
Submitted:
28 September 2023
Posted:
29 September 2023
You are already at the latest version
Abstract
The goal of this article was to evaluate the antioxidant and anti-inflammatory like activities of three species from genus Primula L. growing in Georgia: Primula macrocalyx, Primula woronowii and Primula saguramica . First, fractions containing shoot and root parts were air-dried, grinded and extracted by 80% ethanol followed by condensation and freeze-drying. Second, further chromatographic separations were done on Diaion HP-20 with the following solvents: H2O, methanol (50% and 100 %) and ethylacetate 100% to obtain the expected fractions.
Keywords:
Primula macrocalyx
; Primula woronowii
; Primula saguramica
; Extraction
; Stoichiometric Anticatalytic
; Antioxidant
; ABTS
; DPPH
; Neutrophils
; Myeloperoxidase
; Reactive oxygen species
1. Introduction
The Primulaceae family includes 59 genus of plants and approximately 697 species [1,2]. In Georgia, this family is represented by 7 genera and 46 species. Twenty-two species of the genus Primula L. can be found in Georgia, among them 10 are endemics of the Caucasus, and two - endemics of Georgia: Primula abchasica and Primula saguramica [2]. Phytochemical investigation on various species of genus Primula L. revealed that flavonoids are widely distributed in this genus [3,4,5]. Numerous studies have suggested that flavonoids exhibit biological activities, including antiallergenic, antiviral, anti-inflammatory and vasodilating actions. However, most interest has been devoted to the antioxidant activity of flavonoids, which is due to their ability to reduce free radical formation and to scavenge free radicals [6]. It is recognized that reactive oxygen species (ROS) play different roles in vivo. Some of these effects are beneficial and connected in cell growth as well as to intercellular signaling. However, high ROS production may be very harmful, since they can oxidize lipids, cell membranes, proteins in tissues, enzymes, carbohydrates and DNA [6]. The antioxidant abilities of flavonoids can help to decrease these harmful effects. The hydroxyl groups of aromatic rings in flavonoids donate H atom to various radicals, such as peroxyl, hydroxyl and other radicals, which lose reactivity due to their stabilisation and on the other hand forming a relatively stable flavonoid radical [7]. In addition, the C-2 and C-3 double bonds and OH group at the C-3 position of flavonoids can impove the resonance stability for electron delocalization across the molecule, which determines better antioxidant properties [8]. Several enzymes involved in inflammation, including xanthine oxidase (XO), cyclo-oxygenase (COX), lipoxygenase phosphoinositide 3-kinase and myeloperoxidase (MPO) are also known to be potently inhibited by flavonoids [9,10].
In traditional medicine, different plant species of genus Primula have been used from immemorial times by the inhabitants of Greece as antidote to snake poison and their juice was applied to relieve toothache [11]. Some species of the genus are used traditionally to treat epilepsy and convulsions [12]. Different species of the genus promote functioning of liver and spleen and remove obstructions of these organs, their constituents have been prescribed to relieve pain of kidneys, used to cure boils, scorpion strings and are also used as sedative [13].
In Georgian folk medicine, Primula species were used for cough, especially in children’s practice with boiled grape juice [14]. Zaza Panaskerteli’s book, Karabadine’’ mentions Primula species flower with mixture of honey and vinegar as a wound healer [15].
We are particularly interested by Primula macroxalyx, Primula woronowii being endemic of the Caucasus and Primula saguramica being endemic of Georgia. Although the population of Georgia actively uses these plants as food and for the treatment of various diseases, such as avitaminosis, cough, etc., it is important to understand their biological action mode in relationship with their content of secondary metabolites [16].
Different fractions of these plants were obtained from the crudes and using increasing gradient (0-100%) of MeOH in water or ethylacetate (EtOAc) to separate compounds having different polarities. In the litterature, few data concerns the anti-inflammatory potential of these species. Polymorphonuclear neutrophils (PMNs) play a key role in inflammatory response. PMNs are the major component of phagocytic cells in the human bloodstream. They were stimulated and undergo a respiratory burst when they phacocyte foreign agents as bacteria and damage cells [17]. The engulfed particles are exposed to a high flux of superoxide anion molecules produced via the activity of NADPH-oxidase complex. Once the phagocytic vacuole is formed, fusion with other granules in the neutrophil cytoplasm releases myeloperoxidase (MPO) which uses hydrogen peroxide as a substrate and oxidizes chloride to hypochlorous acid, a strong oxydant molecule that can oxidize in turn many biological molecules, and reduce thiol groups. This leads to serial events resulting in bacterial killing [17].
During excessive inflammation, an important and uncontrolled activation of PMNs occurs leading to an important ROS production and a release of high amount of MPO considered in this context as a pro-oxidant biomarker, triggering and amplifying oxidation, nitration and chlorination of some molecules in the extracellular medium (Davies 2011) . These harmful side effects can also affect molecules of interest such DNA, neurotransmitters, proteins and cause serious diseases. One way to limit the deleterious effect is lowering the ROS overproduction and decreasing MPO activity [17].
In the present work, we evaluated the antioxidant activities of fractions by using four different techniques, (i) cell free system using 2,2-azino-bis (3-ethylbenzothiazoline-6-sulphonic acid (ABTS) and 2,2-diphenyl-1-picrylhydrazyl (DPPH) assays to determine the stoichiometric potential of the extract molecules against free radical production, (ii) a chemiluminescent technique with a cellular model to assess the effect of extracts on the modulation of ROS production by stimulated neutrophils, (iii) an original pharmacological technique to study the anticatalytic activity of the extracts against MPO by targeting its active site (SIEFED), (iv) a pharmacological technique for determination of the electron donor potential of the extracts on the peroxidase cycle of MPO .
2. Results
2.1. Screening of secondary metabolites
In the first step, for screening of some secondary metabolites in Primula species, it was important to perform TLC and preliminary phytochemical tests [18,19,20]. According to the results, the content of flavonoids and triterpene glycosides were determined (see Figure 1A–C). As a reference standard rutin was used at starting concentration of 1 mg/ml and samples with initial concentrations of 20mg/ml. In advance, for TLC, there was prepared system - CHCl3: MeOH: H2O ratio-26:14:3 and silica gel plate was placed in the chromatography tank after about 2 hours. For revelation secondary metabolites (flavonoids and triterpene glycosides) 1% 2-aminoethyl diphenylborinate and 1% vanillin solution was used respectively. According to the results, flavonoids was characterized for 1, 3 and 4 fractions for all species, more intense color seems detected for P.m3, P.w3 and P.s3. On the other hand, triterpenes mostly appear in 1 and 4 fractions.
2.2. Effect of the fractions on the scavenging activity towards radical cation ABTS*+ and DPPH* formation
ABTS and DPPH anti-radical assays were used for the stoichiometric activity assessment of all the fractions. Gallic acid and quercetin used as control standards were more efficient than fractions. Most of the tested fractions exhibited a dose-dependent radical scavenging activity except fractions obtained in H2O and EtOAc. From these results, a calculation of IC50 was performed (see Table 1). The best scavenging activity was found for the standards gallic acid and querectin followed by the plant fractions Pm, Pw, and P.s prepared in MeOH (50%). The fraction from Pw showed the best results (P.w 3 ABTS - IC50=8.51 ± 0.18; DPPH - IC50=34.57 ± 0.47). Results from ABTS indicate that the reaction between studied fraction and ABTS radical species is faster and better than for DPPH assay (see Figure 2A–C)). For some fractions (P.m1,P.m2,P.m4, P.m5; P.w 1DPPH, P.w2, P.w4 DPPH, P.w5; P.s1, P.s2, P.s4 P.s5), the IC50 could not be determined, because at these concentrations, the inhibition of free radicals (ABTS, DPPH) in percentage, did not exceed 50%.
2.3. Effect of the fractions on ROS production by stimulated neutrophils.
One of the most important mechanisms for the determination of anti-inflammatory activity is to study the inhibition of ROS produced by stimulated neutrophils. Gallic acid used as control standard was more efficient than fractions. Considering the 3 species, the fraction 3 and 4 prepared in MeOH (50%) and MeOH (100%) respectively showed the best results. The fractions obtained with ethanol extraction and freeze drying (fraction 1) also showed good results but less effective than for 3 and 4 fractions (Figure 3–C). For plant fractions 3 and 4, additional dilutions were prepared for the detemination of IC50 (see supplementary material). Based on these experiments IC50 were calculated and the best one was determined for: P.w 3 PMN - IC50 0.68 ± 0.04 (see Table 1). For some fractions, the IC50 could not be determined, because at these concentrations, the inhibition of ROS in percentage, did not exceed 50%.
Figure 3 (supplementary material) Relative inhibition of neutrophils ROS production induced by Primula macroxalyx (P.m.) (A), Primula woronowii (P.w) (B) nd Primula saguramica (C) fractions obtained after chromatographic separation with 50 % MeOH and 100 % MeOH (fractions 3 and 4). Gallic acid (GA) and quercetin (Qu) were used as reference compounds. All the powdered extracts were solubilized in DMSO and their ROS inhibition capacity were compared to DMSO control (Ctrl DMSO) and stimulated neutrophils without sample (ST PMN). Results were obtained after 30 min incubation time and expressed in relative percentages vs negative Ctrl set as 100 % (Mean +/- SD, n=3 independent experiments with 3 technical replicates for each experiment).
2.4. Effect of the fractions on MPO activity
Flavonoids are the compounds with different number of hydroxyl groups. Besides their antioxidant activity, another important pharmacological activity in relationship with inflammation is their ability to interfere with the activity of MPO either by penetrating and blocking the active site of the enzyme, or by acting as electron donor molecules for the peroxidase cycle of the enzyme. These abilities were determined by the SIEFED and the classical assay of the enzyme, respectively. First reference molecules, gallic acid and quercetin solubilized in DMSO, showed a better efficiency in the classical assay with similar IC50 in compared to the SIEFED one (Figure 4), suggesting that they act mainly as electron donors for the peroxidase cycle of the enzyme. Nevertheless, SIEFED assay showed that both molecules have the ability to also block the active site of the enzyme but the IC50 for gallic acid (0.53 ± 0.04) was lower than quercetin one (1.37 ± 0.02). For all the plant species, most of fractions showed a dose dependent inhibitory effect but the chromatographic fractions obtained with EtOAc were the less efficient (Figure 4A–C). Whatever the species, it was the chromatographic fraction obtained with 50 % MeOH (3) that gave the best results followed by the crude fraction got with ethanol extraction and freeze drying (1). For these two fractions results obtained by SIEFED and classical assays were quite similar suggesting that compounds from these fractions act mainly by blocking the active site of the enzyme. According to the IC50 from SIEFED and classical enzymatic assays P.w 3 seems to have the strongest anti-MPO activity (Table 1). For some fractions, the IC50 could not be determined, because at these concentrations, the inhibition of MPO and electron donating ability in percentage, did not exceed 50%.
3. Discussion
In this article, we focused on species growing in Georgia: P.m, P.w and P.s. (Primula macrocalyx, Primula woronowii and Primula saguramica). The first step was the development of an optimal extraction condition to obtain secondary metabolites. In the literature, phenols, triterpene saponins, and flavonoids are the most common classes of compounds [21]. Based on these data, EtOH 80% was selected as an extraction solvent. Three times extraction was performed, and as a result, crude extracts were obtained (P.m1, P.w1, and P.s1). For fractionation purposes, column chromatography was carried out with Diaion HP-20 as the stationary phase and solvents with different polarities (water, MeOH 50%, MeOH 100%, and EtOAc 100%) as the mobile phase. Some compounds strengthen each other and, as a result, have synergism; however, in some cases, they interfere with other effects, so the maximum therapeutic effect is not revealed. Based on our results, extraction with highly porous solvents resulted in a high extract yield, but with a low pharmacological effect as compared to non-polar ones. This indicates that fractionation has an important influence on results. In addition, the use of a combination of polar and nonpolar solvents increases the extraction efficiency of phytochemicals with good antioxidant quality from these species. After using column chromatography, the following fractions were obtained: H2O – P.m 2, P.w 2, P.s 2, MeOH 50% - P.m 3, P.w 3, P.s 3, MeOH 100% - P.m 4, P.w 4, P.s 4, EtOAc 100% - P.m 5, P.w 5, P.s 5. For the preliminary screening of phytochemicals, several tests and TLC were performed, which confirmed the content of flavonoids and triterpene saponins. According to TLC, flavonoids mostly are in 3 fractions (P.m 3, P.w 3, P.s 3), while triterpene saponins in 1 and 4. For flavonoids 1% 2-aminoethyl diphenylborinate and for triterpene saponins, 1% vanillin solution in methanol were used, yellow-orange and violet spots were produced respectively [20].
According to the properties of secondary metabolites, several methods have been used to evaluate the antioxidant properties of the fractions in vitro based on the chemical evaluation of the stoichiometric activity of the molecules by measuring their ability to scavenge radical species produced by cell free system (DPPH, ABTS) [22]. ABTS*+ and DPPH* assays are good and complementary techniques for evaluation of the general stoichiometric activities of the molecules, because ABTS*+ includes hydrogen atom transfer assay (HAT), which means donating of hydrogen atom from antioxidants and DPPH – include HAT and single electron transfer mechanism (SET). The latter reaction based on single-electron transfer from antioxidant to free radicals [23]. From ABTS and DPPH results, the 3 species showed stoichiometric antioxidant properties when the plant fraction is prepared in 50% MEOH. According to the results P.w showed the best antiradical activity but with a weaker response for DPPH than for ABTS . According to the TLC results, P.w3 fractions characterized with more type of flavonoids, this is maybe a reason for better antioxidant activity than others. Strong stoichiometric antioxidant properties were already demonstrated in different Primula species (Primula veris, Primula heterochroma Primula vulga ris) indicating that these plants are important source of antioxidants [24,25].
However, ABTS and DPPH methods do not take into consideration the potential effects on the cells. Antioxidant molecules also act by inhibiting the enzymes and pathways responsible for the ROS production (anticatalytic activity). It is evident that the anticatalytic activity is more powerful than the stoichiometric one, because low concentrations of anticatalytic antioxidant molecules are sufficient to block or reduce the source of ROS production [26].
Nowadays, more works are devoted to the antioxidant effects of molecules on the activity of enzymes involved in oxidation process and radical species formation which requires to use enzymatic and extra and intracellular cellular systems [27,28,29,30]. The effect of the crude extract and fractions was investigated on neutrophils and MPO playing key roles in inflammation. We used a luminescent probe, L012, generally used to measure the ROS production by stimulated neutrophils and it is known that the primary ROS is superoxide anion (O2.-) produced by the activity of NADPH oxidase [31]. (see Figure 3–C). Our results showed the strong potential of the 3 Primula species prepared in 50% MeOH to inhibit the ROS production by neutrophils (Fig 3 A,B,C). and confimed our previous results on the antioxidant stoichiometric activities. Theses 3 species are well used in Georgia for their anti-inflammatory properties especially against cough, catarrhs of the respiratory tract and chronic bronchitis. Neutrophils are known to be involved in common respiratory diseases. Stimulation of neutrophils and ROS production also determine the NET formation consisting of decondensed chromatin fibers coated with antimicrobial proteins, such as histones, neutrophil elastase (NE), α-defensins and MPO [32]. Generally excessive NET production increases mucus viscosity, causing the lungs to fill with mucus and affecting lung function. It seems that secondary metabolites containing in primula species can inhibit NET formation [33].
In our cellular model, mostly superoxide anion is measured, however we cannot exclude interaction with cells and therefore modification of cellular metabolism or enzyme activities. Indeed, flavonoids can exert their anti-inflammatory effects at multiple cellular levels, including signalling pathways, cytokines and chemokines production and modulation of the enzymes involved in reactive oxygen and nitrogen species [34,35,36]. Among the enzymes, myeloperoxidase, released by stimulated PMNs, play significant role in inflammation. After its oxidant activity, involving both peroxidase and chlorination cycles, MPO is able to produce strong oxidant species such as HOCl [37]. It is evidenced that oxidant compounds derived from the activity of MPO are necessary employed for the killer function of neutrophils towards bacterial and other pathogen agents, but also cause tissue damage, initiates and propagates acute and chronic inflammatory diseases [38]. The SIEFED (specific immunological extraction followed by enzymatic detection) a licensed method developed by Franck et al. [39,40], which gives us opportunity to study the direct interaction between MPO and molecules from the extracts. The SIEFED consists of the extraction of MPO by immobilized anti-MPO antibodies followed by washing. The washing step allows to eliminate proteins, interfering molecules or testing samples or drugs. After washing, the MPO activity was measured using a detection system containing a fluorogenic substrate, hydrogen peroxide, and sodium nitrite as reaction enhancer (Franck et al. 2005). Any inhibition of the enzyme activity, persisting after the elimination of the molecule evidence that the molecule has interacted with the enzyme, modified its structure, inhibit its active site and thus limited or blocked the access of substrates to this site [39]. On the other hand, with classical enzymatic assay which does not use an immunological capture, we can evaluate electron donor potential of tested molecules in MPO peroxidase cycle [41]. With the classical enzymatic assay there is a risk of reaction between the substrate or the product derived from the enzyme activity and the inhibitor. However it is good technique to determine the electron-donating potential of the molecule for the peroxidase cycle of the enzyme or competitive inhibition toward substrate. For both techniques, the 3 plant species showed good potential to inhibit MPO activity by either interacting with its active site or by substrate competition especially the crude extract and the fraction prepared with 50% MEOH. Therefore, Primula species contained molecules having the potential to modulate the activity of MPO and thus preventing the harmful effect of the oxidant molecules derived from its activity. Flavonoids compounds are known to inhibit the activity of MPO [42]. Further studies are needed to determine which molecules from the extract are responsible for this inhibition.
4. Materials and Methods
4.1. Chemicals and reagents
2,2-Azinobis-(3-ethylbenzohiazoline-6-sulphonic acid (ABTS) was from Fluka (Bornem, Belgium). Sodium, and potassium chloride, ethanol, hydrogen peroxide (H2O2), dimethyl sulfoxide (DMSO), Tween 20 were analytical grade products from Merck (VWRI, Leuven, Belgium). L-012 (8-amino-5-chloro-7-phenyl-pyrido[3,4-d] pyridazine-1,4(2H,3H)dione) was obtained from FujiFilm Wako Chemical Europe (Neuss, Germany) were Amplex Red (10-acetyl-3,7- dihydroxyphenoxazine) was purchased from Invitrogen Phorbol 12-myristate 13-acetate (PMA), gallic acid (2,4,6-trihydroxybenzoic acid) and sodium persulfate (Na2S2O8) DPPH (2,2-diphenyl-1-picrylhydrazyl) was purchased from Aldrich (Steinheim, Germany). Quercetin (3,3,4,5,7-pentahydroxy-2-phenylchromen-4-one) was from ChromaDex (LGC Standard, France). De-oxygenated milliQ water or ultrapure water (EasyPure UV purification system) was used for the preparation of all solutions. Fluoroskan Ascent FL (Fisher Scientific, Tournai, Belgium) and Multiskan Ascent (Thermo Labsystem) were used for determination antioxidant activity.
4.2. Sample collection
The following plants are the research objects from genus Primula. L : Primula macrocalyx Bunge., - was collected in the floristic area of Kartli, in the meadows of the Saguramo-Zedazeni forest-park, in the forest edges E044.788210, at H-990m, in the active flowering phase in 2021, Primula woronowii Losinsk., - was collected in the floristic area of Kartli, in the meadows of the Saguramo-Zedazeni forest-park, E044.785340, at H-891m, in the active flowering phase in 2020 and Primula saguramica Gavr. - was collected in the floristic area of Kartli, on the eastern slope of the Tbilisi ridge, on the slopes of village Karsan in oak-shrub forests E044.712020, at H-798m, in active flowering phase in 2021 [2].
4.3. Extraction procedure and preliminary phytochemicals screening
The first step was the selection of extraction conditions: A shoot and root of all species were dried, grinded and three-time extraction with 80% EtOH was performed, after condensation with rotary evaporator at 70 0C, crude extracts were obtained with lyophilization (P.m 1, P.w 1, P.s 1). From the crude extracts, the following enriched fractions were obtained by column chromatography on Diaion HP-20, with an increasing gradient: H2O – P.m 2, P.w 2, P.s 2, MeOH 50% - P.m 3, P.w 3, P.s 3, MeOH 100% - P.m 4, P.w 4, P.s 4, EtOAc 100% - P.m 5, P.w 5, P.s 5. (see Figure 5)
The ethanol 80%, water, methanol 50%, methanol and ethyl acetate extracts (1 g) were dissolved in 100 mL of its own mother solvents for preparing the stock solution and the latter was used for phytochemical screening following the methodology of Harborne and Kokate [18,19] Thin layer chromatography (TLC) was performed for screening of secondary metabolites [20]. In both cases, flavonoid and triterpene glycosides were revealed. For TLC of flavonoids and triterpene glycosides, the system: CHCl3: MeOH: H2O - 26:14:3 was used [20].
4.4. ABTS Free Radical Scavenging Activity Assay
The first ABTS assay was described in 1993 by Miller [49]. the method was developed based on absorbance of the ABTS.+ radical cation for the evaluation of the total antioxidant capacity of body fluids and drug solutions. This method was first based on the production of ABTS radical cation by activation of metmyoglobin with hydrogen peroxide to generate ferrylmyoglobin radical. The latter reacted with ABTS to form the ABTS°+. ABTS radical cation can also be generated from the reaction between ABTS and sodium persulfate, with direct production of the blue/ green ABTS°+ chromophore [26] [43]. The ABTS free radical-scavenging activity of each extract was measured with a Multiskan Ascent plate reader at 740 nm. The starting concentrations of gallic acid and quercetin were 1 mg/ml and final ranging from 10 to 0.625 µg/ml. The starting concentrations of tested samples were 5mg/ml and final ranging from 50 to 3.125 µg/ml. The samples and standarts were dissolved in DMSO and distributed in 96-well UV-Transparent Microplates control consisted of 0.02 mL DMSO in 1.98 mL of ABTS solution. The measurements were perfromed three times. A control consisted of 0.02 mL DMSO in 1.98 mL of ABTS solution. Samples were prepared and measured in triplicates.
4.5. DPPH Free Radical Scavenging Activity Assay
The DPPH assay was first suggested in the 1950s to find electron donors in natural products [44]. Later it was used to determine the antioxidant activity of phenolic compounds and plant food [45]. The DPPH (2,2-diphenyl-1-picrylhydrazyl) is a radical of organic nitrogen, which has an unpaired valence electron at one atom of nitrogen bridge, and is a stable, soluble organic medium and is an insoluble aqueous medium, with a purple color. The DPPH free radical-scavenging activity of each extract was measured with a Multiskan Ascent plate reader at 510 nm [46]
The starting concentrations of gallic acid and quercetin were 1 mg/ml and final ranging from 10 to 0.625µg/ml. The starting concentrations of tested samples were 5 mg/ml and final ranging from 50 to 3.125 µg/ml. the samples and standarts were dissolved in DMSO and distributed in 96-well UV-Transparent Microplates. A control consisted of 0.02 mL DMSO in 1.98 mL of DPPH solution. The measurements were perfromed three times
4.6. The SIEFED (“Specific Immunological Extraction Followed by Enzymatic Detection”) and classical enzymatic methods for measurement of equine active myeloperoxidase in biological samples.
Myeloperoxidase (MPO) is an abundant heme-containing enzyme, which mostly present in neutrophils. In response to an invading agent in the body, MPO produces hypochlorous acid (HOCl), which damages various cellular structures. Therefore, it is important to determine anticatalytic activity of secondary metabolites.
For determination MPO inhibitory activity SIEFED (specific immunological extraction followed by enzymatic detection) method was performed. This method is specific because cobaye anti-MPO IgG antibodies (3µg/ml) against MPO were bound into the wells of a microplate to capture MPO. 150 microliters of MPO solution were placed in a 96-wells microtiter plate (Combiplate 8 EB), from Thermo Scientific (Breda, Netherlands), coated with cobaye anti-human MPO polyclonal antibodies (CORD, Belgium) following 3 microliters of samples and 147 microliters of PBS. All plates were incubated for 2 h at 370C in the darkness, before the wells were washed up. After washing, the peroxidase activity of MPO was measured with a fluorogenic substrate, hydrogen peroxide, and nitrite as reaction enhancer, [40]. For the classical enzymatic assay, this method reveals the electron donating potential of a molecule in MPO peroxidase cycle. Unlike the previous method, specific antibodies are not used here, washing and incubation steps are not performed. Fluorescence was measured with a Fluoroscan Ascent plate readerc at the excitation and emission wavelengths of 544 and 590 nm, respectively. The fluorescence value was directly proportional to the quantity of active MPO present in the sample [40]. A control assay set as 100% MPO activity was performed with purified MPO solution where an equivalent volume of DMSO solution was added instead of the samples.
The starting concentrations of gallic acid and quercetin were 1mg/ml and final ranging from 10 to 0.078µg/ml. The starting concentrations of tested samples were 5mg/ml and final ranging from 50 to 0.78 µg/ml. Samples were prepared and measured in triplicates.
4.7. Measurement of ROS production
The ROS production by activated neutrophils was measured by L-012-enhanced chemiluminescence (CL) according to the method adapted from Benbarek et al. [47] and Franck et al. [29]. Phorbol 12-myristate 13-acetate (PMA) was used for neutrophil stimulation. The latter one is comonly used to study the neutrophil, because PMA induces pathways to other stimuli, with the participation of PKC, MPO and neutrophil elastase (NE), also PMA induces neutrophil degranulation, metabolic changes, ROS and NET formation [50,51]. For the method, 594 µl neutrophil suspensions (6x104 neutrophils/well) were distributed in a 96-well white microtiter plate (White Combiplate 8, Thermo Labsystems, Finland) by following 6 µl of samples, 10 µl of L-012 and 10 µl of PMA. The plate was incubated for 10 min at 370C with fractions. The CL response of the neutrophils was monitored for 30 min at 37 0C with a Fluoroskan Ascent FL (Fisher Scientific, Tournai, Belgium) and expressed as the integral value of the total CL emission. A control assay set as 100% of CL response was performed with PMA-stimulated neutrophils where an equivalent volume of DMSO solution was added instead of the samples. Samples were prepared and measured in triplicates [48]. The starting concentrations of gallic acid and quercetin were 1mg/ml and final ranging from 10 to 0.019µg/ml. The starting concentrations of tested samples were 5mg/ml and final ranging from 50 to 0.78 µg/ml
4.8. Statistical analysis
For ABTS, DPPH, SIEFED and CL assays, within an experiment each assay was done at least three times. The statistical analysis was performed with GraphPad Instat 3.05 (GraphPad Software, San Diego CA, USA). All the results are expressed as mean ± standard deviation (SD) in percentage vs. controls, which were set as 100%.
5. Conclusion
All three species – P.m, P.w and P.s were extracted with 80% ethanol, followed by fractionation on column chromatography and preliminary phytochemical tests with TLC were performed for screening secondary metabolites. According to the results, triterpene saponins and flavonoids were revealed. Depending on the content of the latter, the antioxidant and anti-inflammatory activity of the fractions were evaluated using different techniques. The results from all methods demonstrate that P.w 3 fraction characterized by both good stoichiometric and anticatalytic activity, in particular, secondary metabolites in P.w 3 can reduce ABTS and DPPH radicals, results from SIEFED - it inhibits the active center of MPO, acts as an electron donor in MPO peroxidase cycle based on the results of the classical enzymatic assay and inhibits reactive oxygen species according to the results conducted on neutrophils.
Overall our results showed that the MeOH 50 % fraction of P.w 3 has both good stoichiometric and anticatalytic activity on neutrophil ROS production and MPO activity. Such results could be partly related with the well known antiinflammatory and medical properties of Primulas species.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
References
- WFO – 7000000497, Primulaceae Batsch ex Borkh. Bot. Wörterb. 2: 240. 1797.
- Microstructural Features of Primula worownoii Losinsk., Primula macrocalyx Bunge. and Primula saguramica Gavr. from Georgian Flora. Nino Sukhishvili1, Ketevan Mchedlidze2, Karen Mulkijanyan3 and Lasha Mskhiladze1. [CrossRef]
- Phytochemistry of European Primula species, Paola S. Colombo, Guido Flamini, Graziella Rodondi, Claudia Giuliani, Laura Santagostini, Gelsomina Fico. Phytochemistry 2017, 143, 132–144.
- Unusual Flavones from Primula macrocalyx as Inhibitors of OAT1 and OAT3 and as Antifungal Agents agains Candida rugosa, Xue Li, Xue Wang, Caiyu Li, Manana Khutsishvili, George Fayvush, Daniel Atha, Youcai Zhang & Robert P. Borris, Published: 25 June 2019.
- Budzianowski, J.; Wollenweber, E. Rare flavones from the glandular leaf exudate of the Oxlip, Primula elatior L. Natural Product Communications 2007, 2, 267–270. [Google Scholar] [CrossRef]
- Pietta, P.G. Flavonoids as antioxidants. J Nat Prod. 2000, 63, 1035–42. [Google Scholar] [CrossRef] [PubMed]
- Heim, K.E.; Tagliaferro, A.R.; Bobilya, D.J. Flavonoid antioxidants: Chemistry, metabolism and structure-activity relationships. J. Nutr. Biochem. 2002, 13, 572–584. [Google Scholar] [CrossRef] [PubMed]
- Nitration of Flavonoids and Tocopherols as Potential Modulators of Nitrosative Stress—A Study Based on Their Conformational Strutures and Energy Content, Jos Manuel Perez de la Lastra, Celia Andres Juan, Francisco J.Plou and Eduardo Perez-Lebena, Received: 9 April 2022 / Revised: 27 April 2022 / Accepted: 5 May 2022 / Published: 9 May 2022. [CrossRef]
- Walker, E.; Pacold, M.; Perisic, O.; et al. Structural determinations of phosphoinositide 3-kinase inhibition by wortmannin, LY294002, quercetin, myricetin, and staurosporine. Mol Cell 2000, 6, 909–919. [Google Scholar] [CrossRef]
- Shiba, Y.; Kinoshita, T.; Chuman, H.; Taketani, Y.; Takeda, E.; Kato, Y.; Naito, M.; Kawabata, K.; Ishisaka, A.; Terao, J.; Kawai, Y. Flavonoids as substrates and inhibitors of myeloperoxidase: molecular actions of aglycone and metabolites. Chem Res Toxicol. 2008, 21, 1600–9. [Google Scholar] [CrossRef]
- Dymock, W.; Warden, C.J.; Hand Hooper, D. Pharmacographical Indica., 451. Published by Thacker Spink and company, Calcutta. Reprinted by Hamdard Institute of Health and Tibbi (Medical) Research Karachi, 1972. 1890. [Google Scholar]
- Jager, A.K.; Gauguyn, B.; Adsersen, A.; Gudyksen, L. Screening of plants used in Danish folk medicine to treat epilepsy and convulsions. Journal of Etnopharmacology 2006, 105, 294–300. [Google Scholar] [CrossRef]
- Saqib, N. Phytochemical studies of some Primulaceous leguminous plants. Ph.D. Thesis, university of Karachi, 1980; pp. 4–6. [Google Scholar]
- Pharmacology in ancient Georgia and the ways of its further development from ancient times to XX century, publishing house, “Art’’ Tbilisi - 1987, Sokrat Salukvadze p.188.
- Zaza Panaskerteli-Tsitsishvili, healing book “karabadine’’, publishing house “Soviet Georgia’’ Tbilisi. 1978, p. 623.
- Kimeridze K. , Georgian Soviet Encyclopedia , vol. 10, ch. , 1986. — p. 429.
- Laboratory techniques in biochemistry and molecular biology. General editors: RH. BURDON and P.H van KNIPPENBERG; techniques in free radical research; Catherine A. Rice-Evan, Anthony T. Diplock, Martyn C.R. Symons. 1991 ELSEVIER. Pg. 30-34.
- Harborne, J.B. Phytochemical methods: A guide to modern techniques of plant analysis, 2nd ed.; Chapman and Hall: London, 1998; pp. 54–84. [Google Scholar]
- Kokate, K.C. Practical pharmacognosy, 4th ed.; Vallabh Prakashan: Delhi, 1997; p. 218. [Google Scholar]
- WAGNERANDBLADT-plant-drug-analysis-a-thin-layer-chromatography-atlas-(359-364) 2001.
- Aslam, K.; Nawchoo, I.A.; Bhat, M.A.; Ganie, A.H.; Aslam, N. Ethno-pharmacological review of genus Primula. International Journal of Advanced Research 2014, 2, 29–34. [Google Scholar]
- Arteaga, J.F.; Ruiz-Montoya, M.; Palma, A.; Alonso-Garrido, G.; Pintado, S.; Rodríguez-Mellado, J.M. Comparison of the simple cyclic voltammetry (CV) and DPPH assays for the determination of antioxidant capacity of active principles. Molecules 2012, 17, 5126–5138. [Google Scholar] [CrossRef]
- Liang, N.; Kitts, D.D. Antioxidant Property of Coffee Components: Assessment of Methods that Define Mechanisms of Action. Molecules 2014, 19, 19180–19208. [Google Scholar] [CrossRef]
- Noroozisharaf, A.; Samizadeh Lahiji, H.; Hatamzadeh, A.; Bakhshi, D. Phytochemical attributes of endemic endangered primrose (Primula heterochroma Stapf.) accessions grown in Iran. Physiol Mol Biol Plants. 2015, 21, 573–81. [Google Scholar] [CrossRef]
- Tarapatskyy, M.; Gumienna, A.; Sowa, P.; Kapusta, I.; Puchalski, C. Bioactive Phenolic Compounds from Primula veris L.: Influence of the Extraction Conditions and Purification. Molecules. 2021, 26, 997. [Google Scholar] [CrossRef]
- Franck, T.; Mouithys-Mickalad, A.; Robert, T.; et al. Differentiation between stoichiometric and anticatalytic antioxidant properties of benzoic acid analogues: a structure/redox potential relationship study. Chemico-biological Interactions 2013, 206, 194–203. [Google Scholar] [CrossRef] [PubMed]
- Derochette, S.; Franck, T.; Mouithys-Mickalad, A.; Deby-Dupont, G.; Neven, P.; Serteyn, D. Serteyn, Intra- and extracellular antioxidant capacities of the new water soluble form of curcumin (NDS27) on stimulated neutrophils and HL-60 cells. Chem. Biol. Interact. 2013, 201, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, K.L.; Liu, R.H. Cellular antioxidant activity (CAA) assay for assessing antioxidants, foods, and dietary supplements. Agric. Food Chem. 2007, 55, 8896–8907. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.A.; Neupane, G.P.; Lee, E.S.; Jeong, B.S.; Park, B.C.; Thapa, P. NADPH oxidase inhibitors: a patent review. Expert Opin. Ther. Pat. 2011, 21, 1147–1158. [Google Scholar] [CrossRef]
- Malle, E.; Furtmüller, P.G.; Sattler, W.; Obinger, C. Myeloperoxidase: a target for new drug development. Br J. Pharmacol. 2007, 152, 838–854. [Google Scholar] [CrossRef]
- Li, Y.; Zhu, H.; Kuppusamy, P.; Roubaud, V.; Zweier, J.L.; Trush, M.A. Validation of lucigenin (bis-N-methylacridinium) as a chemilumigenic probe for detecting superoxide anion radical production by enzymatic and cellular systems. J. Biol. Chem. 1998, 273, 2015–2023. [Google Scholar] [CrossRef]
- Porto, B.N.; Stein, R.T. Neutrophil Extracellular Traps in Pulmonary Diseases: Too Much of a Good Thing? Front Immunol. 2016, 7, 311. [Google Scholar] [CrossRef]
- Seibel, J.; Wonnemann, M.; Werz, O.; et al. A tiered approach to investigate the mechanism of anti-inflammatory activity of an herbal medicinal product containing a fixed combination of thyme herb and primula root extracts. Clin Phytosci 2018, 4, 4. [Google Scholar] [CrossRef]
- González, R.; Ballester, I.; López-Posadas, R.; Suárez, M.D.; Zarzuelo, A.; Martínez-Augustin, O.; Sánchez de Medina, F. Effects of flavonoids and other polyphenols on inflammation. Crit Rev Food Sci Nutr. 2011, 51, 331–62. [Google Scholar] [CrossRef] [PubMed]
- Ciz, M.; Denev, P.; Kratchanova, M.; Vasicek, O.; Ambrozova, G.; Lojek, A. Flavonoids inhibit the respiratory burst of neutrophils in mammals. Oxid Med Cell Longev. 2012, 2012, 181295. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, D.; Freitas, M.; Lima, J.L.; Fernandes, E. Flavonoids inhibit the production of cytokines/chemokines and induce apoptosis in human neutrophils. Free Radic Biol Med. 2014, 75 Suppl 1, S46. [Google Scholar] [CrossRef] [PubMed]
- Davies, M.J. Myeloperoxidase-derived oxidation: mechanisms of biological damage and its prevention. J. Clin. Biochem. Nutr. 2011, 48, 8–19. [Google Scholar] [CrossRef]
- Delporte, C.; Franck, T.; Noyon, C.; Dufour, D.; Rousseau, A.; Madhoun, P.; Desmet, J.M.; Serteyn, D.; Raes, M.; Nortier, J.; Vanhaeverbeek, M. Simultaneous measurement of protein-bound 3-chlorotyrosine and homocitrulline by LCMS/MS after hydrolysis assisted by microwave: application to the study of myeloperoxidase activity during hemodialysis. Talanta 2012, 99, 603–609. [Google Scholar] [CrossRef] [PubMed]
- Franck, T.; Kohnen, S.; Boudjeltia, K.Z.; Van Antwerpen, P.; Bosseloir, A.; Niesten, A.; Gach, O.; Nys, M.; Deby-Dupont, G.; Serteyn, D. A new easy method for specific measurement of active myeloperoxidase in human biological fluids and tissue extracts. Talanta 2009, 80, 723–729. [Google Scholar] [CrossRef]
- Franck, T.; Kohnen, S.; Deby-Dupont, G.; Grulke, S.; Deby, C.; Serteyn, D. A specific method for measurement of equine active myeloperoxidase in biological samples and in in vitro tests. J. Vet. Diagn. Invest. 2006, 18, 326–334. [Google Scholar] [CrossRef]
- Nyssen, P.; Franck, T.; Serteyn, D.; Mouithys-Mickalad, A.; Hoebeke, M. Propofol metabolites and derivatives inhibit the oxidant activities of neutrophils and myeloperoxidase. Free Radic Biol Med. 2022, 191, 164–175. [Google Scholar] [CrossRef] [PubMed]
- Shiba, Y.; Kinoshita, T.; Chuman, H.; Taketani, Y.; Takeda, E.; Kato, Y.; Naito, M.; Kawabata, K.; Ishisaka, A.; Terao, J.; Kawai, Y. Flavonoids as substrates and inhibitors of myeloperoxidase: molecular actions of aglycone and metabolites. Chem Res Toxicol. 2008, 21, 1600–9. [Google Scholar] [CrossRef]
- Phenolic Compounds: Structure, Classification, and Antioxidant Power; Milena Morandi Vuolo1, Verena Silva Lima1,2, Mário Roberto Maróstica Junior. [CrossRef]
- Blois, M. Antioxidant determinations by the use of a stable free radical. Nature. 1958, 181, 1199–1200. [Google Scholar] [CrossRef]
- Bondet, V.; Brand-Williams, W.; Berset, C. Kinetics and mechanisms of antioxidant activity using the DPPH• free radical method. Lebensmittel-Wissenschaft und -Technologie- Food Science and Technology. 1997, 30, 609–615. [Google Scholar] [CrossRef]
- Etsè, K.S.; Etsè, K.D.; Nyssen, P.; Mouithys-Mickalad, A. Assessment of anti-inflammatory-like, antioxidant activities and molecular docking of three alkynyl-substituted 3-ylidene-dihydrobenzo[d]isothiazole 1,1-dioxide derivatives. Chem Biol Interact. 2021, 344, 109513. [Google Scholar] [CrossRef] [PubMed]
- Benbarek, H.; Deby-Dupont, G.; Deby, C.; Caudron, I.; Mathy-Hartert, M.; Lamy, M.; Serteyn, D. Experimental model for the study by chemiluminescence of the activation of isolated equine leucocytes. Res. Vet. Sci. 1996, 61, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Franck, T.; Kohnen, S.; De la Rebière, G.; Deby-Dupont, G.; Deby, C.; Niesten, A.; Serteyn, D. Activation of equine neutrophils by phorbol myristate acetate or N-formyl-methionyl-leucyl-phenylalanine induces a different response in reactive oxygen species production and release of active myeloperoxidase. Vet. Immunol. Immunopathol. 2009, 130, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Miller, N.J.; Rice-Evans, C.; Davies, M.J.; Gopinathan, V.; Milner, A. A novel method for measuring antioxidant capacity and its application to monitoring the antioxidant status in premature neonates. Clinical Science. 1993, 84, 407–412. [Google Scholar] [CrossRef]
- Goel, G.; Makkar, H.P.S.; Francis, G.; Becker, K. Phorbol Esters: Structure, Biological Activity, and Toxicity in Animals. Int. J. Toxicol. 2007, 26, 279–288. [Google Scholar] [CrossRef] [PubMed]
- Pagani, A.; Gaeta, S.; Savchenko, A.I.; Williams, C.M.; Appendino, G. An Improved Preparation of Phorbol from Croton Oil. Beilstein J. Org. Chem. 2017, 13, 1361–1367. [Google Scholar] [CrossRef]
Figure 2.
Relative ABTS and DPPH radical scavenging capacity derived from Primula macroxalyx (P.m.) (A), Primula woronowii (P.w) (B) Primula saguramica (C) fractions obtained after ethanol extraction and freeze drying (1) or after chromatographic separation with H2O, 50 % MeOH, 100 % MeOH and EtOAc (fractions 2, 3, 4 and 5 respectively). Gallic acid (GA) and quercetin (Qu) were used as reference compounds. All the extract powders were solubilized in DMSO and their scavenging capacity were compared to DMSO control (Ctrl DMSO) and ABTS or DPPH solution without sample (Ctrl negative). Results were obtained after 30 min incubation time and expressed in relative percentages vs Ctrl negative set as 100 % (Mean +/- SD, n=3 independent experiments with 3 technical replicates for each experiment).
Figure 2.
Relative ABTS and DPPH radical scavenging capacity derived from Primula macroxalyx (P.m.) (A), Primula woronowii (P.w) (B) Primula saguramica (C) fractions obtained after ethanol extraction and freeze drying (1) or after chromatographic separation with H2O, 50 % MeOH, 100 % MeOH and EtOAc (fractions 2, 3, 4 and 5 respectively). Gallic acid (GA) and quercetin (Qu) were used as reference compounds. All the extract powders were solubilized in DMSO and their scavenging capacity were compared to DMSO control (Ctrl DMSO) and ABTS or DPPH solution without sample (Ctrl negative). Results were obtained after 30 min incubation time and expressed in relative percentages vs Ctrl negative set as 100 % (Mean +/- SD, n=3 independent experiments with 3 technical replicates for each experiment).
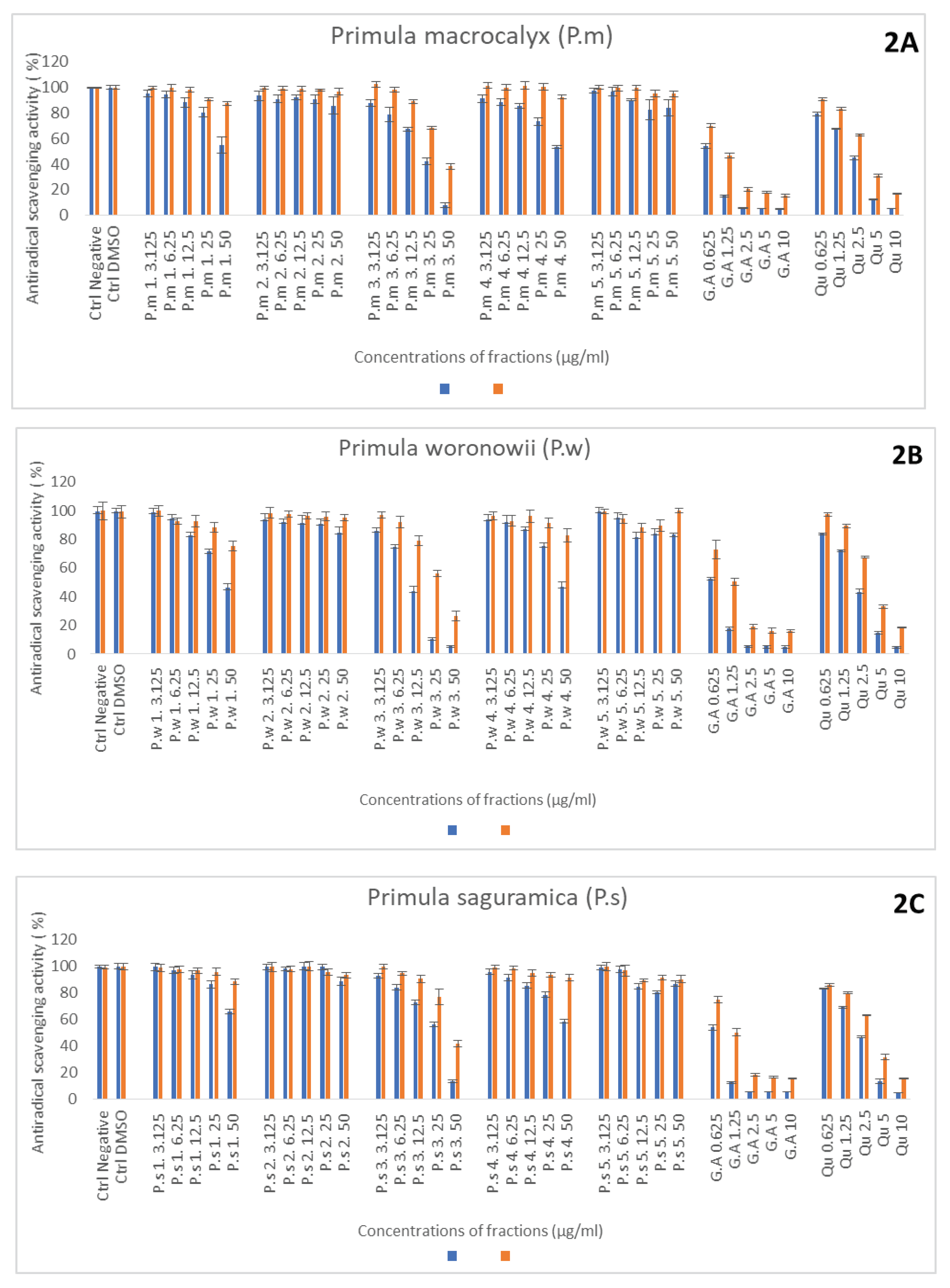
Figure 3.
Relative inhibition of neutrophils ROS production induced by Primula macroxalyx (P.m.) Primula woronowii (P.w) and Primula saguramica fractions obtained after ethanol extraction and freeze drying (fraciton 1) or after chromatographic separation with H2O, MeOH (50% and 100%), and EtOAc (fractions 2, 3, 4 and 5, respectively). Gallic acid (GA) and quercetin (Qu) were used as reference compounds. All the powdered extracts were solubilized in DMSO and their ROS inhibition capacity were compared to DMSO control (Ctrl DMSO) and stimulated neutrophil without sample (ST PMN). Results were obtained after 30 min of neutrophil stimulation with PMA and expressed in relative percentages vs ST PMN set as 100 % response (Mean +/- SD, n=3 independent experiments, with 3 technical replicates for each experiment). NON-ST PMN: Non stimulated PMN. .
Figure 3.
Relative inhibition of neutrophils ROS production induced by Primula macroxalyx (P.m.) Primula woronowii (P.w) and Primula saguramica fractions obtained after ethanol extraction and freeze drying (fraciton 1) or after chromatographic separation with H2O, MeOH (50% and 100%), and EtOAc (fractions 2, 3, 4 and 5, respectively). Gallic acid (GA) and quercetin (Qu) were used as reference compounds. All the powdered extracts were solubilized in DMSO and their ROS inhibition capacity were compared to DMSO control (Ctrl DMSO) and stimulated neutrophil without sample (ST PMN). Results were obtained after 30 min of neutrophil stimulation with PMA and expressed in relative percentages vs ST PMN set as 100 % response (Mean +/- SD, n=3 independent experiments, with 3 technical replicates for each experiment). NON-ST PMN: Non stimulated PMN. .
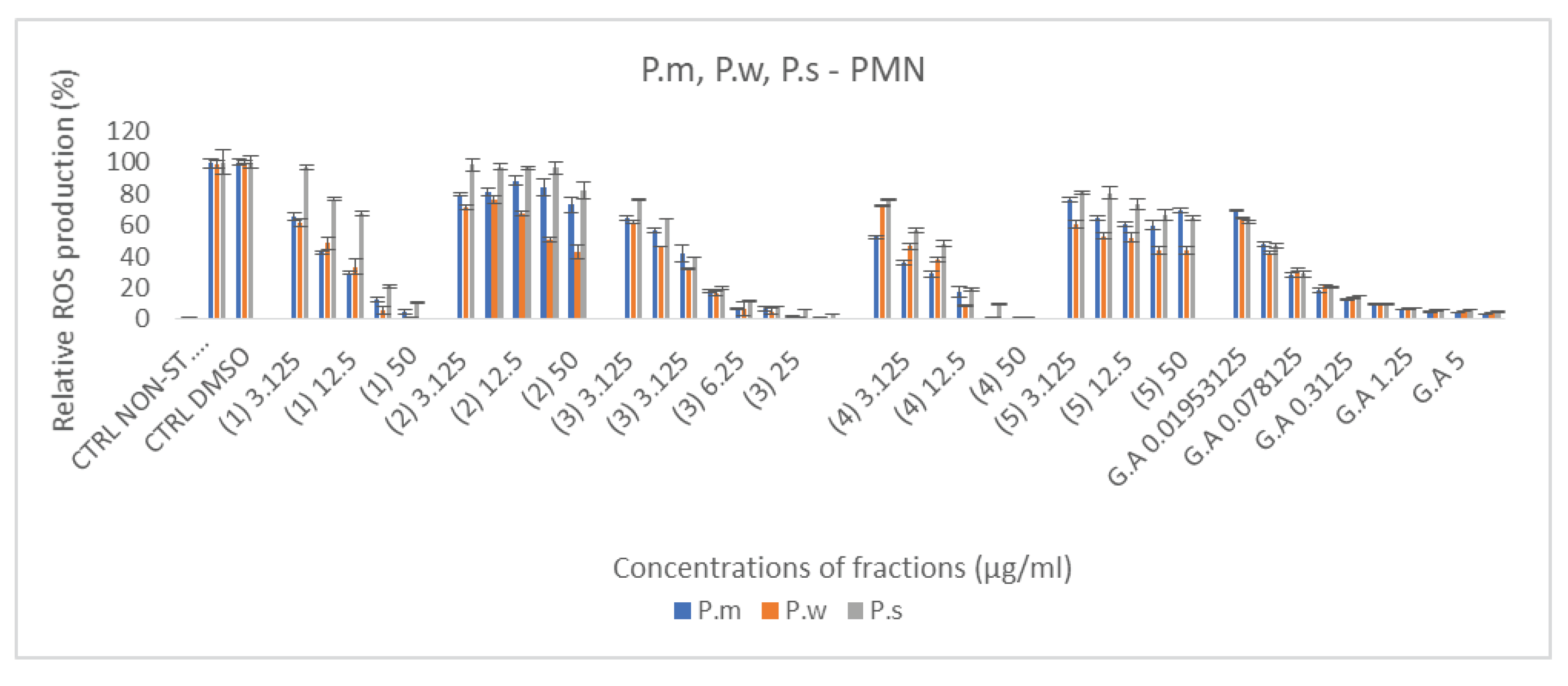
Figure 4.
Inhibition of MPO activity measured by SIEFED and classical assay by Primula macroxalyx (P.m.) Primula woronowii (P.w) and Primula saguramica fractions obtained after ethanol extraction and freeze drying (1) or after chromatographic separation with H2O, 50 % MeOH, 100 % MeOH and EtOAc (fractions 2, 3 4 and 5 respectively). Gallic acid (GA) and quercetin (Qu) were used as reference compounds. All the powedered extracts were solubilized in DMSO and their MPO inhibition was compared to DMSO control (Ctrl DMSO) and MPO control without sample. Results were expressed in relative percentages vs Ctrl MPO set as 100 % (Mean +/- SD, n=3 independent experiments with 3 technical replicates for each experiment).
Figure 4.
Inhibition of MPO activity measured by SIEFED and classical assay by Primula macroxalyx (P.m.) Primula woronowii (P.w) and Primula saguramica fractions obtained after ethanol extraction and freeze drying (1) or after chromatographic separation with H2O, 50 % MeOH, 100 % MeOH and EtOAc (fractions 2, 3 4 and 5 respectively). Gallic acid (GA) and quercetin (Qu) were used as reference compounds. All the powedered extracts were solubilized in DMSO and their MPO inhibition was compared to DMSO control (Ctrl DMSO) and MPO control without sample. Results were expressed in relative percentages vs Ctrl MPO set as 100 % (Mean +/- SD, n=3 independent experiments with 3 technical replicates for each experiment).
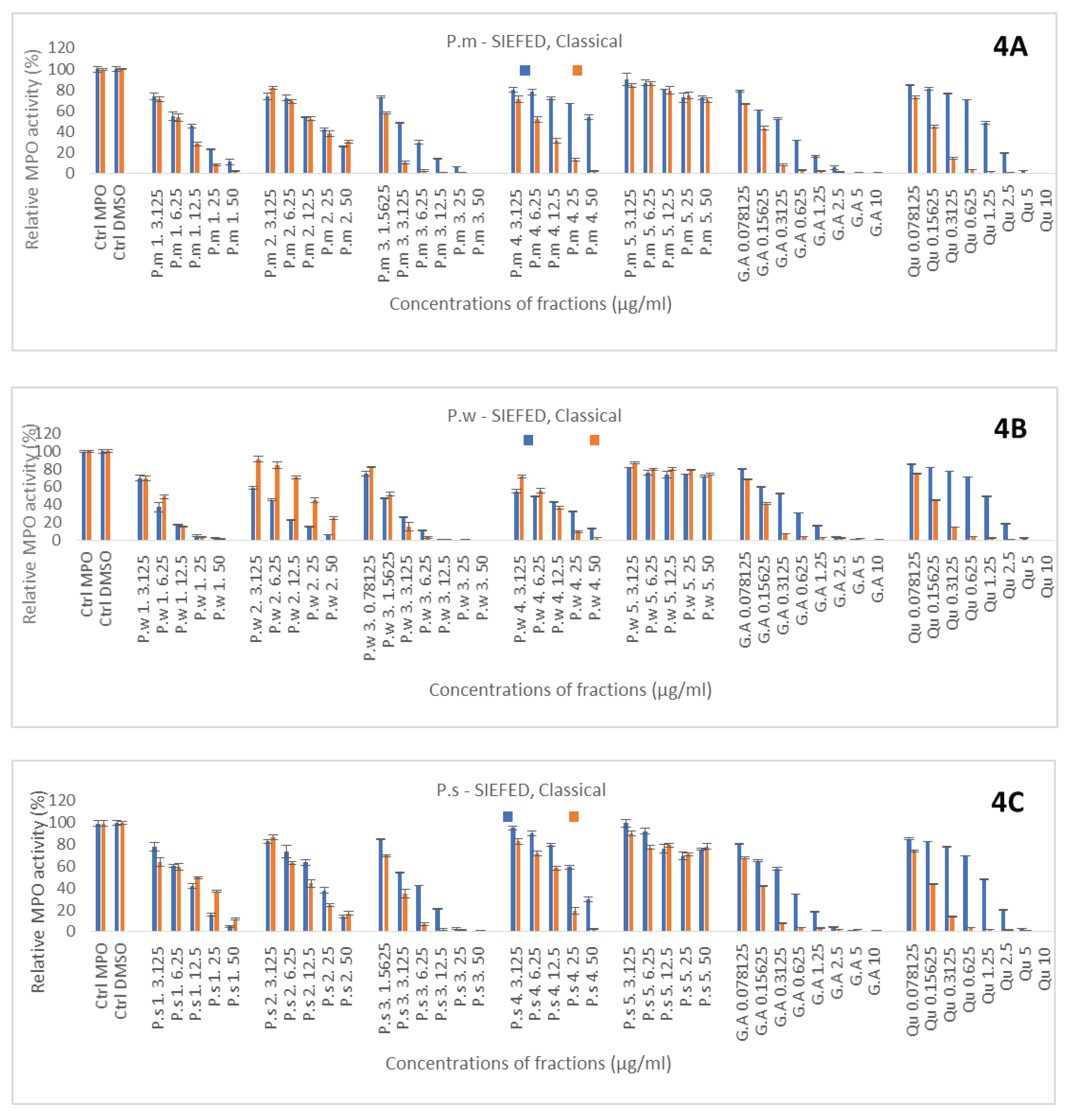
Figure 5.
Extraction procedure and sampling of Primula species fractions.
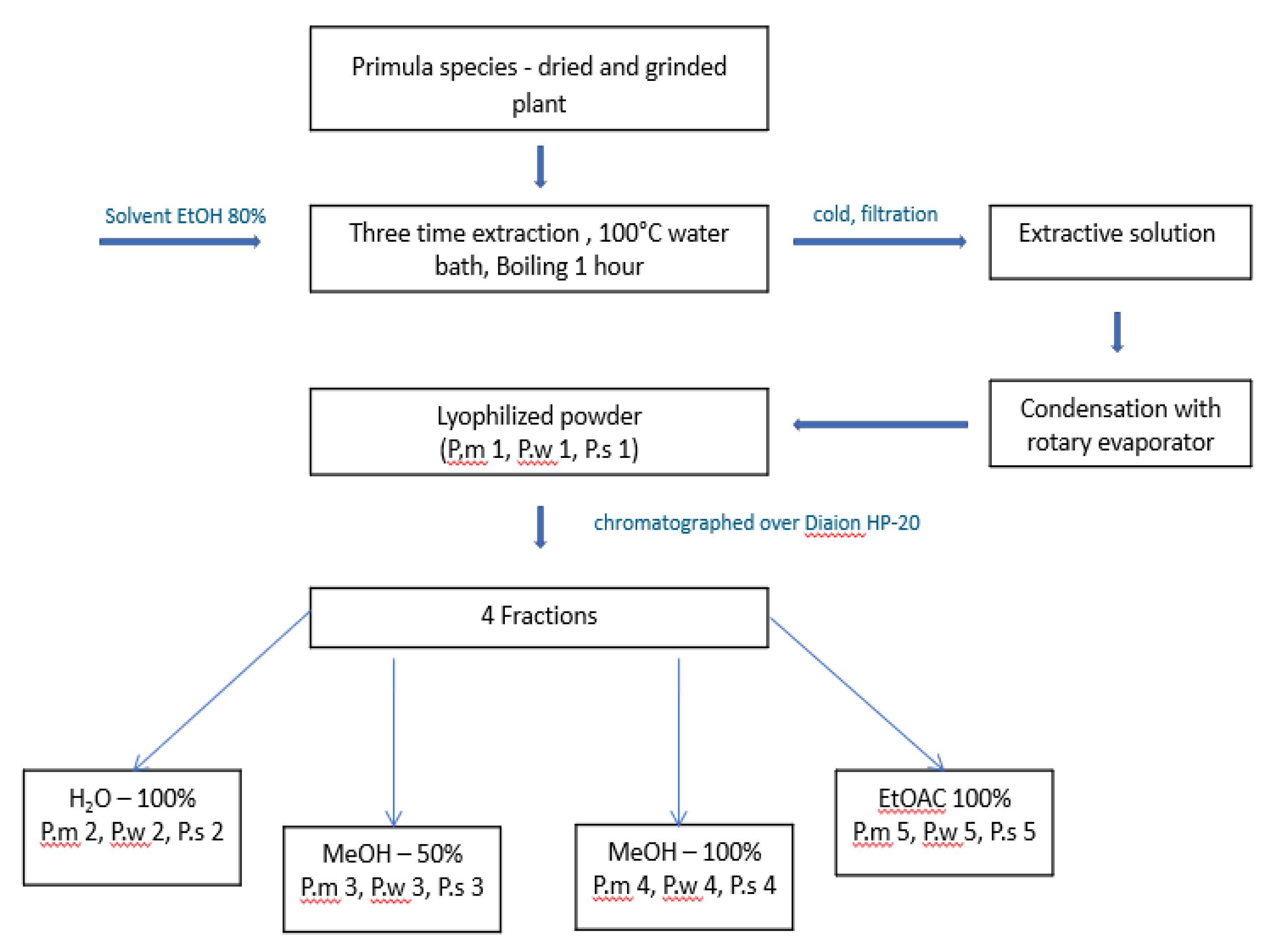

Table 1.
Determination of IC50 (µg/ml).
| Name | ABTS | DPPH | SIEFED | Classical | PMN |
| P.m 1 | > 50 | > 50 | 11.89 ± 0.6 | 9.8 ± 0.1 | 5.1 ± 0.3 |
| P.w 1 | 45.4 ± 2.9 | > 50 | 4.8 ± 0.4 | 5.9 ± 0.7 | 6.44 ± 0.2 |
| P.s 1 | > 50 | > 50 | 11.58 ± 0.7 | 14.1 ± 0.1 | 16.8 ±0.02 |
| P.m 2 | > 50 | > 50 | 17.28 ± 0.8 | 11.9 ± 0.1 | > 50 |
| P.w 2 | > 50 | > 50 | 5.2 ± 0.3 | 17.8 ± 0.1 | 25.4 ± 0.01 |
| P.s 2 | > 50 | > 50 | 22.32 ± 1.02 | 9.25 ± 0.04 | > 50 |
| P.m 3 | 12.88 | 36.47 ± 0.13 | 4.44 ± 0.4 | 1.96 ± 0.2 | 1.17 ± 0.07 |
| P.w 3 | 8.51 ± 0.18 | 34.57 ± 0.47 | 1.49 ± 0.7 | 1.89 ± 0.01 | 0.68 ± 0.04 |
| P.s 3 | 27.9 | 48.88 ± 0.5 | 4.89 ± 0.9 | 2.71 ± 0.9 | 1.1 ± 0.03 |
| P.m 4 | > 50 | > 50 | > 50 | 10.51 ± 0.1 | 1.4 ± 0.03 |
| P.w 4 | 48.1 ± 2.1 | > 50 | 6.02 ± 0.2 | 11.53 ± 0.3 | 2.8 ± 0.2 |
| P.s 4 | > 50 | > 50 | 33.9 ± 0.9 | 15.68 ± 0.1 | 5.3 ± 0.1 |
| P.m 5 | > 50 | > 50 | > 50 | > 50 | > 50 |
| P.w 5 | > 50 | > 50 | > 50 | > 50 | > 50 |
| P.s 5 | > 50 | > 50 | > 50 | > 50 | > 50 |
| Galic acid | 0.66 ± 0.15 | 1.09 ± 0.02 | 0.53 ± 0.04 | 0.18 ± 0.02 | 0.03 ± 0.01 |
| Quercetin | 2.07 ± 0.01 | 2.59 ± 0.07 | 1.37 ± 0.02 | 0.18 ± 0.02 | 0.14 ± 0.01 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



