
Submitted:
29 September 2023
Posted:
29 September 2023
You are already at the latest version
Abstract
This chapter explores the pivotal role of maize (Zea mays L.) in global nutrition, emphasizing its status as a staple food worldwide. It discusses maize's complex nutritional profile, including macronutrients, micronutrients, and dietary fiber. The chapter also covers biofortification efforts to enhance maize with essential nutrients, addresses nutritional concerns related to maize-centric diets, and explores genetic modification and agronomic strategies to boost nutrient-rich maize yields. Additionally, it delves into maize processing traditions, nutrient retention techniques, and the importance of promoting nutrient-rich maize varieties. The chapter outlines post-harvest strategies to combat nutrient loss and prevent contamination while emphasizing the significance of consumer education and diverse diets. Case studies illustrate the impact of improved maize nutrition and provide replicable strategies. The chapter concludes by considering emerging research and technology, identifying challenges, and proposing solutions to enhance maize's nutritional value in the future.
Keywords:
maize
; global nutrition
; macronutrients
; biofortification
; genetic modification
; maize processing
1. Introduction: Maize's Crucial Role in Global Nutrition
Globally, maize (Zea mays) is one of the most widely grown cereal crops. Maize, a high-yielding crop, easy to process, easily digestible, and cost-effective compared with other cereals. It is used not only for human consumption but also as the primary feed for livestock, ethanol production, biogas, and raw material for producing bioplastics, industrial products (e.g., corn syrup, cornstarch). The green fodder of this crop is widely acceptable by dairy farmers because of its higher digestibility than sorghum, bajra, and other non-leguminous forage crops [1]. In terms of acreage and production, it ranks second. A well-aerated soil with a neutral pH and loamy sand or heavy clay is suitable for growing corn [2].
Maize is characterized by its tall, grassy appearance, with leaves arranged spirally along the stem. The seeds, commonly known as kernels, develop within the protective husk and exhibit considerable variability in colour and size, depending on the maize variety. Maize belongs to the Poaceae family, making it a member of the grass family.
Maize is a wind-pollinated (anemophily) plant that relies on cross-pollination (allogamy) to reproduce. It exhibits protandry to minimize self-pollination by ensuring that the male flowers release pollen before the female flowers become receptive. Additionally, maize (Zea mays, 2n = 2x = 20) is a monoecious (inflorescence) plant, where both male and female reproductive structures present on the same individual plant. The male flowers are tassel found at the top of the plant, while the female flowers are ear found at lower on the plant. These characteristics are important considerations in maize cultivation and breeding to ensure optimal pollination and seed production.
The study of maize's origin, evolution, and distribution involves genetic, archaeological, and historical research. Genomic analyses have provided valuable insights into the genetic changes associated with maize domestication and its evolutionary history. Maize, scientifically known as Zea mays L. was domesticated from its wild ancestor, teosinte, around 9,000 to 10,000 years ago in the region that is now southern Mexico. Teosinte (Zea mays ssp. parviglumis) is a grass species that bears little resemblance to modern maize, with small, sparse kernels and a different plant architecture. Maize's evolution is marked by a complex history of genetic changes driven by human selection and natural processes. Over millennia, humans played a pivotal role in shaping maize through selective breeding through hybridization, favouring desirable traits such as larger kernels, greater yield, and improved adaptability to different environments leading to the development of maize as we know it today. Maize is a staple food in many regions, particularly in North and South America, Africa, and parts of Asia. The expansion of maize cultivation can be traced back to European exploration and colonization, which facilitated its introduction to new continents.
Maize goes by several common names in different regions and languages. In North America, it is commonly referred to as ‘corn’ In many English-speaking countries, it's known simply as ‘maize,’ while in Spanish-speaking regions, it's called ‘maíz.’ Across Africa, it often goes by local names, such as ‘mealies’ in South Africa and ‘corn’ in many East African countries. In India and various parts of Asia, maize is commonly known by a variety of regional names. For instances, it is often referred to as ‘makka’ or ‘makki’ in Hindi and other northern languages, ‘bhutta’ in Punjabi, while in southern India, like in Tamil it can be called ‘cholam’ or ‘makka cholam’, ‘makkalu’ in Kannada, and ‘makkalu’ in Telugu, reflecting the linguistic and cultural diversity of the region.
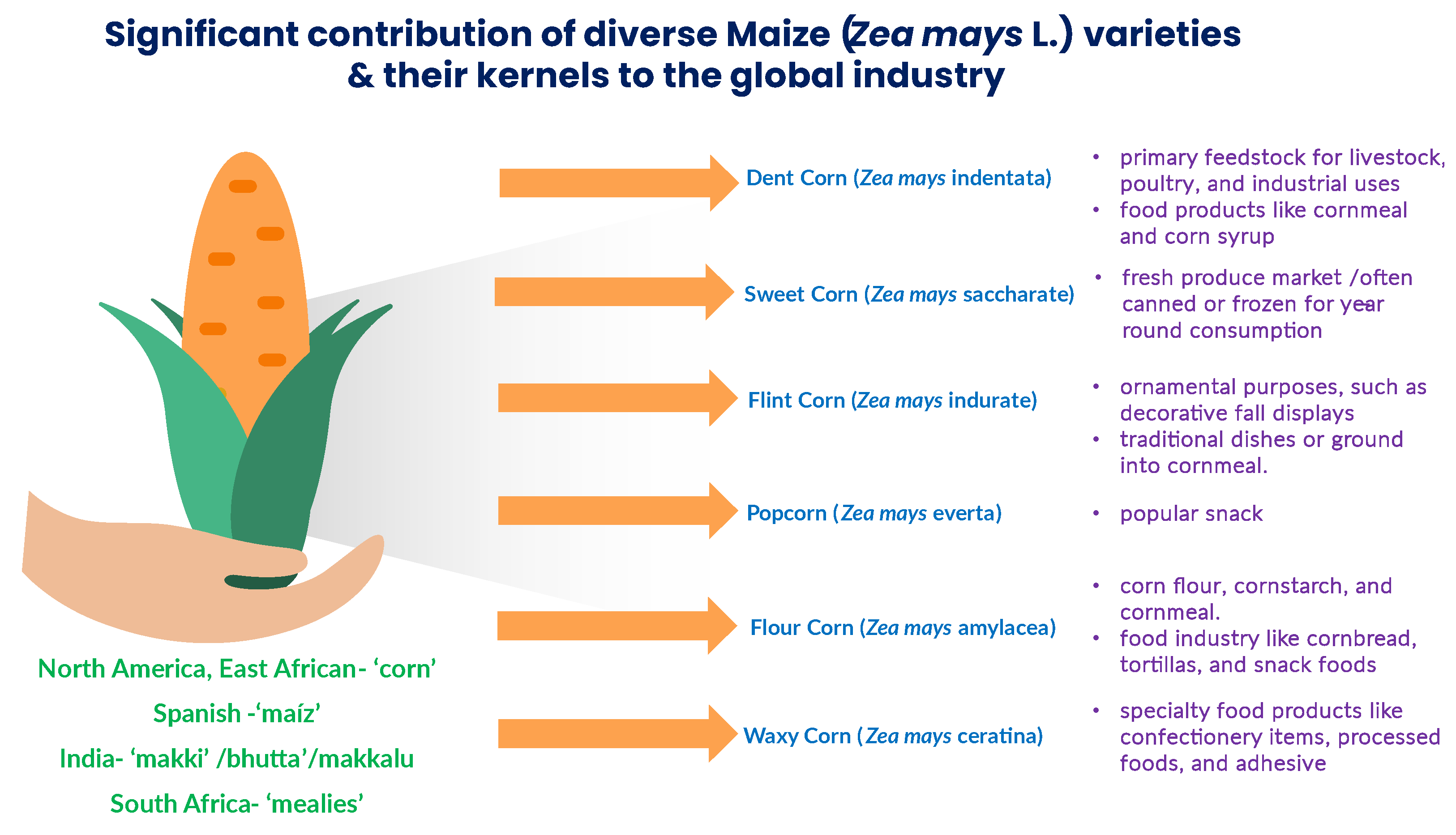
The diversity of maize varieties and their kernels contributes significantly to the global agricultural and food industry. Each of these kernel types has distinct characteristics that lend themselves to various commercial applications, from food and beverages to industrial uses.
Dent Corn (Zea mays indentata):
Kernel Type: Dent corn, also known as field corn, is characterized by a depression or ‘dent’ at the crown of the kernel when it dries. It has hard endosperm and a soft, starchy interior.
Commercial Significance: Dent corn is the most widely cultivated type of maize globally and serves as a primary feedstock for livestock, poultry, and industrial uses. It is also processed into various food products like cornmeal and corn syrup.
Sweet Corn (Zea mays saccharata or rugosa):
Kernel Type: Sweet corn kernels are characterized by their high sugar content and tenderness. They lack the hard endosperm found in dent corn.
Commercial Significance: Sweet corn is popular as a fresh vegetable, consumed on or off the cob. It is a staple in the fresh produce market and is often canned or frozen for year-round consumption.
Flint Corn (Zea mays indurata):
Kernel Type: Flint corn, also known as Indian corn, has hard, vitreous (glassy) endosperm and distinctively colored kernels, often with a hard outer layer.
Commercial Significance: Flint corn is primarily used for ornamental purposes, such as decorative fall displays. In some regions, it may be used for traditional dishes or ground into cornmeal.
Popcorn (Zea mays everta):
Kernel Type: Popcorn has a hard, moisture-sealed hull and a starchy interior. The kernels explode when heated, forming a fluffy, edible product.
Commercial Significance: Popcorn is a popular snack and is commercially grown for its unique popping characteristics. It is enjoyed worldwide in various flavours and forms.
Flour Corn (Zea mays amylacea):
Kernel Type: Flour corn has a soft, starchy endosperm and is typically ground into fine flour, making it suitable for baking and milling.
Commercial Significance: Flour corn is used in the production of corn flour, cornstarch, and cornmeal. It is essential in the food industry for products like cornbread, tortillas, and snack foods.
Waxy Corn (Zea mays ceratina):
Kernel Type: Waxy corn has a high amylopectin content, resulting in a waxy, sticky texture when cooked. It lacks amylose, which is present in regular corn.
Commercial Significance: Waxy corn is used in the production of specialty food products like confectionery items, processed foods, and adhesives due to its unique starch properties.
Europeans began using maize for bread in the 16th century, and it became one of the main grains in South-Eastern Europe by the 19th century. Maize is used in various Spanish food cuisines such as ''talo'' and ''gofio''. Maize flour is used in Italy to prepare the traditional dish ''polenta'' [3]. ''Proja'', an indigenous unfermented bread prepared from pure maize flour, is popular in the Balkan region. Several culinary technical advances have been applied to traditional maize bread, resulting in a variety of unique maize breads such legume-fortified maize loaf, and gluten-free maize toast popular among consumers providing new potential for the bakery business [3].
Figure 1.
Representation of significant contribution of diverse maize varieties and their kernels in the global industry.
Figure 1.
Representation of significant contribution of diverse maize varieties and their kernels in the global industry.
This chapter aims to cater readers a comprehensive overview of maize's nutritional profile, its challenges, and its prospects. It also explores its implications for global health and nutrients. It covers a huge range of topics below numerous subheadings, each contributing to a holistic understanding of maize's role in worldwide nutrients. Throughout this chapter, readers will find more in-depth discussions of topics like micronutrient deficiencies due to maize consumption, biofortification strategies, genetic amendment in maize, agronomic strategies to optimize nutrient availability and techniques for retaining nutrient integrity after harvest. Finally, the chapter concludes by emphasizing the requirement for collective action in enhancing maize's nutritional potential on an international scale. This is done through successful case studies and cutting-edge emerging tools and techniques.
2. Maize's Nutritional Profile: Beyond Basics
2.1. Unveiling the intricate nutrient composition of Maize
Maize kernel is an ingestible and nutritious component. According to [4], the maize kernel contains 71.88g of carbohydrates, 8.84g of protein, 4.57g of fat, 2.15g of fiber, 348mg of phosphorus, 15.9mg of sodium, 114mg of sulphur and 1.78mg of amino acids.
2.2. Implications of macronutrient and micronutrient content
The edible kernels or caryopses of the maize plant account for around half or 42% of its dry weight (DW). Starch, which makes up 72% of the dry weight of maize, is the main carbohydrate in the kernel. The sugars contain sucrose in the range of 1-3%, with minor amounts of maltose, glucose, fructose, and raffinose. Sweet corn has a lower (28%) starch percentage and a greater (18%) sugar content [5]. Amylose and amylopectin, two glucose polymers, make up these complex carbohydrates. According to Nuss and Tanumihardjo [6], amylose ranges from 0% to 73% in various maize genotypes. Recently, waxy maize types have been shown to be an effective source of carbohydrate for preserving blood glucose control [7].
The maize kernels or caryopses account for around 10% of the protein distributed mostly between the endosperm and germ. Prolamins, glutelins, albumins, and globulins make up most crude maize protein, which is frequently calculated based on the nitrogen content. These proteins are distinguished by their solubility characteristics. About 0.5 part of total nitrogen in kernel is provided by prolamin, which is followed by glutelins, which make up 35% of the protein and are both exclusive to the endosperm. Approximately 7% of the nitrogen in the kernel comes from albumins, 5% from globulins, and 6% from nonproteins [8]. About 40% of the proteins from maize have a high protein efficiency ratio (PER). Lysine and tryptophan concentrations in the kernel control PER [9]. Lysine, the most limited amino acid in relation to other amino acids, is present in maize at a rate of about 1%.
Each protein fraction has its own specific amino acid profile. Prolamin, which contributes 50% of the kernel's nitrogen, is a powerful indicator of the total protein quality of maize [9]. After starch and protein, Fat in the oil form, which makes up 3.5-6% of the total weight of the kernel with an average of 4.5%, is the third greatest nutritious component of the kernel. The germ comprises around 45–50% of the oil produced during the wet milling process, which accounts for nearly 85% of the total kernel oil [10]. Germ growth might lead to an increase in oil content. Maize germ oil contains 56% polyunsaturated fatty acids, 30% monounsaturated and 14% saturated. Linoleic acid accounts for 54-60%, oleic acid for 25-31%, stearic acid for 2-3%, palmitic acid for 11-13%, and linolenic acid for 1% of the refined maize oil [11]. The primary storage form of kernel lipid is triacylglycerol, which is glycerol with three fatty acids attached. In addition, phospholipids (1.5%), sterols (1%) and waxes might be included in lipid composition. N-ferrulyl tryptamine and N-p-coumaryl tryptamine are also present, along with vitamin C (0.12 mg), E, K, B1 (0.42 mg), B2, B3, B5, B6; folic acid, and selenium [4].
Water-soluble vitamins are available in the germ and endosperm of maize, with the aleurone layer often containing the highest quantities. Niacin, for instance, is predominantly located in the kernel endosperm, constituting about 80% of its content, followed by seed coat (~4%) and germ (~2%). It's worth noting that unless maize is adequately processed, the niacin it contains may not be biologically accessible to humans, potentially leading to pellagra [12]. Moreover, the levels of thiamin, riboflavin, and niacin can vary among different maize cultivars.
Vitamin E is predominantly located in maize germ oil, constituting approximately 94% of the total tocopherol content. Alpha (α) and gamma (γ) tocopherols are the 2 major types of vitamin E [8]. Maize oil stands out as a rich source of these tocopherols, notably featuring high concentrations of γ-tocopherol, with 21.3 mg/100g of α and 94.1 mg/100g of γ tocopherols [13]. The abundance of tocopherols is absolutely associated with the refined corn oil content boasts a substantial vitamin E concentration of 14.8 mg per 100 g [6].
Sweet corn cultivars vary from conventional maize in provisions of their carotenoid and tocopherol profiles in addition to their distinctive sweetness. Zeaxanthin (0-7.7 g/g DW) and lutein (0-27.6 g/g DW) are the main carotenoids discovered. Genomes of sweet corn, which control the carotenoids and tocopherols synthesis, exhibit significant variation, which suggests that these features are potential target for biofortification efforts targeted at boosting antioxidant levels [14]. γ -tocopherol stands out as the main tocopherol [15].
Plant sterols, commonly referred to as phytosterols, are crucial elements of the cell walls and membranes of plants. Stigmasterol (1.52 mg/100g), Sitosterol (9.91 mg/100g) and Campesterol (3.40 mg/100g) are three phytosterols that are frequently found in maize oil. Their distribution varies among the endosperm, pericarp, and germ sections of maize kernel [16].
2.3. Exploring the value of dietary fiber in Maize
Crude fiber in maize exhibits distinctive characteristics, primarily concentrated within the seed coat, constituting approximately 87% of the seed coat's composition. However, smaller quantities of fiber are also present in the walls of the endosperm and germ. The fiber within the seed coat comprises approximately 67% hemicellulose, 23% cellulose, and a minor 0.1% lignin content. In terms of overall kernel composition, fiber accounts for roughly 7% [5], with predictable proportions of soluble and insoluble fiber. Common maize types typically have less than 2% soluble fiber and roughly 12% insoluble fiber. It's important to keep in mind that processed, dehulled kernels have much less fiber than whole-grain alternatives since fiber mostly gathers in the seed coat, also known as the pericarp.
3. Maize's Nutritional Profile: Beyond Basics
Maize, commonly referred to as corn, ranks among the world's most extensively consumed cereal grains. For centuries, maize has served as a staple food for numerous civilizations, maintaining its vital role in global nutrition [17,18]. Nevertheless, despite the numerous advantages maize provides, it comes with nutritional considerations that individuals and communities must address [19]. In this chapter we will delve into both the benefits and possible obstacles when including maize in our diet. We will also offer advice on maximizing the potential of this versatile grain while maintaining optimal nutrition.
Maize is a potent source of carbohydrates, providing abundant energy for active individuals [17]. Whole-grain maize products like cornmeal and cornbread are rich in dietary fiber, aiding digestion, weight management, and reducing digestive issues [20]. Maize also contains essential vitamins (including C, B vitamins like thiamine and niacin), magnesium, and potassium, vital for overall health [21,22]. Moreover, maize contains essential antioxidants like zeaxanthin and lutein that contribute to maintaining optimal eye health and serve as protective factors against age-related macular degeneration [23]. Other important nutritional content of the consumable part of maize in its dry form Gopalan et al. [24] in Table 1.
Nevertheless, there is growing concern among nutrition advocates regarding corn, as it is often seen as a filler that may lead to excessive carbohydrate consumption at the expense of nutrient-rich foods. Gensberger and colleagues [25] noted that the use of high fructose corn syrup (HFCS), a product derived from corn starch, has generated considerable discussions about the inclusion of sweeteners by manufacturers in a wide range of food items. However, there is no conclusive evidence to suggest that HFCS is inherently more detrimental than alternative sugars [26,27]. Despite the widespread use of GMO corn by American farmers, concerns regarding its safety have been voiced by natural health advocates. Bawa and Anilakumar [28] point out that there is a scarcity of comprehensive data on the potential health effects associated with GMO crops. However, Norris [29] conducted a study that led to the conclusion that GMO crops, including corn, are safe for human consumption. Another notable concern revolves around corn's high cellulose content, an insoluble fiber that the human body cannot digest [30,31]. Nevertheless, it's worth noting that the body does effectively break down other constituents of corn. In summary, while concerns have been raised regarding various aspects of corn, existing research and studies suggest that these concerns may not pose significant health risks. The evidence indicates that corn, in its various forms, is generally safe for human consumption.
4. Biofortification: Empowering Maize with Essential Nutrients
4.1. Pioneering provitamin A enrichment in Maize
Biofortification refers to the process of enhancing the concentration of nutrients in crops, achieved by various methods such as plant breeding, transgenic approaches, and improved farming practices. It represents a viable and cost-efficient approach to provide essential micronutrients to populations with limited access to diverse diets. Biofortification successfully attained targets for micronutrients, specifically raising the provitamin A content to the requisite level of 15 ppm in maize [32]. The regular consumption of biofortified staple crops is expected to yield measurable enhancements in human health [32]. In the pursuit of addressing hidden hunger in developing nations, the introduction of biofortified plant foods aimed at enhancing vitamin A content in maize was initiated by Harvest Plus [8]. Provitamin A maize (PVAM) is a product resulting from the breeding of maize with elevated provitamin A content. Provitamin A generated from biofortified crops has been shown to be extremely bioavailable and has the potential to enhance the vitamin A level in mothers and children [33]. According to a study done in Zambia, children aged 5-7 showed a significant increase in their total body vitamin A reserves after three months of consuming biofortified orange maize [34]. Orange maize eating improved the body's overall vitamin A storage just as well as vitamin A supplements. Additionally, children who had a little vitamin A deficiency showed a notable improvement in their visual function [35]. The danger of hyper-vitaminosis A, which has been connected to the use of vitamin A supplements, may be reduced by biofortification, which is significant [34].
4.2. Unravelling zinc and iron biofortification efforts
Biofortification initiatives hold the potential to address malnutrition, and among the staple crops targeted for biofortification to combat ‘hidden hunger’, maize stands out [36]. Iron (Fe), calcium (Ca), copper (Cu), selenium (Se), magnesium (Mg), iodine (I) and zinc (Zn) are the seven minerals that are typically lacking in human diets [37]. Improved soil uptake, internal transit, accumulation in edible sections, and a decrease in the presence of anti-nutrients are the main objectives of strategies to enhance mineral content in plants. The mobility of iodine (as KIO3), selenium (as Na2SeO4), and zinc (as ZnSO4) in soils and plants is well recognized. Although iron (Fe) is less mobile in soil, direct uptake can be facilitated by foliar sprays (using FeSO4) and iron chelates (Fe-EDDHA, FeEDTA) [38]. Applications of phosphorus (P), nitrogen (N) and potassium (K) in nutrient-poor habitats can improve the transport of zinc (Zn), iron (Fe), and calcium (Ca) from the soil to plant.
The concentration of vital minerals in maize kernels has increased significantly by transgenic technologies. Even though soils contain a lot of Fe (III), many plants have trouble utilizing it without changing it to Fe (II). By overexpressing the gene for nicotianamine aminotransferases known as Yellow Stripe 1 (Ys1), maize has improved Fe accumulation. By creating stable FePS complexes for root uptake, these enzymes aid in the production of phytosiderophores (PS) [39]. Zn and Cu absorption may also involve Ys1 [40]. In combination with enhanced Fe-binding ferritin, overexpression of the Aspergillus phytase (PhyA) gene under the control of a glutelin-1 (Gt-1) promoter unique to the endosperm increases total Fe levels and Fe bioavailability in maize [41]. In comparison to non-transgenic controls, the kernel Fe content increased by twofold to over 40 g/g, while the reduced phytate levels resulted in a threefold increase in Fe bioavailability. Elevating kernel ferritin, the protein that stores iron, can defend against metal chelators and resist denaturation during digestion, resulting in Fe absorption rates that are comparable to those of supplements containing highly bioavailable FeSO4 [42,43].
According to Brown et al. [44], the recommended minimum aim for zinc biofortification in maize is 38 g/g, which would provide about 40% of the physiological needs of non-pregnant women and non-breastfed children who consume a lot of maize. Although the use of zinc-containing fertilizers has successfully improved zinc concentration in wheat and rice, the application of biotechnology to enhance zinc content in maize is still under examining stage. There is little concrete evidence on maize. Zinc-biofortified maize is especially important since populations that depend on grains frequently suffer from zinc insufficiency. The minimum zinc biofortification target levels are anticipated to be attained soon using conventional breeding techniques [45]. The use of biofortification techniques for Cu, Ca, and Mg in maize has received scant attention in the literature. When fortification alternatives are not available, encouraging whole-grain options is advised because most nutrients in maize naturally collect in the aleurone layer and kernel germ. The most effective strategy for increasing vitamin and mineral consumption in people with nutritional deficits continues to be a combination of external and internal fortification approaches.
4.3. Revolutionary breeding techniques for nutrient elevation
During the process of maize domestication, a range of physiological and nutritional traits have been deliberately chosen for improvement. According to the analysis of nucleotide diversity conducted by [46], six genes associated with metabolism of starch (ae1, sh1, sh2, su1, bt2 and wx1) were identified as specific targets during the domestication of maize. Palaisa et al. [47] documented the selection of the Y1 gene, which encodes phytoene synthase, with the aim of enhancing nutritional value. In another study, Karn et al. [48] detected the presence of two, three, and six Quantitative Trait Loci (QTLs) related to starch, oil, and protein, respectively. This study showed that teosinte alleles could be used to improve attributes related to kernel composition in contemporary maize germplasm. Furthermore, in the Chinese waxy maize population, Fan et al. [49] revealed significant selection forces operating on the waxy gene (wx). The discovery of a teosinte-derived allele of teosinte high protein 9 (Thp9), which imparts improved protein levels and higher nitrogen utilization efficiency (NUE), was also revealed in an intriguing recent study. The asparagine synthetase 4 that Thp9 encodes was also discovered. In temperate maize varieties incorrect splicing of Thp9-B73 transcripts was identified as the cause of reduced expression, leading to diminished NUE and protein content [50].
5. Genetic Mastery: Transforming Maize's Nutritional Landscape
5.1. A profound exploration of genetic modification in Maize
Genetic modification in agriculture has primarily concentrated on enhancing desired traits in crops, leading to the creation of plants with increased yields, improved quality, and greater resistance to both pests and environmental stresses. This advancement in biotechnology has facilitated the integration of genetically modified (GM) crops into environmentally sustainable food production methods. As of 2019, soybeans held the largest share of GM crops at 48%, while GM maize covered a global area of 60.9 million hectares, representing approximately 32% of the world's maize production [51]. Biotechnology has enabled the transfer of genetic material not only within the same species but also between different species. This breakthrough led to the introduction of the first GM maize variety, known as Bt (Bacillus thuringiensis) maize, in 1997. Bt maize has significantly reduced the need for pesticides to combat corn-borer infestations, resulting in substantial annual savings for farmers. In 2000, it was estimated that 25% of the maize planted in the USA was genetically modified.
In recent decades, progress in plant biotechnology has enabled the creation of GM maize varieties, which had a substantial impact on agricultural practices and have globally enhanced grain yields. These genetically modified varieties have incorporated characteristics like resistance to herbicides, insects, and diseases, as well as tolerance to environmental stress, increased yield, and enhanced nutritional value. The process of maize transformation, a crucial step in the development of GM maize, is no longer a major obstacle. The use of morphogenic regulators in protocols has seen significant evolution to increase the frequency of transformation and reduce dependence on specific genotypes. Emerging technologies, encompassing both stable and transient expression methods, along with approaches that don't rely on tissue culture like the RNA-guided endonuclease method for direct genome editing, genome editing facilitated by haploid inducers, and the use of pollen transformation, as well as the simultaneous production of double haploids are anticipated to drive significant advancements in maize biotechnology.
5.2. Case studies showcasing nutrient-enriched transgenic Maize strains
Genetic modification of crops is essential for providing the diverse nutrients necessary for promoting human health. Kettenburg et al. [52] presented evidence demonstrating the health benefits derived from Bt maize crops. Moreover, research efforts are concentrated on improving the vitamin-A and provitamin-A (PrVit-A) content in corn [53]. Through the CRISPR/Cas9 system, waxy corn plants with improved amylopectin content have been developed by deactivating an endogenous waxy gene “Wx1” and introduced without regulatory restrictions [54]. Scientists have also engineered corn varieties with significantly increased levels of beta-carotene (169 times more), Vitamin C (six times more), and folate (twice as much) [55]. Various researchers have compared the nutritional profiles of transgenic corn varieties with commercially grown ones and have found no discernible differences. Consequently, transgenic varieties can be utilized for both human consumption and as feed (Table 2).
6. Cultivating Nutrient Excellence: Agronomic Strategies
Agronomic biofortification refers to a strategy aimed at enhancing the micronutrient levels in cereal crops through the application of mineral fertilizers, either at the base or through foliar methods [64]. This approach can enhance crops with various elements, although the most targeted ones include iron (Fe), zinc (Zn), selenium (Se) and iodine (I). Agronomic biofortification (shown in Table 3) has predominantly been applied to essential cereal crops such as rice, wheat, and maize, primarily because they constitute a significant portion of global diets, especially among populations susceptible to mineral nutrient deficiencies. Dimpka and Bindraban [65] advocate for micronutrient fertilization to not only increase nutrient content in crops but also improve overall crop yields. This perspective arises from the fact that fertilization programs in developing nations prioritize phosphorus (P), nitrogen (N) and potassium (K), and sometimes sulphur (S) fertilizers. However, crop productivity can be constrained by deficiencies of various soil micronutrients [66]. Consequently, it has been suggested that applying small quantities of multiple micronutrient elements to the soil as a basal treatment represents a sustainable approach to simultaneously enhance both crop yields and the nutritional quality of the harvested produce [67].
7. Preserving Nutrient Integrity: Maize Processing Traditions
Maize, a dietary staple consumed worldwide, holds a pivotal position in global nutrition [6]. Yet, to harness its complete nutritional value, it is imperative to grasp the traditional techniques for processing maize that safeguard its nutrient content.
7.1. Whole-Grain Maize
The processing of whole-grain maize stands as a vital component within traditional methods, ensuring the preservation of its nutritional quality [71,72]. These techniques play a crucial role in preserving minerals and amino acids while minimizing nutrient loss during the preparation process. Opting for whole-grain maize products, such as cornmeal or tortillas crafted from whole maize, guarantees a richer dietary fiber content and the preservation of essential nutrients found in the bran and germ. Research suggests that the retention of minerals in maize can be affected by different processing techniques. Techniques like hydrothermal treatment and nixtamalization have the potential to safeguard vital minerals in maize, enhancing their bioavailability in the product [20,73]. Traditional processing techniques can effectively lower the levels of phytic acid, a compound known to inhibit mineral absorption. This reduction significantly enhances the nutritional quality of maize-based foods, as noted by Gallego-Castillo et al. [73]. Moreover, it plays a crucial role in preserving the stability of vitamins during storage.
7.2. Nixtamalization
Nixtamalization is a traditional culinary method that involves soaking and cooking maize or other grains in an alkaline solution, usually limewater [74]. This approach finds extensive application across diverse cultures to enhance the nutritional quality, taste, and texture of dishes primarily based on maize [75]. In the nixtamalization process, the grain is immersed in the alkaline solution, resulting in the softening of the outer hull, facilitating its removal [76]. This process has several significant benefits, such as improving the nutritional quality of maize by increasing the availability of essential nutrients through nixtamalization. It enhances the availability of crucial amino acids and vitamins, such as niacin (vitamin B3) [77,6,72]. It also improves the maize's texture, making it better suited for a wide range of culinary uses, such as tortillas, tamales, and other traditional dishes [78,79]. Additionally, nixtamalization enhances the unique taste of these foods [80] and decreases the levels of specific anti-nutrients and toxins, such as mycotoxins, increasing the safety of maize for consumption [80].
7.3. Hydrothermal treatment
In the field of maize processing, hydrothermal treatment is a cooking method that relies on high-temperature hot water or steam. Its primary purpose is to prepare maize for further transformation into a variety of food products, as outlined by research conducted by Rocha-Villarreal et al. [81] and Akinbolade and Igbeka [82]. This comprehensive process involves heating, hydration, starch gelatinization, texture refinement, and the reduction of anti-nutrients. These stages collectively contribute to improving the digestibility, texture, and overall nutritional qualities of maize-based foods [83,81,82].
7.4. Elevating nutrient retention during culinary and industrial processing
Improving the retention of essential nutrients during the cooking and industrial processing of maize is crucial for enhancing the nutritional quality of maize-based foods. Numerous methodologies and approaches have been scrutinized to achieve this goal. For instance, Gallego-Castillo et al. [73] delved into the impact of various maize processing techniques on nutrient retention, identifying the most effective methods for safeguarding essential elements like minerals, amino acids, and other pivotal nutrients. Certain initiatives are dedicated to biofortification, a process that involves growing maize varieties with increased levels of vital nutrients [6,84]. This approach ensures that maize maintains heightened nutrient levels even after undergoing processing. Assessing the quality and consumer satisfaction of processed maize products is integral to evaluating the efficacy of nutrient preservation strategies, ultimately safeguarding the final product’s nutritional quality [85]. Moreover, it is crucial to emphasize the worldwide importance of maize as a staple crop. Raising awareness about maize's importance and promoting proper processing techniques can foster improved practices geared toward preserving essential nutrients [6]. Additionally, prioritizing nutrient management during maize cultivation is essential in guaranteeing that the raw material itself boasts high nutrient content before the processing phase commences [86]. In essence, enhancing nutrient preservation in maize processing encompasses a synergy of effective processing techniques, biofortification, quality evaluation, educational efforts, and nutrient management. This holistic approach aims to optimize the nutritional content of maize-derived food products.
8. Bridging Nutritional Gaps: The Role of Maize Varieties
8.1. Unearthing the spectrum of nutrient content variation within Maize strains
Maize germplasm represents a valuable reservoir of genetic diversity, making it a promising resource for identifying the most suitable materials to create nutritionally enhanced foods. Various studies have underscored the nutritional superiority of ancient varieties. In recent times, there has been a burgeoning interest in landraces, making the preservation and promotion of these genetic sources highly important. For instance, Langyan et al. [87] investigated the nutritional diversity of germplasm collected from Rajasthan, Himachal Pradesh, and Andhra Pradesh. Their analysis of the mineral profile revealed that most of the germplasm exhibited higher iron content. These varieties offer a valuable source of genetic diversity, which is not only significant from a scientific perspective for biodiversity preservation but also holds great potential for biofortification programs aimed at supporting small rural communities where these unique maize germplasms have evolved. The nutritional composition of food depends on the quantities of carbohydrates, proteins, fibres, oils, vitamins, and minerals it contains. Various colorimetric and chromatographic techniques are available for quantitatively analysing these nutrients. However, these methods are invasive, time-consuming, and labour-intensive, often necessitating advanced laboratory equipment. The emergence of near-infrared (NIR) analysers and Raman spectroscopy has enabled entirely non-invasive and non-destructive evaluation of nutrient content in agricultural crops [88,89]. Moreover, these techniques have the capability to determine the nutrient content of individual maize grains within the crop.
8.2. Championing nutrient-rich Maize cultivars for nutritional empowerment
Corn lacks specific essential amino acids, vitamins, and minerals. Maize varieties fortified with essential nutrients might have a significant positive impact in rural regions where individuals lack access to a wide range of diets, nutritional supplements, and enriched foods. Substantial strides have been taken in the development, evaluation, and dissemination of maize varieties enriched by quality protein maize (QPM), PrVit-A, and zinc-enriched kernels. QPM plays a pivotal role in upholding the nutritional welfare of communities heavily reliant on maize. Cereal grains are primarily comprised of carbohydrates (65-75%) and have a low protein content (7-12%), and their proteins lack essential amino acids like lysine (lys) and tryptophan (trp) [90]. QPM is recognized as nutritionally superior maize because it comprises elevated levels of lys and trp, as well as desired kernel characteristics when compared to normal maize varieties. Researchers have identified various mutants in maize, particularly opaque-2 (o2), which lead to higher lys and trp content [91]. Opaque-2 (o2) and floury-2 (fl-2) have improved lys and trp levels by a factor of two compared to standard genotypes.
India has a rich history of biofortified maize cultivar development. In 1970, India introduced three o2 hybrids: “Shakti”, “Rattan”, and “Protina”. Then, in 2000, it approved “Shaktiman-1” and “Shaktiman-2” QPM hybrids for cultivation by utilising CIMMYT's inbred lines [92]. In 2008, “Vivek QPM 9” was generated through Marker-Assisted Selection (MAS) and made available for marketable usage in India during the same year. The upgraded iteration of Vivek QPM 9, known as “Pusa Vivek QPM 9 Improved”, was officially introduced for widespread grown in India by the Central Variety Release Committee in 2017. This improved corn variety, rich in multiple nutrients and combining high-quality protein with PrVit-A, is a remarkable achievement of the Indian Agricultural Research Institute (IARI), New Delhi, making it the world's pioneering corn genotype of its kind. Additionally, China identified an important mutant known as “opaque16” (o16). In 2018, India introduced four QPM hybrid cultivars, namely HQPM-1, HQPM-4, HQPM-5, and HQPM-7, by incorporating the o16 allele into the parental lines of these hybrids using MAS [93]. The description of biofortified corn cultivars released in India presented in Table 4.
9. Safeguarding Nutrients Beyond Harvest: Post-Harvest Strategies
9.1. Unveiling storeroom techniques to counter nutrient depletion
Most of the research efforts in agriculture regarding grains have primarily concentrated on increasing crop productivity in the field, while comparatively less attention has been given to maintaining the quality of grains after harvest, especially when stored with various methods to manage insect pests. It is crucial to balance the focus on enhancing grain production in the field with equally, if not greater, efforts to protect harvested grains from deterioration and maintain their nutritional value during storage. Insect-related postharvest losses are a significant cause of nutritional decline in grains, particularly affecting components like starch and protein. These nutritional losses can have adverse effects on the commodity's market price and can also negatively impact household nutrition and income [94]. Researchers have identified several storage methods that can help preserve the nutritional components of grains even after harvest. The predominant method for managing insect pests in stored food grains, including maize, has been the use of synthetic insecticides, which are now recognized for their associated health and environmental risks. Therefore, alternative nonchemical grain storage techniques have been proposed, including: (1) Combining filter cake powder with corn and storing it in woven polypropylene (WPP) bags, (2) Mixing Triplex dust with corn and storing it in WPP bags, (3) Using metal silos, (4) Employing the Purdue Improved Crop Storage bag, (5) Utilizing the Super GrainPro bag. These extended methods for secure storage empower farmers to preserve excess corn until the market during the lean season.
9.2. Vigilance against mold proliferation and mycotoxin contamination
Due to the conventional storage methods used in smallholder farms, corn is susceptible to significant damages when stored on the farm for an extended period. As a result, farmers are compelled to sell their surplus corn directly after harvest, when its market price is at its lowest point. Additionally, their personal maize stockpiles are prone to substantial quantitative losses when stored conventionally. A reduction in nutritional quality, the growth of mold, and the production of mycotoxins are significant quality deteriorations that can impact the grain reserves of smallholders. Changes in kernel color and the presence of mycotoxins have an adverse effect on the quality of stored grains and pose a threat to human health. The combination of excessive heat, elevated humidity, inadequate ventilation in the storage facilities, and damage from insects and rodents leads to the rapid multiplication and dissemination of fungal spores [95] However, the immediate concern is the missed opportunity for higher earnings due to contamination. One potential avenue for smallholder farmers to access a quality-conscious and guaranteed market for their maize is through the WFP's Purchase for Progress Program (P-P) [96]. The P-P program has established a maximum allowable limit of 20 micrograms/kilogram for aflatoxin in grains [97]. For controlling molds the utilization of flexible hermetic bags, capable of creating a modified atmosphere that can kill or deactivate the molds, along with airtight metal silos, has shown promise as techniques to safeguard the quality of maize.
10. Enlightened Choices: Empowering Consumers Through Education
In the worldwide maize sector, educating consumers plays a crucial role in promoting informed choices. Maize serves as a dietary staple for millions, and having a thorough understanding of its cultivation, processing, and nutritional content is of utmost importance. The focus of "Enlightened Choices for Maize" underscores the importance of educating consumers about this essential crop, aiming to enhance their nutrition and promote sustainable maize production. For instance, the importance of consumers being knowledgeable about maize's essential nutrients, including carbohydrates, fiber, vitamins, and minerals. Instructing them on effective processing techniques like nixtamalization and hydrothermal treatment aids in choosing maize products that are nutrient-rich. Empowering consumers also encompasses providing them with knowledge about sustainable maize farming practices and sourcing. Familiarity with biofortification initiatives motivates consumers to opt for maize products with enhanced nutritional value. Furthermore, offering instruction in various culinary methods and recipes featuring maize improves cooking skills and encourages the incorporation of maize into a wide range of dishes.
11. Championing Change: Success Stories and Global Impact
The first maize reference genome, was unveiled in 2009, initially known as B73 RefGen_v1 primarily assembled using the sequencing of bacterial artificial chromosomes and fosmids [98]. In 2017, a later release, empowered by state-of-the-art single-molecule technologies, marked a significant advancement. Through comparisons with other inbred lines, scientists have detected millions of insertions-deletions (InDels), single nucleotide polymorphisms (SNPs) etc, [99-106].
Plant gene research relies significantly on mutants. In contrast to specific techniques employed in Arabidopsis and rice, researchers in maize predominantly employ transposon mutagenesis for gene identification [107]. Numerous genes associated with significant plant characteristics have been unveiled through this approach, including tb1, which has an impact on branch quantity and flowering [108]. Numerous research initiatives, such as the Trait Utility System for Corn, have employed active Mu elements to tag genes in maize, for detailed study see Brutnell [109]. The primary program employed for this purpose is UniformMu [110]. Nevertheless, it covers approximately 30% of maize genes. Another method, referred to as Targeting Induced Local Lesions in Genomes (TILLING), has also been utilized to create mutant libraries in maize [111]. This approach entails subjecting plants to a chemical agent that induces mutations, subsequently detectable through advanced techniques or Next-Generation Sequencing (NGS).
Derived from teosinte, maize (Zea mays) has evolved into a widespread staple crop worldwide, characterized by its diverse traits, rendering it a fundamental model plant in research. In this section, we delve into maize domestication history, significant genes linked to domestication, genomic research resources, genes that have been cloned for enhancing crops, and provide insights into future directions in crop breeding and functional genomics. Up to the date, thousands of QTL and hundreds of genes had been identified in maize for various traits, including yield, disease resistance, and environmental adaptation [112-119], and numerous marker-trait associations (MTAs) have been identified related to different traits using genome wide association mapping, for detailed study see review published by Shikha et al. [120]. The Table 5 includes crucial domestication genes that have undergone selection, among the identified QTLs, MTAs, and candidate genes, for detailed study see Liu et al. [107].
MAS enhances breeding efficiency, saving time, eliminating extensive phenotyping, enabling early seedling selection, and improving low heritability traits [130]. Its usage is on the rise for hastening the creation of biofortified varieties by either selecting or incorporating key genes/QTLs [131]. In maize, advantageous gene variants have been pinpointed for characteristics such as opaqueness (o2, o16), β-carotene hydroxylase (crtRB1), lycopene epsilon cyclase (lcyE), γ-tocopherol methyl transferase (vte4), as well as lpa1-1 and lpa2-1, for a comprehensive study, refer to Hossain et al. [132]. Molecular markers for these genes are publicly available, facilitating their use in molecular breeding through different MAS approaches.
12. Tomorrow's Promise: Pioneering Nutritional Frontiers
As described in this chapter, emerging research and technology offer promising prospects. The production landscape of maize is transforming thanks to genomic insights, biofortification techniques, smart farming practices, and sustainable agriculture. Aside from alleviating hidden hunger, these advancements also promise to improve the health and well-being of communities that depend on maize for their primary sustenance. However, our journey toward a nourished future with maize is not without its challenges. Biotic and abiotic stress factors, socioeconomic barriers, consumer acceptance, and ethical considerations stand as formidable roadblocks. These challenges require concerted efforts from scientists, policymakers, farmers, and consumers alike to surmount. Hence, in this chapter, we have illuminated the obstacles that lie on the path to enhancing maize's nutritional potency while also presenting potential solutions, offering readers valuable insights into the journey ahead.
12.1. Identifying Roadblocks and Potential Solutions for Elevating Maize's Nutritional Potency
12.1.1. Biotic and Abiotic Stress Factors
Maize faces numerous challenges, including pests, diseases, and climate change-related stresses. These factors can reduce crop yields and affect nutrient accumulation. Integrated pest management and resilient maize varieties bred for stress tolerance are potential solutions to mitigate these issues.
Figure 2.
Pictorial description of common roadblocks and their potential solutions.
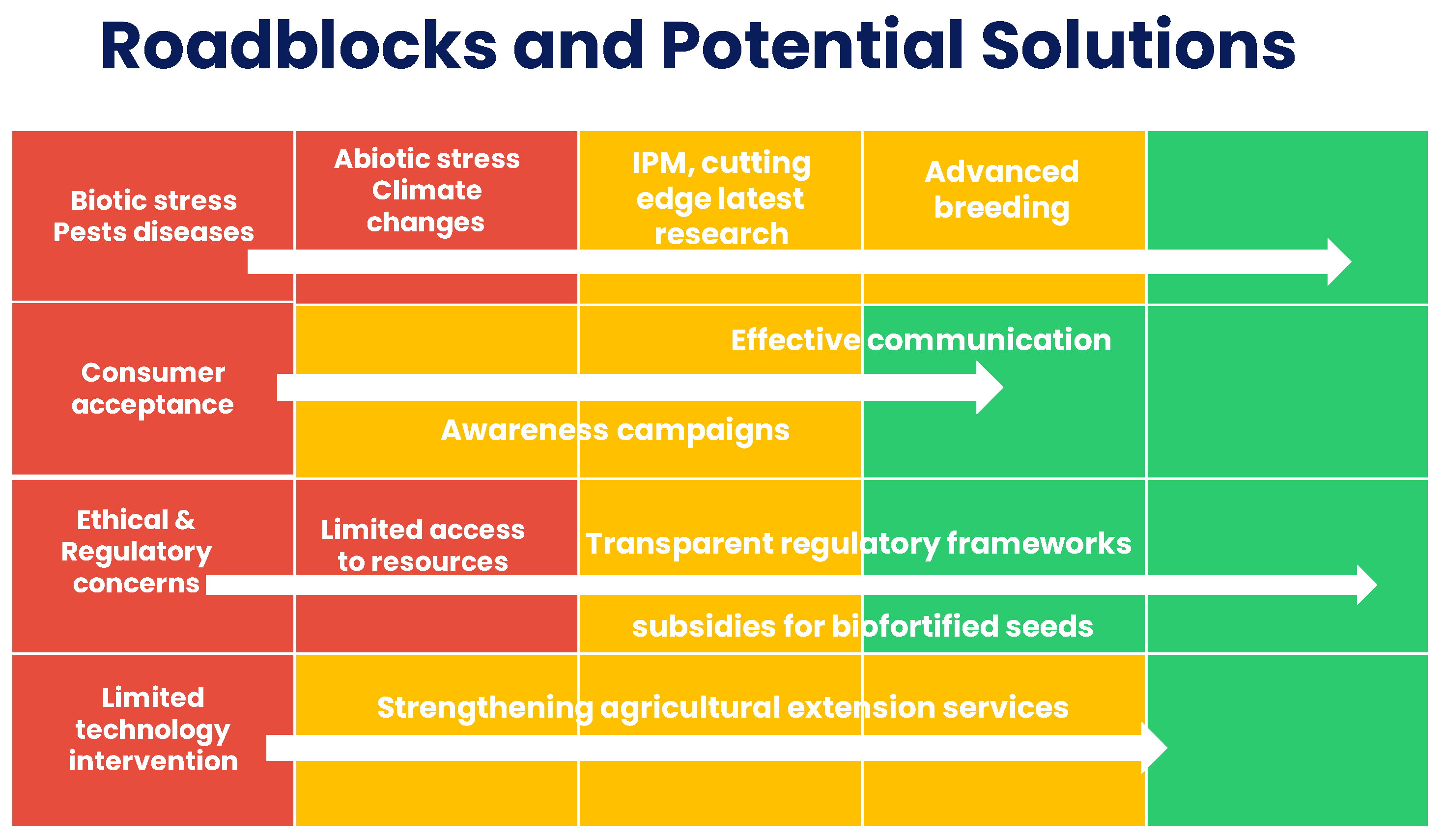
12.1.2. Socioeconomic Barriers
Access to nutrient-rich maize varieties and technologies is often limited in low-resource regions. Strengthening agricultural extension services, providing subsidies for biofortified seeds, and educating farmers on best practices can help bridge this gap and promote widespread adoption.
12.1.3. Consumer Acceptance
Consumer preferences and cultural habits can influence the acceptance of nutrient-enhanced maize varieties. Effective communication and awareness campaigns are essential to inform consumers about the health benefits and safety of these products.
12.1.4. Regulatory and Ethical Considerations
As biotechnology advances, ethical and regulatory concerns must be addressed. Striking a balance between innovation and safety is critical. Transparent regulatory frameworks and active involvement of the public can help establish trust in nutrient enhanced maize.
13. In Conclusion: Forging a Nourished Future with Maize
As researchers strive to enhance the value of maize, they find themselves at the crossroads of innovation, agriculture, and nutrition science. Maize, a food has significant potential to provide essential nutrients to millions of individuals, across the globe. The chapters outlined in this context shed light on the advancements in comprehending enhancing and optimizing maize nutrition. In summary this chapter emphasizes how research and technology hold power in elevating the worth of maize. By leveraging state of the art techniques and addressing challenges proactively maize researchers ensure that maize remains a source of nutrition for people globally especially in regions where it is a dietary mainstay. The vision of cultivating a maize crop goes beyond scientific pursuits; it represents their dedication to enhancing global food security and public health collectively. Collective investigations have the potential to transform tomorrow’s maize from being another grain into hope wellbeing and prosperity, for generations.
References
- Sah RP, KUMAR A, Rana M, Kumar U. Maize (Corn). Forage Crops of the World, 2-volume set: Volume I: Major Forage Crops; Volume II: Minor Forage Crops. 2022 May 29:15.
- Onuwa, G. Fostering Sustainable Productivity through Maize Technology Intensification: Participant responses. In Handbook of Research on Green Technologies for Sustainable Management of Agricultural Resources; IGI Global, 2022; pp. 427–436. [Google Scholar]
- Revilla, P.; Alves, M.L.; Andelković, V.; Balconi, C.; Dinis, I.; Mendes-Moreira, P.; Malvar, R.A. Traditional Foods From Maize (Zea mays L.) in Europe. Front. Nutr. 2022, 8, 683399. [Google Scholar] [CrossRef] [PubMed]
- Rouf Shah, T.; Prasad, K.; Kumar, P. Maize—A potential source of human nutrition and health: A review. Cogent Food Agric. 2016, 2, 1166995. [Google Scholar] [CrossRef]
- USDA. United States Department of Agriculture (USDA). National nutrient database for standard reference. https://fdc.nal.usda.gov/. 2020. [Google Scholar]
- Nuss, E.T.; Tanumihardjo, S.A. Maize: A paramount staple crop in the context of global nutrition. Comprehensive Reviews in Food Science and Food Safety 2010, 9, 417–436. [Google Scholar] [CrossRef] [PubMed]
- Sands, A.L.; Leidy, H.J.; Hamaker, B.R.; Maguire, P.; Campbell, W.W. Consumption of the slow-digesting waxy maize starch leads to blunted plasma glucose and insulin response but does not influence energy expenditure or appetite in humans. Nutr. Res. 2009, 29, 383–390. [Google Scholar] [CrossRef] [PubMed]
- Galani, Y.J.H.; Orfila, C.; Gong, Y.Y. A review of micronutrient deficiencies and analysis of maize contribution to nutrient requirements of women and children in Eastern and Southern Africa. Crit. Rev. Food Sci. Nutr. 2020, 62, 1568–1591. [Google Scholar] [CrossRef]
- Gibbon, B.C.; Larkins, B.A. Molecular genetic approaches to developing quality protein maize. Trends Genet. 2005, 21, 227–233. [Google Scholar] [CrossRef]
- Orthoefer, F.; Eastman, J.; List, G. Corn oil: Composition, processing, and utilization. In Corn: Chemistry and Technology, 2nd ed.; 2003; pp. 671–693. [Google Scholar]
- CRA. Corn oil, 5th ed.; Corn Refiners Association: Washington, DC, 2006. [Google Scholar]
- Kumar, D.; Jhariya, A.N. Nutritional, medicinal and economical importance of corn: A mini review. Res J Pharm Sci 2013, 2319, 555X. [Google Scholar]
- Sen, C.K.; Khanna, S.; Roy, S. Tocotrienols: Vitamin E beyond tocopherols. Life Sci. 2006, 78, 2088–2098. [Google Scholar] [CrossRef]
- Kopsell, D.A.; Armel, G.R.; Mueller, T.C.; Sams, C.E.; Deyton, D.E.; McElroy, J.S.; Kopsell, D.E. Increase in nutritionally important sweet corn kernel carotenoids following mesotrione and atrazine applications. Journal of Agricultural and Food Chemistry 2009, 57, 6362–6368. [Google Scholar] [CrossRef]
- Demeke, K.H. Nutritional Quality Evaluation of Seven Maize Varieties Grown in Ethiopia. Biochem. Mol. Biol. 2018, 3, 45. [Google Scholar] [CrossRef]
- Harrabi, S.; St-Amand, A.; Sakouhi, F.; Sebei, K.; Kallel, H.; Mayer, P.M.; Boukhchina, S. Phytostanols and phytosterols distributions in corn kernel. Food Chemistry 2008, 111, 115–120. [Google Scholar] [CrossRef]
- Murdia, L.K.; Wadhwani, R.; Wadhawan, N.; Bajpai, P.; Shekhawat, S. Maize utilization in India: An overview. American Journal of Food and Nutrition 2016, 4, 169–176. [Google Scholar]
- Eckhoff, S. R.; Paulsen, M. R. Maize. In Cereal grain quality; Springer: Dordrecht, The Netherlands, 1996; pp. 77–112. [Google Scholar]
- Aliyu, K.T.; Huising, J.; Kamara, A.Y.; Jibrin, J.M.; Mohammed, I.B.; Nziguheba, G.; Adam, A.M. Understanding nutrient imbalances in maize (Zea mays L.) using the diagnosis and recommendation integrated system (DRIS) approach in the Maize belt of Nigeria. Scientific Reports 2021, 11, 16018. [Google Scholar] [CrossRef] [PubMed]
- Gwirtz, J.A.; Garcia-Casal, M.N. Processing maize flour and corn meal food products. Ann. New York Acad. Sci. 2013, 1312, 66–75. [Google Scholar] [CrossRef]
- Suri, D.J.; Tanumihardjo, S.A. Effects of different processing methods on the micronutrient and phytochemical contents of maize: From A to Z. Comprehensive Reviews in Food Science and Food Safety 2016, 15, 912–926. [Google Scholar] [CrossRef]
- Brodowska, M.S.; Wyszkowski, M.; Bujanowicz-Haraś, B. Mineral Fertilization and Maize Cultivation as Factors Which Determine the Content of Trace Elements in Soil. Agronomy 2022, 12, 286. [Google Scholar] [CrossRef]
- Abdel-Aal, E.-S.M.; Akhtar, H.; Zaheer, K.; Ali, R. Dietary Sources of Lutein and Zeaxanthin Carotenoids and Their Role in Eye Health. Nutrients 2013, 5, 1169–1185. [Google Scholar] [CrossRef]
- Gopalan, C.; Rama Sastri, B. V.; Balasubramanian, S. C. Nutritive value of Indian foods; National Institute of Nutrition, Indian Council of Medical Research, 1971. [Google Scholar]
- Gensberger, S.; Mittelmaier, S.; Glomb, M.A.; Pischetsrieder, M. Identification and quantification of six major α-dicarbonyl process contaminants in high-fructose corn syrup. Anal. Bioanal. Chem. 2012, 403, 2923–2931. [Google Scholar] [CrossRef]
- Tappy, L.; Lê, K.-A. Metabolic Effects of Fructose and the Worldwide Increase in Obesity. Physiol. Rev. 2010, 90, 23–46. [Google Scholar] [CrossRef]
- Parker, K.; Salas, M.; Nwosu, V.C. High fructose corn syrup: Production, uses and public health concerns. Biotechnol Mol Biol Rev 2010, 5, 71–78. [Google Scholar]
- Bawa, A.S.; Anilakumar, K.R. Genetically modified foods: Safety, risks, and public concerns—A review. Journal of Food Science and Technology 2013, 50, 1035–1046. [Google Scholar] [CrossRef] [PubMed]
- Norris, M. L. Will GMOs hurt my body? The public’s concerns and how scientists have addressed them. Science in the News, 2015. [Google Scholar]
- Li, Y.; Zhang, L.; Liu, H.; Yang, Y.; He, J.; Cao, M.; Zhong, W.; Lin, Y.; Zhuo, Y.; Fang, Z.; et al. Effects of the Ratio of Insoluble Fiber to Soluble Fiber in Gestation Diets on Sow Performance and Offspring Intestinal Development. Animals 2019, 9, 422. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.W.; E Jones, A.; Riddell-Mason, S. Ten Different Dietary Fibers Have Significantly Different Effects on Serum and Liver Lipids of Cholesterol-Fed Rats. J. Nutr. 1994, 124, 78–83. [Google Scholar] [CrossRef] [PubMed]
- Bouis, H.E.; Saltzman, A. Improving nutrition through biofortification: A review of evidence from Harvest Plus, 2003 through 2016. Global food security 2017, 12, 49–58. [Google Scholar] [CrossRef]
- Manjeru, P.; Van Biljon, A.; Labuschagne, M. The development and release of maize fortified with provitamin A carotenoids in developing countries. Crit. Rev. Food Sci. Nutr. 2017, 59, 1284–1293. [Google Scholar] [CrossRef]
- Gannon, B.; Kaliwile, C.; A Arscott, S.; Schmaelzle, S.; Chileshe, J.; Kalungwana, N.; Mosonda, M.; Pixley, K.; Masi, C.; A Tanumihardjo, S. Biofortified orange maize is as efficacious as a vitamin A supplement in Zambian children even in the presence of high liver reserves of vitamin A: A community-based, randomized placebo-controlled trial. Am. J. Clin. Nutr. 2014, 100, 1541–1550. [Google Scholar] [CrossRef]
- Palmer, A.C.; Healy, K.; Barffour, M.A.; Siamusantu, W.; Chileshe, J.; Schulze, K.J.; Labrique, A.B. Provitamin A carotenoid–biofortified maize consumption increases pupillary responsiveness among Zambian children in a randomized controlled trial. The Journal of nutrition 2016, 146, 2551–2558. [Google Scholar] [CrossRef]
- Zuma, M.K.; Kolanisi, U.; Modi, A.T. The Potential of Integrating Provitamin A-Biofortified Maize in Smallholder Farming Systems to Reduce Malnourishment in South Africa. Int. J. Environ. Res. Public Health 2018, 15, 805. [Google Scholar] [CrossRef]
- White, P.; Broadley, M. Biofortifying crops with essential mineral elements. Trends Plant Sci. 2005, 10, 586–593. [Google Scholar] [CrossRef]
- Zhu, C.; Naqvi, S.; Gomez-Galera, S.; Pelacho, A.M.; Capell, T.; Christou, P. Transgenic strategies for the nutritional enhancement of plants. Trends Plant Sci. 2007, 12, 548–555. [Google Scholar] [CrossRef]
- Grusak, M.A. Enhancing Mineral Content in Plant Food Products. In Proceedings of the 42nd Annual Meeting of the American-College-of-Nutrition; pp. 178S–183S.
- Roberts, L.A.; Pierson, A.J.; Panaviene, Z.; Walker, E.L. Yellow Stripe1. Expanded Roles for the Maize Iron-Phytosiderophore Transporter. Plant Physiol. 2004, 135, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Drakakaki, G.; Marcel, S.; Glahn, R.P.; Lund, E.K.; Pariagh, S.; Fischer, R.; Stoger, E. Endosperm-specific co-expression of recombinant soybean ferritin and Aspergillus phytase in maize results in significant increases in the levels of bioavailable iron. Plant Molecular Biology 2005, 59, 869–880. [Google Scholar] [CrossRef] [PubMed]
- Davila-Hicks, P.; Theil, E.C.; Lönnerdal, B. Iron in ferritin or in salts (ferrous sulfate) is equally bioavailable in nonanemic women. Am. J. Clin. Nutr. 2004, 80, 936–940. [Google Scholar] [CrossRef] [PubMed]
- Lönnerdal, B.; Bryant, A.; Liu, X.; Theil, E.C. Iron absorption from soybean ferritin in nonanemic women. Am. J. Clin. Nutr. 2006, 83, 103–107. [Google Scholar] [CrossRef]
- Brown, K.H.; Rivera, J.A.; Bhutta, Z.; Gibson, R.S.; King, J.C.; Lonnerdal, B.; Ruel, B.; Sandtrom, B.; Wasantwisut, E.; Hotz, C. International Zinc Nutrition Consultative Group: Assessment of the risk of zinc deficiency in populations and options for its control. The Food and Nutrition Bulletin 2004, 25, S99–203. [Google Scholar]
- Hotz, C. The Potential to Improve Zinc Status through Biofortification of Staple Food Crops with Zinc. Food Nutr. Bull. 2009, 30, S172–S178. [Google Scholar] [CrossRef]
- Whitt, S.R.; Wilson, L.M.; Tenaillon, M.I.; Gaut, B.S.; Buckler IV, E.S. Genetic diversity and selection in the maize starch pathway. Proceedings of the National Academy of Sciences 2002, 99, 12959–12962. [Google Scholar] [CrossRef]
- Palaisa, K.; Morgante, M.; Tingey, S.; Rafalski, A. Long-range patterns of diversity and linkage disequilibrium surrounding the maize Y1 gene are indicative of an asymmetric selective sweep. Proceedings of the National Academy of Sciences 2004, 101, 9885–9890. [Google Scholar] [CrossRef]
- Karn, A.; Gillman, J.D.; A Flint-Garcia, S. Genetic Analysis of Teosinte Alleles for Kernel Composition Traits in Maize. G3 Genes|Genomes|Genetics 2017, 7, 1157–1164. [Google Scholar] [CrossRef]
- Fan, L.; Bao, J.; Wang, Y.; Yao, J.; Gui, Y.; Hu, W.; Zhu, J.; Zeng, M.; Li, Y.; Xu, Y. Post-Domestication Selection in the Maize Starch Pathway. PLoS ONE 2009, 4, e7612. [Google Scholar] [CrossRef]
- Huang, Y.; Wang, H.; Zhu, Y.; Huang, X.; Li, S.; Wu, X.; Zhao, Y.; Bao, Z.; Qin, L.; Jin, Y.; et al. THP9 enhances seed protein content and nitrogen-use efficiency in maize. Nature 2022, 612, 292–300. [Google Scholar] [CrossRef] [PubMed]
- Turnbull, C.; Lillemo, M.; Hvoslef-Eide, T.A. Global regulation of genetically modified crops amid the gene edited crop boom–a review. Frontiers in Plant Science 2021, 12, 630396. [Google Scholar] [CrossRef] [PubMed]
- Kettenburg, A.J.; Hanspach, J.; Abson, D.J.; Fischer, J. From disagreements to dialogue: Unpacking the Golden Rice debate. Sustain. Sci. 2018, 13, 1469–1482. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Yu, Y.; Li, G.; Xie, L.; Guo, X.; Li, J.; Li, Y.; Hu, J. Genome-wide association study of vitamin E in sweet corn kernels. Crop. J. 2019, 8, 341–350. [Google Scholar] [CrossRef]
- Waltz, E. CRISPR-edited crops free to enter market, skip regulation. Nat. Biotechnol. 2016, 34, 582. [Google Scholar] [CrossRef] [PubMed]
- Zilberman, D.; Holland, T.G.; Trilnick, I. Agricultural GMOs—What we know and where scientists disagree. Sustainability 2018, 10, 1514. [Google Scholar] [CrossRef]
- Chrenková, M.; Sommer, A.; Čerešňáková, Z.; Nitrayová, S.; Prostredná, M. Nutritional evaluation of genetically modified maize corn performed on rats. Archives of Animal Nutrition 2002, 56, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Venneria, E.; Fanasca, S.; Monastra, G.; Finotti, E.; Ambra, R.; Azzini, E.; Durazzo, A.; Foddai, M.S.; Maiani, G. Assessment of the Nutritional Values of Genetically Modified Wheat, Corn, and Tomato Crops. J. Agric. Food Chem. 2008, 56, 9206–9214. [Google Scholar] [CrossRef] [PubMed]
- Baktavachalam, G.B.; Delaney, B.; Fisher, T.L.; Ladics, G.S.; Layton, R.J.; Locke, M.E.; Schmidt, J.; A Anderson, J.; Weber, N.N.; A Herman, R.; et al. Transgenic maize event TC1507: Global status of food, feed, and environmental safety. GM Crop. Food 2015, 6, 80–102. [Google Scholar] [CrossRef]
- EFSA Panel on Genetically Modified Organisms (EFSA GMO Panel); Naegeli, H.; Bresson, J.L.; Dalmay, T.; Dewhurst, I.C.; Epstein, M.M.; Koukoulanaki, M. Assessment of genetically modified maize Bt11× MIR162× MIR604× 1507× 5307× GA21 and subcombinations, for food and feed uses, under Regulation (EC) No 1829/2003 (application EFSA-GMO-DE-2011-103). EFSA Journal 2019, 17, e05635. [Google Scholar]
- Boyle, J.H.; Dalgleish, H.J.; Puzey, J.R. Monarch butterfly and milkweed declines substantially predate the use of genetically modified crops. Proc. Natl. Acad. Sci. 2019, 116, 3006–3011. [Google Scholar] [CrossRef] [PubMed]
- Pellegrino, E.; Bedini, S.; Nuti, M.; Ercoli, L. Impact of genetically engineered maize on agronomic, environmental, and toxicological traits: A meta-analysis of 21 years of field data. Scientific Reports 2018, 8, 3113. [Google Scholar] [CrossRef] [PubMed]
- Woźniak, E.; Tyczewska, A.; Twardowski, T. A Shift Towards Biotechnology: Social Opinion in the EU. Trends Biotechnol. 2020, 39, 214–218. [Google Scholar] [CrossRef]
- Hull, R.; Head, G.; Tzotzos, G. T. (Eds.) Evolution of regulatory systems and national biosafety frameworks. In Genetically Modified Plants, 2nd ed.; Elsevier: Cambridge, Massachusetts, 2021; pp. 127–155. [Google Scholar]
- Cakmak, I. Agronomic Biofortification; International Food Policy Research Institute: Washington, DC, USA, 2014; pp. 56–97. [Google Scholar]
- Dimkpa, C.O.; Bindraban, P.S. Fortification of micronutrients for efficient agronomic production: A review. Agron. Sustain. Dev. 2016, 36. [Google Scholar] [CrossRef]
- Vanlauwe, B.; Descheemaeker, K.; Giller, K.E.; Huising, J.; Merckx, R.; Nziguheba, G.; Zingore, S. Integrated soil fertility management in sub-Saharan Africa: Unravelling local adaptation. Soil 2015, 1, 491–508. [Google Scholar] [CrossRef]
- Teklu, D.; Gashu, D.; Joy, E.J.M.; Amede, T.; Broadley, M.R. Effectiveness of Agronomic Biofortification Strategy in Fighting against Hidden Hunger. Agronomy 2023, 13, 2173. [Google Scholar] [CrossRef]
- Ngigi, P.B.; Lachat, C.; Masinde, P.W.; Du Laing, G. Agronomic biofortification of maize and beans in Kenya through selenium fertilization. Environ. Geochem. Health 2019, 41, 2577–2591. [Google Scholar] [CrossRef]
- Botoman, L.; Chimungu, J.G.; Bailey, E.H.; Munthali, M.W.; Ander, E.L.; Mossa, A.; Young, S.D.; Broadley, M.R.; Lark, R.M.; Nalivata, P.C. Agronomic biofortification increases grain zinc concentration of maize grown under contrasting soil types in Malawi. Plant Direct 2022, 6, e458. [Google Scholar] [CrossRef]
- Umar, W.; Hameed, M.K.; Aziz, T.; Maqsood, M.A.; Bilal, H.M.; Rasheed, N. Synthesis, characterization and application of ZnO nanoparticles for improved growth and Zn biofortification in maize. Arch. Agron. Soil Sci. 2020, 67, 1164–1176. [Google Scholar] [CrossRef]
- Serna-Saldivar, S.O.; Carrillo, E.P. Food uses of whole corn and dry-milled fractions. In Corn; AACC International Press: St. Paul, MN, USA, 2019; pp. 435–467. [Google Scholar]
- Palacios-Rojas, N.; McCulley, L.; Kaeppler, M.; Titcomb, T.J.; Gunaratna, N.S.; Lopez-Ridaura, S.; Tanumihardjo, S.A. Mining maize diversity and improving its nutritional aspects within agro-food systems. Compr. Rev. Food Sci. Food Saf. 2020, 19, 1809–1834. [Google Scholar] [CrossRef]
- Gallego-Castillo, S.; Taleon, V.; Talsma, E.F.; Rosales-Nolasco, A.; Palacios-Rojas, N. Effect of maize processing methods on the retention of minerals, phytic acid and amino acids when using high kernel-zinc maize. Curr. Res. Food Sci. 2021, 4, 279–286. [Google Scholar] [CrossRef] [PubMed]
- Sahasrabudhe, S. N. Corn characterization and development of a convenient laboratory scale alkaline cooking process. 2015. [Google Scholar]
- Sefa-Dedeh, S.; Cornelius, B.; Sakyi-Dawson, E.; Afoakwa, E.O. Effect of nixtamalization on the chemical and functional properties of maize. Food Chem. 2004, 86, 317–324. [Google Scholar] [CrossRef]
- González, R.; Reguera, E.; Mendoza, L.; Figueroa, J.M.; Sánchez-Sinencio, F. Physicochemical Changes in the Hull of Corn Grains during Their Alkaline Cooking. J. Agric. Food Chem. 2004, 52, 3831–3837. [Google Scholar] [CrossRef]
- Mora-Avilés, A.; Lemus-Flores, B.; Miranda-López, R.; Hernández-López, D.; Pons-Hernández, J.L.; Acosta-Gallegos, J.A.; Guzmán-Maldonado, S.H. Effects of common bean enrichment on nutritional quality of tortillas produced from nixtamalized regular and quality protein maize flours. Journal of the Science of Food and Agriculture 2007, 87, 880–886. [Google Scholar] [CrossRef]
- Serna-Saldivar, S.O. Understanding the functionality and manufacturing of nixtamalized maize products. J. Cereal Sci. 2021, 99, 103205. [Google Scholar] [CrossRef]
- Serna-Saldivar SO, Chuck-Hernandez C. Food uses of lime-cooked corn with emphasis in tortillas and snacks. In Corn; AACC International Press, 2019; pp. 469–500.
- Schaarschmidt, S.; Fauhl-Hassek, C. Mycotoxins during the Processes of Nixtamalization and Tortilla Production. Toxins 2019, 11, 227. [Google Scholar] [CrossRef] [PubMed]
- Rocha-Villarreal, V.; Hoffmann, J.F.; Vanier, N.L.; Serna-Saldivar, S.O.; García-Lara, S. Hydrothermal treatment of maize: Changes in physical, chemical, and functional properties. Food Chem. 2018, 263, 225–231. [Google Scholar] [CrossRef]
- Akinbolade, J.A.; Igbeka, J.C. Hydrothermal reactions on selected utilization qualities of parboiled Maize. Agricultural Engineering International: CIGR Journal 2012, 14, 238–245. [Google Scholar]
- Oladiran, D.A.; Emmambux, N.M. Locally Available African Complementary Foods: Nutritional Limitations and Processing Technologies to Improve Nutritional Quality—A Review. Food Rev. Int. 2020, 38, 1033–1063. [Google Scholar] [CrossRef]
- Goredema-Matongera, N.; Ndhlela, T.; Magorokosho, C.; Kamutando, C. N.; van Biljon, A.; Labuschagne, M. Multinutrient biofortification of maize (Zea mays L.) in Africa: Current status, opportunities and limitations. Nutrients 2021, 13, 1039. [Google Scholar] [CrossRef]
- Akinsola, O.T.; Alamu, E.O.; Otegbayo, B.O.; Menkir, A.; Maziya-Dixon, B. Evaluation of Quality and Acceptability of Snack (Kokoro) Produced From Synthetic Provitamin A Maize (Zea mays) Genotypes. Front. Sustain. Food Syst. 2020, 4. [Google Scholar] [CrossRef]
- Batool, M. Nutrient Management of Maize. 2023. [Google Scholar]
- Langyan, S.; Bhardwaj, R.; Kumari, J.; Jacob, S.R.; Bisht, I.S.; Pandravada, S.R.; Singh, A.; Singh, P.B.; Dar, Z.A.; Kumar, A.; et al. Nutritional Diversity in Native Germplasm of Maize Collected From Three Different Fragile Ecosystems of India. Front. Nutr. 2022, 9, 812599. [Google Scholar] [CrossRef] [PubMed]
- Qu, J.-H.; Liu, D.; Cheng, J.-H.; Sun, D.-W.; Ma, J.; Pu, H.; Zeng, X.-A. Applications of Near-infrared Spectroscopy in Food Safety Evaluation and Control: A Review of Recent Research Advances. Crit. Rev. Food Sci. Nutr. 2013, 55, 1939–1954. [Google Scholar] [CrossRef] [PubMed]
- Payne, W.Z.; Kurouski, D. Raman spectroscopy enables phenotyping and assessment of nutrition values of plants: A review. Plant Methods 2021, 17, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Abeshu, M.A.; Lelisa, A.; Geleta, B. Complementary feeding: Review of recommendations, feeding practices, and adequacy of homemade complementary food preparations in developing countries–lessons from Ethiopia. Frontiers in nutrition 2016, 3, 41. [Google Scholar] [CrossRef]
- Maqbool, M.A.; Issa, A.B.; Khokhar, E.S. Quality protein maize (QPM): Importance, genetics, timeline of different events, breeding strategies and varietal adoption. Plant Breed. 2021, 140, 375–399. [Google Scholar] [CrossRef]
- Kumar, P.; Choudhary, M.; Hossain, F.; Singh, N.K.; Choudhary, P.; Gupta, M.; Singh, V.; Chikappa, G.K.; Kumar, R.; Kumar, B.; et al. Nutritional quality improvement in maize (Zea mays): Progress and challenges. Indian J. Agric. Sci. 2019, 89, 895–911. [Google Scholar] [CrossRef]
- Yadava, D. K.; Choudhury, P. R.; Hossain, F.; Kumar, D. Biofortified varieties: Sustainable way to alleviate malnutrition; Indian Council of Agricultural Research: New Delhi, 2017. [Google Scholar]
- Otitodun, G.; Ala, A.; Nwaubani, S.; Omobowale, M.; Ajao, S.; Ogundare, M.; Olenloa, A.; Busari, G.; Abel, G.; Braimah, J.; et al. Assessing efficacies of insect pest management methods to preserve nutritional composition of bagged maize in storehouses located in markets in Nigeria. Afr. J. Food Agric. Nutr. Dev. 2021, 21, 17972–17988. [Google Scholar] [CrossRef]
- Hell, K.; Mutegi, C.; Fandohan, P. Aflatoxin control and prevention strategies in maize for Sub-Saharan Africa. Julius-Kühn-Archiv 2010, 425, 534. [Google Scholar]
- Hodges, R.; Bennett, B.; Bernard, M.; Rembold, F. Tackling post-harvest cereal losses in sub-Saharan Africa. Rural 21: The International Journal for Rural Development 2013, 47, 16–18. [Google Scholar]
- Suleiman, R. A.; Kurt, R. A. Current maize production, postharvest losses, and the risk of mycotoxins contamination in Tanzania. In 2015 ASABE Annual International Meeting; American Society of Agricultural and Biological Engineers, 2015; p. 1. [Google Scholar]
- Schnable, P.S.; Ware, D.; Fulton, R.S.; Stein, J.C.; Wei, F.; Pasternak, S.; Liang, C.; Zhang, J.; Fulton, L.; Graves, T.A.; Minx, P. The B73 maize genome: Complexity, diversity, and dynamics. Science 2009, 20, 1112–1115. [Google Scholar] [CrossRef] [PubMed]
- Bukowski, R.; Guo, X.; Lu, Y.; Zou, C.; He, B.; Rong, Z.; Wang, B.; Xu, D.; Yang, B.; Xie, C.; et al. Construction of the third-generation Zea mays haplotype map. GigaScience 2017, 7, gix134. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Xu, X.; Jin, W.; Xu, M.; Zhao, H.; Xiang, Z.; Song, W.; Ying, K.; Zhang, M.; Jiao, Y.; et al. Genome-wide patterns of genetic variation among elite maize inbred lines. Nat. Genet. 2010, 42, 1027–1030. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.; Liu, H.; He, C.; Fu, J.; Xiao, Y.; Wang, Y.; Xie, W.; Wang, G.; Yan, J. Maize pan-transcriptome provides novel insights into genome complexity and quantitative trait variation. Sci. Rep. 2016, 6, 18936. [Google Scholar] [CrossRef]
- Springer, N.M.; Ying, K.; Fu, Y.; Ji, T.; Yeh, C.-T.; Jia, Y.; Wu, W.; Richmond, T.; Kitzman, J.; Rosenbaum, H.; et al. Maize Inbreds Exhibit High Levels of Copy Number Variation (CNV) and Presence/Absence Variation (PAV) in Genome Content. PLOS Genet. 2009, 5, e1000734. [Google Scholar] [CrossRef]
- Fu, J.; Cheng, Y.; Linghu, J.; Yang, X.; Kang, L.; Zhang, Z.; Zhang, J.; He, C.; Du, X.; Peng, Z.; et al. RNA sequencing reveals the complex regulatory network in the maize kernel. Nat. Commun. 2013, 4, 2832. [Google Scholar] [CrossRef]
- Hirsch, C.N.; Foerster, J.M.; Johnson, J.M.; Sekhon, R.S.; Muttoni, G.; Vaillancourt, B.; Peñagaricano, F.; Lindquist, E.; Pedraza, M.A.; Barry, K.; et al. Insights into the Maize Pan-Genome and Pan-Transcriptome. Plant Cell 2014, 26, 121–135. [Google Scholar] [CrossRef]
- Jiao, Y.; Zhao, H.; Ren, L.; Song, W.; Zeng, B.; Guo, J.; Wang, B.; Liu, Z.; Chen, J.; Li, W.; et al. Genome-wide genetic changes during modern breeding of maize. Nat. Genet. 2012, 44, 812–815. [Google Scholar] [CrossRef]
- Yang, N.; Liu, J.; Gao, Q.; Gui, S.; Chen, L.; Yang, L.; Huang, J.; Deng, T.; Luo, J.; He, L.; et al. Genome assembly of a tropical maize inbred line provides insights into structural variation and crop improvement. Nat. Genet. 2019, 51, 1052–1059. [Google Scholar] [CrossRef]
- Liu, J.; Fernie, A.R.; Yan, J. The Past, Present, and Future of Maize Improvement: Domestication, Genomics, and Functional Genomic Routes toward Crop Enhancement. Plant Commun. 2019, 1, 100010. [Google Scholar] [CrossRef]
- Doebley, J.; Stec, A.; Hubbard, L. The evolution of apical dominance in maize. Nature 1997, 386, 485–488. [Google Scholar] [CrossRef]
- Brutnell, T.P. Transposon tagging in maize. Functional & integrative genomics 2002, 2, 4–12. [Google Scholar]
- Mccarty, D.R.; Mark Settles, A.; Suzuki, M.; Tan, B.C.; Latshaw, S.; Porch, T.; Robin, K.; Baier, J.; Avigne, W.; Lai, J.; et al. Steady-state transposon mutagenesis in inbred maize. Plant J. 2005, 44, 52–61. [Google Scholar] [CrossRef] [PubMed]
- Till, B.J.; Reynolds, S.H.; Weil, C.; Springer, N.; Burtner, C.; Young, K.; Bowers, E.; Codomo, C.A.; Enns, L.C.; Odden, A.R.; Greene, E.A.; Comai, L. Discovery of induced point mutations in maize genes by TILLING. BMC Plant Biology 2004, 4, 12. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.-Y.; Li, Y.; Zhang, C.-Q. QTL mapping for stay-green in maize (Zea mays). Can. J. Plant Sci. 2012, 92, 249–256. [Google Scholar] [CrossRef]
- Zhao, X.; Peng, Y.; Zhang, J.; Fang, P.; Wu, B. Identification of QTLs and Meta-QTLs for Seven Agronomic Traits in Multiple Maize Populations under Well-Watered and Water-Stressed Conditions. Crop. Sci. 2018, 58, 507–520. [Google Scholar] [CrossRef]
- Wang, G.; Zhao, Y.; Mao, W.; Ma, X.; Su, C. QTL Analysis and Fine Mapping of a Major QTL Conferring Kernel Size in Maize (Zea mays). Front. Genet. 2020, 11. [Google Scholar] [CrossRef]
- Badu-Apraku, B.; Adewale, S.; Paterne, A.A.; Gedil, M.; Toyinbo, J.; Asiedu, R. Identification of QTLs for grain yield and other traits in tropical maize under Striga infestation. PLoS ONE 2020, 15, e0239205. [Google Scholar] [CrossRef]
- Yang, J.; Liu, Z.; Chen, Q.; Qu, Y.; Tang, J.; Lübberstedt, T.; Li, H. Mapping of QTL for grain yield components based on a DH population in maize. Scientific Reports 2020, 10, 7086. [Google Scholar] [CrossRef]
- Wang, L.; Zhou, Z.; Li, R.; Weng, J.; Zhang, Q.; Li, X.; Wang, B.; Zhang, W.; Song, W.; Li, X. Mapping QTL for flowering time-related traits under three plant densities in maize. Crop. J. 2020, 9, 372–379. [Google Scholar] [CrossRef]
- Fei, J.; Lu, J.; Jiang, Q.; Liu, Z.; Yao, D.; Qu, J.; Liu, S.; Guan, S.; Ma, Y. Maize plant architecture trait QTL mapping and candidate gene identification based on multiple environments and double populations. BMC Plant Biol. 2022, 22, 110. [Google Scholar] [CrossRef]
- Wang, M.; Zhang, R.; Zhao, Y.; Yao, J.; Li, W.; Yang, Z.; Sun, F.; Yang, X. Identifying QTL and candidate genes for prolificacy in maize. Crop. J. 2023, 11, 531–539. [Google Scholar] [CrossRef]
- Shikha, K.; Shahi, J.P.; Vinayan, M.T.; Zaidi, P.H.; Singh, A.K.; Sinha, B. Genome-wide association mapping in maize: Status and prospects. 3 Biotech 2021, 11, 244. [Google Scholar] [CrossRef] [PubMed]
- Wills, D.M.; Whipple, C.J.; Takuno, S.; Kursel, L.E.; Shannon, L.M.; Ross-Ibarra, J.; Doebley, J.F. From many, one: Genetic control of prolificacy during maize domestication. PLoS Genetics 2013, 9, e1003604. [Google Scholar] [CrossRef] [PubMed]
- Whipple, C.J.; Kebrom, T.H.; Weber, A.L.; Yang, F.; Hall, D.; Meeley, R.; Schmidt, R.; Doebley, J.; Brutnell, T.P.; Jackson, D.P. grassy tillers1 promotes apical dominance in maize and responds to shade signals in the grasses. Proceedings of the National Academy of Sciences 2011, 108, E506–E512. [Google Scholar] [CrossRef]
- Wang, J.; Lin, Z.; Zhang, X.; Liu, H.; Zhou, L.; Zhong, S.; Li, Y.; Zhu, C.; Lin, Z. krn1, a major quantitative trait locus for kernel row number in maize. New Phytol. 2019, 223, 1634–1646. [Google Scholar] [CrossRef]
- Sigmon, B.; Vollbrecht, E. Evidence of selection at the ramosa1 locus during maize domestication. Mol. Ecol. 2010, 19, 1296–1311. [Google Scholar] [CrossRef]
- Studer, A.; Zhao, Q.; Ross-Ibarra, J.; Doebley, J. Identification of a functional transposon insertion in the maize domestication gene tb1. Nature Genetics 2011, 43, 1160–1163. [Google Scholar] [CrossRef]
- Wang, H.; Nussbaum-Wagler, T.; Li, B.; Zhao, Q.; Vigouroux, Y.; Faller, M.; Bomblies, K.; Lukens, L.; Doebley, J.F. The origin of the naked grains of maize. Nature 2005, 436, 714–719. [Google Scholar] [CrossRef]
- Dong, Z.; Li, W.; Unger-Wallace, E.; Yang, J.; Vollbrecht, E.; Chuck, G. Ideal crop plant architecture is mediated by tassels replace upper ears1, a BTB/POZ ankyrin repeat gene directly targeted by TEOSINTE BRANCHED1. Proceedings of the National Academy of Sciences 2017, 114, E8656–E8664. [Google Scholar] [CrossRef]
- Lin, Z.; Li, X.; Shannon, L.M.; Yeh, C.-T.; Wang, M.L.; Bai, G.; Peng, Z.; Li, J.; Trick, H.N.; E Clemente, T.; et al. Parallel domestication of the Shattering1 genes in cereals. Nat. Genet. 2012, 44, 720–724. [Google Scholar] [CrossRef] [PubMed]
- Sosso, D.; Luo, D.; Li, Q.-B.; Sasse, J.; Yang, J.; Gendrot, G.; Suzuki, M.; E Koch, K.; McCarty, D.R.; Chourey, P.S.; et al. Seed filling in domesticated maize and rice depends on SWEET-mediated hexose transport. Nat. Genet. 2015, 47, 1489–1493. [Google Scholar] [CrossRef] [PubMed]
- Cobb, J.N.; Juma, R.U.; Biswas, P.S.; Arbelaez, J.D.; Rutkoski, J.; Atlin, G.; Hagen, T.; Quinn, M.; Ng, E.H. Enhancing the rate of genetic gain in public-sector plant breeding programs: Lessons from the breeder’s equation. Theor. Appl. Genet. 2019, 132, 627–645. [Google Scholar] [CrossRef] [PubMed]
- Prasanna, B.M.; Palacios-Rojas, N.; Hossain, F.; Muthusamy, V.; Menkir, A.; Dhliwayo, T.; Ndhlela, T.; Vicente, F.S.; Nair, S.K.; Vivek, B.S.; et al. Molecular Breeding for Nutritionally Enriched Maize: Status and Prospects. Front. Genet. 2020, 10, 1392. [Google Scholar] [CrossRef]
- Hossain, F.; Zunjare, R.U.; Muthusamy, V.; Bhat, J.S.; Mehta, B.K.; Sharma, D.; Talukder, Z.A.; Chhabra, R.; Katral, A.; Dutta, S.; Chand, G. Biofortification of maize for nutritional security. In Biofortification of Staple Crops; Singapore: Singapore, 2022; pp. 147–174. [Google Scholar]

Table 1.
The nutritional composition of the edible portion of maize in dry form.
| Nutritional content | Amount | Nutritional content | Amount |
|---|---|---|---|
| Phosphorus | 348 | Calcium | 10 |
| Calories | 342 | Fat | 3.6 |
| Potassium | 286 | Fibre | 2.7 |
| Magnesium | 139 | Iron | 2.3 |
| Sulphur | 114 | Amino Acids | 1.78 |
| Carotene | 90 | Minerals | 1.5 |
| Carbohydrates (g) | 66.2 | Thiamine | 0.42 |
| Sodium | 15.9 | Copper | 0.14 |
| Moisture (g) | 14.9 | Vitamin c | 0.12 |
| Protein | 11.1 | Riboflavin | 0.1 |
Table 2.
Comparative studies on nutritional status in transgenic and commercially grown varieties.
| Transgenic lines | Nutrient evaluation | Findings | References |
|---|---|---|---|
| Roundup Ready (RR) contain gene for glyphosate resistance | Crude protein, crude fibre, fatty acid, starch, ash, sugar, fat, amino acids, and macroelement | RR is equivalent in nutrition to isogenic corn | [56] |
| PR33P67 and Pegaso Bt: containing “Cry toxin” gene | fatty acids, vitamin C, antioxidants, total phenols, carotenoids, polyphenols, and mineral composition | Transgenic varieties are nutritionally equivalent to conventional varieties | [57] |
| TC1507: Insect-resistant and herbicide-tolerant | Fatty acid, vitamins, Minerals, amino acids, secondary metabolites, and anti-nutrients | Transgenic variety nutritionally comparable to conventional variety | [58] |
| Genetically modified six-event stack maize: Bt11 x MIR162 x MIR604 x 1507 x 5307 x GA21 | Folic acid, Potassium, zinc, arachidic acid, b-carotene, ferulic acid, and methionine | Nutritionally comparable to its non-GM corn | [59] |
Table 3.
Agronomic biofortification of Selenium (Se) and Zinc (Zn) impact on maize yield and grain concentration.
Table 3.
Agronomic biofortification of Selenium (Se) and Zinc (Zn) impact on maize yield and grain concentration.
| Nutrient | Method of Application | Rate of Application | % Increase in Grain | Yield increase (%) | References |
|---|---|---|---|---|---|
| Se | Basal | 5-20g ha-1 Na2SeO4 | 25–227 | - | [68] |
| Foliar | 423–819 | - | |||
| Zn | Basal | Elemental Zn (30 kg ha−1) | 15 | 11 | [69] |
| ZnO (8 kg Zn ha−1) | 59 | 44 | |||
| 28 | 11 | ||||
| Foliar | ZnO nanoparticle (2% solution) | 82 | 33 | [70] | |
| ZnO (2% solution) | 38 | 11 |
Table 4.
Description of Biofortified varieties of Maize released in India.
| Variety | Nutrition improved | Developed by | Year of Release |
|---|---|---|---|
| Vivek QPM 9 | Lys content: 4.19% of protein. Trp content: 0.83% of protein | ICAR-Vivekananda Parvatiya Krishi Anusandhan Sansthan, Almora | 2008 |
| Pusa HM4 Improved | Lys content: 3.62 % of protein. Trp content: 0.91% of protein | ICAR-IARI, New Delhi | 2017 |
| Pusa HM8 | Lys content: 4.18 % of protein. Trp content: 1.06 % of protein | ICAR-IARI, New Delhi | 2017 |
| Pusa HM9 Improved | Lys content: 2.97 % of protein. Trp content: 0.68 % of protein | ICAR-IARI, New Delhi | 2017 |
| Pusa Vivek QPM9 Improved (India’s first PrVit-A rich corn) | PrVit-A: 8.15 ppm. Lys content: 2.67 % of protein. Trp content: 0.74 % of protein | ICAR-IARI, New Delhi | 2017 |
| Pusa VH 27 Improved | PrVit-A: 5.49 ppm | ICAR-IARI, New Delhi | 2020 |
| Pusa HQPM 5 Improved | PrVit-A: 6.77 ppm. Lys content: 4.25 % of protein. Trp content: 0.94 % of protein | ICAR-IARI, New Delhi | 2020 |
| Pusa HQPM 7 Improved | PrVit-A: 7.10 ppm. Lys content: 4.19 % of protein. Trp content: 0.93 % of protein | ICAR-IARI, New Delhi | 2020 |
| IQMH 201 (LQMH 1) | Lys content: 3.03 % of protein. Trp content: 0.73 % of protein | ICAR-Indian Institute of Maize Research, Ludhiana | 2020 |
| IQMH 202 (LQMH 2) | Lys content: 3.04 % of protein. Trp content: 0.66 % of protein | ICAR-Indian Institute of Maize Research, Ludhiana | 2020 |
| IQMH 203 (LQMH 3) | Lys content: 3.48 % of protein. Trp content: 0.77 % of protein | ICAR-Indian Institute of Maize Research, Ludhiana | 2020 |
Table 5.
Maize's essential genes successfully cloned during domestication.
| Gene | Traits | References |
|---|---|---|
| gt1 | Plant architecture | [121,122] |
| ids1/Ts6 | Kernel row number | [123] |
| ra1 | Inflorescence architecture | [124] |
| tb1 | Plant architecture | [125,108] |
| tga1 | Hardened fruitcase | [126] |
| tru1 | Plant architecture | [127] |
| UB3 | Kernel row number | |
| ZmSh1-5.1+ZmSh1-5.2; ZmSh1-1 | Shattering | [128] |
| ZmSWEET4c | Seed filling | [129] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



