
Submitted:
03 October 2023
Posted:
05 October 2023
You are already at the latest version
Abstract
Phosphorus deficiency highly interferes with plant growth and development. Plants respond to the persistent P deficiency by coordinating the expression of genes involved in the alleviation of the stress. Promoters of phosphate transporter genes are of great choice for the development of genetically modified plants with enhanced phosphate uptake abilities, which improve crop yields in phosphate-deficient soils. In our previous study, the sugarcane phosphate transporter PHT1;2 gene, showed a significantly high expression under salinity stress. In this study, the Erianthus arundinaceus EaPHT1;2 gene was isolated and characterized using various in-silico tools. The deduced 542 amino acid residues have 10 transmembrane domains, with a molecular weight and isoelectric point of 58.9 kDa and 9.80, respectively. They displayed 71–96% similarity with Arabidopsis thaliana, Zea mays, and Saccharum hybrid. To elucidate the function of the 5’ regulatory region, the 1.1kb promoter was isolated and validated in tobacco transgenics under Pi stress. The EaPHT1;2 promoter activity was detected using the β-glucuronidase (GUS) assay. While the EaPHT1;2 promoter showed 3 to 4.2-fold higher expression than the most widely used CaMV35S promoter. The 5’ deletion analysis with and without 5’UTR regions, revealed that a small size 374 bp fragment with the highest promoter activity among 5′ truncated fragments, which was 2.7 and 4.2 times higher than the well-used CaMV35S promoter under normal and Pi-deprivation conditions, respectively. The strong and short promoter of EaPHT1;2 with 374 bp showed significant expression in low Pi-stress conditions and it could be a valuable source for the development of stress-tolerant transgenic crops
Keywords:
Erianthus arundinaceus
; High-affinity phosphate transporter
; PHT1
; 2
; Promoter analysis
; Tobacco transgenic
; Pi stress
1. Introduction
Phosphate (P) is a crucial nutrient essential for plant growth, physiological responses, and stress tolerance, and its efficient uptake is vital for crop productivity. Inorganic phosphate (Pi) is the main source of P for plants, however, it can be challenging for them to absorb because it is in complex with metal ions in the soil [1]. Pi is transported in plants through root cells, xylem, and shoot tissues, involving specific Pi transporters. Phosphate transporters help maintain phosphate homeostasis within plants. These transporters are responsible for Pi uptake, translocation, and remobilization [1,2]. The availability of P in the soil is often limited and therefore plants have evolved various mechanisms to acquire and transport phosphate ions (Pi) from the soil to meet their physiological needs [1,3]. Under limited Pi availability plants employ various strategies to adapt to their environment. These adaptations encompass symbiotic approaches, alterations in root architecture, the release of organic acids, and the production of acid phosphatases by roots [4,5]. However, under stress conditions plants exhibit adaptive mechanisms to optimize the nutrients, but they are not robust enough to prevent the struggle against detrimental effects like stress signaling and stress-response mechanisms [6,7,8,9]. Soil conditions, climate change, fertilizer costs, and plant use efficiency are critical factors that influence P availability to plants [10,11]. Nutrient deficiency, coupled with abiotic stresses such as drought, salinity, and temperature can also have profound effects on plants, impacting their growth, and development, reducing crop yields, which in turn affects the global food security [12].
The PHT1 family has been the most extensively studied plant phosphate transporters. The PHT1 is a plasma membrane protein that plays the main role in direct Pi uptake from soil by plant roots. The movement of Pi via PHT1 is driven by plasma membrane H+-ATPase which belongs to the family of H+/Pi symporters, within the major facilitator superfamily (MFS) and possesses high Pi affinity, and strong expression in roots, especially most of them are up-regulated in Pi-deprived plants [13,14]. Since 1996, functional characterization of PHT1 members has been conducted using mutant lines and transgenic plants. Lately, the precise functions of these members have been examined using various single and multiple mutants produced by synthetic microRNA silencing. Recent studies using GWAS and in-silico analyses of plant genomes revealed PHT1 genes in several crop plants like, barley, rice, maize, foxtail millet, tomato, poplar, apple, wheat, sorghum, rapeseed, flax seed, and tea [15,16,17,18,19,20,21,22,23,24,25,26], while their precise functions are still under investigation.
In plants the majority of PHT1 genes are transcriptionally activated by Pi deficiency and up-regulated in roots, shoots, or both. These responses in PHT genes are regulated by the transcription factors (TFs), linked to consensus cis-acting sequences. The well-known, phosphate starvation response (PHR) transcription factors, from the MYB-CC family is the key regulator of Pi starvation signalling. The PHR TFs, regulate phosphate starvationinduced (PSI) gene expression by binding to P-responsive P1BS element (GNATATNC) in their promoter regions [27,28,29,30,31]. Other regulatory components like microRNA miR339, PHF1, SIZ1, PHR1, MYB62, WRKY75 have also been reported in Arabidopsis and Rice [32,33]. Beyond, Pi starvation signalling, PHT genes are also regulated transcriptionally indicating a complex regulatory mechanism than anticipated. Several cis-elements have been reported for their role in the transcriptional signalling of expression of Pi starvation-responsive genes. Cis-acting elements like P1BS, MBS, W-box, G(E)-box, NIT 2, and PHO elements were reported to be associated with the responsiveness of PHT1s and PSI genes [1,34,35,36,37].
Sugarcane holds significant economic importance as it is cultivated in approximately 121 nations, and produces the impressive cane yield of 70.9 tonnes per hectare [38]. This crop plays a pivotal role in food and industrial uses, nearly 80% of the sugar consumed globally is made from sugarcane. Sugarcane productivity is significantly limited by phosphorus (P), which is crucial for vegetative development and crop durability. P fertilizers address P deficiency in high-input agricultural systems, ~120 kg P ha−1 annually, this legacy phosphorus can indeed become a valuable resource if managed effectively within crop systems [12,39]. Brazil, the largest sugarcane producer, has seen significant growth in stalk yields since 1975 due to improved soil quality, plant breeding, and crop agronomy. However, Brazil's sugarcane production faces challenges due to low phosphorus availability, affecting 20% of fertilizer consumption, making nutrient security vulnerable to future scarcity [40].
We functional characterized the high-affinity phosphate transporter 1 (PHT1) gene promoter from sugarcane (Saccharum spp hybrid) based on our previous report [41,42]. In this current study we isolated the full-length coding sequence and promoter sequences of EaPHT1;2 gene from Erianthus arundinaceus, a wild genus of Saccharum spp. The EaPHT1;2 gene and promoter region were isolated and characterized using various bioinformatics tools, and further the EaPHT1;2 promoter functionally validated in Tobacco. The full-length EaPHT1;2 promoter and a series of truncated promoters with and without 5’UTR region were fused to the GUS reporter gene to identify their expression under Pi stress. We found that in transgenic tobacco, the shortest promoter of 374 bp has significantly strong GUS expression under Low-Pi stress condition.
2. Results
2.1. Isolation and sequence analysis of EaPHT1;2 gene from E. arundinaceus
In our previous study, the PHT1;2 gene sequence was obtained from the Sugarcane genome of Saccharum spp. hybrid (Sh) cultivar R570, thus the gene-specific primer pair was designed based on the conserved regions in Saccharum spp. and Z. mays, was used for isolation of PHT1;2 gene from E. arundinaceus genomic DNA (Figure 1A). The amplicon was cloned and sequenced for further characterization (Figure 1B). BLAST analysis of EaPHT1;2 showed 71–96% similarity with ZmPHT1, SbPHT1, ShPHT1, and AtPHT1. A phylogenetic analysis using MEGA was carried out to confirm the evolutionary relationships among these PHT1 proteins (Figure 1C). The amplified full-length coding sequence of EaPHT1;2 resulted an open reading frame (ORF) of 1626 bp, which encodes a protein of 542 amino acid residues with a molecular weight of 58.9 kDa (Figure S1). Further physico-chemical properties were also predicated using ProtParam tool (Table S2 and Figure S2). The result showed that the EaPHT1;2 protein has theoretical isoelectric point (pI) of 9.80 and Grand average of hydropathicity (GRAVY) of 0.019. Analysis of active domains in the coding sequence confirmed the presence of a functional domain with signature sequences characteristic to the Major facilitator superfamily (MFS, PS50850) family (Figure 2A). The protein was anticipated to contain 10 transmembrane domains (TMs), with a hydrophilic loop in the central part of the domains were initially identified using TMHMM Server v. 2.0 and further verified using Kyte Doolittle Hydropathy Plot (Figure 2B,C). The post-translation modification analysis revealed a total of 39 phosphorylation sites with the distribution of 24:14:1, Ser:Thr:Tyr sites and most of the discovered phosphopeptides are monophosphorylated. N-glycosylation site analysis, identified a potential NSTT site for N-linked glycosylation at 425 amino acids position (Figure S3).
2.2. Structural prediction of EaPHT1;2 protein
The secondary structure prediction using SOPMA (Figure S4) revealed a predominance of an alpha helix with 37.45% followed by random coil (35.79%), extended strand (17.71%), and beta turn (9.04%). As presented in Figure 3A, the 2D structure resulted from the PSIPRED server exhibited 22 α-helices, 1 β-strands, and 20 coils structures. Analysis of PHT1 protein specific signature sequence (GGDYPLSATIxSE) of EaPHT1;2 in comparison with PHT1 proteins of Z. mays, S. bicolor, Arabidopsis and Saccharum hybrid revealed the existences of conserved identical 'GGDYPLSATIMSE' sequence in the protein (Figure 3B). The homology modelling of protein was performed using the Phyre2 and Swiss-Model. The 3D model resulted from Phyre2 analysis with c4j05A as a template having a confidence value of 100% indicated 77% coverage (420 residues) of EaPHT1;2 sequence. However, the protein ID for the Swiss-Model is 7sp5.1.A, showed 22-523 sequence range, and Q-mean of 0.6, indicated that the developed using homology modelling of EaPHT1;2 with 92.4% coverage (Figure 4A,B). The resulting model was validated with Ramachandran plot analysis, 86% and 89% of the residues were located in the most favourable area for the Phyre2 and Swiss-Model structures, respectively (Figure 4C,D). As a result the models produced by Swiss-Model is more acceptable than those produced by Phyre2 (Figure 4E,F). Qualitative evaluation of EaPHT1;2 protein 3D structure was also done using ProSA webserver (Figure S4B,C). The structure of EaPHT1;2 in complex with substrate, PHOSPHATE ION (PO4) an inorganic phosphate was determined to have 4 Å resolution with six phosphate binding sites (A:Y.152, A:Q.179, A:Q.179, A:W.322, A:W.322, A:Y.330) interact through hydrogen bonds (2.44 – 4.0 Å distances) to phosphate in chain A (Figure 4G,H). The electrostatic surface analysis of the EaPHT1;2 protein was visualized using UCSF Chimera.
2.3. Protein association network and subcellular localization of analysis
EaPHT1;2 protein was subjected to a protein-protein interaction (PPI) study using STRING, which was predicated based on interactions between homologous proteins from S. bicolor, Z. mays, and Arabidopsis. The findings showed that the EaPHT1;2 protein interacts with more than 10 possible protein partners of three different genomes. The PPI interaction results indicated that EaPHT1;2 (Red colored node - Pht1, Sb01g020570.1, and PHT1;1 in Figure S5) protein, majority of the proteins are belonging to the Phosphate transporter family, like MFS transporter, phosphodiesterase precursor, SPX domain protein, sodium-phosphate symporter, solute carrier, and sugar transporter. Between ZmPHT1 and EaPHT1;2 interaction, twenty proteins were predicted as functional partners, two of the members (umc1363a and expB8) belonging to the expansin family, and other proteins like cps2, NAR2.2 and pco091084 belongs to the chaperonin (HSP60) family, the component of nitrate transporter and histidine acid phosphatase family proteins, respectively. The PPI analysis revealed that the EaPHT1;2 protein, interacted with phosphate transporter protein families (Figure S5A). The PPI interaction of EaPHT1 based on Arabidopsis, the PHT1 protein, a member of the MYB-CC family, acts as a major integrator of phosphate starvation responses by regulating FER1 expression in response to phosphate deficiency, thereby connecting iron and phosphate homeostasis. RNS1, a member of the ribonuclease T2 family, respond to inorganic phosphate starvation and wound-induced signalling. SPX1 and PHR1 are both in different ways involved in phosphate starvation in plants (Figure S5B). With respect to Sorghum-based EaPHT1 interaction, two members namely Actin 7 and actin7-like proteins were actively engaged in numerous sorts of cell motility processes and are universally found in all eukaryotic cells (Figure S5C). The subcellular localization analysis of EaPHT1;2 protein was predicted using the DeepLoc1.0. The EaPHT1;2 protein localization of various organelles was distinguish by approximate values are presented in Table S3. The analysis anticipated that the EaPHT1;2 protein was more likely to be located in the cell membrane (Figure S6A). The Quick GO analysis also revealed that the EaPHT1;2 protein is a cellular component that present in cell periphery region particularly in the plasma membrane (Figure S6B).
2.4. Isolation of PHT1 promoter region using a genome walking method
The 5’ promoter region present upstream of the translation initiation codon (ATG) of EaPHT1;2 gene was isolated using the random amplification of genome ends (RAGE) approach. Five different genomic DNA libraries were generated by digestion of the Erianthus genomic DNA using the blunt end restriction enzymes (Figure 5A) and ligated with the adapter sequence were used as the template for the primary PCR reaction. The primary PCR was performed using the ASP I and GSP I primers (primers are listed in supplementary Table S2). Further, the secondary PCR reactions were carried out using the primary PCR amplified product as a temple (Figure 5B) using the ASP II and GSP II primers. The amplicon of putative promoter region ~1500 bp, obtained DraI genomic DNA library was isolated, cloned and sequenced. Finally, after sequencing, 1102 bp of 5’ upstream region of EaPHT1;2 promoter region of was obtained through sequences alignments with gene sequence.
2.5. In-silico analysis of Promoter
The EaPHT1;2 promoter sequence was analyzed by means of online tools and software. To identify the transcription start site, the promoter sequence of EaPHT1;2 was analyzed by using NNPP. The closest predicted TSS on the promoter sequence begins 83 base pairs ahead of the ATG codon (Figure 5C). The analysis based on PLACE, and PlantCARE tools indicated the existence of at least 28 different types of regulatory motifs, including conserved motifs such as the TATA box, CAAT box and Pi starvation responsive motifs P1BS (GNATATNC). The root-specific motif like ASF1, OSE1, OSE2, RAV1AAT, SURE core, TAPOX1 and ARFAT were also mapped in Figure 5D. The EaPHT1;2 promoter region was also compared with ZmPHT1 and ShPHT1;2 promoter regions (Figure S7A). In Z. mays and E. arundinaceus two and one conserved regions were spotted with different TFs, respectively (Figure S7B). Multiple cis-elements that enables the stress response, inducible or tissue-specific expression like ABA, salinity, dehydration, light responsive, hormones responsive elements, pathogen/elicitor sensitive motifs were found at numerous positions in EaPHT1;2 promoter sequence.
2.6. Expression analysis of EaPHT1;2 promoters in tobacco transgenic events
To test the promoter activity, a total of 6 constructs namely promoter Full-length (pFL) and Deletions: pD1- pD5 were generated and transformed into tobacco. Putative transgenic plants were validated and confirmed using two sets of specific primer pairs (data not shown). The putative T0 generation tobacco transgenic events were subjected to a GUS histochemical assay and the untransformed wild-type plant serve as control. GUS expression analysis in tobacco events showed a strong constitutive expression in all tissues (Figure 6A). The histochemical assay findings revealed that pFL::GUS were efficient to drive the expression of gene in all tissues of transgenic tobacco events. Comparatively pFL::GUS promoter had a higher level of GUS expression than the pCaMV35S::GUS, and in contrast, it is challenging to detect a substantial variation in expression intensities by using only GUS histochemical analysis. Therefore, fluorometric GUS assay of the leaf, stem, and root tissues of 60-days old-plants (GUS positive events) were taken and the assay was conducted to elucidate the difference in GUS expression levels between the pFL::GUS and pCaMV35S::GUS. Enhanced GUS activities was recorded in the roots, stems, and leaves of pFL::GUS compared to pCaMV35S::GUS promoter transgenic plants. MUG assay revealed that 1.5 times higher GUS activity in pFL::GUS events compared to CaMV35S promoter (Figure 6C). Therefore, to determine the activities of various regulatory regions of the pFL promoter, a series of 5′-deletion analysis with and without 5’ UTR regions were carried out (Figure 6B). The study showed that all EaPHT1;2 promoter fragments drove equally regulated gene expression in all the tissues, despite variation in GUS staining intensity. The flurometric GUS activity showed 1.2 and 2.6 times increased expression in deletion promoter’s, pD1 and pD2, and the promoter’s pD3, pD4 and pD5 showed similar or slight expression as compared to CaMV35S promoter, respectively. The result revealed that plants with 5’UTR region in deletion constructs pD1, and pD2 showed a substantial increase in GUS expression compared to pD3 to pD5 promoters without 5’UTR region. Confirming that the 5′ deletion analysis with or without 5’ UTR region did not result in function loss.
2.7. EaPHT1;2 promoter activity in Pi stress
Twenty-days-old seedlings of the T1 generation were subjected to low (0.1mM) and high (1mM) Pi stress. The quantification of GUS activity in the transformed plants from both treated and untreated was performed. With the increase in Pi concentration (High Pi stress), the activities of all the promoters’ show decreased GUS expression, however, the GUS activity of transgenic plants under low Pi stress displayed significantly increased GUS activity, compared to the untreated control transgenic seedlings. The result showed that under Low-Pi condition, the pD2 promoter has 4.2 times higher expression when compared to CaMV35S promoter and 1.4 times higher expression than the untreated transgenic control (Figure 6C). In the CaMV35S promoter no significant expression was detected in both high and low Pi conditions. Notably, the pD2 sequence still displayed the highest promoter activities among other promoters under both high and low-Pi conditions.
2.8. Relative GUS expression analysis of EaPHT1;2 promoter using RT-PCR
For qRT-PCR analysis, T1 transgenic events with higher levels of GUS expression were used. The results indicate that pFL have constitute expression with 2.5-fold higher expression than the CaMV35S promoter under normal condition. The GUS expression level increased with Low-Pi treatment and thus findings provided evidence that the EaPHT1;2 gene promoter has strong expression under a Low-Pi stress condition. Therefore, the deletion analysis of the promoter might help in deep understating about the tissue specific stress or minimal promoter region analysis. The expression patterns of five EaPHT1;2 promoters under normal, High and Low-Pi stress were subsequently investigated by qRT-PCR in order to validate the stress-induced expression level of the pFL and five pD1 to 5 deletion promoters in comparison with pCaMV35s promoter. According to the findings, the promoter pFL and pD2 genes had higher level of expression than the pCaMV35s promoter, and other EaPHT1;2 promoters (Figure 6D). The pFL showed enhanced expression under low phosphate, the expression levels were 2.5, and 3.0 folds higher than the pCaMV35s promoter under normal and Pi stress. The expression of pD1 had been up-regulated 4.2 folds under Low-Pi stress, and 2.7 fold under normal condition. In contrast, the promoters without 5’UTR regions, pD3, pD4 and pD5 promoters showed 1.0 to 1.5 fold increased expression under low Pi stresses (Figure 6D). Based on these findings, we hypothesized that the phosphate transporter gene promoter have strong gene expression under Low-Pi condition and in addition this study showed the importance of 5’ UTR region which plays a crucial role in promoter regulatory components and act as the positive enhancer that regulates the expression of the promoter region to diverse abiotic stresses.
3. Discussion
Phosphorus (P) is essential for plant growth, development, and abiotic stress tolerance, and their deficiency can limit crop productivity, affects yield and quality. Plants use various mechanisms to cope with low phosphorus availability, by optimizing phosphorus uptake and usage. To satisfy this purpose, phosphate transporters 1 (PHT1) genes, play a crucial role in phosphorus uptake by transporting inorganic phosphate from the soil, which is extensively studied in several plant species. As far as we know, the functional characterization of PHT1 gene promoter in Sugarcane has not been studied, till date. The first study on genome-wide analysis of PHT gene family in sugarcane was reported by us in 2022 [41], revealed that the gene PHT1;2 has higher level of expression under salinity-stress condition in sugarcane root.
In this study, we validated the EaPHT1;2 gene promoter under Pi stress. The EaPHT1;2 gene was isolated and characterized using various bioinformatic tools. The EaPHT1;2 gene belongs to the high-affinity phosphate transporter shares a significant sequence similarity with other plant PHT1 family transporters, particularly ZmPHT1 (Z. mays), followed by SbPHT1 (S. bicolor) and AtPHT1 (A. thaliana) genes [43]. Sugarcane EaPHT1;2 had the most average length and weight than any of the plant phosphate transporters, with 542 aa and 58.9 kDa. The isoelectric point of EaPHT1;2, is similar to the most PHT1 members, and it's conceivable that they are mostly present in plasma membranes. Based on the results obtained and earlier reports, the majority of PHT1s appear to be membrane-bound, which is essential for ion transport [44]. The multiple sequence alignment of EaPHT1;2 protein with the reference genomes was carried out by utilizing the CLUSTALX programme and as illustrated in Figure 3B, the EaPHT1;2 transporter is identical, with other PHT1 protein which possess the characteristic motif sequence GGDYPLSATIMSE (GGDYPLSATIxSE) [45,46,47]. The secondary structure showed 10 transmembrane domains with a hydrophilic loop in the central part of the domains, 39 phosphorylation sites and one N-glycosylation site [13,24,48,49]. The 3D structure of many plant PHT proteins are still not available for the comparative analysis of the phosphate transporter genes, therefore, according to the homology SWISSMODEL, 7sp5.1.A template was chosen to construct the 3D structure with 92.4% of sequence coverage, followed by the validation, the EaPHT1;2 protein structure was analyzed for the binding sites. Figure 3B illustrates the structure of EaPHT1;2 protein which has 4 Å resolution with six phosphate binding sites facing inwards of the protein. Overall, the amino acid sequence and 3D structure of EaPHT1;2 protein exhibit a general conservation with PnPht1;2, CmPht1;2 and the other reported PHT1 protein [47,49,50].
A 1102 bp of 5’ regulatory region was isolated from EaPHT1;2. The cis-regulatory elements present in the promoter region governs several essential roles including biotic and abiotic stress, hormonal stress, and developmental process regulation etc., [51,52]. The in-silico analysis of the 5’ regulatory region found in gene promoters aids in the understanding of the molecular process behind gene expression by using PLACE and PlantCARE. The examination of EaPHT1;2 promoter region, revealed a wide range of previously recognized elements. The conserved promoter elements like P1BS element are mostly present in all of the Pht1 promoters in one or multiple copies. And in EaPHT1;2 promoter two copies of P1BS element was discovered, which is similar as in TaPH2 and HvPht1;2 [53,54]. P1BS element is a conserved Pi-starvation responsive element, typically found as a conserved signalling route for the Pi-starvation response in plants. The P1BS element acts as the binding site for PHR1 protein, a MYB transcription factor, presumably controlling the Pi-starvation response in plants [55]. The substantial conservation of this candidate motif in other Pi transporters promoters in Arabidopsis, rice, barley, wheat, eggplant, and soy bean indicates that it is functionally conserved and may play a role in boosting root expression under Pi-deficient circumstances [53,55,56,57,58,59,60]. Secondly, a number of root specific-regulatory elements, including ROOTMOTIFTAPOX1, OSE1ROOTNODULE and OSE2ROOTNODULE, and other tissue-specific elements like CACTFTPPCA1, CAATBOX1, GTGANTG10, CCGTCC-box, and CAT-box were also discovered. Another major cis-element investigated intensively in PHT gene promoter, the conserved element next to P1BS is W-Box which positively regulates the Pi-starvation and might be present in one or more copies in their promoter regions. Within EgPHT1 promoter many W-box sequences were found. Similarly, the EaPHT1;2 promoter also have 10 W-Box sequences [60]. Along with the conserved cis-element motifs, the EaPHT1;2 gene promoter contains various stress-responsive elements such as LTRE, MYB, MYC, AuxR, and GATA sites.
Previous study in wheat described that under the response to Pi-starvation, the TaPHT1;2 gene was primarily expressed in the roots of wheat seedlings, and its expression levels were higher in the roots of P-deficiency tolerant wheat varieties [61]. In Arabidopsis, the PHT1 family were expressed at the root-soil interface, that it is involved in phosphate absorption and translocation in plants [26]. In Arabidopsis the Pht1;2 phosphate transporter regulates root-specific gene expression, which is significantly enhanced by Pi shortage [62]. High-level transgenic expression may be delivered to almost all tissues and developmental stages in plants via powerful constitutive promoters. Currently, CaMV35s, Rice Actin (OsAct) and maize ubiquitin (Mubi) are the three mostly widely employed promoters in plant genetic transformation [63,64]. In addition, the stress inducible promoters and tissue or organ specific promoter are now widely studied for desired expression in plants. Associated with the abiotic stress, nutrient stress also generate huge impact in plant growth and development, but the combined abiotic stresses with crop nutrient stress still remains unrevealed [12].
In this study, the EaPHT1;2 promoter was functionally characterized under Pi stress. Previous research discovered that Pht1;2 promoter preferentially induces high expression in the epidermis of Pi-deficient roots [62]. The full-length promoter region of EaPHT1;2 promoter showed highly constitute expression which is relatively 2.5-fold higher expression than CaMV35S promoter in transgenic tobacco plants under normal condition and possess higher expression under Low-Pi stress. The pFL and pD2 are the two promoters displayed higher GUS expression under Low Pi and have 3 and 4.2 fold higher expression, compared to CaMV35S promoter. In contrast to the CaMV35S promoter, the EaPHT1;2 pFL and pD2 promoter segments are susceptible to high and low-Pi stress. Additionally the promoter with 5’UTR regions showed better expression compared to promoter without 5’UTR region under same conditions. Notably, under Pi stress and normal conditions, the pD2 promoter continued to exhibit the greatest promoter activities among all other promoters. The in-silico analysis of the promoter regions suggested that the pFL promoter fragment contains three P1BS elements, 8 E-Box/MYCCONSENSUSAT, and 1 MYBST1 motif. Instead, the pD2 promoter fragment has the highest expression contains only one E-Box and P1BS - element. Whereas, pD1 promoter fragment lack the P1BS –element and has no substantial expression in Pi stress. Previous study in HvPHT1 reported that the mutation of the P1BS motif resulted in total lack of gene induction Pi deprivation [65]. Despite the presence of less number of P1BS-element, the Pi- stress response was unaffected. This implies that the gene induction is not particularly regulated by the P1BS element [66]. These findings suggest that the PIBS-motif in the EaPHT1;2 promoter region, is one of the significant regulator (despite of their number) responsible for the Pi stress, and whereas various cis elements situated across the promoter region may also govern the Pi response. Therefore, pFL and pD2 promoter of EaPHT1;2 gene can be the efficient promoter for the development of transgene expression in sugarcane and other crops under Pi stress conditions.
4. Materials and Methods
4.1. Bacteria and Plant Material
Escherichia coli strain TOP10 was used for cloning and propagation of the plasmids, and Agrobacterium tumefaciens strain EHA105 was used for plant transformation. E. coli and A. tumefaciens were grown in Luria-Bertani (LB) and Yeast Extract Peptone (YEP) media at 37°C and 28°C with appropriate antibiotics, respectively [67]. Tobacco seeds (Nicotiana benthamiana var. Bhagyalakshmi) were grown under 16 h light/8 h dark at 25 ± 1°C and maintained for plant transformation [68]. The six-month-old Erianthus arundinaceus plant was used for the study. The total genomic DNA was isolated from young leaf tissues (1mg) of E. arundinaceus without midrib in liquid nitrogen using the CTAB procedure [69].
4.2. EaPHT1;2 Gene isolation and cloning
E. arundinaceus (Ea) Phosphate transporter 1;2 (PHT1;2) gene was isolated from genomic DNA. Based on our previous study [41,42] and Zea mays PHT1 genes as a reference, the gene-specific primer pair was synthesized. The polymerase chain reaction was performed with Platinum Taq DNA polymerase (Invitrogen, USA) on Master-cycler ® nexus (Eppendorf, Germany), and the thermal cycling conditions adopted were as follows: 94°C denaturation for 5 min, followed by 30 cycles of 94 °C for 10 s, 57 °C for 30 s, 72 °C for 1min, and a final extension of 72 °C for 10 min. The resulting PCR product was ligated into a linearized pTZ57R/T-TA cloning vector (Thermo Fisher Scientific, USA), and transformed into E. coli TOP10 competent cells grown under ampicillin selection marker. Plasmids isolated were confirmed or the presence of target gene and further confirmed by Sanger sequencing method.
4.3. Bioinformatics characterization of EaPHT1;2 gene
The EaPHT1;2 nucleotide sequence data was deduced using VecScreen (https://www.ncbi.nlm.nih.gov/tools/vecscreen/, (accessed on 23 November 2022,)) and translated to its corresponding protein primary sequence using the ExPasy translation tool (https://web.expasy.org/translate/, (accessed on 23 November 2022)) [70] and the most suitable reading frame was selected. The consensus sequence was checked for their homology with reported sequences presenting nucleotide database using BLASTn and BLASTx programs of NCBI BLAST (https://blast.ncbi.nlm.nih.gov/Blast.cgi, (accessed on 27 November 2022)) [71]. Multiple Sequence Alignment tool was carried out using EBI CLUSTALW [72] and the evolutionary tree constructed by using the neighbour-joining clustering in MEGA-X [73]. To compute the physicochemical properties of EaPHT1;2 gene ProtParam tool on the ExPASy server was used [70]. The SMART tool (http://smart.embl-heidelberg.de/, (accessed on 28 November 2022)) [74], Pfam server (http://pfam.xfam.org/, (accessed on 02 December 2022)) [75], InterPRO (https://www.ebi.ac.uk/interpro/, (accessed on 02 December 2022)) [76], PROSITE (https://prosite.expasy.org/,(accessed on 02 December 2022) ) [77] and TMHMM 2.0 tool (http://www.cbs.dtu.dk/services/TMHMM/, (accessed on 05 December 2022)) [78] were employed to determine the domains/motif and transmembrane domain of the EaPHT1;2 protein. The secondary structures were analyzed by SOPMA (https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=/NPSA/npsa_sopma.html, (accessed on 05 December 2022)) [79] and PSIPRED (http://bioinf.cs.ucl.ac.uk/psipred/, (accessed on 05 December 2022)) [80]. The three-dimensional structures of EaPHT1;2 proteins were predicted using Phyre2 software (http://www.sbg.bio.ic.ac.uk/phyre2/, (accessed on 05 December 2022)) [81] and Swiss-Model (https://swissmodel.expasy.org/, (accessed on 05 December 2022)) [81]. The models obtained were subjected to PROCHECK [51] and ProSA (https://prosa.services.came.sbg.ac.at, (accessed on 05 December 2022)) [82] and were visualized using UCSF Chimera. The STRING analysis (http://string-db.org/, (accessed on 06 December 2022)) [83] and subcellular localization using DeepLoc1.0. (https://services.healthtech.dtu.dk/services/DeepLoc-1.0/, (accessed on 06 December 2022)) [84] were carried out. Signal peptides, Hydropathy plots, Post-translation modification sites analysis were also carried out using SignalP (http://www.cbs.dtu.dk/services/SignalP/, (accessed on 06 December 2022)) [85], ProtScale (http://web.expasy.org/protscale/, (accessed on 06 December 2022)) [70], phosphorylation (https://services.healthtech.dtu.dk/services/NetPhos-3.1/, (accessed on 10 December 2022)) [86], and glycosylation (https://services.healthtech.dtu.dk/services/NetNGlyc-1.0/, (accessed on 10 December 2022)) [87] sites prediction, respectively [41].
4.4. Isolation of EaPHT1;2 promoter region using a genome walking method
The random amplification of genome ends (RAGE) approach was used to isolate the promoter of the PHT1;2 gene from E. arundinaceus. The genomic DNA was isolated from root tissues of E. arundinaceus using a QIAGEN DNeasy plant mini kit and restricted with DraI, EcoRV, HincII, PvuII, and SspI, to create five genome-walker libraries. Primary PCR amplifications were carried out using each of the libraries as a template and antisense gene-specific primer (GSP I) from the 5′ ends of the ‘coding region of the EaPHT1;2 gene sequence, as well as primary adapter primer ASP I (Table S1), and secondary PCR amplifications were carried out using the nested adapter primer ASP II and a nested EaPHT1;2 gene-specific primer (GPS II) [66,88,89,90]. All PCR amplifications were performed with the Taq polymerase enzyme and cloned in the pTZ57R/T-TA cloning vector and mobilized the ligated product into E. coli TOP10 [67]. Finally, the recombinants carrying the promoter region was screened and plasmid was isolated and sequenced.
4.5. Bioinformatics analysis of the promoter regions
Nucleotide sequences obtained after sequencing were analyzed using NCBI BLAST analysis and the resulting EaPHT1;2 promoter sequence was further characterized using various tools and database search programs. TSS and TATA-Box were analyzed using an online Neural Network Promoter Prediction (NNPP version 2.2) (BDGP: Neural Network Promoter Prediction) (https://fruitfly.org/seq_tools/promoter.html, (accessed on 12 December 2022)) [91] and the TSSPlant (http://www.softberry.com/, (accessed on 12 December 2022)) [92]. The comparative analysis were carried out using PlantPAN 3.0 online tool (http://plantpan3.itps.ncku.edu.tw/, (accessed on 12 December 2022)) [93] and MEME tools (https://meme-suite.org/meme/tools/meme, (accessed on 12 December 2022)) (https://meme-suite.org/tools/meme, (accessed on 15 December 2022 )) [94]. Using PLACE (https://www.dna.affrc.go.jp/PLACE/, (accessed on 15 December 2022 )) [95] and PLANT CARE (https://bioinformatics.psb.ugent.be/webtools/plantcare/html/, (accessed on 15 December 2022 )) [96] web servers, the putative cis-regulatory elements were determined.
4.6. Vector Construction and Tobacco Transformation
The promoter region of 1.1Kb was cloned in the plant transformation binary vector pCAMBIA1305.1 using the promoter region-specific primers. The full-length and a serious of deletion with and without 5’UTR regions were generated using the region-specific truncation primers (Table S1). All the EaPHT1;2 promoter constructs were amplified using each region-specific truncation primers and replaced with the CaMV35S promoter region which drives the GUS gene expression in pCAMBIA1305.1 vector [89,97]. The constructs were transformed into E. coli TOP10 and plasmids were confirmed by 5’ HindIII and 3’ NcoI restriction analysis and sequencing. Further the confirmed plasmids were mobilized into A. tumefaciens strain EHA105 for plant transformation. Consequently, the plasmids were introduced into tobacco by following the transformation procedures as previously described [68,90].
4.7. Low and High -Pi Treatments
The putative tobacco transgenic events of the independent T0 hygromycin-resistant plants expressing GUS were selected and self-fertilized to produce T1 seeds. The seeds were aseptically sown in MS medium containing 25 mg/l hygromycin and 20 days old-seedlings used for the Pi stress analysis [97]. The seedlings of each constructs were treated with 0.1mM (low Pi) and 1mM KH2PO4 (high Pi) for 24 hrs and the samples were collected, frozen using liquid N2, and stored at -80°C for further studies [98].
4.8. Histochemical and Fluorimetric β-glucuronidase (GUS) assays
The histochemical and fluorimetric β-glucuronidase assay for GUS reporter gene expression was performed with some modifications as described in Jefferson [99]. The leaf, stem, and root of the putative transgenic events and T1 seedlings were utilized for GUS assays. For histochemical assay, the samples were dipped in GUS staining solution, and incubated for 12-16 hrs at 37°C. After staining, sections were rinsed in 70% of ethanol to remove chlorophyll and photographed under Leica Microscope (Leica, Germany). The quantitative fluorimetric GUS assays, the total protein was isolated and quantified according to Bradford using bovine serum albumin (BSA) standard [100]. To quantify the GUS activity, the 4-MUG (4-methylumbelliferyl b-d-glucuronide) from Sigma was used as the substrate. The fluorescence was detected using the microplate spectrofluorometer at 365 nm and 455 nm, excitation and emission wavelength, respectively. GUS enzyme activity was presented as pmols of 4-methylumbelliferone (MU), hydrolyzed per milligram of soluble protein per minute.
4.9. qRT-PCR analysis
Total RNA was extracted from control and stress leaves of putative transgenic tobacco plants using the TRIzol method [101] and then treated with RNase-free DNase. The quality and quantity of the RNA was checked on 1.5% agarose gel and NanoDrop method (Thermo Fisher Scientific, US). The cDNA was synthesized by utilizing the RevertAid First strand cDNA synthesis kit from Thermo Fisher Scientific and according to the instruction mentioned in protocol manual. The IDT tools were used to design the primers for qRT PCR (Table S1). Three technical and biological replicates were used for expression analysis and the relative expression was determined by using 2−∆∆Ct method [102]. The native control, used in this study, was the housekeeping gene Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) [103,104,105]. The qPCR reaction conditions explained by Narayan et al. [106,107,108] were followed.
5. Conclusions
Phosphorus (P) is indeed crucial for plant growth and development. It also, regulates physiological responses and enhances abiotic stress tolerance in plants. Its deficiency can limit crop productivity, affecting both yield and quality. Phosphate transporters facilitate the uptake of phosphorus from the soil. Phosphate transporter 1 (PHT1) the widely studied P transporter, and their promoter were validated in various crop plants. But, the PHT1;2 gene promoter has been reported in very few plant species. The present study successfully isolated and functionally characterized the EaPHT1;2 gene promoter under Pi stress condition. The full-length and truncated deletions promoters were validated using the GUS reporter gene expression. The strong and short promoter of EaPHT1;2 with 374 bp shows significant expression in low P condition and in comparison, with the traditional CaMV35S promoter, EaPHT1;2 shows three-fold expression in normal and two-fold expression under Pi stress condition. Therefore, the EaPHT1;2 promoter could be exploited for the development of transgenic crops that produce good yields under P deficient conditions.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1. The nucleotide sequence of Erianthus arundinaceus Phosphate transporter 1;2 (EaPHT1;2). (A) The full-length CDS sequence of the EaPHT1;2 gene; (B) The ExPasy-translated protein sequence of the EaPHT1;2; Figure S2. Physicochemical parameters of the selected protein: (A) Atomic composition of EaPHT1;2 protein, (B) Amino acid composition of EaPHT1;2 protein; Figure S3. Post-translation modification sites of EaPHT1: (A) Phosphorylation sites, (B) Glycosylation site. The amino acids are represented in different lines at their position in the sequence and the scores higher than the threshold value are considered for posttranslational modification; Figure S4. Protein structure prediction and evaluation; (A) Secondary elements in the EaPHT1 protein predicted using the SOPMA tool. (B) Structural evaluation of EaPHT1;2 obtained through Phyre2 model, (B) EaPHT1;2 obtained through Swiss-Modelling; Figure S5. Protein-protein Interaction analysis of EaPHT1;2 protein: (A) PPI of EaPHT1;2 with ZmPHT1 protein, (B) PPI of EaPHT1;2 with AtPHT protein, (C) PPI of EaPHT1;2 with SbPHT protein; Figure S6. Subcellular Localization analysis: (A) EaPHT1;2 protein localization using online tool DeepLoc analysis (B) QuickGo Term analysis of EaPHT1;2 protein; Figure S7. Comparative In-silico analysis of EaPHT1;2 promoter: (A) The EaPHT1;2 promoter conserved regions with ZmPHT1 and (B) One conserved region was ShPHT1;2 promoter regions; Table S1. List of primer sequences used in this study for EaPHT1-2 gene isolation, promoter isolation, promoter deletion constructs, and analysis; Table S2. Physicochemical characteristics of the EaPHT1;2 protein; Table S3. Subcellular localization of EaPHT1-2 gene in different locations.
Author Contributions
Conceptualization, C.A. and M.N.; methodology, M.N.; H.K.M. and C.A.; software, M..N; C.M.; R.V. and M.M., validation, M.N.; H.K.M. and C.M.; formal analysis, M.N.; C.A. and M.A.; resources, C.A.; H.K.M. and G.H.; data curation, M.N. and C.A.; writing—original draft preparation, M.N.; writing—review and editing, C.A.; S.S.K.; G.H.; M.A. and M.M.; visualization M.N.; R.V. and C.A.; supervision, project administration, C.A.; funding acquisition, C.A. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
The data used in this study are available in the NCBI database and as supplementary file of this manuscript.
Acknowledgments
We gratefully acknowledge the Indian Council of Agricultural Research (ICAR)-Sugarcane Breeding Institute (SBI) for supporting the current research. The authors also thank Anilkumar C, Scientist, NRRI, Cuttack for his help in editing the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gu, M.; Chen, A.; Sun, S.; Xu, G. Complex Regulation of Plant Phosphate Transporters and the Gap between Molecular Mechanisms and Practical Application: What Is Missing? Molecular Plant 2016, 9, 396–416. [Google Scholar] [CrossRef]
- Srivastava, S.; Upadhyay, M.K.; Srivastava, A.K.; Abdelrahman, M.; Suprasanna, P.; Tran, L.-S.P. Cellular and Subcellular Phosphate Transport Machinery in Plants. International Journal of Molecular Sciences 2018, 19, 1914. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, F.; Lu, H.; Liu, Y.; Mao, C. Phosphate Uptake and Transport in Plants: An Elaborate Regulatory System. Plant and Cell Physiology 2021, 62, 564–572. [Google Scholar] [CrossRef]
- Crombez, H.; Motte, H.; Beeckman, T. Tackling Plant Phosphate Starvation by the Roots. Developmental Cell 2019, 48, 599–615. [Google Scholar] [CrossRef]
- Vilaça De Vasconcelos, M.J.; Fontes Figueiredo, J.E.; De Oliveira, M.F.; Schaffert, R.E.; Raghothama, K.G. PLANT PHOSPHORUS USE EFFICIENCY IN ACID TROPICAL SOIL. RBMS 2022, 21. [Google Scholar] [CrossRef]
- Balemi, T.; Negisho, K. Management of Soil Phosphorus and Plant Adaptation Mechanisms to Phosphorus Stress for Sustainable Crop Production: A Review. Journal of soil science and plant nutrition 2012, 12, 547–562. [Google Scholar] [CrossRef]
- Akpinar, B.A.; Avsar, B.; Lucas, S.J.; Budak, H. Plant Abiotic Stress Signaling. Plant Signal Behav 2012, 7, 1450–1455. [Google Scholar] [CrossRef] [PubMed]
- Sachdev, S.; Ansari, S.A.; Ansari, M.I.; Fujita, M.; Hasanuzzaman, M. Abiotic Stress and Reactive Oxygen Species: Generation, Signaling, and Defense Mechanisms. Antioxidants (Basel) 2021, 10, 277. [Google Scholar] [CrossRef]
- Satheesh, V.; Tahir, A.; Li, J.; Lei, M. Plant Phosphate Nutrition: Sensing the Stress. Stress Biology 2022, 2, 16. [Google Scholar] [CrossRef]
- Yu, X.; Keitel, C.; Dijkstra, F.A. Global Analysis of Phosphorus Fertilizer Use Efficiency in Cereal Crops. Global Food Security 2021, 29, 100545. [Google Scholar] [CrossRef]
- Elbasiouny, H.; El-Ramady, H.; Elbehiry, F.; Rajput, V.D.; Minkina, T.; Mandzhieva, S. Plant Nutrition under Climate Change and Soil Carbon Sequestration. Sustainability 2022, 14, 914. [Google Scholar] [CrossRef]
- Bechtaoui, N.; Rabiu, M.K.; Raklami, A.; Oufdou, K.; Hafidi, M.; Jemo, M. Phosphate-Dependent Regulation of Growth and Stresses Management in Plants. Frontiers in Plant Science 2021, 12. [Google Scholar] [CrossRef]
- Nussaume, L.; Kanno, S.; Javot, H.; Marin, E.; Nakanishi, T.M.; Thibaud, M.-C. Phosphate Import in Plants: Focus on the PHT1 Transporters. Frontiers in Plant Science 2011, 2. [Google Scholar] [CrossRef]
- Zhang, Y.; Hu, L.; Yu, D.; Xu, K.; Zhang, J.; Li, X.; Wang, P.; Chen, G.; Liu, Z.; Peng, C.; et al. Integrative Analysis of the Wheat PHT1 Gene Family Reveals A Novel Member Involved in Arbuscular Mycorrhizal Phosphate Transport and Immunity. Cells 2019, 8, 490. [Google Scholar] [CrossRef] [PubMed]
- Rae, A.L.; Cybinski, D.H.; Jarmey, J.M.; Smith, F.W. Characterization of Two Phosphate Transporters from Barley; Evidence for Diverse Function and Kinetic Properties among Members of the Pht1 Family. Plant Mol Biol 2003, 53, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Chang, X.-J.; Ye, Y.; Xie, W.-B.; Wu, P.; Lian, X.-M. Comprehensive Sequence and Whole-Life-Cycle Expression Profile Analysis of the Phosphate Transporter Gene Family in Rice. Molecular Plant 2011, 4, 1105–1122. [Google Scholar] [CrossRef] [PubMed]
- Fan, C.; Wang, X.; Hu, R.; Wang, Y.; Xiao, C.; Jiang, Y.; Zhang, X.; Zheng, C.; Fu, Y.-F. The Pattern of Phosphate Transporter 1 Genes Evolutionary Divergence in Glycine maxL. BMC Plant Biol 2013, 13, 48. [Google Scholar] [CrossRef]
- Ceasar, S.A.; Hodge, A.; Baker, A.; Baldwin, S.A. Phosphate Concentration and Arbuscular Mycorrhizal Colonisation Influence the Growth, Yield and Expression of Twelve PHT1 Family Phosphate Transporters in Foxtail Millet (Setaria Italica). PLOS ONE 2014, 9, e108459. [Google Scholar] [CrossRef]
- Chen, A.; Chen, X.; Wang, H.; Liao, D.; Gu, M.; Qu, H.; Sun, S.; Xu, G. Genome-Wide Investigation and Expression Analysis Suggest Diverse Roles and Genetic Redundancy of Pht1 Family Genes in Response to Pi Deficiency in Tomato. BMC Plant Biol 2014, 14, 61. [Google Scholar] [CrossRef]
- Zhang, C.; Meng, S.; Li, M.; Zhao, Z. Genomic Identification and Expression Analysis of the Phosphate Transporter Gene Family in Poplar. Frontiers in Plant Science 2016, 7. [Google Scholar] [CrossRef]
- Sun, T.; Li, M.; Shao, Y.; Yu, L.; Ma, F. Comprehensive Genomic Identification and Expression Analysis of the Phosphate Transporter (PHT) Gene Family in Apple. Frontiers in Plant Science 2017, 8. [Google Scholar] [CrossRef]
- Teng, W.; Zhao, Y.-Y.; Zhao, X.-Q.; He, X.; Ma, W.-Y.; Deng, Y.; Chen, X.-P.; Tong, Y.-P. Genome-Wide Identification, Characterization, and Expression Analysis of PHT1 Phosphate Transporters in Wheat. Frontiers in Plant Science 2017, 8. [Google Scholar] [CrossRef]
- Wang, J.; Yang, Y.; Liao, L.; Xu, J.; Liang, X.; Liu, W. Genome-Wide Identification and Functional Characterization of the Phosphate Transporter Gene Family in Sorghum. Biomolecules 2019, 9, 670. [Google Scholar] [CrossRef]
- Zhou, J.; Lu, M.; Zhang, C.; Qu, X.; Liu, Y.; Yang, J.; Yuan, J. Isolation and Functional Characterisation of the PHT1 Gene Encoding a High-Affinity Phosphate Transporter in Camellia Oleifera. The Journal of Horticultural Science and Biotechnology 2020, 95, 553–564. [Google Scholar] [CrossRef]
- Lhamo, D.; Shao, Q.; Tang, R.; Luan, S. Genome-Wide Analysis of the Five Phosphate Transporter Families in Camelina Sativa and Their Expressions in Response to Low-P. International Journal of Molecular Sciences 2020, 21, 8365. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Liu, J.; Li, Y.; Zhang, J.; Li, S.; An, Y.; Hu, T.; Yang, P. Functional Analysis of the Phosphate Transporter Gene MtPT6 From Medicago Truncatula. Frontiers in Plant Science 2021, 11. [Google Scholar] [CrossRef] [PubMed]
- Rubio, V.; Linhares, F.; Solano, R.; Martín, A.C.; Iglesias, J.; Leyva, A.; Paz-Ares, J. A Conserved MYB Transcription Factor Involved in Phosphate Starvation Signaling Both in Vascular Plants and in Unicellular Algae. Genes Dev. 2001, 15, 2122–2133. [Google Scholar] [CrossRef]
- Zhou, J.; Jiao, F.; Wu, Z.; Li, Y.; Wang, X.; He, X.; Zhong, W.; Wu, P. OsPHR2 Is Involved in Phosphate-Starvation Signaling and Excessive Phosphate Accumulation in Shoots of Plants. Plant Physiology 2008, 146, 1673–1686. [Google Scholar] [CrossRef]
- Bustos, R.; Castrillo, G.; Linhares, F.; Puga, M.I.; Rubio, V.; Pérez-Pérez, J.; Solano, R.; Leyva, A.; Paz-Ares, J. A Central Regulatory System Largely Controls Transcriptional Activation and Repression Responses to Phosphate Starvation in Arabidopsis. PLOS Genetics 2010, 6, e1001102. [Google Scholar] [CrossRef] [PubMed]
- Sega, P.; Pacak, A. Plant PHR Transcription Factors: Put on A Map. Genes (Basel) 2019, 10, 1018. [Google Scholar] [CrossRef]
- Sega, P.; Kruszka, K.; Bielewicz, D.; Karlowski, W.; Nuc, P.; Szweykowska-Kulinska, Z.; Pacak, A. Pi-Starvation Induced Transcriptional Changes in Barley Revealed by a Comprehensive RNA-Seq and Degradome Analyses. BMC Genomics 2021, 22, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Yi, K.; Wu, Z.; Zhou, J.; Du, L.; Guo, L.; Wu, Y.; Wu, P. OsPTF1, a Novel Transcription Factor Involved in Tolerance to Phosphate Starvation in Rice. Plant Physiology 2005, 138, 2087–2096. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-F.; Li, L.-Q.; Xu, Q.; Kong, Y.-H.; Wang, H.; Wu, W.-H. The WRKY6 Transcription Factor Modulates PHOSPHATE1 Expression in Response to Low Pi Stress in Arabidopsis. The Plant Cell 2009, 21, 3554–3566. [Google Scholar] [CrossRef] [PubMed]
- Karthikeyan, A.S.; Varadarajan, D.K.; Mukatira, U.T.; D’Urzo, M.P.; Damsz, B.; Raghothama, K.G. Regulated Expression of Arabidopsis Phosphate Transporters. Plant Physiology 2002, 130, 221. [Google Scholar] [CrossRef] [PubMed]
- Baek, D.; Chun, H.J.; Yun, D.-J.; Kim, M.C. Cross-Talk between Phosphate Starvation and Other Environmental Stress Signaling Pathways in Plants. Mol Cells 2017, 40, 697–705. [Google Scholar] [CrossRef] [PubMed]
- Yong-Villalobos, L.; Cervantes-Pérez, S.A.; Gutiérrez-Alanis, D.; Gonzáles-Morales, S.; Martínez, O.; Herrera-Estrella, L. Phosphate Starvation Induces DNA Methylation in the Vicinity of Cis-Acting Elements Known to Regulate the Expression of Phosphate–Responsive Genes. Plant Signaling & Behavior 2016, 11, e1173300. [Google Scholar] [CrossRef]
- Li, Y.; Wang, X.; Zhang, H.; Wang, S.; Ye, X.; Shi, L.; Xu, F.; Ding, G. Molecular Identification of the Phosphate Transporter Family 1 (PHT1) Genes and Their Expression Profiles in Response to Phosphorus Deprivation and Other Abiotic Stresses in Brassica Napus. PLOS ONE 2019, 14, e0220374. [Google Scholar] [CrossRef]
- Desalegn, B.; Kebede, E.; Legesse, H.; Fite, T. Sugarcane Productivity and Sugar Yield Improvement: Selecting Variety, Nitrogen Fertilizer Rate, and Bioregulator as a First-Line Treatment. Heliyon 2023, 9, e15520. [Google Scholar] [CrossRef]
- Fernandes, G.C.; Rosa, P.A.L.; Jalal, A.; Oliveira, C.E. da S. ; Galindo, F.S.; Viana, R. da S.; De Carvalho, P.H.G.; Silva, E.C. da; Nogueira, T.A.R.; Al-Askar, A.A.; et al. Technological Quality of Sugarcane Inoculated with Plant-Growth-Promoting Bacteria and Residual Effect of Phosphorus Rates. Plants 2023, 12, 2699. [Google Scholar] [CrossRef]
- Soltangheisi, A.; Withers, P.J.A.; Pavinato, P.S.; Cherubin, M.R.; Rossetto, R.; Do Carmo, J.B.; da Rocha, G.C.; Martinelli, L.A. Improving Phosphorus Sustainability of Sugarcane Production in Brazil. GCB Bioenergy 2019, 11, 1444–1455. [Google Scholar] [CrossRef]
- Murugan, N.; Palanisamy, V.; Channappa, M.; Ramanathan, V.; Ramaswamy, M.; Govindakurup, H.; Chinnaswamy, A. Genome-Wide In Silico Identification, Structural Analysis, Promoter Analysis, and Expression Profiling of PHT Gene Family in Sugarcane Root under Salinity Stress. Sustainability 2022, 14, 15893. [Google Scholar] [CrossRef]
- Murugan, N.; Kumar, R.; Pandey, S.K.; Dhansu, P.; Chennappa, M.; Nallusamy, S.; Govindakurup, H.; Chinnaswamy, A. In Silico Dissection of Regulatory Regions of PHT Genes from Saccharum Spp. Hybrid and Sorghum Bicolor and Expression Analysis of PHT Promoters under Osmotic Stress Conditions in Tobacco. Sustainability 2023, 15, 1048. [Google Scholar] [CrossRef]
- Ren, F.; Zhao, C.-Z.; Liu, C.-S.; Huang, K.-L.; Guo, Q.-Q.; Chang, L.-L.; Xiong, H.; Li, X.-B. A Brassica Napus PHT1 Phosphate Transporter, BnPht1;4, Promotes Phosphate Uptake and Affects Roots Architecture of Transgenic Arabidopsis. Plant Mol Biol 2014, 86, 595–607. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Li, G.; Sun, Z.; Chen, Y.; Guo, W.; Li, Y.; Chen, Y.; Yang, J.; Hou, H. Identification, Structure Analysis, and Transcript Profiling of Phosphate Transporters under Pi Deficiency in Duckweeds. International Journal of Biological Macromolecules 2021, 188, 595–608. [Google Scholar] [CrossRef] [PubMed]
- Karandashov, V.; Bucher, M. Symbiotic Phosphate Transport in Arbuscular Mycorrhizas. Trends Plant Sci 2005, 10, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.; Hu, J.; Sun, S.; Xu, G. Conservation and Divergence of Both Phosphate- and Mycorrhiza-Regulated Physiological Responses and Expression Patterns of Phosphate Transporters in Solanaceous Species. New Phytologist 2007, 173, 817–831. [Google Scholar] [CrossRef]
- Cao, G.-H.; Li, Z.-D.; Wang, X.-F.; Zhang, X.; Zhao, R.-H.; Gu, W.; Chen, D.; Yu, J.; He, S. Phosphate Transporters, PnPht1;1 and PnPht1;2 from Panax Notoginseng Enhance Phosphate and Arsenate Acquisition. BMC Plant Biology 2020, 20, 124. [Google Scholar] [CrossRef]
- Wu, Z.; Zhao, J.; Gao, R.; Hu, G.; Gai, J.; Xu, G.; Xing, H. Molecular Cloning, Characterization and Expression Analysis of Two Members of the Pht1 Family of Phosphate Transporters in Glycine Max. PLoS One 2011, 6, e19752. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Su, J.; Stephen, G.K.; Wang, H.; Song, A.; Chen, F.; Zhu, Y.; Chen, S.; Jiang, J. Overexpression of Phosphate Transporter Gene CmPht1;2 Facilitated Pi Uptake and Alternated the Metabolic Profiles of Chrysanthemum Under Phosphate Deficiency. Front Plant Sci 2018, 9, 686. [Google Scholar] [CrossRef]
- Pedersen, B.P.; Kumar, H.; Waight, A.B.; Risenmay, A.J.; Roe-Zurz, Z.; Chau, B.H.; Schlessinger, A.; Bonomi, M.; Harries, W.; Sali, A.; et al. Crystal Structure of a Eukaryotic Phosphate Transporter. Nature 2013, 496, 533–536. [Google Scholar] [CrossRef]
- Kaur, A.; Pati, P.K.; Pati, A.M.; Nagpal, A.K. In-Silico Analysis of Cis-Acting Regulatory Elements of Pathogenesis-Related Proteins of Arabidopsis Thaliana and Oryza Sativa. PLOS ONE 2017, 12, e0184523. [Google Scholar] [CrossRef] [PubMed]
- Lata, C.; Prasad, M. Role of DREBs in Regulation of Abiotic Stress Responses in Plants. Journal of Experimental Botany 2011, 62, 4731–4748. [Google Scholar] [CrossRef] [PubMed]
- Tittarelli, A.; Milla, L.; Vargas, F.; Morales, A.; Neupert, C.; Meisel, L.A.; Salvo-G, H.; Peñaloza, E.; Muñoz, G.; Corcuera, L.J.; et al. Isolation and Comparative Analysis of the Wheat TaPT2 Promoter: Identification in Silico of New Putative Regulatory Motifs Conserved between Monocots and Dicots. Journal of Experimental Botany 2007, 58, 2573–2582. [Google Scholar] [CrossRef] [PubMed]
- Sobkowiak, L.; Bielewicz, D.; Malecka, E.; Jakobsen, I.; Albrechtsen, M.; Pacak, A. The Role of the P1BS Element Containing Promoter-Driven Genes in Pi Transport and Homeostasis in Plants. Frontiers in plant science 2012, 3, 58. [Google Scholar] [CrossRef]
- Schünmann, P.H.D.; Richardson, A.E.; Smith, F.W.; Delhaize, E. Characterization of Promoter Expression Patterns Derived from the Pht1 Phosphate Transporter Genes of Barley (Hordeum Vulgare L. ). Journal of Experimental Botany 2004, 55, 855–865. [Google Scholar] [CrossRef]
- Koyama, T.; Ono, T.; Shimizu, M.; Jinbo, T.; Mizuno, R.; Tomita, K.; Mitsukawa, N.; Kawazu, T.; Kimura, T.; Ohmiya, K.; et al. Promoter of Arabidopsis Thaliana Phosphate Transporter Gene Drives Root-Specific Expression of Transgene in Rice. Journal of Bioscience and Bioengineering 2005, 99, 38–42. [Google Scholar] [CrossRef]
- Chen, A.; Gu, M.; Sun, S.; Zhu, L.; Hong, S.; Xu, G. Identification of Two Conserved Cis-Acting Elements, MYCS and P1BS, Involved in the Regulation of Mycorrhiza-Activated Phosphate Transporters in Eudicot Species. New Phytologist 2011, 189, 1157–1169. [Google Scholar] [CrossRef]
- Li, L.H.; Guo, N.; Wu, Z.Y.; Zhao, J.M.; Sun, J.T.; Wang, X.T.; Xing, H. P1BS, a Conserved Motif Involved in Tolerance to Phosphate Starvation in Soybean. Genet. Mol. Res. 2015, 14, 9384–9394. [Google Scholar] [CrossRef]
- Peñaloza, E.; Santiago, M.; Cabrera, S.; Muñoz, G.; Corcuera, L.J.; Silva, H. Characterization of the High-Affinity Phosphate Transporter PHT1;4 Gene Promoter of Arabidopsis Thaliana in Transgenic Wheat. Biologia plant. 2017, 61, 453–462. [Google Scholar] [CrossRef]
- Zhao, Q.; Luo, Z.; Chen, J.; Jia, H.; Ai, P.; Chen, A.; Li, Y.; Xu, G. Characterization of Two Cis-Acting Elements, P1BS and W-Box, in the Regulation of OsPT6 Responsive to Phosphors Deficiency. Plant Growth Regul 2021, 93, 303–310. [Google Scholar] [CrossRef]
- Miao, J.; Sun, J.; Liu, D.; Li, B.; Zhang, A.; Li, Z.; Tong, Y. Characterization of the Promoter of Phosphate Transporter TaPHT1. 2 Differentially Expressed in Wheat Varieties. Journal of Genetics and Genomics 2009, 36, 455–466. [Google Scholar] [CrossRef] [PubMed]
- Mudge, S.R.; Smith, F.W.; Richardson, A.E. Root-Specific and Phosphate-Regulated Expression of Phytase under the Control of a Phosphate Transporter Promoter Enables Arabidopsis to Grow on Phytate as a Sole P Source. Plant Science 2003, 165, 871–878. [Google Scholar] [CrossRef]
- Hou, J.; Jiang, P.; Qi, S.; Zhang, K.; He, Q.; Xu, C.; Ding, Z.; Zhang, K.; Li, K. Isolation and Functional Validation of Salinity and Osmotic Stress Inducible Promoter from the Maize Type-II H+-Pyrophosphatase Gene by Deletion Analysis in Transgenic Tobacco Plants. PLOS ONE 2016, 11, e0154041. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Jing, R.; Mao, X. Functional Characterization of TaSnRK2. 8 Promoter in Response to Abiotic Stresses by Deletion Analysis in Transgenic Arabidopsis. Front Plant Sci 2017, 8, 1198. [Google Scholar] [CrossRef]
- Schünmann, P.H.D.; Richardson, A.E.; Vickers, C.E.; Delhaize, E. Promoter Analysis of the Barley Pht1;1 Phosphate Transporter Gene Identifies Regions Controlling Root Expression and Responsiveness to Phosphate Deprivation. Plant Physiol 2004, 136, 4205–4214. [Google Scholar] [CrossRef]
- Li, W.; Liu, B.; Zhao, M.; Zhang, K.; He, Q.; Zhao, X.; Cheng, W.; Ding, Z.; Zhang, K.; Li, K. Isolation and Characterization of a 295-Bp Strong Promoter of Maize High-Affinity Phosphate Transporter Gene ZmPht1; 5 in Transgenic Nicotiana Benthamiana and Zea Mays. Planta 2020, 251, 106. [Google Scholar] [CrossRef]
- Kasirajan, L.; Valiyaparambth, R.; Kamaraj, K.; Sebastiar, S.; Hoang, N.V.; Athiappan, S.; Srinivasavedantham, V.; Subramanian, K. Deep Sequencing of Suppression Subtractive Library Identifies Differentially Expressed Transcripts of Saccharum Spontaneum Exposed to Salinity Stress. Physiologia Plantarum 2022, 174, e13645. [Google Scholar] [CrossRef]
- Horsch, R.B.; Fry, J.E.; Hoffmann, N.L.; Wallroth, M.; Eichholtz, D.; Rogers, S.G.; Fraley, R.T. A Simple and General Method for Transferring Genes into Plants. Science 1985, 227, 1229–1231. [Google Scholar] [CrossRef]
- Doyle, J. DNA Protocols for Plants. In Molecular Techniques in Taxonomy; Hewitt, G.M., Johnston, A.W.B., Young, J.P.W., Eds.; NATO ASI Series; Springer: Berlin, Heidelberg, 1991; pp. 283–293 ISBN 978-3-642- 83962-7.
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Duvaud, S.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. Protein Identification and Analysis Tools on the ExPASy Server. In The Proteomics Protocols Handbook; Walker, J.M., Ed.; Springer Protocols Handbooks; Humana Press: Totowa, NJ, 2005; pp. 571–607 ISBN 978-1-59259- 890-8.
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J Mol Biol 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the Sensitivity of Progressive Multiple Sequence Alignment through Sequence Weighting, Position-Specific Gap Penalties and Weight Matrix Choice. Nucleic Acids Research 1994, 22, 4673–4680. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol Biol Evol 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Letunic, I.; Khedkar, S.; Bork, P. SMART: Recent Updates, New Developments and Status in 2020. Nucleic Acids Research 2021, 49, D458–D460. [Google Scholar] [CrossRef]
- Finn, R.D.; Bateman, A.; Clements, J.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Heger, A.; Hetherington, K.; Holm, L.; Mistry, J.; et al. Pfam: The Protein Families Database. Nucleic Acids Research 2014, 42, D222–D230. [Google Scholar] [CrossRef]
- Paysan-Lafosse, T.; Blum, M.; Chuguransky, S.; Grego, T.; Pinto, B.L.; Salazar, G.A.; Bileschi, M.L.; Bork, P.; Bridge, A.; Colwell, L.; et al. InterPro in 2022. Nucleic Acids Research 2023, 51, D418–D427. [Google Scholar] [CrossRef] [PubMed]
- Hulo, N.; Bairoch, A.; Bulliard, V.; Cerutti, L.; De Castro, E.; Langendijk-Genevaux, P.S.; Pagni, M.; Sigrist, C.J.A. The PROSITE Database. Nucleic Acids Res 2006, 34, D227–D230. [Google Scholar] [CrossRef] [PubMed]
- Sonnhammer, E.L.; von Heijne, G.; Krogh, A. A Hidden Markov Model for Predicting Transmembrane Helices in Protein Sequences. Proc Int Conf Intell Syst Mol Biol 1998, 6, 175–182. [Google Scholar]
- Geourjon, C.; Deléage, G. SOPMA: Significant Improvements in Protein Secondary Structure Prediction by Consensus Prediction from Multiple Alignments. Bioinformatics 1995, 11, 681–684. [Google Scholar] [CrossRef]
- McGuffin, L.J.; Bryson, K.; Jones, D.T. The PSIPRED Protein Structure Prediction Server. Bioinformatics 2000, 16, 404–405. [Google Scholar] [CrossRef]
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J.E. The Phyre2 Web Portal for Protein Modeling, Prediction and Analysis. Nat Protoc 2015, 10, 845–858. [Google Scholar] [CrossRef]
- Wiederstein, M.; Sippl, M.J. ProSA-Web: Interactive Web Service for the Recognition of Errors in Three-Dimensional Structures of Proteins. Nucleic Acids Research 2007, 35, W407–W410. [Google Scholar] [CrossRef]
- Jensen, L.J.; Kuhn, M.; Stark, M.; Chaffron, S.; Creevey, C.; Muller, J.; Doerks, T.; Julien, P.; Roth, A.; Simonovic, M.; et al. STRING 8--a Global View on Proteins and Their Functional Interactions in 630 Organisms. Nucleic Acids Res 2009, 37, D412–416. [Google Scholar] [CrossRef] [PubMed]
- Almagro Armenteros, J.J.; Sønderby, C.K.; Sønderby, S.K.; Nielsen, H.; Winther, O. DeepLoc: Prediction of Protein Subcellular Localization Using Deep Learning. Bioinformatics 2017, 33, 3387–3395. [Google Scholar] [CrossRef] [PubMed]
- Petersen, T.N.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 4. 0: Discriminating Signal Peptides from Transmembrane Regions. Nat Methods 2011, 8, 785–786. [Google Scholar] [CrossRef]
- Blom, N.; Sicheritz-Pontén, T.; Gupta, R.; Gammeltoft, S.; Brunak, S. Prediction of Post-Translational Glycosylation and Phosphorylation of Proteins from the Amino Acid Sequence. PROTEOMICS 2004, 4, 1633–1649. [Google Scholar] [CrossRef]
- Gupta, R.; Brunak, S. Prediction of Glycosylation across the Human Proteome and the Correlation to Protein Function. Pac Symp Biocomput 2002, 310–322. [Google Scholar]
- Potenza, C.; Aleman, L.; Sengupta-Gopalan, C. Targeting Transgene Expression in Research, Agricultural, and Environmental Applications: Promoters Used in Plant Transformation. In Vitro Cell.Dev.Biol.-Plant 2004, 40, 1–22. [Google Scholar] [CrossRef]
- Ahmadi, F.; Akmar Abdullah, S.N.; Kadkhodaei, S.; Ijab, S.M.; Hamzah, L.; Aziz, M.A.; Rahman, Z.A.; Rabiah Syed Alwee, S.S. Functional Characterization of the Gene Promoter for an Elaeis Guineensis Phosphate Starvation-Inducible, High Affinity Phosphate Transporter in Both Homologous and Heterologous Model Systems. Plant Physiology and Biochemistry 2018, 127, 320–335. [Google Scholar] [CrossRef]
- Mohan, C. Isolation and Characterization of Constitutive and Wound Inducible Promoters and Validation of Designed Synthetic Stem/Root Specific Promoters for Sugarcane Transformation, 2015.
- Reese, M.G. Application of a Time-Delay Neural Network to Promoter Annotation in the Drosophila Melanogaster Genome. Computers & Chemistry 2001, 26, 51–56. [Google Scholar] [CrossRef]
- Shahmuradov, I.A.; Umarov, R.K.; Solovyev, V.V. TSSPlant: A New Tool for Prediction of Plant Pol II Promoters. Nucleic Acids Research 2017, 45, e65. [Google Scholar] [CrossRef]
- Chow, C.-N.; Lee, T.-Y.; Hung, Y.-C.; Li, G.-Z.; Tseng, K.-C.; Liu, Y.-H.; Kuo, P.-L.; Zheng, H.-Q.; Chang, W.-C. PlantPAN3. 0: A New and Updated Resource for Reconstructing Transcriptional Regulatory Networks from ChIP-Seq Experiments in Plants. Nucleic Acids Research 2019, 47, D1155–D1163. [Google Scholar] [CrossRef]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME Suite: Tools for Motif Discovery and Searching. Nucleic Acids Research 2009, 37, W202–W208. [Google Scholar] [CrossRef] [PubMed]
- Higo, K.; Ugawa, Y.; Iwamoto, M.; Korenaga, T. Plant Cis-Acting Regulatory DNA Elements (PLACE) Database: 1999. Nucleic Acids Research 1999, 27, 297–300. [Google Scholar] [CrossRef]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a Database of Plant Cis-Acting Regulatory Elements and a Portal to Tools for in Silico Analysis of Promoter Sequences. Nucleic Acids Res 2002, 30, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Aquino, S.; Carneiro, F.; Rêgo, E.; Alves, G.; Andrade, A.; Marraccini, P. Functional Analysis of Different Promoter Haplotypes of the Coffee (Coffea Canephora) CcDREB1D Gene through Genetic Transformation of Nicotiana Tabacum. Plant Cell, Tissue and Organ Culture (PCTOC) 2018, 132, 1–16. [Google Scholar] [CrossRef]
- Remy, E.; Cabrito, T.R.; Batista, R.A.; Teixeira, M.C.; Sá-Correia, I.; Duque, P. The Pht1;9 and Pht1;8 Transporters Mediate Inorganic Phosphate Acquisition by the Arabidopsis Thaliana Root during Phosphorus Starvation. New Phytologist 2012, 195, 356–371. [Google Scholar] [CrossRef] [PubMed]
- Jefferson, R.A.; Kavanagh, T.A.; Bevan, M.W. GUS Fusions: Beta-Glucuronidase as a Sensitive and Versatile Gene Fusion Marker in Higher Plants. EMBO J 1987, 6, 3901–3907. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal Biochem 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Chomczynski, P.; Sacchi, N. The Single-Step Method of RNA Isolation by Acid Guanidinium Thiocyanate–Phenol–Chloroform Extraction: Twenty-Something Years On. Nat Protoc 2006, 1, 581–585. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Selvarajan, D.; Mohan, C.; Dhandapani, V.; Nerkar, G.; Jayanarayanan, A.N.; Vadakkancherry Mohanan, M.; Murugan, N.; Kaur, L.; Chennappa, M.; Kumar, R.; et al. Differential Gene Expression Profiling through Transcriptome Approach of Saccharum Spontaneum L. under Low Temperature Stress Reveals Genes Potentially Involved in Cold Acclimation. 3 Biotech 2018, 8, 195. [Google Scholar] [CrossRef]
- Manoj, V.M.; Anunanthini, P.; Swathik, P.C.; Dharshini, S.; Ashwin Narayan, J.; Manickavasagam, M.; Sathishkumar, R.; Suresha, G.S.; Hemaprabha, G.; Ram, B.; et al. Comparative Analysis of Glyoxalase Pathway Genes in Erianthus Arundinaceus and Commercial Sugarcane Hybrid under Salinity and Drought Conditions. BMC Genomics 2019, 19, 986. [Google Scholar] [CrossRef] [PubMed]
- Dharshini, S.; Manoj, V.M.; Suresha, G.S.; Narayan, J.A.; Padmanabhan, T.S.S.; Kumar, R.; Meena, M.R.; Manickavasagam, M.; Ram, B.; Appunu, C. Isolation and Characterization of Nuclear Localized Abiotic Stress Responsive Cold Regulated Gene 413 (SsCor413) from Saccharum Spontaneum. Plant Mol Biol Rep 2020, 38, 628–640. [Google Scholar] [CrossRef]
- Narayan, J.A.; Dharshini, S.; Manoj, V.M.; Padmanabhan, T.S.S.; Kadirvelu, K.; Suresha, G.S.; Subramonian, N.; Ram, B.; Premachandran, M.N.; Appunu, C. Isolation and Characterization of Water-Deficit Stress-Responsive α-Expansin 1 (EXPA1) Gene from Saccharum Complex. 3 Biotech 2019, 9, 186. [Google Scholar] [CrossRef]
- Ashwin Narayan, J.; Chakravarthi, M.; Nerkar, G.; Manoj, V.M.; Dharshini, S.; Subramonian, N.; Premachandran, M.N.; Arun Kumar, R.; Krishna Surendar, K.; Hemaprabha, G.; et al. Overexpression of Expansin EaEXPA1, a Cell Wall Loosening Protein Enhances Drought Tolerance in Sugarcane. Industrial Crops and Products 2021, 159, 113035. [Google Scholar] [CrossRef]
- Narayan, A.; V M, M.; Nerkar, G.; Mohan, C.; Dharshini, S.; Subramonian, N.; Premachandran, M.; Valarmathi, R.; Kumar, R.; Gomathi, R.; et al. Transgenic Sugarcane with Higher Levels of BRK1 Showed Improved Drought Tolerance. Plant Cell Reports 2023, 42, 1–18. [Google Scholar] [CrossRef]
Figure 1.
EaPHT1;2 Gene isolation and Phylogenetic analysis. (A) EaPHT1 gene amplification. Lanes: M- 1kb DNA Ladder; and 1 to 3- EaPHT1;2 (∼1650 bp). (B) E. coli transformants screened with EaPHT1;2 gene-specific primer, Lanes; M - 1kb DNA Ladder; and 1 to 8-Random transformants selected for screening showing amplification of EaPHT1 gene. The red arrow indicates the specific amplicon of 1650bp. (C) Phylogenetic analysis of PHT1 gene obtained from E. arundinaceus, Z. mays, S. bicolor, Arabidopsis and Saccharum hybrid using MEGA.
Figure 1.
EaPHT1;2 Gene isolation and Phylogenetic analysis. (A) EaPHT1 gene amplification. Lanes: M- 1kb DNA Ladder; and 1 to 3- EaPHT1;2 (∼1650 bp). (B) E. coli transformants screened with EaPHT1;2 gene-specific primer, Lanes; M - 1kb DNA Ladder; and 1 to 8-Random transformants selected for screening showing amplification of EaPHT1 gene. The red arrow indicates the specific amplicon of 1650bp. (C) Phylogenetic analysis of PHT1 gene obtained from E. arundinaceus, Z. mays, S. bicolor, Arabidopsis and Saccharum hybrid using MEGA.
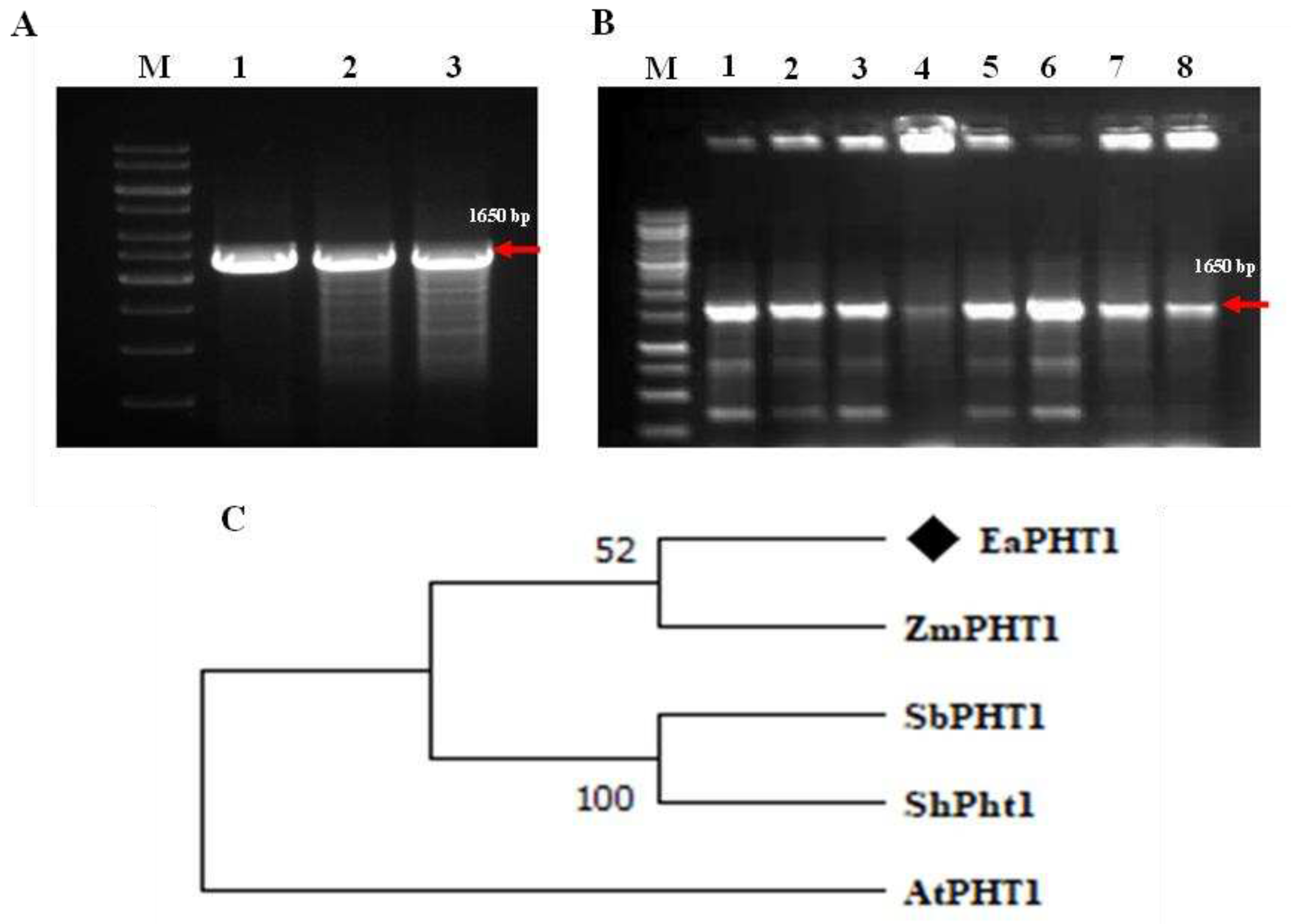
Figure 2.
Protein domain and Topology analysis of EaPHT1:2. (A) The Major facilitator superfamily (MFS) domain, denotes in the 20-511 a.a sequence, is a gene conserved domain represented in all PHT1 genes. (B) The topology of EaPHT1;2 was predicted based on its protein sequence. Loops and coils are denoted by lines. The yellow blocks represent transmembrane domains S1-S10. The numbers at the top and bottom of each domain indicate the positions of amino acid residues. (C) Hydropathy plots of the EaPHT1;2 protein. The X-axis represents the location of the protein starting from the N-terminus region and the Y-axis indicates the hydrophobicity score of the protein. The peaks denote that EaPHT1;2 is hydrophobic in nature.
Figure 2.
Protein domain and Topology analysis of EaPHT1:2. (A) The Major facilitator superfamily (MFS) domain, denotes in the 20-511 a.a sequence, is a gene conserved domain represented in all PHT1 genes. (B) The topology of EaPHT1;2 was predicted based on its protein sequence. Loops and coils are denoted by lines. The yellow blocks represent transmembrane domains S1-S10. The numbers at the top and bottom of each domain indicate the positions of amino acid residues. (C) Hydropathy plots of the EaPHT1;2 protein. The X-axis represents the location of the protein starting from the N-terminus region and the Y-axis indicates the hydrophobicity score of the protein. The peaks denote that EaPHT1;2 is hydrophobic in nature.
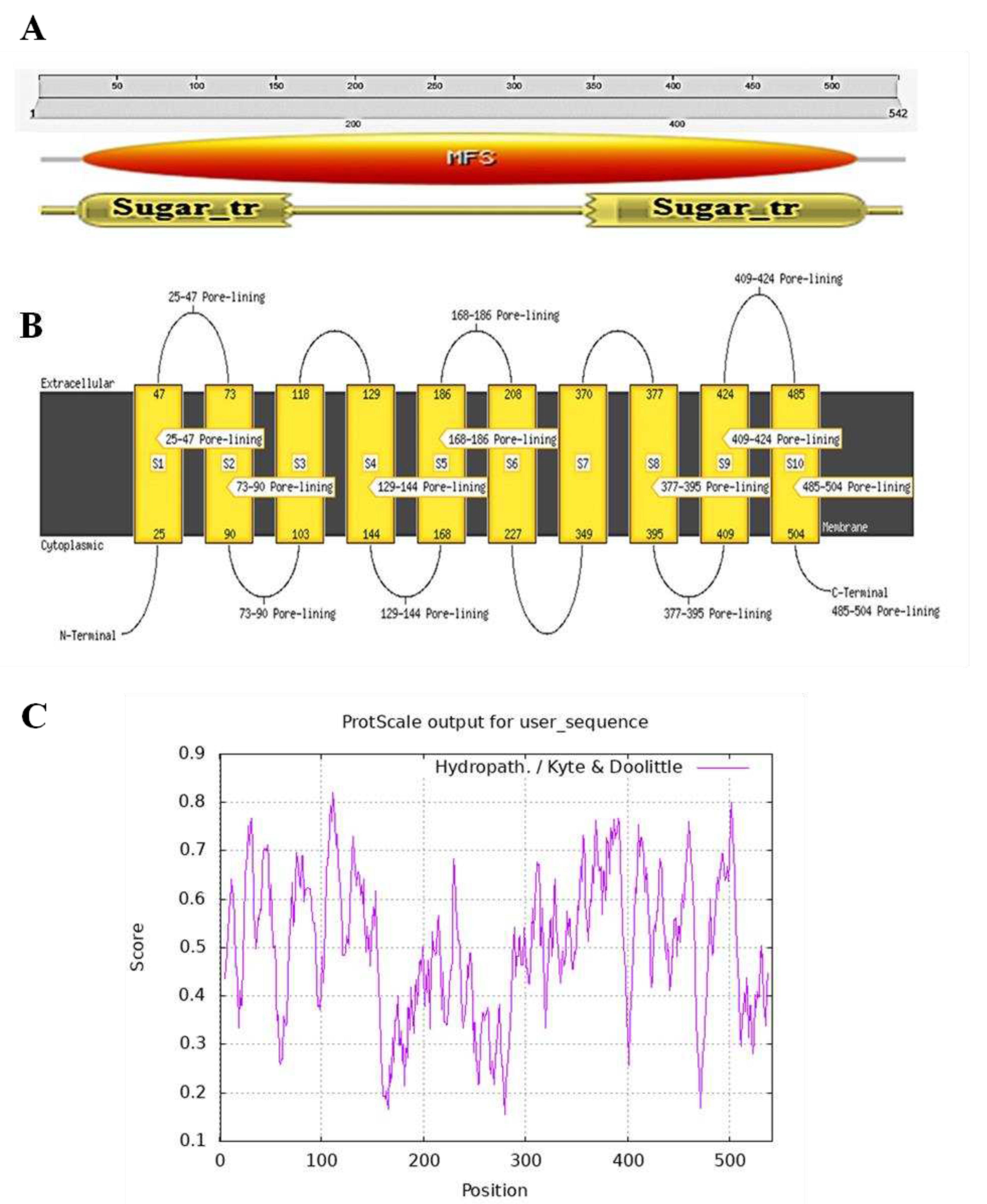
Figure 3.
Protein structure and Homologous sequence analysis. (A) Secondary structure of EaPHT1;2 protein predicted using PSIPRED. (B) Homologous sequence alignment and conservation among the PHT1 proteins from E. arundinaceus, Z. mays, S. bicolor, Arabidopsis and Saccharum hybrid. The violet highlighted position in a black box represents the conserved sequence of the PHT1 gene family.
Figure 3.
Protein structure and Homologous sequence analysis. (A) Secondary structure of EaPHT1;2 protein predicted using PSIPRED. (B) Homologous sequence alignment and conservation among the PHT1 proteins from E. arundinaceus, Z. mays, S. bicolor, Arabidopsis and Saccharum hybrid. The violet highlighted position in a black box represents the conserved sequence of the PHT1 gene family.
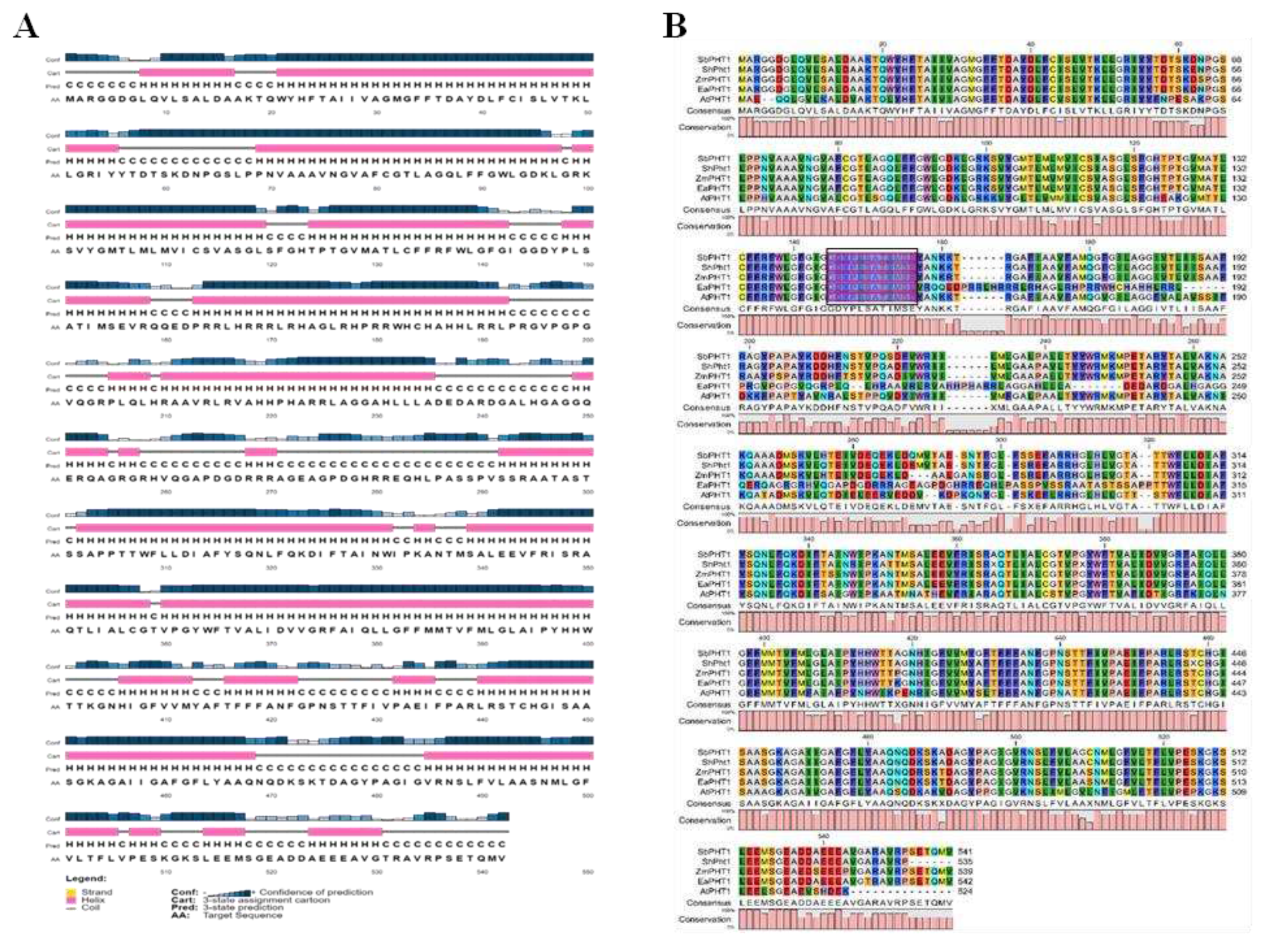
Figure 4.
3D structural analysis and Validation. (A) EaPHT1;2 3D structure obtained using the Phyre2 analysis. (B) The model was obtained using Swiss modelling. (C), and (D) RamPlot representation for the Phyre2 and Swiss-modelling structure validation using VADAR. (E) and (F) Qualitative evaluation of EaPHT1;2 3D structure using ProSA webserver for Phyre2 model and Swiss-modelling. (G) Electrostatic surface representation of EaPTT1;2 and (H) The structure of EaPHT1;2 in complex with substrate, PHOSPHATE ION (PO4).
Figure 4.
3D structural analysis and Validation. (A) EaPHT1;2 3D structure obtained using the Phyre2 analysis. (B) The model was obtained using Swiss modelling. (C), and (D) RamPlot representation for the Phyre2 and Swiss-modelling structure validation using VADAR. (E) and (F) Qualitative evaluation of EaPHT1;2 3D structure using ProSA webserver for Phyre2 model and Swiss-modelling. (G) Electrostatic surface representation of EaPTT1;2 and (H) The structure of EaPHT1;2 in complex with substrate, PHOSPHATE ION (PO4).
Figure 5.
EaPHT1;2 Promoter isolation and In-silico validation. (A) Restriction of the E. arundinaceus genomic ‘DNA. Lanes: M - 1kb DNA marker; 1 - Unrestricted gDNA; 2 to 6 - restricted gDNA (Lane 2-6 as follows: DraI, EcoRV, HincII, PvuII, and SspI). (B) RAGE PCR Amplification of EaPHT1-2 promoter; Lanes: M- 1 kb DNA marker. Lane: 1-5, Primary PCR products; and Lane: 6-10, Secondary PCR products of 1.5kb. (C) Promoter sequence prediction by NNPP tool. The position of the TSS is highlighted with a larger font size. (D) In-silico analysis of Cis-elements in EaPHT1;2 promoter, common motifs, Pi related motif and root-specific motifs were highlighted and marked bold in different colors.
Figure 5.
EaPHT1;2 Promoter isolation and In-silico validation. (A) Restriction of the E. arundinaceus genomic ‘DNA. Lanes: M - 1kb DNA marker; 1 - Unrestricted gDNA; 2 to 6 - restricted gDNA (Lane 2-6 as follows: DraI, EcoRV, HincII, PvuII, and SspI). (B) RAGE PCR Amplification of EaPHT1-2 promoter; Lanes: M- 1 kb DNA marker. Lane: 1-5, Primary PCR products; and Lane: 6-10, Secondary PCR products of 1.5kb. (C) Promoter sequence prediction by NNPP tool. The position of the TSS is highlighted with a larger font size. (D) In-silico analysis of Cis-elements in EaPHT1;2 promoter, common motifs, Pi related motif and root-specific motifs were highlighted and marked bold in different colors.
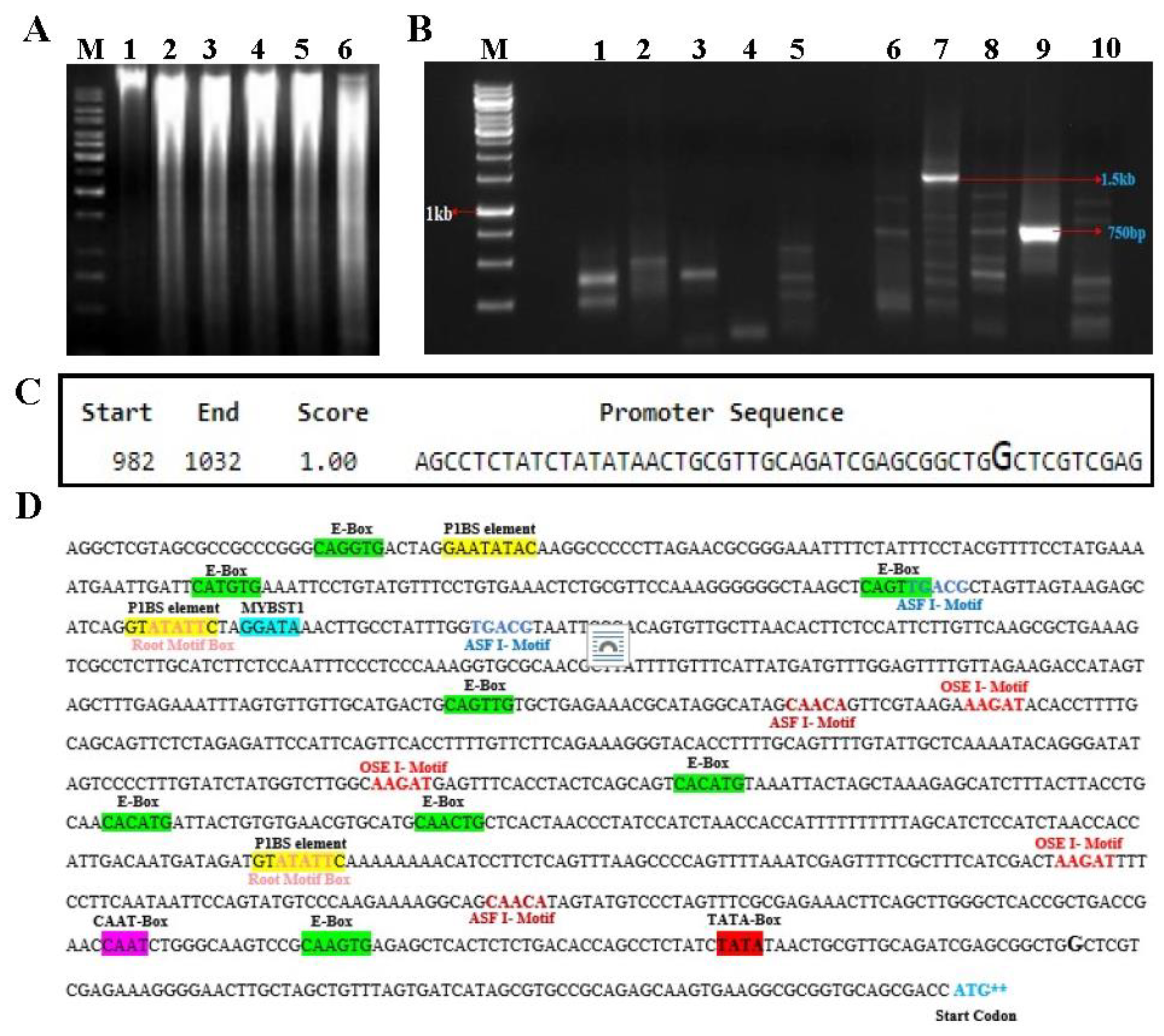
Figure 6.
GUS Expression Analysis of EaPHT1;2 Promoter. (A) Histochemical GUS expression analysis in transgenic pFL promoter. (B) Systemic representation of EaPHT1;2 regulatory sequence deletion constructs. The size of each construct was mentioned in numbers and deletions are names as pD1 to pD5. The red color highlighted region denotes the 5’UTR region. (C) Fluorometric analysis of putative transgenic events under Pi stress. (D) qRT-PCR analysis of GUS activity in transgenic tobacco lines under Pi stress. (WT- Wild Type, UTC-Untreated Control, L-Pi – Low Pi, H-Pi – High Pi).
Figure 6.
GUS Expression Analysis of EaPHT1;2 Promoter. (A) Histochemical GUS expression analysis in transgenic pFL promoter. (B) Systemic representation of EaPHT1;2 regulatory sequence deletion constructs. The size of each construct was mentioned in numbers and deletions are names as pD1 to pD5. The red color highlighted region denotes the 5’UTR region. (C) Fluorometric analysis of putative transgenic events under Pi stress. (D) qRT-PCR analysis of GUS activity in transgenic tobacco lines under Pi stress. (WT- Wild Type, UTC-Untreated Control, L-Pi – Low Pi, H-Pi – High Pi).
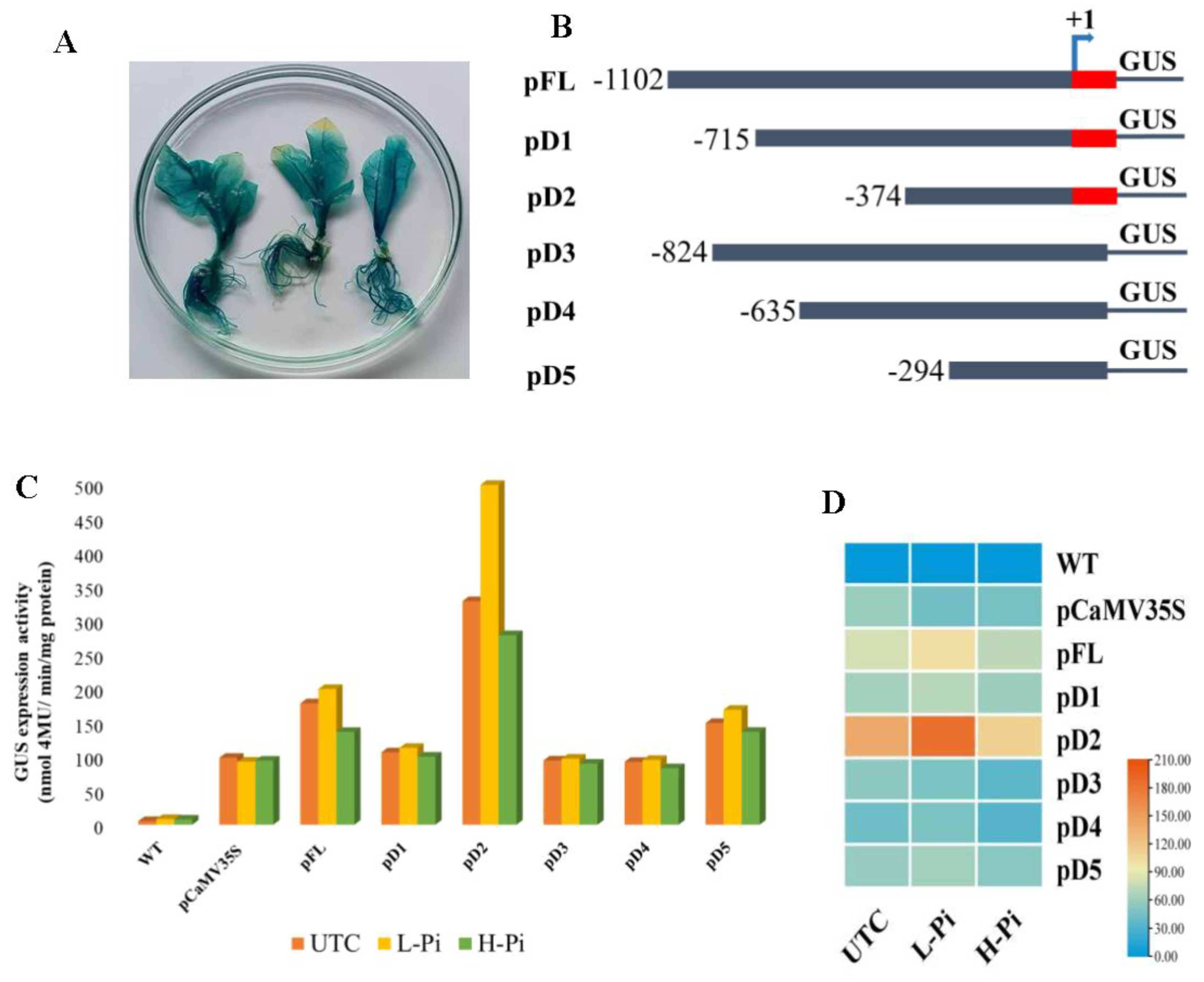
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



