
Submitted:
29 September 2023
Posted:
01 October 2023
You are already at the latest version
Abstract
The Iberian wolf (Canis lupus signatus) is recolonizing historical distribution areas after decades of absence. As in other human-dominated landscapes, finding a balance to protect this species by favoring recolonization and mitigating human-wildlife conflicts is a challenge. Since wolves are often generalist opportunistic predators, we studied their diet composition in the Central Spain to evaluate the consumption of domestic ungulates and provide reliable data that help local authorities to deal with the current wolf-cattle ranchers conflict and coexistence. Diet composition (% prey occurrence, % prey ingested biomass) was analyzed through the identification of prey hairs present in 671 scats collected between 2017 and 2021. Wolves fed more on wild ungulates (82% occurrence) than domestic ones (18%). Wild boar (Sus scrofa, 44% occurrence) and roe deer (Capreolus capreolus, 35%) were the most consumed prey. Wolves positively selected these two species. The wolf's diet varied between seasons, years, and forest regions, but a diet based on wild ungulates predominated over domestic ones. Food niche breadth and showed variations depending on seasons and years. Preserving the availability and diversity of wild ungulates may favor reducing livestock attacks and would be an achievable goal that help to conserve this species and reduce conservation conflicts.
Keywords:
Canis lupus signatus
; domestic ungulates
; wild ungulates
; Iberian wolf
; recolonization
1. Introduction
Habitat loss and fragmentation are scenarios often associated with anthropic factors such as infrastructures and human activities [1,2,3,4]. These scenarios entail the decrease or disappearance of populations and the contraction of large carnivores’ ranges in areas where they were previously widely distributed [5]. The Grey wolf (Canis lupus lupus) is an example of these carnivores with completely disappeared populations in some regions of Europe in the 18th century [6,7], due to direct persecution and prey abundance decrease [8]; which is recolonizing semi-desert and agricultural lands in many industrialized countries in recent years [9]. This European recolonization has been favored by endowing the wolf with a strict protection status from Council Directive 92/43/EEC of the EU Habitats Directive, the conservation strategies of the European Commission such as the European Life Program (European Commission, 2020), and the habitat restoration strategies of some countries [10,11,12], constituting a relevant milestone in the rewilding process [13].
Overall, the wolf’s diet is broadly influenced by the structure of prey communities. Studies performed in several European countries showed a diversity of species in their diet: red deer, roe deer and wild boar in Poland [14]; roe deer and wild boar in Italy [15]; moose (Alces alces) predominantly and Eurasian beaver (Castor fiber), European badger (Meles meles) and brown hare (Lepus europaeus) as more sporadic consumption in the Scandinavian Peninsula [16]; roe deer, red deer, wild boar, mouflon and brown hares in Germany [17]. In some parts of the world, the gray wolf feeds on smaller prey species such as rodents, birds, and invertebrates [18,19]. Additionally, the species can be an opportunistic predator depending on the group and body size of the prey [20,21]. The wolf feeds mainly on medium and large ungulates that coexist in its distribution range [22], becoming skilled in the selective hunting of a particular species depending on its availability in each habitat [23,24]. Furthermore, diet is also influenced by various factors such as genetic structure of populations, prey ecology, climate change, recreational hunting, and agricultural policies [25,26,27,28]. It should be noted that the wolf, as a top predator, plays a fundamental role in maintaining balance in its ecosystem, since it modulates the abundance of mesocarnivores and large herbivores. Thus, an uncontrolled increase in ungulates due to removal of the top predator from the ecosystem can lead to a loss of biodiversity [8](Ripple et al. 2014). If, on the other hand, the population of wild ungulates were to be drastically reduced, wolves could become interested in more abundant and easily preyed domestic prey, leading to a human-wildlife conflict [29,30].
In the Iberian Peninsula, food resources availability is distributed in a variable way, mainly influenced by an anthropogenic use of the land that are carried out in each area [31,32,33]. For this reason, the feeding habits of the wolf are highly variable depending on the area studied [23,34]. Wolves coexist with humans in Spain, as in other Western European countries [33,35]. Although wolves tend to avoid anthropic areas, they benefit from resources associated with humans, such as livestock [36]. Predation on domestic livestock is often associated with areas where wild ungulates abundance has been relatively low for decades [37]. However, a study conducted on the north face of the Central System Mountain range did not find an association between a greater abundance of wild ungulates and a reduction in attacks on livestock [12]. Nowadays, the wolf is expanding towards the southern face of the Central System Mountain range, located in Comunidad Autónoma de Madrid [38]. These are areas recently recolonized where more relaxed extensive livestock farming practices are carried out and where protection against predators is scarcer than in other areas where wolves have inhabited for years [11]. Therefore, the objective of this study was to examine the feeding habits of the wolf by analyzing its diet composition, the diversity of prey in its habitat and niche breadth in Sierra de Guadarrama National Park, Sierra del Rincón and surroundings (located to the south of the Duero River) using a non-invasive methodology. The hypotheses and predictions proposed were:
i) As wild ungulates are abundant in the study area, wolves would mainly feed on these prey types, as occurs in most studies carried out in Europe [7,22]. Considering the results of various studies [39,40,41,42], wolves will select wild ungulates.
ii) Among wild ungulates, the wild board (Sus scrofa) has large litters compared to the roe deer with one or two calves (Capreolus capreolus, [43]). Autumn censuses have shown a well-distributed population throughout the pine and oak forests in the study area ([44], Comunidad de Madrid, unpublished data). Therefore, wild boar is expected to be the most consumed wild prey, as revealed in many studies carried out in Europe [17,45].
iii) Considering that cattle (Bos taurus) are the predominant livestock species in the study area, it is forecast to be the most consumed prey among domestic ungulates, especially calves.
iv) Based on data from previous studies showing wolf’s selection of certain prey [11,23], it is expected a narrow niche breadth and, therefore, a specialist diet.
v) Environmental factors that have been previously reported may drive the consumption of different prey species [46,47], thus, season and year are predicted to potentially influence wolf diet in the study area. We predict that in spring the consumption of wild ungulates will be higher than domestic ones due to the greater availability of young prey of different species. In addition, we also expect similar consumption of wild and domestic ungulates in all years because neither wild ungulate abundance nor livestock management changed substantially.
vi) As wolf´s diet variations were previously observed between localities or regions within an area [27,48], we expected variations in wolves’ feeding habits between forest regions of the study area. We expect a high consumption of domestic ungulates in the regions where more attacks on livestock were reported.
2. Materials and Methods
2.1. Study Area
We conducted the study in the Sierra de Guadarrama National Park (PNSG, declared by the Spanish Law 7/2013, of 25 June), Sierra del Rincón Biosphere Reserve (Designated by UNESCO on June 29 2005, and expanded in 2022) and bordering areas (Figure 1). This area covered the territory of several packs of Iberian wolves [Barja et al. unpublished data], a protected species (Law 42/2007) whose conservation is a priority (Directive 97/62/EC). The sampled surface was a mountain range with an extension of 100,775 ha and presents strong slopes and discontinuities, as well as numerous perennials and temporary watercourses. In this area, the average annual temperature ranged from -3.2ºC to 22.4ºC with annual average rainfall records of 1,223 mm (AEMet, www.opendata.aemet.es). So, climate was described as a continental Mediterranean climate, with dry and temperate summer and cold and humid winter. The study area contained different forest regions, delimited by geographic, ecological, forestry and socioeconomic criteria; established for the adequate planning and execution of the actions that articulate the management of mountains, forests, and cattle trails (Consejería de Medio Ambiente y Ordenación del Territorio 2007). The forest regions included in the study area were: PN Peñalara, Lozoya, Buitrago, Montejo, PRCAM Norte, El Espinar, Navafría y Riaza.
Scotch pine (Pinus sylvestris L.) is the predominant forest species (35.4%) between 1,200-1,900 m, unlike the vegetation located at 1,100 m where Pyrenean oak (Quercus pyrenaica Willd.; 15.8%) is the most abundant species. In addition, to high mountain pastures (23.9%), the study area also had an undergrowth (24.9%) formed by Cytisus oromediterraneus C. purgans auct. non (L.) Spach, Common juniper (Juniperus communis L.), Common holly (Ilex aquifolium L.) and Adenocarpus hispanicus (Lam.) DC. The predominant wild fauna species, that are susceptible to be preyed by wolf, were: wild boar, roe deer, and mountain goat (Capra pyrenaica). The ungulates species abundance in the area, included densities ranging from 3-5 ind/km2 of wild boars, 3-6 ind/km2 of roe deer, and high mountain enclaves with 15-36 ind/km2 of mountain goats (Comunidad de Madrid, unpublished data). However, domestic ungulates were also present in the study area, especially in summer, which takes advantage of pastures coming from a multitude of livestock huts such as cattle, goat (Capra aegagrus hircus), sheep (Ovis orientalis aries) and horse. The number of domestic ungulates in the area was 100,793 animals, of which 58,454 individuals corresponded to cattle and the rest to goats and sheep [INE 2020].
2.2. Protection Measures for the Iberian Wolf and Its Conservation Conflicts
The presence of wolves in Spain was hardly recorded throughout the 20th century, reaching the minimum occurrence in the 1970s. At this time, protection measures began to be applied and favored the expansion of the remaining population in the mountains of the northeast and the recolonization of the southeast of Spain, reaching the south of the Duero River in 1990[49,50,51]. The EU Habitats Directive granted the strict protection status of Annex IV to wolf populations inhabiting in the south of the Duero River, while populations located in the north of the river benefit from a more flexible status of Annex V [52,53]. This means that northern populations can be legally hunted with restrictions, while southern populations are fully protected. The different protection on each side of the Duero River is due to the different damage caused by the wolf to livestock depending on the area [53]. However, after reviewing the situation of the Iberian wolf, the Spanish Government considered that, although this species did not meet the requirements to qualify as “vulnerable”, it was appropriate to equalize its legal status throughout Spain. Thus, the Spanish Government included the wolf in the list of protected species and prohibited its hunting throughout the country in September 2021, and it may be hunted exceptionally if it has been shown that other “preventive or wildlife protection” measures have been “adequately” applied and these have been “ineffective” (Order TED/980/2021, RD 239/2011).
Wolf damage to livestock is a constant source of socioeconomic conflict according to some authors [54,55,56]. However, this problem must be considered a conservation challenge since the wolf are not knowingly antagonists in conflict [57]. Livestock consumption (17.8-38.9 % occurrence) was demonstrated by dietary studies carried out at the end of the 20th century in the northwestern mountains (north of the Duero River), which are highly populated and intensively used areas where livestock in extensive grazing conditions is habitual and the presence of wolves is considered high [58,59,60,61]. Recent studies, also carried out in the north of the Duero River, place the consumption of livestock between 10.6-62.3% of occurrence, varying noticeably between areas [23,34]. However, the recolonization of the southern Duero River is currently being studied and yielding mixed results. For example, an investigation carried out in central Portugal determined that wolf´s diet depended on domestic livestock by more than 90%, a result that the authors related with the low diversity and density of wild ungulates [11]. Conversely, results from other research conducted in central Spain (Segovia) revealed that at higher elevations cattle were subjected to increased attacks irrespective of wild prey abundance [12]. In any case, wolf attacks substantially increased to the south of the Douro River from 2007 to 2017, while the increase was only moderate in the north. In 2017, for example, >73% of all attacks (n = 1989) occurred in the south (Junta de Castilla-León 2017).
2.3. Collection of Faecal Samples
The collection of faecal samples in the field is a non-invasive and affordable technique that allows the collection of many samples to perform a qualitative and quantitative diet analysis [23]. The collection of wolf faecal samples was carried out by establishing 15 itineraries along forest trails and firebreaks, places where wolves move and deposit their scats, either with a marking function or as simple excretion [62,63,64]. Since the probability of defecation at cross-roads is higher [62,65], these were also included in the sampling itineraries. In each itinerary, the presence of the species was recorded using signs of its activity (scats, tracks, and/or scratches). The surveys were carried out on foot and the average length of each itinerary was 3,12 ± 0.44 km (range between 2.47-4.60 km). The samplings were carried out monthly for five years (2017-2021), obtaining a total of 671 scats in individual bags which were identified with a numerical code and recorded by a portable GPS devise [66]. The samplings were refrigerated in the laboratory until their subsequent analysis.
2.4. Identification of Wolf Prey Species
The diet of the wolves was determined from the analysis of the hair found in the scats collected in the study area. To do this, 5 to 8 hairs strands were collected from each faecal sample to be washed in a petri dish with soap and water. After rinsing with water, the hair was left drying on filter paper. The hairs selected from each sample were identified at the species level based on their macroscopic and microscopic characteristics [66](Ferretti et al. 2019). The macroscopic characteristics, comprising the coloration, shape, length, and thickness of the hair, were observed in a binocular magnifying glass (model Olympus TL2 SZ30). For the correct identification, a comparative study was carried out with hair of known species from the personal collection of Dr Isabel Barja. The microscopic characteristics (shape, arrangement, margin, and distance between margins of the scales) correspond to the cuticular pattern of the hair (which varies between species, [67]. To observe these cuticular patterns, the dry hair was fixed on a slide spraying a thin and homogeneous layer of hairspray. Each hair was placed leaving a free end that would facilitate the detachment of the hairspray layer after 10-15 min, enough time to leave the cuticular pattern imprinted in a microscope slide. Cuticular patterns were observed using an optical microscope (Olympus CX41 model) attached to a camera (Color view); we took photographs at different magnifications (10x, 20x, 40x) with the Soft Imaging system five software (AnalySIS getIT). The prey species identification was made with a manual on macroscopic and cuticular patterns of mammalian hairs from the Iberian Peninsula published by Barja et al. [67].
2.5. Mapping Using Kernel Densities
A Kernel density map was made to differentiate the areas with the highest concentration of wolf scats within the study area, considering the number of collected scats. In this way we could know the possible number of reproductive packs of Iberian wolves in study the area which is useful for the management of a newly recolonized area. Zub et al. [68] showed the overlaps of home ranges of the 4 wolf packs in Poland, comparing the areas comprising 75% of fecal mark locations (plotted using the Kernel method) with the distribution of radiolocations. Kernel density was calculated by quantifying the relationships of points within a radius of influence by analyzing patterns of a specific data set. The place of the occurrences was recorded by means of a coordinate system that allowed a count of all the points within a region of influence to be weighted by the distance of each one from the place of interest. The density of each region of the study area was calculated by interpolation. Interpolation made it possible to build a continuous surface of the variables (a smoothed surface), inferring the spatial variation of the variable for the entire study area, even in regions where the process has not generated any real occurrence, allowing to verify possible data trends [69,70,71]. Kernel density is based on two defining parameters: the radius of influence (R) and the estimation function (k). The radius of influence was defined as the area centered on the estimation point P that indicates how many events contribute to the estimate of the intensity function λ. The estimation function (k) takes care of the properties to smooth the density calculated by the Kernel technique and was calculated through the formula:
where k is a bivariate and symmetric Kernel function called the estimation or smoothing function and the parameter R > 0 is known as the width of the band (or radius of influence) and determines the degree of smoothing. This is the radius of a disk centered at P (P represents any location in R), where Pi will contribute significantly [70,71,72,73].
2.6. Statistical Analysis
The results are showed as frequency and percentage of occurrence, and ingested biomass. The frequency of occurrence of the prey species was determined by counting the number of scats that presented hair of each prey species. The ingested biomass (in kg and %) was calculated based on the average weight of each prey species (Table 1) and using the equation of Floyd et al. [74], revised and adjusted by Weaver [75] (see Appendix A), already used in studies on wolf diet in the Iberian Peninsula [11,76,77].
Subsequently, we ran a goodness-of-fit chi-square test (χ2) to verify the adjustment between observed and expected frequencies of consumed prey species hypothesis. In addition, we used contingency tables to evaluate differences in the relative frequencies of prey species in relation to the seasonality and years. We used the Pearson’s χ2 test (in 2 x 2 tables, where df. = 1, we applied Yates’ continuity correction) for cases in which less than 20% of the expected frequencies in the table were less than 5 records, and, for cases in which more than 20% of the expected frequencies in the table were less than 5 records; additionally, we used the Monte Carlo exact test (Fisher’s exact statistic was used in 2 x 2 tables and in the rest of the cases the χ2 statistic).
Additionally, we calculated the Shannon diversity index to estimate dietary diversity according to seasonality and years (see Appendix A). We tested significant differences between pairs of Shannon indices using Hutcheson’s t-statistic. Hutcheson’s t-test is a modified version of the classical t-test that provides a way to compare two samples using the variance of the Shannon index [78](Hutcheson 1970).
We estimated niche breadth of the wolf in terms of diet resources according to the frequency of occurrence of prey consumed and the biomass ingested over the seasons and years. We used the Levin’s food niche breadth Index (FNB) [79](see Appendix A) to quantitatively measure specialization in the composition of the wolf’s diet.
Finally, we calculated the Ivlev’s electivity index modified by Jacobs [80] to assess whether wolves selected preys positively or negatively (see Appendix A). This index was applied to evaluate the selection of prey throughout the study area and secondly, to evaluate the selection among domestic ungulates by forest regions. The level of significance to reject the null hypothesis was p<0.05. Statistical tests were carried out using SPSS v.23.00 (SPSS Inc, Chicago, IL, U.S.A.).
3. Results
3.1. General Remarks
The analysis of 671 scats showed that the wolf consumed more wild ungulates compared to domestic ones (Table 1). On the one hand, differences in the consumption of different prey species were statistically significant in relation to the percentage of occurrence (χ2=1282.56; df=8; p=0.001; n=671). Specifically, the wild ungulates most consumed were wild boar and roe deer (χ2=476.69; df=3; p=0.001; n=550; Table 1). Among domestic ungulates, cattle were the prey most predated (χ2=64.08; df=4; p=0.001; n=121; Table 1). In few occasions, the small amount of hair and its fragmentation in the sample led to doubts between two domestic species. Consequently, to calculate the biomass ingested (kg and %), we did not consider the occurrence obtained of these unidentified domestic samples. Overall, the biomass percentage provided by domestic ungulates was slightly higher (Table 1).
Ivlev’s index showed that wolves positively selected the wild board (D=0.92) and the roe deer (D=0.89) and avoided the mountain goat (D= -0.27) and domestic ungulates (cattle: D= -0.84; sheep and goat: D= -0.80). There was a higher consumption of roe deer from 2017 to 2019 than wild boar, but from 2020 to 2021 wild boar was more frequently found in wolf’s scats (roe deer frequency in scats: mean=56.2; SD=47.42), (wild boar frequency in scats: mean=43.2; SD=33.95). Furthermore, wild boar represented a greater biomass contribution to wolfs diet with 186,6 kg on average; SD=146.62, while roe deer contributed with 95.73 Kg on average; SD=80.80 on an annual basis.
3.2. Seasonal Trends
The consumption of wild ungulates was greater than on domestic ungulates in all seasons. On the one hand, the highest percentage of occurrence of wild ungulates was in summer, while the lowest percentage was in autumn, contrary to the occurrence trend of domestic ungulates (Figure 2A). The percentage of occurrence of roe deer was the highest with respect to the rest of the species in all seasons except in autumn, when wild boar was the predominant species. Regarding domestic ungulates, cattle was the most frequent species in all seasons (see Appendix B). Seasonal differences in the percentage of occurrence of the different prey species were statistically significant (χ2=66.07; df=24; p=0.001; n=637; see Suplementary Material 2). On the other hand, ingested biomass of wild ungulates was higher in winter than in the rest of the seasons, while ingested biomass corresponding to domestic ungulates was higher in autumn compared to other seasons (Fig 2B). The wild boar contributed the most biomass to the wolf´s diet in winter while cattle did it in the rest of seasons (Table 2).
The diversity in the diet of the wolf varied according to seasonality, being higher in autumn (H’=1.42), followed by spring (H’=1.27) and summer (H’=1.22), and lowering in winter (H’=1.10). Seasonal differences were statistically significant both between autumn and winter (t=2.85; df=196; p=0.01), and between autumn and summer (t=1.98; df =294; p=0.05). Between autumn and spring, no significant differences were observed (t=1.66; df=380; p=0.09).
The Food Niche Breadth (FNB), calculated according to the frequency of occurrence of prey species in the collected scats, had a relatively small variation according to seasons (see Appendix B). Considering two types of ungulate prey (wild vs domestic), wolfs’ niche adjusted to a diet specialized in wild ungulates in all seasons. Yet, predation over domestic ungulates increase with a marginal widening in autumn and winter (B standardized = 0.31 and 0.21); while wolfs’ diet was more focused over wild ungulates in spring and summer (B standardized = 0.26 and 0.24). However, when considering four wild prey species (roe deer, wild boar, red deer ˗ Cervus elaphus ˗, and mountain goat), we observed a narrower FNB in winter. On the other hand, when considering four domestic species (cattle, sheep, domestic goat, and horse), the wolf showed the narrowest FNB in summer, compared to the other seasons in which it turned to a more generalist strategy. FNB calculated according to the amount of biomass (kg) also varied according to seasons. Considering two types of ungulate prey (wild/domestic), FNB showed a generalist diet in all seasons (B<0.6). However, the values of FNB obtained showed a specialist diet when was calculated considering the four wild species (B<0.6). The same occurred when considering the four domestic species.
3.3. Annual Trends
The consumption of wild ungulates by wolves was higher compared to domestic ungulates in all years, considering that the abundance of each species in the study area did not change substantially during the evaluated period. On the one hand, the consumption of wild ungulates increased over time until 2019 yet decreasing for the two following years. In contrast, domestic ungulate consumption decreased from 2019 to 2021, being higher in 2017 that corresponded with the first year of the study (Figure 3A). According to the percentage of occurrence, roe deer predominated in 2017, 2018 and 2019, while the wild boar did in 2020 and 2021. Regarding domestic ungulates, the consumption of goat and cattle was similar in 2017; later, cattle was the most preyed domestic species during the 2018-2020 period, while in 2021 the goat was more preyed (see Appendix B). The percentage of occurrence of the different prey species according to the years was statistically significant for roe deer (χ2=327.61; df=32; p<0.001; n=637; see Appendix B). On the other hand, regarding the ingested biomass, values provided by wild were higher in all years. Overall, biomass from wild ungulates was similar between years, but a maximum was detected in 2021. Conversely, although biomass provided by domestic ungulates was also similar between years, a noticeable decrease occurred in 2020 (Fig 3B). The wild prey species that contributed most to the wolf diet in terms of biomass were wild boar and roe deer while cattle was the most consumed domestic ungulate according to the percentage of biomass in all years except 2020 (see Appendix B).
The year also influenced the diversity in wolf´s diet. 2019 showed the greatest diversity of diet (H’=1.39), unlike 2021 when the lowest diversity was noted (H’=1.02). Overall, the diversity was similar in the rest of the years (2017: H’=1.17; 2018: H’=1.14; 2020: H’=1.15) (Figure 7). Significant differences were observed in the diversity of the wolf diet between 2017-2019 (t Hutcheson =2.37; df=186; p=0.02), 2018-2019 (t Hutcheson =2.90; df=319; p=0.01), 2019-2020 (t Hutcheson =2.07; df=46; p=0.04), and 2019-2021 (t Hutcheson =3.02; df=61; p=0.01).
Food Niche Breadth (FNB) of the wolf was calculated according to the frequency of occurrence of prey species in the collected scats varied over time (Appendix B). Considering two types of ungulate prey (wild and domestic) as food categories, a specialized diet in wild prey was observed every year (B standardized < 0.4). On the one hand, when considering the four wild prey species (roe deer, wild boar, red deer, and mountain goat) in the FNB estimation, we observed a broader FNB due to a less strict specialist diet of wolf in 2019 and 2020. Instead, the wolf’s diet became specialized again, feeding primarily on roe deer and wild boar in 2021, as was the case in 2017 and 2018 when the FNB was narrower. On the other hand, regarding FNB estimation considering the four domestic preys (cattle, sheep, goat, and horse), a generalist diet was observed in 2017-2018. In 2019 the wolf began to restrict the consumption of a greater variety of domestic livestock prey and, its diet began to be considered specialist (feeding almost exclusively goat and sheep) from 2020 to 2021.
FNB estimated according to ingested biomass (kg), obtained from the collected scats, corroborated the results obtained with the calculations based on frequency of prey occurrence (see Appendix B). A generalist diet was observed in all years except 2021 (year in which FNB was narrower) when two types of prey (wild and domestic) were considered for calculation of index. The wolf diet was specialist in all years when considering wild ungulates, being the lowest FNB in 2021. FNB estimations from domestic ungulates showed a very specialized diet in cattle in all years, since it was the species that contributed the greatest amount of biomass to the wolf’s diet.
3.4. Forest Regions Trends
The consumption of wild ungulates by wolves was higher compared to domestic ungulates in all forest regions (Figure 4A). Roe deer was the predominant prey in all these regions, except in Navafría where wild board was the predominant prey. Among domestic ungulates, cattle were the most consumed animals (see Appendix B). The percentage of occurrence of the different prey species according to the forest regions was statistically significant (χ2=134.66; df=56; p=0.025; n=613; see Appendix B). The ingested biomass corresponding to wild ungulates was higher in Navafría than in other forest regions, without consider El Espinar, PRCAM Norte and Riaza where the sample size was n <5. (Figure 4B). The greatest biomass contribution to wolf diet from wild prey species, came from wild boar in all forest regions except in Montejo and Buitrago where the most consumed prey was roe deer (see Appendix B).
Wolfs’ diet diversity also varied according to the forest regions. Lozoya accounted the greatest diversity of diet (H’=1.33), unlike PRCAM Norte and Riaza, which were the ones with the lowest diversity (H’= 0.69 and H’=0.00, respectively). All other forest regions showed similar diversity values (El Espinar: H’=1.04; Montejo: H’=1.17; Navafría: H’=1.21; PN Peñarala: H’=1.22; Buitrago: H’=1.27) (Figure 7). However, the only significant differences were observed between Lozoya and Navafría (t Hutcheson =2.24; df=226; p=0.03).
The Ivlev’s index for domestic ungulates according to forest regions showed that wolves positively selected sheep and goats (El Espinar: D=0.91; Lozoya: D=0.76; Buitrago: D=0.98) in most regions over cows (El Espinar: D= 0.00; Lozoya: D=0.04; Buitrago: D=0.23). In PN Peñalara wolves selected cattle (D=0.40) but no sheep and goats. In Montejo, the selectivity was similar (cattle: D=0.52; sheeps: and goats: D=0.61). However, in Navafría wolves positively selected cattle (D=0.94) and negatively selected sheep and goats (D=-0.12).
The local authorities gave us information about canid attacks on cattle over 2020 and 2021. The year that ranchers reported the most attacks was 2021. The forest regions where the most attacks were reported were Montejo, PN Peñalara and Buitrago (Table 2).
4. Discussion
- The diet of wolves in the Sierra de Guadarrama, Sierra del Rincón and surroundings primarily consists of wild ungulates, like other regions in Europe [41,77,84,85]. However, there were differences in wolf diet compared to areas south of the Duero River in the Iberian Peninsula [11]. Our study area is characterized by a multi-prey ecosystem and well-distributed wild ungulates, such as roe deer and wild boar. This could explain why these species were the main prey, while consumption of domestic livestock, particularly free-roaming cattle, was minimal. Additionally, the larger size of adult cattle makes them a challenging target for wolves, with attacks primarily targeting calves.
- Wild boar was the predominant prey, in terms of the most frequently encountered species in wolf scat samples, followed by roe deer, which is consistent with aligns with Mori et al. [86] in Italy. Wild boar populations in the Iberian Peninsula increased throughout the region since the 21st century [87,88,89], even at high elevations [90], while roe deer populations are strongly associated with forested areas [44,91]. This difference in the availability distribution explains why the most consumed prey was wild boar, although wolves heavily rely on roe deer as well. Ivlev’s index supports this conclusion, indicating positive selection for roe deer and wild boar compared to other prey species. When considering the percentage of ingested biomass, cattle were the most consumed species, but this finding should be interpreted with caution due to the difference in body size between cattle and other prey species (e.g., cattle outweigh wild boar by a factor of 10).
- Regarding seasonal patterns, the consumption of wild ungulates based on the percentage of appearance of prey was higher in spring and summer (reproductive and breeding season), decreasing its consumption in autumn and winter. In contrast, the consumption of domestic ungulates was higher in autumn and winter. Pups abound in spring and summer and are an easy prey for wolves, given their inexperience [23,92]. The scarcity of this prey species during the colder seasons can compromise cattle and they may be perceived more attractive to wolves. Specifically, roe deer was the most consumed prey in all seasons, except spring when wild boar became the predominant prey, likely due to the high reproductive rate and larger litters (ẋ=3.5 ind.; [43]) of wild boar compared to roe deer (ẋ=1.46 ind.; [93]). When considering ingested biomass, cattle consistently contribute the highest percentage in all seasons due to their larger body size. FNB findings suggest that wolves exhibit a specialized diet during spring and summer, while showing a more general feeding pattern in autumn and winter. During the reproductive seasons (spring and summer), wolves have a larger pool of prey to choose from, due to the increased population of ungulates resulting from birthing season. This allows them to selectively target specific prey and specialize their diet according to resources availability. Conversely, in winter, when food availability is limited, the wolf’s diet becomes more generalized, consuming both domestic species and wild ones. This suggests that the wolf in the study area is a facultative specialist species, adapting its feeding behavior depending on the seasons.
- In terms of annual patterns, the consumption of wild ungulates is higher than domestic ungulates. Wolf’s diet was mainly based on roe deer from 2017 to 2019, while wild boar prevailed in recent years, possibly due to a decrease in roe deer populations (F. Horcajada, unpublished data) since the establishment of wolves in the area. The presence of red deer in wolf scats in 2019, despite not being generally present in the study area (although it is present in the eastern and southern surroundings), could be attributed to the dispersal behavior of wolves [94,95] or bait placement by hunters [96] or intended for study by researchers [36]. Consumption on mountain goats was sporadic, except for a slight increase in 2019. Mountain goat frequent rough areas that are difficult to access and/or guarantee a successful attack from wolves [97]. Wolves prefer steep slopes and open habitats where wild ungulates are more easily detectable and accessible [98], which may explain their lower consumption of mountain goats. The consumption of domestic horse in 2019 was a sporadic event, likely consumed as carrion. Based on FNB, and considering the % occurrence of prey in scats, the wolf followed a specialist diet every year feeding on wild ungulates instead of domestic ones. However, based on the biomass ingested, the wolf diet could be considered generalist, except in 2021. This could be due to the different size of the preys, as previously discussed.
- Regarding the forest region pattern, wolves fed mainly on wild ungulates, especially roe deer and wild boar, in all forest regions. Among species of domestic ungulates, cattle were the most consumed prey. In most of the forest regions, the presence of cattle is greater than that of sheep and goats, except Navafría where goats and sheep are predominant (INE 2020). Although cattle are more numerous in most forest regions, the prey selection index was higher for sheep and goats. However, in the case of Navafría, where the presence of cattle is lower, wolves positively selected cattle and avoided sheep and goats.
- Finally, an important concern in the study area is the inconsistency between official data on canid attacks on livestock provided by the Comunidad de Madrid and the findings regarding the wolf’s diet in different forest regions. For instance, the consumption of wild ungulates in PN Peñalara was significantly higher (2020: 78.6%, n=6; 2021: 88.2%, n=45) compared to domestic ungulates (2020: 21.4%, n=3; 2021: 10.2%, n=6 in PN Peñalara; Table 2). In Montejo, where the highest number of attacks was recorded, it paradoxically had one of the lowest consumption rates of domestic ungulates from 2017 to 2019. Despite the lack of diet data for Montejo in 2020-2021, which coincides with the peak number of attacks, the pattern of low domestic ungulates consumption in previous years suggests that the attacks may be primarily caused by other canids such as dogs rather than wolves. The higher number of attacks registered in 2020 and 2021 compared to the detection of domestic ungulate remains in wolf scats similar prey diversity indices in other regions like Buitrago (H’=1.27) and PN Peñalara (H’=1.22), further supports this hypothesis. We were unable to draw conclusions about the El Espinar, PRCAM Norte and Riaza forest regions due to low sample size.
5. Conclusions
In conclusion, the diet of the Iberian wolf in the Sierra de Guadarrama National Park, Sierra del Rincón and neighboring areas is mainly made up of wild ungulate species and a minority of domestic ones. The wolves exhibited selective feeding behavior, preferring roe deer and wild boar while avoiding other ungulates, especially domestic ones. These findings contradict the high number of reported attacks on livestock by forest rangers (Table 10). Therefore, we recommend against making management decisions or implementing control measures for wolf populations based solely on the argument of livestock attacks, as it lacks scientific support. Instead, we suggest focusing on protecting livestock in the area, considering the coexistence of wild and domestic ungulates, particularly during periods when wild ungulate juveniles are less available, and livestock remains exposed in the field. Roe deer and wild boar were the most frequently preyed upon species in the study area, with their ranking alternating depending on the year. Among domestic ungulates, cattle were the most targeted prey, contributing the highest biomass percentage in the wolf’s diet. The diversity of the wolf’s diet varied seasonally and annually, with a decreasing trend in the consumption of livestock over time. Maintaining a diverse and abundant wild prey population, especially during conflicting seasons when domestic animals are present in the field, can help reduce or prevent attacks on livestock, as supported by other studies [7,22]. Therefore, we recommend actions that benefit roe deer and wild boar populations, particularly related to forest and hunting management. This study highlights the effectiveness of non-invasive methods for monitoring the wolf’s trophic ecology and obtaining valuable information for the species management and conservation. We emphasize the importance of conducting long-term monitoring to collect extensive data, which can provide reliable and precise conclusions. This approach enables the exploration of alternative solutions for conservation conflicts and promotes better coexistence between large predators and humans.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
I.B. and F.H.: conceptualization. They have conceived and designed the study. All authors: faecal samples collection in field, but mainly F.H. and SRS. A.N.C. and I.B.: diet analysis. I.B. and L.O.J: data analysis. I.B. and L.O.J.; writing original draft. J.V. and I.B.: funding acquisition.
Funding
Financial support was provided by Consejería de Medio Ambiente, Administración Local y Ordenación del Territorio de la Comunidad de Madrid and Sierra de Guadarrama National Park (projects: 10/329633.9/18, 10/122964.9/19, 10/180884.9/19; contracts: 2020/0112, 2020/0169, 2020/0379, 2020/0528, 2021/0138).
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
We thank Consejería de Medio Ambiente, Administración Local y Ordenación del Territorio de la Comunidad de Madrid, Sierra de Guadarrama National Park and Sierra del Rincón Biosphere Reserva (Madrid, Spain), specially to Luis del Olmo and Pablo Sanjuanbenito, for the permits required to conduct this study. We are very grateful to Centro de Investigación, Seguimiento y Evaluación of the Sierra de Guadarrama National Park for their help during the fieldwork. Thanks are due to gamekeepers of the “Sierra del Rincón” Biosphere Reserva for their collaboration in the wolf scats collection in this zone. Ilad G. Vivas for his comments to improve the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A
Equation of Floyd et al. (1978), revised and adjusted by Weaver (1993)
This equation describes the relationship between the body mass of the prey (kg) and the mass of prey consumed (kg) per scat collected, where is the mass of prey consumed per scat collected and is the body mass average of an individual of a given prey species. The use of average mass is because the age of prey consumed leads to an overestimation of the contribution of smaller prey species to the diet due to their greater amount of hair and other indigestible matter per unit of body mass, which it produces more scats per unit mass of prey consumed (Weaver 1993). The estimate of the total biomass of each of the prey species in the scats was obtained by multiplying the calculated value of y by the number of collected scats containing each prey species. The biomass percentage of each prey item consumed was calculated by dividing the total weight of a particular prey item consumed by the weight of all mammalian prey items consumed.
Shannon Diversity Index Calculation
Shannon diversity index was calculated to estimate dietary diversity according to seasonality and years.
Shannon diversity index: Significant differences between pairs of Shannon indices were tested using Hutcheson’s t-statistic. Hutcheson’s t-test is a modified version of the classical t-test that provides a way to compare two samples using the variance of the Shannon index (Hutcheson 1970).
Hutcheson’s t-test: In the formula H represents the Shannon diversity index (Shannon 1948) for each of the two samples (subscript a and b). The lower part of the formula refers to the variance of each of the samples. The calculation of the variance of the Shannon diversity is done using the formula shown below:
In the formula S is the total number of species, while N is the total abundance. The p is the proportion that each species makes with respect to the total.
Niche Breadth (Levin’s Index) Calculation
Niche breadth and niche overlap of the wolf’s diet were calculated according to the frequency of occurrence of prey consumed and the biomass ingested over the seasons and years. Levin’s food niche breadth (FNB) Index (Levins 1968) was used to quantitatively measure specialization in the composition of the wolf’s diet.
Levins’s index: In the formula represents food niche breadth of wolf and the proportion of contribution of each group of wolf prey in the total biomass of food consumed by the canid (Nowak et al. 2011). The Levins’s index was standardized with the following equation to express the level of specialization on a scale from 0 to 1.
Levin’s index standardized: Where represents a high specialization and represents the equitable consumption of all prey. In the formula represents the Levin’s index standardized, represents the Levin’s index and represents the number of prey species consumed by wolf (Müller et al. 2006). If the values obtained are <0.6 means that the diet analysed presents a low diversity of prey considering the species under study a specialist predator. On the other hand, if the values are >0.6 they indicate that the species shows generalist eating habits, that is, it is a generalist predator (Krebs 1999; Cruz-Escalona et al. 2000).
We calculated the FNB in each of the four seasons and in each of the five years using both the data in the form of % occurrence and % biomass ingested. This index was calculated in three ways:
1. Wild/Dom: two categories of pi, one corresponding to the total proportion of wild ungulates and the other corresponding to the total proportion of domestic ungulates that appear in wolf feces at each season.
2. Wild/Wild: four categories of pi, corresponding to the proportion of each wild ungulate species (roe deer, red deer, wild boar and mountain goat) appearing in wolf feces at each season.
3. Dom/Dom: four categories of pi, corresponding to the proportion of each species of domestic ungulates (cattle, sheep, goat and horse) that appear in wolf feces in each season.
Ivelev’s Electivity Index Calculation
Ivlev’s electivity index modified by Jacobs (1974) was calculated to assess whether wolves selected preys positively or negatively. This index was applied, on the one hand, to evaluate the selection of prey throughout the study area and, on the other hand, to evaluate the selection among domestic ungulates by forest regions.
Ivlev’s index: In the formula is the proportion of a given prey species and is the proportion of available prey i in the study area. This index generates values ranging from -1 to 1. Negative values indicate prey inaccessibility or total avoidance of a species, value 0 indicates random prey consumption or not selection, and positive values indicate selection of a specific prey item.
Appendix B
Seasonal variation of wolf diet (n=637; pooled years)

| Prey occurrence | Ingested biomass | |||||
| Prey | Season | n | % | Kg | % | |
| Wild ungulates | Roe deer | Autumn | 83 | 43.9 | 141.4 | 13.2 |
| Winter | 49 | 44.1 | 83.5 | 20.1 | ||
| Spring | 76 | 38.0 | 129.4 | 10.7 | ||
| Summer | 73 | 53.3 | 124.3 | 15.5 | ||
| Red deer | Autumn | 1 | 0.5 | 5.0 | 0.5 | |
| Winter | 0 | 0.0 | 5.0 | 1.2 | ||
| Spring | 1 | 0.5 | 5.1 | 0.4 | ||
| Summer | 0 | 0.0 | 0.0 | 0.0 | ||
| Wild boar | Autumn | 54 | 28.5 | 233.2 | 21.8 | |
| Winter | 38 | 34.2 | 164.1 | 39.5 | ||
| Spring | 86 | 43.0 | 371.4 | 30.8 | ||
| Summer | 38 | 27.7 | 164.1 | 20.5 | ||
| Mountain goat | Autumn | 7 | 3.7 | 23.2 | 2.2 | |
| Winter | 1 | 0.9 | 3.3 | 0.8 | ||
| Spring | 4 | 2.0 | 13.3 | 1.1 | ||
| Summer | 7 | 5.1 | 23.2 | 2.9 | ||
| Total | Autumn | 145 | 76.7 | 402.8 | 37.7 | |
| Winter | 88 | 79.3 | 255.9 | 61.6 | ||
| Spring | 167 | 83.5 | 519.2 | 43.1 | ||
| Summer | 118 | 86.1 | 311.6 | 38.9 | ||
| Domestic ungulates | Cattle | Autumn | 18 | 9.5 | 630.7 | 59.1 |
| Winter | 4 | 3.6 | 140.2 | 33.7 | ||
| Spring | 19 | 9.5 | 665.7 | 55.2 | ||
| Summer | 13 | 9.5 | 455.5 | 56.9 | ||
| Domestic goat | Autumn | 12 | 6.3 | 20.34 | 1.9 | |
| Winter | 3 | 2.7 | 5.1 | 1.2 | ||
| Spring | 8 | 4.0 | 13.6 | 1.1 | ||
| Summer | 5 | 3.6 | 8.5 | 1.1 | ||
| Sheep | Autumn | 8 | 4.2 | 14.3 | 1.3 | |
| Winter | 3 | 2.7 | 14.2 | 3.5 | ||
| Spring | 4 | 2.0 | 7.1 | 0.6 | ||
| Summer | 0 | 0.0 | 0.0 | 0.0 | ||
| Horse | Autumn | 0 | 0.0 | 0.0 | 0.0 | |
| Winter | 0 | 0.0 | 0.0 | 0.0 | ||
| Spring | 0 | 0.0 | 0.0 | 0.0 | ||
| Summer | 1 | 0.7 | 24.8 | 3.1 | ||
| Unidentified Livestock |
Autumn | 6 | 3.2 | 0.0 | 0.0 | |
| Winter | 13 | 11.7 | 0.0 | 0.0 | ||
| Spring | 2 | 1.0 | 0.0 | 0.0 | ||
| Summer | 0 | 0.0 | 0.0 | 0.0 | ||
| Total | Autumn | 44 | 23.3 | 665.3 | 62.3 | |
| Winter | 23 | 20.7 | 159.5 | 38.4 | ||
| Spring | 33 | 16.5 | 686.4 | 56.9 | ||
| Summer | 19 | 13.9 | 488.8 | 61.1 | ||
| Season | Faecal samples collected (n) | % Wild ungulates occurrence | % Domestic ungulates occurrence |
Statistical results in each season (wild vs domestic) |
| Autumn | 189 | 76.7 | 23.3 | χ2= 53.97; df=1; p=0.001 |
| Winter | 111 | 79.3 | 20.7 | χ2= 8.06; df=1; p=0.001 |
| Spring | 200 | 83.5 | 16.5 | χ2= 89.78; df=1; p=0.001 |
| Summer | 137 | 86.1 | 13.9 | χ2= 71.54; df=1; p=0.001 |
|
Statistical results in all seasons (Wild species vs domestic species) |
χ2=16.81; df=9; p=0.037, n=518 | χ2= 43.59; df=12; p=0.001, n=119 | ||
Percentage of occurrence of wild and domestic ungulates according to seasons. Statistical results according to season comparing the consumption of domestic or wild ungulates, respectively.
Food niche breadth (A) wolf’s diet according to seasons. // Number of preys = number of prey species found in collected wolf scats. // Wild/Dom calculates the indicators based on two types of prey species (wild species and domestic species); Wild/Wild calculates the indicators based on four species of wild prey (roe deer, wild boar, red deer and mountain goat); Dom/Dom calculates the indicators based on four domestic prey species (cattle, sheep, domestic goat, and horse).
| Occurrence (n) | Biomass (Kg) | |||||
| B’ | B’ | |||||
| Wild/Dom | Wild/Wild | Dom/Dom | Wild/Dom | Wild/Wild | Dom/Dom | |
| Autumn | 0.31 | 0.37 | 0.57 | 0.90 | 0.40 | 0.00 |
| Winter | 0.21 | 0.33 | 0.64 | 0.90 | 0.30 | 0.10 |
| Spring | 0.26 | 0.37 | 0.44 | 1.00 | 0.20 | 0.00 |
| Summer | 0.24 | 0.35 | 0.28 | 0.90 | 0.40 | 0.00 |
| Number of preys | 2 | 4 | 4 | 2 | 4 | 4 |
Yearly changes of Iberian wolf diet (n=637; pooled seasons)
| Prey occurrence | Ingested biomass | |||||
| Prey | Years | n | % | Kg | % | |
| Wild ungulates | Roe deer | 2017 | 53 | 48.7 | 90.3 | 20.0 |
| 2018 | 87 | 51.5 | 148.2 | 20.4 | ||
| 2019 | 119 | 41.5 | 202.7 | 10.8 | ||
| 2020 | 9 | 30.0 | 15.3 | 5.3 | ||
| 2021 | 13 | 31.0 | 22.1 | 16.5 | ||
| Red deer | 2017 | 0 | 0.0 | 0.0 | 0.0 | |
| 2018 | 0 | 0.0 | 0.0 | 0.0 | ||
| 2019 | 2 | 0.7 | 10.1 | 0.6 | ||
| 2020 | 0 | 0.0 | 0.0 | 0.0 | ||
| 2021 | 0 | 0.0 | 0.0 | 0.0 | ||
| Wild boar | 2017 | 31 | 28.4 | 133.9 | 29.6 | |
| 2018 | 47 | 27.8 | 203.0 | 27.9 | ||
| 2019 | 100 | 34.8 | 431.9 | 23.0 | ||
| 2020 | 14 | 46.7 | 60.5 | 20.9 | ||
| 2021 | 24 | 57.1 | 103.7 | 77.1 | ||
| Mountain goat | 2017 | 0 | 0.0 | 0.0 | 0.0 | |
| 2018 | 1 | 0.6 | 3.3 | 0.5 | ||
| 2019 | 17 | 5.9 | 56.4 | 3.0 | ||
| 2020 | 1 | 3.3 | 3.3 | 1.1 | ||
| 2021 | 0 | 0.0 | 0.0 | 0.0 | ||
| Total | 2017 | 84 | 77.1 | 224.2 | 49.6 | |
| 2018 | 135 | 79.9 | 354.5 | 48.8 | ||
| 2019 | 238 | 82.9 | 701.1 | 37.4 | ||
| 2020 | 24 | 80.0 | 79.1 | 27.3 | ||
| 2021 | 37 | 88.1 | 125.8 | 93.6 | ||
| Domestic ungulates | Cattle | 2017 | 6 | 5.5 | 210.2 | 46.5 |
| 2018 | 10 | 5.9 | 350.4 | 48.3 | ||
| 2019 | 32 | 11.2 | 1121.3 | 59.8 | ||
| 2020 | 6 | 20.0 | 210.2 | 72.7 | ||
| 2021 | 0 | 0.0 | 0.0 | 0.0 | ||
| Domestic goat | 2017 | 6 | 5.5 | 10.2 | 2.3 | |
| 2018 | 8 | 4.7 | 13.5 | 1.9 | ||
| 2019 | 11 | 3.8 | 18.7 | 0.9 | ||
| 2020 | 0 | 0.0 | 0.0 | 0.0 | ||
| 2021 | 3 | 7.1 | 5.1 | 3.8 | ||
| Sheep | 2017 | 4 | 3.7 | 7.1 | 1.6 | |
| 2018 | 4 | 2.4 | 7.1 | 0.9 | ||
| 2019 | 5 | 1.7 | 8.9 | 0.5 | ||
| 2020 | 0 | 0.0 | 0.0 | 0.0 | ||
| 2021 | 2 | 4.8 | 3.6 | 2.7 | ||
| Horse | 2017 | 0 | 0.0 | 0.0 | 0.0 | |
| 2018 | 0 | 0.0 | 0.0 | 0.0 | ||
| 2019 | 1 | 0.3 | 24.8 | 1.6 | ||
| 2020 | 0 | 0.0 | 0.0 | 0.0 | ||
| 2021 | 0 | 0.0 | 0.0 | 0.0 | ||
| Unidentified livestock | 2017 | 9 | 8.3 | 0.0 | 0.0 | |
| 2018 | 12 | 7.1 | 0.0 | 0.0 | ||
| 2019 | 0 | 0.0 | 0.0 | 0.0 | ||
| 2020 | 0 | 0.0 | 0.0 | 0.0 | ||
| 2021 | 0 | 0.0 | 0.0 | 0.0 | ||
| Total | 2017 | 25 | 22.9 | 227.5 | 50.4 | |
| 2018 | 34 | 20.1 | 371.0 | 51.1 | ||
| 2019 | 49 | 17.1 | 1173.7 | 62.6 | ||
| 2020 | 6 | 20.0 | 210.2 | 72.7 | ||
| 2021 | 5 | 11.9 | 8.7 | 6.4 | ||
| Year | Faecal samples collected (n) | % Wild ungulates occurrence | % Domestic ungulates occurrence |
Statistical results in each year (wild vs domestic) |
| 2017 | 109 | 77.1 | 22.9 | χ2=31.94; df=1; p=0.001 |
| 2018 | 169 | 79.9 | 20.1 | χ2=60.36; df=1; p=0.001 |
| 2019 | 287 | 82.9 | 17.1 | χ2=124.46; df=1; p=0.001 |
| 2020 | 30 | 80.0 | 20.0 | χ2=10.80; df=1; p=0.001 |
| 2021 | 42 | 88.1 | 11.9 | χ2=24.38; df=1; p=0.001 |
|
Statistical results in all years (wild species vs domestic species) |
χ2=16.81; df=9; p=0.037, n=518 | χ2= 43.59; df=12; p=0.001, n=119 | ||
Percentage of occurrence of wild and domestic ungulates according to years. Statistical results according to year comparing the consumption of domestic and wild ungulates separately.
Food niche breadth (A) of wolf diet according to years. // Number of preys = number of prey species found in collected wolf scats. // Wild/Dom calculates the indicators based on two types of prey species (wild species and domestic species); Wild/Wild calculates the indicators based on four species of wild prey (roe deer, wild boar, red deer and mountain goat); Dom/Dom calculates the indicators based on four domestic prey species (cattle, sheep, domestic goat, and horse).
| Ocurrence (n) | Biomass (Kg) | |||||
| B’ | B’ | |||||
| Wild/Dom | Wild/Wild | Dom/Dom | Wild/Dom | Wild/Wild | Dom/Dom | |
| 2017 | 0.22 | 0.29 | 0.63 | 1.00 | 0.30 | 0.10 |
| 2018 | 0.21 | 0.28 | 0.56 | 1.00 | 0.30 | 0.00 |
| 2019 | 0.32 | 0.44 | 0.35 | 0.90 | 0.40 | 0.00 |
| 2020 | 0.33 | 0.46 | 0.00 | 0.70 | 0.20 | 0.00 |
| 2021 | 0.18 | 0.27 | 0.30 | 0.10 | 0.10 | 0.30 |
| Number of preys | 2 | 4 | 4 | 2 | 4 | 4 |
Comparative of composition of Iberian wolf diet between forest regions based on 637 scats. The ingested prey biomass (in kg) was calculated using body masses.
| Prey occurrence | Ingested biomass | |||||
| Prey | Forest region | n | % | Kg | % | |
| Wild ungulates | Roe deer | El Espinar | 1 | 25.0 | 1.7 | 18.1 |
| PRCAM Norte | 1 | 50.0 | 1.7 | 28.3 | ||
| PN Peñalara | 114 | 43.5 | 194.1 | 13.2 | ||
| Lozoya | 62 | 48.9 | 105.6 | 15.1 | ||
| Montejo | 30 | 60.0 | 51.1 | 21.4 | ||
| Buitrago | 16 | 53.3 | 27.2 | 16.3 | ||
| Navafría | 46 | 34.1 | 78.3 | 11.0 | ||
| Riaza | 3 | 100.0 | 5.1 | 100 | ||
| Red deer | El Espinar | 0 | 0.0 | 0.0 | 0.0 | |
| PRCAM Norte | 0 | 0.0 | 0.0 | 0.0 | ||
| PN Peñalara | 1 | 0.4 | 5.0 | 0.4 | ||
| Lozoya | 0 | 0.0 | 0.0 | 0.0 | ||
| Montejo | 0 | 0.0 | 0.0 | 0.0 | ||
| Buitrago | 0 | 0.0 | 0.0 | 0.0 | ||
| Navafría | 1 | 0.7 | 5.0 | 0.7 | ||
| Riaza | 0 | 0.0 | 0.0 | 0.0 | ||
| Wild boar | El Espinar | 1 | 25.0 | 4.3 | 45.9 | |
| PRCAM Norte | 1 | 50.0 | 4.3 | 71.7 | ||
| PN Peñalara | 106 | 40.5 | 457.8 | 31.2 | ||
| Lozoya | 28 | 22.0 | 120.9 | 17.3 | ||
| Montejo | 4 | 8.0 | 17.3 | 7.3 | ||
| Buitrago | 6 | 20.0 | 25.9 | 15.5 | ||
| Navafría | 67 | 49.6 | 289.4 | 40.7 | ||
| Riaza | 0 | 0.0 | 0.0 | 0.0 | ||
| Mountain goat | El Espinar | 0 | 0.0 | 0.0 | 0.0 | |
| PRCAM Norte | 0 | 0.0 | 0.0 | 0.0 | ||
| PN Peñalara | 4 | 1.5 | 13.3 | 0.9 | ||
| Lozoya | 3 | 2.4 | 10.0 | 1.4 | ||
| Montejo | 8 | 16.0 | 26.6 | 11.1 | ||
| Buitrago | 0 | 0.0 | 0.0 | 0.0 | ||
| Navafría | 3 | 2.2 | 9.9 | 1.4 | ||
| Riaza | 0 | 0.0 | 0.0 | 0.0 | ||
| Total | El Espinar | 2 | 50.0 | 6.0 | 64.0 | |
| PRCAM Norte | 2 | 100.0 | 6.0 | 100.0 | ||
| PN Peñalara | 225 | 85.9 | 670.2 | 45.7 | ||
| Lozoya | 93 | 73.2 | 236.5 | 33.8 | ||
| Montejo | 42 | 84.0 | 95.0 | 39.8 | ||
| Buitrago | 22 | 73.4 | 53.1 | 31.8 | ||
| Navafría | 117 | 86.6 | 382.6 | 53.8 | ||
| Riaza | 3 | 100.0 | 5.1 | 100.0 | ||
| Domestic ungulates | Cattle | El Espinar | 0 | 0.0 | 0.0 | 0.0 |
| PRCAM Norte | 0 | 0.0 | 0.0 | 0.0 | ||
| PN Peñalara | 22 | 8.4 | 770.9 | 52.5 | ||
| Lozoya | 12 | 9.4 | 420.5 | 60.0 | ||
| Montejo | 4 | 8.0 | 140.2 | 58.8 | ||
| Buitrago | 3 | 10.0 | 105.1 | 63.0 | ||
| Navafría | 9 | 6.7 | 315.3 | 44.3 | ||
| Riaza | 0 | 0.0 | 0.0 | 0.0 | ||
| Domestic goat | El Espinar | 2 | 50.0 | 3.4 | 36.0 | |
| PRCAM Norte | 0 | 0.0 | 0.0 | 0.0 | ||
| PN Peñalara | 9 | 3.4 | 15.3 | 1.0 | ||
| Lozoya | 6 | 4.7 | 10.2 | 1.4 | ||
| Montejo | 1 | 2.0 | 1.7 | 0.7 | ||
| Buitrago | 4 | 13.3 | 6.8 | 4.1 | ||
| Navafría | 7 | 5.3 | 11.9 | 1.7 | ||
| Riaza | 0 | 0.0 | 0.0 | 0.0 | ||
| Sheep | El Espinar | 0 | 0.0 | 0.0 | 0.0 | |
| PRCAM Norte | 0 | 0.0 | 0.0 | 0.0 | ||
| PN Peñalara | 6 | 2.3 | 10.7 | 0.7 | ||
| Lozoya | 5 | 3.9 | 8.9 | 1.3 | ||
| Montejo | 1 | 2.0 | 1.8 | 0.7 | ||
| Buitrago | 1 | 3.3 | 1.9 | 1.1 | ||
| Navafría | 1 | 0.7 | 1.8 | 0.2 | ||
| Riaza | 0 | 0.0 | 0.0 | 0.0 | ||
| Horse | El Espinar | 0 | 0.0 | 0.0 | 0.0 | |
| PRCAM Norte | 0 | 0.0 | 0.0 | 0.0 | ||
| PN Peñalara | 0 | 0.0 | 0.0 | 0.0 | ||
| Lozoya | 1 | 0.9 | 24.8 | 3.5 | ||
| Montejo | 0 | 0.0 | 0.0 | 0.0 | ||
| Buitrago | 0 | 0.0 | 0.0 | 0.0 | ||
| Navafría | 0 | 0.0 | 0.0 | 0.0 | ||
| Riaza | 0 | 0.0 | 0.0 | 0.0 | ||
| Unidentified livestock | El Espinar | 0 | 0.0 | - | - | |
| PRCAM Norte | 0 | 0.0 | - | - | ||
| PN Peñalara | 0 | 0.0 | - | - | ||
| Lozoya | 10 | 7.9 | - | - | ||
| Montejo | 2 | 4.0 | - | - | ||
| Buitrago | 0 | 0.0 | - | - | ||
| Navafría | 1 | 0.7 | - | - | ||
| Riaza | 0 | 0.0 | - | - | ||
| Total | El Espinar | 2 | 50.0 | 3.4 | 36.0 | |
| PRCAM Norte | 0 | 0.0 | 0 | 0.0 | ||
| PN Peñalara | 37 | 14.1 | 796.9 | 54.3 | ||
| Lozoya | 34 | 26.8 | 464.4 | 66.2 | ||
| Montejo | 8 | 16.0 | 143.7 | 60.2 | ||
| Buitrago | 8 | 26.6 | 113.8 | 68.2 | ||
| Navafría | 18 | 13.4 | 329.0 | 46.2 | ||
| Riaza | 0 | 0.0 | 0.0 | 0.0 | ||
Percentage of occurrence of wild and domestic ungulates according to forest regions. Statistical results according to forest region comparing the consumption of domestic and wild ungulates.
| Forest region | Faecal samples collected (n) | % Wild ungulates occurrence | % Domestic ungulates occurrence |
Statistical results (wild vs domestic) |
| El Espinar | 4 | 50.0 | 50.0 | - |
| PRCAM Norte | 2 | 100.0 | 0.0 | - |
| PN Peñalara | 262 | 85.9 | 14.1 | χ2=134.90; df=1; p=0.001 |
| Lozoya | 127 | 73.2 | 26.8 | χ2=27.40; df=1; p=0.001 |
| Montejo | 50 | 84.0 | 16.0 | χ2=23.12; df=1; p=0.001 |
| Buitrago | 30 | 73.4 | 26.6 | χ2=6.53; df=1; p=0.11 |
| Navafría | 135 | 86.6 | 13.4 | χ2=72.6; df=1; p=0.001 |
| Riaza | 3 | 100.0 | 0.0 | - |
| Statistical results WYXYWX(wild species vs domestic species) | χ2=69.68; df=21; p=0.038, n=506 | χ2=30.94; df=20; p=0.065, n=107 | ||
References
- Keller, I.; Largiadèr, C.R. Recent Habitat Fragmentation Caused by Major Roads Leads to Reduction of Gene Flow and Loss of Genetic Variability in Ground Beetles. Proc. R. Soc. Lond. B Biol. Sci. 2003, 270, 417–423. [Google Scholar] [CrossRef]
- Geneletti, D. Using Spatial Indicators and Value Functions to Assess Ecosystem Fragmentation Caused by Linear Infrastructures. Int. J. Appl. Earth Obs. Geoinformation 2004, 5, 1–15. [Google Scholar] [CrossRef]
- Mancebo Quintana, S.; Martín Ramos, B.; Casermeiro Martínez, M.A.; Otero Pastor, I. A Model for Assessing Habitat Fragmentation Caused by New Infrastructures in Extensive Territories—Evaluation of the Impact of the Spanish Strategic Infrastructure and Transport Plan. J. Environ. Manage. 2010, 91, 1087–1096. [Google Scholar] [CrossRef] [PubMed]
- Mullu, D. A Review on the Effect of Habitat Fragmentation on Ecosystem. J. Nat. Sci. Res. 2016, 6, 1. [Google Scholar]
- Laliberte, A.; Ripple, W. Range Contractions of North American Carnivores and Ungulates. Bioscience 2004, 54. [Google Scholar] [CrossRef]
- Aybes, C.; Yalden, D. Mammal Review. 1995, pp. 201–227.
- Zlatanova, D.; Ahmed, A.; Vlasseva, A.; Genov, P. Adaptive Diet Strategy of the Wolf (Canis lupus L.) in Europe: A Review. Acta Zool. Bulg. 2014, 66, 439–452. [Google Scholar]
- Ripple, W.; Estes, J.; Beschta, R.; Wilmers, C.; Ritchie, E.; Hebblewhite, M.; Berger, J.; Elmhagen, B.; Letnic, M.; Nelson, M.; et al. Status and Ecological Effects of the World’s Largest Carnivores. Science 2014, 343, 1241484. [Google Scholar] [CrossRef]
- Louvrier, J.; Duchamp, C.; Lauret, V.; Marboutin, E.; Cubaynes, S.; Choquet, R.; Miquel, C.; Gimenez, O. Mapping and Explaining Wolf Recolonization in France Using Dynamic Occupancy Models and Opportunistic Data. Ecography 2018, 41, 647–660. [Google Scholar] [CrossRef]
- Gazzola, A.; Capitani, C.; Mattioli, L.; Apollonio, M. Livestock Damage and Wolf Presence. J. Zool. 2008, 274, 261–269. [Google Scholar] [CrossRef]
- Tinoco Torres, R.; Silva, N.; Brotas, G.; Fonseca, C. To Eat or Not To Eat? The Diet of the Endangered Iberian Wolf (Canis lupus Signatus) in a Human-Dominated Landscape in Central Portugal. PLoS ONE 2015, 10. [Google Scholar] [CrossRef]
- Recio, M.R.; Sand, H.; Virgós, E. Promoting Grazing or Rewilding Initiatives against Rural Exodus? The Return of the Wolf and Other Large Carnivores Must Be Considered. Environ. Conserv. 2020, 47, 269–276. [Google Scholar] [CrossRef]
- Boitani, L.; Linnell, J. Bringing Large Mammals Back: Large Carnivores in Europe. Rewilding Eur. Landsc. 2015, 67–84. [Google Scholar] [CrossRef]
- Nowak, S.; Mysłajek, R.; Jedrzejewska, B. Patterns of Wolf Canis lupus Predation on Wild and Domestic Ungulates in the Western Carpathian Mountains (S Poland). Acta Theriol. (Warsz.) 2005, 50, 263–276. [Google Scholar] [CrossRef]
- Meriggi, A.; Lovari, S. A Review of Wolf Predation in Southern Europe: Does the Wolf Prefer Wild Prey to Livestock? J. Appl. Ecol. 1996, 33, 1561–1571. [Google Scholar] [CrossRef]
- Müller, S. Diet Composition of Wolves (Canis lupus) on the Scandinavian Peninsula Determined by Scat Analysis., School of Forest Science and Resource Management, Technical University of München, Germany., 2006.
- Ansorge, H.; Kluth, G.; Hahne, S. Feeding Ecology of WolvesCanis lupus Returning to Germany. Acta Theriol. (Warsz.) 2006, 51, 99–106. [Google Scholar] [CrossRef]
- Stahler, D.R.; Smith, D.W.; Guernsey, D.S. Foraging and Feeding Ecology of the Gray Wolf (Canis lupus): Lessons from Yellowstone National Park, Wyoming, USA. J. Nutr. 2006, 136, 1923S–1926S. [Google Scholar] [CrossRef]
- Daria Octenjak; Lana Pađen; Valentina Šilić; Slaven Reljić; Tajana Trbojević Vukičević; Josip Kusak Wolf Diet and Prey Selection in Croatia. Mammal Res. 2020, 65, 647–654. [CrossRef]
- Wolves: Behavior, Ecology, and Conservation; Mech, L. D., Boitani, L., Eds.; University of Chicago Press: Chicago, IL, 2007; ISBN 978-0-226-51697-4. [Google Scholar]
- Lanszki, J.; Márkus, M.; Újváry, D.; Szabó, Á.; Szemethy, L. Diet of Wolves Canis lupus Returning to Hungary. Acta Theriol. (Warsz.) 2012, 57, 189–193. [Google Scholar] [CrossRef]
- Newsome, T.; Boitani, L.; Chapron, G.; Ciucci, P.; Dickman, C.; Dellinger, J.; López-Bao, J.V.; Peterson, R.; Shores, C.; Wirsing, A.; et al. Food Habits of the World’s Grey Wolves. Mammal Rev. 2016, 46, n. [Google Scholar] [CrossRef]
- Barja, I. Prey and Prey-Age Preference by the Iberian Wolf Canis lupus Signatus in a Multiple-Prey Ecosystem. Wildl. Biol. 2009, 15, 147–154. [Google Scholar] [CrossRef]
- Stahlberg, S.; Bassi, E.; Viviani, V.; Apollonio, M. Quantifying Prey Selection of Northern and Southern European Wolves (Canis lupus). Mamm. Biol.—Z. Für Säugetierkd. 2016, 83. [Google Scholar] [CrossRef]
- Sidorovich, V.E.; Tikhomirova, L.L.; Jędrzejewska, B. Wolf Canis lupus Numbers, Diet and Damage to Livestock in Relation to Hunting and Ungulate Abundance in Northeastern Belarus during 1990–2000. Wildl. Biol. 2003, 9, 103–111. [Google Scholar] [CrossRef]
- Wilmers, C.C.; Post, E.; Hastings, A. The Anatomy of Predator-Prey Dynamics in a Changing Climate. J. Anim. Ecol. 2007, 76, 1037–1044. [Google Scholar] [CrossRef]
- Jędrzejewski, W.; Niedziałkowska, M.; Hayward, M.W.; Goszczyński, J.; Jędrzejewska, B.; Borowik, T.; Bartoń, K.A.; Nowak, S.; Harmuszkiewicz, J.; Juszczyk, A.; et al. Prey Choice and Diet of Wolves Related to Ungulate Communities and Wolf Subpopulations in Poland. J. Mammal. 2012, 93, 1480–1492. [Google Scholar] [CrossRef]
- Mech, L.D.; Fieberg, J. Re-Evaluating the Northeastern Minnesota Moose Decline and the Role of Wolves. J. Wildl. Manag. 2014, 78, 11431150. [Google Scholar] [CrossRef]
- Bocci, A.; Lovari, S.; Khan, M.Z.; Mori, E. Sympatric Snow Leopards and Tibetan Wolves: Coexistence of Large Carnivores with Human-Driven Potential Competition. Eur. J. Wildl. Res. 2017, 63, 92. [Google Scholar] [CrossRef]
- Assessing the Relationship between Illegal Hunting of Ungulates, Wild Prey Occurrence and Livestock Depredation Rate by Large Carnivores—Soofi—2019—Journal of Applied Ecology—Wiley Online Library Available online:. Available online: https://besjournals.onlinelibrary.wiley.com/doi/full/10.1111/1365-2664.13266 (accessed on 26 September 2023).
- Blanco, J.C.; Cuesta, L.; Reig, S. El Lobo (Canis lupus) En Espana. Situacion, Problematica y Apuntes Sobre Su Ecologia.; Madrid, 1990; p. 118.
- Cuesta, L.; Barcena, F.; Palacios, F.; Reig, S. The Trophic Ecology of the Iberian Wolf (Canis lupus signatus Cabrera, 1907). A New Analysis of Stomach’s Data. Mammalia 1991, 55, 239–254. [Google Scholar] [CrossRef]
- Lagos, L.; Bárcena, F. Spatial Variability in Wolf Diet and Prey Selection in Galicia (NW Spain). Mammal Res. 2018. [Google Scholar] [CrossRef]
- Llaneza, L.; López-Bao, J.V. Indirect Effects of Changes in Environmental and Agricultural Policies on the Diet of Wolves. Eur. J. Wildl. Res. 2015, 61, 895–902. [Google Scholar] [CrossRef]
- Chapron, G.; Kaczensky, P.; Linnell, J.; von Arx, M.; Huber, D.; Andrén, H.; López-Bao, J.V.; Adamec, M.; Álvares, F.; Anders, O.; et al. Recovery of Large Carnivores in Europe’s Modern Human-Dominated Landscapes. Science 2014, 346, 1517–1519. [Google Scholar] [CrossRef]
- Iliopoulos, Y.; Sgardelis, S.; Koutis, V.; Savaris, D. Wolf Depredation on Livestock in Central Greece. Mammal Res. 2009, 54, 11–22. [Google Scholar] [CrossRef]
- Nores, C.; González, F.; García, P. Wild Boar Distribution Trends in the Last Two Centuries: An Example in Northern Spain. IBEX J. Mt. Ecol. 1995, 3, 137–140. [Google Scholar]
- Silva, P.; López-Bao, J.V.; Llaneza, L.; Álvares, F.; Lopes, S.; Blanco, J.C.; Cortés, Y.; García, E.; Palacios, V.; Rio-Maior, H.; et al. Cryptic Population Structure Reveals Low Dispersal in Iberian Wolves. Sci. Rep. 2018, 8, 14108. [Google Scholar] [CrossRef]
- Gazzola, A.; Bertelli, I.; Avanzinelli, E.; Tolosano, A.; Bertotto, P.; Apollonio, M. Predation by Wolves (Canis lupus) on Wild and Domestic Ungulates of the Western Alps, Italy. J. Zool. 2005, 266, 205–213. [Google Scholar] [CrossRef]
- Meriggi, A.; Brangi, A.; Schenone, L.; Signorelli, D.; Milanesi, P. Changes of Wolf (Canis lupus) Diet in Italy in Relation to the Increase of Wild Ungulate Abundance. Ethol. Ecol. Evol. 2011, 23, 195–210. [Google Scholar] [CrossRef]
- Milanesi, P.; Alberto, M.; Merli, E. Selection of Wild Ungulates by Wolves Canis lupus (L. 1758) in an Area of the Northern Apennines (North Italy). Ethol. Ecol. Evol.—ETHOL ECOL EVOL 2012, 24, 81–96. [Google Scholar] [CrossRef]
- Guimarães, N.F.; Álvares, F.; Ďurová, J.; Urban, P.; Bučko, J.; Iľko, T.; Brndiar, J.; Štofik, J.; Pataky, T.; Barančeková, M.; et al. What Drives Wolf Preference towards Wild Ungulates? Insights from a Multi-Prey System in the Slovak Carpathians. PloS One 2022, 17, e0265386. [Google Scholar] [CrossRef]
- FernáNdez-Llario, P.; Carranza, J.; Mateos-Quesada, P. Sex Allocation in a Polygynous Mammal with Large Litters: The Wild Boar. Anim. Behav. 1999, 58, 1079–1084. [Google Scholar] [CrossRef]
- Horcajada-Sánchez, F.; Barja, I. Evaluating the Effectiveness of Two Distance-Sampling Techniques for Monitoring Roe Deer (Capreolus Capreolus) Densities. Ann. Zool. Fenn. 2015, 52, 167–176. [Google Scholar] [CrossRef]
- Valdmann, H.; Andersone-Lilley, Z.; Koppa, O.; Ozolins, J.; Bagrade, G. Winter Diets of WolfCanis lupus and LynxLynx Lynx in Estonia and Latvia. Acta Theriol. (Warsz.) 2005, 50, 521–527. [Google Scholar] [CrossRef]
- Imbert, C.; Caniglia, R.; Fabbri, E.; Milanesi, P.; Randi, E.; Serafini, M.; Torretta, E.; Meriggi, A. Why Do Wolves Eat Livestock?: Factors Influencing Wolf Diet in Northern Italy. Biol. Conserv. 2016, 195, 156–168. [Google Scholar] [CrossRef]
- Ciucci, P.; Artoni, L.; Crispino, F.; Tosoni, E.; Boitani, L. Inter-Pack, Seasonal and Annual Variation in Prey Consumed by Wolves in Pollino National Park, Southern Italy. Eur. J. Wildl. Res. 2018, 64. [Google Scholar] [CrossRef]
- Smith, D.; Drummer, T.; Murphy, K.; Guernsey, D.; Evans, S. Winter Prey Selection and Estimation of Wolf Kill Rates in Yellowstone National Park, 1995-2000. J. Wildl. Manag. 2004, 68, 153–166. [Google Scholar] [CrossRef]
- Blanco, J.; Reig, S.; Cuesta, L. Distribution, Status and Conservation Problems of the Wolf Canis lupus in Spain. Biol. Conserv. 1992, 60, 73–80. [Google Scholar] [CrossRef]
- Blanco, J.C.; Cortés, Y. Ecología, Censos, Percepción y Evolución Del Lobo En España: Análisis de Un Conflicto. Sociedad Española Para La Conservación y Estudio de Los Mamíferos, Málaga, Spain. In Proceedings of the Sociedad Española para la Conservación y Estudio de los Mamíferos; Málaga; 2002. [Google Scholar]
- Blanco, J.; Cortés, Y. Dispersal Patterns, Social Structure and Mortality of Wolves Living in Agricultural Habitats in Spain. J. Zool. 2007, 273, 114–124. [Google Scholar] [CrossRef]
- Trouwborst, A. The EU Habitats Directive and Wolf Conservation and Management on the Iberian Peninsula: A Legal Perspective. Galemys Span. J. Mammal. 2014, 26, 15–30. [Google Scholar] [CrossRef]
- Delibes-Mateos, M. Wolf Media Coverage in the Region of Castilla y León (Spain): Variations over Time and in Two Contrasting Socio-Ecological Settings. Animals 2020, 10, 736. [Google Scholar] [CrossRef]
- Espirito-Santo, C. Human Dimensions in Iberian Wolf Management in Portugal: Attitudes and Beliefs of Interest Groups and the Public toward a Fragmented Wolf Population. masters, Memorial University of Newfoundland, 2007.
- Houston, M.; Bruskotter, J.; Fan, D. Attitudes Toward Wolves in the United States and Canada: A Content Analysis of the Print News Media, 1999–2008. Hum. Dimens. Wildl. 2010, 15, 389–403. [Google Scholar] [CrossRef]
- López-Bao, J.V.; Sazatornil, V.; Llaneza, L.; Rodriguez, A. Indirect Effects on Heathland Conservation and Wolf Persistence of Contradictory Policies That Threaten Traditional Free- Ranging Horse Husbandry. Conserv. Lett. 2013, 6, 448–455. [Google Scholar] [CrossRef]
- Redpath, S.M.; Gutiérrez, R.J.; Wood, K.A.; Sidaway, R.; Young, J.C. An Introduction to Conservation Conflicts. In Conflicts in Conservation: Navigating Towards Solutions; Cambridge University Press, 2015; pp. 3–18.
- Braña, F.; Campo, J.; Palomero, G. cta Biológica Montana. 1982, pp. 33–52.
- Salvador, A.; Abad, P.L. Food Habits of a Wolf Population (Canis lupus) in León Province, Spain. 1987, 51, 45–52. [CrossRef]
- Urios, V.; Vila, C.; Bernáldez, E.; Delibes, M. Contribución al Conocimiento de La Alimentación Del Lobo Ibérico Canis lupus En La Sierra de La Culebra (Zamora). In Proceedings of the El lobo ibérico; Adm. Prov. Salamanca; pp. 47–56.
- Fernández, A.; Fernández, JM; Palomero, G. El lobo en Cantabria. Situación, problemática y apuntes sobre su ecología. 33–44.
- Barja, I.; de Miguel, F.J.; Bárcena, F. The Importance of Crossroads in Faecal Marking Behaviour of the Wolves (Canis lupus). Naturwissenschaften 2004, 91, 489–492. [Google Scholar] [CrossRef]
- Barja, I.; de Miguel, F.; BÁRCENA, F. Faecal Marking Behaviour of Iberian Wolf in Different Zones of Their Territory. Folia Zool. -Praha- 2005, 54, 21–29. [Google Scholar]
- Steenweg, R.; Gillingham, M.; Parker, K.; Heard, D. Considering Sampling Approaches When Determining Carnivore Diets: The Importance of Where, How, and When Scats Are Collected. Mammal Res. 2015, 60. [Google Scholar] [CrossRef]
- Barja, I. Decision Making in Plant Selection during the Faecal-Marking Behaviour of Wild Wolves. Anim. Behav. 2009, 77, 489–493. [Google Scholar] [CrossRef]
- Ferretti, F.; Lovari, S.; Mancino, V.; Burrini, L.; Rossa, M. Food Habits of Wolves and Selection of Wild Ungulates in a Prey-Rich Mediterranean Coastal Area. Mamm. Biol. 2019, 99, 119–127. [Google Scholar] [CrossRef]
- Núñez, I.B.; González, M.C.H.; Castilla, Á.N. Manual de los patrones macroscópicos y cuticulares del pelo en mamíferos de la península ibérica; UAM Ediciones, 2021.
- Zub, K.; Theuerkauf, J.; Jędrzejewski, W.; Jędrzejewska, B.; Schmidt, K.; Kowalczyk, R. Wolf Pack Territory Marking in the Białowieża Primeval Forest (Poland). Behaviour 2003, 140, 635–648. [Google Scholar] [CrossRef]
- Bergamaschi, R. SIG Aplicado à Segurança No Trânsito–Estudo de Caso No Município de Vitória–ES. Monogr. Apresentada Ao Dep. Geogr. UFES 2010.
- Kawamoto, M. Análise de Técnicas de Distribuição Espacial Com Padrões Pontuais e Aplicação a Dados de Acidentes de Trânsito e a Dados Da Dengue de Rio Claro-SP, Universidade Estadual Paulista Julio de Mesquita Filho, Instituto de Biociências de Botucatu, 2012.
- Rizzatti, M.; Batista, N.; Cezar Spode, P.; Bouvier Erthal, D.; Faria, R.; Scotti, A.; Trentin, R.; Petsch, C.; Turba, I.; Quoos, J. Mapeamento Da COVID-19 Por Meio Da Densidade de Kernel. Metodol. E Aprendizado 2020, 3, 44–53. [Google Scholar] [CrossRef]
- Carvalho, M.; Câmara, G. Análise de Eventos Pontuais. In Análise Espacial de Dados Geográficos; In: Druck S, Carvalho MS, Câmara G.; Monteiro AVM.: Brasília, EMBRAPA, 2004 ISBN 85-7383-260-6.
- BRASIL. Ministério da Saúde. Secretaria de Vigilância em Saúde. Fundação Oswaldo Cruz. Introdução à Estatística Espacial para a Saúde Pública. Brasília: Ministério da Saúde, 2007.
- Floyd, T.J.; Mech, L.D.; Jordan, P.A. Relating Wolf Scat Content to Prey Consumed. J. Wildl. Manag. 1978, 42, 528–532. [Google Scholar] [CrossRef]
- Weaver, J.L. Refining the Equation for Interpreting Prey Occurrence in Gray Wolf Scats. J. Wildl. Manag. 1993, 57, 534–538. [Google Scholar] [CrossRef]
- Llaneza, L.; Fernández, A.; Nores, C. Dieta Del Lobo En Dos Zonas de Asturias (España) Que Difieren En Carga Ganadera. Doñana Acta Vertebr. 1996, 23, 201–213. [Google Scholar]
- Figueiredo, A.; Valente, A.; Barros, T.; Carvalho, J.; Silva, D.; Fonseca, C.; Madeira de Carvalho, L.; Tinoco Torres, R. What Does the Wolf Eat? Assessing the Diet of the Endangered Iberian Wolf (Canis lupus Signatus) in Northeast Portugal. PLOS ONE 2020, 15, e0230433. [Google Scholar] [CrossRef] [PubMed]
- Hutcheson, K. A Test for Comparing Diversities Based on the Shannon Formula. J. Theor. Biol. 1970, 29, 151–154. [Google Scholar] [CrossRef] [PubMed]
- Levins, R. Evolution in Changing Environments: Some Theoretical Explorations. (MPB-2) (Monographs in Population Biology, 2)—Levins, Richard: 9780691080628—AbeBooks; Princeton University Press, 1968.
- Jacobs, J. Quantitative Measurement of Food Selection. Oecologia 1974, 14, 413–417. [Google Scholar] [CrossRef] [PubMed]
- Urios, V. Eto-Ecología de La Depredación Del Lobo Canis lupus Signatus En El NO de La Península Ibérica. PhD Thesis, Tesis doctoral. Universidad de Barcelona, 1995.
- Blanco, J.C. Mamíferos de España; Planeta, 1998.
- Mateos-Quesada, P. Biología y Comportamiento Del Corzo Ibérico; Universidad de Extremadura, Servicio de Publicaciones, 2002.
- Wagner, C.; Holzapfel, M.; Kluth, G.; Reinhardt, I.; Ansorge, H. Wolf (Canis lupus) Feeding Habits during the First Eight Years of Its Occurrence in Germany. Mamm. Biol. 2012, 77, 196–203. [Google Scholar] [CrossRef]
- Alberto, M.; DAGRADI, V.; Dondina, O.; PERVERSI, M.; Milanesi, P.; LOMBARDINI, M.; RAVIGLIONE, S.; REPOSSI, A. Short-Term Responses of Wolf Feeding Habits to Changes of Wild and Domestic Ungulate Abundance in Northern Italy. Ethol. Ecol. Evol. 2014, 27, 1–23. [Google Scholar] [CrossRef]
- Mori, E.; Benatti, L.; Lovari, S.; Ferretti, F. What Does the Wild Boar Mean to the Wolf? Eur. J. Wildl. Res. 2017, 63. [Google Scholar] [CrossRef]
- Rosell, C.; Herrero, J. Sus Scrofa Linnaeus, 1758. Atlas Libro Rojo Los Mamíferos Esp. Dir. Gen. Para Biodivers.-SECEM-SECEMU Madr. 2007, 348–351.
- Fonseca, C.; Correia, F. Fonseca C, Correia F (2008). O Javali. Colecção Património Natural Transmontano. João Azevedo Editor (1.a Edição). Mirandela.; 2008; p. 168.
- Giménez-Anaya, A.; Bueno, C.G.; Fernández-Llario, P.; Fonseca, C.; García-González, R.; Herrero, J.; Nores, C.; Rosell, C. What Do We Know About Wild Boar in Iberia. In Problematic Wildlife II; Angelici, F.M., Rossi, L., Eds.; Springer International Publishing: Cham, 2020; ISBN 978-3-030-42334-6. [Google Scholar]
- Rosell, C.; Muñoz, A.; Navàs, F.; Fernández-Bou, M.; Sunyer, P.; Bonal, R.; Espelta, J.M. Principals Resultats Del Projecte “Impactboar” de Seguiment de l’activitat Del Senglar al Parc Nacional d’Aigüestortes i Estany de Sant Maurici. X Jornades Sobre Recer. Al Parc Nac. D’Aigüestortes Estany St. Mauric Dep. Territ. Sostenibilitat–Parc Nac. D’Aigüestortes Estany St. Maurici 2016, 209–218.
- Horcajada-Sánchez, F.; Barja, I. Local Ecotypes of Roe Deer Populations (Capreolus Capreolus L.) in Relation to Morphometric Features and Fur Colouration in the Centre of the Iberian Peninsula. Pol. J. Ecol. 2016, 64, 113–124. [Google Scholar] [CrossRef]
- Mattioli, L.; Capitani, C.; Gazzola, A.; Scandura, M.; Apollonio, M. Prey Selection and Dietary Response by Wolves in a High-Density Multi-Species Ungulate Community. Eur. J. Wildl. Res. 2011, 57, 909–922. [Google Scholar] [CrossRef]
- Mateos-Quesada, P.; Carranza, J. Reproductive Patterns of Roe Deer in Central Spain. Etología 2000, 8, 17–20. [Google Scholar]
- Fuller, T.K.; Mech, L.D.; Cochrane, J.F. Wolf Population Dynamics. Wolves: Behavior, Ecology, and Conservation. Univ. Chic. Press Chic. IL USA Pac. Clim. Eff. Ungulate Recruit. 2003, 481, 161–191. [Google Scholar]
- Kirilyuk, A.; Kirilyuk, V.; Minaev, A. Daily Activity Patterns of Wolves in Open Habitats in the Dauria Ecoregion, Russia. Nat. Conserv. Res. 2021, 6. [Google Scholar] [CrossRef]
- Cromsigt, J.P.G.M.; Kuijper, D.P.J.; Adam, M.; Beschta, R.L.; Churski, M.; Eycott, A.; Kerley, G.I.H.; Mysterud, A.; Schmidt, K.; West, K. Hunting for Fear: Innovating Management of Human-Wildlife Conflicts. J. Appl. Ecol. 2013, 50, 544–549. [Google Scholar] [CrossRef]
- McPhee, H.M.; Webb, N.F.; Merrill, E.H. Hierarchical Predation: Wolf (Canis lupus) Selection along Hunt Paths and at Kill Sites. Can. J. Zool. 2012, 90, 555–563. [Google Scholar] [CrossRef]
- Torretta, E.; Caviglia, L.; Serafini, M.; Alberto, M. Wolf Predation on Wild Ungulates: How Slope and Habitat Cover Influence the Localization of Kill Sites. Curr. Zool. 2017, 64. [Google Scholar] [CrossRef]
Figure 1.
Scats densities in forest regions of the study area using Kernel density technique. The colors represent varying densities of collected scats. Red lines mark the forest regions, black lines separate Castilla y León (northwest) from Madrid (southeast), green lines represent Sierra de Guadarrama National Park, and pink lines represent Sierra del Rincón Biosphere Reserve.
Figure 1.
Scats densities in forest regions of the study area using Kernel density technique. The colors represent varying densities of collected scats. Red lines mark the forest regions, black lines separate Castilla y León (northwest) from Madrid (southeast), green lines represent Sierra de Guadarrama National Park, and pink lines represent Sierra del Rincón Biosphere Reserve.
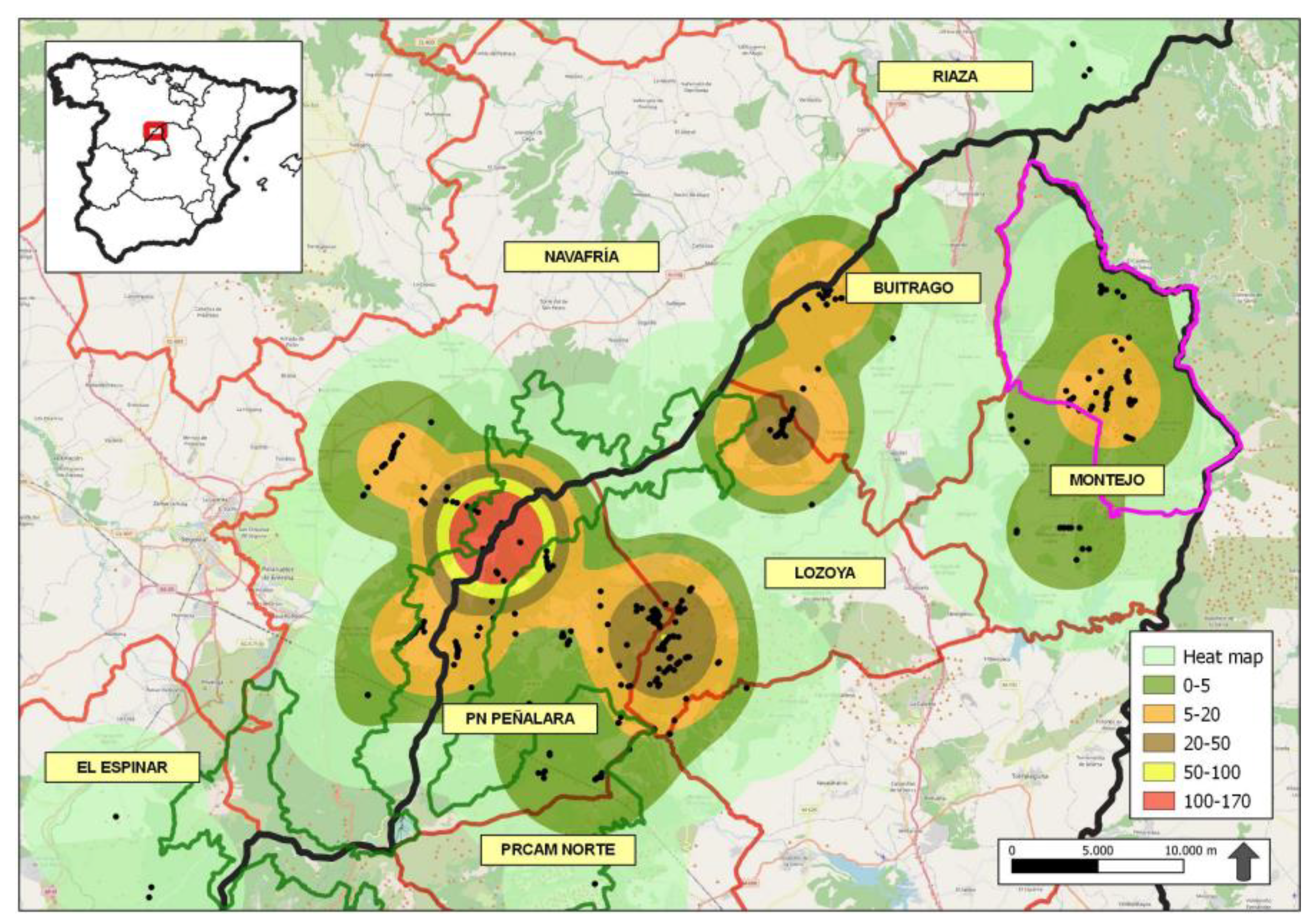
Figure 2.
Consumption of wild and domestic ungulates in relation to seasons: A) % prey occurrence; B) % ingested biomass. Sample size in Autumn: n=189; Winter: n=111; Spring: 200; Summer: n=137.
Figure 2.
Consumption of wild and domestic ungulates in relation to seasons: A) % prey occurrence; B) % ingested biomass. Sample size in Autumn: n=189; Winter: n=111; Spring: 200; Summer: n=137.
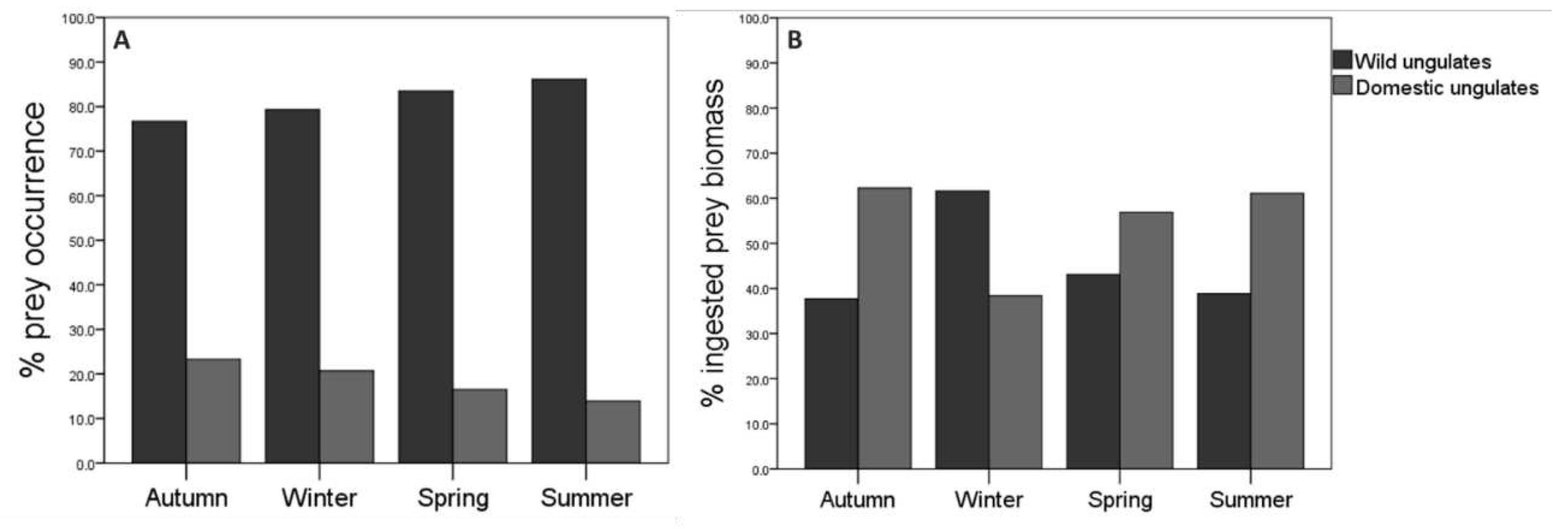
Figure 3.
Consumption of wild and domestic ungulates in relation to years: A) % prey occurrence; B) % prey biomass ingested. Sample size 2017: n=109; 2018: n=169; 2019: n=287; 2020: n=30; 2021: n=42.
Figure 3.
Consumption of wild and domestic ungulates in relation to years: A) % prey occurrence; B) % prey biomass ingested. Sample size 2017: n=109; 2018: n=169; 2019: n=287; 2020: n=30; 2021: n=42.
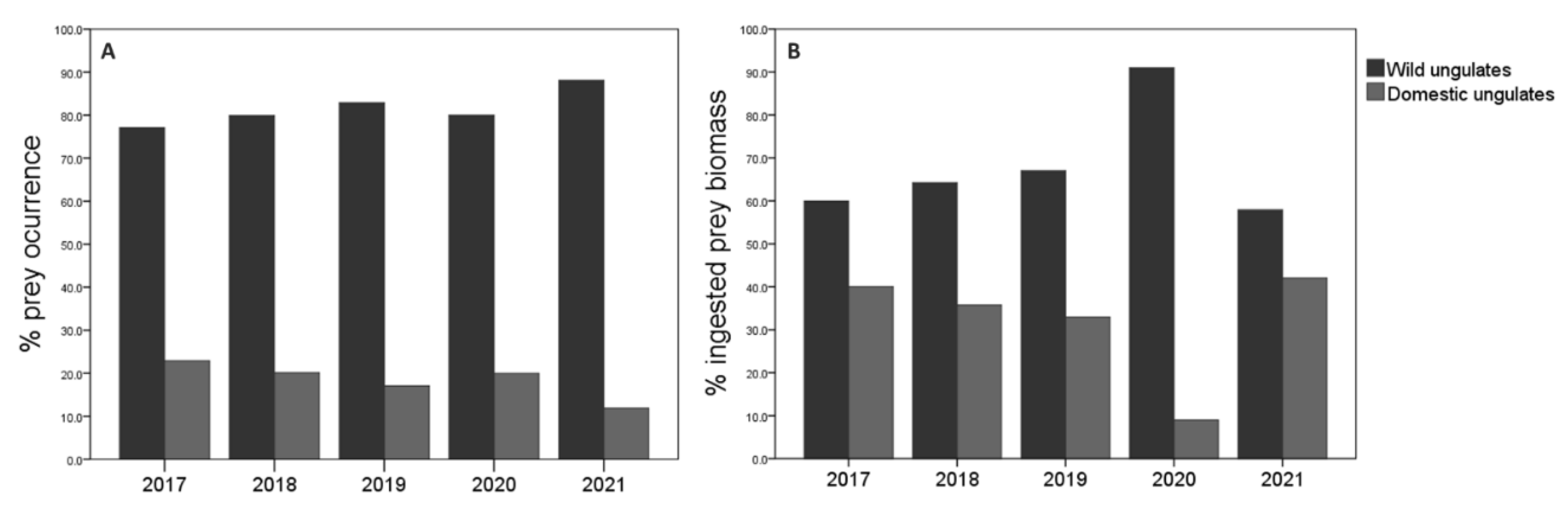
Figure 4.
Consumption of wild and domestic ungulates in relation to forest regions: A) % occurrence; B) % ingested biomass. Sample size in El Espinar: n=4; PRCAM Norte: n=2; PN Peñalara: n=262; Lozoya: n=127; Montejo: n=50; Buitrago: n=30; Navafría: n=135; Riaza: n=3.
Figure 4.
Consumption of wild and domestic ungulates in relation to forest regions: A) % occurrence; B) % ingested biomass. Sample size in El Espinar: n=4; PRCAM Norte: n=2; PN Peñalara: n=262; Lozoya: n=127; Montejo: n=50; Buitrago: n=30; Navafría: n=135; Riaza: n=3.
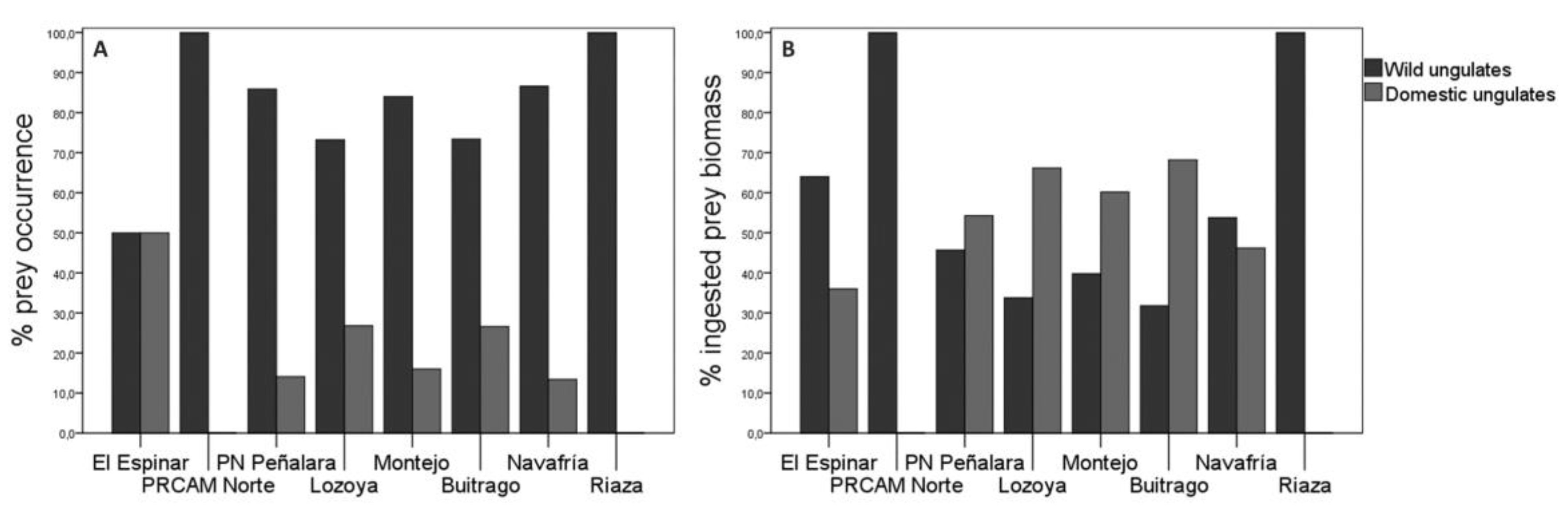
Table 1.
Composition of Iberian wolf diet for 5 years (2017-2021) in Sierra de Guadarrama National Park and surroundings based on 671 scats. The ingested biomass (kg) was calculated using body masses obtained from the literature ([23,76,81,82,83] Horcajada pers. comm. 2022).
| Prey occurrence | Ingested biomass | Prey mean mass (Kg) | ||||||
|---|---|---|---|---|---|---|---|---|
| Prey | N | % | Kg | % | Adult | Youth | Mean | |
| Wild ungulates | Roe deer | 233 | 34.7 | 500.7 | 13.8 | 24.5 | 7.0 | 15.8 |
| Red deer | 2 | 0.3 | 10.1 | 0.3 | 90.0 | 25.0 | 57.5 | |
| Wild boar | 294 | 43.9 | 1006.3 | 27.7 | 75.0 | 22.0 | 48.5 | |
| Mountain goat | 21 | 3.1 | 69.7 | 1.9 | 61.0 | 11.0 | 36.0 | |
| Total | 550 | 82.0 | 1586.7 | 43.7 | ||||
| Domestic ungulates | Cattle | 55 | 8.2 | 1927.2 | 53.0 | 750.0 | 115.0 | 432.5 |
| Domestic goat | 28 | 4.2 | 47.5 | 1.3 | 26.3 | 5.0 | 15.7 | |
| Sheep | 15 | 2.2 | 26.8 | 0.7 | 28.5 | 5.0 | 16.8 | |
| Horse | 2 | 0.3 | 49.7 | 1.3 | 550.0 | 60.0 | 305.0 | |
| Unidentified livestock | 21 | 3.1 | - | - | - | - | - | |
| Total | 121 | 18.0 | 2051.0 | 56.3 | ||||
| Total | 671 | 100.0 | 3637.8 | 100.0 | ||||
Table 2.
Number of canid attacks reported by gamekeepers in 2020 and 2021 registered by the Consejería de Medio Ambiente, Vivienda y Agricultura de la Comunidad de Madrid (Comunidad de Madrid, unpublished data) compared to occurrence of domestic ungulates in scats from 2017 to 2021 and to total scats collected each year in each region.
Table 2.
Number of canid attacks reported by gamekeepers in 2020 and 2021 registered by the Consejería de Medio Ambiente, Vivienda y Agricultura de la Comunidad de Madrid (Comunidad de Madrid, unpublished data) compared to occurrence of domestic ungulates in scats from 2017 to 2021 and to total scats collected each year in each region.
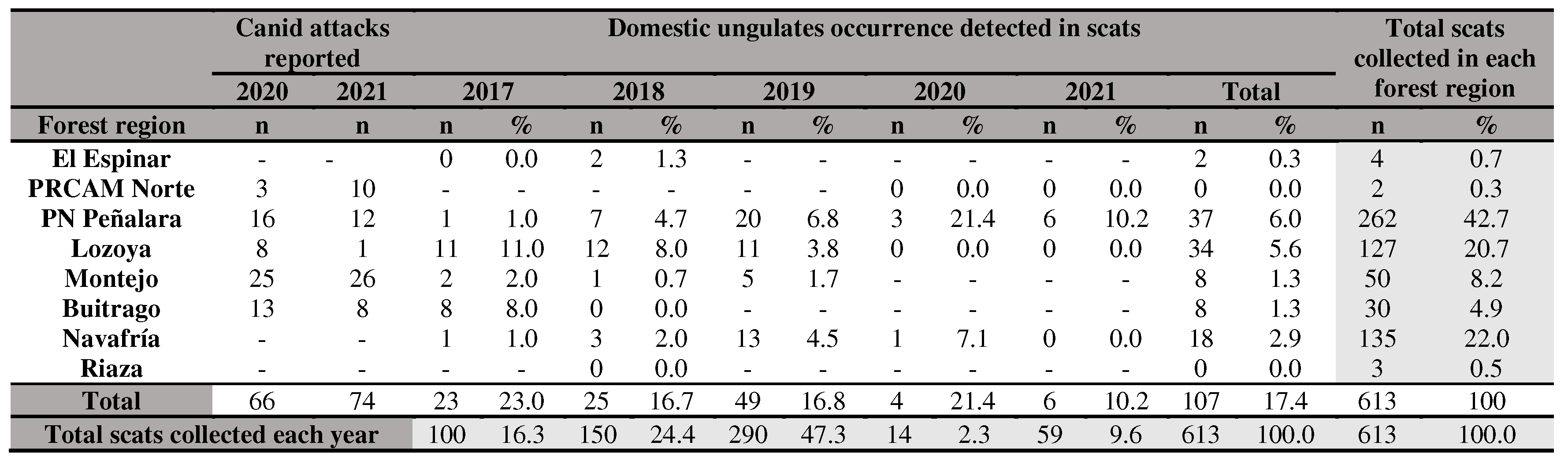
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



