C1 ORFs of CLCuKoV [20,21,84]. Four distinct prediction algorithms (miRanda, RNA22, psRNATarget and RNAhybrid) predicted hybridization binding site of ghr-miR2950 at consensus genomic locus 82 of CLCuKoV targeting LIR (Figure 3 A-D). In addition, LIR was targeted by three miRNAs: ghr-miR7484 (a, b) and ghr-miR7497, as indicated by psRNATarget (Figure 3C). RNAhybrid predicted cotton ghr-miRNAs in the LIR: ghr-miR399 (a, b, c), ghr-miR2948-5p, ghr-miR2950, at nucleotide positions 2633, 2616, 2611, and 82, respectively (Figure 3D) and (Table 2, Table S3-S4, File S1).
3.9. Predicting Common Cotton miRNAs
3.10. Predicting Consensual Cotton miRNAs
Of the 80 targeting mature G.hirsutum locus-derived ghr-miRNAs investigated, 7 G.hirsutum ghr-miRNAs: ghr-miR390 (a, b, c), ghr-miR7484 (a, b) and ghr-miR7503, ghr-miR7512 at nucleotide positions 2281, 1081, 2214 and 917, respectively were predicted to have potential binding sites within the CLCuKoV-Lu genome based on consensus genomic loci by at least two online miRNA prediction tools ( Table 2- 4). Out of 80 cotton miRNAs, three consensual G. hirsutum ghr-miRNAs: ghr-miR7486 (a, b) and ghr-miR7513 were predicted to have consensus genomic binding sites at nucleotide positions 2488 and 1350, respectively, was identified by union of consensus between the multiple algorithms used in this study. In this current study, only one G.hirsutum ghr-miRNA (ghr-miR2950) was predicted to have target binding site at common genomic position 82 by all the algorithms used (Table 2- 4, Figure 5). In terms of the CLCuKoV-Lu genome, ghr-miR2950 was predicted to target the non-coding LIR, ghr-miR2488 (a, b) were predicted to target the coding region C1 gene while ghr-miR7513 targets the overlapping region of ORFs C2/C3 (Figure 5 and Table 3).
Out of eleven consensual G. hirsutum ghr-miRNAs investigated, only one ghr-miRNA of G.hirsutum (ghr-miR2950 at nucleotide position 78-97), with a MFE of −27.38 Kcal/mol, was detected as top effective candidate (Table 4-5). The ‘cleavage’ efficacy of the ghr-miR2950 was verified against CLCuKoV-Lu by the RNAi-mediated suppression as concluded by Brodersen [85].
Figure 5.
Intersection plot of predicted consensus binding sites of cotton ghr-miRNAs by multiple algorithms. Binding sites were predicted at least three algorithms at consensus genomic positions targeting specific genes of CLCuKoV-Lu.
Figure 5.
Intersection plot of predicted consensus binding sites of cotton ghr-miRNAs by multiple algorithms. Binding sites were predicted at least three algorithms at consensus genomic positions targeting specific genes of CLCuKoV-Lu.
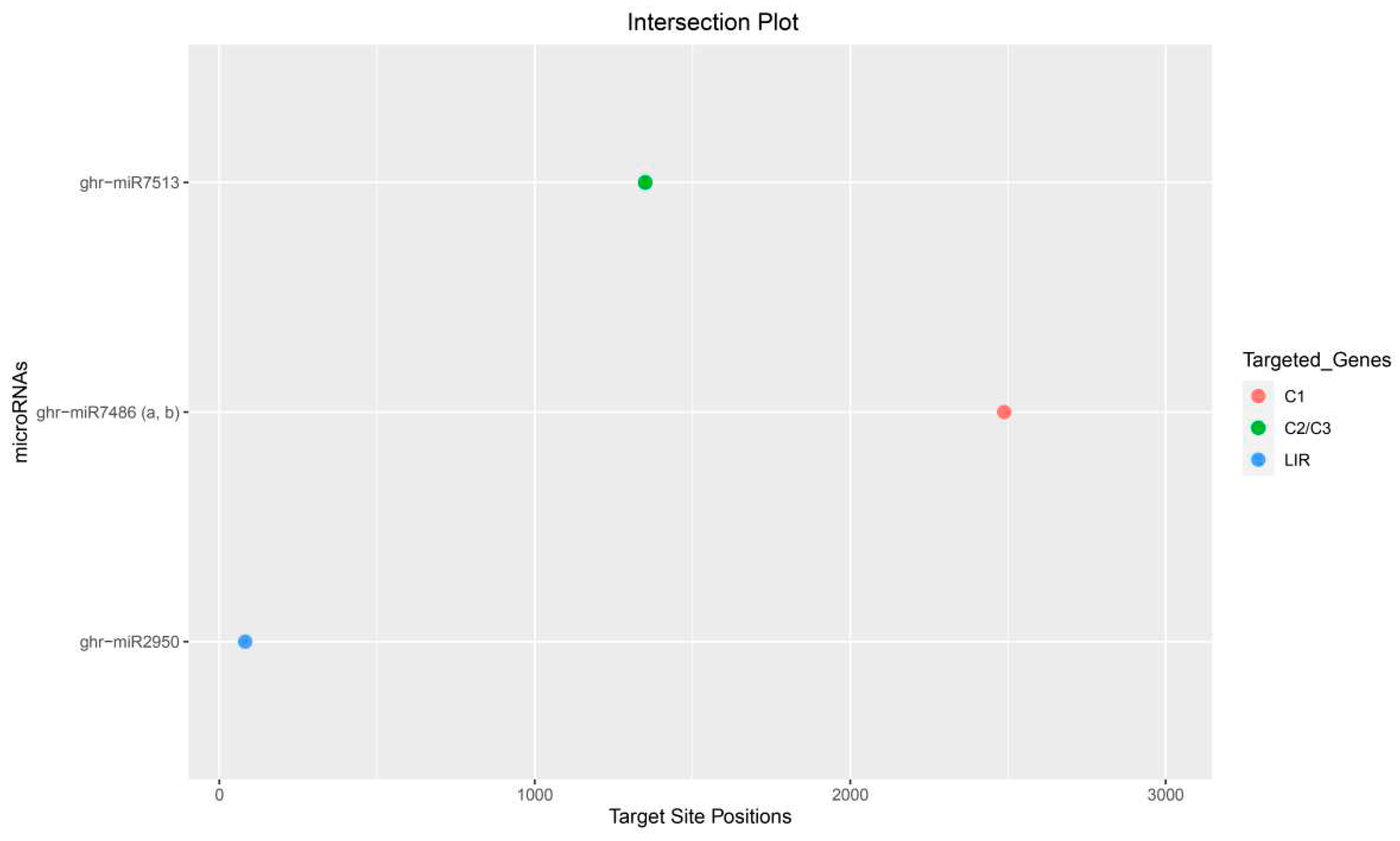

Table 3.
Target binding sites of predicted consensus cotton ghr-miRNAs in the CLCuKoV-Lu genome were detected by different algorithms. .
Table 3.
Target binding sites of predicted consensus cotton ghr-miRNAs in the CLCuKoV-Lu genome were detected by different algorithms. .
| Cotton miRNA |
Target Site miRanda |
Target Site RNA22 | Target Site psRNATarget |
Target Site RNAhybrid |
MFE * miRanda |
MFE ** RNA22 |
Expectation psRNATarget |
MFE * RNAhybrid |
| ghr-miR390 (a, b, c) | 2278 | 2281 | −21.48 | −18.00 | ||||
| ghr–miR2950 | 78 | 78 | 78 | 82 | −27.38 | −23.70 | 6.5 | −30.20 |
| ghr–miR7484 (a, b) | 1081 | 1077 | 6.5 | −20.90 | ||||
| ghr-miR7486 (a, b) | 2488/846 | 2488 | 2488 | 849 | −23.15/−29.28 | −21.48 | 5.0 | −30.70 |
| ghr-miR7503 | 2214 | 2214 | −23.35 | −27.00 | ||||
| ghr-miR7512 | 917 | 917 | −16.70 | −23.50 | ||||
| ghr-miR7513 | 1350 | 1350 | 1351 | −21.45 | −17.50 | −26.80 |
Table 4.
Binding sites of predicted consensus cotton ghr-miRNAs targeting different genes.
| miRNA ID | Accession ID | Mature Sequence (5′–3′) |
Target Genes ORF(s) |
Target Binding Locus Position |
|---|---|---|---|---|
| ghr-miR390a | MIMAT0005815 | AAGCUCAGGAGGGAUAGCGCC | C1/C4 | 2278–2298 |
| ghr-miR390b | MIMAT0005816 | AAGCUCAGGAGGGAUAGCGCC | C1/C4 | 2278–2298 |
| ghr-miR390c | MIMAT0005817 | AAGCUCAGGAGGGAUAGCGCC | C1/C4 | 2278–2298 |
| ghr-miR2950 | MIMAT0014348 | UGGUGUGCAGGGGGUGGAAUA | LIR | 78–97 |
| ghr-miR7484a | MIMAT0029124 | UUUGUAUAUUAGAUCAAAGAGCAA | C3 | 1081–1105 |
| ghr-miR7484b | MIMAT0029125 | UUUGUAUAUUAGAUCAAAGAGCAA | C3 | 1081–1105 |
| ghr-miR7486a | MIMAT0029127 | AAGGAAGCGCUUUGUCCACGUGGA | C1/V1 | 2488–2510/846-871 |
| ghr-miR7486b | MIMAT0029128 | AAGGAAGCGCUUUGUCCACGUGGA | C1/V1 | 2488–2510/871 |
| ghr-miR7503 | MIMAT0029150 | AGAUCGAUGGCUGAACAAGUUAGA | C4/C1 | 2214–2237 |
| ghr-miR7512 | MIMAT0029161 | UGCUACUUGUAGUUAUGCAUG | V1 | 917–938 |
| ghr-miR7513 | MIMAT0029162 | AAUCAGCCAGGAAUCGUUUGA | C2/C3 | 1350–1372 |
Table 5.
Features of predicted consensus G.hirsutum ghr-miRNA-target pairs.
| Cassava miRNA |
miRNA-Target Pair | Locus Position |
MFE (Kcal/mol) |
Score | Complementarity (%) |
Mode of Inhibition | |
|---|---|---|---|---|---|---|---|
| ghr-miR2950 | Query: 3' auaagGUGGGGGACGUGUGGu 5' | : | : | | | | | | | | | : Ref: 5' aataaCGCTCCC-GCACACTa 3' |
78-97 | −27.38 | 142 | 93.33 | Cleavage | |
| ghr-miR7486 (a, b) | Query: 3' aggUGCACCUGUUUCGCGAAGGAa 5' | : | | | : | | | | : | | | | | | | Ref: 5' tgaATTTGGG-AAAGTGCTTCCTc 3' |
2488-2510 | −23.15 | 171 | 90.00 | Cleavage | |
| ghr-miR7513 | Query: 3' agUUUGCUAA--GGACCGACUAa 5' : : | | | : | | | | | | | | | | | Ref: 5' atGGACGGTTGACGTGGCTGATg 3' |
1350-1372 | −21.45 | 162 | 85.00 | Cleavage | |
3.10.1. Visualization of miRNA Target
In order to integrate miRNA-mRNA target gene interaction analysis, we constructed Circos plots to identify novel host miRNA targets. The mapped mature cotton locus-derived ghr-miRNAs are depicted in the CLCuKoV-Lu genome.
3.10.2. Secondary Structure Analysis
In this study, we validated the efficacy of predicted consensus miRNAs by producing their secondary structures. The secondary structures were developed using manually curated cotton precursors. The MFE is a standard feature for stability of the map of the CLCuKoV-Lu RNA structurome. The current study validated and determined characteristic features of seven consensus precursors (Table 4).
Table 6.
Characteristics features of precursor of cotton locus-derived ghr-miRNAs were predicted.
| miRNA ID | Accession ID |
Length Precursor |
MFE*/Kcal/mol | AMFE** | MFEI*** | (G+C)% |
|---|---|---|---|---|---|---|
| ghr-MIR2950 | MI0013555 | 108 nt | −48.10 | −44.53 | −1.002 | 44.44 |
| ghr-MIR7486a | MI0024169 | 105 nt | −81.90 | −78.00 | −1.436 | 54.29 |
| ghr-MIR7486b | MI0024170 | 101 nt | −69.50 | −68.81 | −1.336 | 51.49 |
| ghr-MIR7513 | MI0024204 | 103 nt | −36.70 | −35.63 | −0.965 | 36.89 |
3.10.3. Free Energy (ΔG) Computation
In order to validate the predicted miRNAs we evaluated the Free Energy (ΔG) of duplex. The free energies (ΔG) of four seven consensual cotton locus-derived ghr-miRNAs were computed (Table 7).
3.10.4. Conserved Genomic Binding Sites Analysis
Among all the predicted genomic binding sites, highest level of conservation was obserevd in ghr-miR2950 (78-97) in different strains of CLCuKoV-Lu (Figure 6).
Figure 6.
A Circos plot showing cotton locus-derived ghr-miRNAs targeting CLCuKoV-Lu ORFs. CLCuKoV-Lu ORFs are represented with colored lines. (A) Showing interaction Circos map by seed-based algorithm miRanda. (B) Representing interaction Circos map by non-seed based algorithm psRNATarget.
Figure 6.
A Circos plot showing cotton locus-derived ghr-miRNAs targeting CLCuKoV-Lu ORFs. CLCuKoV-Lu ORFs are represented with colored lines. (A) Showing interaction Circos map by seed-based algorithm miRanda. (B) Representing interaction Circos map by non-seed based algorithm psRNATarget.
4. Discussion
CLCuD is a multifaceted disease. It has very complex molecular mechanism of development and progression. The current study has identified, by computational approach, and validated, by different algorithms, a combination of 3 cotton miRNAs in 80 mature miRNAs with potential to target CLCuKoV-Lu genome. For the last three decades, as a primitive virus, CLCuKoV can severely affect cotton production in Pakistan [9,11,12,86,87]. CLCuKoV-Bu infection triggered widespread gene silencing as an adaptive defence using cotton miRNAs. We filtered the false positive prediction data and considered to validate using the computational algorithms. We presented an equitable and integrative approach for the evaluation of predicted miRNA biological data at three different levels. Performance of data-driven algorithms is validated at union of intersections levels for interpretable predicted biological data. Assessing the potential cotton miRNA–mRNA targets interactions could be consensually validated (Figure 5). The present study predicts mature cotton genome encoded miRNAs (ghr-miR2950, ghr-miR7486 (a, b) targeting CLCuKoV-Lu to develop resistance in cotton cultivar. The predicted miRNAs have their interactions with the C1 and LIR of CLCuKoV-Lu. Our data indicated that evolutionary conserved cotton miRNA (ghr-miR2950) was selectively employed by CLCuKoV-Lu. Previous studies has reported host-virus interaction using online computational tools to identify binding affinity of genome encoded miRNAs in RTV1 [88], SCBV [89], SCYLV [90], ZYMV [91], SCBGAV [92], ToBRFV and PhCMoV [93], RYMV [94], MCMV [95] and ICMV-Ker[96]. We have reported similar findings using online computational tools for best target prediction against sugarcane and rubber tree viruses [88,89,90,92]. In this study, we employed in-silico algorithms for computational prediction, i.e. consensual target binding sites of ghr-miR2950 at locus (locus 78), ghr-miR7484 (a, b) at locus (1081) and ghr-miR7486 (a, b) at locus (2488), while TAPIR was predicted no binding site. Host delivered plant miRNAs are primary source of inducing the degradation in the viral target and are responsible for typical pattern of base pairing. This study demonstrated that CLCuKoV-Lu genomic components (C1 and LIR) are highly susceptible to be targeted by consensual miRNAs. Among the 80 cotton miRNAs investigated, ghr-miR2950 was predicted to have consensus genomic binding site within the LIR of the CLCuKoV-Lu genome (Figure 7). LIR is the key component of CLCuKoV genome to govern the bidirectional mode of transcription of C1 and V1 genes and works as a bidirectional promoter [20,21].
Union and intersection approaches of prediction are important to control false-positive prediction. Union level of prediction approach relies on combination of target prediction algorithms to find true targets. In this case, sensitivity level of predicted data was increased at the cost of lower level of specificity. Whereas an intersectional approach was predicted a different mode and combines two or more algorithmic tools at specific level at the cost of lower sensitivity [97,98,99,100]. Here, we present overview computational methods for miRNA target interaction prediction achieved the best outcomes (Figure 2 and 5). Previous studies on plant host-delivered miRNAs had also predicted the silencing viral genome using list of in silico tools. Experimental evaluation of potential plant host genome encoded miRNAs targeting different plant viruses have been reported successfully [31,51,101].
This study was designed an equal computational approach to predict cotton miRNAs in the CLCuKoV-Lu genome to combat of Begomovirus in cotton cultivars. Application of RNAi discovery to varietal improvement in cotton to control CLCuD infection is the highly recommended strategy to decrease yield losses [102,103,104,105]. However, gene pyramiding for enhanced resistance to CLCuKoV-Lu for sustainable upland cotton is highly complicated due to the complex allotetraploid nature of the cotton genome. Lower regeneration efficiency of cotton callus is another constraint for the development of CLCuD-resistant allotetraploid upland cotton. The differential expression profile of ghr-miR2950 was reported against at early stages of V.dahlliae infection [48,106,107]. The cotton miRNA (ghr-miR2950) exhibited a differential expression in PHYA1 RNAi cotton [108]. The ghr-miR2950 is a key role to encoding a gibberellin 3 hydroxylase [109]. This enzyme was experimentally validated to accumulate in fibers at high level and is responsible for fiber cell elongation using GA signaling in the PHYA1 RNAi cotton [108,110]. The ghr-miR2950 was characterized and identified during the growth and development of ovule and fiber in cotton. The ghr-miR2950 is also involved in inducing Root-knot nematodes (RKN) infection [111,112]. RNAi is a technique used to screen host-delivered factors for identifying various cellular functions against viruses [113,114,115]. In this study, we employed 80 experimentally validated mature cotton miRNAs with annotated targets in the CLCuKoV-Lu genome. In the current study, we cover bioinformatics workflows for CLCuKoV-Lu genome silencing provides evidence for the generation of antiviral agents. The design, construction and validation of the amiRNA-based construct was presented to harbor a modified miRNA/miRNA* duplex of the precursor (ghr-MIR-2950) (Figure 8). Furthermore, we summarized current knowledge to demonstrate a novel methodology to minimize the antiviral effects of host genome encoded miRNAs against CLCuKoV-Lu.
5. Conclusions and Recommendations
CLCuKoV-Lu is an emerging pathogen associated with the CLCuD pandemic. CLCuD reduces yield and quality of all cotton cultivars in Pakistan. Overall this study provides an optimized prediction tools and parameters to identify the best-candidate miRNAs against CLCuKoV-Lu begomovirus. Prior to molecular cloning, we employed in silico tools and approaches for predicting effective binding affinity of mature candidate cotton miRNAs in the CLCuKoV-Lu genome. Among the 80 cotton miRNAs investigated, we predicted 3 consensual cotton locus-derived ghr-miRNAs that have miRNA-mRNA hybridization pairings of allotetraploid upland cotton miRNAs within the CLCuKoV-Lu. We identified ghr-miR2950 (out of 80 miRNAs) as the miRNA with highest affinity for the CLCuKoV-Lu genome. By way of comparison to other CLCuKoV strains, we have also established a reliable microRNA-binding region in the CLCuKoV genome that we predicted in course of an analysis of multiple sequence alignment. Hence, our efforts in mapping the cotton miRNA–mRNA target interaction may be aided in untangling molecular underpinnings of hereditary and CLCuD.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: List of mature G.hirsutum ghr-miRNA; Table S2: List of G. hirsutum precursor miRNAs; Table S3: Identification of binding sites of ghr-miRNAs in the CLCuKoV-Lu genome; Table S4: Gene wise analysis of predicted miRNA and File S1: Prediction results analyzed by computational algorithms.
Author Contributions
M.A.A., J.K.B and N.Y. conceived the original idea of the work. All the authors performed, analyzed and interpreted the in silico data. M.A.A wrote the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Central Public-interest Scientific Institution Basal Research Fund (1630052023003). The research work was completed under a MoU among the three institutes mentioned.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors acknowledge the IPFP of HEC and TYSP of China to support M.A.A. The authors acknowledge to Hainan Provincial Natural Science Foundation (321RC640), National Key R&D Program of China (2019YFD1000500) for partial financial support. The research was conducted in collaboration with Khwaja Fareed University of Engineering and Information Technology, Chinese Academy of Tropical Agricultural Sciences and University of Arizona under a MoU.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Li, S.; Geng, S.; Pang, B.; Zhao, J.; Huang, Y.; Rui, C.; Cui, J.; Jiao, Y.; Zhang, R.; Gao, W. Revealing Genetic Differences in Fiber Elongation between the Offspring of Sea Island Cotton and Upland Cotton Backcross Populations Based on Transcriptome and Weighted Gene Coexpression Networks. Genes 2022, 13, 954. [Google Scholar] [CrossRef]
- Jan, M.; Liu, Z.; Guo, C.; Sun, X. Molecular Regulation of Cotton Fiber Development: A Review. International Journal of Molecular Sciences 2022, 23, 5004. [Google Scholar] [CrossRef]
- Townsend, T. World natural fibre production and employment. In Handbook of natural fibres; Elsevier: 2020; pp. 15-36.
- Beasley, J. The origin of American tetraploid Gossypium species. The American Naturalist 1940, 74, 285–286. [Google Scholar] [CrossRef]
- COTTON, C.S.I. IV. CHROMOSOME CONJUGATION IN INTERSPECIFIC HYBRIDS. Jour. Genetics 1937, 34, 5–42. [Google Scholar]
- Zhang, T.; Hu, Y.; Jiang, W.; Fang, L.; Guan, X.; Chen, J.; Zhang, J.; Saski, C.A.; Scheffler, B.E.; Stelly, D.M. Sequencing of allotetraploid cotton (Gossypium hirsutum L. acc. TM-1) provides a resource for fiber improvement. Nature biotechnology 2015, 33, 531–537. [Google Scholar] [CrossRef]
- Briddon, R.W. Cotton leaf curl disease, a multicomponent begomovirus complex. Molecular Plant Pathology 2003, 4, 427–434. [Google Scholar] [CrossRef]
- Rahman, M.-u.-. .; Khan, A.Q.; Rahmat, Z.; Iqbal, M.A.; Zafar, Y. Genetics and genomics of cotton leaf curl disease, its viral causal agents and whitefly vector: a way forward to sustain cotton fiber security. Frontiers in Plant Science 2017, 8, 1157. [Google Scholar] [CrossRef]
- Zubair, M.; Zaidi, S.S.-e.-A.; Shakir, S.; Farooq, M.; Amin, I.; Scheffler, J.A.; Scheffler, B.E.; Mansoor, S. Multiple begomoviruses found associated with cotton leaf curl disease in Pakistan in early 1990 are back in cultivated cotton. Scientific reports 2017, 7, 1–11. [Google Scholar] [CrossRef]
- Biswas, K.K.; Bhattacharyya, U.K.; Palchoudhury, S.; Balram, N.; Kumar, A.; Arora, R.; Sain, S.K.; Kumar, P.; Khetarpal, R.K.; Sanyal, A. Dominance of recombinant cotton leaf curl Multan-Rajasthan virus associated with cotton leaf curl disease outbreak in northwest India. PLoS One 2020, 15, e0231886. [Google Scholar] [CrossRef]
- Amrao, L.; Amin, I.; Shahid, M.S.; Briddon, R.W.; Mansoor, S. Cotton leaf curl disease in resistant cotton is associated with a single begomovirus that lacks an intact transcriptional activator protein. Virus research 2010, 152, 153–163. [Google Scholar] [CrossRef]
- Ashraf, M.A.; Shahid, A.A.; Mohamed, B.B.; Dahab, A.A.; Bajwa, K.S.; Rao, A.Q.; Khan, M.A.U.; Ilyas, M.; Haider, M.S.; Husnain, T. Molecular characterization and phylogenetic analysis of a variant of highly infectious cotton leaf curl Burewala virus associated with CLCuD from Pakistan. Australian Journal of Crop Science 2013, 7, 1113–1122. [Google Scholar]
- Shuja, M.N.; Tahir, M.; Briddon, R.W. Occurrence of a recombinant molecule carrying sequences derived from an alphasatellite and the helper virus in cotton affected with cotton leaf curl disease. Tropical Plant Pathology 2017, 42, 397–402. [Google Scholar] [CrossRef]
- Chen, T.; Tang, Y.; Zhao, R.; He, Z. Identification of the cryptic species of Bemisia tabaci transmitting Cotton leaf curl Multan virus. Acta Phytophylacica Sinica 2016, 43, 91–98. [Google Scholar]
- Nigam, D. Genomic variation and diversification in begomovirus genome in implication to host and vector adaptation. Plants 2021, 10, 1706. [Google Scholar] [CrossRef]
- Chen, T.; Saeed, Q.; He, Z.; Lu, L. Transmission efficiency of Cotton leaf curl Multan virus by three cryptic species of Bemisia tabaci complex in cotton cultivars. PeerJ 2019, 7, e7788. [Google Scholar] [CrossRef]
- Paredes-Montero, J.R.; Hameed, U.; Zia-Ur-Rehman, M.; Rasool, G.; Haider, M.S.; Herrmann, H.-W.; Brown, J.K. Demographic expansion of the predominant Bemisia tabaci (Gennadius)(Hemiptera: Aleyrodidae) mitotypes associated with the Cotton leaf curl virus epidemic in Pakistan. Annals of the Entomological Society of America 2019, 112, 265–280. [Google Scholar] [CrossRef]
- Brown, J.K.; Zerbini, F.M.; Navas-Castillo, J.; Moriones, E.; Ramos-Sobrinho, R.; Silva, J.C.; Fiallo-Olivé, E.; Briddon, R.W.; Hernández-Zepeda, C.; Idris, A. Revision of Begomovirus taxonomy based on pairwise sequence comparisons. 2015.
- Kumar, J.; Kumar, A.; Roy, J.; Tuli, R.; Khan, J. Identification and molecular characterization of begomovirus and associated satellite DNA molecules infecting Cyamopsis tetragonoloba. Virus Genes 2010, 41, 118–125. [Google Scholar] [CrossRef]
- Ashraf, M.A.; Shahid, A.A.; Rao, A.Q.; Bajwa, K.S.; Husnain, T. Functional characterization of a bidirectional plant promoter from cotton leaf curl Burewala virus using an Agrobacterium-mediated transient assay. Viruses 2014, 6, 223–242. [Google Scholar] [CrossRef]
- Ashraf, M.A.; Shahid, A.A.; Rao, A.Q.; Brown, J.K.; Husnain, T. Development and Evaluation of the Cotton Leaf Curl Kokhran Virus-Burewala Bidirectional Promoter for Enhanced Cry1Ac Endotoxin Expression in Bt Transgenic Cotton. Applied Sciences 2022, 12, 11275. [Google Scholar] [CrossRef]
- Bornancini, V.A.; Irazoqui, J.M.; Flores, C.R.; Vaghi Medina, C.G.; Amadio, A.F.; López Lambertini, P.M. Reconstruction and characterization of full-length begomovirus and alphasatellite genomes infecting pepper through metagenomics. Viruses 2020, 12, 202. [Google Scholar] [CrossRef]
- Palchoudhury, S.; Khare, V.; Balram, N.; Bhattacharyya, U.; Das, S.; Shukla, P.; Chakraborty, P.; Biswas, K. A multiplex polymerase chain reaction for the simultaneous detection of the virus and satellite components associated with cotton leaf curl begomovirus disease complex. Journal of Virological Methods 2022, 300, 114369. [Google Scholar] [CrossRef]
- Gawande, S.; Raghavendra, K.; Monga, D.; Nagrale, D.; Prabhulinga, T.; Hiremani, N.; Meshram, M.; Kranthi, S.; Gokte-Narkhedkar, N.; Waghmare, V. Development of Loop Mediated Isothermal Amplification (LAMP): A new tool for rapid diagnosis of cotton leaf curl viral disease. Journal of Virological Methods 2022, 114541. [Google Scholar] [CrossRef] [PubMed]
- Inoue-Nagata, A.K.; Albuquerque, L.C.; Rocha, W.B.; Nagata, T. A simple method for cloning the complete begomovirus genome using the bacteriophage φ29 DNA polymerase. Journal of virological methods 2004, 116, 209–211. [Google Scholar] [CrossRef] [PubMed]
- Shafiq, M.; Iqbal, Z.; Ali, I.; Abbas, Q.; Mansoor, S.; Briddon, R.W.; Amin, I. Real-time quantitative PCR assay for the quantification of virus and satellites causing leaf curl disease in cotton in Pakistan. Journal of virological methods 2017, 248, 54–60. [Google Scholar] [CrossRef]
- Haible, D.; Kober, S.; Jeske, H. Rolling circle amplification revolutionizes diagnosis and genomics of geminiviruses. Journal of virological methods 2006, 135, 9–16. [Google Scholar] [CrossRef]
- Ahmed, N.; Amin, I.; Zaidi, S.S.-e.-A.; Rahman, S.U.; Farooq, M.; Fauquet, C.M.; Mansoor, S. Circular DNA enrichment sequencing reveals the viral/satellites genetic diversity associated with the third epidemic of cotton leaf curl disease. Biology Methods and Protocols 2021, 6, bpab005. [Google Scholar] [CrossRef] [PubMed]
- Pan, L.-L.; Cui, X.-Y.; Chen, Q.-F.; Wang, X.-W.; Liu, S.-S. Cotton leaf curl disease: which whitefly is the vector? Phytopathology 2018, 108, 1172–1183. [Google Scholar] [CrossRef] [PubMed]
- Deng, Z.; Ma, L.; Zhang, P.; Zhu, H. Small RNAs Participate in Plant–Virus Interaction and Their Application in Plant Viral Defense. International Journal of Molecular Sciences 2022, 23, 696. [Google Scholar] [CrossRef]
- Zhou, L.; Yuan, Q.; Ai, X.; Chen, J.; Lu, Y.; Yan, F. Transgenic Rice Plants Expressing Artificial miRNA Targeting the Rice Stripe Virus MP Gene Are Highly Resistant to the Virus. Biology 2022, 11, 332. [Google Scholar] [CrossRef]
- Villegas-Estrada, B.; Sánchez, M.A.; Valencia-Jiménez, A. Foliar Infiltration of Virus-Derived Small Hairpin RNAs Triggers the RNAi Mechanism against the Cucumber Mosaic Virus. International journal of molecular sciences 2022, 23, 4938. [Google Scholar] [CrossRef]
- Akbar, S.; Wei, Y.; Zhang, M.-Q. RNA Interference: Promising Approach to Combat Plant Viruses. International Journal of Molecular Sciences 2022, 23, 5312. [Google Scholar] [CrossRef]
- Kim, Y.J.; Zheng, B.; Yu, Y.; Won, S.Y.; Mo, B.; Chen, X. The role of Mediator in small and long noncoding RNA production in Arabidopsis thaliana. The EMBO journal 2011, 30, 814–822. [Google Scholar] [CrossRef] [PubMed]
- Fang, X.; Cui, Y.; Li, Y.; Qi, Y. Transcription and processing of primary microRNAs are coupled by Elongator complex in Arabidopsis. Nature Plants 2015, 1, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Manavella, P.A.; Koenig, D.; Weigel, D. Plant secondary siRNA production determined by microRNA-duplex structure. Proceedings of the National Academy of Sciences 2012, 109, 2461–2466. [Google Scholar] [CrossRef]
- Hu, B.; Zhong, L.; Weng, Y.; Peng, L.; Huang, Y.; Zhao, Y.; Liang, X.-J. Therapeutic siRNA: state of the art. Signal transduction and targeted therapy 2020, 5, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Hung, Y.-H.; Slotkin, R.K. The initiation of RNA interference (RNAi) in plants. Current Opinion in Plant Biology 2021, 61, 102014. [Google Scholar] [CrossRef] [PubMed]
- Reinhart, B.J.; Weinstein, E.G.; Rhoades, M.W.; Bartel, B.; Bartel, D.P. MicroRNAs in plants. Genes & development 2002, 16, 1616–1626. [Google Scholar]
- Cui, J.; You, C.; Chen, X. The evolution of microRNAs in plants. Current opinion in plant biology 2017, 35, 61–67. [Google Scholar] [CrossRef]
- Carrington, J.C.; Ambros, V. Role of microRNAs in plant and animal development. Science 2003, 301, 336–338. [Google Scholar] [CrossRef]
- Wang, M.; Sun, R.; Li, C.; Wang, Q.; Zhang, B. MicroRNA expression profiles during cotton (Gossypium hirsutum L) fiber early development. Scientific reports 2017, 7, 1–13. [Google Scholar] [CrossRef]
- Zhao, T.; Xu, X.; Wang, M.; Li, C.; Li, C.; Zhao, R.; Zhu, S.; He, Q.; Chen, J. Identification and profiling of upland cotton microRNAs at fiber initiation stage under exogenous IAA application. BMC genomics 2019, 20, 1–15. [Google Scholar]
- Zhang, J.; Ma, M.; Liu, Y.; Ismayil, A. Plant Defense and Viral Counter-Defense during Plant–Geminivirus Interactions. Viruses 2023, 15, 510. [Google Scholar] [PubMed]
- Li, J.; Hull, J.J.; Liang, S.; Wang, Q.; Chen, L.; Zhang, Q.; Wang, M.; Mansoor, S.; Zhang, X.; Jin, S. Genome-wide analysis of cotton miRNAs during whitefly infestation offers new insights into plant-herbivore interaction. International journal of molecular sciences 2019, 20, 5357. [Google Scholar]
- Zhang, T.; Zhao, Y.-L.; Zhao, J.-H.; Wang, S.; Jin, Y.; Chen, Z.-Q.; Fang, Y.-Y.; Hua, C.-L.; Ding, S.-W.; Guo, H.-S. Cotton plants export microRNAs to inhibit virulence gene expression in a fungal pathogen. Nature plants 2016, 2, 1–6. [Google Scholar]
- Wang, W.; Liu, D.; Chen, D.; Cheng, Y.; Zhang, X.; Song, L.; Hu, M.; Dong, J.; Shen, F. MicroRNA414c affects salt tolerance of cotton by regulating reactive oxygen species metabolism under salinity stress. RNA biology 2019, 16, 362–375. [Google Scholar]
- He, X.; Sun, Q.; Jiang, H.; Zhu, X.; Mo, J.; Long, L.; Xiang, L.; Xie, Y.; Shi, Y.; Yuan, Y. Identification of novel microRNAs in the Verticillium wilt-resistant upland cotton variety KV-1 by high-throughput sequencing. SpringerPlus 2014, 3, 1–9. [Google Scholar]
- Dong, Z.; Zhang, J.; Zhu, Q.; Zhao, L.; Sui, S.; Li, Z.; Zhang, Y.; Wang, H.; Tian, D.; Zhao, Y. Identification of microRNAs involved in drought stress responses in early-maturing cotton by high-throughput sequencing. Genes & genomics 2018, 40, 305–314. [Google Scholar]
- Zhang, H.; Feng, H.; Lu, X.; Wang, C.; Yang, W.; Li, F. An asymmetric bulge enhances artificial microRNA-mediated virus resistance. Plant Biotechnology Journal 2020, 18, 608. [Google Scholar]
- Niu, Q.-W.; Lin, S.-S.; Reyes, J.L.; Chen, K.-C.; Wu, H.-W.; Yeh, S.-D.; Chua, N.-H. Expression of artificial microRNAs in transgenic Arabidopsis thaliana confers virus resistance. Nature biotechnology 2006, 24, 1420–1428. [Google Scholar] [PubMed]
- Kozomara, A.; Birgaoanu, M.; Griffiths-Jones, S. miRBase: from microRNA sequences to function. Nucleic acids research 2019, 47, D155–D162. [Google Scholar] [PubMed]
- Sayers, E.W.; Beck, J.; Bolton, E.E.; Bourexis, D.; Brister, J.R.; Canese, K.; Comeau, D.C.; Funk, K.; Kim, S.; Klimke, W. Database resources of the national center for biotechnology information. Nucleic acids research 2021, 49, D10. [Google Scholar] [CrossRef]
- Enright, A.; John, B.; Gaul, U.; Tuschl, T.; Sander, C.; Marks, D. MicroRNA targets in Drosophila. Genome biology 2003, 4, 1–27. [Google Scholar] [CrossRef]
- John, B.; Enright, A.J.; Aravin, A.; Tuschl, T.; Sander, C.; Marks, D.S. Human microRNA targets. PLoS biology 2004, 2, e363. [Google Scholar]
- Miranda, K.C.; Huynh, T.; Tay, Y.; Ang, Y.-S.; Tam, W.-L.; Thomson, A.M.; Lim, B.; Rigoutsos, I. A pattern-based method for the identification of MicroRNA binding sites and their corresponding heteroduplexes. Cell 2006, 126, 1203–1217. [Google Scholar] [PubMed]
- Loher, P.; Rigoutsos, I. Interactive exploration of RNA22 microRNA target predictions. Bioinformatics 2012, 28, 3322–3323. [Google Scholar] [CrossRef]
- Dai, X.; Zhao, P.X. psRNATarget: a plant small RNA target analysis server. Nucleic acids research 2011, 39, W155–W159. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Zhuang, Z.; Zhao, P.X. psRNATarget: a plant small RNA target analysis server (2017 release). Nucleic acids research 2018, 46, W49–W54. [Google Scholar] [CrossRef] [PubMed]
- Krüger, J.; Rehmsmeier, M. RNAhybrid: microRNA target prediction easy, fast and flexible. Nucleic acids research 2006, 34, W451–W454. [Google Scholar] [CrossRef]
- Lorenz, R.; Bernhart, S.H.; Höner zu Siederdissen, C.; Tafer, H.; Flamm, C.; Stadler, P.F.; Hofacker, I.L. ViennaRNA Package 2.0. Algorithms for molecular biology 2011, 6, 1–14. [Google Scholar]
- Bernhart, S.H.; Tafer, H.; Mückstein, U.; Flamm, C.; Stadler, P.F.; Hofacker, I.L. Partition function and base pairing probabilities of RNA heterodimers. Algorithms for Molecular Biology 2006, 1, 1–10. [Google Scholar]
- Krzywinski, M.; Schein, J.; Birol, I.; Connors, J.; Gascoyne, R.; Horsman, D.; Jones, S.J.; Marra, M.A. Circos: an information aesthetic for comparative genomics. Genome research 2009, 19, 1639–1645. [Google Scholar] [CrossRef] [PubMed]
- MEGA, X. molecular evolutionary genetics analysis across computing platforms; S Kumar, G Stecher, M Li, C Knyaz, K Tamura. Molecular Biology and Evolution 2018, 35, 1547–1549. [Google Scholar]
- Edgar, R.C. MUSCLE: a multiple sequence alignment method with reduced time and space complexity. BMC bioinformatics 2004, 5, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Błażewicz, J.; Formanowicz, P.; Kasprzak, M.; Michalak, K.; Wierzejewski, P. Clustal W algorithm for multiple sequence alignment revisited. Foundations of Computing and Decision Sciences 2001, 26, 189–196. [Google Scholar]
- Gandrud, C. Reproducible research with R and RStudio; Chapman and Hall/CRC: 2018.
- Fan, Y.-Y.; Zhong, Y.-W.; Zhao, J.; Chi, Y.; Bouvaine, S.; Liu, S.-S.; Seal, S.E.; Wang, X.-W. Bemisia tabaci vesicle-associated membrane protein 2 interacts with begomoviruses and plays a role in virus acquisition. Cells 2021, 10, 1700. [Google Scholar] [CrossRef]
- Götz, M.; Popovski, S.; Kollenberg, M.; Gorovits, R.; Brown, J.K.; Cicero, J.M.; Czosnek, H.; Winter, S.; Ghanim, M. Implication of Bemisia tabaci heat shock protein 70 in begomovirus-whitefly interactions. Journal of virology 2012, 86, 13241–13252. [Google Scholar] [CrossRef]
- Poornima Priyadarshini, C.; Ambika, M.; Tippeswamy, R.; Savithri, H. Functional characterization of coat protein and V2 involved in cell to cell movement of Cotton leaf curl Kokhran virus-Dabawali. PLoS One 2011, 6, e26929. [Google Scholar] [CrossRef]
- Bahari, A.; Castillo, A.G.; Safaie, N.; Bejarano, E.R.; Luna, A.P.; Shams-Bakhsh, M. Functional analysis of V2 protein of Beet curly top Iran virus. Plants 2022, 11, 3351. [Google Scholar] [CrossRef]
- Li, M.; Li, C.; Jiang, K.; Li, K.; Zhang, J.; Sun, M.; Wu, G.; Qing, L. Characterization of pathogenicity-associated V2 protein of tobacco curly shoot virus. International Journal of Molecular Sciences 2021, 22, 923. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, Y.; Gong, Q.; Ismayil, A.; Yuan, Y.; Lian, B.; Jia, Q.; Han, M.; Deng, H.; Hong, Y. Geminiviral V2 protein suppresses transcriptional gene silencing through interaction with AGO4. Journal of virology 2019, 93, e01675–01618. [Google Scholar] [CrossRef]
- Hipp, K.; Rau, P.; Schäfer, B.; Gronenborn, B.; Jeske, H. The RXL motif of the African cassava mosaic virus Rep protein is necessary for rereplication of yeast DNA and viral infection in plants. Virology 2014, 462, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Zhang, Q.; Hong, J.; Li, Z.; Zhang, X.; Zhou, X. Cryo-EM structure of a begomovirus geminate particle. International Journal of Molecular Sciences 2019, 20, 1738. [Google Scholar] [CrossRef] [PubMed]
- Torres-Herrera, S.I.; Romero-Osorio, A.; Moreno-Valenzuela, O.; Pastor-Palacios, G.; Cardenas-Conejo, Y.; Ramírez-Prado, J.H.; Riego-Ruiz, L.; Minero-García, Y.; Ambriz-Granados, S.; Argüello-Astorga, G.R. A lineage of begomoviruses encode Rep and AC4 proteins of enigmatic ancestry: hints on the evolution of geminiviruses in the New World. Viruses 2019, 11, 644. [Google Scholar] [CrossRef] [PubMed]
- Guerrero, J.; Regedanz, E.; Lu, L.; Ruan, J.; Bisaro, D.M.; Sunter, G. Manipulation of the plant host by the geminivirus AC2/C2 protein, a central player in the infection cycle. Frontiers in plant science 2020, 11, 591. [Google Scholar] [CrossRef]
- Rosas-Díaz, T.; Macho, A.P.; Beuzón, C.R.; Lozano-Durán, R.; Bejarano, E.R. The C2 protein from the geminivirus Tomato yellow leaf curl Sardinia virus decreases sensitivity to jasmonates and suppresses jasmonate-mediated defences. Plants 2016, 5, 8. [Google Scholar] [CrossRef]
- Veluthambi, K.; Sunitha, S. Targets and mechanisms of geminivirus silencing suppressor protein AC2. Frontiers in Microbiology 2021, 12, 645419. [Google Scholar] [CrossRef]
- Settlage, S.B.; See, R.G.; Hanley-Bowdoin, L. Geminivirus C3 protein: replication enhancement and protein interactions. Journal of virology 2005, 79, 9885–9895. [Google Scholar] [CrossRef]
- Fondong, V.N. Geminivirus protein structure and function. Molecular plant pathology 2013, 14, 635–649. [Google Scholar] [CrossRef]
- Medina-Puche, L.; Orílio, A.F.; Zerbini, F.M.; Lozano-Durán, R. Small but mighty: Functional landscape of the versatile geminivirus-encoded C4 protein. PLoS Pathogens 2021, 17, e1009915. [Google Scholar] [CrossRef]
- Dai, K.-W.; Tsai, Y.-T.; Wu, C.-Y.; Lai, Y.-C.; Lin, N.-S.; Hu, C.-C. Identification of Crucial Amino Acids in Begomovirus C4 Proteins Involved in the Modulation of the Severity of Leaf Curling Symptoms. Viruses 2022, 14, 499. [Google Scholar] [CrossRef]
- Khan, Z.A.; Abdin, M.Z.; Khan, J.A. Functional characterization of a strong bi-directional constitutive plant promoter isolated from cotton leaf curl Burewala virus. PLoS One 2015, 10, e0121656. [Google Scholar] [CrossRef] [PubMed]
- Brodersen, P.; Sakvarelidze-Achard, L.; Bruun-Rasmussen, M.; Dunoyer, P.; Yamamoto, Y.Y.; Sieburth, L.; Voinnet, O. Widespread translational inhibition by plant miRNAs and siRNAs. Science 2008, 320, 1185–1190. [Google Scholar] [CrossRef] [PubMed]
- Siddique, Z.; Akhtar, K.P.; Hameed, A.; Sarwar, N.; Imran-Ul-Haq; Khan, S. A. Biochemical alterations in leaves of resistant and susceptible cotton genotypes infected systemically by cotton leaf curl Burewala virus. Journal of Plant Interactions 2014, 9, 702–711. [Google Scholar] [CrossRef]
- Amrao, L.; Akhter, S.; Tahir, M.N.; Amin, I.; Briddon, R.W.; Mansoor, S. Cotton leaf curl disease in Sindh province of Pakistan is associated with recombinant begomovirus components. Virus research 2010, 153, 161–165. [Google Scholar] [CrossRef]
- Ashraf, M.A.; Tariq, H.K.; Hu, X.-W.; Khan, J.; Zou, Z. Computational Biology and Machine Learning Approaches Identify Rubber Tree (Hevea brasiliensis Muell. Arg.) Genome Encoded MicroRNAs Targeting Rubber Tree Virus 1. Applied Sciences 2022, 12, 12908. [Google Scholar] [CrossRef]
- Ashraf, M.A.; Feng, X.; Hu, X.; Ashraf, F.; Shen, L.; Iqbal, M.S.; Zhang, S. In silico identification of sugarcane (Saccharum officinarum L.) genome encoded microRNAs targeting sugarcane bacilliform virus. PloS one 2022, 17, e0261807. [Google Scholar] [CrossRef]
- Ashraf, M.A.; Ashraf, F.; Feng, X.; Hu, X.; Shen, L.; Khan, J.; Zhang, S. Potential targets for evaluation of sugarcane yellow leaf virus resistance in sugarcane cultivars: in silico sugarcane miRNA and target network prediction. Biotechnology & Biotechnological Equipment 2021, 35, 1980–1991. [Google Scholar]
- Shahid, M.N.; Rasheed, S.; Iqbal, M.S.; Jamal, A.; Khalid, S.; Shamim, Z. IN SILICO PREDICTION OF POTENTIAL miRNAs TO TARGET ZYMV IN CUCUMIS MELO. Pak. J. Bot 2022, 54, 1319–1325. [Google Scholar]
- Ashraf, F.; Ashraf, M.A.; Hu, X.; Zhang, S. A novel computational approach to the silencing of Sugarcane Bacilliform Guadeloupe A Virus determines potential host-derived MicroRNAs in sugarcane (Saccharum officinarum L.). PeerJ 2020, 8, e8359. [Google Scholar] [CrossRef]
- Gaafar, Y.Z.A.; Ziebell, H. Novel targets for engineering Physostegia chlorotic mottle and tomato brown rugose fruit virus-resistant tomatoes: in silico prediction of tomato microRNA targets. PeerJ 2020, 8, e10096. [Google Scholar] [CrossRef]
- Jabbar, B.; Iqbal, M.S.; Batcho, A.A.; Nasir, I.A.; Rashid, B.; Husnain, T.; Henry, R.J. Target prediction of candidate miRNAs from Oryza sativa for silencing the RYMV genome. Computational biology and chemistry 2019, 83, 107127. [Google Scholar] [CrossRef]
- Iqbal, M.S.; Jabbar, B.; Sharif, M.N.; Ali, Q.; Husnain, T.; Nasir, I.A. In silico MCMV silencing concludes potential host-derived miRNAs in maize. Frontiers in plant science 2017, 8, 372. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, M.A.; Ali, B.; Brown, J.K.; Shahid, I.; Yu, N. In Silico Identification of Cassava Genome-Encoded MicroRNAs with Predicted Potential for Targeting the ICMV-Kerala Begomoviral Pathogen of Cassava. Viruses 2023, 15, 486. [Google Scholar] [CrossRef] [PubMed]
- Quillet, A.; Anouar, Y.; Lecroq, T.; Dubessy, C. Prediction methods for microRNA targets in bilaterian animals: Toward a better understanding by biologists. Computational and Structural Biotechnology Journal 2021, 19, 5811–5825. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, A.C.; Bovolenta, L.A.; Nachtigall, P.G.; Herkenhoff, M.E.; Lemke, N.; Pinhal, D. Combining results from distinct microRNA target prediction tools enhances the performance of analyses. Frontiers in genetics 2017, 8, 59. [Google Scholar] [CrossRef]
- Min, H.; Yoon, S. Got target?: computational methods for microRNA target prediction and their extension. Experimental & molecular medicine 2010, 42, 233–244. [Google Scholar]
- Riffo-Campos, Á.L.; Riquelme, I.; Brebi-Mieville, P. Tools for sequence-based miRNA target prediction: what to choose? International journal of molecular sciences 2016, 17, 1987. [Google Scholar] [CrossRef]
- Petchthai, U.; Yee, C.S.L.; Wong, S.-M. Resistance to CymMV and ORSV in artificial microRNA transgenic Nicotiana benthamiana plants. Scientific reports 2018, 8, 1–8. [Google Scholar] [CrossRef]
- Akhtar, S.; Tahir, M.N.; Amin, I.; Mansoor, S. Amplicon-based RNAi construct targeting beta-C1 gene gives enhanced resistance against cotton leaf curl disease. 3 Biotech 2021, 11, 1–10. [Google Scholar] [CrossRef]
- Ahmad, A.; Zia-Ur-Rehman, M.; Hameed, U.; Qayyum Rao, A.; Ahad, A.; Yasmeen, A.; Akram, F.; Bajwa, K.S.; Scheffler, J.; Nasir, I.A. Engineered disease resistance in cotton using RNA-interference to knock down Cotton leaf curl Kokhran virus-Burewala and Cotton leaf curl Multan betasatellite expression. Viruses 2017, 9, 257. [Google Scholar] [CrossRef]
- Yasmeen, A.; Kiani, S.; Butt, A.; Rao, A.Q.; Akram, F.; Ahmad, A.; Nasir, I.A.; Husnain, T.; Mansoor, S.; Amin, I. Amplicon-based RNA interference targeting V2 gene of cotton leaf curl Kokhran Virus-Burewala strain can provide resistance in transgenic cotton plants. Molecular biotechnology 2016, 58, 807–820. [Google Scholar] [CrossRef]
- Ali, I.; Amin, I.; Briddon, R.W.; Mansoor, S. Artificial microRNA-mediated resistance against the monopartite begomovirus Cotton leaf curl Burewala virus. Virology journal 2013, 10, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Yin, Z.; Li, Y.; Han, X.; Shen, F. Genome-wide profiling of miRNAs and other small non-coding RNAs in the Verticillium dahliae–inoculated cotton roots. PloS one 2012, 7, e35765. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, W.; Chen, J.; Liu, J.; Xia, M.; Shen, F. Identification of miRNAs and their targets in cotton inoculated with Verticillium dahliae by high-throughput sequencing and degradome analysis. International Journal of Molecular Sciences 2015, 16, 14749–14768. [Google Scholar] [CrossRef] [PubMed]
- Miao, Q.; Deng, P.; Saha, S.; Jenkins, J.N.; Hsu, C.-Y.; Abdurakhmonov, I.Y.; Buriev, Z.T.; Pepper, A.; Ma, D.-P. Genome-wide identification and characterization of microRNAs differentially expressed in fibers in a cotton phytochrome A1 RNAi line. PLoS One 2017, 12, e0179381. [Google Scholar] [CrossRef]
- Pang, M.; Woodward, A.W.; Agarwal, V.; Guan, X.; Ha, M.; Ramachandran, V.; Chen, X.; Triplett, B.A.; Stelly, D.M.; Chen, Z.J. Genome-wide analysis reveals rapid and dynamic changes in miRNA and siRNA sequence and expression during ovule and fiber development in allotetraploid cotton (Gossypium hirsutum L.). Genome biology 2009, 10, 1–21. [Google Scholar] [CrossRef]
- Hedden, P.; Kamiya, Y. Gibberellin biosynthesis: enzymes, genes and their regulation. Annual review of plant biology 1997, 48, 431–460. [Google Scholar] [CrossRef]
- Cai, C.; Li, C.; Sun, R.; Zhang, B.; Nichols, R.L.; Hake, K.D.; Pan, X. Small RNA and degradome deep sequencing reveals important roles of microRNAs in cotton (Gossypium hirsutum L.) response to root-knot nematode Meloidogyne incognita infection. Genomics 2021, 113, 1146–1156. [Google Scholar] [CrossRef]
- Pan, X.; Nichols, R.L.; Li, C.; Zhang, B. MicroRNA-target gene responses to root knot nematode (Meloidogyne incognita) infection in cotton (Gossypium hirsutum L.). Genomics 2019, 111, 383–390. [Google Scholar] [CrossRef]
- Hirsch, A.J. The use of RNAi-based screens to identify host proteins involved in viral replication. Future microbiology 2010, 5, 303–311. [Google Scholar] [CrossRef]
- Kampmann, M.; Horlbeck, M.A.; Chen, Y.; Tsai, J.C.; Bassik, M.C.; Gilbert, L.A.; Villalta, J.E.; Kwon, S.C.; Chang, H.; Kim, V.N. Next-generation libraries for robust RNA interference-based genome-wide screens. Proceedings of the National Academy of Sciences 2015, 112, E3384–E3391. [Google Scholar] [CrossRef] [PubMed]
- Martin, S.; Chiramel, A.I.; Schmidt, M.L.; Chen, Y.-C.; Whitt, N.; Watt, A.; Dunham, E.C.; Shifflett, K.; Traeger, S.; Leske, A. A genome-wide siRNA screen identifies a druggable host pathway essential for the Ebola virus life cycle. Genome medicine 2018, 10, 1–14. [Google Scholar] [CrossRef] [PubMed]
Figure 3.
Predicted target sites of cotton ghr-miRNAs. Four widely used in silico miRNA-mRNA target prediction algorithms were used. A. Prediction of miRNA binding sites by miRanda. B miRNA target sites were predicted by RNA22. C psRNATarget predicted target sites. D. Prediction of miRNA binding sites by RNAhybrid. miRNA target sites are represented by colored dots.
Figure 3.
Predicted target sites of cotton ghr-miRNAs. Four widely used in silico miRNA-mRNA target prediction algorithms were used. A. Prediction of miRNA binding sites by miRanda. B miRNA target sites were predicted by RNA22. C psRNATarget predicted target sites. D. Prediction of miRNA binding sites by RNAhybrid. miRNA target sites are represented by colored dots.
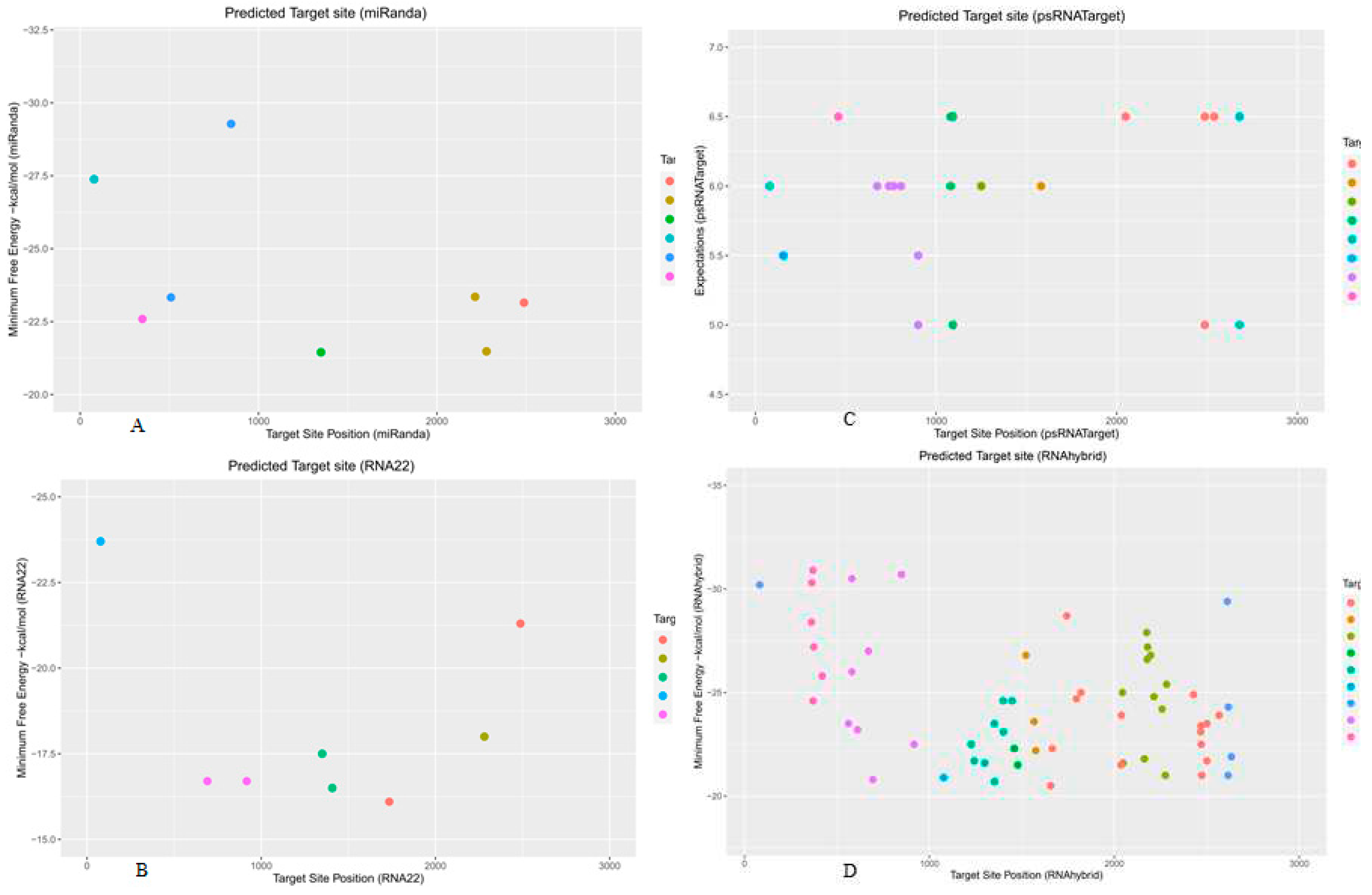
Figure 4.
Union plot of prediction shows predicted miRNAs results by all algorithms. miRNA target sites are represented by colored dots.
Figure 4.
Union plot of prediction shows predicted miRNAs results by all algorithms. miRNA target sites are represented by colored dots.
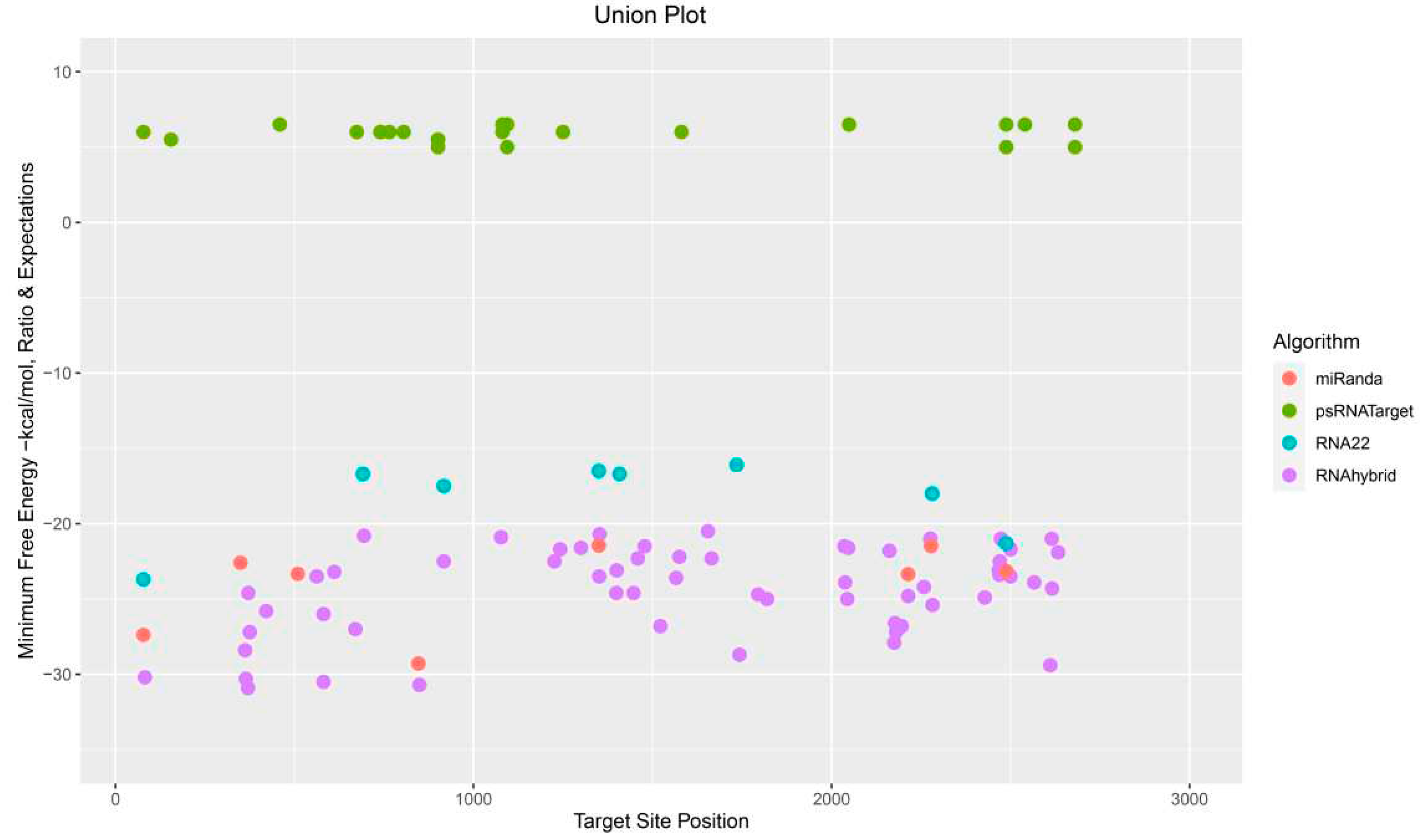
Figure 7.
Respective genomic region of CLCuKoV strains was targeted by ghr-miR2950. Multiple sequence alignment of CLCuKoV genomes showing the conservation of target site.
Figure 7.
Respective genomic region of CLCuKoV strains was targeted by ghr-miR2950. Multiple sequence alignment of CLCuKoV genomes showing the conservation of target site.

Figure 8.
Mechanism of miRNA-mRNA gene silencing pathway for the generation of CLCuKoV-resistant cotton. A. representing the candidate consensus cotton precursor miRNAs (ghr-MIR2950). B. describing miRNA/miRNA duplex replacement. C. representing showing miRNA expression construct harboring precursor sequenced driven by a promoter at 5’ end. The construct was terminated with a NOS gene. D. Indicating generation of mature amiRNA/amiRNA*duplex. E. representing RISC for processing amiRNA. F. indicating amiRNA degrades mRNA. .
Figure 8.
Mechanism of miRNA-mRNA gene silencing pathway for the generation of CLCuKoV-resistant cotton. A. representing the candidate consensus cotton precursor miRNAs (ghr-MIR2950). B. describing miRNA/miRNA duplex replacement. C. representing showing miRNA expression construct harboring precursor sequenced driven by a promoter at 5’ end. The construct was terminated with a NOS gene. D. Indicating generation of mature amiRNA/amiRNA*duplex. E. representing RISC for processing amiRNA. F. indicating amiRNA degrades mRNA. .
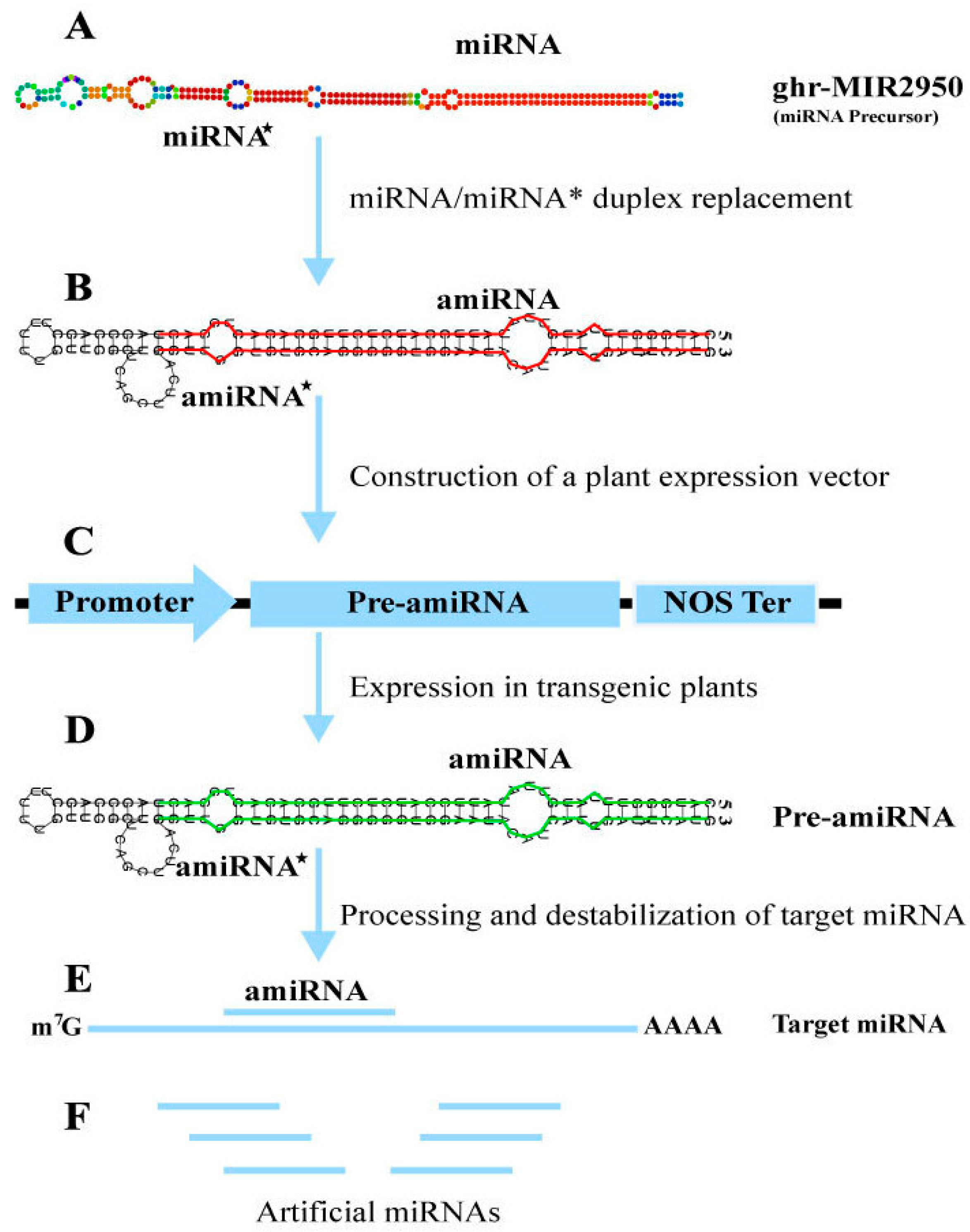
Table 1.
Comparison and overview of distinctive parameters of popular miRNA- mRNA target prediction algorithms were considered in this study.
Table 1.
Comparison and overview of distinctive parameters of popular miRNA- mRNA target prediction algorithms were considered in this study.
| Tools | Algorithms | Seed Pairing | Multiple Target Sites | Translation inhibition | Free Energy | Availability (web /code) |
|---|---|---|---|---|---|---|
| miRanda | Local alignment | + | + | + | + | +/+ |
| RNA22 | FASTA | − | + | − | + | +/− |
| psRNATarget | Smith-Waterman | − | + | + | − | +/− |
| RNAhybrid | Intermolecular hybridization | + | + | + | + | +/− |
| Tapirhybrid | FASTA | + | + | − | + | +/+ |
| TargetSpy | FASTA | − | + | − | + | −/+ |
| Targetfinder | FASTA | + | − | − | + | −/+ |
Table 2.
The cotton ghr-miRNAs were predicted to target each ORF/gene.
| CLCuKoV-Lu gene |
miRanda | RNA22 | psRNATarget | RNAhybrid | |
| V1 | ghr-miR7486 (a, b), ghr-miR7506 | ghr-miR169a, ghr-miR7512 | ghr-miR827 (a, b, c), ghr-miR3476-5p | ghr-miR393, ghr-miR482 (a, b), | |
| ghr-miR7492 (a, b, c), ghr-miR7500 ghr-miR7510a |
ghr-miR7486 (a, b), ghr-miR7490, ghr-miR7504a, ghr-miR7510a, ghr-miR7512 |
||||
| V1/V2 | ghr-miR7497 | ghr-miR7497 | ghr-miR164, ghr-miR479, ghr-miR3476-5p, ghr-miR7497, ghr-miR7498, ghr-miR7507 | ||
| C1 | ghr-miR7486 (a, b) | ghr-miR398, ghr-miR7486 (a, b) | ghr-miR7486 (a, b) | ghr-miR156 (a, b, c, d), ghr-miR162a, ghr-miR166b, ghr-miR169a, ghr-miR398, ghr-miR827 (a, b, c), ghr-miR2949-3p, ghr-miR3476-3p, ghr-miR7491, ghr-miR7492 (a, b, c), ghr-miR7500, ghr-miR7501, ghr-miR7505, ghr-miR7506 |
|
| C2 | ghr-miR394 (a, b), ghr-miR7504b | ||||
| C1/C2 | ghr-miR7510b | ghr-miR7485, ghr-miR7487, ghr-miR7514 | |||
| C3 | ghr-miR7484 (a, b), ghr-miR7492 (a, b, c) | ghr-miR7484 (a, b) | |||
| C2/C3 | ghr-miR7513 | ghr-miR7489, ghr-miR7513 | ghr-miR396 (a, b) | ghr-miR167 (a, b), ghr-miR396 (a, b), ghr-miR2949(a-5p, b, c),ghr-miR7489, ghr-miR7493, ghr-miR7494, ghr-miR7511, ghr-miR7513 | |
| C4/C1 | ghr-miR390 (a, b, c), ghr-miR7503 | ghr-miR390 (a, b, c) | ghr-miR160, ghr-miR172, ghr-miR390 (a, b, c), ghr-miR399d ghr-miR7488, ghr-miR7495 (a, b), ghr-miR7503, ghr-miR7508, ghr-miR7509, ghr-miR7510b |
||
| LIR | ghr-miR2950 | ghr-miR2950 | ghr-miR2950 | ghr-miR399 (a, b, c, e), ghr-miR2948-5p, ghr-miR2950, | |
Table 7.
The free energy (ΔG) of the consensus cotton ghr-miRNA-mRNA were estimated.
| miRNA ID | miRNA-mRNA Sequence (5′–3′) |
ΔG Duplex (Kcal/mol) |
ΔG Binding(Kcal/mol) | |
| ghr-miR2950 | 5′ UGGUGUGCAGGGGGUGGAAUA 3′ 5′ AATAACGCTCCCGCACACTA 3′ |
−24.80 | −24.37 | |
| ghr-miR7486 (a, b) | 5′ AAGGAAGCGCUUUGUCCACGUGGA 3′ 5′ TGAATTTGGGAAAGTGCTTCCTC3′ |
−22.70 | −17.41 | |
| ghr-miR7513 | 5′AAUCAGCCAGGAAUCGUUUGA 3’ 5′ ATGGACGGTTGACGTGGCTGATG 3’ |
−20.90 | −17.74 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



