
Submitted:
29 September 2023
Posted:
30 September 2023
You are already at the latest version
Abstract
Powdery mildew (PM), caused by the fungal pathogen Blumeria graminis f. sp. tritici (Bgt), significantly threatens global bread wheat production. Although the use of resistant cultivars is an effective strategy for managing PM, currently available wheat cultivars lack sufficient levels of resistance. To tackle this challenge, we conducted a comprehensive genome-wide association study (GWAS) using a diverse panel of 286 bread wheat genotypes. Over three consecutive years (2020-21, 2021-22, and 2022-23), these genotypes were extensively evaluated for PM severity under field conditions following inoculation with virulent Bgt isolates. The panel was previously genotyped using the Illumina 90K SNP Infinium iSelect SNP assay to obtain genome-wide SNP marker coverage. By applying FarmCPU, a multilocus mixed model, we identified a total of 113 MTAs located on chromosomes 1A, 1B, 2B, 3A, 3B, 4A, 4B, 5A, 5B, 6B, 7A, and 7B at a significance level of p≤0.001. Notably, four novel MTAs on chromosome 6B were consistently detected in 2020-21 and 2021-22 environments. Furthermore, within the confidence intervals of the identified SNPs, we identified 96 candidate genes belonging to different proteins including 12 disease resistance/host-pathogen interactions related protein families. Among these, protein kinases, leucine-rich repeats, and zinc finger proteins, were of particular interest due to their potential roles in PM resistance. These identified loci can serve as targets for breeding programs aimed at developing disease-resistant wheat cultivars.
Keywords:
powdery mildew
; wheat
; GWAS
; FarmCPU
; MTAs
; candidate genes
1. Introduction
Bread wheat (Triticum aestivum L.) is a vital source of nutrients and serves as the primary staple food crop for approximately 35% of the global population [1]. In 2022, worldwide wheat consumption increased from 783 million metric tons to 792.69 million metric tons [2]. This rise in wheat consumption is necessary for global food security, but has led to extra burdens to wheat production system which is already challenged by the prevalence of pests and diseases such as powdery mildew (PM), a devastating wheat disease.
PM is caused by the fungal pathogen Blumeria graminis (DC.) E.O. Speer f. sp. tritici Em. Marchal, (syn. Erysiphe graminis f. sp. tritici) (Btg). It is the third most destructive disease of wheat, causing yield losses ranging from 13% to 34% under low infestation and 50% to 100% under severe infestation in various wheat-growing regions [3,4]. Environmental factors like temperature and relative humidity significantly influence the development and spread of this disease, contributing to epidemics [5]. In the Himalayan regions of India, PM is a significant factor contributing to reduced grain yield [6]. Breeding approaches have proven effective in developing PM-resistant wheat varieties to overcome yield losses and increase economic production. Using resistant varieties is currently considered an efficient, environmentally safe, and successful strategy to control PM and mitigate production losses.
Major effect genes and minor-to-moderate effects PM resistance genes are generally inherited qualitatively and quantitatively, respectively, and have also been identified in wheat and integrated into wheat resistance breeding programs. These includes over 100 alleles distributed over >60 major loci (Pm1–Pm68: Pm8 is allelic to Pm17; Pm18 = Pm1c; Pm22 = Pm1e; Pm23 = Pm4c, and Pm31 = Pm21) and many valuable quantitative trait loci (QTLs) distributed across wheat chromosomes [7,8,9,10,11,12,13,14,15,16,17,18,19,20,21,22,23,24,25]. Whereas some of the PM resistance genes, such as Pm38, Pm39, and Pm46, are inherited quantitatively [26,27,28] , there are other genes which possesses with many resistance alleles. For instance, Pm3 exhibits the highest allelic diversity, with 17 variants, followed by Pm1 and Pm5, each having 5 alleles. Pm4 and Pm2, on the other hand, possess 4 and 3 alleles, respectively [11].
The PM resistance genes have primarily been transferred from both domesticated and wild relatives and from other species such as rye (Secale cereale) to wheat [29]. However, their utilization in wheat breeding is challenged by linkage drags [15]. For instance, Pm8 gene, which derived from S. cereale and also significantly contributed to PM resistance in wheat breeding during the 1990s, is linked with secalin glycopeptide locus on 1RS chromosome segment which causes a decline in flour quality [30]. Furthermore, many of the identified resistance genes have been overcome by the pathogen in different countries, leaving only a few that are still effective against current Bgt isolates in the field [31,32]. Thus, the continuous identification, characterization and deployment of new resistance genes is essential to minimize economic losses.
The declining cost of genotyping and the increasing availability of diverse single nucleotide polymorphisms (SNPs) fingerprinting platforms has furthered the identification and deployment of new resistance genes. Many genetic mapping methods are now available to accommodate different types of traits, mapping populations, and study objectives [33]. These methods can be categorized into three main groups based on the principle and genetic material used: (i) QTL mapping using a biparental population, (ii) association mapping or genome-wide association studies (GWAS) in a diverse set of germplasm, and (iii) genetic mapping utilizing the multi-parent populations. The first two approaches, QTL mapping and GWAS, have successfully identified genes and their markers associated with disease resistance [34,35,36,37]. However, the QTL mapping approach, which relies on a segregating population generated from parents exhibiting contrasting trait performances [12], faces limitations in QTL detection power. The scarcity of recombination events and genetic variation in such populations often leads to identifying only a few QTLs. To overcome this limitation, advanced wheat populations such as ‘multiparent advanced generation inter-cross’ (MAGIC) have been introduced to enhance the resolution of traditional mapping [38]. Nevertheless, challenges arise in validating map order and analyzing recombination due to using bi-allelic markers in these populations [39]. Conversely, GWAS circumvents the need for developing populations by conducting analyses on diverse germplasm [40] and offers several advantages such as the utilization of larger sample sizes, which facilitates the exploration of a broader range of genetic variations including those with minor effects and allelic diversity [36,41]. Additionally, GWAS enables higher-resolution mapping, identifying candidate genes associated with the trait of interest [42].
Thus, the objectives of the present study were to identify potentially novel loci that confer effective PM resistance at the adult plant stage utilizing a panel of 286 wheat accessions known as ‘wheat association mapping initiative’ (WAMI) panel created by CIMMYT, Mexico, identify potential PM resistance-associated markers and candidate genes, and compare identified loci with the previously reported genes/QTLs and identify candidate genes using comparative analyses. Leveraging landmark resources such as the high-density 90K SNP array [43] and the Ensemble plants database [44], we aims to address the aforementioned objectives and shed light on the genetic basis of PM resistance in wheat using a GWAS study.
2. Results
2.1. Phenotypic Analysis
The phenotypic analysis conducted in specific environments showed significant variation for disease severity in the WAMI panel. Table 1 provides a summary of the phenotypic variation for PM disease severity. The broad sense heritability for PM disease severity ranged from 65% to 89% (Table 1). The analysis of variance indicated that both genotypic and environment as well as genotype-environment interactions had highly significant (P ≤ 0.001) estimated variance components for disease response to PM (Table 2). However, despite environmental variation, significant positive correlations (> 0.6 for each environment) were observed for the disease severity among the three environments (Figure 1A).
2.2. Population Structure Analysis
Principal components analysis indicated the existence of five distinct sub-groups within the WAMI panel (Figure 2B). PC1 accounted for 43.3% of the total variance, while PC2 explained 18.8%. These results suggest that the combination of PC1 and PC2 captures a significant portion of the underlying genetic variation within the panel, allowing for identifying and differentiating the sub-groups present in the WAMI population.
2.3. Genome-wide Marker Trait Association (MTA) Analysis
A scatter plot for r2 values of pairwise markers showing genome-wide linkage disequilibrium (LD) decay for 286 genotypes of the WAMI panel assessed using the Hill and Weir formula [45] is given in Figure 1C. The average r2 values of the genome was 0.12. The LD decay curve intersected with the threshold (r2 = 0.23) at 3.0 cM (Figure 1C). This showed the ± 3.0 cM as the genome-wide critical distance to detect linkage and single QTLs as described earlier [46].
The comprehensive genome-wide MTA analysis encompassed three distinct models: GLM, MLM, and FarmCPU; however, the final GWAS analysis exclusively incorporated the best-fitting FarmCPU model. Over the course of the three-year investigation, a total of 19, 85, and 9 MTAs were successfully identified in each respective year (Figure 2 and Table 3). Figure 2 displays the frequency distribution of the best linear unbiased prediction (BLUP) values for PM disease severity for three consecutive years namely, 2020-21, 2021-22, and 2022-23, the Manhattan plot illustrating the results of genome-wide association scans, and the Quantile-Quantile (Q-Q) plot of p-values, comparing the observed -lg (P) for PM resistance to Bgt isolates in FarmCPU method.
In the initial year (2020-21), the distribution of MTAs spanned multiple chromosomes, with notable occurrences on the chromosome 1A (2 MTAs), 2B (4 MTAs), 6B (9 MTAs), and 7A (4 MTAs). Notably, a striking concentration of 9 MTAs emerged at a specific genetic position (59-60 cM) on chromosome 6B. This intriguing observation suggests the presence of a potential genomic “hotspot” linked to PM resistance, signifying the collective impact of multiple genetic variants.
Transitioning to the subsequent year (2021-22), the landscape of MTAs extended across various chromosomes: chromosome 2B (1 MTA), 3B (9 MTAs), 4A (19 MTAs), 5A (1 MTA), 5B (2 MTAs), 6B (48 MTAs), and 7B (3 MTAs). Noteworthy is the aggregation of MTAs at 48 cM on chromosome 4A, underscoring another genomic hotspot exerting influence on PM resistance. The 6B chromosome also exhibited remarkable association richness, hosting 49 MTAs. Within this cluster, 8 MTAs were anchored at 59 cM, 77 MTAs at 64 cM, and 3 MTAs at 65 cM, further reinforcing the pivotal role of the 6B chromosome region in contributing to the observed phenotypic variance for PM resistance.
Concluding the study in the final year (2022-23), a reduced yet significant number of MTAs were identified, totaling 9 associations distributed across chromosomes 1B (1 MTA), 3A (1 MTA), 3B (3 associations), 7A (2 MTAs), and 7B (1 MTA).
Figure 2.
PM disease severity distribution, marker-trait associations and quantile-quantile (Q-Q) plots distributions. Histograms on left (A, D and G) display the frequency distribution of BLUP estimated for PM disease severity for three consecutive years, 2020-21, 2021-22, and 2022-23. Manhattan plots in the middle (B, E, and H) are illustrating the results of genome-wide association scans for PM resistance using BLUP of the 286 lines of WAMI panel across 2020-21, 2021-22, and 2022-23, respectively. The significant thresholds are represented by solid yellow lines, while the number of effective markers is indicated by the yellow lines. The Q-Q plots of p-values on right (C, F, and I) comparing the observed -lg (P) for PM resistance to Bgt isolates in FarmCPU analyses to the expected distribution of -lg (P) under a uniform distribution.
Figure 2.
PM disease severity distribution, marker-trait associations and quantile-quantile (Q-Q) plots distributions. Histograms on left (A, D and G) display the frequency distribution of BLUP estimated for PM disease severity for three consecutive years, 2020-21, 2021-22, and 2022-23. Manhattan plots in the middle (B, E, and H) are illustrating the results of genome-wide association scans for PM resistance using BLUP of the 286 lines of WAMI panel across 2020-21, 2021-22, and 2022-23, respectively. The significant thresholds are represented by solid yellow lines, while the number of effective markers is indicated by the yellow lines. The Q-Q plots of p-values on right (C, F, and I) comparing the observed -lg (P) for PM resistance to Bgt isolates in FarmCPU analyses to the expected distribution of -lg (P) under a uniform distribution.
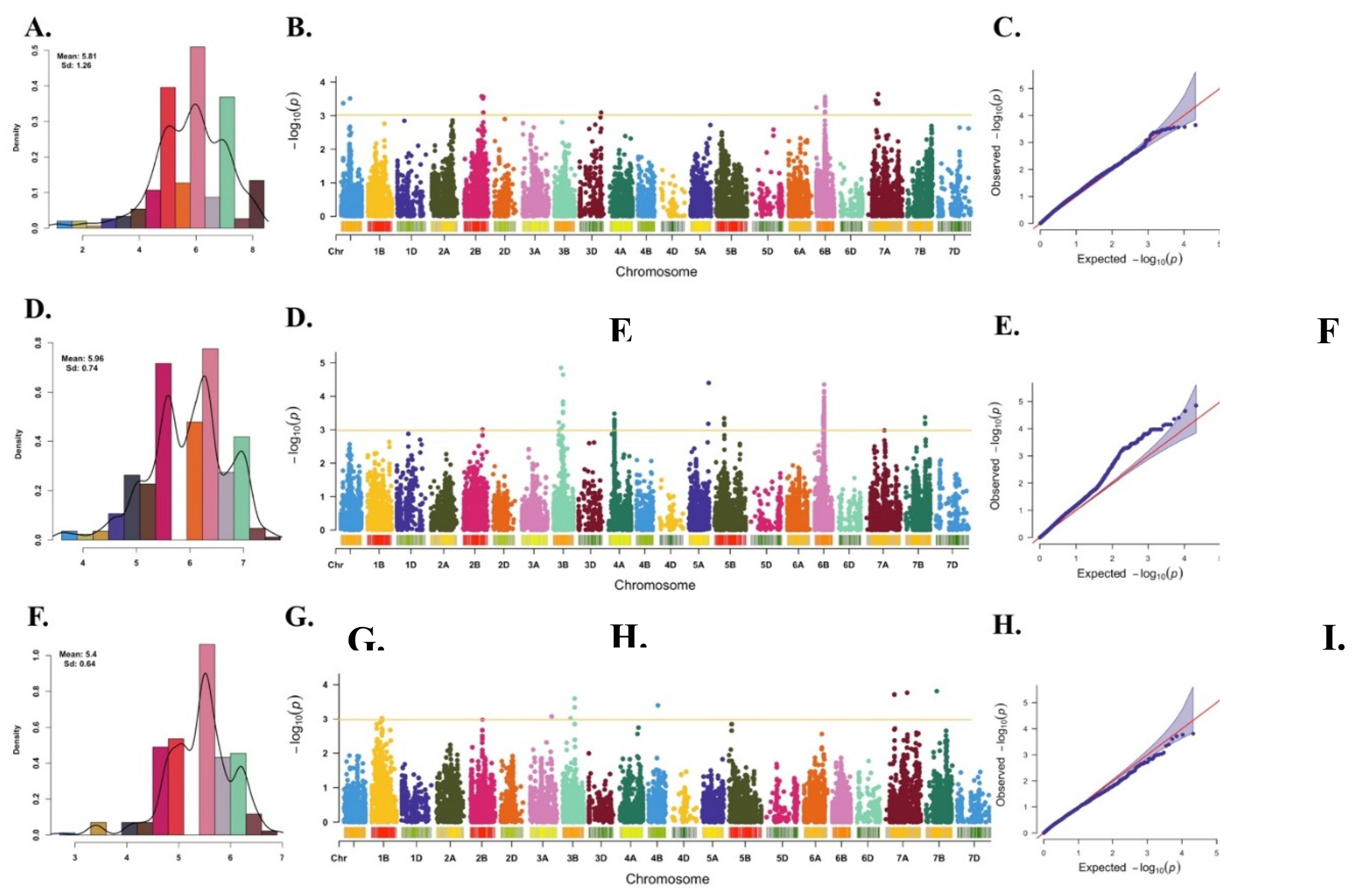
Interestingly, four common MTAs were consistently observed in crop seasons 2020-21 and 2021-22 located on chromosome 6B (Table 3). Notably, certain chromosomal regions, such as 2B (132 – 144 cM), 3B (57 – 71 cM), 6B (59 – 65 cM), and 7A (57 – 74 cM), also exhibited repeated associations with PM resistance, indicating their crucial role in the genetic variation of PM resistance. These findings provide valuable insights for future studies aiming to understand the genetic architecture of PM resistance and for targeted breeding efforts to enhance it in commercial cultivars through marker-assisted selection.

Table 3.
Details of marker-trait associations (MTAs) detected for resistance against PM in WAMI panel during 2020-21, 2021-22, and 2022-23.
Table 3.
Details of marker-trait associations (MTAs) detected for resistance against PM in WAMI panel during 2020-21, 2021-22, and 2022-23.
| Marker | Chr | Pos (cM)1 | Pos (bp)2 | Effect | p value |
|---|---|---|---|---|---|
| 2020-21 | |||||
| RAC875_c1710_376$ | 1A | 27.0 | 9103721 | -0.55 | 0.0004 |
| Kukri_c35200_895$ | 1A | 74.0 | 459514896 | -0.59 | 0.0003 |
| BS00080318_51$ | 2B | 132.0 | 763842555 | 0.32 | 0.0003 |
| Excalibur_c5438_274$ | 2B | 142.0 | 774958099 | 0.30 | 0.0003 |
| Excalibur_rep_c109577_698$ | 2B | 144.0 | 775368259 | -0.27 | 0.0008 |
| RAC875_rep_c83950_222$ | 2B | 144.0 | 775053184 | -0.29 | 0.0003 |
| Ra_c22493_190$ | 6B | 0.0 | 6196794 | -0.48 | 0.0006 |
| Excalibur_rep_c106789_271 | 6B | 59.0 | 153957925 | -0.33 | 0.0004 |
| RAC875_c25489_1208 | 6B | 59.0 | 153959656 | -0.31 | 0.0008 |
| wsnp_Ex_c39304_46635517 | 6B | 59.0 | 151609970 | -0.34 | 0.0003 |
| wsnp_Ex_rep_c102044_87296690 | 6B | 59.0 | 153959656 | -0.32 | 0.0005 |
| wsnp_Ex_rep_c102044_87297599 | 6B | 59.0 | 153957925 | -0.34 | 0.0003 |
| wsnp_Ra_c48999_54089942 | 6B | 59.0 | 153956896 | -0.31 | 0.0008 |
| Excalibur_c3795_198$ | 6B | 60.0 | 156266046 | -0.31 | 0.0005 |
| Ra_c14852_1487 | 6B | 60.0 | 156266570 | -0.32 | 0.0003 |
| RAC875_c66770_208 | 7A | 57.0 | 26879337 | -0.58 | 0.0004 |
| Excalibur_c7538_2718$ | 7A | 65.0 | 35602223 | -0.48 | 0.0004 |
| wsnp_Ex_c35_77935$ | 7A | 73.0 | 41960564 | -0.36 | 0.0002 |
| BS00023055_51 | 7A | 74.0 | 41959666 | -0.32 | 0.0004 |
| 2021-22 | |||||
| Excalibur_c7971_1573 | 2B | 144.0 | 775371388 | 0.04 | 0.0009 |
| BS00011728_51$ | 3B | 57.0 | 59064340 | -0.28 | 0.0000 |
| BS00022741_51$ | 3B | 61.0 | 66953509 | -0.26 | 0.0009 |
| Excalibur_c21372_142 | 3B | 61.0 | 66954023 | -0.26 | 0.0009 |
| Tdurum_contig51993_52$ | 3B | 61.0 | 66953509 | -0.26 | 0.0009 |
| BS00011869_51 | 3B | 71.0 | 426452313 | -0.23 | 0.0003 |
| Excalibur_c80041_400$ | 3B | 71.0 | 429610611 | -0.26 | 0.0000 |
| Kukri_c21818_519$ | 3B | 71.0 | 556059780 | -0.23 | 0.0002 |
| RAC875_c58159_989 | 3B | 71.0 | 569425136 | -0.19 | 0.0008 |
| wsnp_Ku_c21818_31604716$ | 3B | 71.0 | 556059565 | -0.24 | 0.0001 |
| BobWhite_c27944_234$ | 4A | 48.0 | 109443802 | -0.23 | 0.0007 |
| Ex_c17894_1159$ | 4A | 48.0 | 654418148 | -0.22 | 0.0005 |
| Excalibur_rep_c66815_273 | 4A | 48.0 | 104375678 | -0.22 | 0.0005 |
| GENE-2637_94$ | 4A | 48.0 | 164633479 | -0.23 | 0.0008 |
| IAAV3906$ | 4A | 48.0 | 113854919 | -0.22 | 0.0006 |
| IAAV8784 | 4A | 48.0 | 135358615 | -0.22 | 0.0005 |
| IACX1896$ | 4A | 48.0 | 109431746 | -0.22 | 0.0007 |
| Kukri_c44469_1240 | 4A | 48.0 | 136432865 | -0.22 | 0.0005 |
| Kukri_c48155_158$ | 4A | 48.0 | 120605031 | -0.22 | 0.0005 |
| RAC875_c22562_429$ | 4A | 48.0 | 111292238 | -0.23 | 0.0003 |
| RAC875_rep_c74695_101$ | 4A | 48.0 | 164632278 | -0.23 | 0.0008 |
| tplb0035b22_184 | 4A | 48.0 | 135614180 | -0.22 | 0.0005 |
| wsnp_BE442869A_Ta_2_1 | 4A | 48.0 | 140134106 | -0.22 | 0.0005 |
| wsnp_Ex_c10527_17198865$ | 4A | 48.0 | 115570013 | -0.22 | 0.0005 |
| wsnp_Ex_c1387_2659020$ | 4A | 48.0 | 115912754 | -0.22 | 0.0005 |
| wsnp_Ex_c14529_22547438 | 4A | 48.0 | 115913618 | -0.22 | 0.0005 |
| wsnp_Ex_c1865_3515470$ | 4A | 48.0 | 136894495 | -0.22 | 0.0005 |
| wsnp_Ex_c36141_44153175$ | 4A | 48.0 | 111091261 | -0.23 | 0.0007 |
| wsnp_Ex_c4286_7734046 | 4A | 48.0 | 114744375 | -0.22 | 0.0006 |
| wsnp_Ex_c43734_49968808$ | 4A | 48.0 | 211000047 | -0.24 | 0.0005 |
| wsnp_Ex_rep_c69890_68851948$ | 4A | 48.0 | 105249095 | -0.24 | 0.0005 |
| Tdurum_contig51134_191$ | 5A | 144.0 | 698501228 | -0.36 | 0.0000 |
| RAC875_c19099_308$ | 5B | 68.0 | 519153210 | 0.24 | 0.0007 |
| Tdurum_contig53926_455$ | 5B | 68.0 | 516469300 | 0.26 | 0.0006 |
| BS00046963_51 | 6B | 59.0 | 150665070 | -0.19 | 0.0006 |
| Excalibur_rep_c106789_271 | 6B | 59.0 | 153957925 | -0.20 | 0.0005 |
| Kukri_c38732_225$ | 6B | 59.0 | 151131387 | -0.18 | 0.0007 |
| wsnp_Ex_rep_c102044_87296690 | 6B | 59.0 | 153959656 | -0.19 | 0.0007 |
| wsnp_Ex_rep_c102044_87297599 | 6B | 59.0 | 153957925 | -0.20 | 0.0004 |
| Ra_c14852_1487$ | 6B | 60.0 | 156266570 | -0.18 | 0.0006 |
| BS00040868_51$ | 6B | 63.0 | 228977167 | -0.24 | 0.0001 |
| BobWhite_c34920_228 | 6B | 64.0 | 249498169 | -0.21 | 0.0006 |
| BS00003955_51$ | 6B | 64.0 | 260548319 | -0.24 | 0.0001 |
| BS00021686_51$ | 6B | 64.0 | 174566651 | -0.24 | 0.0003 |
| BS00045761_51$ | 6B | 64.0 | 227070547 | -0.24 | 0.0002 |
| BS00067871_51$ | 6B | 64.0 | 261585651 | -0.24 | 0.0002 |
| BS00067873_51$ | 6B | 64.0 | 261585682 | -0.23 | 0.0003 |
| BS00080544_51$ | 6B | 64.0 | 234560063 | -0.22 | 0.0007 |
| Ex_c49055_617$ | 6B | 64.0 | 260548488 | -0.22 | 0.0005 |
| Excalibur_c20503_382$ | 6B | 64.0 | 257568556 | -0.24 | 0.0001 |
| Excalibur_c47738_334 | 6B | 64.0 | 262473204 | -0.22 | 0.0004 |
| Excalibur_c5136_2314$ | 6B | 64.0 | 231677537 | -0.22 | 0.0003 |
| Excalibur_c53834_416$ | 6B | 64.0 | 257993802 | -0.25 | 0.0000 |
| Excalibur_c79066_165$ | 6B | 64.0 | 232168465 | -0.24 | 0.0002 |
| GENE-2606_197$ | 6B | 64.0 | 257567774 | -0.25 | 0.0001 |
| Kukri_c25377_106$ | 6B | 64.0 | 174567929 | -0.23 | 0.0005 |
| Kukri_c38058_532 | 6B | 64.0 | 257567774 | -0.22 | 0.0009 |
| Kukri_c52515_442 | 6B | 64.0 | 259877872 | -0.21 | 0.0005 |
| Ra_c77985_260$ | 6B | 64.0 | 214089373 | -0.22 | 0.0008 |
| RAC875_c12805_908$ | 6B | 64.0 | 234559907 | -0.23 | 0.0005 |
| RAC875_c24962_1326 | 6B | 64.0 | 229280173 | -0.23 | 0.0003 |
| RFL_Contig311_951 | 6B | 64.0 | 231354317 | -0.23 | 0.0003 |
| TA005139-0719$ | 6B | 64.0 | 234559694 | -0.24 | 0.0004 |
| wsnp_BQ161448B_Ta_2_1$ | 6B | 64.0 | 261530574 | -0.23 | 0.0002 |
| wsnp_Ex_c1603_3056226 | 6B | 64.0 | 221813724 | -0.23 | 0.0004 |
| wsnp_Ex_c23474_32717535$ | 6B | 64.0 | 276221225 | -0.25 | 0.0001 |
| wsnp_Ex_c27934_37093614 | 6B | 64.0 | 259881611 | -0.24 | 0.0002 |
| wsnp_Ex_c42372_48966781 | 6B | 64.0 | 229280173 | -0.21 | 0.0007 |
| wsnp_Ex_c46160_51746546 | 6B | 64.0 | 229278919 | -0.23 | 0.0002 |
| wsnp_Ex_rep_c103466_88415738 | 6B | 64.0 | 274209042 | -0.24 | 0.0002 |
| wsnp_Ex_rep_c103466_88415994 | 6B | 64.0 | 274208315 | -0.22 | 0.0006 |
| wsnp_Ex_rep_c103497_88437811$ | 6B | 64.0 | 277165618 | -0.24 | 0.0001 |
| wsnp_Ex_rep_c68480_67305954 | 6B | 64.0 | 259292578 | -0.24 | 0.0001 |
| wsnp_JD_c6448_7610859 | 6B | 64.0 | 257993802 | -0.24 | 0.0001 |
| wsnp_Ku_c27423_37369145 | 6B | 64.0 | 259883491 | -0.21 | 0.0005 |
| wsnp_Ra_c16850_25605248 | 6B | 64.0 | 262473278 | -0.23 | 0.0002 |
| wsnp_Ra_c33358_42248399$ | 6B | 64.0 | 257993727 | -0.22 | 0.0004 |
| wsnp_Ra_rep_c111161_93528347$ | 6B | 64.0 | 277163269 | -0.21 | 0.0005 |
| wsnp_Ra_rep_c73725_71801179 | 6B | 64.0 | 274207889 | -0.21 | 0.0006 |
| wsnp_Ra_rep_c73725_71801237 | 6B | 64.0 | 274207831 | -0.21 | 0.0005 |
| wsnp_Ra_rep_c73731_71807419 | 6B | 64.0 | 257996714 | -0.24 | 0.0001 |
| BobWhite_c32911_243$ | 6B | 65.0 | 255268901 | -0.22 | 0.0003 |
| Kukri_c24148_254$ | 7B | 136.0 | 705270901 | -0.16 | 0.0007 |
| TA005284-0990$ | 7B | 136.0 | 704271881 | -0.16 | 0.0006 |
| wsnp_JD_c13673_13606066$ | 7B | 136.0 | 704271648 | -0.17 | 0.0004 |
| 2022-23 | |||||
| IACX3595$ | 1B | 76.0 | 539562016 | 0.17 | 0.0009 |
| BS00030652_51 | 3A | 158.0 | 693002205 | -0.25 | 0.0008 |
| Excalibur_c8284_580$ | 3B | 57.0 | 736672449 | -0.20 | 0.0009 |
| IAAV6566 | 3B | 84.0 | 736712583 | -0.17 | 0.0003 |
| Tdurum_contig8365_433 | 3B | 84.0 | 736672399 | -0.17 | 0.0005 |
| Kukri_c2526_1375$ | 4B | 66.0 | 523447728 | -0.28 | 0.0004 |
| Excalibur_c25471_225 | 7A | 64.0 | 34539197 | 0.17 | 0.0002 |
| BS00094965_51$ | 7A | 150.0 | 669729091 | -0.30 | 0.0002 |
| Kukri_c30836_582 | 7B | 73.0 | 699427808 | 0.16 | 0.0002 |
1 = Marker position (genetic) on linkage map in cM; 2 = Marker position (physical) on chromosome in bp, and $ = Marker with candidate genes that encode a protein.
2.4. Candidate Genes
In our study, we investigated genomic regions to find candidate genes within a specific window of 200 kilobases (kb) surrounding each MTA for their association with PM resistance. This window consisted of 100 kb on each side of the MTA. We discovered a total of 94 unique CGs (89 encoding known protein domains) in 51 genomic intervals for 65 MTAs (Table S1), while no CG were spotted in defined genomic regions of the remaining 48 MTAs.
Further analysis of these 94 CGs involved screening them for the presence of genes that are already known to play a role in various pathways related to the interactions between pathogens and their host organisms. The results of this screening, as presented in Table S2 revealed that 30 CGs encoded a diverse range of protein domains relevant to plant defense and the interactions between pathogens and hosts. The identified proteins encompassed a wide array of functional domains and families. They included the following: (i) ABC transporter-like, ATP-binding domain (ii) Ankyrin repeat (iii) Aspartic peptidase A1 family (iv) Peroxidase (v) Cytochrome P450 (vi) Disease resistance protein (NB-LRR) (vii) Glutathione S-transferase (viii) P-loop containing nucleoside triphosphate hydrolase (ix) Kinase like domain superfamily (including Serine-threonine/tyrosine/cysteine-protein kinase) (x) Wall-associated receptor kinase, galacturonan-binding domain (xi) Zinc finger proteins and (xii) F-box superfamily. These CGs and their corresponding proteins are directly or indirectly involved in the response of the host organism to pathogen attacks.
3. Discussion
To uncover novel PM resistance genes/MTAs, a series of field experiments were conducted over three years: 2020-21, 2021-22, and 2022-23, utilizing the diverse WAMI panel of common wheat. The analysis of PM disease severity unveiled a wide spectrum of phenotypic variations, displaying normal distributions in two of the three environments (2020-21 and 2022-23), and a near binomial distribution in the remaining environment (2021-22) (Figure 2). This distribution pattern suggests the involvement of major and multiple quantitative trait loci (QTLs) that contribute to PM resistance. As indicated by the phenotypic analysis, GWAS analysis also identified a total of 113 MTAs associated with adult plant resistance against PM (Table 3). This can be explained by the extensive diversity present for PM resistance within the WAMI panel [47]. The presence of multiple PM resistance QTLs/MTAs in wheat germplasm sourced from diverse countries has also been reported previously [16], underlining the intricate and diverse genetic foundation of PM resistance. These findings highlight the complexity and diversity of the genetic basis underlying PM resistance and reinforce the importance of the WAMI panel in dissecting PM resistance and emphasizing the need for further investigations to unravel the specific PM resistance mechanisms involved in different wheat populations. All the important MTAs are discussed in greater detail below.
3.1. Investigating the Concordance among Significant MTAs and Previously Mapped QTL/Pm Resistance Genes
This study identified a total of 113 MTAs (19, 85 and 9 MTAs in 2020-21, 2021-22 and 2022-23, respectively) with some notable findings such as clusters of MTAs at the same position (Table 3). Upon comparing the position of markers associated with significant MTAs identified in this study with the previously reported MTAs/QTLs and Pm resistance genes, it was observed that several MTAs were located to the positions where Pm resistance genes/QTLs had been reported before while others represent novel loci. For instance, on chromosome 1A, two MTAs at positions 27.0 and 74.0 cM (detected during 2020-21), were found to share similar locations with previously identified Pm resistance loci such as resistance genes Pm3 [48] and Pm223389 [49], QTLs Qpm.osu.1A [50], Qpm.mgb-1AS [20] and QPm.Caas-1A [51], SNP markers SNP_BS00021714_51 [46], SNP_tplb0041a22_935, SNP_Excalibur_c15098_59116 [17] and five other MTAs identified by Liu et al. [12], and a meta-QTL MQTL1 [29].
Similarly, on chromosome 1B, an MTA was detected at 76.0 cM during 2022-23 which shared the chromosomal region with previously identified loci Qaprpm.cgb-1B [52] and QPm.osu-1B [50], as well as five MTAs identified by Liu et al. [12].
Our analysis revealed that five SNPs detected between 132.0 to 144.0 cM on chromosome 2B during 2020-21 and 2021-22 were found in close proximity to previously detected QTLs QPm.crag-2B and Qaprpm.cgb-2B, and major resistance genes such as a recessive gene pm42 and PmWE99 [10,53].
On chromosome 3B, we detected twelve PM resistance associated SNPs between 57.0 and 84.0 cM during 2021-22 and 2022-23. Comparison of the chromosome intervals with the previously identified loci such as PM_3B1 and PM_3B2 [16], QPm.inra-3B [54], QPm.osu-3B [50], QPm.caas-3BS [55], CP2 [29], Qpm.mgb-3BL.3 [20], as well as other MTAs identified by Liu et al. [12] and Alemu et al. [46], showed that markers identified in our study possibly shared chromosomal intervals with QPm.inra-3B [54], QPm.osu-3B [50], QPm.caas-3BS [55] and CP2 [29].
On chromosome 4A, an MTA was detected at 48.0 cM during 2021-22. This MTA seems to share same chromosome region as previously identified QTLs QPm.inra-4A [56] and Qpm.mgb-4AL [20] and a meta-QTL MQTL11 [29].
The presence of two MTAs at 68.0 cM on chromosome 5B (detected during 2021-22) seems associated with previously reported loci QPm.nuls-5B and QPm.umb-5BS [27], and a major gene Pm16 [57], highlights its potential for marker-assisted selection in breeding programs targeting PM resistance.
Among the chromosomes examined, chromosome 6B stood out with the largest number of MTAs, totaling 57, one at 0.0 cM while remaining within the 59.0 to 65.0 cM region. PM resistance MTA at 0.0 cM seems to be associated with QPm.umb-6BS [27]. However no previously identified loci were found in second region, suggesting the presence of novel genes or regulatory elements associated with PM resistance in this region.
On chromosome 7A, six MTAs were identified at different positions (at 57.0, 64.0, 65.0, 73.0, 74.0, and 150.0 cM) during 2020-21 and 2022-23. It seems that MTAs at 57.0, 64.0, 65.0, 73.0, and 74.0 represent a previously repetitively detected locus associated with QTLs QPm.caas-7AS at 26.0 to 28.0 cM [58] and meta-QTL MQTL21 at 22.6 to 53.2 cM [29], while MTA at 150.0 seems associated with Pm1, the SNP marker RAC875_c37085_317 (at 152.8 cM) linked with QPm.icg.7A cM [17], and QTL Qpm.mgb-7AS at 111 cM or Qpm.mgb-7AL at 191.4 cM [20]. The second region, which appears to be associate with Pm1, present a valuable target for marker-assisted selection.
Chromosome 7B exhibited four MTAs, with one MTA detected at 73.0 cM during 2022-23 and the other three detected at 136.0 cM during 2021-22. Several major QTLs such as Qpm.mgb-7BS1, Qpm.mgb-7BS.2, Qpm.mgb-7BL [20], and major PM resistance genes such as PmE [59], Pm47 [60] and Pm40 [61] have been reported on in vicinity of the MTA detected at 73.0 cM on chromosome 7B. It seems markers detected at 136.0 cM represent a novel PM resistance locus since no resistance loci has been observed in this region previously.
In addition to the above-discussed MTAs, a number of other novel MTAs were detected on chromosomes 3A (at 158.0 cM, detected during 2022-23), 4B (at 66.0 cM, detected during 2022-23), and 5A (at 144.0 cM, detected during 2021-22). No QTLs/MTAs or genes were previously reported by other studies in their vicinity. These findings emphasize the importance of further investigations focused on these and other novel MTA regions such as those on chromosome 6B (between 59.0 to 65.0 cM, detected during 2020-21 and 2021-22) for a deeper understanding of the genetic mechanisms underlying PM resistance.
The co-localization of MTAs with previously identified genetic markers, MTAs, QTLs and genes provides further evidence for their involvement in the resistance mechanism against PM. Identified novel MTAs could also be valuable for diversifying the sources of genetic resistance in breeding programs and provide valuable targets to enhance PM resistance by combining with repetitively detected loci using the associated SNP markers identified in this study. Moreover, these findings provide valuable information for breeders to prioritize genomic regions and genetic markers associated with PM resistance and contribute to a better understanding of the genetic basis of PM resistance in common wheat.
3.2. Candidate Genes for PM Resistance
To identify potential target genes for breeding, GWAS is often used as a starting point [62]. However, a challenge arises when significant markers associated with the trait of interest encompass a wide range of genes within their confidence intervals, making it difficult to pinpoint the exact causal genes [36]. Thus, in this study, we addressed this issue by focusing on MTAs found associated with genomic intervals carrying disease resistance associated candidate genes. Identified CGs (30 out of the 96 detected) broadly represent twelve plant protein families that exhibited clear connections to disease resistance and host-pathogen interactions. These proteins have been previously reported to impart disease resistance to various pathogens such as Blumeria graminis f. sp. Tritici, Fusarium graminearum, Puccinia striiformis, Bipolaris sorokiniana, Puccinia triticina, Parastagonospora nodorum and Pyrenophora tritici-repentis in plants [63,64,65,66,67,68,69,70,71,72].
It may be recalled that the 30 CGs encode proteins with distinctive domains, such as the ABC transporter-like domain, ATP-binding domain, ankyrin repeat, aspartic peptidase A1 family, peroxidase, cytochrome P450, disease resistance protein (NB-LRR), Glutathione S-transferase, NB-ARC P-loop containing nucleoside triphosphate hydrolase, kinase-like domain superfamily, wall-associated receptor kinase, galacturonan-binding domain, zinc finger proteins, and F-box superfamily domain (Table S2), which are widely recognized as essential components of disease resistance mechanisms. These results reinforce findings from previous research. For instance, Peng and Yang's [73] comprehensive analysis delved into ABC, NLR, and START genes in hexaploid wheat, revealing their co-localization with disease resistance quantitative trait loci (QTLs) associated with adult leaf rust resistance. Molecular characterization of the leaf rust-resistance gene Lr34, which encodes an ABC transporter with transmembrane (TM) and nucleotide binding site (NB) domains, was elucidated in studies by Krattinger et al. [74,75]. Similarly, Lr14a, a race-specific leaf rust resistance gene, was found to encode an ANKTM protein [76]. The YrU1 protein, conferring wheat stripe rust resistance, also featured an integrated ANK domain from ANKTM proteins, suggesting their role in resistance [76,77]. The up-regulation of A1 aspartic peptidase and G1 families during Zymoseptoria tritici infection [65] underscores their defensive roles. Similarly, peroxidases' positive contribution against Puccinia striiformis infection [78] highlights their significance in immunity. Disease sensitivity genes like Tsn1 which encode a protein with S/TPK and NBS-LRR domains [64,67] reveal their significance in recognizing necrotrophic effectors. The critical role of the NB-ARC domain in regulating R protein activity [63] deepens our understanding of pathogen recognition. Wall-associated receptor kinases (WAKs), exemplified by Stb6 [66] and TaWAK6 [69], emerge as potent defenders against STB and leaf rust. During defense, identified proteins exhibit versatile functions [68,72], encompassing stress resilience, apoptosis, transcription, and interactions. The F-box family protein, known for its role in numerous biological processes, including biotic stress resistance [79], highlights the multifaceted engagement of these proteins in wheat's resistance responses.
Despite identification of candidate genes and their association with disease resistance, no candidate gene were found in genomic intervals of 48 MTAs which may potentially carry other novel structural variants responsible for regulating plant defense against Bgt and wheat-Bgt interaction; however, further investigations are required to determine the significance and potential functional relevance of these specific SNPs in Bgt resistance.
In conclusion, our findings unveiled the intricate genetic landscape of PM resistance within the WAMI panel. This collection showcases a valuable pool of significant PM resistance MTAs/genes embedded in elite genetic backgrounds. Finally, through a comparison with previous studies on disease resistance in wheat, we have validated the functions of key MTAs.
4. Materials and Methods
4.1. Plant Material
WAMI panel consisting of 286 genetically diverse and elite advanced wheat lines was utilized for GWAS of PM resistance. The panel was assembled and distributed through the International Wheat Improvement Network (IWIN) the by International Maize and Wheat Improvement Center (CIMMYT) and possesses a narrow range of variation for days to heading and plant height which is appropriate for gene discovery without the confounding effects of phenology and plant height [47]. The seed of WAMI panel was obtained from CIMMYT. WL711, a highly PM susceptible common wheat cultivar [80], was used as a susceptible check and disease spreader.
4.2. Experimental Design and Trait Evaluation
The field evaluations of genotypes were conducted at the research farm of Eternal University, Baru Sahib, Himachal Pradesh, India, spanning three consecutive growing seasons (2020-21, 2021-2022, and 2022-23). The Baru Sahib PM disease nursery research farm in situated in a north Indian hilly state, Himachal Pradesh, and is known for climatic conditions favorable for the natural infection, growth, and the development of PM [82].
The experiments adhered to a randomized block design and were replicated twice. Random assignment of lines to each replication was carried out using the Fisher and Yates Random Table method [81]. Fertilizers included 120 kg N, 60 kg P2O5, and 40 kg K2O. Except N, fertilizers were thoroughly applied at the time of sowing. The application of nitrogen was divided into three doses: half at sowing, one-fourth at the first irrigation (21 days after sowing), and the remaining one-fourth at the second irrigation (45 days after sowing).
The genotypes under investigation were exposed to natural infections of PM. To enhance disease stress, two rows of WL711, a highly PM susceptible cultivar, were planted around the field, with an additional row seeded between the plots one month before sowing the WAMI panel in November each year. The WL711 was also sown alongside the experimental materials during main season sowing. Planting distances were maintained at 20 cm between rows and 5 cm between plants.
Disease severity was scored using a rating scale of 1 to 9 developed by Bennett and Westcott [83] on randomly chosen five plants from each plot. Once maximum expression of PM (when reached a score of 9) observed on susceptible check cultivar WL711, whole panel was rated following the established scoring criteria [83]. This approach enabled the tracking of disease progression over time and the identification of an appropriate growth stage exhibiting varying degrees of resistance or susceptibility to PM.
4.3. DNA Extraction and Genotyping
DNA extraction, genotyping of samples and data processing was performed as described previously [84]. Briefly, DNA was extracted from fresh leaves of each line using the CTAB procedure, as outlined by Saghai-maroof et al. [85]. Subsequently, genotyping was conducted at the USDA-ARS Small Grain Genotyping Center, Fargo (http://wheat.pw.usda.gov/GenotypingLabs), utilizing the Illumina iSelect 90K beadchip assay [43]. The SNP calling process utilized the default clustering algorithm integrated into Genome Studio v2011.1, resulting in the identification of a total of 26,814 bi-allelic SNPs [84,86]. To uphold data quality standards, SNPs characterized by a minor allele frequency (MAF) lower than 0.05 were omitted from the analysis, alongside monomorphic and low-quality SNPs. This meticulous filtration procedure led to the retention of approximately 21,132 polymorphic SNPs [84,87], which were subsequently harnessed for the purpose of conducting the GWAS in this study.
4.4. Descriptive Statistics Analyses
ANOVA and correlation coefficients analyses were performed using the Agricolae (version 1.2–4) package of the R (version 4.0.3) software [88]. The BLUP values were obtained using the 'lme4' package [89] in R [88]. Descriptive statistics, such as mean, standard deviation, and coefficient of variation (CV), were calculated using SPSS v. 17.0 (SPSS Inc 2008).
4.5. Population Structure, Kinship Matrix and Principal Components Analyses
Population structure (population structure matrix or Q matrix) and relatedness matrix (Kinship or K matrix) were modeled using principal components analysis (PCA) utilizing the genotypic data belonging to a total of 21,132 high-quality SNPs as described earlier [90]. The optimal numbers of PCAs were determined using the Bayesian information criterion (BIC) [91]. The analysis was performed with R (version 4.0.3) software [84,88] employing the parameters suggested by VanRaden [92] and Yin et al. [93]. The first two principal components were used to create a scatter plot that visualized the distribution of genotypes into sub-groups.
4.6. Linkage Disequilibrium and Genome-Wide Association Analyses
Linkage disequilibrium (LD) and genome-wide association analyses were conducted using a dataset of 21,132 high-quality single nucleotide polymorphisms (SNPs) available from the CIMMYT, Mexico website [84]. Pairwise squared allele-frequency correlations (r^2) between SNP markers were calculated using the TASSEL (Trait Analysis by Association, Evolution, and Linkage) software with a sliding window size of 100. To assess LD between loci, r^2 values were plotted against physical distance in centimorgans (cM). The LD decay curve was fitted using a smoothing spline regression line at the genome level, following the method outlined by Hill and Weir [45] and implemented in the R environment with a script previously employed by Marroni et al. [94].
Single- and multilocus models including the general linear model (GLM) [90], mixed linear model (MLM) [95], and the fixed and random model circulating probability unification (FarmCPU) [96] method were fitted to identify marker-trait associations (MTAs). Among these, FarmCPU was selected based on the best fit and used for the final GWAS. In FarmCPU, both fixed and random effects are incorporated to improve the accuracy of association mapping. It is a two-step procedure where the fixed effect model is initially fitted to control for population structure, and then the random effect model is used to further account for relatedness among individuals. The rMVP (R based Memory-efficient, Visualization-enhanced, and Parallel-accelerated Tool For Genome-Wide Association Study) software package [93] was employed for the analysis.
4.7. Identification of Candidate Genes
To identify potential candidate genes (CGs), we focused on the most significant MTAs. The marker sequences associated with these MTAs were aligned with the wheat genome assembly IWGSC v.1 downloaded from the Ensembl database. Specifically, we analyzed a 200 kb window around each MTA marker to extract highly significant and annotated CGs. For the gene ontology (GO) annotation of these CGs, we utilized IWGSC (http://www.wheatgenome.org), which offered the requisite information for annotating the GO terms associated with the identified candidate genes.
5. Conclusions
Incorporating the insights gleaned from the three-year investigation, a focused set of significant MTAs emerged, each holding the potential to drive advancements in resistance to PM disease in wheat. Notably, recurrent novel MTAs, consistently observed on chromosome 6B across 2020-21 and 2021-22 (Table 3), underscore the enduring influence of key genomic regions on PM resistance. By harnessing the knowledge embedded within these MTAs, breeders can refine their selection processes, develop tailored markers, and ultimately expedite the development of superior PM-resistant cultivars through precision-guided approaches. This study not only sheds light on the genetic architecture underpinning PM resistance but also paves the way for innovative and accelerated wheat breeding programs that align with the demands of sustainable and resilient agricultural practices. Moreover, we identified potential candidate genes involved in disease resistance mechanisms in wheat by analyzing significant SNPs and their associated gene sequences.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Supplementary File 1.xlsx. Table S1: Details of candidate genes identified in genomic intervals of marker trait associations detected during experimental years 2020-21, 2021-22 and 2022-23.; Table S2: Details of defense and host-pathogen associated proteins encoded by candidate genes identified in genomic intervals of marker trait associations detected during this study.
Author Contributions
Conceptualization, N.K.V.; methodology, R.K. and N.K.V.; formal analysis, R.K.; investigation, R.K.; resources, N.K.V, V.K.M., S.S., A.K.J. and M.T.; data curation, R.K., V.K.R. and N.K.V.; writing—original draft preparation, R.K. and N.K.V.; writing—review and editing, R.D., A.K.J. and N.K.V; visualization, R.D. and N.K.V; supervision, N.K.V.; project administration, N.K.V.; funding acquisition, N.K.V; intellectual input, R.D. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Science and Engineering Research Board, New Delhi, grant number SRG/2020/000091.
Data Availability Statement
The single nucleotide polymorphism (SNP) genotyping data of the Wheat Association Mapping Initiative (WAMI) germplasm panel of spring wheat, which is used for the present study, has been published previously by Sukumaran et al. [84], and is available publicly to download from the link: http://hdl.handle.net/11529/10714. All other data generated or analyzed during this study are included in this published article.
Acknowledgments
We sincerely acknowledge Matthew P. Reynolds of CIMMYT, Mexico, for generously providing the WAMI population and granting access to the molecular data used in this study. Additionally, we extend our gratitude to the Department of Genetics, Plant Breeding, and Biotechnology, Dr. K. S. Gill Akal College of Agriculture, Eternal University, Baru Sahib, HP, India, for their valuable support and provision of facilities to NKV and RK throughout the course of this research.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Grote, U.; Fasse, A.; Nguyen, T.T.; Erenstein, O. Food Security and the Dynamics of Wheat and Maize Value Chains in Africa and Asia. Frontiers in Sustainable Food Systems 2021, 4. [CrossRef]
- Shahbandeh, M. Total wheat consumption worldwide 2022/23. Available online: https://www.statista.com/statistics/1094056/total-global-rice-consumption/ (accessed on August 8).
- Alam, M.A., Mandal, M.S.N., Wang, C. and Wanquan, J. Chromosomal location and SSR markers of a powdery mildew resistance gene in common wheat line N0308. Afr. J. Microbiol Res. 2013, 7, 477-482.
- Mwale, V.M., Chilembwe H.C., Uluko H.C. Wheat powdery mildew (Blumeria graminis f. sp. tritici): Damage effects and genetic resistance developed in wheat (Triticum aestivum). Journal of Plant Science 2014, 5, 1-16.
- Te Beest, D.E.; Paveley, N.D.; Shaw, M.W.; van den Bosch, F. Disease-weather relationships for powdery mildew and yellow rust on winter wheat. Phytopathology 2008, 98, 609-617. [CrossRef]
- Mehta, A.B.A.K.B.D.K.B.D. Effect of weather parameters on powdery mildew development of wheat at different location in Himachal Pradesh. Indian Phytopathology 2018, 71, 349-353. [CrossRef]
- Hartl, L.; Weiss, H.; Stephan, U.; Zeller, F.J.; Jahoor, A. Molecular identification of powdery mildew resistance genes in common wheat (Triticum aestivum L.). Theoretical and Applied Genetics 1995, 90, 601-606. [CrossRef]
- Keller, M.; Keller, B.; Schachermayr, G.; Winzeler, M.; Schmid, J.E.; Stamp, P.; Messmer, M.M. Quantitative trait loci for resistance against powdery mildew in a segregating wheat×spelt population. Theoretical and Applied Genetics 1999, 98, 903-912. [CrossRef]
- Hysing, S.-C.; Merker, A.; Liljeroth, E.; Koebner, R.M.D.; Zeller, F.J.; Hsam, S.L.K. Powdery mildew resistance in 155 Nordic bread wheat cultivars and landraces. Hereditas 2007, 144, 102-119. [CrossRef]
- Hua, W.; Liu, Z.; Zhu, J.; Xie, C.; Yang, T.; Zhou, Y.; Duan, X.; Sun, Q.; Liu, Z. Identification and genetic mapping of pm42, a new recessive wheat powdery mildew resistance gene derived from wild emmer (Triticum turgidum var. dicoccoides). Theoretical and Applied Genetics 2009, 119, 223-230. [CrossRef]
- Hao, Y.; Parks, R.; Cowger, C.; Chen, Z.; Wang, Y.; Bland, D.; Murphy, J.P.; Guedira, M.; Brown-Guedira, G.; Johnson, J. Molecular characterization of a new powdery mildew resistance gene Pm54 in soft red winter wheat. Theoretical and Applied Genetics 2015, 128, 465-476. [CrossRef]
- Liu, N.; Bai, G.; Lin, M.; Xu, X.; Zheng, W. Genome-wide Association Analysis of Powdery Mildew Resistance in U.S. Winter Wheat. Scientific Reports 2017, 7, 11743. [CrossRef]
- Li, G.; Cowger, C.; Wang, X.; Carver, B.F.; Xu, X. Characterization of Pm65, a new powdery mildew resistance gene on chromosome 2AL of a facultative wheat cultivar. Theoretical and Applied Genetics 2019, 132, 2625-2632. [CrossRef]
- Li, G.; Xu, X.; Tan, C.; Carver, B.F.; Bai, G.; Wang, X.; Bonman, J.M.; Wu, Y.; Hunger, R.; Cowger, C. Identification of powdery mildew resistance loci in wheat by integrating genome-wide association study (GWAS) and linkage mapping. The Crop Journal 2019, 7, 294-306. [CrossRef]
- Jia, M.; Xu, H.; Liu, C.; Mao, R.; Li, H.; Liu, J.; Du, W.; Wang, W.; Zhang, X.; Han, R.; et al. Characterization of the Powdery Mildew Resistance Gene in the Elite Wheat Cultivar Jimai 23 and Its Application in Marker-Assisted Selection. Frontiers in Genetics 2020, 11. [CrossRef]
- Kang, Y.; Barry, K.; Cao, F.; Zhou, M. Genome-wide association mapping for adult resistance to powdery mildew in common wheat. Molecular Biology Reports 2020, 47, 1241-1256. [CrossRef]
- Leonova, I.N. Genome-Wide Association Study of Powdery Mildew Resistance in Russian Spring Wheat (T. aestivum L.) Varieties. Russian Journal of Genetics 2019, 55, 1360-1374. [CrossRef]
- McIntosh R. A., D.J., Rogers W. J., Xia X. C., Raupp W. J. Catalogue of gene symbols for wheat: 2020 supplement. In Proceedings of the Annual Wheat Newsletter, Manhattan, USA, 2020; pp. 98–113.
- McIntosh, R.A.; Hart, G.E.; Devos, K.M.; Gale, M.D.; Rogers, W.J. Catalogue of Gene Symbols for Wheat. 1998. https://wheat.pw.usda.gov/ggpages/wgc/98/.
- Simeone, R.; Piarulli, L.; Nigro, D.; Signorile, M.A.; Blanco, E.; Mangini, G.; Blanco, A. Mapping Powdery Mildew (Blumeria graminis f. sp. tritici) Resistance in Wild and Cultivated Tetraploid Wheats. Int J Mol Sci 2020, 21. [CrossRef]
- Pang, Y.; Wu, Y.; Liu, C.; Li, W.; St. Amand, P.; Bernardo, A.; Wang, D.; Dong, L.; Yuan, X.; Zhang, H.; et al. High-resolution genome-wide association study and genomic prediction for disease resistance and cold tolerance in wheat. Theoretical and Applied Genetics 2021, 134, 2857-2873. [CrossRef]
- Du, X.; Xu, W.; Peng, C.; Li, C.; Zhang, Y.; Hu, L. Identification and validation of a novel locus, Qpm-3BL, for adult plant resistance to powdery mildew in wheat using multilocus GWAS. BMC Plant Biology 2021, 21, 357. [CrossRef]
- He, H.; Liu, R.; Ma, P.; Du, H.; Zhang, H.; Wu, Q.; Yang, L.; Gong, S.; Liu, T.; Huo, N.; et al. Characterization of Pm68, a new powdery mildew resistance gene on chromosome 2BS of Greek durum wheat TRI 1796. Theoretical and Applied Genetics 2021, 134, 53-62. [CrossRef]
- Hinterberger, V.; Douchkov, D.; Lück, S.; Kale, S.; Mascher, M.; Stein, N.; Reif, J.C.; Schulthess, A.W. Mining for New Sources of Resistance to Powdery Mildew in Genetic Resources of Winter Wheat. Frontiers in Plant Science 2022, 13. [CrossRef]
- Jin, P.; Guo, X.; Guo, M.; Li, R.; Li, Q.; Cheng, P.; Wang, B. Genome-Wide Association Mapping of Resistance to Powdery Mildew in Regional Trials of Wheat Mainly from China. Plant Dis 2022, 106, 2701-2710. [CrossRef]
- Spielmeyer, W.; McIntosh, R.A.; Kolmer, J.; Lagudah, E.S. Powdery mildew resistance and Lr34/Yr18 genes for durable resistance to leaf and stripe rust cosegregate at a locus on the short arm of chromosome 7D of wheat. Theor Appl Genet 2005, 111, 731-735. [CrossRef]
- Lillemo, M.; Asalf, B.; Singh, R.P.; Huerta-Espino, J.; Chen, X.M.; He, Z.H.; Bjørnstad, Å. The adult plant rust resistance loci Lr34/Yr18 and Lr46/Yr29 are important determinants of partial resistance to powdery mildew in bread wheat line Saar. Theoretical and Applied Genetics 2008, 116, 1155-1166. [CrossRef]
- Herrera-Foessel, S.A.; Singh, R.P.; Lillemo, M.; Huerta-Espino, J.; Bhavani, S.; Singh, S.; Lan, C.; Calvo-Salazar, V.; Lagudah, E.S. Lr67/Yr46 confers adult plant resistance to stem rust and powdery mildew in wheat. Theor Appl Genet 2014, 127, 781-789. [CrossRef]
- Marone, D.; Russo, M.A.; Laidò, G.; De Vita, P.; Papa, R.; Blanco, A.; Gadaleta, A.; Rubiales, D.; Mastrangelo, A.M. Genetic basis of qualitative and quantitative resistance to powdery mildew in wheat: from consensus regions to candidate genes. BMC Genomics 2013, 14, 562. [CrossRef]
- Lee, J.H.; Graybosch, R.A.; Peterson, C.J. Quality and biochemical effects of a IBL/IRS wheat-rye translocation in wheat. Theoretical and Applied Genetics 1995, 90, 105-112. [CrossRef]
- Zeng, F.-s.; Yang, L.-j.; Gong, S.-j.; Shi, W.-q.; Zhang, X.-j.; Wang, H.; Xiang, L.-b.; Xue, M.-F.; Yu, D.-z. Virulence and Diversity of Blumeria graminis f. sp. tritici Populations in China. Journal of Integrative Agriculture 2014, 13, 2424-2437. [CrossRef]
- Bapela, T.; Shimelis, H.; Terefe, T.; Bourras, S.; Sánchez-Martín, J.; Douchkov, D.; Desiderio, F.; Tsilo, T.J. Breeding Wheat for Powdery Mildew Resistance: Genetic Resources and Methodologies—A Review. Agronomy 2023, 13. [CrossRef]
- Dhariwal, R.; Randhawa, H.S. Mapping Quantitative Trait Loci in Wheat: Historic Perspective, Tools, and Methods for Analysis. In Accelerated Breeding of Cereal Crops, Bilichak, A., Laurie, J.D., Eds.; Springer Protocols Handbooks; Springer US: New York, NY, 2022; pp. 31-75.
- Bartoli, C.; Roux, F. Genome-Wide Association Studies In Plant Pathosystems: Toward an Ecological Genomics Approach. Frontiers in Plant Science 2017, 8. [CrossRef]
- Vagndorf, N.; Nielsen, N.H.; Edriss, V.; Andersen, J.R.; Orabi, J.; Jørgensen, L.N.; Jahoor, A. Genomewide association study reveals novel quantitative trait loci associated with resistance towards Septoria tritici blotch in North European winter wheat. Plant Breeding 2017, 136, 474-482. [CrossRef]
- Juliana, P.; Singh, R.P.; Poland, J.; Mondal, S.; Crossa, J.; Montesinos-López, O.A.; Dreisigacker, S.; Pérez-Rodríguez, P.; Huerta-Espino, J.; Crespo-Herrera, L.; et al. Prospects and Challenges of Applied Genomic Selection—A New Paradigm in Breeding for Grain Yield in Bread Wheat. The Plant Genome 2018, 11, 180017. [CrossRef]
- Muqaddasi, Q.H.; Zhao, Y.; Rodemann, B.; Plieske, J.; Ganal, M.W.; Röder, M.S. Genome-wide Association Mapping and Prediction of Adult Stage Septoria tritici Blotch Infection in European Winter Wheat via High-Density Marker Arrays. The Plant Genome 2019, 12, 180029. [CrossRef]
- Stadlmeier, M.; Jørgensen, L.N.; Corsi, B.; Cockram, J.; Hartl, L.; Mohler, V. Genetic Dissection of Resistance to the Three Fungal Plant Pathogens Blumeria graminis, Zymoseptoria tritici, and Pyrenophora tritici-repentis Using a Multiparental Winter Wheat Population. G3 Genes|Genomes|Genetics 2019, 9, 1745-1757. [CrossRef]
- Huang, B.E.; Verbyla, K.L.; Verbyla, A.P.; Raghavan, C.; Singh, V.K.; Gaur, P.; Leung, H.; Varshney, R.K.; Cavanagh, C.R. MAGIC populations in crops: current status and future prospects. Theor Appl Genet 2015, 128, 999-1017. [CrossRef]
- Buckler, E.S.t.; Thornsberry, J.M. Plant molecular diversity and applications to genomics. Curr Opin Plant Biol 2002, 5, 107-111. [CrossRef]
- Zhu, C.; Gore, M.; Buckler, E.S.; Yu, J. Status and Prospects of Association Mapping in Plants. 2008, 1, 5-20. [CrossRef]
- Nelson, R.; Wiesner-Hanks, T.; Wisser, R.; Balint-Kurti, P. Navigating complexity to breed disease-resistant crops. Nat Rev Genet 2018, 19, 21-33. [CrossRef]
- Wang, S.; Wong, D.; Forrest, K.; Allen, A.; Chao, S.; Huang, B.E.; Maccaferri, M.; Salvi, S.; Milner, S.G.; Cattivelli, L.; et al. Characterization of polyploid wheat genomic diversity using a high-density 90 000 single nucleotide polymorphism array. Plant Biotechnology Journal 2014, 12, 787-796. [CrossRef]
- Bolser, D.; Staines, D.M.; Pritchard, E.; Kersey, P. Ensembl Plants: Integrating Tools for Visualizing, Mining, and Analyzing Plant Genomics Data. Methods in molecular biology (Clifton, N.J.) 2016, 1374, 115-140. [CrossRef]
- Hill, W.G.; Weir, B.S. Variances and covariances of squared linkage disequilibria in finite populations. Theor Popul Biol 1988, 33, 54-78. [CrossRef]
- Alemu, A.; Brazauskas, G.; Gaikpa, D.S.; Henriksson, T.; Islamov, B.; Jørgensen, L.N.; Koppel, M.; Koppel, R.; Liatukas, Ž.; Svensson, J.T.; et al. Genome-Wide Association Analysis and Genomic Prediction for Adult-Plant Resistance to Septoria Tritici Blotch and Powdery Mildew in Winter Wheat. Frontiers in Genetics 2021, 12. [CrossRef]
- Lopes, M.S.; Dreisigacker, S.; Peña, R.J.; Sukumaran, S.; Reynolds, M.P. Genetic characterization of the wheat association mapping initiative (WAMI) panel for dissection of complex traits in spring wheat. Theoretical and Applied Genetics 2015, 128, 453-464. [CrossRef]
- Zeller, F.J.; Lutz, J.; Stephan, U. Chromosome location of genes for resistance to powdery mildew in common wheat (Triticum aestivum L.) 1. Mlk and other alleles at the Pm3 locus. Euphytica 1993, 68, 223-229. [CrossRef]
- Li, G.; Carver, B.F.; Cowger, C.; Bai, G.; Xu, X. Pm223899, a new recessive powdery mildew resistance gene identified in Afghanistan landrace PI 223899. Theor Appl Genet 2018, 131, 2775-2783. [CrossRef]
- Chen, Y.; Hunger, R.M.; Carver, B.F.; Zhang, H.; Yan, L. Genetic characterization of powdery mildew resistance in U.S. hard winter wheat. Mol Breeding 2009, 24, 141-152. [CrossRef]
- Liang, S.S.; Suenaga, K.; He, Z.H.; Wang, Z.L.; Liu, H.Y.; Wang, D.S.; Singh, R.P.; Sourdille, P.; Xia, X.C. Quantitative trait Loci mapping for adult-plant resistance to powdery mildew in bread wheat. Phytopathology 2006, 96, 784-789. [CrossRef]
- Huang, Q.; Ruilian, J.; Xinyuan, W. QTL mapping for adult-plant resistance to powdery mildew in common wheat. Scientia Agricultura Sinica 2008.
- Ma, P.; Xu, H.; Han, G.; Luo, Q.; Xu, Y.; Zhang, X.; An, D.; Li, L.; Sun, Y. Characterization of a Segregation Distortion Locus with Powdery Mildew Resistance in a Wheat-Thinopyrum intermedium Introgression Line WE99. Plant Dis 2016, 100, 1541-1547. [CrossRef]
- Bougot, Y.; Lemoine, J.; Pavoine, M.T.; Guyomar'ch, H.; Gautier, V.; Muranty, H.; Barloy, D. A major QTL effect controlling resistance to powdery mildew in winter wheat at the adult plant stage. Plant Breeding 2006, 125, 550-556. [CrossRef]
- Jia, A.; Ren, Y.; Gao, F.; Yin, G.; Liu, J.; Guo, L.; Zheng, J.; He, Z.; Xia, X. Mapping and validation of a new QTL for adult-plant resistance to powdery mildew in Chinese elite bread wheat line Zhou8425B. Theoretical and Applied Genetics 2018, 131, 1063-1071. [CrossRef]
- Xu, H.; Yao, G.; Xiong, L.; Yang, L.; Jiang, Y.; Fu, B.; Zhao, W.; Zhang, Z.; Zhang, C.; Ma, Z. Identification and mapping of pm2026: a recessive powdery mildew resistance gene in an einkorn (Triticum monococcum L.) accession. Theor Appl Genet 2008, 117, 471-477. [CrossRef]
- Chen, X.M.; Luo, Y.H.; Xia, X.C.; Xia, L.Q.; Chen, X.; Ren, Z.L.; He, Z.H.; Jia, J.Z. Chromosomal location of powdery mildew resistance gene Pm16 in wheat using SSR marker analysis. Plant Breeding 2005, 124, 225-228. [CrossRef]
- Lan, C.; Liang, S.; Wang, Z.; Yan, J.; Zhang, Y.; Xia, X.; He, Z. Quantitative Trait Loci Mapping for Adult-Plant Resistance to Powdery Mildew in Chinese Wheat Cultivar Bainong 64. Phytopathology® 2009, 99, 1121-1126. [CrossRef]
- Ma, Q.; Luo, P.; Ren, Z.; Jiang, H.; Yang, Z. Genetic analysis and chromosomal location of two new genes for resistance to powdery mildew in wheat (Triticum aestivum L.). Acta Agronomica Sinica 2010, 33, 1-8.
- Xiao, M.; Song, F.; Jiao, J.; Wang, X.; Xu, H.; Li, H. Identification of the gene Pm47 on chromosome 7BS conferring resistance to powdery mildew in the Chinese wheat landrace Hongyanglazi. Theor Appl Genet 2013, 126, 1397-1403. [CrossRef]
- Zhong, S.; Ma, L.; Fatima, S.A.; Yang, J.; Chen, W.; Liu, T.; Hu, Y.; Li, Q.; Guo, J.; Zhang, M.; et al. Collinearity Analysis and High-Density Genetic Mapping of the Wheat Powdery Mildew Resistance Gene Pm40 in PI 672538. PLOS ONE 2016, 11, e0164815. [CrossRef]
- Dellaporta, S.L.W., J.; Hicks, J.B. Isolation of DNA from higher plants. Plant Mol Biol Rep 1983, 4, 19–21.
- van Ooijen, G.; Mayr, G.; Kasiem, M.M.; Albrecht, M.; Cornelissen, B.J.; Takken, F.L. Structure-function analysis of the NB-ARC domain of plant disease resistance proteins. J Exp Bot 2008, 59, 1383-1397. [CrossRef]
- Faris, J.D.; Zhang, Z.; Lu, H.; Lu, S.; Reddy, L.; Cloutier, S.; Fellers, J.P.; Meinhardt, S.W.; Rasmussen, J.B.; Xu, S.S.; et al. A unique wheat disease resistance-like gene governs effector-triggered susceptibility to necrotrophic pathogens. Proceedings of the National Academy of Sciences 2010, 107, 13544-13549. [CrossRef]
- Krishnan, P.; Ma, X.; McDonald, B.A.; Brunner, P.C. Widespread signatures of selection for secreted peptidases in a fungal plant pathogen. BMC Evolutionary Biology 2018, 18, 7. [CrossRef]
- Saintenac, C.; Lee, W.-S.; Cambon, F.; Rudd, J.J.; King, R.C.; Marande, W.; Powers, S.J.; Bergès, H.; Phillips, A.L.; Uauy, C.; et al. Wheat receptor-kinase-like protein Stb6 controls gene-for-gene resistance to fungal pathogen Zymoseptoria tritici. Nature Genetics 2018, 50, 368-374. [CrossRef]
- Gupta, P.K.; Chand, R.; Vasistha, N.K.; Pandey, S.P.; Kumar, U.; Mishra, V.K.; Joshi, A.K. Spot blotch disease of wheat: the current status of research on genetics and breeding. Plant Pathology 2018, 67, 508-531. [CrossRef]
- Noman, A.; Aqeel, M.; Khalid, N.; Islam, W.; Sanaullah, T.; Anwar, M.; Khan, S.; Ye, W.; Lou, Y. Zinc finger protein transcription factors: Integrated line of action for plant antimicrobial activity. Microbial Pathogenesis 2019, 132, 141-149. [CrossRef]
- Dmochowska-Boguta, M.; Kloc, Y.; Zielezinski, A.; Werecki, P.; Nadolska-Orczyk, A.; Karlowski, W.M.; Orczyk, W. TaWAK6 encoding wall-associated kinase is involved in wheat resistance to leaf rust similar to adult plant resistance. PLOS ONE 2020, 15, e0227713. [CrossRef]
- Yates, S.; Mikaberidze, A.; Krattinger, S.G.; Abrouk, M.; Hund, A.; Yu, K.; Studer, B.; Fouche, S.; Meile, L.; Pereira, D.; et al. Precision Phenotyping Reveals Novel Loci for Quantitative Resistance to Septoria Tritici Blotch. Plant Phenomics 2019. [CrossRef]
- Mahmoudi, Z.; Taliei, F.; Ahangar, L.; Kheyrgoo, M. Assessment of salicylic acid-induced resistance against Septoria tritici blotch disease on wheat using real-time PCR. mdrsjrns 2021, 10, 151-165.
- Lin, P.-C.; Pomeranz, M.C.; Jikumaru, Y.; Kang, S.G.; Hah, C.; Fujioka, S.; Kamiya, Y.; Jang, J.-C. The Arabidopsis tandem zinc finger protein AtTZF1 affects ABA- and GA-mediated growth, stress and gene expression responses. The Plant Journal 2011, 65, 253-268. [CrossRef]
- Peng, F.Y.; Yang, R.C. Prediction and analysis of three gene families related to leaf rust (Puccinia triticina) resistance in wheat (Triticum aestivum L.). BMC Plant Biol 2017, 17, 108. [CrossRef]
- Krattinger, S.G.; Lagudah, E.S.; Spielmeyer, W.; Singh, R.P.; Huerta-Espino, J.; McFadden, H.; Bossolini, E.; Selter, L.L.; Keller, B. A putative ABC transporter confers durable resistance to multiple fungal pathogens in wheat. Science 2009, 323, 1360-1363. [CrossRef]
- Krattinger, S.G.; Lagudah, E.S.; Wicker, T.; Risk, J.M.; Ashton, A.R.; Selter, L.L.; Matsumoto, T.; Keller, B. Lr34 multi-pathogen resistance ABC transporter: molecular analysis of homoeologous and orthologous genes in hexaploid wheat and other grass species. The Plant journal : for cell and molecular biology 2011, 65, 392-403. [CrossRef]
- Kolodziej, M.C.; Singla, J.; Sánchez-Martín, J.; Zbinden, H.; Šimková, H.; Karafiátová, M.; Doležel, J.; Gronnier, J.; Poretti, M.; Glauser, G.; et al. A membrane-bound ankyrin repeat protein confers race-specific leaf rust disease resistance in wheat. Nature Communications 2021, 12, 956. [CrossRef]
- Wang, H.; Zou, S.; Li, Y.; Lin, F.; Tang, D. An ankyrin-repeat and WRKY-domain-containing immune receptor confers stripe rust resistance in wheat. Nature Communications 2020, 11, 1353. [CrossRef]
- Yang, Y.; Yu, Y.; Bi, C.; Kang, Z. Quantitative Proteomics Reveals the Defense Response of Wheat against Puccinia striiformis f. sp. tritici. Scientific Reports 2016, 6, 34261. [CrossRef]
- Kim, H.S.; Delaney, T.P. Arabidopsis SON1 Is an F-Box Protein That Regulates a Novel Induced Defense Response Independent of Both Salicylic Acid and Systemic Acquired Resistance. The Plant Cell 2002, 14, 1469-1482. [CrossRef]
- Sharma, M.; Kaur, S.; Saluja, M.; Chhuneja, P. Mapping and characterization of powdery mildew resistance gene in synthetic wheat. Czech Journal of Genetics and Plant Breeding 2016, 52, 120-123.
- Panse, V.G.S., P.V. . Statistical methods for Agricultural workers. 1985.
- Gautam, A.K. Studies on some powdery mildew of Himachal Pradesh, India. Australasian Mycologist 2015, 32, 10-13.
- Bennett, F.G.A.; Westcott, B. Field assessment of resistance to powdery mildew in mature wheat plants. Plant Pathology 1982, 31, 261-268. [CrossRef]
- Sukumaran, S.; Crossa, J.; Jarquín, D.; Lopes, M.; Reynolds, M.P. Genomic and pedigree prediction with genotype × environment interaction in spring wheat grown in South and Western Asia, North Africa, and Mexico. 2016, hdl:11529/10714. [CrossRef]
- Saghai-Maroof, M.A.; Soliman, K.M.; Jorgensen, R.A.; Allard, R.W. Ribosomal DNA spacer-length polymorphisms in barley: mendelian inheritance, chromosomal location, and population dynamics. Proceedings of the National Academy of Sciences 1984, 81, 8014-8018. [CrossRef]
- Sukumaran, S.; Lopes, M.; Dreisigacker, S.; Reynolds, M. Genetic analysis of multi-environmental spring wheat trials identifies genomic regions for locus-specific trade-offs for grain weight and grain number. Theoretical and Applied Genetics 2018, 131, 985-998. [CrossRef]
- Ahirwar, R.N.; Mishra, V.K.; Chand, R.; Budhlakoti, N.; Mishra, D.C.; Kumar, S.; Singh, S.; Joshi, A.K. Genome-wide association mapping of spot blotch resistance in wheat association mapping initiative (WAMI) panel of spring wheat (Triticum aestivum L.). PLOS ONE 2018, 13, e0208196. [CrossRef]
- R Core Team R: A language and environment for statistical computing., R Foundation for Statistical Computing: Vienna, Austria., 2013.
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. Journal of Statistical Software 2015, 67, 1-48.
- Price, A.L.; Patterson, N.J.; Plenge, R.M.; Weinblatt, M.E.; Shadick, N.A.; Reich, D. Principal components analysis corrects for stratification in genome-wide association studies. Nature Genetics 2006, 38, 904-909. [CrossRef]
- Schwarz, G. Estimating the Dimension of a Model. The Annals of Statistics 1978, 6, 461-464. [CrossRef]
- VanRaden, P.M. Efficient methods to compute genomic predictions. J Dairy Sci 2008, 91, 4414-4423. [CrossRef]
- Yin, L.; Zhang, H.; Tang, Z.; Xu, J.; Yin, D.; Zhang, Z.; Yuan, X.; Zhu, M.; Zhao, S.; Li, X.; et al. rMVP: A Memory-efficient, Visualization-enhanced, and Parallel-accelerated Tool for Genome-wide Association Study. Genomics, Proteomics & Bioinformatics 2021, 19, 619-628. [CrossRef]
- Marroni, F.; Pinosio, S.; Zaina, G.; Fogolari, F.; Felice, N.; Cattonaro, F.; Morgante, M. Nucleotide diversity and linkage disequilibrium in Populus nigra cinnamyl alcohol dehydrogenase (CAD4) gene. Tree Genetics & Genomes 2011, 7, 1011-1023. [CrossRef]
- Yu, J.; Pressoir, G.; Briggs, W.H.; Vroh Bi, I.; Yamasaki, M.; Doebley, J.F.; McMullen, M.D.; Gaut, B.S.; Nielsen, D.M.; Holland, J.B.; et al. A unified mixed-model method for association mapping that accounts for multiple levels of relatedness. Nature Genetics 2006, 38, 203-208. [CrossRef]
- Liu, X.; Huang, M.; Fan, B.; Buckler, E.S.; Zhang, Z. Iterative Usage of Fixed and Random Effect Models for Powerful and Efficient Genome-Wide Association Studies. PLOS Genetics 2016, 12, e1005767. [CrossRef]
- Maccaferri, M.; El-Feki, W.; Nazemi, G.; Salvi, S.; Canè, M.A.; Colalongo, M.C.; Stefanelli, S.; Tuberosa, R. Prioritizing quantitative trait loci for root system architecture in tetraploid wheat. J Exp Bot 2016, 67, 1161-1178. [CrossRef]
- Singh, S.; Gaurav, S.S.; Vasistha, N.K.; Kumar, U.; Joshi, A.K.; Mishra, V.K.; Chand, R.; Gupta, P.K. Genetics of spot blotch resistance in bread wheat (Triticum aestivum L.) using five models for GWAS. Frontiers in Plant Science 2023, 13. [CrossRef]
Figure 1.
Correlation coefficient, population structure and linkage disequilibrium (LD). A. A graphical representation of correlation coefficient among three environmental years for PM disease severity. B. Scatterplots showing the results of principal component analysis (PCA) conducted on genotypic data obtained from WAMI panel containing 286 wheat accessions. The figure highlights the population structure of the WAMI panel, as revealed by the first two principal components (PC1 and PC2). C. A scatter plot for r2 values of pairwise markers showing genome-wide linkage disequilibrium decay in 286 genotypes of the WAMI panel assessed using the Hill and Weir formula [45]. The red curved line depicts the fitted model for LD decay (non-linear regression of r2 against distance), while the blue horizontal line represents a threshold (0.12) for the QTLs. The vertical green line depicts a genetic distance confidence interval of 3 cM at which the LD half-decay (r2 = 0.23, the vertical green line) intersect with the LD decay curve.
Figure 1.
Correlation coefficient, population structure and linkage disequilibrium (LD). A. A graphical representation of correlation coefficient among three environmental years for PM disease severity. B. Scatterplots showing the results of principal component analysis (PCA) conducted on genotypic data obtained from WAMI panel containing 286 wheat accessions. The figure highlights the population structure of the WAMI panel, as revealed by the first two principal components (PC1 and PC2). C. A scatter plot for r2 values of pairwise markers showing genome-wide linkage disequilibrium decay in 286 genotypes of the WAMI panel assessed using the Hill and Weir formula [45]. The red curved line depicts the fitted model for LD decay (non-linear regression of r2 against distance), while the blue horizontal line represents a threshold (0.12) for the QTLs. The vertical green line depicts a genetic distance confidence interval of 3 cM at which the LD half-decay (r2 = 0.23, the vertical green line) intersect with the LD decay curve.
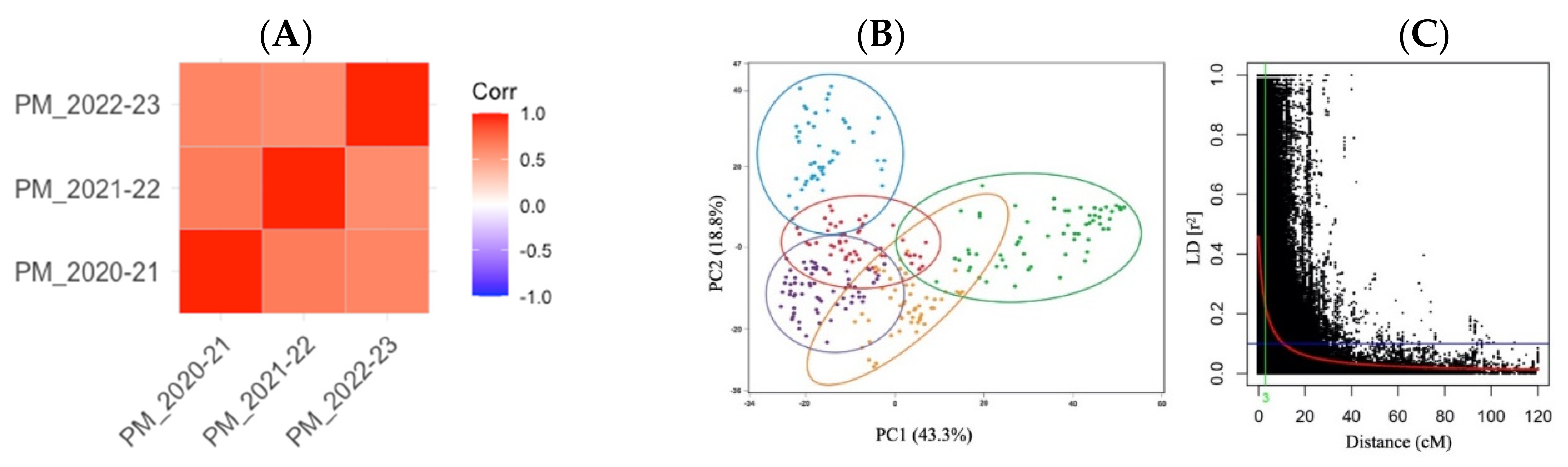
Table 1.
The descriptive statistics and heritability for the PM disease severity in wheat association mapping initiative (WAMI) panel containing 286 wheat accessions screened at Baru Sahib PM disease nurseries in Himachal Pradesh, India during three field seasons, spanning from 2020 to 2023.
Table 1.
The descriptive statistics and heritability for the PM disease severity in wheat association mapping initiative (WAMI) panel containing 286 wheat accessions screened at Baru Sahib PM disease nurseries in Himachal Pradesh, India during three field seasons, spanning from 2020 to 2023.
| Source | 2020-21 | 2021-22 | 2022-23 |
|---|---|---|---|
| Min | 1 | 2.5 | 1.5 |
| Max | 8.5 | 8.3 | 9 |
| Mean | 5.3 | 5.1 | 5.3 |
| LSD | 1.2 | 1.4 | 1.3 |
| CV | 8.4 | 12.2 | 9.0 |
| Heritability | 0.89 | 0.65 | 0.81 |
Min: minimum; Max: maximum; LSD: least significant difference; CV: coefficient of variation.
Table 2.
Analysis of variance (ANOVA) for PM disease severity in wheat association mapping initiative (WAMI) panel containing 286 wheat accessions.
Table 2.
Analysis of variance (ANOVA) for PM disease severity in wheat association mapping initiative (WAMI) panel containing 286 wheat accessions.
| Source of variation | Df | MSS |
|---|---|---|
| Genotypes | 285 | 4.9 ** |
| Environments (years) | 2 | 52.7 ** |
| Replications | 1 | 8.5 |
| Genotype × Environment | 570 | 1.22 ** |
Df: degrees of freedom; MSS: mean squares significant at **p ≤ 0.001.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



