
Submitted:
27 September 2023
Posted:
30 September 2023
You are already at the latest version
Abstract
Maize (Zea mays L.) stands as one of the most extensively cultivated crops globally, offering significant contributions to food, animal feed, and biofuel production. Maize yield is significantly impacted by diverse climate and soil variables, yet its productivity faces hindrances due to abiotic stresses, including drought, salinity, extreme temperatures, and cold conditions. In confronting these hurdles, the field of crop breeding has undergone a transformation thanks to high-throughput sequencing technologies (HSTs). These advancements have streamlined the identification of beneficial quantitative trait loci (QTL), associations between markers and traits (MTAs), as well as genes and alleles that contribute to crop improvement. Presently, well-established omics techniques such as genomics, transcriptomics, proteomics, and metabolomics are being amalgamated into maize breeding studies. These approaches have unveiled new biological markers that are being employed to bolster maize's capacity to withstand an array of challenges. In this chapter, we explore the current knowledge regarding the morpho-physiological and molecular mechanisms that underlie the resistance and tolerance of maize to biotic and abiotic stresses. The focus is on utilizing omics techniques to augment maize's ability to withstand these challenges. Moreover, it emphasizes the significant potential of integrating various omics techniques to tackle the challenges presented by biotic and abiotic stress in maize productivity, contrasting with singular approaches.
Keywords:
Maize
; Omics
; Stresses
; Resistance
; Crop Improvement
1. Introduction
Zea mays L., commonly referred to as maize, holds a prominent and cherished status as one of the world's most essential crops. Its importance extends beyond geographical borders, influencing cultures, economies, and dietary habits worldwide. Maize, serving as a staple food source for humans, providing nourishment, supporting livelihoods, fermentation substrate, and a valuable commodity in numerous industrial applications [1], particularly in its dry grain form, occupies a critical role on the global scale. Wheat, maize, and rice are the primary staple cereals worldwide, each cultivated on approximately 200 million hectares. Corn, frequently referred to as maize, underwent domestication more than 9,000 years ago within the southern Mexico/Mesoamerica region [2]. Together, these three primary global staple cereals, namely wheat, rice, and maize, make up a substantial portion of the human diet, accounting for approximately 40 percent of the world's calorie intake and 35 percent of protein consumption [3]. Maize fulfils a versatile and continually evolving function within the global agricultural and food systems, making substantial contributions to food and nutrition security [4,5]. About 56% of its output is used as livestock feed, while one-fifth finds application in non-food sectors, and 13% is designated for human consumption. Notably, maize is distinguished by its high starch content, constituting roughly 65% of its composition [6]. Currently, maize has risen as a viable alternative to rice and wheat. Around 35% of its harvest is directed towards human consumption, while 25% serves as feed for poultry and cattle, and another 15% is used in food processing [1]. It has achieved the status of a significant global commodity, with 15% of the world's maize production currently being exported, marking an increase from the 11% reported in the previous decade [3]. It is on the verge of overtaking wheat as the most heavily traded cereal. Leading net-exporting nations such as the USA, Brazil, Argentina, Ukraine, and Romania are consistently shipping substantial quantities, ranging from 5 to 54 million metric tons annually [7]. Over the last century, maize yield has surged by a factor of eight, thanks to innovations in yield per plant and plant density optimization achieved through harnessing heterosis.
Throughout history, maize has been a quintessential subject in the realms of genetics, developmental biology, physiology, and, more recently, genomic research. The genetic investigation of Zea mays L. commenced with Edward East's pioneering research in 1908, which explored topics like inbreeding depression and hybrid vigor. A significant advancement in cytogenetics occurred in the 1940s when transposable elements (TEs) were discovered, as exemplified by Barbara McClintock's pioneering work [8]. In 2009, the first maize genome was made public [9]. Then, Jiao et al. [10] embarked on a re-sequencing project focused on the B73 maize variety. This effort revealed that a substantial 74% of its genome is comprised of long-terminal repeat retrotransposons (LTRs), which predominantly contribute to its enlarged size in comparison to other grass species. The main factor responsible for the maize genome's expansion relative to other grasses is the widespread increase in LTRs [10]. The cumulative cytogenetic, genetic, and genomic studies of maize have yielded rich insights into its genome's structure, function, and evolution. Resequencing wild relatives, traditional landraces, and improved maize lines, and aligning them with the reference genome, suggests that introgression from wild relatives contributes to post-domestication maize diversity. Through this method, genes with a wide range of biological functions that experienced selective pressure during the domestication process have been pinpointed [11].
The foundation of maize breeding relies on leveraging heterosis, which involves genome-wide allelic interactions, interactions among quantitative trait loci (QTLs), and inter-genomic interactions that occur when the two parental genomes combine in the F1 hybrid. The functional understanding of many maize genes, especially those linked to heterosis, is less advanced compared to other cereal crops, notably wheat and rice. Integrating whole-genome markers into genomic-based breeding represents a viable approach for improving maize breeding and holds significant promise. Genomic selection (GS) serves as a notable illustration of this genomic design breeding strategy as it doesn't require an extensive comprehension of gene functions or the precise assessment of each individual marker's efficacy [12]. In this contemporary genomics era, the integration of various strategies and methodologies promises to boost maize productivity. This includes the amalgamation of modern genomics, phenomics, gene editing, synthetic biology, and the utilization of AI technology. The integration of an extensive array of maize omics data, spanning genomics, phenomics, epigenomics, transcriptomics, proteomics, and metabolomics, will form a vital foundation for machine learning approaches to build network models illustrating the interactions among various genetic components. In the context of a maize breeding program, a key strategic approach for achieving the desired goals of increased production and enhanced quality traits revolves around reducing the frequency of hybridizations while maximizing the incorporation of superior alleles. Swift progress toward trait enhancement objectives can be attained by executing two or three iterations of small-scale population development, thereby making the most of the available genetic diversity. This chapter highlights the importance of incorporating comprehensive strategies to enhance maize production, utilizing various modern techniques such as molecular breeding, marker-assisted selection (MAS), GS, the role of genome editing (CRISPR-Cas), and transgenic approaches. Additionally, the chapter delves into future prospects and significant challenges in the field of maize improvement.
2. Traditional Approaches in Maize Crop Improvement
Based on recent molecular analysis, it is now believed that the process of maize domestication commenced in the Central Balsas River Valley approximately 8,700 long ago in southwestern Mexico. This domestication process occurred rapidly, originating from Zea mays ssp. parviglumis wild precursor, a subspecies of teosinte. This information is supported by studies conducted by Liu et al. [13], Piperno et al. [14], and Ranere et al. [15]. At the International Institute of Tropical Agriculture (IITA), maize underwent enhancements in various quantitative traits through classical or conventional methods. These improvements encompassed traits such as Striga resistance, nitrogen utilization efficiency, drought tolerance, resilience to stem borers, mitigation of aflatoxin accumulation, yield potential, and enhancement of nutritional quality [16,17]. In traditional maize breeding, the approach entails the development of new plant cultivars by adhering to the principles of natural inheritance. This involves the selection of plants based on their exceptional performance in specific traits or characteristics, as discussed by Lamichhane and Thapa [18]. Conventional breeding methods have been employed for the two, self-pollinated and cross-pollinated plants for quite shortly. One example is the concept of pure line selection, which was introduced by Johannsen [19], as documented by Poehlman [20]. This method involves the creation of pure lines through the self-pollination of a single superior homozygous parental genotype. Following several years of conducting multi-locational trials, typically spanning approximately 6 to 7 years and involving the comparison with established check varieties, superior genotypes are officially introduced as new maize varieties. Pure-line selection is less effective due to low heritability caused by environmental effects, as genetic makeup closely resembles parental genotypes [21]. Mass selection, akin to pure-line selection, relies on highly heritable traits for plant choice [22]. Mass selection can be executed in two ways; first is single-parental, where one kind of gamete is controlled, and second one is bi-parental, where the couple of gametes, female and male are controlled. The chosen individuals are then planted in the crop land and harvested when they reach maturity. After harvest, seeds are mixed and sowed for the next generation. In the next year, crop plants grown from mixed seeds are justified with a check variety for variance. Selected plants are released as new varieties after multi-location trials. Backcross breeding introduces desirable traits from one plant into another without affecting other traits by crossing with a homozygous parent [23]. In this method, donor parents possess the desired trait, and recurrent parents receive these selected genes. After five to six generations of repeated backcrossing with the recurrent parent, the backcrossed progeny should inherit approximately 98% of the recurrent parent's genome [24]. In backcross breeding, the newly formed variety typically inherits a majority of its genes from the recurrent parent, with only a few coming from the donor parent, as noted by Singh [25]. Another method is recurrent selection, a term introduced by Hull [26], primarily applied in maize breeding but later extended to other cereal crops, as discussed by Ramya et al. [27]. This process entails the continued selection of favourable traits over multiple generations, with the aim of increasing their prevalence through crosses between high-performing individuals from the heterozygous recurrent parent and inbred individuals, as discussed by Bangarwa [28]. Hybridization is another method for creating hybrids with desirable traits by mating genetically distant parents within the same species (Intraspecific hybridization) or between different species (Interspecific hybridization). It involves combining characteristics from different parents to produce genetically superior offspring, whether through natural or artificial means [21].
In a remarkable long-term study conducted with conventional breeding techniques, researchers at the Illinois Agricultural Experiment Station successfully enhanced the oil concentration in maize. They started with a base of approximately 5% oil content and, over the course of 100 generations, developed high oil-producing maize lines, which now boast an impressive 20% oil content [29]. However, conventional breeding methods do have their limitations. For instance, identical parents do not produce variation due to lack of segregation of gametes in conventional breeding [30]. Additionally, this process is often time-consuming, typically spanning over a decade or more before a new cultivar is ready for release, as noted by Bharti and Chimata [31]. Moreover, conventional breeding heavily relies on the cultivars phenotypic expressions to identify superior ones. Hence, the chosen cultivars may not consistently be without errors, given that phenotypes are significantly affected by genotype-environment interactions [32]. The selection process involves choosing individuals for breeding based on their differences in desired features, which are usually measurable or observable traits [33]. It's worth noting that conventional breeding is an applied science that heavily depends on the observations, skills, and experiences of breeders for judgment, as highlighted by Allard [34].
3. Molecular Breeding and Marker-Assisted Selection (MAS)
Traditional plant breeding involves the iterative practice of selecting both parents and their offspring based on desirable characteristics. The significance of molecular breeding is substantial in the contemporary world, especially in developed countries where a small proportion of the population is engaged in agriculture. This minority group bears the demanding responsibility of providing sustenance for the majority of the country's population [35]. This achievement is feasible in view of the fact that plant breeding has effectively enhanced crop yields without the need to expand the cultivated land or the workforce engaged in agriculture. Achieving this objective can be readily accomplished through crop enhancement via molecular breeding techniques. Molecular breeding employs various approaches such as the identification of simple traits or QTLs within breeding lines/populations, the integration of genes from breeding lines or wild relatives, gene pyramiding, marker-assisted recurrent selection (MARS), and Marker-assisted backcross (MABC).
Molecular MAS, often referred to as marker-aided selection or MAS, is an indirect approach to nomination or selection wherein a specific trait is targeted through the use of a marker [29]. Within the context of MAS, a marker is located in vicinity to a gene responsible for controlling the trait, thereby signifying the presence of a desirable allele when the marker is detected [36]. Knowing the alleles in key loci allows for the creation of optimal allele combinations to enhance the agronomic value of the genotype. Marker-assisted selection is commonly used for resistance gene pyramiding, which can provide complete resistance for several plant generations until it's challenged by pathogen strains. Achieving gene pyramiding for resistance becomes exceedingly challenging through classical breeding methods for certain traits like pathogen-induced disease resistance when dominant resistance genes are present [35]. There are primarily three categories of genetic markers. The first comprises visible or morphological markers, which are characteristics or phenotypic traits. The second category includes biochemical markers, which involve enzymes allelic variations referred to as isozymes. The third category consists of molecular or DNA markers, which unveil points of variation in the DNA sequence [37,38]. For a nucleotide sequence to be useful as a molecular marker in molecular breeding, it usually requires polymorphism within its sequence. These variations in nucleotide sequences are unveiled through molecular methods like restriction fragment length polymorphism (RFLP), amplified fragment length polymorphism (AFLP), random amplified polymorphic DNA (RAPD), single nucleotide polymorphism (SNP), microsatellite or simple sequence repeat polymorphism (SSRP), sequence-tagged site (STS), single-strand conformation polymorphism (SSCP), and cleavable amplified polymorphic sequences (CAPS) among others [39]. Utilizing this marker set relies on extensive prior research. This typically involves various research stages for each trait, commencing with QTL mapping, progressing to fine mapping, and ultimately culminating in positional cloning [40]. For a successful MAS program, essential components include dependable markers, a robust DNA extraction method, well-constructed genetic maps, swift and efficient data handling, an interpretation of marker and trait connections, and knowledge to access tools for high-throughput marker detection [41].
MABC represents a specialized example of MAS, wherein the process of backcrossing is aided by molecular markers to expedite the selection of the recurrent parent and enhance genome recovery speed. The MABC technique has found extensive application in eliminating undesirable traits, such as susceptibility to insects and diseases, as well as anti-nutritional factors, from widely adopted high-yielding varieties by introducing QTLs or genes of interest from donor parents [42,43]. Using DNA markers in a breeding program recommended a variety of benefits. For instances, DNA marker-based screening facilitating early selection for traits due to evaluation of plant genotypes at the seedling stage or even from seeds, that may only manifest in adult plants, such as male sterility, fruit or grain quality, and photoperiod sensitivity. It expedite the selection of alleles that are difficult to assess phenotypically, especially for environmentally sensitive traits, simplifying and enhancing the breeding process. Individual plant selection, which may be impractical through phenotypic means, becomes feasible when relying on marker information. The issue of low heritability becomes inconsequential when using marker-based selection. Additionally, in traits with intricate inheritance patterns, it becomes possible to select each individual genetic component contributing to the trait independently. Using molecular markers, multiple characters that typically exhibit epistatic interactions can be preserved and ultimately stabilized. Moreover, the preservation of recessive genes does not necessitate progeny testing in every generation since homozygous and heterozygous plants can be differentiated using (codominant) markers, as explained by Lema [32]. Molecular markers have a important role in enhancing maize's genetic traits, such as addressing the intricate inheritance patterns related to drought tolerance [44,45,46], improving nutrient utilization [47,48,49,50,51,52,53], and enhancing diseases, parasitic and insect pests plant resistance in maize [17,54,55,56,57]. Additional details can be found in the works of Hossain et al. [58] and Muntean et al. [33].
4. Genomic Selection
The GS represents an advanced iteration of marker-assisted selection, enhancing the efficiency of selection and expediting the progress in selective breeding within a shorter timeframe. It achieves this by employing markers across the entire genome to predict the impact of quantitative gene loci, subsequently calculating genomic estimated breeding values (GEBVs) [59,60]. The GS breeding approach was initially proposed by Meuwissen et al. [61]. GS has the capability to swiftly and cost-effectively forecast the yield potential of individual plants, ultimately leading to a reduction in both the time and expenses associated with a breeding cycle. In contrast to GWAS (genome-wide association studies) and linkage analysis, the primary goal of GS is not to pinpoint specific QTLs but rather to make predictions about the future performance of offspring based on the DNA information gathered in the present. In GS, breeders can predict a plant's breeding value by utilizing data from all markers without the need for direct phenotype evaluation. This prediction relies on statistical models developed using a “training population” where both genotypes and phenotypes have been recorded. In mixed model analysis for genomic selection, markers are treated as random factors. This approach is necessary because the number of markers often exceeds the number of individuals in the training population, making it impractical to estimate the effect of each marker individually due to limited degrees of freedom [35,62]. MAS proves valuable when choosing traits such as grain yields, flower colours, seed characteristics, and others that manifest primarily during the later reproductive stages. Through the application of GS, these traits can be detected by employing DNA markers in a genotype even at preliminary stages of plant development as highlighted by Madhusudhana [63]. It offers numerous advantages compared to traditional MAS. Notably, it doesn't require QTL mapping, as it efficiently estimates breeding values using a comprehensive set of molecular markers that ideally spans the whole genome [64]; At early selection, it is more precise as it estimates all QTLs effect by utilizing high-density molecular markers and explains genetic variance for desirable traits. This stands in contrast to MAS, which relies on a limited number of markers for trait selection [65]; it shortens generation intervals, accelerates genetic progress (4-25% farther up phenotypic selection), and reduces breeding costs (26-56% lesser traditional methods) [65]; it exhibits superior efficiency in selecting traits with low heritability compared to MAS; in GS, breeding values serve as the selection criteria which are the sum of all allele genetic effects for each individual,. This approach is known for its superior accuracy because it assesses the average performance of the offspring, rather than relying solely on the parents' performance [67]. In crops like maize, research conducted by the CIMMYT (International Maize and Wheat Improvement Center) [68], suggests that the breeding interval could be shortened to as little as half of the conventional timeframe. GS has been effectively applied to enhance various traits in maize, including shelling percentage, grain yield, grain moisture, ear length, ear width, ear rows, tassel branch number, kernels per spike, hundred-grain weight, kernel number per ear, and kernel depth [69,70]. Challenges in GS arise from factors including the training population's size and variability, as well as the heritability of the traits being predicted. The statistical intricacies in GS are connected to the vast amount of marker data, where the number of markers far exceeds the number of observations [33]. In conclusion, GS has proven to be a valuable tool for the improvement of multiple important traits in maize, offering promising prospects for enhancing the overall performance and quality of maize crops.
5. Genome Editing and CRISPR-Cas9
Genome editing is a form of genetic engineering that involves the intentional alteration of DNA within living cells through the insertion, deletion, or modification of genetic material. Genome editing, previously referred to as gene targeting, is a precise method for modifying the nucleotide sequence of the genome with an exceptional level of specificity down to the individual base pair. It encompasses a range of strategies and methods designed to make deliberate and customized modifications to the genetic makeup of an [71]. The essence of this editing technology is its reliance on site-specific nucleases (SSNs), which are customizable enzymes proficient in precisely targeting specific gene sequences. By utilizing these modified nucleases, it becomes feasible to precisely remove, insert, or replace particular gene sequences, demonstrating the benefits of site-directed mutagenesis when compared to random mutagenesis methods [72]. Genome editing techniques encompass a range of approaches, including tailored homing nucleases (meganucleases), zinc-finger nucleases (ZFNs), and transcription activator-like effector nucleases (TALENs). These methods employ protein-based systems that can be tailored to possess precise DNA-binding functions, allowing them to pinpoint specific gene sequences. The most widely adopted platform in recent times is clustered regularly interspaced short palindromic repeats-CRISPR-associated 9 nuclease (CRISPR-Cas9), which relies on RNA as a targeting element directing the nuclease to a specific DNA sequence [73,74].
5.1. CRISPR-Cas9 in Maize
Maize (Zea mays L.) is recognized as the most widely grown grain crop globally. Its versatility in terms of applications and adaptability to diverse environmental and soil conditions have contributed to its popularity as a desirable crop across the globe [75]. The drawbacks of random mutagenesis sparked investigations into precise genome modification methods, resulting in notable progress over the last decade. These methods have significantly increased the accuracy of gene editing, enhancing fidelity by nearly a thousand fold [76,77]. The initial generation of targeted genome editing methods, including ZFNs and TALENs, achieved partial success but exhibited specific limitations [78].
In contrast, the CRISPR/Cas9 system, a relatively recent addition, has revolutionized the field of genome editing due to its simple design, operational flexibility, and cost-effectiveness [79,80,81]. The system consists of a universally used Cas9 nuclease protein and a solitary guide RNA (sgRNA) that includes a 20-base pair (bp) target site sequence along with a hairpin structure. The Cas9 protein induces a double-strand break (DSB) at the 20 bp genomic locus specified by the sgRNA, occurring near the NGG sequence called Protospacer Adjacent Motif (PAM), where N can represent any nucleotide. Cas9 consists of two catalytic nuclease domains, RuvC and HNH, responsible for creating DSBs at precise target sites guided by sgRNA. These DSBs can subsequently be repaired through two main mechanisms: Non-homologous end-joining (NHEJ) or Homology-directed repair (HDR) (see Figure 1). It's worth noting that a mere 20 bp sequence is ample for achieving allele specificity within single-copy regions of a genome, such as in maize. The compact size of the sequence simplifies the creation of sgRNA [82,83].
In maize protoplasts, the CRISPR/Cas9 system achieved a targeted mutation efficiency of 13.1% in the phytic acid biosynthesis gene, ZmIPK, whereas TALENs achieved 9.1% [84]. Furthermore, the CRISPR technology demonstrated a mutation frequency in maize that was 10-20 times greater than that observed with homing endonucleases [85]. In a more recent advancement, a user-friendly public sector system known as “ISU Maize CRISPR” has been established to facilitate efficient site-specific mutagenesis in maize. It employs an E. coli cloning vector and an Agrobacterium binary vector, enabling the incorporation of as many as four single guide RNAs (sgRNAs) for single or multiplex mutagenesis. This development marks a significant stride in applying CRISPR/Cas9 for multifaceted gene editing in crops, with a specific focus on maize [86].
Delivering the CRISPR/Cas9 system, comprising the sgRNA and Cas9 protein, can be accomplished through transient techniques or by employing a long-term maize transformation process into the cell. Furthermore, this method allows for multiplexing [83]. Unlike some other nucleases, CRISPR/Cas9 is capable of targeting methylated DNA, rendering it a versatile tool for editing plant genomes [87].
5.2. Potential Benefits of Genome Editing and Ethical Considerations
Genome editing starts from a molecular understanding of the target gene and utilizes a targeted and precise approach based on specific molecular knowledge. This allows genome editing to achieve highly accurate and intentional modifications in a controlled manner. One of the notable concerns linked to genome editing in plants is the potential for unintended genetic changes arising from off-target mutations [88,89]. These off-target mutations refer to unintended changes in the DNA sequence of the plant's genome that occurs at sites other than the intended target. Such unintended alterations can have unpredictable effects on the plant's characteristics and may raise safety and environmental concerns. The concerns surrounding genome editing also stem from the limited understanding of its principles and applications among the general public. There is a need for clear communication and education to address the knowledge gap and foster informed discussions about the technology. Differentiating between various categories of genetically modified plants, such as transgenic plants and genome-edited plants [90,91], is essential. This distinction aids in comprehending the precise methods employed, the scope of genetic alterations made, and the possible consequences regarding safety, regulation, and public perception. By addressing these concerns and promoting transparency, scientists, policymakers, and stakeholders can work together to ensure responsible and ethically sound use of genome editing technologies in plant research and crop improvement. The risks related to the emerging techniques in genome editing encompass various interconnected aspects, including environmental, health-related, agricultural, economic, social, and political concerns. Among these, only a limited subset of risks is directly associated with the new techniques. One notable risk is the potential for bioterrorism, although it is currently only a theoretical concern when it comes to plants. Genome editing can have both positive and negative implications for agricultural risks, particularly in relation to biodiversity. On one hand, it has the potential to contribute to a reduction in biodiversity. On the other hand, it can also be utilized to enhance diversity and address emerging threats in agriculture.
6. Use of Transgenic Approaches to Introduce Foreign Genes into Maize
The adoption of genetically modified (GM) traits has been rapidly embraced by farmers worldwide, making it one of the fastest innovations in agriculture. As per findings presented by Brookes and Barfoot, [92], the worldwide economic benefits derived from genetically modified crop varieties amounted to US$ 225 billion during the period spanning from 1996 to 2018, with developing countries accounting for 52% of these gains. In 2019, transgenic crops were grown across 190.4 million hectares in 29 countries, intended for both food and feed purposes. This represents a substantial rise from 1.7 million hectares in 1996, marking a remarkable 112-fold increase. The most widely adopted GM crops include soybean, maize, cotton, and canola. Of these crops, maize takes the lead. In 2019, 31% of the total global maize cultivation area, encompassing 60.9 million hectares across 14 countries, was allocated for growing genetically modified maize varieties [93].
In the last three decades, GM maize varieties have been effectively introduced to the market, providing farms with traits like resistance to herbicides and insects. The initial wave of genetically modified maize featured a single gene with a precise mechanism targeting a particular order of insects to confer insect resistance. In subsequent generations, the approach included crossbreeding herbicide and insect-resistant traits, as well as diverse insect resistance traits, to establish multiple mechanisms of action against a range of insect orders. Farmers have found these stacked varieties to be remarkably effective, delivering evident and comprehensive phenotypic results [94]. Developing traits linked to quantitative characteristics such as tolerance to abiotic stress, efficient nutrient utilization, and increased yield presents a more intricate challenge. These characteristics are influenced by numerous genes and are sensitive to environmental factors, which adds complexity to the development process. To examine how individual genes affect complex traits, companies have established extensive biotechnology pipelines that involve evaluating genes in real field conditions on a large scale [95]. A standard biotechnology pipeline comprises various stages, which encompass discovery, demonstrating feasibility, initial development, advanced development, pre-launch, and the eventual release of commercial varieties. Certain stages might briefly coincide, particularly when a promising lead is identified early in the discovery phase, leading to the initiation of optimization efforts before validation is entirely finished. The phase of gene discovery entails the quest for potential genes, which can be arduous, expensive, and uncertain, especially for traits such as drought tolerance and yield, which demand clearly defined phenotypic reactions. Extensive phenotypic screening of model plants like Arabidopsis thaliana and Oryza sativa is performed to assess hundreds of candidate genes [96,97]. The proof-of-concept stage involves creating events for each candidate gene and conducting preliminary phenotypic evaluations in controlled settings as well as small-scale field experiments. During the early development phase, there is a focus on optimizing the lead to enhance stability and increase protein expression. Candidates demonstrating favourable agronomic performance, consistent trait expression, and heritability are chosen. These chosen candidates are subsequently subjected to molecular-level characterization and extensive field trials conducted across various locations and over multiple years [98].
In the advanced development stage, the validated leads are incorporated into commercial lines, often employing molecular markers to expedite the breeding process and guarantee the successful transfer of traits. In this phase, regulatory data related to the toxicity of gene products, allergenic potential, compositional analysis, as well as environmental and human safety aspects are collected. During the prelaunch stage, the production of seeds for the novel GM variety is expanded, quality control protocols are instituted to guarantee trait consistency and purity, a regulatory report is submitted, and arrangements are made for the commercial release of the new GM trait hybrid. The duration to finalize the pipeline, which varies according to the trait and available resources, generally averages around 11-13 years.
6.1. Enhancing Traits in Genetically Modified (GM) Maize Varieties
Twenty-five years ago, the debut of the initial commercially accessible insect-resistant GM maize [99,100] marked the beginning of a journey that has led to the approval of 148 GM maize events for global commercial utilization [94]. By 2019, the worldwide GM maize cultivation had expanded to cover 61 million hectares, with the most substantial acreage located in the USA (33 million hectares), Brazil (15 million hectares), Argentina (6 million hectares), and South Africa (2 million hectares) [93]. Among crops, maize holds the record for the largest number of approved GM events, totalling 148 events across 35 countries. Most of these events integrate traits like insect resistance and herbicide tolerance. Furthermore, approved traits for maize encompass fertility restoration, male sterility, heightened drought tolerance, phytase production, modified amino acids and alpha amylase expression, improved photosynthesis, and increased ear biomass. These authorized traits encompass a total of 39 individual genes, with the largest proportion associated with insect resistance (18 genes) and herbicide tolerance (11 genes). The forthcoming generation of GM maize varieties poised for market release incorporates events featuring novel insecticidal proteins, including Vpb4Da2, DvSnf7 RNA, and IPDO72Aa. These proteins are designed to manage insect populations that have developed resistance to Bt [101,102,103]. Additional prospective varieties seek to enhance grain yield by upregulating the zmm28 and ZM-BG1H1 genes [104,105], and to bolster drought tolerance through the overexpression of ARGOS8 [106].
6.2. Concerns and Regulations Related to GM Crops
It's crucial to emphasize that scientific evidence confirms that GM crops do not present any heightened risks to both humans and the environment in comparison to conventional crops (National Academies of Sciences, Engineering, and Medicine, 2016). Nevertheless, public apprehensions and restrictions related to GM technology, notably within the European Union, continue to hold substantial importance [107]. In an effort to tackle these concerns and potentially shift public opinion, there is ongoing exploration of new plant breeding technologies (NPBTs), including cisgenesis, intragenesis, and genome editing. Proficient communication of these technologies to the public holds the potential to impact public approval [108,109]. Following seven years of GM crop cultivation with no observable health impacts, apprehensions about potential environmental hazards, particularly gene transfer to other species, have gained more prominence than concerns regarding food safety. Pollen and seeds released into the environment may convey genetic characteristics to neighbouring crops or wild relatives. Self-pollinating crops like wheat, barley, and potatoes have minimal chances of gene transfer, whereas cross-pollinating crops like sugar beets and corn are of greater concern in this regard. Although numerous cultivated crops lack wild counterparts in their present cultivation regions, the places of origin for these crop species are notably vulnerable to the infiltration of transgenic traits into native varieties or landraces. There is apprehension that transgenic varieties possessing a competitive edge might progressively supplant valuable genetic diversity. Consequently, Mexico, a nation harbouring over 100 distinct corn varieties, has enforced a ban on the cultivation of transgenic corn.
7. Omics Technologies in Maize Improvement
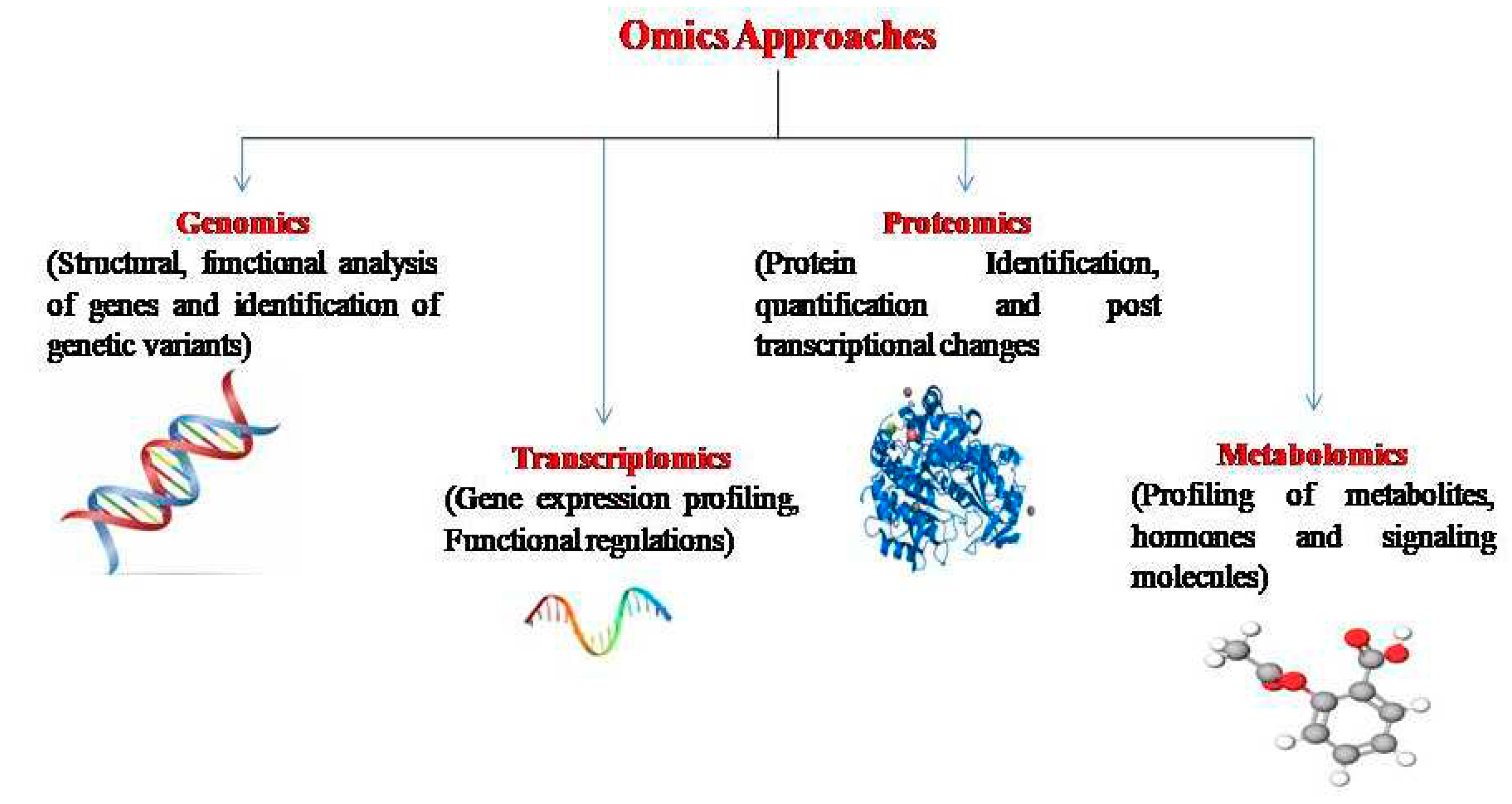
Advancements in the fields of biotechnology and computational sciences have paved the way for the generation of omics data on a large scale for various plant sets, including different varieties and species [109]. The application of diverse omics techniques has facilitated the discovery of genes, their respective functions, the specific types of RNA or proteins involved, their structural attributes, and the pathways influencing the development of final morphological traits. These identified genes can be subject to manipulation or transfer to create novel varieties or hybrids possessing advantageous traits. The multi-omics approaches has proven successful in enhancing crop yields and developing resistance to stresses in agriculture (Figure 2). Molecular biology methods encompassing various omics technologies, such as genomics, transcriptomics, proteomics, and metabolomics, have played instrumental roles in advancing these research endeavours [111,112].
Genomics is dedicated to the sequencing, characterization, and comprehensive exploration of a plant's genetic makeup, encompassing its composition, structure, functions, and intricate networks within the genome [113]. Novel approaches to plant breeding have been made possible by developments in plant genomics, which effectively enhancing and expediting various aspects of the breeding process. These innovations include techniques such as marker-assisted selection, gene pyramiding, association mapping, breeding by design, genomic selection, and more [114]. In a study conducted by Vinayan and colleagues [115], genomic regions linked with fodder traits were pinpointed, and a prediction study on genomic regions was carried out using 1026 DH lines and 276 elite lines as prediction sets from bi-parental crosses.
In order to determine the expression profiles of both coding and non-coding RNA in response to different stresses, high-throughput sequencing platforms were used by transcriptomics to generate transcript data. It also incorporates RNA sequencing, microarray and serial analysis of gene expression (SAGE) [116]. The genetic makeup of the transcripts that have differential gene expression in particular cells has been revealed by a number of studies. These transcripts can affect phenotypic variations in maize, such as growth, yield components, disease tolerance, environmental response and quality traits. For instance, transcriptome correlation and comparisons signalling network analysis were used in a study by Liu and Zhang [117] to identify six genes essential to the control of the MAP Kinase cascade and HY5 module in the presence of blue light. In maize, these genes are essential for regulating stomata formation and dispersion. qRT-PCR and transcriptome analysis studies in maize roots, that are infected by Holotrichia parallela larvae, were the focus of another study project conducted in 2020 by Pan and colleagues [118,119]. This showed the expression of twelve differently expressed genes linked to the pathways of benzoxazinoid production and Jasmonic acid-mediated signalling, which are in charge of maize roots' defense mechanisms against invaders. Zhou and colleagues [120] used bulked sergeant transcriptome analysis (BSTA) to study the mechanism behind maize's resistance to drought stress. On chromosome 2, four highly expressed candidate genes that confer Gibberella ear rot disease resistance in maize were found by transcriptome profiling of several inbred lines of maize [121]. Together, these results highlight the significance of transcriptomics in maize research, as it facilitates the discovery of essential regulatory components for enduring abiotic and biotic challenges, as well as the annotation of gene functions and the identification of candidate genes. Breeders will be able to solve present and future economic, ecological, and environmental concerns and ensure food security by using this information to gain the insights they need to create improved varieties of maize [70,122,123].
Proteomics involves a comprehensive investigation of proteins within a biological system, encompassing plants and animals, at a specific moment in time [124]. The analysis of proteomics serves the purpose of quantifying the abundance of various proteins, discerning alterations resulting from diverse post-translational modifications, elucidating their following functions and localization [125]. It offers a snapshot of diverse metabolic processes, their ensuing interactions, and impacts on other regulatory pathways. Consequently, proteomic studies are indispensable for deciphering the diverse reactions within pathways under various stress conditions and timeframes [126]. Proteomics field has attracted considerable interest from scientists seeking to examine physiological differences at the proteomic level under varying stress conditions. For example, Zhang et al. [127] performed a proteomic analysis of maize leaves in an attempt to evaluate proteome-level changes in corn when infected by the Ostrinia furnacalis (Asian corn borer). A total of 62 defense-responsive proteins were found, with a special focus on thioredoxin M-type and pathogenesis-related protein 1 (PR1), a chloroplastic precursor that significantly impacted the development of corn borer larvae and pupae. Comparative proteome profiling was done on resistant and susceptible lines exposed to Puccinia polysora (southern corn rust) in a study by Wang et al. [128]. This study demonstrated that resistance in the resistant lines was inhibited by a particular remorin protein (ZmREM 1.3). A comparative proteome profiling of drought-tolerant and susceptible maize lines was carried out by Dong et al. [129]. Plants use the development of defense-associated proteins (DAPs) in conjunction with the down-regulation of redundant proteins as a stress-reduction and energy-saving strategy.
Metabolomics is a cutting-edge biotechnique that seeks to identify metabolites that are functionally active, clarify their functions, and provide insight into the various biochemical processes that occur in plant genotypes and the phenotypic expressions that follow (Führs et al. [130]. All metabolites, primary and secondary, with a molecular weight of less than 1500 Da, as well as their precursors and intermediates within the corresponding metabolic processes, are included in the metabolomes. Based on their particular goals, metabolic investigations can be divided into 2 categories: targeted and untargeted. The goal of targeted metabolomics is to precisely quantify one or a small number of metabolites from a predetermined list of recognized compounds. This method helps identify metabolites linked to particular features because of its high sensitivity and quantitative nature. Untargeted metabolomics, on the other hand, increases the possibility of identifying unintentional impacts by measuring the mass spectrometric properties of metabolites with unknown identities [131]. The application of metabolomics has proven invaluable in understanding how maize plants respond to various stress conditions i.e heat, salinity and drought. For example, a metabolomic investigation involving salt-tolerant and salt-sensitive maize genotypes revealed differences in metabolite accumulation in both roots and seedlings under salt stress. In seedlings, salt stress induced glucose and acid metabolism. Thirty common chemicals, including metabolites linked to basic metabolism such as deoxyadenosine, adenine, L-pyroglutamic acid, cis-9-palmitoleic acid, and galactinol compounds, were found in the roots of both salt sensitive and tolerant cultivars [132]. Heat stress effects on pollen male sterility in maize, especially at the most vulnerable tetrad stage, has also been clarified by metabolic pathway study. A reduction in pyruvate levels and an enhancement in sucrose levels were found by this research. In the meantime, other genes linked to signalling, unfolded protein stress, and auxin synthesis did not alter [133]. More importantly, a study by Ganie et al. [134] revealed metabolic pathways impacted by phosphorus stress situations, offering insights into strategies for improving phosphorus efficiency. The analysis, conducted using gas chromatography-mass spectroscopy, the investigation revealed a drop in fatty acids like cholesterol and stigmasterol, which are critical for membrane fluidity, and an increase in sugar alcohols like glucitol and mannitol under P-limitation, which are essential for membrane fluidity. In cases of severe phosphorus starvation, plants will scavenge phosphorus from these fatty acids, disrupting membrane fluidity. Additionally, elevated levels of serine and glycine indicated an increase in photorespiration rates [135].
8. Conclusions
The notable decrease in maize production resulting from various biotic and abiotic stresses raises substantial concerns. To address these challenges, farmers frequently turn to the application of chemical pesticides as they offer a rapid remedy. The negative consequences of widespread pesticide use on both human health and the environment have spurred the exploration of alternative pest control methods. Host plant resistance, recognized as an eco-friendly strategy, has emerged as a vital component within Integrated Pest Management (IPM) initiatives. Maize varieties with stress resistance or tolerance offer a sustainable and environmentally conscious means of managing pests. Although strides have been made in pinpointing resistance sources for both biotic and abiotic stress in crops, traditional approaches for crafting such resilient or tolerant maize varieties are arduous and time-consuming. This is mainly attributable to the intricate character of quantitative traits, which are influenced by numerous genetic loci. Nevertheless, contemporary biotechnological tools, particularly various omics techniques, present encouraging prospects for creating sustainable, multi-faceted resistance to both biotic and abiotic stresses. Omics methodologies are currently being harnessed to develop novel plant resistance attributes that provide robust protection against various stresses in maize cultivation. This entails the utilization of innovative molecules, resistant genes, and the alteration of gene expression. The anticipated progress in biotechnological innovations, encompassing genome editing, genetic transformation, and marker-assisted breeding, among other methods, is poised to expedite the development of disease-resistant crops, both in the present and in the times ahead. RNA interference and genome editing through CRISPR/Cas9 represent novel approaches for creating disease-resistant crops. Biotechnology has emerged as a valuable instrument for tackling the worldwide pest challenge, resulting in the production of economically efficient, pesticide-resistant, and environmentally friendly insect-resistant crops. When employed judiciously and ethically, biotechnology holds the promise of delivering substantial advantages.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Erenstein, O., Jaleta, M., Sonder, K., Mottaleb, K. and Prasanna, B.M. Global maize production, consumption and trade: Trends and R&D implications. Food Security 2022, 14, 1295–1319.
- Awika, J.M., 2011. Major cereal grains production and use around the world. In Advances in cereal science: implications to food processing and health promotion (pp. 1-13). American Chemical Society.
- FAOStat. (2022). FAO Stat. FAO, Rome. http:// www. fao. org/ faost at.
- Grote, U., Fasse, A., Nguyen, T.T. and Erenstein, O. Food security and the dynamics of wheat and maize value chains in Africa and Asia. Frontiers in Sustainable Food Systems 2021, 4, 617009. [CrossRef]
- Poole N, Donovan J, Erenstein O. Agri-nutrition research: Revisiting the contribution of maize and wheat to human nutrition and health. Food Policy. 2021, 100, 101976. [Google Scholar] [CrossRef]
- Serna-Saldivar SO, Perez-Carrillo E, Heredia-Olea E. Soybean-fortified wheat flour tortillas. InFlour and breads and their fortification in health and disease prevention 2019 Jan 1 (pp. 291-306). Academic Press.
- Tiwari, A., Choudhary, S., Padiya, J., Ubale, A., Mikkilineni, V. and Char, B., 2022. Recent Advances and Applicability of GBS, GWAS, and GS in Maize. Genotyping by Sequencing for Crop Improvement, pp.188-217.
- Walbot, V., 2008. Maize genome in motion.
- Schnable, P.S., Ware, D., Fulton, R.S., Stein, J.C., Wei, F., Pasternak, S., Liang, C., Zhang, J., Fulton, L., Graves, T.A. and Minx, P. The B73 maize genome: complexity, diversity, and dynamics. Science 2009, 326, 1112–1115.
- Jiao, Y., Peluso, P., Shi, J., Liang, T., Stitzer, M.C., Wang, B., Campbell, M.S., Stein, J.C., Wei, X., Chin, C.S. and Guill, K. Improved maize reference genome with single-molecule technologies. Nature 2017, 546, 524–527.
- Chapman, M.A., He, Y. and Zhou, M. Beyond a reference genome: pangenomes and population genomics of underutilized and orphan crops for future food and nutrition security. New Phytologist 2022, 234, 1583–1597. [CrossRef]
- Voss-Fels, K.P., Cooper, M. and Hayes, B.J. Accelerating crop genetic gains with genomic selection. Theoretical and Applied Genetics 2019, 132, 669–686. [CrossRef] [PubMed]
- Liu C, Li X, Meng D, Zhong Y, Chen C, Dong X, Xu X, Chen B, Li W, Li L, Tian X. A 4-bp insertion at ZmPLA1 encoding a putative phospholipase A generates haploid induction in maize. Molecular plant. 2017, 10, 520–522. [Google Scholar] [CrossRef]
- Piperno DR, Ranere AJ, Holst I, Iriarte J, Dickau R. Starch grain and phytolith evidence for early ninth millennium BP maize from the Central Balsas River Valley, Mexico. Proceedings of the National Academy of Sciences. 2009, 106, 5019–5024. [Google Scholar] [CrossRef]
- Ranere AJ, Piperno DR, Holst I, Dickau R, Iriarte J. The cultural and chronological context of early Holocene maize and squash domestication in the Central Balsas River Valley, Mexico. Proceedings of the National Academy of Sciences. 2009, 106, 5014–5018. [Google Scholar] [CrossRef] [PubMed]
- Alene AD, Menkir A, Ajala SO, Badu-Apraku B, Olanrewaju AS, Manyong VM, Ndiaye A. The economic and poverty impacts of maize research in West and Central Africa. Agricultural Economics. 2009, 40, 535–550. [Google Scholar] [CrossRef]
- Gedil M, Menkir A. An integrated molecular and conventional breeding scheme for enhancing genetic gain in maize in Africa. Frontiers in Plant Science. 2019, 10, 1430. [Google Scholar] [CrossRef]
- Lamichhane S, Thapa S. Advances from conventional to modern plant breeding methodologies. Plant breeding and biotechnology. 2022, 10, 1–4. [Google Scholar] [CrossRef]
- Johannsen, W. Heredity in populations and pure lines. Classic papers in genetics. 1903, 20–26. [Google Scholar]
- Poehlman, JM. Breeding field crops. Springer Science & Business Media; 2013 Apr 17.
- Begna, T. Conventional Breeding Methods Widely used to Improve Self-Pollinated Crops. International Journal of Research. 2021, 7, 1–6. [Google Scholar]
- Brown J, Caligari P. An introduction to plant breeding. John Wiley & Sons; 2011 Aug 26.
- Aleksoski, J. The effect of backcross method in tobacco breeding. Journal of Agriculture and Plant Sciences 2018, 16, 9–19. [Google Scholar]
- Vogel, KE. Backcross breeding. Transgenic Maize: Methods and Protocols. 2009, 161–169.
- Singh, SP. Alternative methods to backcross breeding. Annu. Rep. Bean Improv. Coop. 1982, 25, 11–12. [Google Scholar]
- Hull, FH. Recurrent selection for specific combining ability in corn 1. Agronomy Journal. 1945, 37, 134–145. [Google Scholar] [CrossRef]
- Ramya P, Singh GP, Jain N, Singh PK, Pandey MK, Sharma K, Kumar A, Prabhu KV. Effect of recurrent selection on drought tolerance and related morpho-physiological traits in bread wheat. PloS one. 2016, 11, e0156869. [Google Scholar]
- Bangarwa, S. 2021. Recurrent Selection - Definition and Types. Biotecharticles.com. 19 October.
- Schlegel, RH. History of plant breeding. CRC press; 2017 Dec 15.
- Acquaah, G. Principles of plant genetics and breeding. John Wiley & Sons; 2009 Mar 12.
- Bharti G, Chimata M K. Review on New Plant Breeding Techniques. International Journal of Science and Research 2019, 8, 723–730. [Google Scholar]
- Lema, M. Marker assisted selection in comparison to conventional plant breeding. Agric Res Technol. 2018, 14, 555914. [Google Scholar]
- Muntean L, Ona A, Berindean I, Racz I, Muntean S. Maize Breeding: From Domestication to Genomic Tools. Agronomy. 2022, 12, 2365. [Google Scholar] [CrossRef]
- Allard, RW. Principles of plant breeding. Soil Science. 1961, 91, 414. [Google Scholar] [CrossRef]
- Breseghello F, Coelho AS. Traditional and modern plant breeding methods with examples in rice (Oryza sativa L.). Journal of agricultural and food chemistry. 2013, 61, 8277–8286. [Google Scholar] [CrossRef] [PubMed]
- Stevens, R. Prospects for using marker-assisted breeding to improve maize production in Africa. Journal of the Science of Food and Agriculture. 2008, 88, 745–755. [Google Scholar] [CrossRef]
- Jones CJ, Edwards KJ, Castaglione S, Winfield MO, Sala F, Van de Wiel C, Bredemeijer G, Vosman B, Matthes M, Daly A, Brettschneider R. Reproducibility testing of RAPD, AFLP and SSR markers in plants by a network of European laboratories. Molecular breeding. 1997, 3, 381–390. [Google Scholar] [CrossRef]
- Winter P, Kahl G. Molecular marker technologies for plant improvement. World Journal of Microbiology and Biotechnology. 1995, 11, 438–448. [Google Scholar] [CrossRef] [PubMed]
- Mohan M, Nair S, Bhagwat A, Krishna TG, Yano M, Bhatia CR, Sasaki T. Genome mapping, molecular markers and marker-assisted selection in crop plants. Molecular breeding. 1997, 3, 87–103. [Google Scholar] [CrossRef]
- Bhullar NK, Zhang Z, Wicker T, Keller B. Wheat gene bank accessions as a source of new alleles of the powdery mildew resistance gene Pm3: a large scale allele mining project. BMC plant biology. 2010, 10, 1–3. [Google Scholar]
- Kumawat G, Kumawat CK, Chandra K, Pandey S, Chand S, Mishra UN, Lenka D, Sharma R. Insights into marker assisted selection and its applications in plant breeding. In Plant Breeding-Current and Future Views 2020 Nov 30. Intechopen.
- Ribaut JM, Hoisington D. Marker-assisted selection: new tools and strategies. Trends in Plant Science. 1998, 3, 236–239. [Google Scholar] [CrossRef]
- Hasan MM, Rafii MY, Ismail MR, Mahmood M, Rahim HA, Alam MA, Ashkani S, Malek MA, Latif MA. Marker-assisted backcrossing: a useful method for rice improvement. Biotechnology & Biotechnological Equipment. 2015, 29, 237–254. [Google Scholar]
- Ribaut JM, Ragot M. Marker-assisted selection to improve drought adaptation in maize: the backcross approach, perspectives, limitations, and alternatives. Journal of experimental botany. 2007, 58, 351–360. [Google Scholar]
- Wang X, Wang H, Liu S, Ferjani A, Li J, Yan J, Yang X, Qin F. Genetic variation in ZmVPP1 contributes to drought tolerance in maize seedlings. Nature genetics. 2016, 48, 1233–1241. [Google Scholar] [CrossRef] [PubMed]
- Prasanna BM, Palacios-Rojas N, Hossain F, Muthusamy V, Menkir A, Dhliwayo T, Ndhlela T, San Vicente F, Nair SK, Vivek BS, Zhang X. Molecular breeding for nutritionally enriched maize: status and prospects. Frontiers in genetics. 2020, 10, 1392. [Google Scholar] [CrossRef]
- Bhatt V, Muthusamy V, Jha S, Zunjare RU, Baveja A, Dosad S, Hossain F. Development of low phytic acid maize through marker assisted introgression of lpa1-1 and lpa2-1 genes. In Abstracts: 13th Asian Maize Conference on and Expert Consultation on Maize for Food, Feed, Nutrition and Environmental Security, Ludhiana, India, -10 2018 Oct 8 (pp. 143-144). 8 October.
- Goswami R, Zunjare R, Khan S, Baveja A, Muthusamy V, Hossain F. Marker-assisted introgression of rare allele of crtRB1 gene into elite quality protein maize inbred for combining high lysine, tryptophan and provitamin A in maize. Plant Breed. 2019, 138, 174–183. [Google Scholar] [CrossRef]
- Mehta BK, Muthusamy V, Zunjare RU, Baveja A, Chauhan HS, Chhabra R, Singh AK, Hossain F. Biofortification of sweet corn hybrids for provitamin-A, lysine and tryptophan using molecular breeding. Journal of Cereal Science. 2020, 96, 103093. [Google Scholar] [CrossRef]
- Prasanna BM, Palacios-Rojas N, Hossain F, Muthusamy V, Menkir A, Dhliwayo T, Ndhlela T, San Vicente F, Nair SK, Vivek BS, Zhang X. Molecular breeding for nutritionally enriched maize: status and prospects. Frontiers in genetics. 2020, 10, 1392. [Google Scholar] [CrossRef]
- Baveja A, Muthusamy V, Panda KK, Zunjare RU, Das AK, Chhabra R, Mishra SJ, Mehta BK, Saha S, Hossain F. Development of multinutrient-rich biofortified sweet corn hybrids through genomics-assisted selection of shrunken2, opaque2, lcyE and crtRB1 genes. Journal of Applied Genetics. 2021, 62, 419–429. [Google Scholar] [CrossRef]
- Das AK, Gowda MM, Muthusamy V, Zunjare RU, Chauhan HS, Baveja A, Bhatt V, Chand G, Bhat JS, Guleria SK, Saha S. Development of maize hybrids with enhanced vitamin-E, vitamin-A, lysine, and tryptophan through molecular breeding. Frontiers in plant science. 2021, 12, 659381. [Google Scholar] [CrossRef]
- Singh J, Sharma S, Kaur A, Vikal Y, Cheema AK, Bains BK, Kaur N, Gill GK, Malhotra PK, Kumar A, Sharma P. Marker-assisted pyramiding of lycopene-ε-cyclase, β-carotene hydroxylase1 and opaque2 genes for development of biofortified maize hybrids. Scientific Reports. 2021, 11, 12642. [Google Scholar] [CrossRef]
- Zhao X, Tan G, Xing Y, Wei L, Chao Q, Zuo W, Lübberstedt T, Xu M. Marker-assisted introgression of qHSR1 to improve maize resistance to head smut. Molecular breeding. 2012, 30, 1077–1088. [Google Scholar] [CrossRef]
- Li, P. Understanding Maize Biology for Better Crop Improvement. Molecular Plant. 2017, 10, 1014–1017. [Google Scholar] [CrossRef] [PubMed]
- Yang Q, He Y, Kabahuma M, Chaya T, Kelly A, Borrego E, Bian Y, El Kasmi F, Yang L, Teixeira P, Kolkman J. A gene encoding maize caffeoyl-CoA O-methyltransferase confers quantitative resistance to multiple pathogens. Nature Genetics. 2017, 49, 1364–1372. [Google Scholar] [CrossRef]
- Xu Z, Hua J, Wang F, Cheng Z, Meng Q, Chen Y, Han X, Tie S, Liu C, Li X, Wang Z. Marker-assisted selection of qMrdd8 to improve maize resistance to rough dwarf disease. Breeding Science. 2020, 70, 183–192. [Google Scholar] [CrossRef]
- Hossain F, Muthusamy V, Bhat JS, Zunjare RU, Kumar S, Prakash NR, Mehta BK. Maize Breeding. InFundamentals of Field Crop Breeding 2022 (pp. 221-258). Singapore: Springer Nature Singapore. 6 May.
- Wang X, Xu Y, Hu Z, Xu C. Genomic selection methods for crop improvement: Current status and prospects. The Crop Journal. 2018, 6, 330–340. [Google Scholar] [CrossRef]
- Sun Q, Wang P, Li W, Li W, Lu S, Yu Y, Zhao M, Meng Z. Genomic selection on shelling percentage and other traits for maize. Breeding science. 2019, 69, 266–271. [Google Scholar] [CrossRef] [PubMed]
- Meuwissen TH, Hayes BJ, Goddard M. Prediction of total genetic value using genome-wide dense marker maps. genetics. 2001, 157, 1819–1829. [Google Scholar] [CrossRef] [PubMed]
- Bernardo R, Yu J. Prospects for genomewide selection for quantitative traits in maize. Crop Science. 2007, 47, 1082–1090. [Google Scholar] [CrossRef]
- Madhusudhana, R. Marker-assisted breeding in sorghum. InBreeding sorghum for diverse end uses 2019 Jan 1 (pp. 93-114). Woodhead Publishing.
- Jannink JL, Lorenz AJ, Iwata H. Genomic selection in plant breeding: from theory to practice. Briefings in functional genomics. 2010, 9, 166–177. [Google Scholar] [CrossRef]
- Heffner EL, Sorrells ME, Jannink JL. Genomic selection for crop improvement. Crop Science. 2009, 49, 1–2. [Google Scholar] [CrossRef]
- Mayor PJ, Bernardo R. Genome wide selection and marker-assisted recurrent selection in doubled haploid versus F2 populations. Crop Science. 2009, 49, 1719–1725. [Google Scholar] [CrossRef]
- Massman JM, Jung HJ, Bernardo R. Genomewide selection versus marker-assisted recurrent selection to improve grain yield and stover-quality traits for cellulosic ethanol in maize. Crop Science. 2013, 53, 58–66. [Google Scholar] [CrossRef]
- Crossa J, Pérez-Rodríguez P, Cuevas J, Montesinos-López O, Jarquín D, De Los Campos G, Burgueño J, González-Camacho JM, Pérez-Elizalde S, Beyene Y, Dreisigacker S. Genomic selection in plant breeding: methods, models, and perspectives. Trends in plant science. 2017, 22, 961–975. [Google Scholar] [CrossRef]
- Wang T, Ma X, Li Y, Bai D, Liu C, Liu Z, Tan X, Shi Y, Song Y, Carlone M, Bubeck D. Changes in yield and yield components of single-cross maize hybrids released in China between 1964 and 2001. Crop Science. 2011, 51, 512–525. [Google Scholar] [CrossRef]
- Sun, G.; Yu, H.; Wang, P.; Guerrero, M.L.; Mural, R.V.; Mizero, O.N.; Grzybowski, M.; Song, B.; van Dijk, K.; Schachtman, D.P.; et al. A role for heritable transcriptomic variation in maize adaptation to temperate environments. BioRxiv 2022, 39. [Google Scholar] [CrossRef] [PubMed]
- Zaidi SK, Frietze SE, Gordon JA, Heath JL, Messier T, Hong D, Boyd JR, Kang M, Imbalzano AN, Lian JB, Stein JL. Bivalent epigenetic control of oncofetal gene expression in cancer. Molecular and cellular biology. 2017, 37, e00352–17. [Google Scholar]
- Osakabe K, Osakabe Y, Toki S. Site-directed mutagenesis in Arabidopsis using custom-designed zinc finger nucleases. Proceedings of the National Academy of Sciences. 2010, 107, 12034–12039. [Google Scholar] [CrossRef]
- Voytas DF, Gao C. Precision genome engineering and agriculture: opportunities and regulatory challenges. PLoS biology. 2014, 12, e1001877. [Google Scholar]
- Sander JD, Joung JK. CRISPR-Cas systems for editing, regulating and targeting genomes. Nature biotechnology. 2014, 32, 347–355. [Google Scholar] [CrossRef]
- Pegoraro C, Mertz LM, da Maia LC, Rombaldi CV, de Oliveira AC. Importance of heat shock proteins in maize. J Crop Sci Biotechnol. 2011, 14, 85–95. [Google Scholar] [CrossRef]
- Puchta H, Dujon B, Hohn B. Homologous recombination in plant cells is enhanced by in vivo induction of double strand breaks into DNA by a site-specific endonuclease. Nucl Acids Res. 1993, 21, 5034–5040. [Google Scholar] [CrossRef]
- Svitashev S, Schwartz C, Lenderts B, Young JK, Cigan AM. Genome editing in maize directed by CRISPR–Cas9 ribonucleoprotein complexes. Nat Commun. 2016, 7, 13274. [Google Scholar] [CrossRef] [PubMed]
- Joung JK, Sander JD. TALENs: a widely applicable technology for targeted genome editing. Nat Rev Mol Cell Biol. 2013, 14, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Jinek M, Chylinski K, Fonfara I, Hauer M, Doudna JA, Charpentier E. A programmable dual-RNA–guided DNA endonuclease in adaptive bacterial immunity. Science. 2012, 337, 816–821. [Google Scholar] [CrossRef] [PubMed]
- Gilbert LA, Larson MH, Morsut L, Liu Z, Brar GA, Torres SE, Stern-Ginossar N, Brandman O, Whitehead EH, Doudna JA, Lim WA. CRISPR-mediated modular RNA-guided regulation of transcription in eukaryotes. Cell. 2013, 154, 442–451. [Google Scholar] [CrossRef]
- O’Connell MR, Oakes BL, Sternberg SH, East-Seletsky A, Kaplan M, Doudna JA. Programmable RNA recognition and cleavage by CRISPR/Cas9. Nature. 2014, 516, 263–266. [Google Scholar] [CrossRef] [PubMed]
- Newman M, Ausubel FM. Introduction to gene editing and manipulation using CRISPR/Cas9 technology. Curr Protoc Mol Biol. 2016, 115, 1–6. [Google Scholar]
- Shan Q, Wang Y, Li J, Gao C. Genome editing in rice and wheat using the CRISPR/Cas system. Nat Protoc. 2014, 9, 2395–2410. [Google Scholar] [CrossRef] [PubMed]
- Liang Z, Zhang K, Chen K, Gao C. Targeted mutagenesis in Zea mays using TALENs and the CRISPR/Cas system. J Genet Genomics. 2014, 41, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Svitashev S, Young JK, Schwartz C, Gao H, Falco SC, Cigan AM. Targeted mutagenesis, precise gene editing, and site-specific gene insertion in maize using Cas9 and guide RNA. Plant Physiol. 2015, 169, 931–945. [Google Scholar] [CrossRef]
- Char SN, Neelakandan AK, Nahampun H, Frame B, Main M, Spalding MH, Becraft PW, Meyers BC, Walbot V, Wang K, Yang B. An Agrobacterium-delivered CRISPR/Cas9 system for high-frequency targeted mutagenesis in maize. Plant Biotechnol J. 2017, 15, 257–268. [Google Scholar] [CrossRef] [PubMed]
- Bortesi L, Fischer R. The CRISPR/Cas9 system for plant genome editing and beyond. Biotechnol Adv. 2015, 33, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Liang, Z., Chen, K., Zhang, Y., Liu, J., Yin, K., Qiu, J. L., et al. Genome editing of bread wheat using biolistic delivery of CRISPR/Cas9 in vitro transcripts or ribonucleoproteins. Nat. Protoc. 2018, 13, 413–430. [CrossRef]
- Pineda, M., Lear, A., Collins, J. P., Kiani, S. Safe CRISPR: challenges and possible solutions. Trends Biotechnol. 2019, 37, 389–401. [CrossRef] [PubMed]
- Garcia Ruiz, M. T., Knapp, A. N., Garcia-Ruiz, H. Profile of genetically modified plants authorized in Mexico. GM Crops Food 2018, 9, 152–168. [CrossRef] [PubMed]
- Eckerstorfer, M. F., Engelhard, M., Heissenberger, A., Simon, S., Teichmann, H. Plants developed by new genetic modification techniques—comparison of existing regulatory frameworks in the EU and non-EU countries. Front. In Bioeng. Biotechnol. 2019, 7, 26. [CrossRef]
- Brookes, G., and Barfoot, P. GM Crops: Global Socio-economic and Environmental Impacts 1996–2018. 2020. Available online: https://pgeconomics.co.uk/ pdf/globalimpactfinalreportJuly2020.pdf (accessed on 28 August 2021).
- ISAAA Brief (2019). Global Status of Commercialized Biotech/GM Crops in 2019. ISAAA Ithaca: NY.
- ISAAA database (2021). GM Approval Database. Available online: https://www. isaaa.org/gmapprovaldatabase/default.asp (accessed on 10 August 2021).
- . Simmons, C. R., Lafitte, H. R., Reimann, K. S., Brugière, N., Roesler, K., Albertsen, M. C., et al. Successes and insights of an industry biotech program to enhance maize agronomic traits. Plant Sci. 2021, 307, 110899. [CrossRef]
- Wang A, Shu X, Jing X, Jiao C, Chen L, Zhang J, Ma L, Jiang Y, Yamamoto N, Li S, Deng Q. Identification of rice (Oryza sativa L.) genes involved in sheath blight resistance via a genome-wide association study. Plant Biotechnology Journal. 2021, 19, 1553–1566. [Google Scholar] [CrossRef]
- Kumar V, Singh A, Mithra SA, Krishnamurthy SL, Parida SK, Jain S, Tiwari KK, Kumar P, Rao AR, Sharma SK, Khurana JP. Genome-wide association mapping of salinity tolerance in rice (Oryza sativa). DNA research. 2015, 22, 133–145. [Google Scholar] [CrossRef]
- Alam MA, Rahman M, Ahmed S, Jahan N, Khan MA, Islam MR, Alsuhaibani AM, Gaber A, Hossain A. Genetic variation and genotype by environment interaction for agronomic traits in maize (Zea mays L.) hybrids. Plants. 2022, 11, 1522. [Google Scholar] [CrossRef]
- Tabashnik, B. E., and Carrière, Y. Surge in insect resistance to transgenic crops and prospects for sustainability. Nat. Biotechnol. 2017, 35, 926–935. [CrossRef]
- Pellegrino, E. , Bedini, S., Nuti, M., and Ercoli, L. Impact of genetically engineered maize on agronomic, environmental and toxicological traits: a metaanalysis of 21 years of field data. Sci. Rep. 2018, 8, 3113. [CrossRef]
- Schellenberger, U., Oral, J., Rosen, B. A., Wei, J. Z., Zhu, G., Xie, W., et al. A selective insecticidal protein from pseudomonas for controlling corn rootworms. Science 2016, 354, 634–637. [CrossRef]
- Moar, W., Khajuria, C., Pleau, M., Ilagan, O., Chen, M., Jiang, C., et al. Cry3Bb1-resistant western corn rootworm, diabrotica virgifera virgifera (LeConte) does not exhibit cross-resistance to DvSnf7 dsRNA. PLoS One 2017, 12, 1–15. [CrossRef]
- Yin, Y., Flasinski, S., Moar, W., Bowen, D., Chay, C., Milligan, J., et al. A new bacillus thuringiensis protein for Western corn rootworm control. PLoS One 2020, 15, 1–16. [CrossRef]
- Wu, J., Lawit, S. J., Weers, B., Sun, J., Mongar, N., Van Hemert, J., et al. Overexpression of zmm28 increases maize grain yield in the field. Proc. Natl. Acad. Sci. 2019, 116, 23850–23858. [CrossRef]
- Simmons, C. R., Weers, B. P., Reimann, K. S., Abbitt, S. E., Frank, M. J., Wang, W., et al. Maize BIG GRAIN1 homolog overexpression increases maize GRAIN yield. Plant Biotechnol. J. 2020, 18, 2304–2315. [CrossRef]
- Shi, J., Habben, J. E., Archibald, R. L., Drummond, B. J., Chamberlin, M. A., Williams, R. W., et al. Overexpression of ARGOS genes modifies plant sensitivity to ethylene, leading to improved drought tolerance in Both Arabidopsis and maize. Plant Physiol. 2015, 69, 266–282. [CrossRef]
- Woźniak, E., Tyczewska, A., and Twardowski, T. A shift towards biotechnology: social opinion in the EU. Trends Biotechnol. 2021, 39, 214–218. [CrossRef]
- Cardi, T. Cisgenesis and genome editing: combining concepts and efforts for a smarter use of genetic resources in crop breeding. Plant Breed. 2016, 135, 139–147. [Google Scholar] [CrossRef]
- Harfouche, A. L., Petousi, V., Meilan, R., Sweet, J., Twardowski, T., and Altman, A. Promoting ethically responsible use of agricultural biotechnology. Trends Plant Sci. 2021, 26, 546–559. [CrossRef]
- Li, Q.; Yan, J. Sustainable agriculture in the era of omics: Knowledge-driven crop breeding. Genome Biol. 2020, 21, 154. [Google Scholar] [CrossRef] [PubMed]
- Somegowda, V.K.; Rayaprolu, L.; Rathore, A.; Deshpande, S.P.; Gupta, R. Genome-Wide Association Studies (GWAS) for Traits Related to Fodder Quality and Biofuel in Sorghum: Progress and Prospects. Protein Pept. Lett. 2021, 28, 843–854. [Google Scholar] [CrossRef] [PubMed]
- Al-Qurainy, F.; Alshameri, A.; Gaafar, A.-R.; Khan, S.; Nadeem, M.; Alameri, A.A.; Tarroum, M.; Ashraf, M. Comprehensive Stress-Based De Novo Transcriptome Assembly and Annotation of Guar (Cyamopsis tetragonoloba (L. ) Taub.): An Important Industrial and Forage Crop. Int. J. Genomics 2019, 2019, 7295859. [Google Scholar] [PubMed]
- Abdurakhmonov, I.Y. Plant Genomics; InTech: London, UK, 2016; ISBN 978-953-51-2455-9. [Google Scholar]
- Pérez-de-Castro, A.M.; Vilanova, S.; Cañizares, J.; Pascual, L.; Blanca, J.M.; Díez, M.J.; Prohens, J.; Picó, B. Application of genomic tools in plant breeding. Curr. Genom. 2012, 13, 179–195. [Google Scholar] [CrossRef] [PubMed]
- Vinayan, M.T.; Seetharam, K.; Babu, R.; Zaidi, P.H.; Blummel, M.; Nair, S.K. Genome wide association study and genomic prediction for stover quality traits in tropical maize (Zea mays L.). Sci. Rep. 2021, 11, 686. [Google Scholar] [CrossRef]
- Cramer, G.R.; Urano, K.; Delrot, S.; Pezzotti, M.; Shinozaki, K. Effects of abiotic stress on plants: A systems biology perspective. BMC Plant Biol. 2011, 11, 163. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, X. Transcriptome and metabolomic analyses reveal regulatory networks controlling maize stomatal development in response to blue light. Int. J. Mol. Sci. 2021, 22, 5393. [Google Scholar] [CrossRef]
- Pan Y, Zhao SW, Tang XL, Wang S, Wang X, Zhang XX, Zhou JJ, Xi JH. Transcriptome analysis of maize reveals potential key genes involved in the response to belowground herbivore Holotrichia parallela larvae feeding. Genome. 2020, 63, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Pan Y, Zhao S, Wang Z, Wang X, Zhang X, Lee Y, Xi J. Quantitative proteomics suggests changes in the carbohydrate metabolism of maize in response to larvae of the belowground herbivore Holotrichia parallela. PeerJ. 2020, 8, e9819. [Google Scholar] [CrossRef]
- Zhou, K.; Zeng, X.; Zhang, B.; Aslam, M.; Xin, H.; Liu, W.; Zou, H. Bulk segregant transcriptome analysis based differential expression of drought response genes in maize. Pak. J. Agric. Sci. 2020, 57, 909–923. [Google Scholar]
- Kebede, A.Z.; Johnston, A.; Schneiderman, D.; Bosnich, W.; Harris, L.J. Transcriptome profiling of two maize inbreds with distinct responses to Gibberella ear rot disease to identify candidate resistance genes. BMC Genom. 2018, 19, 131. [Google Scholar] [CrossRef]
- He, W.; Zhu, Y.; Leng, Y.; Yang, L.; Zhang, B.; Yang, J.; Zhang, X.; Lan, H.; Tang, H.; Chen, J.; et al. Transcriptomic analysis reveals candidate genes responding maize gray leaf spot caused by Cercospora zeina. Plants 2021, 10, 2257. [Google Scholar] [CrossRef]
- Wu, X.; Wang, B.; Xie, F.; Zhang, L.; Gong, J.; Zhu, W.; Li, X.; Feng, F.; Huang, J. QTL mapping and transcriptome analysis identify candidate genes regulating pericarp thickness in sweet corn. BMC Plant Biol. 2020, 20, 117. [Google Scholar] [CrossRef] [PubMed]
- Suhre, K.; McCarthy, M.I.; Schwenk, J.M. Genetics meets proteomics: Perspectives for large population-based studies. Nat. Rev. Genet. 2021, 22, 19–37. [Google Scholar] [CrossRef] [PubMed]
- Mustafa, G.; Komatsu, S. Plant proteomic research for improvement of food crops under stresses: A review. Mol. Omics 2021, 17, 860–880. [Google Scholar] [CrossRef]
- Guo, J.; Li, C.; Zhang, X.; Li, Y.; Zhang, D.; Shi, Y.; Song, Y.; Li, Y.; Yang, D.; Wang, T. Transcriptome and GWAS analyses reveal candidate gene for seminal root length of maize seedlings under drought stress. Plant Sci. 2020, 292, 110380. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.T.; Zhang, Y.L.; Chen, S.X.; Yin, G.H.; Yang, Z.Z.; Lee, S.; Liu, C.G.; Zhao, D.D.; Ma, Y.K.; Song, F.Q.; et al. Proteomics of methyl jasmonate induced defense response in maize leaves against Asian corn borer. BMC Genom. 2015, 16, 224. [Google Scholar] [CrossRef]
- Wang J, Lin Z, Zhang X, Liu H, Zhou L, Zhong S, Li Y, Zhu C, Lin Z. krn1, a major quantitative trait locus for kernel row number in maize. New Phytologist. 2019, 223, 1634–1646. [Google Scholar] [CrossRef] [PubMed]
- Dong, A.; Yang, Y.; Liu, S.; Zenda, T.; Liu, X.; Wang, Y.; Li, J.; Duan, H. Comparative proteomics analysis of two maize hybrids revealed drought-stress tolerance mechanisms. Biotechnol. Biotechnol. Equip. 2020, 34, 763–780. [Google Scholar] [CrossRef]
- Führs, H.; Götze, S.; Specht, A.; Erban, A.; Gallien, S.; Heintz, D.; Van Dorsselaer, A.; Kopka, J.; Braun, H.-P.; Horst, W.J. Characterization of leaf apoplastic peroxidases and metabolites in Vigna unguiculata in response to toxic manganese supply and silicon. J. Exp. Bot. 2009, 60, 1663–1678. [Google Scholar] [CrossRef]
- Kaur, B.; Sandhu, K.S.; Kamal, R.; Kaur, K.; Singh, J.; Röder, M.S.; Muqaddasi, Q.H. Omics for the improvement of abiotic, biotic, and agronomic traits in major cereal crops: Applications, challenges, and prospects. Plants 2021, 10, 1989. [Google Scholar] [CrossRef] [PubMed]
- Yue, J.Y.; Wang, L.H.; Dou, X.T.; Wang, Y.J.; Wang, H.Z. Comparative metabolomic profiling in the roots of salt-tolerant and salt-intolerant maize cultivars treated with NaCl stress. Biol. Plant. 2020, 64, 569–577. [Google Scholar] [CrossRef]
- Begcy, K.; Nosenko, T.; Zhou, L.-Z.; Fragner, L.; Weckwerth, W.; Dresselhaus, T. Male sterility in maize after transient heat stress during the tetrad stage of pollen development. Plant Physiol. 2019, 181, 683–700. [Google Scholar] [CrossRef]
- Ganie, A.H.; Ahmad, A.; Pandey, R.; Aref, I.M.; Yousuf, P.Y.; Ahmad, S.; Iqbal, M. Metabolite Profiling of Low-P Tolerant and Low-P Sensitive Maize Genotypes under Phosphorus Starvation and Restoration Conditions. PLoS ONE 2015, 10, e0129520. [Google Scholar] [CrossRef]
Figure 1.
A visual model depicting the genome modification process of the CRISPR/Cas9 system.
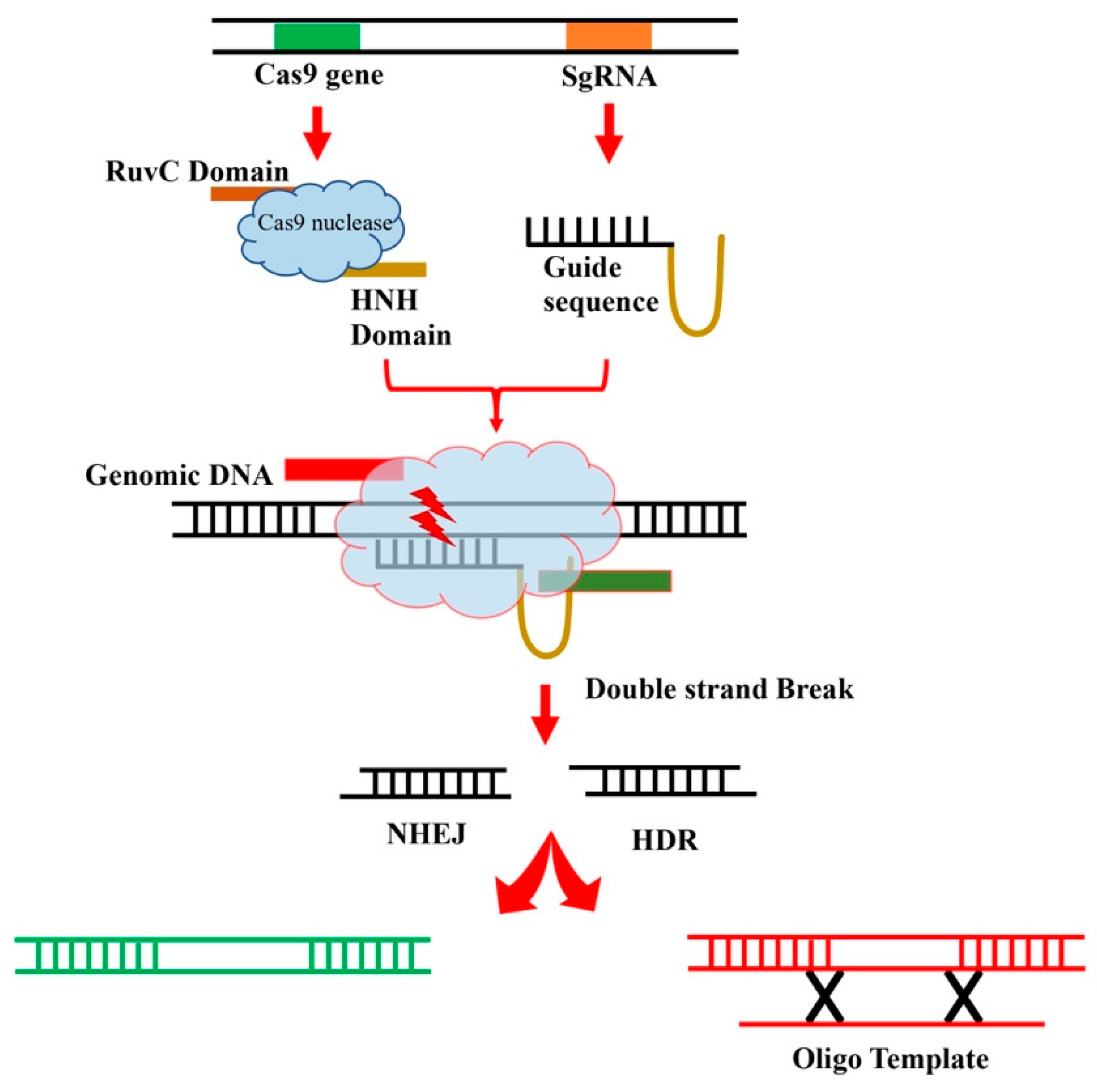
Figure 2.
Representation of different omics approaches used for maize crop improvement.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



