
Submitted:
29 September 2023
Posted:
29 September 2023
You are already at the latest version
Abstract
Hemilauxania parvula sp. nov., a new fossil species of the family Lauxaniidae (Diptera: Acalyptratae), is described and illustrated from Oise amber, France (Eocene, lower Ypresian, ca 53 Ma) and its relationship is discussed. Inasmuch as this first finding of a member of Schizophora in Oise amber probably represents the oldest known record of this group of Diptera, the age of Schizophora, based on the known fossil records, is discussed.
Keywords:
extinct lauxaniid flies
; age of Acalyptratae
; Paleogene
1. Introduction
The recent Lauxaniidae are a rich and diversified group of Diptera Acalyptratae [1]. However, they are scarce in the fossil record. Only two genera and two species have been described from Baltic amber (Eocene, 48–34 Ma), viz. Chamaelauxania succini Hennig, 1965 and Hemilauxania incurviseta Hennig, 1965 [2] but there is a series of lauxaniid specimens of the same origin in collections probably containing also unnamed species, see [3] and [4]. Hong [5] described Trypaneiodes ellipticus from the Eocene (Lutetian, 41.2–47.8 Ma) Fushun amber (China) and included it to Lauxaniidae which he subsequently transferred to a new genus Eolausanites Hong, 2001 under the family „Lausaniidae“, see [6] (p. 412). Both these names were subsequently corrected to Eolauxanites and Lauxaniidae in an atlas by Hong [7] (p. 206) where also photos of the holotype E. ellipticus (Hong, 1981) are presented. However, the affiliation of this fossil taxon to Lauxaniidae is not certain. According to Gaimari and Miller [1] (p. 1764), at least 15 undescribed species of Lauxaniidae are known from the early Miocene Dominican amber (Burdigalian, 16–18 Ma) but they all belong to extant New World genera. Also two compression fossils of Lauxaniidae from the Oligocene are described in literature, one from USA, Colorado, viz. Sapromyza veterana Melander, 1949 [8], the other from Canada, British Columbia, viz. Lonchaea senescens Scudder, 1877 (originally placed in Lonchaeidae [9] but see [10,11]) but their taxonomic status and systematic classification are in need of revision. Consequently, all known fossil Lauxaniidae are from the Paleogene and none of them is older than 48 Ma.
A new fossil species of Lauxaniidae (Figure 1), described below, belongs to the genus Hemilauxania Hennig, 1965, being formerly only known from Baltic amber [2,3]. It was found in a sample of Oise amber (France). The fauna and flora of the amber from Le Quesnoy at Oise department, Paris Basin, France is relatively well documented. Amber from Le Quesnoy, usually addressed as Oise amber, is of lowermost Eocene age (Sparnacian, lower Ypresian, basal Eocene, 53 Ma). An inventory list of taxa described from Oise amber was given by Brasero et al. [12]. All main arthropod orders known from other Eocene amber deposits like that from the Baltic Bay of Gdańsk are present in this list except for scorpions and myriapods. Diptera were represented by only 12 species belonging to 6 families, but, subsequently, a series of further taxa from several families have been added [13,14,15,16,17,18,19,20,21,22]. Currently, more than 50 species belonging to 14 families of Diptera are known from Oise amber: nematocerous Bibionidae, Anisopodidae, Mycetophilidae, Cecidomyiidae, Keroplatidae (as Lygistorrhinidae), Chironomidae, Psychodidae and Scatopsidae and brachycerous Athericidae, Rhagionidae, Bombyliidae, Mythicomyiidae, Scenopinidae, Hybotidae and Dolichopodidae. Consequently, no representative of higher flies (Schizophora) has hitherto been recorded from this amber deposit.
The oldest records of ancient Schizophora (both Acalyptratae and Calyptratae) have recently been reviewed in this journal by us [23] who considered as such those from the Eocene Baltic amber (48–34 Ma). However, there is an older fossil species of the group, viz. Pareuthychaeta eoindica Grimaldi, 2012, belonging to Ephydroidea [24] and originating from Cambay amber (India) of the Early Eocene (Ypresian, 50–52 Ma) age, see [25,26]. Most recently, the oldest representative of Calyptratae, belonging to Hippoboscidae, viz. Eornithoica grimaldii Nel, Garrouste & Engel, 2023, has been described from a compression fossil found in USA, Colorado, Green River Formation (lower Eocene, Ypresian, 46.2–40.4 Ma) but authors suggested its age to be around 52 Ma based on radiometric timing of the same fossil deposit [27].
The aim of this paper is not only to describe the oldest known species of Lauxaniidae but also to discuss its relationships and palaeohabitat as well as the assumed age of Acalyptratae and Schizophora as a whole.
2. Materials and Methods
2.1. Material
Origin and repository. The amber sample with the Hemilauxania inclusion is currently registered in the Hoffeins' collection under inventory number CCHH #729-11, to be deposited at the Senckenberg Deutsches Entomologisches Institut, Müncheberg, Germany (SDEI). The inclusion was a donation to the junior author from an amber friend who purchased a large lot of Oise amber with inclusions at the mineral fair at Sainte-Marie-aux-Mines (France) somewhat in the beginning of the 2000s. The very first collector of this amber from Oise is unknown [28].
2.2. Preparation of Amber Specimen
The amber with inclusion was cut, ground and polished and subsequently embedded in Polyester resin for conservation purpose following the method described in [29]. However, for a better view and photography the artificial resin has been removed from one side of preparatum (Figures 2A,B) and this side of the amber has been cleaned and polished during the examination process (Figure 2C). After the study of the sample is finished, the preparatum will again be completed with a layer of artificial resin on this side.
2.3. Techniques of Investigation
The amber inclusion has been examined, drawn and measured using two types of binocular stereoscopic microscopes (Reichert, Olympus). The specimen was photographed by means of a Canon EOS 60D digital camera with macro lens Canon MPE 65 mm 1–5× and, particularly, by a Canon EOS 5D Mark III digital camera with a Nikon CFI Plan 4× /0.10NA 30 mm WD objective attached to a Canon EF 70–200 mm f/4L USM zoom lens. During photography by the latter equipment the specimen was repositioned upwards between each exposure using a Cognisys StackShot Macro Rail and the final photograph was compiled from multiple layers (35–40) using Helicon Focus Pro 7.0.2. The final images were edited in Adobe Photoshop CS6. Other illustrations were drawn on the basis of macrophotographs in which details were inked based on direct observation at higher magnification using a binocular microscope. Description of the species is prepared following the sequence of morphological structures and characters as developed for fossil acalyptrates by [30] and [23]. Measurements: Six characteristics were measured – body length (measured from anterior margin of head to end of cercus, thus excluding the antenna), wing length (from wing base to wing tip), wing width (maximum width), index Cs3:Cs4 (=ratio of length of 3rd costal sector:length of 4th costal sector), index r-m\dm-cu:dm-cu (=ratio of length of section between r-m and dm-cu on discal cell:length of dm-cu) and index r-m\dm-cu:CuA1 (=ratio of length of section between r-m and dm-cu on discal cell:length of apical portion of CuA1).
2.4. Morphological Terminology
Follows that used in [23] for ancient Clusiomitidae to be in continuation with [30] on fossil Anthomyzidae. Morphological terms of the female abdomen are depicted in Figure 5 and Figure 8. The synonymous morphological terms of adult structures and their abbreviations as used in the most recent manual of Afrotropical Diptera [31] are given in parentheses in the list of abbreviations (used in text and/or figures) below.
A1–first branch of anal vein (= CuA+CuP)
A2–second branch of anal vein (= A1)
ac–acrostichal (setulae)
bm–basal medial cell
C–costa
ce–cercus
Cs2, Cs3, Cs4–2nd, 3rd, 4th costal sector
CuA1–cubitus (=M4)
cup–posterior cubital cell (= cua)
cx1, cx2, cx3–fore, mid, hind coxa
dc–dorsocentral setae
dm–discal medial cell
dm-cu–discal medial-cubital (= dm-m, posterior, tp) cross-vein
f1, f2, f3–fore, mid, hind femur
frt–frontal triangle
h–humeral cross-vein
hu–humeral (= postpronotal) (seta)
ia–posterior intra-alar
M–media (= M1)
mspl–mesopleural (= anepisternal) (seta)
npl–notopleural (seta)
oc–ocellar (seta)
ori–lower fronto-orbital (seta)
ors–upper fronto-orbital (seta)
pa–postalar (seta)
ppl–propleural (= proepisternal) (seta)
pra–pre-alar (= anterior supra-alar)(seta)
prs–presutural (= presutural intra-alar) (seta)
pvt–postvertical (= poc, postocellar) (seta)
R1–1st branch of radius
R2+3–2nd branch of radius
R4+5–3rd branch of radius
r-m–radial-medial (= anterior, ta) cross-vein
S1–S10–abdominal sterna
sa–supraalar (seta)
sc–scutellar (seta)
Sc–subcosta
sctl–scutellum
stpl–sternopleural (= katepisternal) (seta)
T1–T10–abdominal terga
t1, t2, t3–fore, mid, hind tibia
vte–outer vertical (seta)
vti–inner vertical (seta)
3. Results
3.1. Systematic Palaeontology
Class Insecta Linnaeus, 1758.
Order Diptera Linnaeus, 1758
Cyclorrhapha Brauer, 1863
Schizophora Becher, 1882
Acalyptratae Macquart, 1835
Superfamily Lauxanioidea Macquart, 1835
Family Lauxaniidae Macquart, 1835
Genus Hemilauxania Hennig, 1965
3.2. Hemilauxania parvula sp. nov. (Figure 1 and Figure 3–8)
LSID
Type material. Holotype female, labelled: “Faszination Bernstein, Christel Hoffeins, Hans Werner Hoffeins” (framed on obverse), ”729-11, Diptera, Acalyptratae, Lauxaniidae” (handwritten by C. Hoffeins, on reverse), ”Oise amber” (handwritten on pale green label) and “Holotypus ♀, Hemilauxania parvula sp.n., J. Roháček & C. Hoffeins det. 2023” (red label), currently deposited Hoffeins' collection, inventory no. 729-11. A female in good preservation, embedded in a multi-layered piece of amber of light orange colour; inclusion almost complete, tibiae and tarsi of fore legs and tarsus of one mid leg cut-off, head very close to the surface, face, antennae and anterior part of eyes destroyed, apex of the right wing cut-off. [block-shaped /prism with quadrangular base/ amber piece ca 7.3 × 5.1 × 3.4 mm, embedded in polyester resin, actual size 11.2 × 9.4 × 6 mm] (Figure 2); syninclusions: fragment of Chironomidae, female. The holotype will be subsequently deposited in SDEI.
Type locality: France: Paris Basin, Oise department, Le Quesnoy.
Horizon and age: Sparnacian, lower Ypresian, basal Eocene, ca 53 Ma.
Etymology: The new species is named “parvula” (= small, minor, from Latin, adjective) to refer to its small body size.
Diagnosis. The new species differs from the only other representative of the genus Hemilauxania, viz. H. incurviseta Hennig, 1965, by a distinctly smaller size (2.7 mm vs 3.5–5 mm), laterally yellow mesonotum and yellow scutellum, frons finely striated between orbits and frontal triangle, pvt convergent but not crossed, oc shorter, postocular setulae in single row, only 1 robust mspl seta, f1 with 2 dorsal setae in distal half, f2 with 3 anterodorsal setae near the middle, f3 with no anterodorsal setae, t3 lacking a group of enlarged anterodorsal setae, C with spinulae ending in front of apex of R2+3, costal and subcostal cell distinctly wider, and female T7 with lateral parts extended onto ventral side of postabdomen.
Description: Male unknown. Female. Total body length about 2.7 mm; general colour apparently bicolourous but mostly brown, with only some parts of body ochreous to yellow; legs pale brown to yellow; thorax and abdominal sclerites microtomentose and relatively dull (Figure 1 and Figure 3).
Head (Figure 3 and Figure 4) slightly higher than long, dorsally about as broad as thorax (not precisely measurable due to damaged eyes); dorsal part of occiput very slightly concave. Head largely brown (almost entire frons, occiput darkest), only some small anterior and ventral parts yellow. Frons about as long as wide and as wide as eye in dorsal view, very slightly tapering anteriorly, largely brown, with only very anterior part of orbit and (probably) anterior margin of frons yellowish ochreous. Orbit delimited by a distinctly striated area separated medially by the frontal triangle. Frontal triangle relatively narrow (but wider than ocellar triangle), brown, with plain and dull surface, with anterior corner poorly visible, probably almost reaching anterior margin of frons. Ocellar triangle not clearly delimited (margined) but distinctly protruding; ocelli medium-sized. Anterior part of head, including frontal lunule, face (praefrons), parafacialia and both antennae damaged, and, therefore, not described. Gena also obscured; postgena pale brown, relatively small, not expanded, with several small setae at posterior margin. Cephalic chaetotaxy (Figure 3 and Figure 4): pvt distinct but shorter than oc, strongly convergent but not crossed; vti, vte and posterior ors relatively strong, subequal (longest cephalic setae); oc somewhat shorter and weaker, inserted between ocelli close to each other; 3 (on right) or 4 (on left) strong fronto-orbital setae, composed of 2 or 3 reclinate ors and 1 inclinate ori (sensu Hennig 1965), the hindmost ors only slightly longer and the foremost ors smaller than others (Figure 4); no microsetulae on frons medially or in front of ors but 3 microsetae inserted between ocelli on ocellar triangle; postocular setulae numerous, in single long row surrounding posterior eye margin and ranging from vti to postgena. Setosity of damaged anteroventral part of head not visible. Eye relatively large, bare, convex, suboval to subcircular, with longest diameter somewhat oblique and only 1.1–1.2 times as long as shortest diameter. Gena probably low (damaged, not precisely visible); palpus not preserved in the fossil. Mouthparts also damaged, distorted and poorly preserved, yellow. Both antennae heavily damaged, with pedicel and 1st flagellomere (including arista) lost. Only scape partly visible, small, short and yellow, with series of 6 or 7 microsetulae at distal margin dorsally.
Thorax hardly wider than head, bicolourous but largely brown, with only some parts paler, ochreous to yellow (Figure 1 and Figure 3). Mesonotum mostly brown, darker medially and posteriorly, but anteromedially with 3 short dark stripes separated by yellow spots (see Figure 3); also lateral parts of mesonotum and scutellum yellow to pale ochreous, subscutellum brown. Humeral (postpronotal) callus narrowly brown only anteriorly, otherwise yellow; also notopleural area pale brown anteriorly, yellow posteriorly; pleural part of thorax with all sclerites brown (Figure 1). Scutellum rounded trapezoid, wider than long, slightly convex dorsally; subscutellum poorly visible but probably not protruding. Thoracic chaetotaxy (Figure 4 and Figure 5): 1 distinct (as long as posterior npl) hu, plus several microsetae on humeral callus; 2 npl, anterior strong, about 1.5 times as long as posterior; 1 prs (presutural intra-alar), 1 pra (pre-alar = anterior supra-alar) and 1 sa, all well developed but thin; 1 distinct but thin ia (posterior intra-alar); 2 pa, anterior robust, posterior weak; 3 (on right) or 4 (on left) dc, all postsutural and becoming shorter anteriorly; the hindmost dc long and robust (almost as long as sc) and shifted more laterally than others; scutum otherwise covered by uniform and relatively dense microsetae (more than 15 dc microsetae in row in front of anterior dc); ac microsetae also dense, arranged in 8–10 irregular rows in front of suture, in about 6 rows at level of posterior dc; prescutellar ac strong and as long as middle dc and having a pair of ac microsetae between them; 2 strong and long sc (longest thoracic setae), subequal in length; apical sc distinctly crossed, also laterobasal sc more or less inclinate; scutellum with 2 very minute microsetae at margin between laterobasal and apical sc but none on disc (Figure 4); 1 short and fine upcurved ppl (Figure 6A); mesopleuron with only 1 robust but relatively short mspl plus 2 weak setae above and 1 below it at posterior margin (Figure 6A), otherwise with a series of microsetae in posterodorsal half of sclerite; sternopleuron with genus-specific chaetotaxy, i.e. with 3 strong and long stpl at dorsal margin of posterior half and with a number of scattered microsetae on most of surface (including also 2 longer fine setulae on ventral corner).
Legs distinctly paler than thorax (Figure 1 and Figure 7), pale brown to yellow, all relatively robust. Fore and mid legs largely dirty yellow, with some brownish tinge on coxae and femora; hind leg pale brown, with yellowish trochanter and tarsus. Both fore legs with tibia and tarsus missing. cx1 with 5 longer fine setae at ventral margin distally; also cx2 with a tuft (or short row) of longer fine setae; cx3 with only short setulae. f1 (Figure 6C) with 2 dorsal setae in distal half, 6 posterodorsal setae in proximal half and a row of 6 or 7 fine posteroventral setae along entire length, all these setae relatively short; f2 (Figure 6D) longer and more robust than f1, finely densely setulose but with 3 short but distinct anterodorsal setae about the middle, also 2 subapical posteroventral setae somewhat enlarged; f3 (Figure 6E) yet more stout than f2, also uniformly densely finely setulose but with 1 robust posteroventral seta in distal sixth, surrounded by 2 thicker but short setae. t1 lost, undescribed, but probably with 1 short dorsopreapical seta as known in H. incurviseta Hennig, 1965 (see [2], fig. 147]; t2 (Figure 6D) besides dense fine setulae with 1 short dorsopreapical seta and 1 robust and long (longer than width of tibia) ventroapical seta; t3 (Figure 6E) also with 1 small dorsopreapical seta and with 1 (yet shorter) ventroapical seta, otherwise uniformly finely setulose. Tarsi simple and short; mid and hind basitarsi (fore tarsi missing) almost as long as other tarsal segments together (see Figure 7); claws relatively small.
Wing (Figure 1, Figure 5 and Figure 6B) proximally widest, distally tapered; veins ochreous; membrane uniformly pale ochreous because densely microtrichose. C reaching to apex of M, besides microsetae provided with series of small spines reaching from beyond apex of humeral cross-vein almost to apex of R2+3 (Figure 6B). No costal break. Sc complete, well developed, separate along its entire length, distally ending into C far from apex of R1. R1 of medium length, bare, ending into C in basal third of wing. Both costal and subcostal cell relatively broad (Figure 5). R2+3 long, very slightly sinuate, apically somewhat upcurved to C and ending farther from wing apex than M. R4+5 shallowly bent posteriorly, distally subparallel with M (see Figure 6B). Distal part of M also slightly bent posteriorly and ending in C. Cell dm long, gradually slightly widened distally; its upper distal corner rectangular; anterior cross-vein (r-m) situated in the middle of cell dm. Distal part of CuA1 slightly (right wing) to distinctly (left wing) shorter than dm-cu cross-vein and almost reaching wing margin. Cells bm and cup closed. A1 very shortened, almost stump-like. A2 distinctly developed, markedly longer than A1 but not reaching wing margin (Figure 5). Anal lobe well developed. Alula small and narrow. Wing measurements: length ca 2.4 mm, width 0.95 mm, Cs3:Cs4 = ca 1.2, r-m\dm-cu:dm-cu = 2.21–2.44, r-m\dm-cu:CuA1 = 1.12–1.32. Haltere (Figures 1 and 5) dirty yellow, knob relatively large, rounded.
Abdomen broad, subovoid in ventral view (Figure 7 and Figure 8), posteriorly more tapered. Preabdominal terga short and strongly transverse, all brown and relatively densely but shortly setose, with longest setae at posterior margins (Figure 5). Preabdominal sterna (but only S4 and S5 visible, Figure 8) markedly narrower than associated terga but also short and transverse, sparsely shortly setose. T1 pale brown and narrower than T2, more or less distinctly separate from the latter; T2–T5 darker brown; T2 somewhat widened posteriorly and about as long as T3; T3 widest tergum, T4 of subequal length but slightly narrower; T5 slightly shorter and narrower than T4, distinctly tapered posteriorly, with posterior corners rounded. S1–S3 not visible but probably narrower than S4. S4 and S5 brown, of about the same width but S4 distinctly longer (Figure 8).
Postabdomen. Terga only partly visible in lateral (Figure 5) and ventral view (Figure 8). T6–T8 brown. T6 similarly transverse but somewhat shorter and narrower than T5, posteriorly tapered. T7 dorsally distinctly shorter and narrower than T6 but laterally extended to reach on ventral side almost to S7 (see Figure 8), dorsally similarly setose as T6. T8 much smaller than T7, simply transverse, with sides not projecting ventrally. T10 (supra-anal plate) not clearly visible, probably small and pale. Ventral side of postabdomen as in Figure 8. S6 short and very transverse, distinctly shorter than S5 but also brown and with sparse short setae. S7 pale ochreous, narrow (only half width of S6), roughly trapezoidal (anteriorly narrower), laterally rounded. S8 also pale but yet narrower (half width of S7), only slightly wider than long, suboblong. Both S7 and S8 sparsely shortly setose. S10 (subanal plate) small, short transversely subtriangular, with a group of very short dense setulae medially. Cerci (Figure 8) visible as slender, elongately subconical and pale appendages, with a few very short setulae.
Comments. Hemilauxania parvula sp. nov. is the second species of the formerly monotypic genus Hemilauxania Hennig, 1965. However, there are a few more unnamed species of the genus in collections [3,4], all from Baltic amber. The new species, described above, although with anterior part of head and fore legs damaged, undoubtedly belong to Hemilauxania sharing almost all generic characters presented by Hennig [2]. It seems to be closely allied to H. incurviseta, the type species of the genus, but differs from it not only by characters listed above in the Diagnosis, but also by further features, e.g. in having a few microsetae between ocelli, mesonotum anteromedially with distinctive brown and yellow pattern (Figure 3), mesonotal microsetae (acrostichal, dorsocentral and others) distinctly more numerous and denser, scutellum without microsetae on disc but with 2 marginal ones between sc setae (Figure 4), f3 more robust than f2 and legs with markedly shorter dorsopreapical setae on tibiae (Figures 6D,E). Hennig [2] did not see vein A2 in H. incurviseta as being so distinct in H. parvula (Figures 1, 5) but it could be obscured and invisible in the two specimens he examined.
Some variability in setae of head and thorax of the holotype of H. parvula should also be commented on here. This specimen has 4 fronto-orbital (3 ors + 1 ori) setae on the left side of head and only 3 (2 ors + 1 ori) on the right side. Similarly, there are 4 postsutural dc macrosetae on the left side of mesonotum and only 3 on the right side (see Figure 4). Comparing this with chaetotaxy found in H. incurviseta, see [2] (figs 138–140, 142, 144) it is most plausible that 1 ors has been occasionally lost on the right orbital plate of the H. parvula holotype (thus, that 3 ors is the normal number) while 1 dc (the foremost) seta seems to be supernumerary on the left side of the mesonotum of that specimen and normally there should only be 3 dc setae.
4. Discussion
The discovery of a new lauxaniid fly in the Oise amber and its affiliation to the genus Hemilauxania Hennig lead us to discuss several topics. The general rarity of Lauxaniidae in the fossil record (for review of a few published findings see Introduction) is reflected in very insufficient knowledge of the evolution of this currently very diverse family of Diptera Acalyptratae. If we ignore the more or less dubious published data (see [5,6,7,8,9]) the only reliable ancient taxa of Lauxaniidae are those based on inclusions in Baltic amber. Hennig [2,32] investigated only 5 specimens, two males and one female of Chamaelauxania succini Hennig, 1965 and two females of Hemilauxania incurviseta. In the amber collection of Hoffeins (Hamburg) 28 specimens of Lauxaniidae are actually housed in a total of more than 1,100 acalyptrate fossil flies, of which 7 are placed in Chamaelauxania and 14 in Hemilauxania whereas 3 cannot be identified to species level because of poor or incomplete preservation and 4 may represent an undescribed taxon [4].
4.1. Relationships and classification of Hemilauxania
According to [2] Hemilauxania differs from all recent members of Lauxaniidae in having 4 fronto-orbital (including 1 inclinate ori) setae while only 2 (more rarely 1) ors, sometimes including 1 ori, are present in extant taxa of the family [1,10]. Hennig [33] (p. 33), however, considered the more numerous fronto-orbital setae to belong to ground plan of Lauxanioidea because 3 (albeit small to hair-like) fronto-orbitals also occur in the closely related family Celyphidae. Thus, the reduced number of fronto-orbitals can be considered a synapomorphy of recent Lauxaniidae.
The presence of 3 strong sternopleural setae is another distinctive difference against all recent Lauxaniidae (which possess only 1 or 2 stpl) and this character seems to be shared with the other Baltic amber fossil genus, Chamaelauxania Hennig, 1965 having these setae yet more numerous (5 strong stpl), see [2] (p. 107, fig. 131). It is peculiar that Hennig [2,33] has not discussed this important feature as diagnostic for both his fossil genera but, instead, he stressed the presence of 1 pre-alar (= anterior supra-alar) seta in Hemilauxania as a unique character within all known Lauxaniidae. Similarly, the presence of a well-developed vein A2 was not recorded by him (in both Chamaelauxania and Hemilauxania) although this vein occurs also on wings of recent Lauxaniidae, cf. [34,35]. It should be remarked that the presence of A2 in (many) extant genera of Lauxaniidae has not been listed (neglected or overlooked?) in most Diptera manuals, see [1,10,36].
Although undoubtedly monophyletic, the suprageneric classification of Lauxaniidae ‘is in its infancy’ [1]. As stated already by Hennig [2] Hemilauxania and Chamaelauxania have no distinct relatives among recent genera of Lauxaniidae and cannot be affiliated to any of the currently recognized subfamilies, viz. Homoneurinae or Lauxaniinae. Judging from ancestral cephalic chaetotaxy with 3 or 4 fronto-orbital setae (with distinct inclinate ori) and cup cell with a slightly or not concave distal margin (= cross-vein CuA2) (see also [2]) Hemilauxania should be treated as an extinct lineage of the family, branching very basally from the common stem of Lauxaniidae. The systematic position of Chamaelauxania remains unclear; it could either be related to Hemilauxania or represents another separate lineage.
4.2. Palaeohabitat and palaeobiology of Hemilauxania
The family Lauxaniidae has almost cosmopolitan distribution, most of the species occurring in tropical regions of Asia and the Americas and species diversity declines strongly towards the more temperate regions. Most species inhabit humid forests, where the adults are usually found sitting on leaves of the understory. Larvae are saprophagous and develop in rooting wood and, particularly, leaf-detritus [10,35,37].
The Eocene Baltic amber and the contemporaneous Rovno amber from the Ukraine, were formed in a warm-temperate ('subtropical summerwet') palaeoenvironment from resin produced mainly by conifers in the so-called 'Baltic amber forest'. Alekseev & Alekseev [38] described this palaeohabitat as a thermophilic, humid-mixed forest resembling the recent subtropical forests in eastern and southeastern Asia. However, other authors found that various forest types evolved in this area [39,40,41]. The vegetation obviously had first (in the early Eocene) a tropical character that gradually changed to subtropical and later to warm-temperate due subsequent cooling. On the other hand, the older and more southerly situated (at 42–47°N latitude) Oise amber forest was growing close to the early Eocene warming maximum (ca 49 Ma) and, therefore, plausibly had a tropical character; moreover, the Oise amber had an angiosperm origin, because it was formed from resin produced by the ancient tree Aulacoxylon sparnacense Combes, 1907 (Combretaceae or Caesalpiniaceae or Fabaceae), see [42,43]. Nel & Brasero [42] wrote about this palaeohabitat: "In Oise, the dominance of an arborescent amber-producing tree and the presence of freshwater suggest a semi-deciduous forest, with a mosaic of gallery-forest mixed with dryer plant communities, in a deltaic 'paratropical' region."
However, the palaeohabitats with riparian forests and deciduous trees under high humidity conditions, most suitable for development of Lauxaniidae, were documented both for the Oise [12,42] and the Baltic amber [41] (Sadowski 2020). Consequently, the presence of Hemilauxania species in amber from both these deposits is not so surprising, particularly when some (or all) Baltic amber samples with Hemilauxania inclusions could be older than currently dated (see below). Nevertheless, the occurrence of Hemilauxania species in Ukrainian Rovno amber can be predicted with high probability.
4.3. Age of Hemilauxania and other oldest fossils in Schizophora
The new Hemilauxania species originates from a stratum of the Oise or Le Quesnoy amber being of the Sparnacian, lower Ypresian, basal Eocene age (ca 53 Ma). Factually, it represents the oldest known species not only of Acalyptratae but also of the whole Schizophora. The previously oldest records of schizophoran flies were also lower Ypresian Pareuthychaeta eoindica Grimaldi, 2012 (Ephydroidea) from Cambay amber (India) dated 50–52 Ma [24] and Eornithoica grimaldii Nel, Garrouste & Engel, 2023 (Calyptratae: Hippoboscidae), a compression fossil from the Green River Formation (USA, Colorado) dated about 52 Ma [27].
In the geological timescale the Ypresian is the oldest period or lowest stratigraphic stage of the Eocene. It spans the time between 56 and 47.8 Ma, and is followed by the Eocene Lutetian Age. Besides the above ambers and shales, also the Fur Formation in Denmark and the Messel shales in Germany are of the same Ypresian age.
However, Hemilauxania incurviseta was found in the Baltic amber [2] whose deposit and age must also be considered. Baltic amber was deposited in marine sediments in the Eocene epoch. Amber bearing layers were sedimented in the Lower Blue Earth (Ypresian), the Wild Earth and Blue Earth (Lutetian) and the Green Wall (Bartonian) horizon. The Lutetian Blue Earth is the main amber horizon but also the Lower Blue Earth and the Wild Earth contains scattered amber [44]. The Lower Blue Earth is an amber bearing layer of Ypresian age, 54.5–49 Ma.
The age of these Baltic sediments covers a range of roughly 15 Ma. Amber found in these sediments was redeposited in Eocene marine sediments as well as Neogene fluvial and glacigenic deposits [45], consequently, it is never found in its original stratigraphic position. Moreover, the stratigraphic age does not correlate with the ‘real’ age of the resin which was transformed into amber. If an individual amber nugget is obtained, nothing can be stated about which strata it may have come from, if Ypresian, Lutetian or Bartonian sediments. We just can state the age of the sediments ranging between 54.5 Ma to 37 Ma. Amber as an exudate of Eocene conifers must have been produced somewhat before it was sedimented, this means it is in fact older than the sediment itself. Consequently, although the main amber bearing layer from Baltic area is of Lutetian age, we cannot exclude a possibility that H. incurviseta and/or other hitherto undescribed Hemilauxania specimens of the same origin also are of Ypresian age as is H. parvula from Oise amber.
The origin of Schizophora (and also Acalyptratae) is estimated by Grimaldi & Engel [46] to occur immediately after the K-T (K-Pg) boundary, being associated with the Cretaceous–Paleogene extinction event dated ca 66 Ma [47]. Based on molecular-based time-calibrated phylogeny Wiegmann et al. [48] reached almost the same conclusion, dating the Schizophora split off at 65 Ma. Considering the age of the oldest known fossils of schizophoran flies (ca 53 Ma, see above) we can agree with the latter dating of the origin of Schizophora because 1 million years after the Chicxulub bolide impact causing the massive K-Pg extinction of fauna the atmospheric conditions had already been stabilized [47] to enable development of a new group of Diptera.
Author Contributions
Conceptualization, J.R.; methodology, J.R. and C.H.; investigation, J.R. and C.H.; writing—original draft preparation, J.R and C.H..; writing—review and editing, J.R. and C.H.; illustrations and visualization, J.R.; funding acquisition, J.R. All authors have read and agreed to the published version of the manuscript.
Funding
The research of J.R. was supported by the Ministry of Culture of the Czech Republic by institutional financing of long-term conceptual development of the research institution (the Silesian Museum, MK000100595).
Data Availability Statement
The data presented in this study are available in this article.
Acknowledgments
C.H. is grateful to Rainer Ohlhoff (Saarbrücken, Germany) for donation of the Oise amber with the Lauxaniidae inclusion and to her husband for careful preparation of this rare amber specimen. We would like to thank also to P.J. Chandler (Melksham, England, U.K.) for critical comments and valuable help with the English presentation of the paper.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Gaimari, S.D.; Miller, R.M. Lauxaniidae (Lauxaniidae Flies). In Manual of Afrotropical Diptera; Kirk-Spriggs, A.H, Sinclair, B.J., Eds.; South African National Biodiversity Institute: Pretoria, South Africa, 2021; Volume 3, Brachycera–Cyclorrhapha, pp. 1757–1781.
- Hennig, W. Die Acalyptratae des Baltischen Bernsteins und ihre Bedeutung für die Erforschung der phylogenetischen Entwicklung dieser Dipteren-Gruppe. Stuttg. Beiträge Nat. 1965, 145, 1–215.
- Von Tschirnhaus, M.; Hoffeins, C. Fossil flies in Baltic amber—Insight in the diversity of Tertiary Acalyptratae (Diptera, Schizophora), with new morphological characters and a key based on 1000 collected inclusions. Denisia 2009, 26, 171–212.
- Hoffeins, C. (Hamburg, Germany). Personal communication, 2023.
- Hong, Y.-C. Eocene fossil Diptera Insecta in amber of Fushun coalfield. Geological Publishing House: Beijing, China, 1981; pp. 1–166 + plates 1–27 (in Chinese only).
- Hong, Y.-C. Amber insects of China. Huayu Nature Book Trade Co. Ltd: Beijing, China, 2002, pp. 1–653 + plates 1–48 (in Chinese only).
- Hong, Y.-C. Atlas of amber insects of China. Henan Scientific and Technological Publishing House: Zhengzhou, China, 2002, pp. 1–394 (in Chinese only).
- Melander, A.L. A report on some Miocene Diptera from Florissant, Colorado. Am. Mus. Novit. 1949, 1407, 1–63. http://npshistory.com/publications/flfo/amn-n1407-1949.pdf.
- Scudder, S.H. The insects of the Tertiary beds at Quesnel (British Columbia). Rep. Prog. Geol. Surv. Terr. 1877, 1875–1876, 266–280. https://publications.gc.ca/collections/collection_2019/rncan-nrcan/M41-1-1-1876-eng.pdf.
- Shewell, G.E.; 87. Lauxaniidae. In Manual of Nearctic Diptera. McAlpine J. F., Peterson, B.V., Shewell, G.E., Teskey, H.J., Vockeroth, J.R., Wood, D.M., Eds.; Minister of Supply and Services: Ottawa, Canada; Volume 2, Agriculture Canada Monograph No. 28, pp. 993–1006.
- Evenhuis, N.L. Catalogue of the fossil flies of the world (Insecta: Diptera). Backhuys Publisher: Leiden, The Netherlands, pp. 1–600.
- Brasero, N.; Nel, A.; Michez, D. Insects from the Early Eocene amber of Oise (France): diversity and palaeontological significance. Denisia 2009, 26, 41–52.
- Doitteau, G.; Nel, A. Chironomid midges from early Eocene amber of France (Diptera: Chironomidae). Zootaxa 2007, 1404, 1–66.
- Blagoderov, V.; Hippa, H.; Nel, A. Parisognoriste, a new genus of Lygistorrhinidae (Diptera, Sciaroidea) from the Oise amber with redescription of Palaeognoriste Meunier. ZooKeys 2010, 50, 79–90. [CrossRef]
- Myskowiak, J.; Nel, A. A new genus and species of ibis fly in the Lowermost Eocene amber of Oise (France) (Diptera: Athericidae). Zootaxa 2014, 3869(4): 372–382. [CrossRef]
- Garrouste, R.; Azar, D.; Nel, A. The oldest accurate record of Scenopinidae in the Lowermost Eocene amber of France (Diptera: Brachycera). Zootaxa 2016, 4093(3): 444–450. [CrossRef]
- Nel, A.; Perreau, Z.; Doitteau, G. The oldest representative of the modern snipe fly genus Symphoromyia (Diptera: Rhagionidae). Zootaxa 2016, 4196(1), 144–150. [CrossRef]
- Bramuzzo, S.; Coty, D.; Nel, A. A new species of Ferneiella from the Eocene French amber (Diptera: Scatopsidae). Zootaxa 2017, 4350(1), 177–184. [CrossRef]
- Bramuzzo, S.; Nel, A. Youngest representative of the extinct genus Microphorites in the Eocene amber of France (Diptera: Dolichopodidae: Microphorinae). Zootaxa 2017, 4231(4), 590–594. [CrossRef]
- Myskowiak, J.; Garrouste, R.; Nel, A. Eodromyia pumilio gen. et sp. nov., a new empidoid fly from the Earliest Eocene amber of France (Diptera: Hybotidae: Tachydromiinae). Zootaxa 2018, 4379(2): 279–286. [CrossRef]
- Camier, M.; Nel, A. First Mycetobiinae of the lowermost Eocene Oise amber (France) (Diptera: Anisopodidae). Palaeoentomology 2019, 2(3), 245–250. [CrossRef]
- Camier, M.; Nel, A. The oldest fungus gnat of the tribe Exechiini in the lowermost Eocene Oise amber (Diptera: Mycetophilidae). Zootaxa 2020, 4722(1): 91–98. [CrossRef]
- Roháček, J.; Hoffeins, C. Clusiomitidae, a new family of Eocene fossil Acalyptratae, with revision of Acartophthalmites Hennig and Clusiomites gen. nov. (Diptera). Insects 2021, 12(12) 1123: 1–52. [CrossRef]
- Grimaldi, D.A.; Singh, H. The extinct genus Pareuthychaeta in Eocene ambers (Diptera: Schizophora: Ephydroidea). Can. Entomologist 2012, 144(1): 17–28.
- Rust, J.; Singh, H.; Rana, R.S.; McCanna, T.; Singh, L.; Anderson, K.; Sarkare, N.; Nascimbene, P.C.; Stebner, F.; Thomas, J.C.; Solorzano Kraemer, M.; Williams, C.J.; Engel, M.S.; Sahni, A.; Grimaldi, D. Biogeographic and evolutionary implications of a diverse paleobiota in amber from the early Eocene of India. Proc. Natn. Acad. Sci. U. S. Am. 2010, 107, 18360–18365.
- Roháček, J.; Hammel, J.U.; Baranov, V. Christelenkidae, a new extinct family based on a new taxon from Eocene Baltic amber (Diptera: Acalyptratae), with X-ray synchrotron microtomography imaging of its structures. Arthropod Syst. Phyl., 2023, 81, 475–498. [CrossRef]
- Nel, A.; Garrouste, R.; Engel, M.S. The earliest Pupipara (Diptera: Hippoboscoidea): A new genus and species from the lower Eocene of the Green River Formation. Palaeoentomology 2023, 6(1): 58–63. [CrossRef]
- Ohlhoff, R. Bernstein aus Frankreich. In Bernstein Magazin, Ausgabe 2022; Ribbecke, H.-W., Ed.; Verein zur Förderung des Geologisch-Paläontologischen Museums der Universität Hamburg e.V. „Arbeitskreis Bernstein“: Hamburg, Germany, 2022; pp. 40–41.
- Hoffeins, H.W. On the preparation and conservation of amber inclusions in artificial resin. Pol. Pismo Entomol. 2001, 70, 215–219.
- Roháček, J. New amber fossil Anthomyzidae (Diptera): An unexpected Eocene diversity. J. Syst. Palaeontol. 2013, 11, 431–473. [CrossRef]
- Cumming, J.M.; Wood, D.M. 2. Adult morphology and terminology. In Manual of Afrotropical Diptera. Introductory chapters and Keys to Diptera Families; Kirk-Spriggs, A.H., Sinclair, B.J., Eds.; Suricata 4—South African National Biodiversity Institute: Pretoria, South Africa, 2017; Volume 1, pp. 89–133.
- Hennig, W. Neue Übersicht über die aus dem Baltischen Bernstein bekannten Acalyptratae (Diptera: Cyclorrhapha). Stuttg. Beiträge Nat. 1969, 209, 1–42.
- Hennig, W. Neue Untersuchungen über die Familien der Diptera Schizophora. Stuttg. Beiträge Nat. 1971, 226, 1–76.
- Papp, L.; Shatalkin, A.I. 3.37. Family Lauxaniidae. In Contribution to a manual of Palaearctic Diptera; Papp, L., Darvas, B., Eds.; Science Herald: Budapest, 1998; Volume 3, pp. 383–400.
- Semelbauer, M. Lauxaniidae – tieňovkovité (Diptrera, Cyclorrhapha). Veda: Bratislava, Slovakia, 2016, pp. 1–183 (in Slovak).
- Gaimari, S.D.; Silva, V.C. 71. Lauxaniidae (Lauxaniid flies). In Manual of Central American Diptera; Brown, B.V.; Borkent, A.; Cumming, J.M.; Wood, D.M.; Woodley, N.E.; Zumbado, M.A., Eds.; NRC Research Press: Ottawa, Canada, 2010; Volume 2, pp. 971–995.
- Miller, R.M. Ecology of Lauxaniidae (Diptera: Acalyptratae) I. Old and new rearing records with biological notes and discussion. Annls Natal Mus. 1977, 23(1), 215–238.
- Alekseev, V.I.; Alekseev, P.I. New approaches for reconstruction of the ecosystem of an Eocene amber forest. Biology Bulletin, 2016, 43, 75–86. https://link.springer.com/article/10.1134/S1062359016010027.
- Szwedo, J. Life in the Eocene forests. In World Amber Council Seminar. Gdańsk the World Amber Capital, Gdańsk (Poland), May 18-19.2012; Pytlos, R., Szadziewski, R., Zbierska, E., Adamska, G., Dmowska, A., Eds.; Mayor’s Office for City Promotion, City Hall of Gdańsk, Gdańsk, Poland, 2012; pp. 56–70.
- Słodkowska, B.; Kramarska, R.; Kasiński, J.R. The Eocene Climatic Optimum and the formation of the Baltic amber deposits. In The international amber researcher symposium Amber. Deposits–collections–the market. Gdańsk (Poland), 22–23.03.2013; Kosmowska-Ceranowicz, B., Gierlowski, W., Sontag, E., Eds.; Gdańsk International Fair Co. Amberif: Gdańsk, Poland, 2013; pp. 28–32.
- Sadowski. E.-M.; Schmidt, A.R.; Denk, T. Staminate inflorescences with in situ pollen from Eocene Baltic amber reveal high diversity in Fagaceae (oak family). Willdenowia 2020, 50, 405–517. [CrossRef]
- Nel, A; Brasero, N. Oise Amber. In Biodiversity of fossils in amber from the major world deposits; Penney, D., Ed.; Siri Scientific Press: Manchester, U.K., 2010; pp. 137–148.
- Nohra, Y.; Perrichot, V.; Jeanneau, L.; Le Pollès; L.; Azar, D. Chemical characterization and botanical origin of French ambers. J. Nat. Products 2015, 78, 1284–1293. [CrossRef]
- Ritzkowski, S.: Das geologische Alter der bernsteinführenden Sedimente in Sambia (Bezirk Kaliningrad), bei Bitterfeld (Sachsen-Anhalt) und bei Helmstedt (SE-Niedersachsen). In Investigations into amber. Proceedings of the International Interdisciplinary Symposium: Baltic amber and other fossil resins, 2-6 September 1997; Kosmowska-Ceranowicz, B., Paner, H., Eds.; The Archaeological Museum in Gdansk, Museum of the Earth, Polish Academy of Sciences: Gdańsk, Poland, 1999; pp. 33–40.
- Wolfe, A.P.; McKellar, R.C.; Tappert, R.; Sodhi, R.N.S.; Muehlenbachs, K. Bitterfeld amber is not Baltic amber: three geochemical tests and further constraints on the botanical affinities of succinite. Rev. Palaeobot. Palynol. 2016, 225, 21–32.
- Grimaldi, D.; Engel M.S. Evolution of the Insects. Cambridge University Press: New York, USA, 2005; pp. 1–755.
- Renne, P.R; Deino, A.L.; Hilgen, F.J.; Kuiper, K.F.; Mark, D.F.; Mitchell III, W.S.; Morgan, L.E.; Mundil, R.; Smit J. Time Scales of Critical Events Around the Cretaceous-Paleogene Boundary. Science, 2013, 339(6120), 684–687. [CrossRef]
- Wiegmann, B.M.; Trautwein, M.D.; Winkler, I.S.; Barr, N.B.; Kim, J.-W.; Lambkin, C.; Bertone, M.A.; Cassel, B.K.; Bayless, K.M.; Heimberg, A.M.; Wheeler, B.M.; Peterson, K.J.; Pape, T.; Sinclair, B.J.; Skevington, J.H.; Blagoderov, V.; Caravas, J.; Kutty, S.N.; Schmidt-Ott, U.; Kampmeier, G.E.; Thompson, F.C.; Grimaldi, D.A.; Beckenbach, A.T.; Courtney, G.W.; Friedrich, M.; Meier, R.; Yeates, D.K. Episodic radiations in the fly tree of life. Proc. Natn. Acad. Sci. U. S. Am. 2011, 108 (14), 5690-5695. [CrossRef]
Figure 1.
Hemilauxania parvula sp. nov., female holotype, right laterally. Scale: 0.5 mm. Photo by J. Roháček
Figure 1.
Hemilauxania parvula sp. nov., female holotype, right laterally. Scale: 0.5 mm. Photo by J. Roháček

Figure 2.
Hemilauxania parvula sp. nov., female holotype, preparatum: (A) whole amber sample (preparatum in polyester resin), in situ; (B) ditto, from largest side; (C), ditto, close-up of amber sample. Scales: 0.2 mm. Photos by J. Roháček
Figure 2.
Hemilauxania parvula sp. nov., female holotype, preparatum: (A) whole amber sample (preparatum in polyester resin), in situ; (B) ditto, from largest side; (C), ditto, close-up of amber sample. Scales: 0.2 mm. Photos by J. Roháček

Figure 3.
Hemilauxania parvula sp. nov., female holotype, dorsally to left dorsolaterally. Scale: 0.5 mm. Photo by J. Roháček
Figure 3.
Hemilauxania parvula sp. nov., female holotype, dorsally to left dorsolaterally. Scale: 0.5 mm. Photo by J. Roháček

Figure 4.
Hemilauxania parvula sp. nov., female holotype, head and thorax dorsally, with chaetotaxy. Scale: 0.5 mm. For abbreviations see paragraph 2.4.
Figure 4.
Hemilauxania parvula sp. nov., female holotype, head and thorax dorsally, with chaetotaxy. Scale: 0.5 mm. For abbreviations see paragraph 2.4.
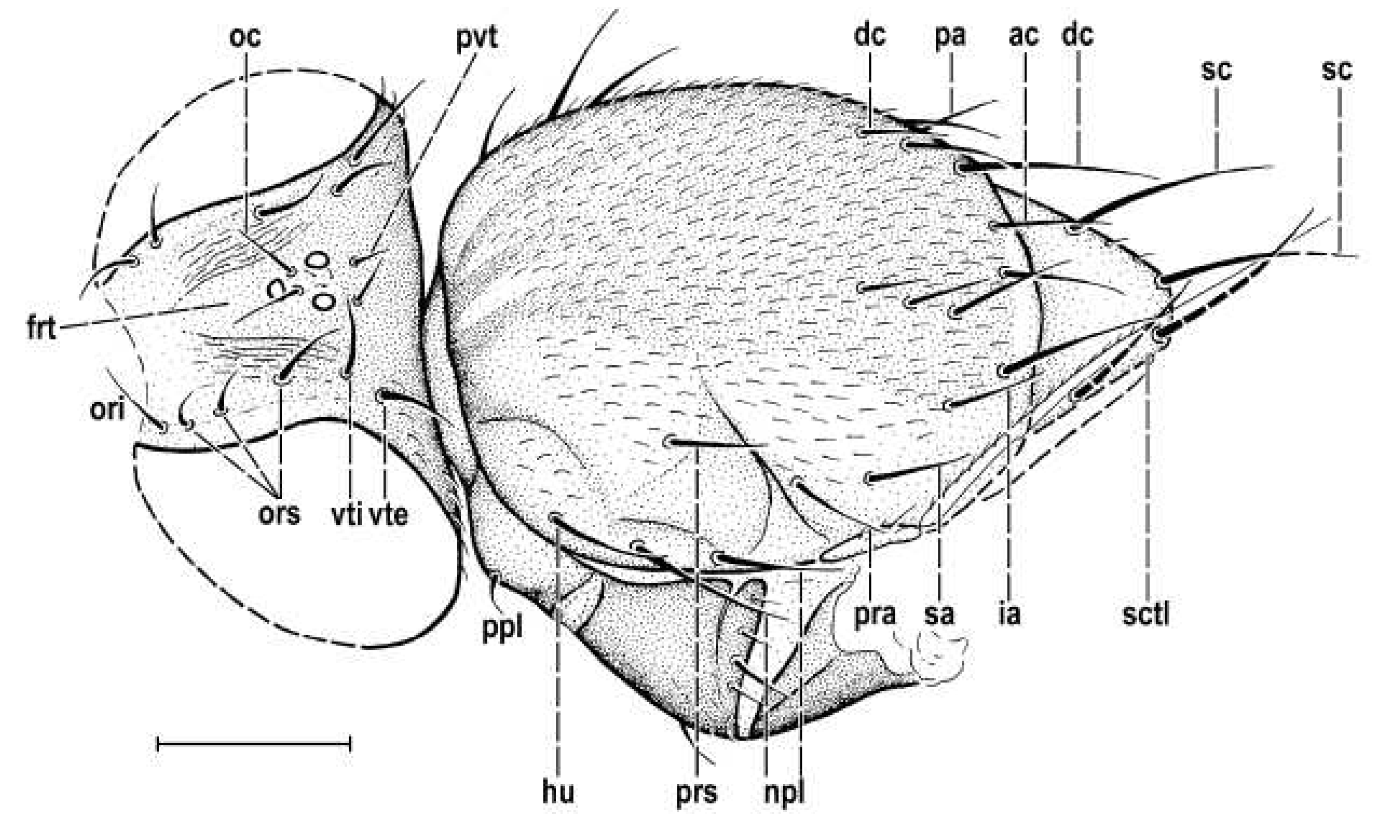
Figure 5.
Hemilauxania parvula sp. nov., female holotype, laterally, with wing venation. Scale: 0.5 mm. For abbreviations see paragraph 2.4.
Figure 5.
Hemilauxania parvula sp. nov., female holotype, laterally, with wing venation. Scale: 0.5 mm. For abbreviations see paragraph 2.4.
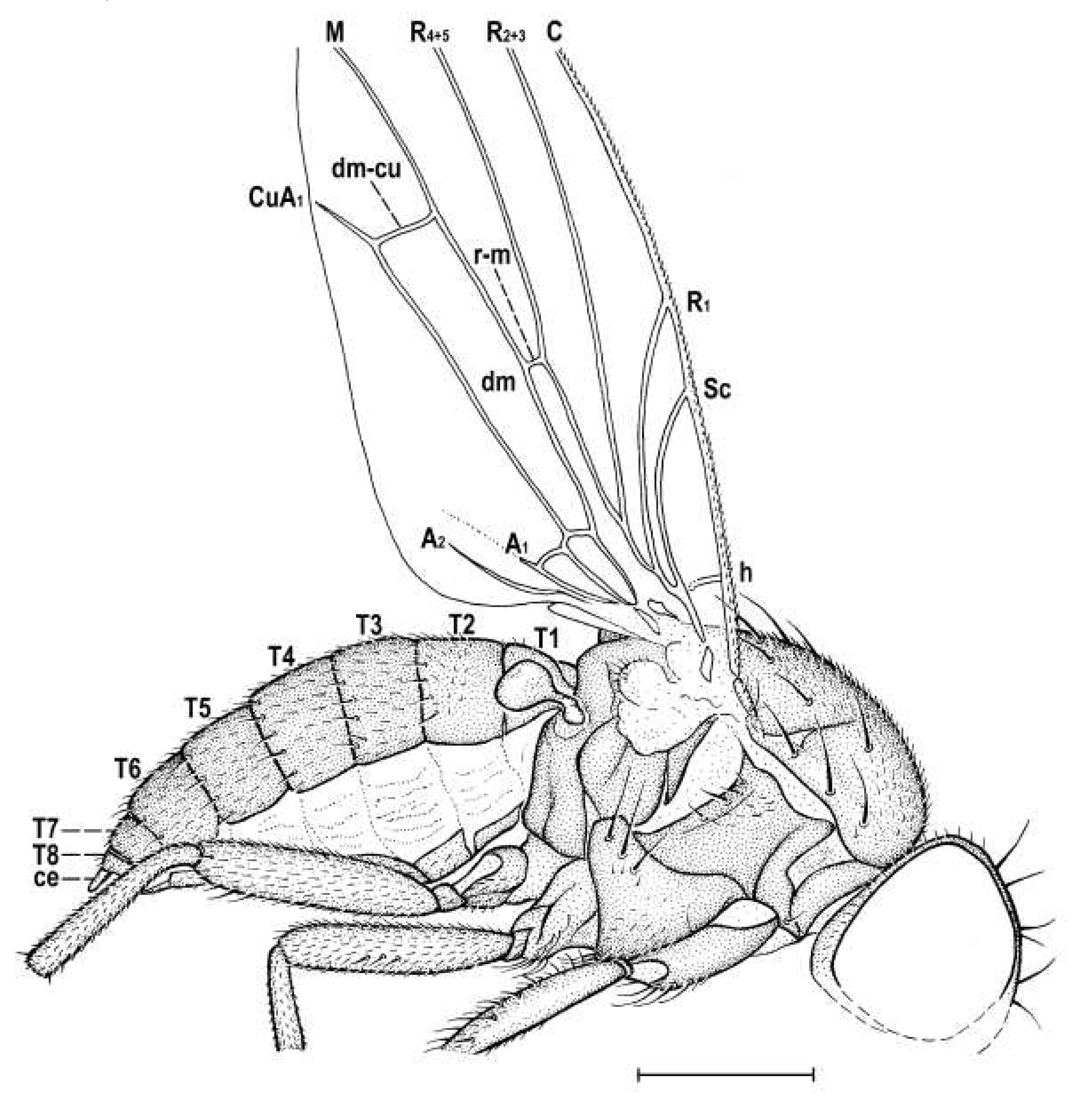
Figure 6.
Hemilauxania parvula sp. nov., female holotype, laterally, structures of thoracic pleuron, wing and legs: (A) anterior part of pleuron, right laterally; (B) distal part of left wing; (C) right fore femur, dorsally; (D) right mid femur and tibia, anteriorly; (E) left hind femur and tibia, anteriorly. Scales: 0.3 mm (A, C, D, E) and 0.5 mm (B). For abbreviations see paragraph 2.4.
Figure 6.
Hemilauxania parvula sp. nov., female holotype, laterally, structures of thoracic pleuron, wing and legs: (A) anterior part of pleuron, right laterally; (B) distal part of left wing; (C) right fore femur, dorsally; (D) right mid femur and tibia, anteriorly; (E) left hind femur and tibia, anteriorly. Scales: 0.3 mm (A, C, D, E) and 0.5 mm (B). For abbreviations see paragraph 2.4.
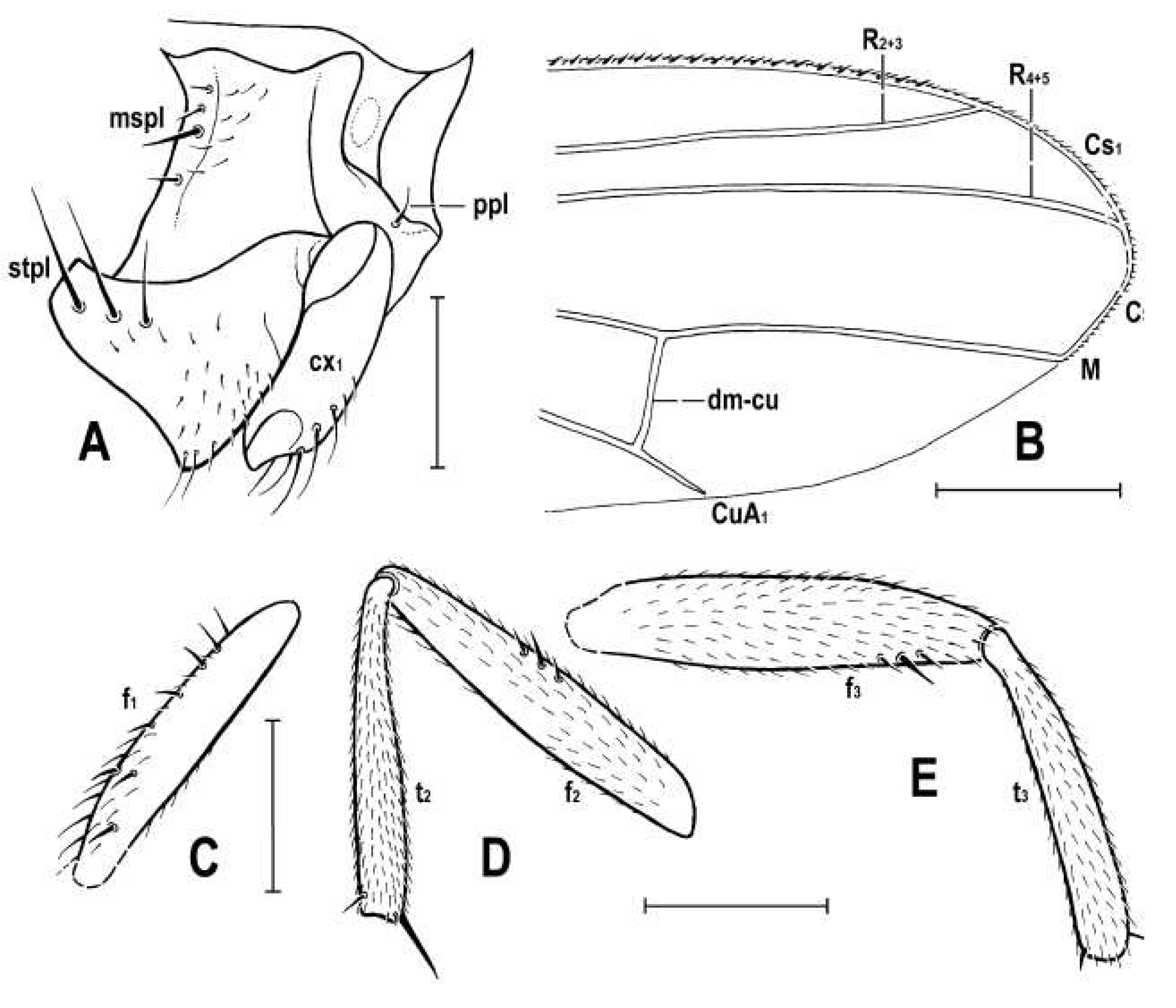
Figure 7.
Hemilauxania parvula sp. nov., female holotype, ventrally. Scale: 0.5 mm. Photo by J. Roháček
Figure 7.
Hemilauxania parvula sp. nov., female holotype, ventrally. Scale: 0.5 mm. Photo by J. Roháček

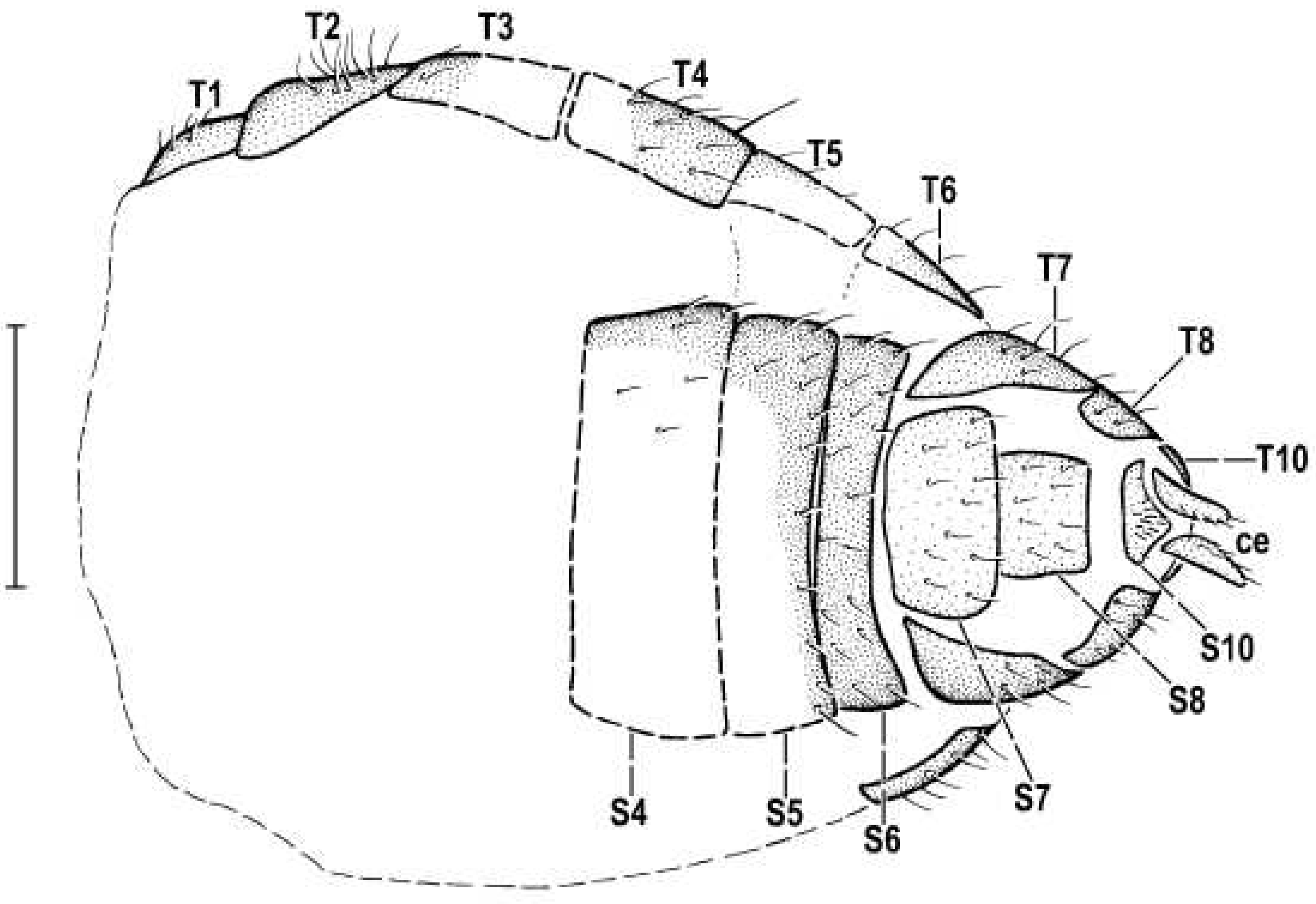
Figure 8.
Hemilauxania parvula sp. nov., female holotype, abdomen ventrally. Scale: 0.3 mm. For abbreviations see paragraph 2.4.
Figure 8.
Hemilauxania parvula sp. nov., female holotype, abdomen ventrally. Scale: 0.3 mm. For abbreviations see paragraph 2.4.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



