
Submitted:
28 September 2023
Posted:
29 September 2023
You are already at the latest version
Abstract
Castration of stallions is traditionally performed after puberty around the age of 2 years old. No studies have focused on the effects of early castration on osteoarticular metabolism. Thus, we sought to compare early castration (3 days after birth) with traditional castration (18 months of age) in horses. Testosterone and estradiol levels were monitored from birth to 33 months in these two groups. We quantified the levels of biomarkers of cartilage and bone anabolism (CPII and N-MID) and catabolism (CTX-I and CTX-II), of osteoarthritis (HA and COMP) and inflammation (IL-6 and PGE2). We revealed a lack of parallelism between testosterone and estradiol syntheses after birth and during puberty in both groups. An extra-gonadal synthesis of steroids was observed around the 28 month-mark, regardless of the castration age. We found the expression of estrogen receptor (ESR1) in cartilage and bone, whereas androgen receptor (AR) expression appeared to be restricted to bone. Nevertheless, with regards to osteoarticular metabolism, steroid hormone deprivation resulting from early castration showed no discernable impact on the levels of biomarkers related to bone and cartilage metabolism, nor on those associated with OA and inflammation. Consequently, our research demonstrated that early castration does not disrupt bone and cartilage homeostasis.
Keywords:
horse
; castration
; steroid hormones
; cartilage
; bone
; in vivo metabolism
1. Introduction
Steroid hormones, such as testosterone and estradiol, are derived from cholesterol during steroidogenesis. Testosterone is a direct precursor of estradiol and is converted by the enzymatic complex composed of cytochrome P450 and aromatase, which is encoded by a single gene, Cyp19a1 [1,2]. Two distinct postnatal periods of testosterone synthesis have been observed in males. Specifically, a transient peak in synthesis is observed during the perinatal period [3]. Subsequently, the production of testosterone decreases significantly. Afterward, testosterone levels increase to a peak during puberty [4]. In adulthood, these levels remain high until a decline in elderly men. In contrast to testosterone, the synthesis of estradiol is characterized by a single increase in production during puberty. Following this period, estradiol levels remain low in men. This estradiol synthesis is carried on by many tissues in men, it is estimated that the testis is responsible for 20% of the total synthesis [5].
The endocrine function of the testis along with the widespread distribution of steroid hormone receptors in various tissues, highlights the range of physiological processes regulated by the steroid hormones [6]. Numerous studies conducted in human and rodent have shown that testosterone and estradiol play a significant role in the physiology of both bone and cartilage [7,8,9,10]. The biological effects of these two steroids are mediated through their specific receptors: the classical estrogen receptors ESR1 and ESR2, and the androgen receptor AR. Specifically , ESR1, ESR2, and AR have been demonstrated to be expressed in osteocytes, osteoblasts, and osteoclasts for bone, as well as in chondrocytes for cartilage [11,12,13,14,15].
Testosterone and estradiol play essential roles in the growth, development, and maintenance of the skeleton in various species [16,17]. During the pubertal stage, the elevation of steroid hormones is linked to the peak in bone growth observed during this period [18,19]. Moreover, sex steroid hormones play a significant role in skeletal sexual dimorphism [15,20,21]. In addition, testosterone plays a crucial role in growth and contributes to periosteal apposition, which is radial growth. These differences result in longer (8%) and wider bones, as well as 25% more bone mass in men compared to women [22]. Puberty concludes with the closure of the epiphyseal growth plate. In fact, at the culmination of puberty, the elevated concentrations of estrogens stimulate the closure of the epiphyses through their influence on growth plate chondrocytes, occurring in both boys and girls [18].
Throughout adult life, there exists a balance between bone formation performed by osteoblasts and resorption carried out by osteoclasts [23,24]. The decline in circulating levels of sex steroid hormones observed with aging is linked to bone loss in both men and women [25,26]. Consequently, the prevalence of osteoporosis dramatically increases with age [27]. The pivotal role of steroid hormones in maintaining bone homeostasis has also been demonstrated in clinical instances of gonadal failure [28,29], cases involving ESR1 mutations [30,31], or CYP19A1 mutations [32], as well as in mouse models [33,34,35,36]. Finally, it has been established that estrogens play a considerably more significant role in human bone metabolism than testosterone [25]. Steroid hormones also have a role in maintaining articular cartilage homeostasis. This avascular tissue is primarily composed of chondrocytes, which synthesize a specialized and abundant extracellular matrix (ECM). In osteoarthritis (OA), a degenerative disease of articular cartilage, the balance of tissue homeostasis is disturbed, favoring catabolism [37]. The leading risk factors for OA are age, obesity, and joint injury. A difference in the prevalence of OA between men and women has also been well established. Indeed OA rarely appears in women before the age of 45, but its frequency increases significantly after menopause, suggesting that estrogen deficiency could play a role in the disease [8,10]. In addition, it has been demonstrated that postmenopausal estrogen replacement therapy might offer protection against OA of the hip in elderly women [38]. Furthermore, it has been described that estradiol stimulates the expressions of type II collagen, a specific marker of hyaline cartilage, and UDP-glucose dehydrogenase, a key enzyme in glycosaminoglycan (GAG) synthesis, in articular chondrocytes [39,40]. Moreover, castration in rabbits resulted in an increase in apoptosis and a decrease in chondrocytes proliferation in the growth plate [41].
In horses, castration is a common surgical procedure performed to mitigate the fiery and sometimes aggressive character of stallions. This practice entails suppressing the main production of gonadal steroid, in particular testosterone, which affects behavior through its impact on the brain [42]. The stallion is the male mammal that produces the highest levels of testicular estrogens, distinguishing it from other species [43,44]. Currently, castration is typically carried out after puberty during the phase of skeletal growth (before the age of 2) or later, around the age of 3, once growth has concluded, aiming to attain the morphological characteristics of the stallion [45,46]. Performing this procedure at an earlier stage is infrequent, and limited information is available regarding its potential consequences.
In our previous study, the impact of early-age castration in neonates (at 3 days of age) compared to traditional castration (at 18 months of age) on the physical and behavioral development of horses was evaluated [47]. They found no pre-, intra-, or post-operative complications related to early castration in foals. Moreover, early castration did not disrupt the morphological or behavioral development of horses followed up to the age of 3 years. In this context, the objective of the present study was to assess the medium-term side effects of early castration compared to traditional castration on osteoarticular metabolism, focusing on the circulating levels of testosterone and estradiol in the same cohort of horses. First of all, the expression of steroid receptors (AR and ESR1) in bone and cartilage tissues was investigated to identify if these tissues are steroid targets. The presence of enzymes responsible of last step synthesis of testosterone (CYP17A1) and estradiol (CYP19) were also investigated in neonatal testes to determine the contribution of testis to circular rate of these hormones. Finally, the impact of early or traditional castration on bone anabolism and catabolism was investigated. Thus, we quantified biomarkers indicating bone (Cross-linked C-telopeptide of Type I Collagen or CTX-I) and cartilage (Cross-linked C-telopeptide of Type II Collagen or CTX-II), catabolism, biomarkers of bone (N-terminal midfragment of osteocalcin or N-MID) and cartilage (carboxypropetide of type II collagen or CPII) anabolism, and biomarkers of OA (hyaluronic acid or HA, cartilage oligomeric matrix protein or COMP) and inflammation (prostaglandin E2 or PGE2, Interleukin 6 or IL-6) in serum or plasma from birth to 33 months of age for both groups of horses.
2. Results
2.1. Investigation of ESR1 and AR detection in horse bone and cartilage tissues
To explore equine skeletal as steroid target, the expression of ESR1 and AR was examined in the cartilage and bone tissues of adult horses using immunohistochemistry (Figure 1). Staining for ESR1 was observed in chondrocytes (Figure 1A) as well as in osteocytes within Havers systems (osteon) and the interstitial lamella of compact bone (Figure 1B). For AR, no signal was detected in cartilage tissue (Figure 1C), while a small number of cells within bone tissue exhibited expression of this receptor (Figure 1D). These data suggest that estradiol could influence both bone and cartilage metabolism, whereas testosterone may only affect bone metabolism.
2.2. Analysis of circulating levels and testicular synthesis of testosterone in both study groups
Circulating testosterone levels were measured from birth up to 33 months using plasma samples collected from both study groups (Figure 2). As expected, a rate fluctuation was observed in group 1 (horses castrated at 18 months) between 0 and 33 months. Indeed, an initial increase in circulating levels of testosterone was observed during the first months after birth, peaking around 2 months. Subsequently, a second significant increase in plasma testosterone levels occurred between 7 and 17 months, reaching its maximum synthesis at 12 months (456.1 pg/mL ± 40.02). Following castration at 18 months, a decline in circulating testosterone levels was noted, eventually reaching levels slightly above those in horses castrated at birth (22 months: 22.37 pg/mL ± 3.43).
In group 2 (horses castrated at birth), the circulating testosterone level was stable and low but remain detectable, registering as soon as 1 month (12.85 pg/mL ± 1.33), in contrast to the control group, and it remained stable until 22 months. Then, in both groups, an increase in testosterone levels was noted at 28 months, even after castration, reaching levels comparable to those measured at birth (43.66 pg/mL ± 6.58 for group 1 and 66.37 pg/mL ± 8.66 for group 2).
To ascertain whether plasma testosterone levels were linked to gonadal synthesis in neonates, we examined the protein expression of the enzyme CYP17A1, which plays a role in the last step of androgen synthesis. Through immunohistological analysis, we successfully identified the expression of this enzyme in testes of neonates (Figure 3A). Specifically, CYP17A1 was immunolocalized within the interstitial tissue of the testes in 3-day-old foals, as observed in adult testes (Figure 3B), confirming the capacity of the testis to synthesize androgens throughout the perinatal period.
2.3. Analysis of circulating levels and testicular synthesis of estradiol in both study groups
Circulating estradiol levels were quantified from birth up to 33 months using plasma samples collected from both study groups (Figure 4).
High estradiol levels were observed at birth in both study groups (1866.75 pg/mL ± 390.96 for group 1, castrated at 18 months and 1202.6 pg/mL ± 445.5 for group 2, castrated at birth). A substantial reduction (approximately 8.5-fold) in plasma estradiol levels was noted in both groups starting from 1 month after birth, with no significant difference between the two groups (group 1: 211.17 pg/mL ± 35.78 and group 2: 146 pg/mL ± 33.80). Following this pronounced decline, these levels gradually decreased for up to 3 months and remained consistently low in both groups until 12 months (about 56 pg/mL). Subsequently, after reaching one year of age, a significant increase in estradiol levels was observed in group 1, peaking at 14 months (409.80 pg/mL ± 109.67). After castration of horses at 18 months (group 1), the reduction in estradiol levels continued reaching levels equivalent to those of horses castrated at birth by 22 months (group 2: 44.48 pg/mL ± 12.04). In both study groups, estradiol levels increased after 22 months, regardless of early or late castration, reaching levels by 33 months similar to those observed during the pubertal peak at 14 months (362 pg/mL ± 42.49 for group 1 at 33 months and 544.04 pg/mL ± 84.07 for group 2). These findings indicate an extragonadal synthesis of estradiol.
To determine whether plasma estradiol levels at birth could be linked to gonadal synthesis, the presence of aromatase enzyme, responsible for the aromatization of testosterone into estradiol, was examined in the testes of 3-day-old animals (Figure 5A). However, the immunohistological analysis did not reveal the presence of aromatase in the testes of 3-day-old foals contrary to adults. Indeed, an interstitial expression of aromatase was observed (Figure 5B).
2.4. Analysis of circulating levels of osteoarticular metabolism biomarkers in both study groups
In addition to gonadal biomarkers, aspects of the osteo-articular status were studied to evaluate the impact of sex steroid deprivation on osteoarticular metabolism. Thus, biomarkers of bone anabolism (N-MID; Figure 6A) and catabolism (CTX-I; Figure 6B), cartilage anabolism (CPII; Figure 7A) and catabolism (CTX-II; Figure 7B), as well as biomarkers of osteoarthritis (COMP and HA; Figure 7C and 7D), were quantified from birth to 33 months using blood samples collected from both groups of castrated horses. The profiles of these biomarkers were consistent between both groups regardless of the castration stage, showing no statistically significant differences. Thus, the levels of different biomarkers were not affected by castration. However, their levels exhibited variation over time.
For bone markers, N-MID levels gradually decreased over time in a similar manner for both study groups (from an average level of 117.9 ng/mL at birth to an average level of 6.3 ng/mL at 33 months for the entire group, indicating an 18.7-fold decrease; Figure 6A). In contrast, CTX-I levels increased progressively until 33 months (from an average level of 0.118 ng/mL at birth to an average level of 0.327 ng/mL at 33 months for the entire group, representing a 2.7-fold increase; Figure 6B) with no significant difference between the two groups. We observed a peak in CTX-I levels at 22 months (with an average level of 0.493 ng/ml). These findings indicate an enhancement of bone matrix catabolism during foal growth, irrespective of whether castration was performed early or late. These results may also suggest an increase in the turnover of bone ECM (osteoformation/osteolysis imbalance) during foal growth.
For cartilage markers, CPII levels progressively increased by approximately 2-fold from birth to 33 months, and this trend was consistent in both study groups (with average levels of 1862.5 ng/mL at birth and 3815.9 ng/mL at 33 months for the entire group; figure 7A). CTX-II levels continuously and significantly decreased by about 44-fold over the same period, and this decline occurred similarly in both study groups (with average levels of 2243.4 ng/mL and 51.3 ng/mL at birth and at 33 months for the entire group; Figure 7B). These findings support the notion of cartilage anabolism during foal growth, regardless of whether castration was performed early or later.
In addition, serum osteoarthritis markers, such as COMP and HA, were also assessed. COMP levels remained stable and comparable between both study groups throughout the analysis (with average levels of 62.2 ng/mL for group 1 castrated at 18 months and of 69.1 ng/mL for group 2 castrated at birth; Figure 7C). HA levels exhibited a gradual decrease in both study groups from birth to 33 months (with average levels of 110.7 ng/mL at birth and 37.6 ng/mL at 33 months for the entire group, representing a decrease of approximately 3-fold) and this reduction showed no significant difference between the two groups (Figure 7D).
Moreover, pro-inflammatory biomarkers such as IL-6 (Sup data 1A) and PGE2 (Sup data 1B) were measured in both study groups. These markers are produced in osteoarticular pathologies such as arthritis or, to a lesser extent, OA. Elevated levels of these markers were detected at birth, reflecting the animal’s inflammatory state following parturition. The levels of these two markers similarly decreased after birth in both study groups, and their levels did not appear to be influenced by the chirurgical procedure of castration. There was an observed progressive decrease in IL-6 levels (approximately 2.8-fold decrease from birth to 33 months for the entire group), while PGE2 levels exhibited a significant decrease during the first 2 months of the animal’s life (approximately 3.8-fold decrease for the entire group), followed by a gradual decline until 33 months (approximately 5.5-fold decrease from two months to 33 months, with average levels of 80.6 pg/mL at 33 months for the entire group). Overall, these findings suggested that neither the act of castration nor the timing of castration affected the levels of osteoarthritis and inflammatory biomarkers.
3. Discussion
In equines, castration is a common breeding practice. However, due to the lack of consensus among veterinarians and breeders, this procedure is conducted at various ages. Castration is seldom carried out during the early stages, and its effects on osteoarticular development remain unexplored. Castration could be one of the mechanisms underlying osteoarticular pathologies [10,28,48], which could impact animal welfare and escalate costs for breeders. However, unlike other farmed species (such as bovine and pigs) which are commonly castrated to enhance the farming industry performance before slaughter, horses are intended for a prolonged career. This study is part of a project that seeks to compare the medium-term effects of equine castration at 3 days of age (early castration, before pubertal action of steroids) with castration at 18 months of age (traditional castration, after pubertal action of steroids). Thus, we have already carried out two comparative studies on the surgical procedure of early and late castration in horses. We have highlighted that early castration does not impact physical and behavioral development, as well as the non-contribution of the equine testis on vitamin D metabolism [47,49]. In this study, we assessed the osteoarticular metabolism’s side effects resulting from early castration in comparison to traditional castration up until the age of 33 months.
First of all, perinatal synthesis of testicular androgens was confirmed by the immunodetection of the CYP17A1 enzyme in the testes of 3-day-old horses. Our study revealed a lack of parallelism between testosterone and estradiol plasma levels both post-birth and during puberty in horses. Testosterone synthesis in stallions is characterized by an initial transient perinatal elevation, a phenomenon described in various species and referred to as « mini puberty » [3,50,51]. The absence of steroid exposure during mini puberty in group 2 of horses castrated at birth could potentially impact animal behavior. Indeed, it has been described that mini puberty allows for masculinization and defeminization of the central nervous system in males [52,53]. This period of development involves changes in total brain volume, cortex thickness, and cortical network development [54,55,56]. As a result, early castration could influence the behavior of horses due to the absence of mini puberty. However, our previous study within the same cohort demonstrated no differences between early or traditional castration in terms of temperament and behavior at either 1 or 3 years of age [47]. Several parameters were measured, including reactivity to humans, sensory sensitivity, gregariousness, fearfulness, and activity.
Conversely, estrogen synthesis in stallions was characterized by high estradiol levels at birth, which markedly decreased during the months of life. These elevated estradiol levels noted post-birth probably reflected a maternal origin. Indeed, it has been described within the 48h preceding parturition, that 17β-estradiol levels are twice as high in mares [57]. This hypothesis was confirmed by the absence of the CYP19A1 protein, as demonstrated through immunohistochemistry in testes of 3-day-old animals. Following this, estradiol levels increased at the onset of puberty; however, this pubertal estradiol peak was both delayed and more transient (spanning from 14 to 17 months) than the testosterone pubertal peak. This delayed occurrence of the estradiol peak could potentially be attributed to different regulations of steroidogenesis enzymes in equine testes.
The monitoring of plasma steroid levels (testosterone and estradiol) revealed that horses castrated at birth (group 2) are not exposed to mini puberty or the pubertal peak in testosterone and estradiol, unlike horses subjected to traditional castration at 18 months (group 1). Moreover, even after castration at birth, measurable levels of steroids persist. The circulating testosterone levels in castrated horses exhibited another dynamic alteration following early or late orchidectomy. Notably, a slight and transient testosterone peak was observed at 28 months in both groups. Subsequently, testosterone levels returned to baseline.
The temporal fluctuations in steroid levels following castration reveal a considerable synthesis of androgen by extra-gonadal tissues. However, the question arises regarding the origin of the steroids detected post-orchidectomy. The activity of the CYP17A1 enzyme, which is involved in androgen synthesis, has been reported to be abundant in the testes, with low levels detected in the adrenal glands [58,59]. While the testes are responsible for 95 % of circulating levels of testosterone in males, the adrenal cortex has been shown to possess the ability to synthesize androgens like dehydroepiandrosterone (DHEA). DHEA can subsequently be converted into testosterone and, consequently, into estradiol in tissues that express various steroidogenic enzymes responsible for this synthesis, including bones [15,60]. Moreover, in humans, DHEA, dehydroepiandrosterone sulfate (DHEAS), and androstenedione are primarily secreted by the reticularis zona of the adrenal cortex [61]. These hormones serve as essential substrate source for the extragonadal synthesis of sex steroids. In humans, it has been observed that the secretion of adrenal androgens increases during adrenarche in children, peaking between the ages of 20 and 30 [62,63]. If we extend this timeframe to horses, it could potentially correspond to the peak in testosterone levels observed in 28-month-old horses. Adrenal androgens might assume a role in relaying testicular androgens after castration in horses, as androstenedione has been suggested to enhance bone accretion during growth [64]. Moreover, low levels of DHEA have been linked to a higher incidence of osteoporosis [65]. Besides to the aromatization of adrenal androgens, estrogens can also originate from adipose tissue. Indeed, a study conducted on ovariectomized rats demonstrated an increase in extragonadal aromatization by adipose and liver tissues, as well as by the adrenal glands, resulting in elevated blood estradiol levels [66]. The increase in estradiol levels occurring around 28 months, regardless of the age of castration, could be attributed to variations in body composition during the horse’s growth period. It has indeed been noted that the proportion of adipose tissue increases between the ages of 12 and 30 months in horses [67]. Adipose tissue has been identified as having the capability to synthesize certain steroids like estrogens [68]. Therefore, this tissue could potentially serve as the origin of the elevated estradiol levels documented in our study. Others tissues expressing aromatase (skin, bone, brain) [68] could also produce estrogens in the absence of gonadal source.
Regarding osteoarticular metabolism, we have demonstrated that the depravation of steroid hormones due to early castration does not affect the levels of biomarkers associated with bone and cartilage metabolism (there was no differences observed in the levels of N-MID, CTX-I, CPII and CTX-II between both study groups). This suggests that castration has no impact on bone loss and cartilage homeostasis. This original result goes against the findings observed in humans [7,26,28,29,69] and rodents [33,34,70], as well as in studies related to cartilage [8,10,41,71]. Nevertheless, this is in accordance with our previous results that demonstrated a low or negligible contribution of the testes to vitamin D bioactivation in horses, whereas vitamin D plays a crucial role in maintaining bone health [49]. It is possible that the low levels of steroids quantified after early castration might still be sufficient to exert their action on bone and cartilage tissues. The absence of AR expression in cartilaginous tissue and the low expression of AR in bone tissue, compared to ESR1, support the idea that estrogens play a much more important role than androgens in the regulation of bone and cartilage metabolism. This aligns with findings from human clinical studies [10,26,69,72,73]. As a result, testosterone could potentially act as a preferred substrate for estrogen synthesis, which exerts its effects through nuclear receptors like ESR1.
Among the bone markers investigated, N-MID is a specific marker of bone formation, while CTX-I is used as a marker of bone resorption, aiding in the assessment of osteoporosis risk [74,75]. We observed that N-MID levels exhibited a gradual decrease, whereas CTX-I levels increased from birth to 33 months, regardless of early or late castration. These findings rather imply a potential bone loss, which contrasts with typical bone growth in foals. Another explanation we can suggest is that these data may also reflect an increase in the turnover, in the metabolic activity of the ECM of bone tissue, in other words an enhancement in the osteolysis/osteoformation balance. Moreover, it is important to consider that these data need to be correlated with a juvenile population. Indeed, serum levels of bone turnover markers are not stable across the lifespan and tend to be higher in infants and children compared to adults [76,77]. Unlike adults, children have elevated concentrations of bone markers due to their rapid skeletal growth and heightened bone turnover rate [78]. As a result, complementary measurements in adult horses indicated that levels of these two bone markers at 33 months tend to align with adult values (data not shown). Regardless, we did not identify any differences between the two study groups. It might have been informative to complement these results with bone densitometry data and the measurement of other bone markers, such as alkaline phosphatase, for example.
In terms of cartilage markers, CP-II is a specific marker of cartilage anabolism, and CTX-II is a specific marker for cartilage degradation [79,80]. Our observations indicate that CPII levels experience a gradual and consistent increase, whereas CTX-II levels exhibit a continuous and significant decrease from birth to 33 months, regardless of the group to which the castrated horses belong. These findings support the notion of cartilage anabolism during foal growth, regardless of early or late castration. This could be correlated with the establishment of the growth plate in foals. Given cartilage degradation observed in OA is primarily and mainly correlated with aging, to our knowledge, no study has yet correlated the levels of cartilage markers with OA in a pediatric population. To address this gap, we expanded our analyses by measuring COMP and HA levels in both groups of horses. Indeed, serum COMP levels exhibit a strong correlation with the degree of OA [79,81], and serum HA levels may serve as a valuable predictor of OA progression [82,83]. We showed that COMP levels remained consistent, and HA levels gradually decreased in both study groups from birth to 33 months, regardless of the age of castration. Additionally, pro-inflammatory biomarkers (IL-6 and PGE2) were not affected by the age of castration. Taken together, these findings imply that neither castration nor the timing of castration impacted cartilage homeostasis and fate.
4. Materials and Methods
4.1. Animals and samples collection
The animal requirements adhered to the European Community Council Directive 2010/63/UE.
This study focused on two groups of 22 male Welsh ponies from an experimental herd within the PAO experimental unit (UEPAO; INRAE Val de Loire, France). All experimental procedures received approval from the Val de Loire Ethic committee (authorization N° APAFlS#4530-20l603l4150l1475 v3) [47,49]. These animals were categorized based on their age at the time of castration:
- GROUP 1: Traditional castration: foals castrated at 18 months. Horses from this group were castrated at 18 months and served as the control group for the other experimental group.
- GROUP 2: Early castration: neonates castrated at birth. Horses underwent early castration 3 days after birth.
Blood sample (S) were collected concurrently with morphometric measurements from birth until 33 months (S0 to S12: n=11, S14 to S17: n=8, S28 to S33: n=5). The blood samples were collected in either dry tubes or tubes containing anticoagulants (Heparin or EDTA) to obtain serum or plasma, respectively. Subsequently, the tubes were centrifuged at 3000 r.p.m. for 10 min at 4°C to separate the supernatants, which were then stored at -80°C until ELISA analysis.
The testicular tissues, obtained following the castration of horses from the two groups of Welsh ponies, were derived from the middle of the testis. Testes from adult horses (from an average age of 6.6 years) were collected during routine castrations (Saint-Michel du Livet veterinary clinic, Calvados, France) as positive control for immunohistological analysis. Testes were fixed in 4% PFA and embedded in paraffin for subsequent histological analysis.
Osteochondral sections were harvested from the medial part of the distal aspect of the metacarpal condyle of French Standardbred horses (median age of 3 years) as part of another study [84]. The study protocol received approval from the ComEth Anses/ENVA/UPEC Ethical Committee (permit number: 10/03/15-12). Osteochondral sections from joints treated with a physiological saline placebo were fixed with a formalin solution (SIGMA-Aldrich, Saint-Louis, MO, USA), decalcified (RDO, Eurobio Scientific, Courtaboeuf, France) and embedded in paraffin for subsequent histological analysis.
4.2. Biomarker assays
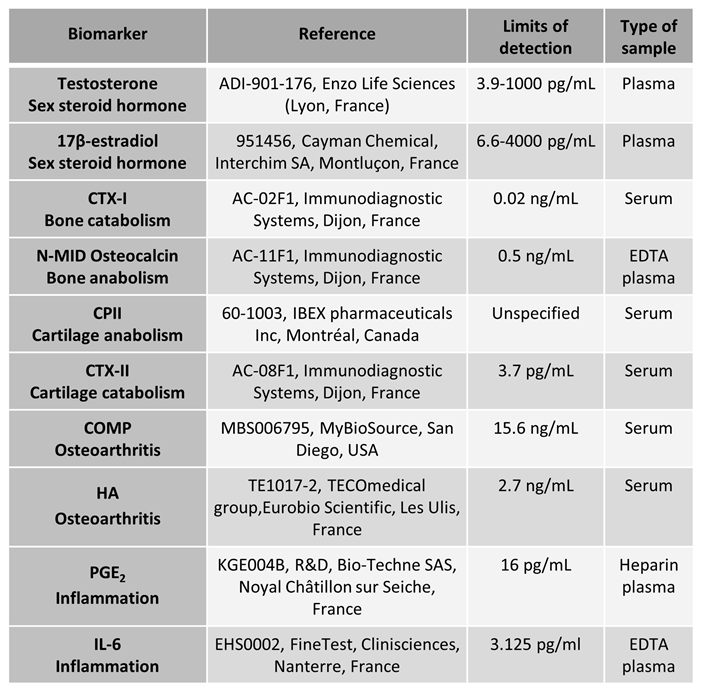
Testicular, osteoarticular, and inflammation biomarker levels were measured using ELISA kits in accordance with the manufacturer’s instructions, using plasma or serum samples. Each sample was quantified in duplicate. The kit references used in this study, along with their respective detection limits, are detailed in table 1.

Table 1.
References for the ELISA kits used.
 |
4.3. Immunohistochemistry
4.3.1. Testicular tissue
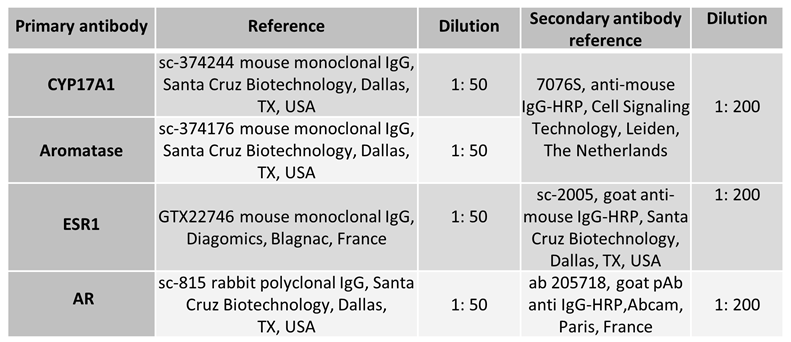
Immunohistochemistry was conducted on deparaffinized and rehydrated 5 μm testicular sections mounted on glass slides, following the previously described procedure [49]. To inhibit endogenous peroxidase activity, all sections underwent a 10-minute incubation in 3% H2O2/PBS solution. Then, antigen retrieval was achieved by immersing the sections in a sodium citrate buffer (0,1M; pH 6) and heating them in a microwave oven at 90W for 10 minutes. Following the heating step, the sections were allowed to cool for 40 minutes and then rinsed with PBS. For permeabilization, the glass slides holding the sections were treated with 0.2% Triton / PBS for 20 minutes. Then, the testicular sections were blocked with PBS containing 3% BSA for 20 minutes and then incubated with the appropriate primary antibody at 4°C overnight in a humidified chamber. Following three washes in PBS, the sections were exposed to the compatible secondary antibody for 1h30. Immunoreactivity was visualized using an indirect immunoperoxidase method following a brief rinse. A positive signal is identified through the response of the diaminobenzidine chromogen (Sigma-Aldrich, Saint-Louis, USA). Following counterstaining with Mayer’s hematoxylin (BIOCYC GmbH & CO. KG, Potsdam, Germany), the sections were dehydrated and covered with Eukitt mounting medium (ORSAtec GMBH, VWR International SAS, Rosny-sous-Bois, France). The primary and secondary antibodies used are outlined in table 2. As a negative control, the primary antibody was excluded.
Table 2.
Antibodies used for histological analyses.
 |
4.3.2. Osteochondral tissue
Immunohistochemistry was conducted on deparaffinized and rehydrated 5 μm osteochondral sections mounted on glass slides. An additional decalcification step required a 10 minutes incubation in 9% formic acid (Sigma-Aldrich, Saint-Louis, USA). Subsequently, bone sections underwent a 30-minute retrieval/blocking procedure in a solution containing 0.5% hyaluronidase and 3% BSA (Sigma-Aldrich, Saint-Louis, USA). To facilitate permeabilization, the glass slides holding the sections were exposed to a 0.2% Tween 20 / PBS solution for 10 minutes. Following this, the sections were incubated in a 1.5% H2O2 / PBS solution (Sigma-Aldrich, Saint-Louis, USA) for 20 minutes. After being rinsed thrice in PBS, the sections were placed in a humidified chamber and incubated with the appropriate primary antibody at 4°C overnight. After three PBS washes, the sections were exposed to a compatible secondary antibody for a duration of 1h30. Post-secondary antibody exposure, the osteochondral sections were washed in PBS, and immunoreactivity was visualized using the diaminobenzidine detection system (Sigma-Aldrich, Saint-Louis, USA). After counterstaining with Mayer’s hematoxylin (BIOCYC GmbH & CO. KG, Potsdam, Germany), the sections were dehydrated and then covered with Eukitt mounting medium (ORSAtec GMBH, VWR International SAS, Rosny-sous-Bois, France). The antibodies used are summarized in table 2. As a negative control, the primary antibody was excluded.
4.4. Statistical analyses
All data were reported as mean ± SEM. Statistical analyses were conducted using GraphPad Prism 8.0.1 software (San Diego, CA, USA). A Kruskal-Wallis test, followed by Dunn’s multiple comparisons test, was performed to compare biomarker concentration levels with P0 within each experimental group (castrated at 18 months and at birth). Additionally, a Mann-Whitney test was used for a statistical analysis to compare samples at the same time between the two groups. Differences were deemed significant at p ≤ 0.05.
5. Conclusions
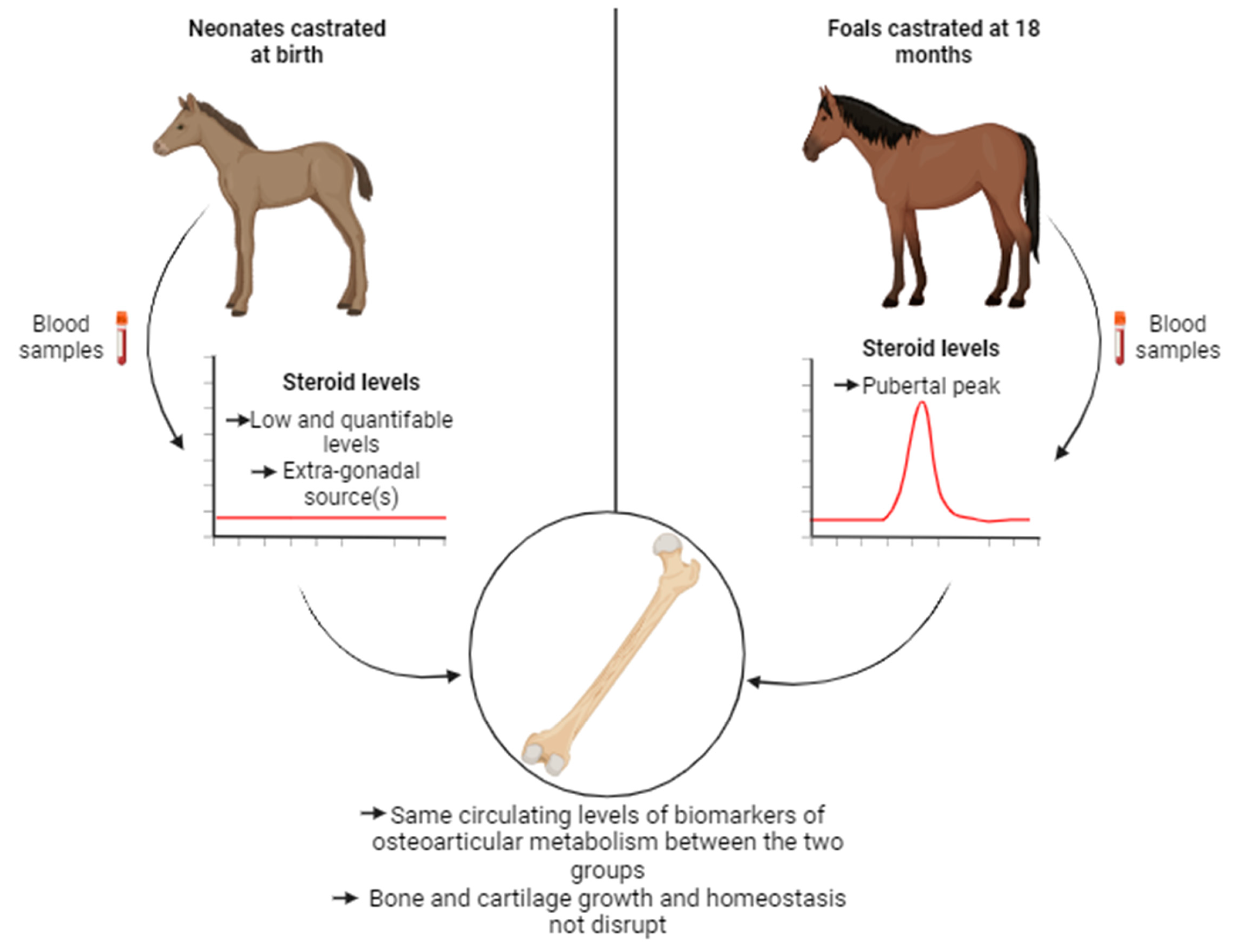
This study yielded results indicating that there are no discernable side effects of castration on osteoarticular metabolism in horses, regardless of whether the procedure was carried out 3 days after birth (early castration) or at 18 months (traditional castration) (Figure 8). In particular, the removal of the pubertal source of gonadal steroid synthesis did not disrupt the levels of biomarkers associated with bone and cartilage metabolism during growth and maturation of skeleton. We have highlighted that extra-gonadal source(s) of steroids is (are) non-negligible and could allow the maintenance of bone and cartilage homeostasis. As such, our study serves to complement the other investigations associated with this project and reinforces the idea that horses constitute an unique model in endocrinology [47,49]. Taken together with our previous studies, we have confirmed that there are no contraindications to practicing early castration. This is supported by the absence of complications related to the surgical procedure, the behavior and morphology of the animals, as well as the osteoarticular metabolism.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: Circulating levels of inflammatory biomarkers in plasma samples from horses castrated at birth and at 18 months.
Author Contributions
Conceptualization: C. Delalande, J. Cognié, F. Reigner, P. Barrière, S. Deleuze, F. Legendre, P. Galéra, V. Hanoux, and H. Bouraïma-Lelong. Methodology: J. Cognié, F. Reigner, P. Barrière, and S. Deleuze. Validation: M. Rouge, F. Legendre, C. Delalande, J. Cognié, F. Reigner, P. Barrière, V. Hanoux, P. Galéra and H. Bouraïma-Lelong. Formal analysis: M. Rouge, F. Legendre and R. Elkhatib. Investigation: M. Rouge, F. Legendre and R. Elkhatib. Ressources: J. Cognié, F. Reigner, P. Barrière and S. Deleuze. Experimentation: M. Rouge, F. Legendre and R. Elkhatib. Writing -Original Draft Preparation: M. Rouge and F. Legendre. Writing - Review and editing: M. Rouge, F. Legendre, C. Delalande, V. Hanoux, P. Galéra and H. Bouraïma-Lelong. Visualization: M. Rouge, F. Legendre, V. Hanoux, P. Galéra and H. Bouraïma-Lelong. Supervision: F. Legendre, V. Hanoux, P. Galéra and H. Bouraïma-Lelong. Funding acquisition: P. Galéra, H. Bouraïma-Lelong, J. Cognié, F. Reigner and S. Deleuze.
Funding
This work was supported by the «Région Normandie », «Institut Français du Cheval et de l’Equitation » (IFCE), by the «Ministère de l’Enseignement Supérieur, de la Recherche et de l’Innovation » (Ministry of higher education and research), by Fonds Eperon (EQUISTEM, N80-2014, 917CB194) and by the GIS CENTAURE equine research (EQUISTEM-G,014CJ061). These funding sources had no involvement in the study design, collection, analysis and interpretation of data, in the writing of the manuscript, or in the decision to submit the manuscript for publication.
Institutional Review Board Statement
This study was conducted according to European Community Council Directive 2010/63/UE and approved by the Val de Loire Ethic committee (authorization N° APAFlS#4530-20l603l4150l1475 v3) and by the ComEth Anses/ENVA/UPEC Ethical Committee (permit number: 10/03/15-12).
Informed Consent Statement
Not applicable.
Data Availability Statement
The data are presented in this study.
Acknowledgments
We thanks M. Melin (Novotec, Bron, France) for her help in preparing osteochondral sections and C. Ballandone (BIOTARGEN, UNICAEN) for her help in ELISA. We also thank the “Carré International” of the University of Caen Normandie (France) for editing the article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Carreau, S.; Genissel, C.; Bilinska, B.; Levallet, J. Sources of Oestrogen in the Testis and Reproductive Tract of the Male. Int. J. Androl. 1999, 22, 211–223. [Google Scholar] [CrossRef]
- Payne, A.H.; Hales, D.B. Overview of Steroidogenic Enzymes in the Pathway from Cholesterol to Active Steroid Hormones. Endocr. Rev. 2004, 25, 947–970. [Google Scholar] [CrossRef]
- Andersson, A.-M.; Toppari, J.; Haavisto, A.-M.; Petersen, J.H.; Simell, T.; Simell, O.; Skakkeb, N.E. Longitudinal Reproductive Hormone Profiles in Infants: Peak of Inhibin B Levels in Infant Boys Exceeds Levels in Adult Men. 1998, 83, 7.
- Zirkin, B.R.; Papadopoulos, V. Leydig Cells: Formation, Function, and Regulation. Biol. Reprod. 2018, 99, 101–111. [Google Scholar] [CrossRef]
- Vermeulen, A.; Kaufman, J.M.; Goemaere, S.; Van Pottelberg, I. Estradiol in Elderly Men. Aging Male 2002, 5, 98–102. [Google Scholar] [CrossRef]
- Fagerberg, L.; Hallström, B.M.; Oksvold, P.; Kampf, C.; Djureinovic, D.; Odeberg, J.; Habuka, M.; Tahmasebpoor, S.; Danielsson, A.; Edlund, K.; et al. Analysis of the Human Tissue-Specific Expression by Genome-Wide Integration of Transcriptomics and Antibody-Based Proteomics. Mol. Cell. Proteomics 2014, 13, 397–406. [Google Scholar] [CrossRef]
- Cauley, J.A. Estrogen and Bone Health in Men and Women. Steroids 2015, 99, 11–15. [Google Scholar] [CrossRef]
- Gokhale, J.A.; Frenkel, S.R.; Dicesare, P.E. Estrogen and Osteoarthritis. Am. J. Orthop. Belle Mead NJ 2004, 33, 71–80. [Google Scholar]
- Lorentzon, M.; Swanson, C.; Andersson, N.; Mellström, D.; Ohlsson, C. Free Testosterone Is a Positive, Whereas Free Estradiol Is a Negative, Predictor of Cortical Bone Size in Young Swedish Men: The good study: estradiol predicts crotical bone size in men. J. Bone Miner. Res. 2005, 20, 1334–1341. [Google Scholar] [CrossRef]
- Roman-Blas, J.A.; Castañeda, S.; Largo, R.; Herrero-Beaumont, G. Osteoarthritis Associated with Estrogen Deficiency. Arthritis Res. Ther. 2009, 11, 241. [Google Scholar] [CrossRef]
- Bord, S. Estrogen Receptors and Are Differentially Expressed in Developing Human Bone. J. Clin. Endocrinol. Metab. 2001, 86, 2309–2314. [Google Scholar] [CrossRef]
- Khalid, A.B.; Krum, S.A. Estrogen Receptors Alpha and Beta in Bone. Bone 2016, 87, 130–135. [Google Scholar] [CrossRef] [PubMed]
- Nasatzky, E.; Schwartz, Z.; Soskolne, W.A.; Brooks, B.P.; Dean, D.D.; Boyan, B.D.; Ornoy, A. Evidence for Receptors Specific for 17 Beta-Estradiol and Testosterone in Chondrocyte Cultures. Connect. Tissue Res. 1994, 30, 277–294. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, O.; Chrysis, D.; Pajulo, O.; Boman, A.; Holst, M.; Rubinstein, J.; Martin Ritzén, E.; Sävendahl, L. Localization of Estrogen Receptors-Alpha and -Beta and Androgen Receptor in the Human Growth Plate at Different Pubertal Stages. J. Endocrinol. 2003, 177, 319–326. [Google Scholar] [CrossRef] [PubMed]
- Vanderschueren, D.; Vandenput, L.; Boonen, S.; Lindberg, M.K.; Bouillon, R.; Ohlsson, C. Androgens and Bone. Endocr. Rev. 2004, 25, 389–425. [Google Scholar] [CrossRef]
- Khosla, S.; Monroe, D.G. Regulation of Bone Metabolism by Sex Steroids. Cold Spring Harb. Perspect. Med. 2018, 8, a031211. [Google Scholar] [CrossRef]
- Nakamura, T.; Imai, Y.; Matsumoto, T.; Sato, S.; Takeuchi, K.; Igarashi, K.; Harada, Y.; Azuma, Y.; Krust, A.; Yamamoto, Y.; et al. Estrogen Prevents Bone Loss via Estrogen Receptor α and Induction of Fas Ligand in Osteoclasts. Cell 2007, 130, 811–823. [Google Scholar] [CrossRef]
- Juul, A. The Effects of Oestrogens on Linear Bone Growth. Hum. Reprod. Update 2001, 7, 303–313. [Google Scholar] [CrossRef]
- Tanner, J.M.; Davies, P.S.W. Clinical Longitudinal Standards for Height and Height Velocity for North American Children. J. Pediatr. 1985, 107, 317–329. [Google Scholar] [CrossRef]
- Cadogan, J.; Blumsohn, A.; Barker, M.E.; Eastell, R. A Longitudinal Study of Bone Gain in Pubertal Girls: Anthropometric and Biochemical Correlates. J. Bone Miner. Res. 1998, 13, 1602–1612. [Google Scholar] [CrossRef]
- Wen, P.; Cowell, T.; Lloyd-Jones, A.; Briody, N. Volumetric Bone Mineral Density in Normal Subjects, Aged 5-27 Years. M. 1996, 5. [Google Scholar]
- Boot, A.M.; de Ridder, M.A.J.; van der Sluis, I.M.; van Slobbe, I.; Krenning, E.P.; de Muinck Keizer-Schrama, S.M.P.F. Peak Bone Mineral Density, Lean Body Mass and Fractures. Bone 2010, 46, 336–341. [Google Scholar] [CrossRef] [PubMed]
- Harada, S.; Rodan, G.A. Control of Osteoblast Function and Regulation of Bone Mass. Nature 2003, 423, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Rodan, G.A.; Martin, T.J. Therapeutic Approaches to Bone Diseases. Science 2000, 289, 1508–1514. [Google Scholar] [CrossRef] [PubMed]
- Khosla, S.; Amin, S.; Orwoll, E. Osteoporosis in Men. Endocr. Rev. 2008, 29, 441–464. [Google Scholar] [CrossRef]
- Khosla, S.; Melton, L.J.; Atkinson, E.J.; O’Fallon, W.M.; Klee, G.G.; Riggs, B.L. Relationship of Serum Sex Steroid Levels and Bone Turnover Markers with Bone Mineral Density in Men and Women: A Key Role for Bioavailable Estrogen. J. Clin. Endocrinol. Metab. 1998, 83, 2266–2274. [Google Scholar] [CrossRef]
- Looker, A.C.; Orwoll, E.S.; Johnston, C.C.; Lindsay, R.L.; Wahner, H.W.; Dunn, W.L.; Calvo, M.S.; Harris, T.B.; Heyse, S.P. Prevalence of Low Femoral Bone Density in Older U.S. Adults from NHANES III. J. Bone Miner. Res. 1997, 12, 1761–1768. [Google Scholar] [CrossRef]
- Behre, H.M.; Kliesch, S.; Leifke, E.; Link, T.M.; Nieschlag, E. Long-Term Effect of Testosterone Therapy on Bone Mineral Density in Hypogonadal Men. J. Clin. Endocrinol. Metab. 1997, 82, 2386–2390. [Google Scholar] [CrossRef]
- Bhasin, S.; Cunningham, G.R.; Hayes, F.J.; Matsumoto, A.M.; Snyder, P.J.; Swerdloff, R.S.; Montori, V.M. ; Task Force, Endocrine Society Testosterone Therapy in Men with Androgen Deficiency Syndromes: An Endocrine Society Clinical Practice Guideline. J. Clin. Endocrinol. Metab. 2010, 95, 2536–2559. [Google Scholar] [CrossRef]
- Smith, E.; Korach, K. Oestrogen Receptor Deficiency: Consequences for Growth. Acta Paediatr. 1996, 85, 39–43. [Google Scholar] [CrossRef]
- Smith, E.P.; Boyd, J.; Frank, G.R.; Takahashi, H.; Cohen, R.M.; Specker, B.; Williams, T.C.; Lubahn, D.B.; Korach, K.S. Estrogen Resistance Caused by a Mutation in the Estrogen-Receptor Gene in a Man. N. Engl. J. Med. 1994, 331, 1056–1061. [Google Scholar] [CrossRef]
- Carani, C.; Qin, K.; Simoni, M.; Faustini-Fustini, M.; Serpente, S.; Boyd, J.; Korach, K.S.; Simpson, E.R. Effect of Testosterone and Estradiol in a Man with Aromatase Deficiency. N. Engl. J. Med. 1997, 337, 91–95. [Google Scholar] [CrossRef] [PubMed]
- Moverare, S.; Venken, K.; Eriksson, A.-L.; Andersson, N.; Skrtic, S.; Wergedal, J.; Mohan, S.; Salmon, P.; Bouillon, R.; Gustafsson, J.-A.; et al. Differential Effects on Bone of Estrogen Receptor and Androgen Receptor Activation in Orchidectomized Adult Male Mice. Proc. Natl. Acad. Sci. 2003, 100, 13573–13578. [Google Scholar] [CrossRef] [PubMed]
- Notini, A.J.; McManus, J.F.; Moore, A.; Bouxsein, M.; Jimenez, M.; Chiu, W.M.; Glatt, V.; Kream, B.E.; Handelsman, D.J.; Morris, H.A.; et al. Osteoblast Deletion of Exon 3 of the Androgen Receptor Gene Results in Trabecular Bone Loss in Adult Male Mice. J. Bone Miner. Res. 2007, 22, 347–356. [Google Scholar] [CrossRef] [PubMed]
- Peng, Z.; Li, X.; Mäkelä, S.; Väänänen, H.K.; Poutanen, M. Skeletal Changes in Transgenic Male Mice Expressing Human Cytochrome P450 Aromatase. J. Bone Miner. Res. Off. J. Am. Soc. Bone Miner. Res. 2004, 19, 1320–1328. [Google Scholar] [CrossRef]
- Vico, L.; Vanacker, J.-M. Sex Hormones and Their Receptors in Bone Homeostasis: Insights from Genetically Modified Mouse Models. Osteoporos. Int. 2010, 21, 365–372. [Google Scholar] [CrossRef] [PubMed]
- Demoor, M.; Ollitrault, D.; Gomez-Leduc, T.; Bouyoucef, M.; Hervieu, M.; Fabre, H.; Lafont, J.; Denoix, J.-M.; Audigié, F.; Mallein-Gerin, F.; et al. Cartilage Tissue Engineering: Molecular Control of Chondrocyte Differentiation for Proper Cartilage Matrix Reconstruction. Biochim. Biophys. Acta 2014, 1840, 2414–2440. [Google Scholar] [CrossRef]
- Nevitt, M.C.; Cummings, S.R.; Lane, N.E.; Hochberg, M.C.; Scott, J.C.; Pressman, A.R.; Genant, H.K.; Cauley, J.A. Association of Estrogen Replacement Therapy with the Risk of Osteoarthritis of the Hip in Elderly White Women. Study of Osteoporotic Fractures Research Group. Arch. Intern. Med. 1996, 156, 2073–2080. [Google Scholar] [CrossRef]
- Maneix, L.; Servent, A.; Porée, B.; Ollitrault, D.; Branly, T.; Bigot, N.; Boujrad, N.; Flouriot, G.; Demoor, M.; Boumediene, K.; et al. Up-Regulation of Type II Collagen Gene by 17β-Estradiol in Articular Chondrocytes Involves Sp1/3, Sox-9, and Estrogen Receptor α. J. Mol. Med. Berl. Ger. 2014, 92, 1179–1200. [Google Scholar] [CrossRef]
- Maneix, L.; Beauchef, G.; Servent, A.; Wegrowski, Y.; Maquart, F.X.; Boujrad, N.; Flouriot, G.; Pujol, J.P.; Boumediene, K.; Galéra, P.; et al. 17β-Oestradiol up-Regulates the Expression of a Functional UDP-Glucose Dehydrogenase in Articular Chondrocytes: Comparison with Effects of Cytokines and Growth Factors. Rheumatology 2008, 47, 281–288. [Google Scholar] [CrossRef]
- Irie, T.; Aizawa, T.; Kokubun, S. The Role of Sex Hormones in the Kinetics of Chondrocytes in the Growth Plate: A STUDY IN THE RABBIT. J. Bone Joint Surg. Br. 2005, 87-B, 1278–1284. [CrossRef]
- Mhaouty-Kodja, S.; Naulé, L.; Capela, D. Sexual Behavior: From Hormonal Regulation to Endocrine Disruption. Neuroendocrinology 2018, 107, 400–416. [Google Scholar] [CrossRef]
- Eisenhauer, K.M.; McCue, P.M.; Nayden, D.K.; Osawa, Y.; Roser, J.F. Localization of Aromatase in Equine Leydig Cells. Domest. Anim. Endocrinol. 1994, 11, 291–298. [Google Scholar] [CrossRef]
- Raeside, J.I. The Isolation of Estrone Sulfate and Estradiol-17 Beta Sulfate from Stallion Testes. Can. J. Biochem. 1969, 47, 811–815. [Google Scholar] [CrossRef] [PubMed]
- Line, S.W.; Hart, B.L.; Sanders, L. Effect of Prepubertal versus Postpubertal Castration on Sexual and Aggressive Behavior in Male Horses. J. Am. Vet. Med. Assoc. 1985, 186, 249–251. [Google Scholar] [PubMed]
- Moll, H.D.; Pelzer, K.D.; Pleasant, R.S.; Modransky, P.D.; May, K.A. A Survey of Equine Castration Complications. J. Equine Vet. Sci. 1995, 15, 522–526. [Google Scholar] [CrossRef]
- Cognie, J.; Freret, S.; Lansade, L.; Parias, C.; Barriere, P.; Gesbert, A.; Reigner, F.; Deleuze, S. Early Castration in Foals: Consequences on Physical and Behavioural Development. Equine Vet. J. 2023, 55, 214–221. [Google Scholar] [CrossRef] [PubMed]
- Sims, N.A.; Clément-Lacroix, P.; Minet, D.; Fraslon-Vanhulle, C.; Gaillard-Kelly, M.; Resche-Rigon, M.; Baron, R. A Functional Androgen Receptor Is Not Sufficient to Allow Estradiol to Protect Bone after Gonadectomy in Estradiol Receptor–Deficient Mice. J. Clin. Invest. 2003, 111, 1319–1327. [Google Scholar] [CrossRef]
- Rouge, M.; Elkhatib, R.; Delalande, C.; Cognié, J.; Reigner, F.; Barriere, P.; Deleuze, S.; Cousty, M.; Legendre, F.; Galera, P.; et al. Investigation of Equine Testis Contribution to Vitamin D Bioactivation. Domest. Anim. Endocrinol. 2022, 79, 106691. [Google Scholar] [CrossRef]
- Chen, L.; Wang, R.; Wang, W.; Lu, W.; Xiao, Y.; Wang, D.; Dong, Z. Hormone Inhibition During Mini-Puberty and Testicular Function in Male Rats. Int. J. Endocrinol. Metab. 2015, 13, e25465. [Google Scholar] [CrossRef]
- Winter, J.S.; Hughes, I.A.; Reyes, F.I.; Faiman, C. Pituitary-Gonadal Relations in Infancy: 2. Patterns of Serum Gonadal Steroid Concentrations in Man from Birth to Two Years of Age. J. Clin. Endocrinol. Metab. 1976, 42, 679–686. [Google Scholar] [CrossRef]
- Jost, A.; Vigier, B.; Prépin, J.; Perchellet, J.P. Studies on Sex Differentiation in Mammals. In Proceedings of the 1972 Laurentian Hormone Conference; Elsevier; 1973; pp. 1–6. [Google Scholar]
- Wilson, C.A.; Davies, D.C. The Control of Sexual Differentiation of the Reproductive System and Brain. Reproduction 2007, 133, 331–359. [Google Scholar] [CrossRef]
- Gao, W.; Alcauter, S.; Elton, A.; Hernandez-Castillo, C.R.; Smith, J.K.; Ramirez, J.; Lin, W. Functional Network Development During the First Year: Relative Sequence and Socioeconomic Correlations. Cereb. Cortex 2015, 25, 2919–2928. [Google Scholar] [CrossRef] [PubMed]
- Knickmeyer, R.C.; Gouttard, S.; Kang, C.; Evans, D.; Wilber, K.; Smith, J.K.; Hamer, R.M.; Lin, W.; Gerig, G.; Gilmore, J.H. A Structural MRI Study of Human Brain Development from Birth to 2 Years. J. Neurosci. 2008, 28, 12176–12182. [Google Scholar] [CrossRef] [PubMed]
- Lyall, A.E.; Shi, F.; Geng, X.; Woolson, S.; Li, G.; Wang, L.; Hamer, R.M.; Shen, D.; Gilmore, J.H. Dynamic Development of Regional Cortical Thickness and Surface Area in Early Childhood. Cereb. Cortex 2015, 25, 2204–2212. [Google Scholar] [CrossRef] [PubMed]
- Barnes, R.J.; Nathanielsz, P.W.; Rossdale, P.D.; Comline, R.S.; Silver, M. Plasma Progestagens and Oestrogens in Fetus and Mother in Late Pregnancy. J. Reprod. Fertil. Suppl. 1975, 617–623. [Google Scholar]
- Legacki, E.L.; Ball, B.A.; Corbin, C.J.; Loux, S.C.; Scoggin, K.E.; Stanley, S.D.; Conley, A.J. Equine Fetal Adrenal, Gonadal and Placental Steroidogenesis. Reprod. Camb. Engl. 2017, 154, 445–454. [Google Scholar] [CrossRef]
- Schlinger, B.A.; Lane, N.I.; Grisham, W.; Thompson, L. Androgen Synthesis in a Songbird: A Study of Cyp17 (17alpha-Hydroxylase/C17,20-Lyase) Activity in the Zebra Finch. Gen. Comp. Endocrinol. 1999, 113, 46–58. [Google Scholar] [CrossRef]
- Witchel, S.F.; Pinto, B.; Burghard, A.C.; Oberfield, S.E. Update on Adrenarche. Curr. Opin. Pediatr. 2020, 32, 574–581. [Google Scholar] [CrossRef]
- Turcu, A.F.; Auchus, R.J. Adrenal Steroidogenesis and Congenital Adrenal Hyperplasia. Endocrinol. Metab. Clin. North Am. 2015, 44, 275–296. [Google Scholar] [CrossRef]
- Labrie, F.; Bélanger, A.; Cusan, L.; Gomez, J.L.; Candas, B. Marked Decline in Serum Concentrations of Adrenal C19 Sex Steroid Precursors and Conjugated Androgen Metabolites during Aging. J. Clin. Endocrinol. Metab. 1997, 82, 2396–2402. [Google Scholar] [CrossRef]
- Migeon, C.J.; Keller, A.R.; Lawrence, B.; Shepard, T.H. Dehydroepiandrosterone and Androsterone Levels in Human Plasma: Effect of Age and Sex; Day-to-Day and Diurnal Variations. J. Clin. Endocrinol. Metab. 1957, 17, 1051–1062. [Google Scholar] [CrossRef]
- Remer, T.; Manz, F.; Hartmann, M.F.; Schoenau, E.; Wudy, S.A. Prepubertal Healthy Children’s Urinary Androstenediol Predicts Diaphyseal Bone Strength in Late Puberty. J. Clin. Endocrinol. Metab. 2009, 94, 575–578. [Google Scholar] [CrossRef] [PubMed]
- Holló, I.; Fehér, T.; Szücs, J. Serum Dehydroepiandrosterone, Androsterone and Cortisol Level in Primary Postmenopausal and Other Type Osteoporosis. Acta Med Acad Sci Hung. 1970. [Google Scholar]
- Zhao, H.; Tian, Z.; Hao, J.; Chen, B. Extragonadal Aromatization Increases with Time after Ovariectomy in Rats. Reprod. Biol. Endocrinol. 2005, 3, 6. [Google Scholar] [CrossRef]
- The Growing Horse: Nutrition and Prevention of Growth Disorders; Juliand, V., Martin-Rosset, W., Eds.; EAAP Scientific Series; Wageningen Academic Publishers: The Netherlands, 2005; Vol. 114; ISBN 978-90-76998-62-6. EAAP Scientific Series.
- Nelson, L.R.; Bulun, S.E. Estrogen Production and Action. J. Am. Acad. Dermatol. 2001, 45, S116–S124. [Google Scholar] [CrossRef] [PubMed]
- Falahati-Nini, A.; Riggs, B.L.; Atkinson, E.J.; O’Fallon, W.M.; Eastell, R.; Khosla, S. Relative Contributions of Testosterone and Estrogen in Regulating Bone Resorption and Formation in Normal Elderly Men. J. Clin. Invest. 2000, 106, 1553–1560. [Google Scholar] [CrossRef] [PubMed]
- Verhas, M.; Schoutens, A.; L’hermite-Baleriaux, M.; Dourov, N.; Verschaeren, A.; Mone, M.; Heilporn, A. The Effect of Orchidectomy on Bone Metabolism in Aging Rats. Calcif. Tissue Int. 1986, 39, 74–77. [Google Scholar] [CrossRef]
- Sniekers, Y.H.; Weinans, H.; Bierma-Zeinstra, S.M.; van Leeuwen, J.P.T.M.; van Osch, G.J.V.M. Animal Models for Osteoarthritis: The Effect of Ovariectomy and Estrogen Treatment - a Systematic Approach. Osteoarthritis Cartilage 2008, 16, 533–541. [Google Scholar] [CrossRef]
- Peshkova, M.; Lychagin, A.; Lipina, M.; Di Matteo, B.; Anzillotti, G.; Ronzoni, F.; Kosheleva, N.; Shpichka, A.; Royuk, V.; Fomin, V.; et al. Gender-Related Aspects in Osteoarthritis Development and Progression: A Review. Int. J. Mol. Sci. 2022, 23, 2767. [Google Scholar] [CrossRef]
- Slemenda, C.W.; Longcope, C.; Zhou, L.; Hui, S.L.; Peacock, M.; Johnston, C.C. Sex Steroids and Bone Mass in Older Men. Positive Associations with Serum Estrogens and Negative Associations with Androgens. J. Clin. Invest. 1997, 100, 1755–1759. [Google Scholar] [CrossRef]
- Eastell, R.; Hannon, R.A. Biomarkers of Bone Health and Osteoporosis Risk. Proc. Nutr. Soc. 2008, 67, 157–162. [Google Scholar] [CrossRef]
- Nagasue, K.; Inaba, M.; Okuno, S.; Kitatani, K.; Imanishi, Y.; Ishimura, E.; Miki, T.; Kim, M.; Nishizawa, Y. Serum N-Terminal Midfragment vs. Intact Osteocalcin Immunoradiometric Assay as Markers for Bone Turnover and Bone Loss in Hemodialysis Patients. Biomed. Pharmacother. Biomedecine Pharmacother. 2003, 57, 98–104. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, D.; Intemann, T.; Lauria, F.; Mårild, S.; Molnár, D.; Moreno, L.A.; Sioen, I.; Tornaritis, M.; Veidebaum, T.; Pigeot, I.; et al. Reference Values of Bone Stiffness Index and C-Terminal Telopeptide in Healthy European Children. Int. J. Obes. 2014, 38, S76–S85. [Google Scholar] [CrossRef]
- Rauchenzauner, M.; Schmid, A.; Heinz-Erian, P.; Kapelari, K.; Falkensammer, G.; Griesmacher, A.; Finkenstedt, G.; Högler, W. Sex- and Age-Specific Reference Curves for Serum Markers of Bone Turnover in Healthy Children from 2 Months to 18 Years. J. Clin. Endocrinol. Metab. 2007, 92, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Ambroszkiewicz, J.; Gajewska, J.; Rowicka, G.; Klemarczyk, W.; Chelchowska, M. Assessment of Biochemical Bone Turnover Markers and Bone Mineral Density in Thin and Normal-Weight Children. Cartilage 2018, 9, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Bleasel, J.F.; Poole, A.R.; Heinegård, D.; Saxne, T.; Holderbaum, D.; Ionescu, M.; Jones, P.; Moskowitz, R.W. Changes in Serum Cartilage Marker Levels Indicate Altered Cartilage Metabolism in Families with the Osteoarthritis-Related Type II Collagen Gene COL2A1 Mutation. Arthritis Rheum. 1999, 42, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Mouritzen, U.; Christgau, S.; Lehmann, H.-J.; Tankó, L.B.; Christiansen, C. Cartilage Turnover Assessed with a Newly Developed Assay Measuring Collagen Type II Degradation Products: Influence of Age, Sex, Menopause, Hormone Replacement Therapy, and Body Mass Index. Ann. Rheum. Dis. 2003, 62, 332–336. [Google Scholar] [CrossRef] [PubMed]
- Hoch, J.M.; Mattacola, C.G.; Medina McKeon, J.M.; Howard, J.S.; Lattermann, C. Serum Cartilage Oligomeric Matrix Protein (SCOMP) Is Elevated in Patients with Knee Osteoarthritis: A Systematic Review and Meta-Analysis. Osteoarthritis Cartilage 2011, 19, 1396–1404. [Google Scholar] [CrossRef]
- Saruga, T.; Sasaki, E.; Inoue, R.; Chiba, D.; Ota, S.; Iwasaki, H.; Uesato, R.; Nakaji, S.; Ishibashi, Y. Usefulness of Serum Hyaluronic Acid Levels as a Predictor of Incidence of Hand Osteoarthritis Analyzed by Longitudinal Analysis from the Iwaki Cohort. Sci. Rep. 2021, 11, 4074. [Google Scholar] [CrossRef]
- Sasaki, E.; Tsuda, E.; Yamamoto, Y.; Maeda, S.; Inoue, R.; Chiba, D.; Fujita, H.; Takahashi, I.; Umeda, T.; Nakaji, S.; et al. Serum Hyaluronic Acid Concentration Predicts the Progression of Joint Space Narrowing in Normal Knees and Established Knee Osteoarthritis – a Five-Year Prospective Cohort Study. Arthritis Res. Ther. 2015, 17, 283. [Google Scholar] [CrossRef]
- Bertoni, L.; Jacquet-Guibon, S.; Branly, T.; Legendre, F.; Desancé, M.; Mespoulhes, C.; Melin, M.; Hartmann, D.-J.; Schmutz, A.; Denoix, J.-M.; et al. An Experimentally Induced Osteoarthritis Model in Horses Performed on Both Metacarpophalangeal and Metatarsophalangeal Joints: Technical, Clinical, Imaging, Biochemical, Macroscopic and Microscopic Characterization. PloS One 2020, 15, e0235251. [Google Scholar] [CrossRef]
Figure 1.
Immunolocalization of ESR1 and AR in cartilage (A and C) and bone from horse (B and D). Sections from cartilage and bone showed immunostaining for ESR1 in chondrocytes (A; arrow) and in osteocytes (B; arrow). Sections from cartilage and bone showed immunostaining for AR in few osteocytes (D; arrow), but not in chondrocytes (C). The insert did not show immunostaining for ESR1 and AR when the section was incubated with secondary antibody only. Images shown are representative of the experimental results obtained from three horses (100 μm scale bar).
Figure 1.
Immunolocalization of ESR1 and AR in cartilage (A and C) and bone from horse (B and D). Sections from cartilage and bone showed immunostaining for ESR1 in chondrocytes (A; arrow) and in osteocytes (B; arrow). Sections from cartilage and bone showed immunostaining for AR in few osteocytes (D; arrow), but not in chondrocytes (C). The insert did not show immunostaining for ESR1 and AR when the section was incubated with secondary antibody only. Images shown are representative of the experimental results obtained from three horses (100 μm scale bar).
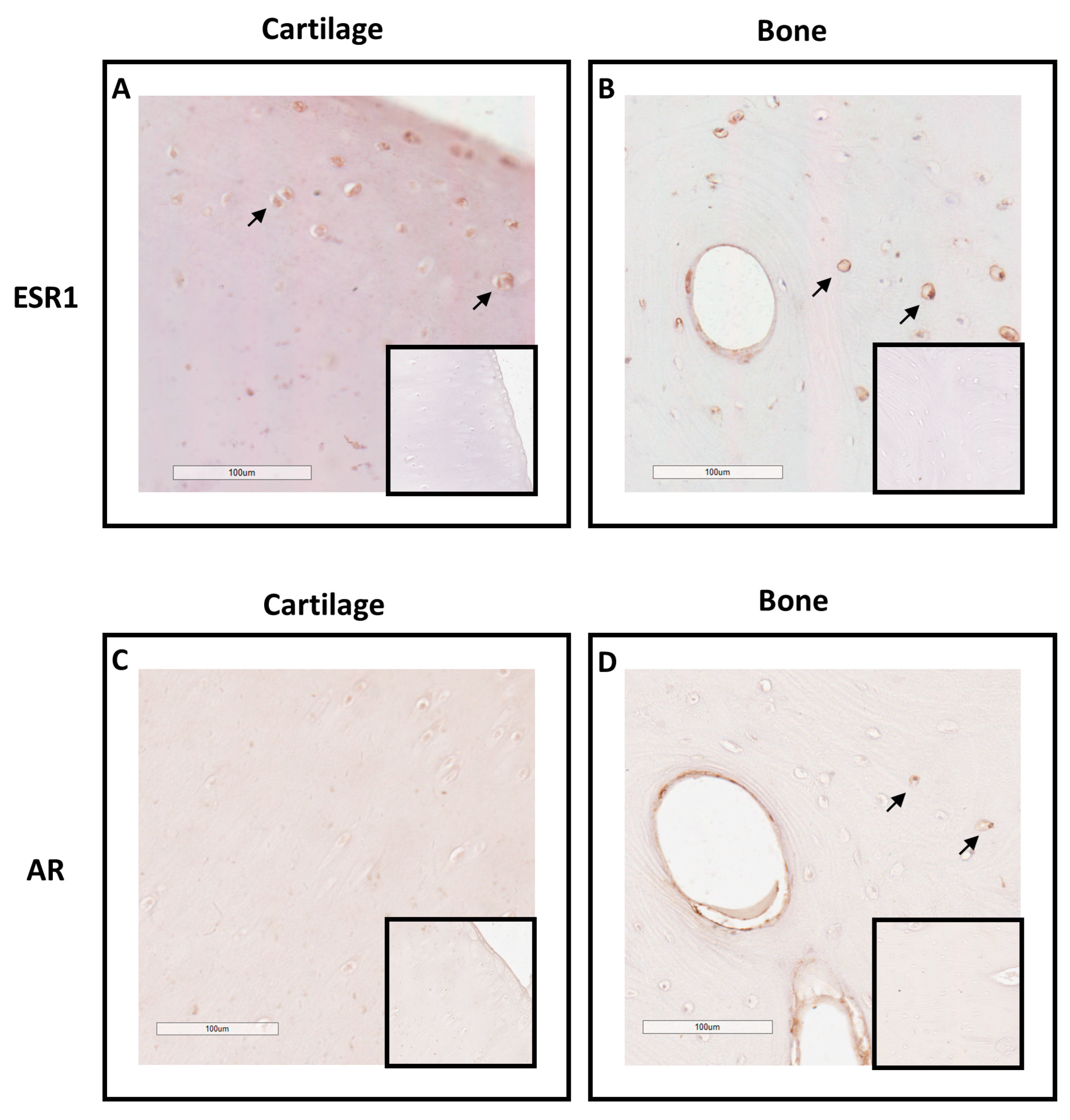
Figure 2.
Circulating levels of testosterone in plasma samples from horses castrated at birth and at 18 months. Plasma testosterone levels were measured for 33 months after birth from the two experimental groups by ELISA. Data were presented as mean ± SEM. A variation analysis in the testosterone levels compared to P0 within the same group was performed with a Kruskal-Wallis test followed by a Dunn’s comparison (a ≠ b; c ≠ d). A Mann-Witney test was performed to compare the testosterone levels at the same time between the two groups (* p ≤ 0.05; ** p ≤ 0.01; *** p ≤ 0.001).
Figure 2.
Circulating levels of testosterone in plasma samples from horses castrated at birth and at 18 months. Plasma testosterone levels were measured for 33 months after birth from the two experimental groups by ELISA. Data were presented as mean ± SEM. A variation analysis in the testosterone levels compared to P0 within the same group was performed with a Kruskal-Wallis test followed by a Dunn’s comparison (a ≠ b; c ≠ d). A Mann-Witney test was performed to compare the testosterone levels at the same time between the two groups (* p ≤ 0.05; ** p ≤ 0.01; *** p ≤ 0.001).
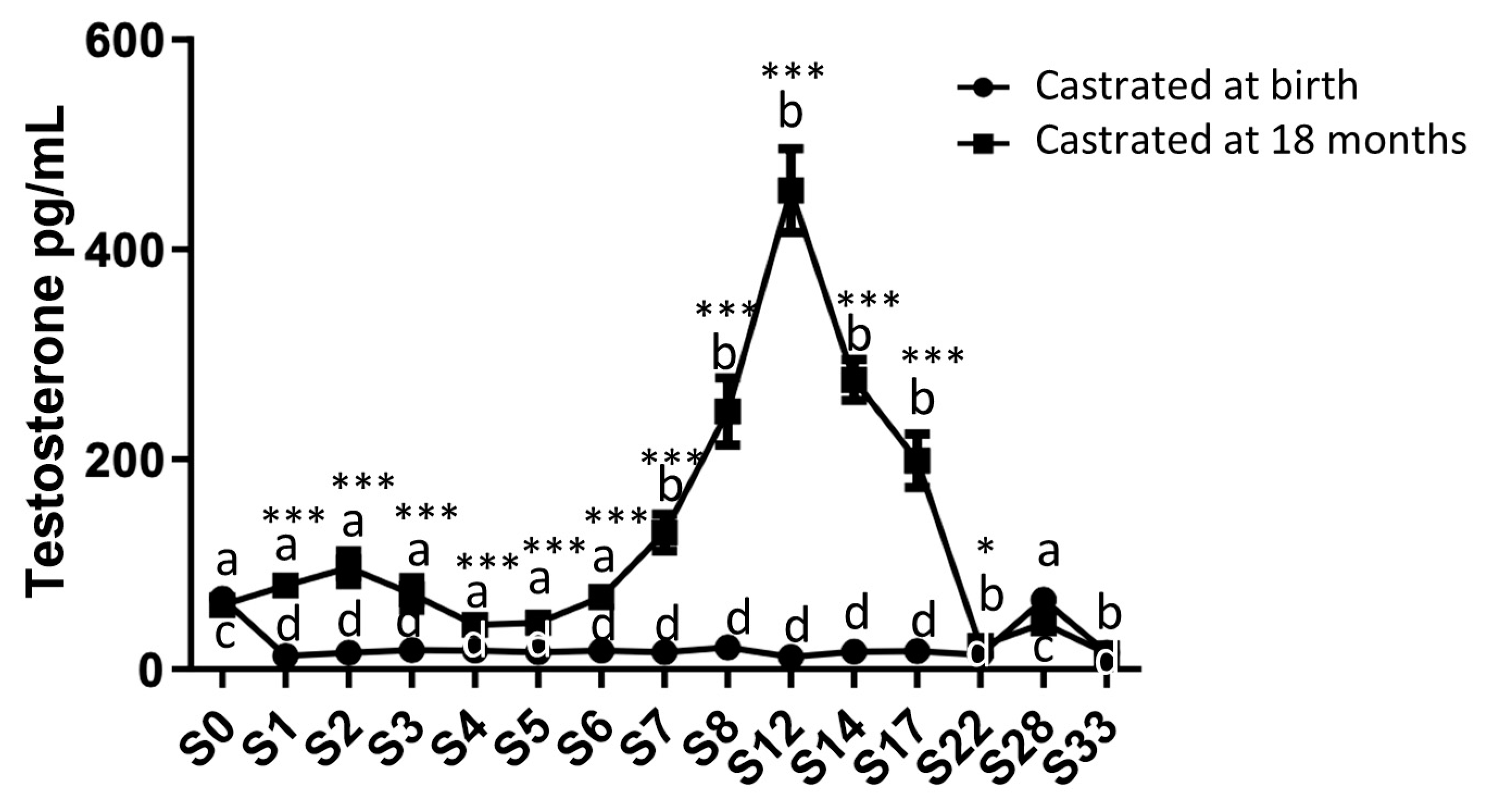
Figure 3.
Immunodetection of CYP17A1 protein in neonatal (A) and adult (B) testes. A positive CYP17A1 signal was observed in the neonatal testicular interstitial tissue with 10X magnification (A). Adult testis was used as a positive control (B). The insert did not show immunostaining for CYP17A1 when the section was incubated with secondary antibody only. Images shown are representative of the experimental results obtained from five horses (100 μm scale bar).
Figure 3.
Immunodetection of CYP17A1 protein in neonatal (A) and adult (B) testes. A positive CYP17A1 signal was observed in the neonatal testicular interstitial tissue with 10X magnification (A). Adult testis was used as a positive control (B). The insert did not show immunostaining for CYP17A1 when the section was incubated with secondary antibody only. Images shown are representative of the experimental results obtained from five horses (100 μm scale bar).
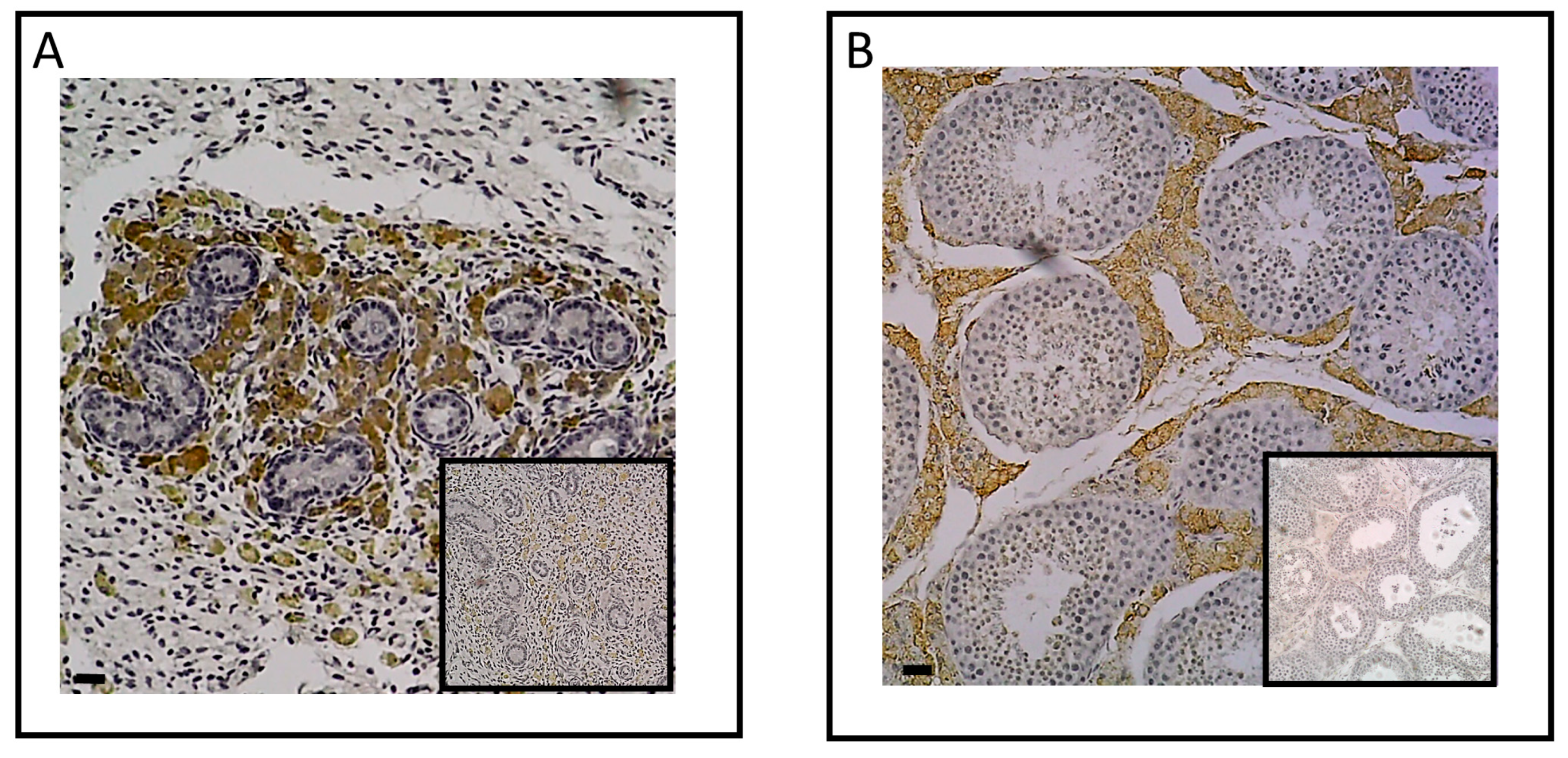
Figure 4.
Circulating levels of estradiol in plasma samples from horses castrated at birth and at 18 months. Plasma estradiol levels were measured during 33 months after birth from the two experimental groups by ELISA. Data were presented as mean ± SEM. A variation analysis in the estradiol levels compared to P0 within the same group was performed with a Kruskal-Wallis test followed by a Dunn’s comparison (a ≠ b; c ≠ d). A Mann-Witney test was performed to compare the estradiol levels at the same time between the two groups (* p ≤ 0.05; ** p ≤ 0.01; *** p ≤ 0.001).
Figure 4.
Circulating levels of estradiol in plasma samples from horses castrated at birth and at 18 months. Plasma estradiol levels were measured during 33 months after birth from the two experimental groups by ELISA. Data were presented as mean ± SEM. A variation analysis in the estradiol levels compared to P0 within the same group was performed with a Kruskal-Wallis test followed by a Dunn’s comparison (a ≠ b; c ≠ d). A Mann-Witney test was performed to compare the estradiol levels at the same time between the two groups (* p ≤ 0.05; ** p ≤ 0.01; *** p ≤ 0.001).
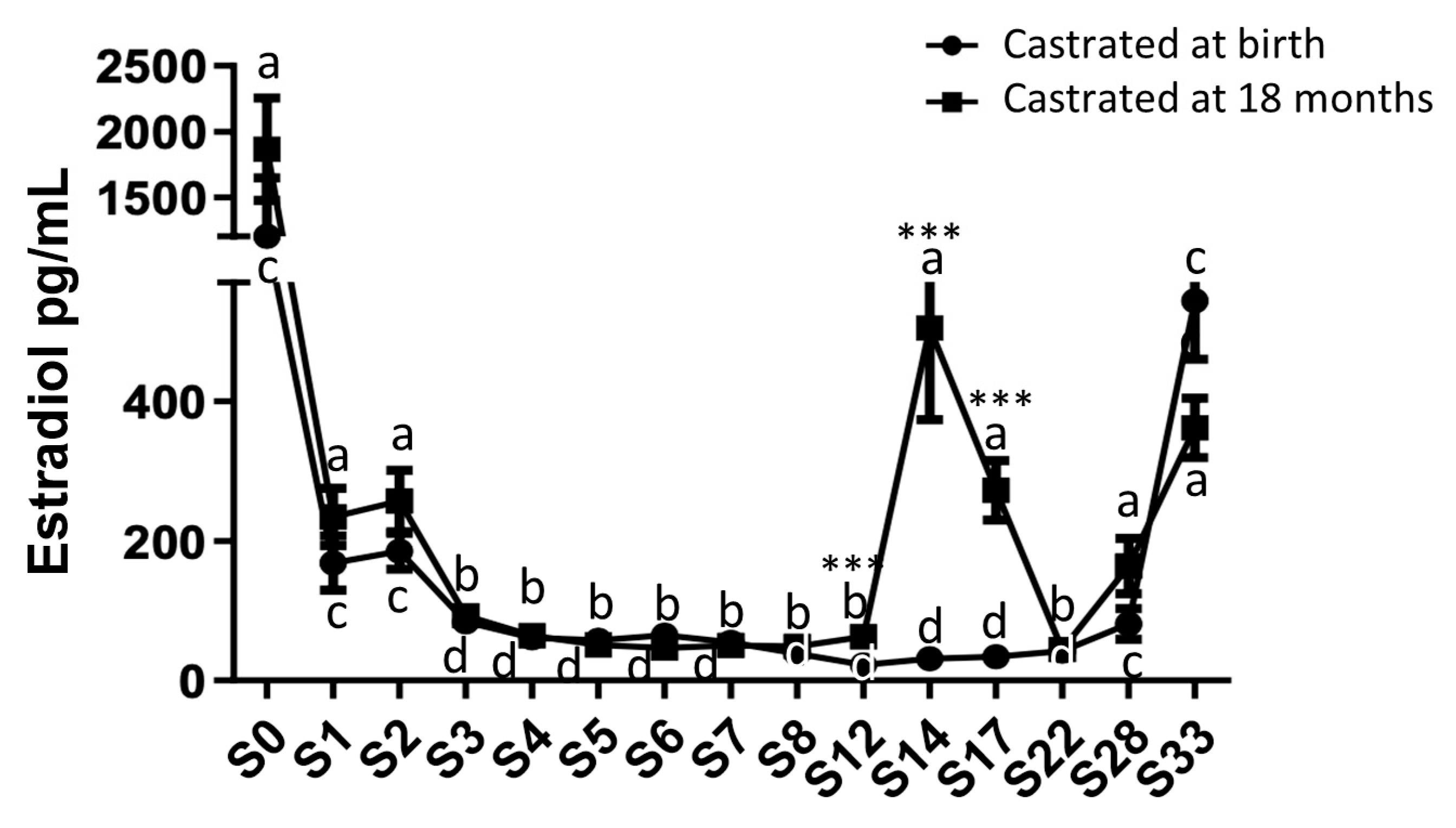
Figure 5.
Immunodetection of aromatase in neonatal (A) and adult (B) testes. No positive aromatase signal was observed in the neonatal testicular interstitial tissue with 10X magnification (A). Adult testis was used as a positive control (B). The insert did not show immunostaining for aromatase when the section was incubated with secondary antibody only. Images shown are representative of the experimental results obtained from five horses (100 μm scale bar).
Figure 5.
Immunodetection of aromatase in neonatal (A) and adult (B) testes. No positive aromatase signal was observed in the neonatal testicular interstitial tissue with 10X magnification (A). Adult testis was used as a positive control (B). The insert did not show immunostaining for aromatase when the section was incubated with secondary antibody only. Images shown are representative of the experimental results obtained from five horses (100 μm scale bar).
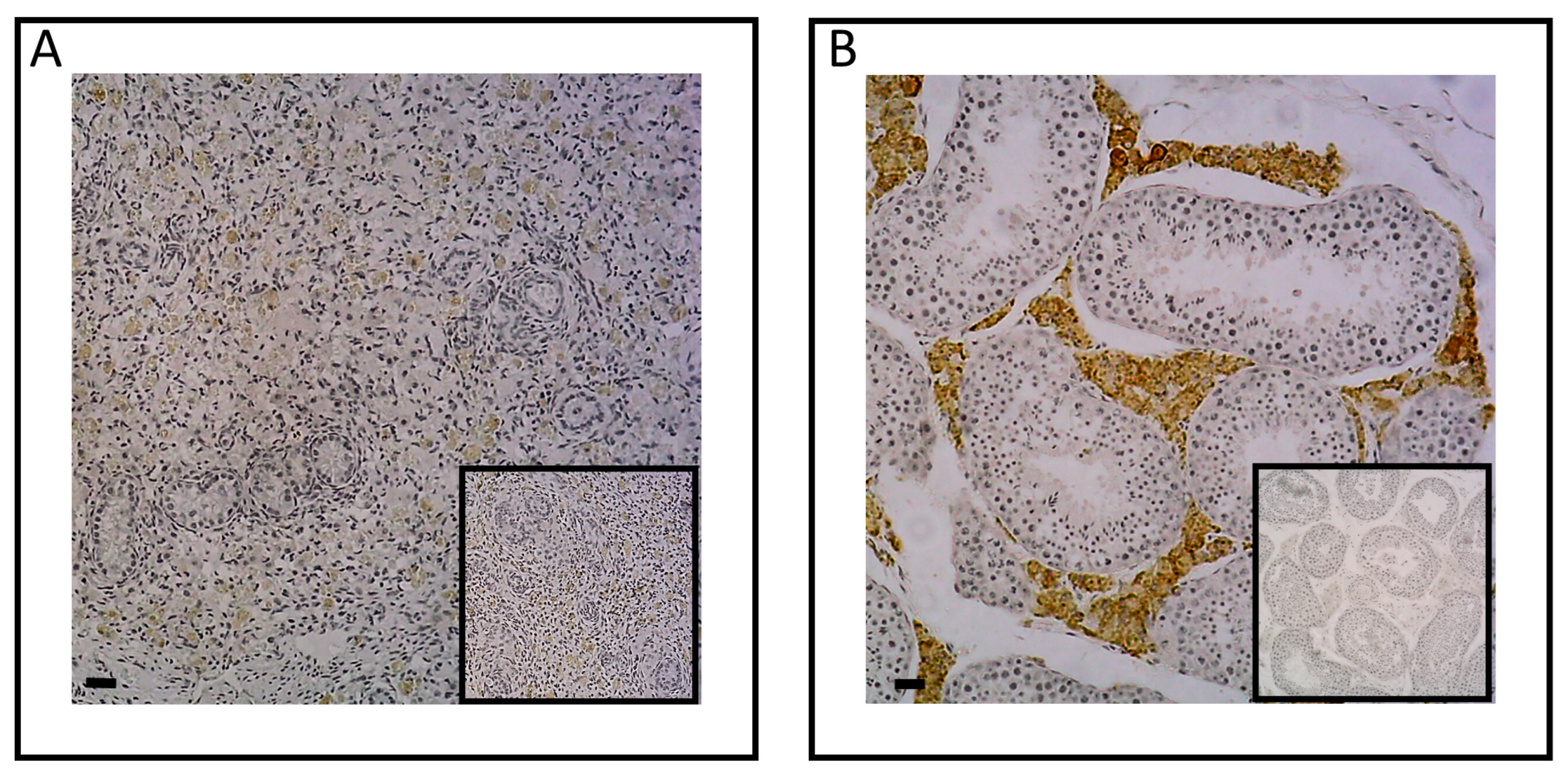
Figure 6.
Circulating levels of biomarkers of bone metabolism in blood samples from horses castrated at birth and at 18 months. Plasma N-MID levels (A) as well as serum CTX-I levels (B) were measured for 33 months after birth from the two experimental groups by ELISA. Data were presented as mean ± SEM. A variation analysis in the biomarker levels compared to P0 within the same group was performed with a Kruskal-Wallis test followed by a Dunn’s comparison (a ≠ b; c ≠ d). A Mann-Witney test was performed to compare the biomarkers levels at the same time between the two groups: no difference was observed.
Figure 6.
Circulating levels of biomarkers of bone metabolism in blood samples from horses castrated at birth and at 18 months. Plasma N-MID levels (A) as well as serum CTX-I levels (B) were measured for 33 months after birth from the two experimental groups by ELISA. Data were presented as mean ± SEM. A variation analysis in the biomarker levels compared to P0 within the same group was performed with a Kruskal-Wallis test followed by a Dunn’s comparison (a ≠ b; c ≠ d). A Mann-Witney test was performed to compare the biomarkers levels at the same time between the two groups: no difference was observed.
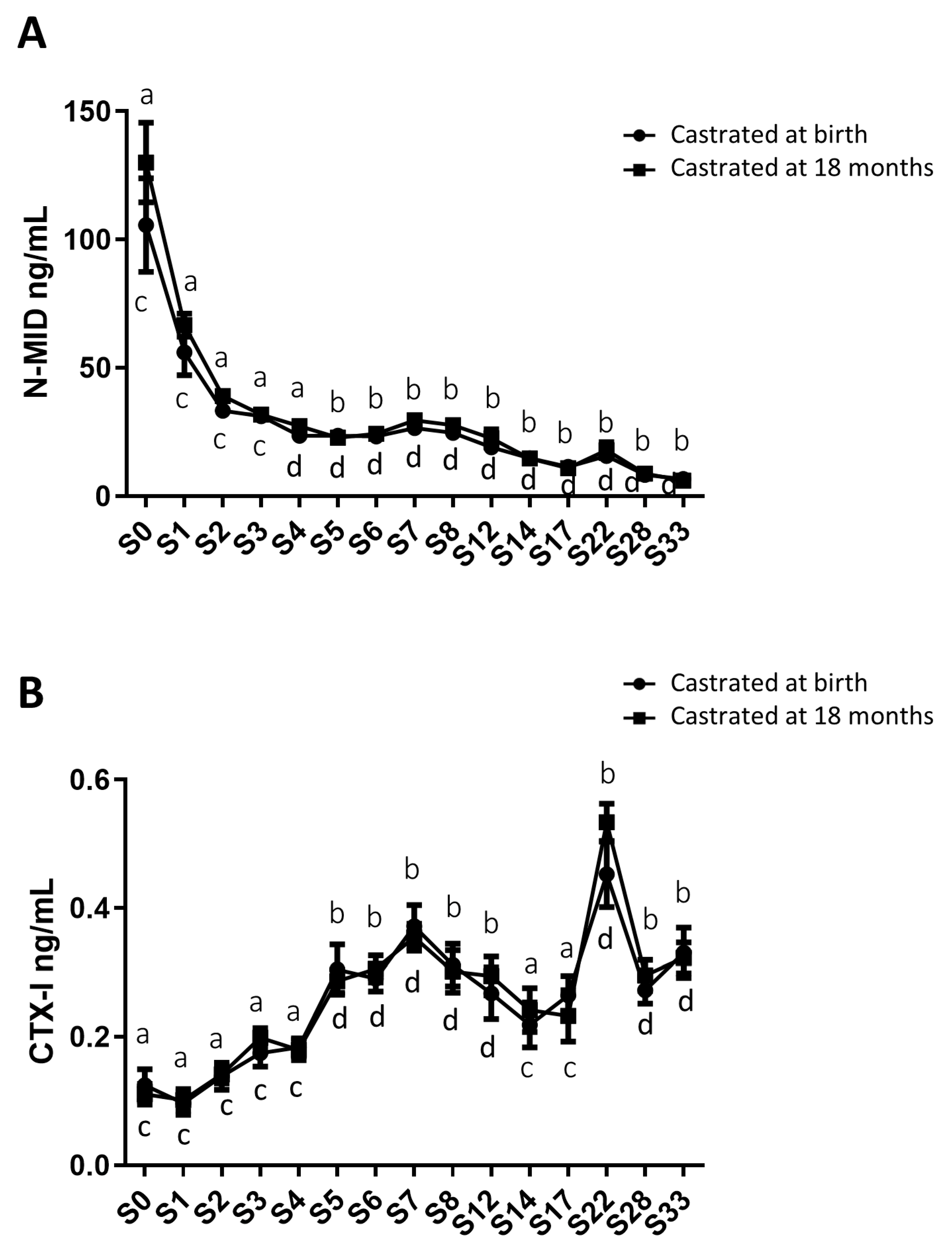
Figure 7.
Circulating levels of cartilage and osteoarthritis biomarkers in serum samples from horses castrated at birth and at 18 months. Serum CPII (A), CTX-II (B), COMP (C) and HA (D) levels were measured for 33 months after birth from the two experimental groups by ELISA. Data were presented as mean ± SEM. A variation analysis in the biomarker levels compared to P0 within the same group was performed with a Kruskal-Wallis test followed by a Dunn’s comparison (a ≠ b; c ≠ d). A Mann-Witney test was performed to compare the biomarkers levels at the same time between the two groups: no difference was observed.
Figure 7.
Circulating levels of cartilage and osteoarthritis biomarkers in serum samples from horses castrated at birth and at 18 months. Serum CPII (A), CTX-II (B), COMP (C) and HA (D) levels were measured for 33 months after birth from the two experimental groups by ELISA. Data were presented as mean ± SEM. A variation analysis in the biomarker levels compared to P0 within the same group was performed with a Kruskal-Wallis test followed by a Dunn’s comparison (a ≠ b; c ≠ d). A Mann-Witney test was performed to compare the biomarkers levels at the same time between the two groups: no difference was observed.
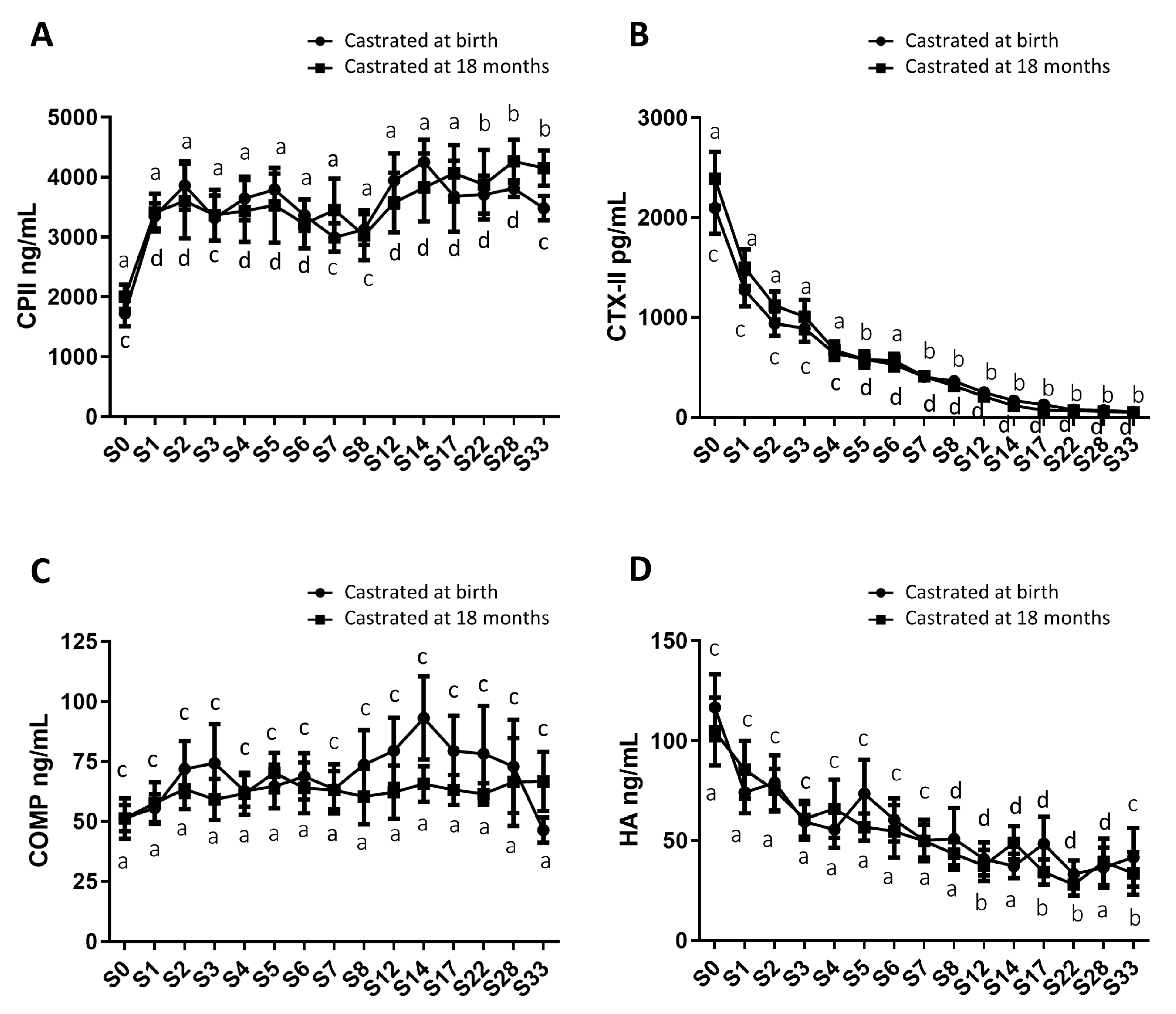
Figure 8.
Early castration has no short- and medium-term effects on bone and cartilage growth in foals.
Figure 8.
Early castration has no short- and medium-term effects on bone and cartilage growth in foals.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



