
Submitted:
02 October 2023
Posted:
03 October 2023
You are already at the latest version
Abstract
The plant water status is crucial for growth and production, but the current climate change scenario makes it challenging to match the water plant demand. Blueberry is an economically important crop and plays an acknowledged role in human health due to its antioxidant compounds. This research aimed to determine whether exogenous methyl jasmonate application improves the antioxidant defense mechanisms for protecting the photosynthetic performance in blueberry plants under the stress condition of water deficit. A greenhouse experiment was carried out under a 16-h light period; 20ºC; and 60-80% relative humidity for 2 weeks before treatment application of methyl jasmonate (MeJA) to blueberry plants (Vaccinium corymbosum, Brigitta cultivar). The following treatments were maintained for 7-days: (i) 80% field capacity (NoWD); (ii) 80% field capacity plus MeJA application (NoWD + MeJA); (iii) 20% field capacity (WD); and (iv) 20% field capacity plus MeJA application (WD + MeJA). The MeJA was sprayed as an aqueous solution of 10 µM MeJA (Sigma-Aldrich) over the plant's foliar system. At the end of the assay; blueberry leaves were analyzed for the relative water content; specific leaf area; lipid peroxidation; total antioxidant activity; total phenols; total anthocyanins; anthocyanidin compounds and photosynthetic performance. Brigitta cultivar showed a significant decrease in the oxidative stress at leaf levels; with an increase in antioxidant activity; phenolic compounds; total anthocyanins; delphinidin; petunidin; antheraxanthin; zeaxanthin; and an improvement in photosynthetic performance parameters. Blueberry Brigitta cultivar was shown to be susceptible to WD decreased mainly photosynthesis. However; the MeJA application on leaves induced metabolic changes; through an increase of antioxidant strategy within the plant to counteract the negative effects of WD; protecting the photosynthetic apparatus; which allows the Brigitta cultivar to withstand the period of WD.
Keywords:
abiotic stress
; antioxidant system
; drought
; jasmonates
; photosynthesis
1. Introduction
A large surface of the planet is covered by water (70%), but most of this volume is salty water (97.5%), leaving only 2.5% of fresh unsalted water for human consumption [1]. Most of this freshwater is unavailable (98.8% as groundwater, glaciers, and ice caps), and 1.2% of superficial water (lakes and rivers) supports the global population’s water necessity. Moreover, the surface freshwater fraction is naturally unequal distributed worldwide, and anthropogenic activities such as agriculture have contributed to intensifying the limiting conditions in some regions [2,3]. The impact of climate change on the precipitation pattern has affected water availability, and some croplands are facing severe water restrictions, resulting in long-term water deficit (WD) episodes and decreasing crop production. Crops are highly water-demanding, being affected by WD due to climate change [4,5].
The WD plant stress condition affects biochemical and physiological processes, in-ducing plant morphological changes. Metabolic processes involved in electron transport and energy production are mainly affected by WD, such as photosynthesis, dark respiration, and photorespiration [6]. Therefore, the production of highly antioxidant fruit species, like blueberries, can be seriously affected by WD. These crops are attractive for securing food production reasons. Under a WD condition, plant organelles are prone to undergo a sensitive imbalance, resulting in an exacerbated production and accumulation of reactive oxygen species (ROS) [7,8]. These partially reduced oxygen derivatives components are highly reactive and toxic, causing irreversible oxidation of cellular organelles, and detrimental cell metabolic activities [8]. The consequence of the oxidative damage in the membrane of stomata cell guards, produces an imbalance of the intra- and extra-cellular calcium, chloride and potassium concentrations, extending the membrane depolarization and causing permanent stomata closure. In addition, ROS are reported as crucial participants in guard cell signaling; where H2O2 is involved in ABA-induced stomatal closure [9]. This stomata closure restricts the fixation of atmospheric CO2, down-regulates photosynthesis, limits the water absorption, promote the photorespiration and stimulate even more the production of ROS [10,11]. Thus, WD through ROS production is considered the major cause of crop productivity loss [12,13], including blueberry plant orchards.
On the other hand, under abiotic stress conditions such as WD, the increased ROS formation can be linked to signals derived from changes in the regulation of phytohormones [14]. ROS are a common signal to down-regulate the release of phytohormones such as salicylic acid (SA), abscisic acid (ABA), ethylene (ET), jasmonic acid (JA) and its methyl ester the methyl jasmonate (MeJA) involved in the stress control [14,15,16]. MeJA is a phytohormone derived from the acetate pathway and is known for its role controlling aluminum stress in blueberry plants [17,18]; however, the effect of MeJA application to overcome WD has not yet been reported. Thus, this might be a useful solution to avoid the plant stress through WD periods in blueberry cultivars, under the current uncertain climate change scenarios. Therefore, this research aimed to determine whether exogenous methyl jasmonate application improves the antioxidant defense mechanisms for protecting the photosynthetic performance in blueberry plants under the stress condition of water deficit.
2. Materials and Methods
Plant material
Twenty plants of two-year-old Vaccinium corymbosum Brigitta cultivar were selected based on the resilience of the plants to abiotic stresses [19,20]. Brigitta cultivar plants were provided by Berries San Luis (Quillém, Lautaro, Chile; 38° 29` S, 72° 23` W). Healthy plants with a similar size were selected to set up the WD experiment.
Plant water deficit (WD) condition assay
The experiment was carried out in a greenhouse at the Instituto de Agroindustria, Universidad de La Frontera, Temuco, Chile (38°45’S, 72°40’W). The greenhouse growing conditions were established at 16-h light period, 20ºC, and 60-80% relative humidity. Plants were placed into pots containing 1.5 kg of dried and sieved Andisol soil. The ex-perimental design was completely randomized with five replicates per treatment. To minimize any positional effects, the pots with plants were re-randomized daily.
Water irrigation was applied periodically for plant conditioning, using distilled wa-ter during 14 days in order to maintain 80% field capacity (FC). After that, plants were subjected to different treatments as follow: (i) 80% field capacity (NoWD), (ii) 80% field capacity plus MeJA application (NoWD + MeJA), (iii) 20% field capacity (WD), and (iv) 20% field capacity plus MeJA application (WD + MeJA). Plants were kept as control (NoWD) and MeJA treatment applications (NoWD + MeJA, WD, and WD + MeJA) for 7 days. In this period, an aqueous solution of 10 µM MeJA (Sigma-Aldrich) was sprayed at the start of the experiment about the foliar system on treatments (NoWD + MeJA) and (WD), using the method described by Ulloa-Inostroza et al. [18]. Irrigation was applied twice a day using distilled water to maintain the differential water content treatments. At the end of the experimental period, in vivo photosynthesis and fluorescence measure-ments were done. Plant leaves and roots were harvested, separating enough fresh material for immediate lipid peroxidation (LP) analysis, and frozen the remaining material for subsequent biochemical analyses at -80°C (Revco Elite Series Ultra-Low Temperature, Thermo Scientific™).
Physiological analyses:
The relative water content (RWC) was determined based on the equation described by Larcher [21] as follow:
where, FW = fresh leaf weight; DW (g) = dry weight of leaves (g); and SW= saturation weight of the leaves (g). FW and DW were determined after drying at 60 °C and SW after soaking in water for 24 h at 20 °C.
RWC = [(FW - DW) / (SW - DW)] x 100
The specific leaf area (SLA) was determined by randomly selecting two leaves from each individual plant, which were placed on a white background and photographed with a reference point of 1 cm2. The SLA was calculated using the “Aurea” software developed by the Facultad de Ingeniería y Ciencias, Universidad de La Frontera, Chile, dividing each leaf area (LA) derived from the software by the dry weight (DW).
Biochemical analyses
The LP of membranes was determined as an indicator of oxidative stress. The meas-urement was carried out from fresh leaves and root, using thiobarbituric acid reacting substances (TBARS) according to the modified method described by Du and Bramlage [22] and measured spectrophotometrically at 440, 532, and 600 nm.
The total antioxidant activity (AA) in leaves and roots was determined by using the 2.1-diphenyl1-1-picrylhydrazyl (DPPH) method according to Chinnici et al. [23], measured at 515 nm and expressed in Trolox equivalents (TE).
Total phenols (TP) concentration was determined following the procedure described by Slinkard and Singleton [24] using Folin-Ciocalteau reagent and measured spectro-photometrically at 765 nm.
Total anthocyanins (TA) were estimated from leaves by the pH differential method as described by Cheng and Breen [25], and measured spectrophotometrically at 530 nm and at 657 nm with a molar extinction coefficient for cyanidin-3-glucoside of 29,600 nm. Results were expressed as micrograms of cyanidin 3-O-glycoside equivalent per g of fresh weight (µg cyanidin 3-O-glycoside eq. g−1 FW).
The anthocyanin compounds analyses were carried out by HPLC as described in Castrejón et al. [26]. The signals were detected at 530 nm and the data were expressed as milli- or micro-grams per g of fresh weight (mg or μg g−1 FW). The mobile phase was performed using acidified water (A) and 100% methanol (B). The binary gradient was applied as follows: 0-39.9 min of 90% A - 10% B, 40-41.9 min of 60% A - 40% B, 42-48.9 min of 20% A - 80% B, and 49-62 min of 90% A - 10% B according to Ribera-Fonseca et al. [27].
Photosynthetic performance
In vivo photochemical efficiency of leaves was determined by chlorophyll a fluores-cence with a modulated fluorometer (FMS 2; Hansatech Instruments, King's Lynn, UK). The evaluated parameters were: maximal photochemical efficiency of PSII [Fv/Fm = (Fm – Fo)/Fm], effective photochemical efficiency of PSII (ΦPSII), electron transport rate (ETR), and non-photochemical quenching (NPQ). These parameters were measured according to Reyes-Díaz et al. [28], and calculations were done according to Maxwell and Johnson [29].
In vivo measurements of net photosynthesis, transpiration, and stomatal conduct-ance were measured with a portable infrared gas analyzer (Licor-6800, LI-COR Bioscience, Inc., Lincoln, Nebraska, USA) according Reyes-Díaz et al. [19]. Intrinsic water-use effi-ciency of photosynthesis (WUEPh) was calculated according to Locke and Ort [30] as follow:
WUEPh = net photosynthesis / transpiration (µmol CO2 m-2s-1/mmol H2O m-2s-1).
Photosynthetic pigment concentrations in the leaves were carried out with 100 (v/v) HPLC grade acetone at 4ºC under a green safelight and centrifuged at 4ºC. The pigments were quantified using a high-performance liquid chromatography (HPLC) system (Agilent technologies 1200 series, column C-18 Waters spherisorb 5.0 μm ODS1 4.6 × 250 mm) according to Garcia-Plazaola and Becerril [31]. All standards for the pigments Vx, Ax, Zx, neoxanthin (Nx), chlorophyll (Chl) a, b, 𝛽-carotene (𝛽Ca) and lu-tein (Lt) were purchased from Sigma-Aldrich (Sigma Chemical Co. St. Louis, MO).
Statistical analyses
The data set was tested for normality applying equal variance and Kolmogo-rov-Smirnov normality tests. Then, one-way ANOVA parametric test was carried out, and differences between treatments were detected using the Tukey HSD test at 0.05 level. The analyses were performed with Statistical software SAS v. 8.01.
3. Results
3.1. Physiological Parameters
Differences for RLA and RWC among the control treatment (NoWD) and the treatment at water deficit (WD) were observed (p ≤ 0.05). However, no significant differences in RLA and RWC between control and treatment under normal water regimen with MeJA application (NoWD + MeJA) were observed (Table 1). WD decrease RLA by about 33% compared to NoWD and NoWD+MeJA, increasing 12% by MeJA application. A decrease of 22% was observed by WD in RWC, recovering 12% with MeJA application (Table 1).
3.2. Biochemical Determinations
Regarding LP, this parameter increased directly according to the WD in leaves (18.2%) and roots (2.4-fold) of each treatment compared with the control. However, MeJA application notoriously decreased LP in leaves of NoWD + MeJA treatment and WD+ MeJA compared to WD treatment (Figure 1A). While, in root all treatments LP were increased, being higher in the WD and WD + MeJA treatments (Figure 1C).
Highest values of AA were obtained in leaves and roots of treatment WD + MeJA compared with the remaining treatments and NoWD (Figure 1B,D, p ≤ 0.05). Further, AA from roots of treatment NoWD + MeJA decreased at 25% respect to the NoWD (Figure 1B,D). Non statistical significant differences were observed in leaves of treatments NoWD + MeJA and WD + MeJA, increasing around 32% this parameter respect to the control (Figure 1B,D).
The highest decrease of TP (26.5%) was observed for treatment WD compared with the NoWD (p ≤0.05). However, no significant differences among treatments NoWD + MeJA and WD+ MeJA with the control were observed (Table 2).
The lowest TA concentrations were observed for leaves of treatment WD compared with the remaining treatments and NoWD (p ≤ 0.05). However, MeJA application increased TA in treatment NoWD + MeJA (50%) compared with the NoWD, respectively (p ≤ 0.05, Table 2).
Regarding the anthocyanin profile in leaves, it was observed that delphinidin concentration decreased 56% for treatment WD compared with the NoWD. However, MeJA application in NoWD + MeJA treatment seemed to improve the response by a lower decline in this parameter (38%) compared with the NoWD of this compound in leaves. Likewise, cyanidin and petunidin concentrations decreased notoriously under WD and absence of MeJA (59.3%), being alleviated by MeJA application in treatment NoWD + MeJA (32%). On the other hand, for peonidin concentration, the main affecting factor seemed to be WD, because its lowest levels were found in treatment WD compared with the remaining treatments and NoWD, which did not show significant statistical differences among themselves (Table 2).
3.3. Photosynthetic Performance
Maximum quantum yield (Fv/Fm) for each WD and treatments (0.75 - 0.8) scored among the normal ranges (Figure 2). Thereby, the ΦPSII, ETR, and NPQ results did not show significant statistical differences between NoWD and treatment NoWD + MeJA. In contrast, ΦPSII and ETR were decreased in 28.5% and ~41% with WD, respectively (Figure 2B,C), while an alleviation (around at 33%) when MeJA [WD + MeJA] was applied (Figure 2B,C).
The photosynthetic parameters measured in the Blueberry control plants (NoWD) were within the range informed for healthy plants. For the plant treatments with MeJA application, i.e. (NoWD + MeJA) and (WD + MeJA), these parameters were improved (p ≤ 0.05, Figure 3). Specifically, the MeJA application contrasted with WD-plants caused the in-crease of net photosynthesis by 4-fold, 2-fold the stomatal conductance, and also transpiration, and water use efficiency (p ≤ 0.05, Figure 3).
The chlorophylls concentration decreased 20% in Brigitta cultivar with WD treatment without significant differences with MeJA application (Table 3). β-carotene and lutein decreased in all treatments compared to control plants, while zeaxanthin increased 1.8-fold with MeJA application and WD compared to WD (Table 3).
4. Discussion
In the present study, blueberry plants improve their performance with the application of MeJA (Table 1, Table 2 and Table 3; Figure 1, Figure 2 and Figure 3). Furthermore, these differences were observed for leaves of plants without WD plus the MeJA application, which notoriously decreased its oxidative stress level and increased its antioxidant activity (Figure 1).
The MeJA application notoriously increased TA, cyanidin, and peonidin in the Brigitta cultivar leaves (Table 2). Several studies report the increase of secondary metabolites and antioxidant compounds after the application of MeJA, but not on blueberry plants. Li et al. [32] evidenced a significantly increased concentration of phenolic compounds at the cuticular level on sunflower (Helianthus annuus), tomato (Solanum lycopersicum), and soja (Glycine max) plants after treatment with 0.1, 0.5, 1.0, and 2.5 mM of MeJA. In turn, for blueberry plants, the effect of MeJA has been investigated on foliar tissue from cultivars Legacy and Bluegold for other abiotic stress. Ulloa-Inostroza et al. [18] reported an increase in total antioxidant activity, the enzymatic activity of SOD and CAT, and total phenols for Legacy and Bluegold cultivars treated with 5 μM of exogenous MeJA under Al stress. Ribera-Fonseca et al. [33] evidenced that leaf extracts of Legacy cultivars treated with 0.05 mM MeJA significantly increased their antioxidant properties, mainly attributed to anthocyanins. Besides, the photosynthetic performance was concordant with typical values of MeJA-treated plants and similar to the reported by Ulloa-Inostroza et al. [18] in Legacy and Bluegold cultivars. Thus, the results from the Brigitta cultivar reinforce the capacity of this cultivar to respond to MeJA application, since the volatile nature of this compound efficiently permeates and enters the cell intraspace and makes itself available to participate in the biochemical secondary metabolic route. It seems that an intracellular concentration change scenario of MeJA is transcripted as an alert or secondary message that activates an antioxidant mechanism triggered by a stress condition.
As a response at seven days of WD, the Brigitta cultivar presented notorious shifts in the physiological and biochemical parameters, especially the increment of oxidative damage and antioxidant activity, and the reduction in the relative foliar area and photosynthesis (Table 1, Figure 1, Figure 2 and Figure 3), which are indicatives of a serial of biochemical imbalances induced by an exacerbate production and accumulation of ROS. As a result, the oxidative damage to the membrane of stomatal cells increases, driving the permanent closure of stomas by differences in the intra- and extra-cellular levels of calcium (Ca2+), chloride (Cl-), and potassium (K+) and prolonged membrane depolarization. Consequently, the stomatal closure restricts photosynthesis, CO2 fixation, and water consumption, whereas photorespiration is promoted and ROS production is potentiated [7,8,9]. Furthermore, the results showed that the Brigitta cultivar promoted in leaves and roots increment of antioxidant activity as a response to the water stress, to ameliorate the damage induced by WD (Figure 1B–D). However, our results showed that levels of total phenolic compounds and anthocyanins decrease under WD, and the total antioxidant activity could not be dependent on these compounds (Table 2). On the other, we also can explain it was previously described by Inostroza-Blancheteau et al. [20] for aluminum toxicity. They demonstrated that the antioxidant response of Brigitta cultivars was mainly mediated by antioxidant enzymes such as SOD and CAT, which transform ROS into water. In this context, the strategy of Brigitta could be to increase enzymatic antioxidants against water deficit, since this cultivar is prone to decrease its physiological activity under these stressing conditions compared to other cultivars such as Elliot, Bluegold, Biloxi, and Sharpblue. Those are more efficient in keeping their RWC under drought [34]. This evidence was supported by Améglio et al. [35], who determined that some cultivars, such as Bluecrop, can rapidly close their stomata and decrease evapotranspiration, which finally restricts water loss. Likewise, Molnar et al. [36] concluded that blueberry cultivar Goldtraube had the highest drought tolerance efficiency, followed by Bluecrop, Hortblue Petite, Duke, and Brigitta Blue.
The results from this research can support agronomic management decisions for the blueberry Brigitta cultivar, particularly under the climate change scenario, where the availability of water for production is uncertain. This cultivar is sensitive to water stress, and critical production stages like transplanting, orchard settling, and under certain phe-nological periods, could benefit from MeJA application.
In recent years, MeJA has received particular attention due to its ability to increase the production of antioxidant compounds against the oxidative effect of ROS at photosyn-thetic levels. In this way, our results demonstrated a significant (p < 0.05) reduction of the ROS damage in Brigitta cultivars treated with MeJA (Table 1, Table 2 and Table 3; Figure 1, Figure 2 and Figure 3). This effect was notorious at the leaf level, highlighting the increase in antioxidant activity, production of phenolic compounds, TA, delphinidin, petunidin, antheraxanthin, zeaxanthin, and photosynthetic performance parameters in the Brigitta cultivar (Table 1, Table 2 and Table 3; Figure 1, Figure 2 and Figure 3). Our results are in concordance with those shown by Wang [37], who observed that the increasing water retention by strawberry plants under water deficit was improved after MeJA application. Ma et al. [38] found that treating wheat plants with MeJA had a protective effect on their photosynthetic performance under water deficit. This protecting effect was attributed to the decreasing effect on ROS damage by MeJA, which ameliorated the decreasing effect on the net photosynthetic ratio and stomatal conductance, while at the same time, the efficiency in water uptake was improved. Additionally, the incremental on the photo-dissipator of energy elements such as delphinidin, petunidin, antheraxanthin, and zeaxanthin derived from MeJA application, could protect the performance of chloroplasts (Table 2 and Table 3). However, the activation of the antioxidant mechanism by MeJA is dependent on the water deficit degree [39]. Probably, the antioxidant response could be not only by MeJA but also by a summative effect of the stress condition. In this context, Su et al. [40] reported that ryegrass plants under heat stress treated with MeJA could activate the exact antioxidant mechanisms and elements for photo-dissipation of energy than observed for water deficit stress. This was at the same time associated with an increased expression of genes related to the biosynthetic pathway of jasmonate compounds (LpLOX2, LpAOC, LpOPR3, and LpJMT).
5. Conclusions
This study has demonstrated that the blueberry Brigitta cultivar was susceptible to WD, but the effect was ameliorated by applying MeJA on plant leaves. The stress condition caused by WD triggered plant metabolic changes associated with a strategy focused on the antioxidants to ameliorate the harmful effects of drought, by protecting the photosynthetic apparatus and its functionality in highbush blueberry, allowing the Brigitta cultivar to endure WD. On the other hand, our study has great relevance in the crops with economic importance since our results demonstrated that the MeJA application could be an interesting tool to improve the resistance of crops subjected to changing climatic conditions.
Author Contributions
E.M. U.-I., and M. R.-D.; methodology, formal analysis, investigation, and writing the original draft, E.M. U.-I., M. R.-D., C. C. and M. C.; review and editing, E.M. U.-I., M. R.-D., C.C. and M. C.; supervision, M. R.-D., and E.M. U.-I.; funding acquisition. All authors have read and agreed to the published version of the manuscript.
Funding
The publication was supported by funds from the FONDECYT 3180754, and 1211856 projects from ANID (Chile), FONDEQUIP-UAysén Ulloa-2018 project, URY 20993 “Centro de Innovación e Incubadora de Ideas U. Aysén (3i U. Aysén)”, Chilean Ministry of Education, and ANID/FONDAP/15130015.
Data Availability Statement
Not applicable.
Acknowledgments
We thank to the Centro de Innovación e Incubadora de Ideas U. Aysén (3i U. Aysén, Universidad de Aysén) and the Center of Plant, Soil Interaction and Natural Resources Biotechnology, Scientific and Technological Bioresource Nucleus (BIOREN-UFRO), Universidad de La Frontera.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Fekete, B.M. State of the world's water resources. In Climate Vulnerability: Understanding and Addressing Threats to Essential Resources; Pielke Sr., R.A., Ed.; Elsevier Inc.,: Amsterdam, 2013; pp. 11–23. [Google Scholar]
- Westmacott, J.R.; Burn, D.H. Climate change effects on the hydrologic regime within the Churchill-Nelson River Basin. J. Hydrol. 1997, 202, 263–279. [Google Scholar] [CrossRef]
- Wolf, A.T. Criteria for equitable allocations: The heart of international water conflict. Nat. Resour. Forum. 1999, 23, 3–30. [Google Scholar] [CrossRef]
- Boyer, J. Plant productivity and environment. Science 1982, 218, 443. [Google Scholar] [CrossRef]
- Rosa, L.; Chiarelli, D.D.; Rulli, M.C.; Dell’Angelo, J.; D’Odorico, P. Global agricultural economic water scarcity. Sci. Adv. 2020, 6, 1–10. [Google Scholar] [CrossRef]
- Urban, L.; Aarrouf, J.; Bidel, L.P.R. Assessing the effects of water deficit on photosynthesis using parameters derived from measurements of leaf gas exchange and of chlorophyll a fluorescence. Front. Plant Sci. 2017, 8, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Borhannuddin Bhuyan, M.H.M.; Parvin, K.; Bhuiyan, T.F.; Anee, T.I.; Nahar, K.; Hossen, Md.S.; Zulfiqar, F.; Alam, Md.M.; Fujita, M. Regulation of ROS metabolism in plants under environmental stress: A review of recent experimental evidence. Int. J. Mol. Sci. 2020, 21, 8695. [Google Scholar] [CrossRef]
- Mansoor, S.; Wani, O.A.; Lone, J.K.; Manhas, S.; Kour, N.; Alam, P.; Ahmad, A.; Ahmad, P. Reactive oxygen species in plants: From source to sink. Antioxidants 2022, 11, 225. [Google Scholar] [CrossRef]
- Singh, R.; Parihara, P.; Singha, S.; Mishra, R.K.; Singh, V.P.; Prasad, S.M. Reactive oxygen species signaling and stomatal movement: Current updates and future perspectives. Redox Biol. 2017, 11, 213–218. [Google Scholar] [CrossRef]
- Keles, Y.; Oncel, I. Response of antioxidative defence system to temperature and water stress combinations in wheat seedlings. Plant Sci. 2002, 163, 783–790. [Google Scholar] [CrossRef]
- Luna, C.M.; Pastori, G.M.; Driscoll, S.; Groten, K.; Bernard, S.; Foyer, C.H. Drought controls on H2O2 accumulation, catalase (CAT) activity and CAT gene expression in wheat. J Exp Bot. 2005, 56, 417. [Google Scholar] [CrossRef]
- Fahad, S.; Bajwa, A.A.; Nazir, U.; Anjum, S.A.; Farooq, A.; Zohaib, A.; Sadia, S.; Nasim, W.; Adkins, S.; Saud, S.; Ihsan, M.Z.; Alharby, H.; Wu, C.; Wang, D.; Huang, J. Crop Production under drought and heat stress: Plant responses and management options. Front. Plant Sci. 2017, 8, 1147. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Borhannuddin Bhuyan, M.H.M.; Zulfiqar, F.; Raza, A.; Mohsin, S.M.; Al Mahmud, J.; Fujita, M.; Fotopoulos, V. Reactive oxygen species and antioxidant defense in plants under abiotic stress: Revisiting the crucial role of a universal defense regulator. Antioxidants 2020, 9, 681. [Google Scholar] [CrossRef]
- Iqbal, S.; Wang, X.; Mubeen, I.; Kamran, M.; Kanwal, I.; Díaz, G.A.; Abbas, A.; Parveen, A.; Atiq, M.N.; Alshaya, H.; Zin El-Abedin, T.K.; Fahad, S. Phytohormones trigger drought tolerance in crop plants: Outlook and future perspectives. Front. Plant Sci. 2022, 12, 799318. [Google Scholar] [CrossRef] [PubMed]
- Kwak, J.M.; Nguyen, V.; Schroeder, J.I. The role of reactive oxygen species in hormonal responses1. Plant Physiol. 2006, 141, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Verma, V.; Ravindran, P.; Kumar, P.P. 2016. Plant hormone-mediated regulation of stress responses. BMC Plant Biol. 2016, 16, 86. [Google Scholar] [CrossRef] [PubMed]
- Reyes-Díaz, M.; Ulloa-Inostroza, E.M.; Gonzalez-Villagra, J.; Ivanov, A.G.; Kurepin, L.V. Phytohormonal responses to soil acidity in plants. In Plant hormones under challenging environmental factors; Ahammed, G.J., Yu, J.-Q., Eds.; Springer-Verlag: Berlin Heidelberg; New York, 2016; pp. 133–155. [Google Scholar]
- Ulloa-Inostroza, E.M.; Alberdi, M.; Meriño-Gergichevich, C.; Reyes-Díaz, M. Low doses of exogenous methyl jasmonate applied simultaneously with toxic aluminum improve the antioxidant performance of Vaccinium corymbosum. Plant Soil 2017, 412, 81–96. [Google Scholar] [CrossRef]
- Reyes-Díaz, M.; Inostroza-Blancheteau, C.; Millaleo, R.; Cruces, E.; Wulff-Zottele, C.; Alberdi, M.; Mora, M.L. Long-term aluminum exposure effects on physiological and biochemical features of highbush blueberry cultivars. J. Am. Soc. Hortic. Sci. 2010, 135, 1–11. [Google Scholar] [CrossRef]
- Inostroza-Blancheteau, C.; Reyes-Diaz, M.; Aquea, F.; Nunes-Nesi, A.; Alberdi, M.; Arce-Johnson, P. Biochemical and molecular changes in response to aluminium-stress in highbush blueberry (Vaccinium corymbosum L). Plant Physiol. Bioch. 2011, 49, 1005–1012. [Google Scholar] [CrossRef]
- Larcher, W. Physiological plant ecology. Ecophysiology and stress physiology of functional groups, 4th edition; Springer-Verlag: Berlin Heidelberg; New York, 2003; 504p. [Google Scholar]
- Du, Z.; Bramlage, W.J. Modified thiobarbituric acid assay for measuring lipidoxidation in sugar-rich plant tissue extracts. J. Agr. Food Chem. 1992, 40, 1556–1570. [Google Scholar] [CrossRef]
- Chinnici, F.; Bendini, A.; Gaiani, Y.; Riponi, C. Radical scavenging activities of peels and pulps from cv. Golden delicious apples as related to their phenolic composition. J. Agr. Food Chem. 2004, 52, 4684–4689. [Google Scholar] [CrossRef]
- Slinkard, K.; Singleton, V.A. Total phenol analysis: automation and comparison with manual methods. Am. J. Enol. Vitic. 1977, 28, 29–55. [Google Scholar] [CrossRef]
- Cheng, G.W.; Breen, P.J. Activity of phenylalanine ammonia-lyase (PAL) and concentrations of anthocyanins and phenolics in developing strawberry fruit. J. Am. Soc. Hort. Sci. 1991, 116, 865–869. [Google Scholar] [CrossRef]
- Castrejón, A.D.R.; Eichholz, I.; Rohn, S.; Kroh, L.W.; Huyskens-Keil, S. Phenolic profile and antioxidant activity of highbush blueberry (Vaccinium corymbosum L.) during fruit maturation and ripening. Food Chem. 2008, 109, 564–572. [Google Scholar] [CrossRef]
- Ribera, A.E.; Reyes-Díaz, M.; Alberdi, M.; Zuñiga, G.E.; Mora, M.L. Antioxidant compounds in skin and pulp of fruits change among genotypes and maturity stages in highbush blueberry (Vaccinium corymbosum L.) grown in southern Chile. J. Soil. Sci. Plant Nutr. 2010, 10, 509–536. [Google Scholar] [CrossRef]
- Reyes-Díaz, M.; Alberdi, M.; Mora, M.L. Short-term aluminum stress differentially affects the photochemical efficiency of photosystem II in highbush blueberry genotypes. J. Am. Soc. Hortic. Sci. 2009, 134, 1–8. [Google Scholar] [CrossRef]
- Maxwell, K.; Johnson, G. Chlorophyll fluorescence. A practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Locke, A.M.; Ort, D.R. Leaf hydraulic conductance declines in coordination with photosynthesis, transpiration and leaf water status as soybean leaves age regardless of soil moisture. J. Exp. Bot. 2014, 65(22), 6617–6627. [Google Scholar] [CrossRef]
- Garcia-Plazaola, J.I.; Becerril, J.M. A rapid HPLC method to measure liphophilic antioxidant in stressed plants: simultaneous determination of carotenoids and tocopherols. Phytocheml. Anal. 1999, 10, 307–313. [Google Scholar] [CrossRef]
- Li, C.; Wang, P.; Menzies, N.W.; Lombi, E.; Kopittke, P.M. Effects of methyl jasmonate on plant growth and leaf properties. J. Plant Nutr. Soil Sci. 2018, 181(3), 1–10. [Google Scholar] [CrossRef]
- Ribera-Fonseca, A.; Jiménez, D.; Leal, P.; Riquelme, I.; Roa, J.C.; Alberdi, M.; Peek, R.M.; Reyes-Díaz, M. The anti-proliferative and anti-invasive effect of leaf extracts of blueberry plants treated with methyl jasmonate on human gastric cancer in vitro is related to their antioxidant properties. Antioxidants 2020, 9, 45. [Google Scholar] [CrossRef]
- Balboa, K.; Ballesteros, G.I.; Molina-Montenegro, M.A. Integration of physiological and molecular traits would help to improve the insights of drought resistance in highbush blueberry cultivars. Plants 2020, 9, 1457. [Google Scholar] [CrossRef] [PubMed]
- Améglio, T.; Roux, L.; Perrier, M.C. Water relations of highbush blueberry under drought conditions. Acta Hortic. 2000, 537, 273–278. [Google Scholar] [CrossRef]
- Molnar, S.; Clapa, D.; Viorel, M. Response of the five highbush blueberry cultivars to in vitro induced drought stress by polyethylene glycol. Agronomy 2022, 12(3), 732. [Google Scholar] [CrossRef]
- Wang, S.Y. Methyl jasmonate reduces water stress in strawberry. Plant Growth Regul. 1999, 18, 127–134. [Google Scholar] [CrossRef]
- Ma, C.; Wang, Z.Q.; Zhang, L.T.; Sun, M.M.; Lin, T.B. Photosynthetic responses of wheat (Triticum aestivum L.) to combined effects of drought and exogenous methyl jasmonate. Photosynthetica 2014, 52, 377–385. [Google Scholar] [CrossRef]
- Ðurić, M.; Subotić, A.; Prokić, L.; Trifunović-Momčilov, M.; Milošević, S. Alterations in physiological, biochemical, and molecular responses of Impatiens walleriana to drought by methyl jasmonate foliar application. Genes 2023, 14, 1072. [Google Scholar] [CrossRef]
- Su, Y.; Huang, Y.; Dong, X.; Wang, R.; Tang, M.; Cai, J.; Chen, J.; Zhang, X.; Nie, G. Exogenous methyl jasmonate improves heat tolerance of perennial ryegrass through alteration of osmotic adjustment, antioxidant defense, and expression of jasmonic acid-responsive genes. Front. Plant Sci. 2021, 12, 664519. [Google Scholar] [CrossRef]
Figure 1.
The lipid peroxidation (LP) and antioxidant activity (AA) in Brigitta cultivar under WD and MeJA application in leaves. Lowercase letters show the significant statistical differences among control and treatments (p ≤ 0.05). Each value is the mean of five replicates with standard deviation of the mean.
Figure 1.
The lipid peroxidation (LP) and antioxidant activity (AA) in Brigitta cultivar under WD and MeJA application in leaves. Lowercase letters show the significant statistical differences among control and treatments (p ≤ 0.05). Each value is the mean of five replicates with standard deviation of the mean.
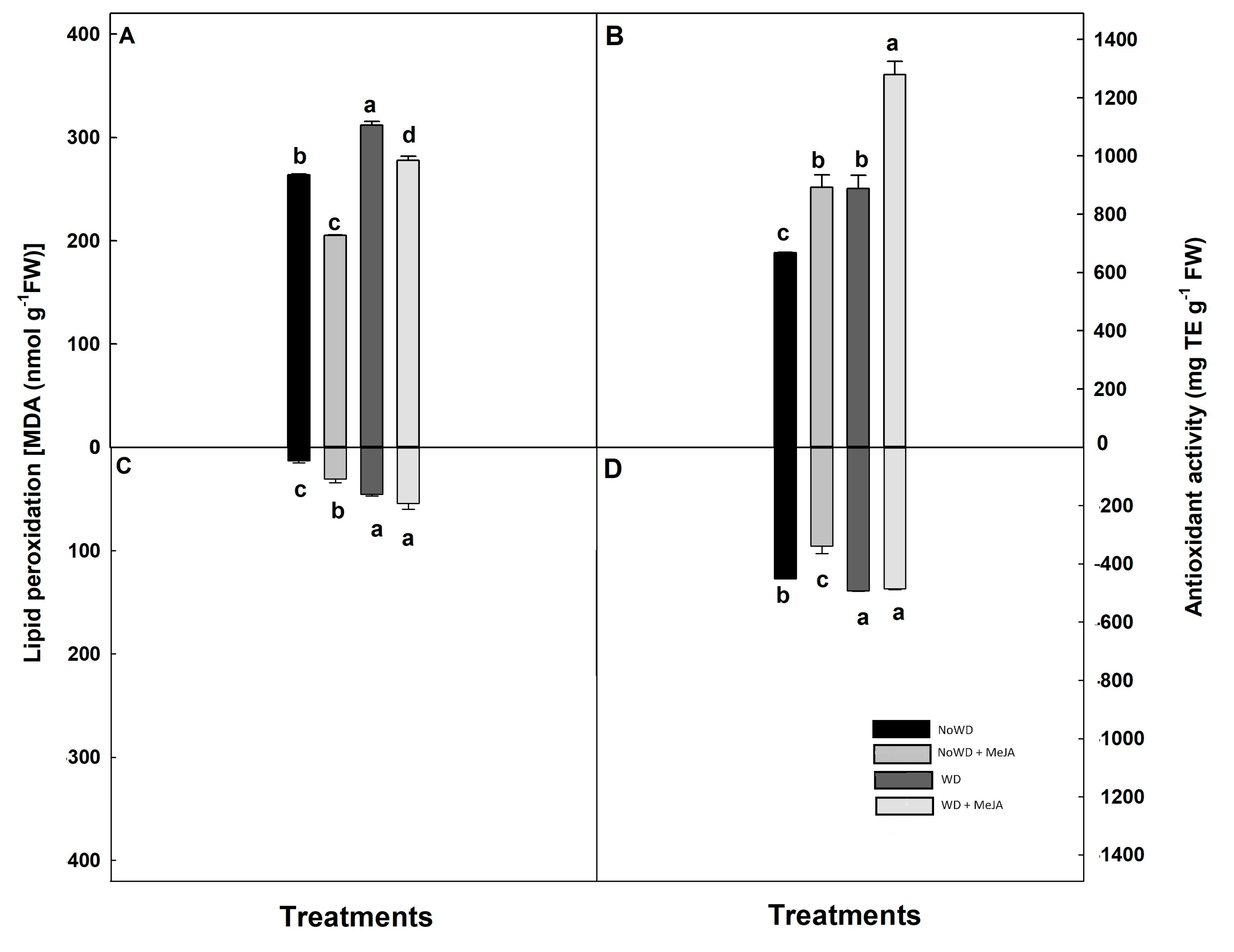
Figure 2.
Maximum quantum yield (Fv/Fm), effective quantum yield (ΦPSII), electron transport rate (ETR), and non-photochemical (NPQ) in Brigitta cultivar under WD and MeJA application leaves. Lowercase letters show the significant statistically differences among control and treat-ments (p ≤ 0.05). Each value is the mean of five replicates with standard deviation of the mean.
Figure 2.
Maximum quantum yield (Fv/Fm), effective quantum yield (ΦPSII), electron transport rate (ETR), and non-photochemical (NPQ) in Brigitta cultivar under WD and MeJA application leaves. Lowercase letters show the significant statistically differences among control and treat-ments (p ≤ 0.05). Each value is the mean of five replicates with standard deviation of the mean.
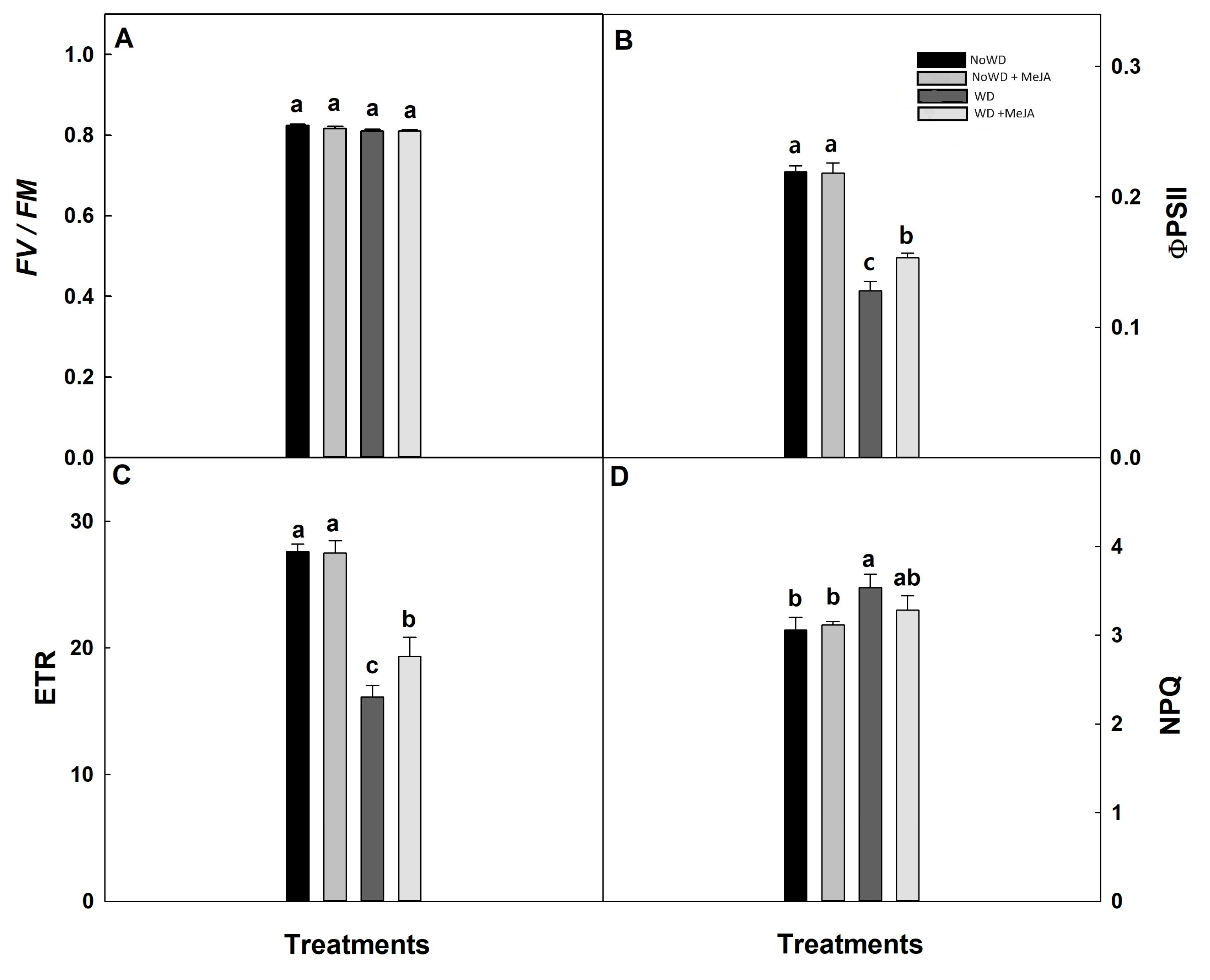
Figure 3.
Photosynthetic rate (A), stomatal conductance (B), transpiration rate (C) and water use efficiency (D) in Brigitta cultivar under WD and MeJA application in leaves. Lowercase letters show the significant statistical differences among control and treatments (p ≤ 0.05). Each value is the mean of five replicates with standard deviation of the mean.
Figure 3.
Photosynthetic rate (A), stomatal conductance (B), transpiration rate (C) and water use efficiency (D) in Brigitta cultivar under WD and MeJA application in leaves. Lowercase letters show the significant statistical differences among control and treatments (p ≤ 0.05). Each value is the mean of five replicates with standard deviation of the mean.
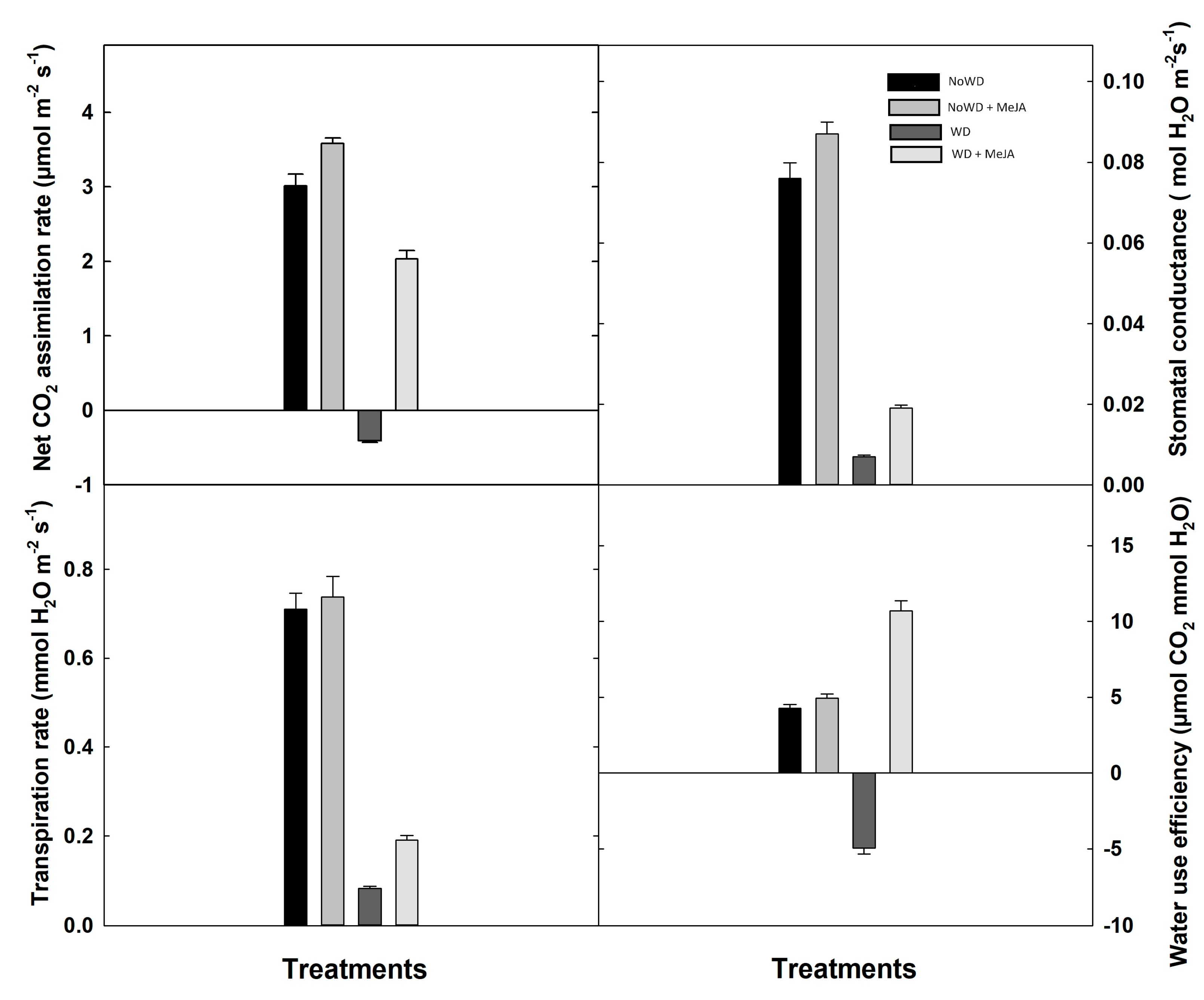

Table 1.
Relative leaf area (RLA) and Relative water content (RWC) in Blueberry Brigitta cultivar under WD and MeJA application in leaves. Lowercase letters show the significant statistical differences among control and treatments (p ≤ 0.05). Each value is the mean of five replicates with standard deviation of the mean.
Table 1.
Relative leaf area (RLA) and Relative water content (RWC) in Blueberry Brigitta cultivar under WD and MeJA application in leaves. Lowercase letters show the significant statistical differences among control and treatments (p ≤ 0.05). Each value is the mean of five replicates with standard deviation of the mean.
| Treatments | FC (%) |
MeJA (mM) |
RLA [Mean ± SE (cm2/g)] |
RWC [Mean ± SE (%)] |
|---|---|---|---|---|
| NoWD | 80 | 0 | 197.6 ± 3.3 a | 63.3 ± 3.4 ab |
| NoWD + MeJA | 80 | 10 | 197.7 ± 10.9 a | 69.3 ± 3.0 a |
| WD | 20 | 0 | 132.0 ± 8.1 c | 54.2 ± 3.5 c |
| WD + MeJA | 20 | 10 | 150.9 ± 7.3 b | 61.4 ± 2.1 b |
Table 2.
Total phenols, total anthocyanins, delphinidin, cyanidin, petunidin, peonidin in Brigitta cultivar under WD and MeJA application in leaves. Lowercase letters show the significant statistical differences among control and treatments (p ≤ 0.05). Each value is the mean of five replicates.
Table 2.
Total phenols, total anthocyanins, delphinidin, cyanidin, petunidin, peonidin in Brigitta cultivar under WD and MeJA application in leaves. Lowercase letters show the significant statistical differences among control and treatments (p ≤ 0.05). Each value is the mean of five replicates.
| Treatments | FC (%) | MeJA (µM) | Total Phenols [Chlorogenic Acid eq (μg g−1 FW)] | Total Anthocyanins (mg Cyanidin 3-O-Glycoside/g−1 FW)] | Delphinidin (mg/g−1 PF) | Cyanidin (mg/g−1 PF) | Petunidin (µg/g−1 PF) | Peonidin (µg/g−1 PF) |
|---|---|---|---|---|---|---|---|---|
| NoWD | 80 | 0 | 1433.7 ± 33.2 a | 1.0 ± 0.0 b | 50.4 ± 3.1 a | 3.2 ± 0.0 b | 146.5 ± 6.1 a | 77.0 ± 2.7 b |
| NoWD + MeJA | 80 | 10 | 1524.1 ± 154.5 a | 1.5 ± 0.0 a | 41.3 ± 0.1 b | 3.5 ± 0.0 a | 142.9 ± 5.0 a | 92.3 ± 4.2 a |
| WD | 20 | 0 | 1053.8 ± 47.9 b | 0.2 ± 0.0 c | 22.1 ± 2.0 d | 1.3 ± 0.1 c | 99.9 ± 7.0 b | 48.9 ± 5.9 c |
| WD + MeJA | 20 | 10 | 1345.9 ± 123.9 a | 0.9 ± 0.1 b | 31.6 ± 1.8 c | 1.5 ± 0.0 c | 144.9 ± 4.8 a | 52.4 ± 3.4 c |
Table 3.
The Chl a+b and Chl a/b concentration, and carotenoids concentration in Brigitta cultivar under WD and MeJA application leaves. Lowercase letters show the significant statistical differences among control and treatments (p ≤ 0.05). Each value is the mean of five replicates with standard deviation of the mean.
Table 3.
The Chl a+b and Chl a/b concentration, and carotenoids concentration in Brigitta cultivar under WD and MeJA application leaves. Lowercase letters show the significant statistical differences among control and treatments (p ≤ 0.05). Each value is the mean of five replicates with standard deviation of the mean.
| FC (%) | MeJA (µM) | Chlorophyll a+b | Chlorophyll a/b | Beta Carotene | Lutein | Violaxanthin | Antheraxanthin | Zeaxanthin | Neoxanthin | |
|---|---|---|---|---|---|---|---|---|---|---|
| NoWD | 80 | 0 | 1751.3 ± 126.6 a | 2.6 ± 0.2 ab | 110.2 ± 5.9 a | 188.1 ± 10.2 a | 81.4 ± 6.0 a | 27.3 ± 1.5 b | 70.3 ± 3.0 b | 87.9 ± 5.9 a |
| NoWD + MeJA | 80 | 10 | 1101.3 ± 60.7 c | 2.7 ± 0.0 ab | 63.7 ± 3.8 b | 116.7 ± 3.6 b | 66.4 ± 1.5 ab | 16.8 ± 1.3 c | 59.3 ± 1.4 b | 47.4 ± 1.6 c |
| WD | 20 | 0 | 1409.5 ± 49.9 b | 2.6 ± 0.0 b | 72.8 ± 4.9 b | 139.7 ± 9.3 b | 39.8 ± 2.9 c | 42.8 ± 0.7 a | 83.8 ± 9.5 b | 62.1 ± 3.5 b |
| WD + MeJA | 20 | 10 | 1461.2 ± 31.6 b | 2.8 ± 0.1 a | 82.7 ± 4.5 b | 149.5 ± 8.4 b | 52.5 ± 5.4 bc | 36.7 ± 0.5 a | 127.4 ± 13.7 a | 66.1 ± 1.7 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



