
Submitted:
03 October 2023
Posted:
05 October 2023
You are already at the latest version
Abstract
Monitoring water supply requires, among other quality indicators, the identification of the cyanobacte-ria community taking into account their potential impact in terms of water quality. In this work, cya-nobacteria strains were isolated from Cheffia reservoir and identified based on morphological features, 16S rRNA gene, phylogenetic analysis, and toxin production by PCR screening of the genes involved in the biosynthesis of cyanotoxins (mcyA, mcyE, sxtA, sxtG, sxtI, cyrJ, and anaC). Thirteen strains repre-senting six different genera: Aphanothece, Microcystis, Geitlerinema, Lyngbya, Microcoleus, and Pseudanabaena were obtained. The results demonstrated the importance of morphological features in determining the genus or the species when incongruence between the morphological and phylogenetic analysis occurs and only the utility of the 16S rRNA gene on determining higher taxonomic levels. The phylogenetic analysis confirmed the polyphyly of cyanobacteria for the Microcystis and Oscillatoriales genera. Unexpectedly, Aphanothece sp. CR 11 had the genetic potential to produce microcystins. Our study gives new insight instead of picocyanobacteria toxic genotype in this ecosystem. Thus, conven-tional water treatment methods in this ecosystem have to be adapted.
Keywords:
Algeria
; cyanobacteria
; mcyE gene
; morphology
; Picocyanobacteria
; phylogeny
1. Introduction
Cyanobacteria are photosynthetic prokaryotes found in aquatic and terrestrial habitats [1]. These organisms are prolific producers of natural products recognized as toxins with hepatotoxic, neurotoxic, and dermatotoxic effects, harmful to humans and animals [2]. For people, the Caruraru syndrome, which occurred in Brazil (Caruaru City), was the most severe event. During this incident, 76 patients died in a hemodialysis clinic; analysis of liver tissue and serum of the victims led to the identification of microcystins (MCs, hepatotoxic cyanotoxins) [3]. Nowadays, monitoring of water intended for public use takes account of the identification of toxic cyanobacteria [4].
Morphological analyses have been the basis of cyanobacterial classification systems [5]. However, because of limitations due to the extreme phenotypic flexibility depending on different environmental and culture conditions [6], a polyphasic approach combining morphological and molecular information has been used for accurate cyanobacteria identification [7]. Nevertheless, this approach cannot differentiate between toxic and non-toxic cyanobacteria strains. Accordingly, various immunological and analytical approaches such as enzyme-linked immunosorbent assays (ELISA), the protein phosphatase inhibition (PPI) assay, chromatography, and mass spectrometry [4] have been used for this objective. However, sensitivity, specificity, and high-cost limitations have made them inadequate for routine use [8]. The capacity of cyanobacteria to produce toxins is associated with specific metabolic processes that are encoded by complex gene operons. Toxin-related genes PCR amplification is a reliable technique for detecting potential toxic strains [9].
In Algeria, several reports have described the occurrence of cyanobacteria in ecosystems with socio-economic importance [10,11,12]. Cheffia reservoir is a water body in the North-East of Algeria; the water is used for drinking and irrigation. Previous reports have been particularly limited to analyzing environmental samples and toxins in this ecosystem and identifying cyanobacteria by culture-independent technique [10]. Cultivation approach has advantages over the cultivation-independent studies of cyanobacteria; it allows detailed phenotypic, genetic, physiological and biochemical characterization [13]. Genetic research with axenic cultures are critical for cyanobacteria polyphasic taxonomy investigations, particularly for understudied polyphyletic taxa [14]. Recent studies have shown that numerous newly described cyanobacteria are exclusively based on isolated strains [15,16,17,18]. Thus, the aim of this study was focused specifically on cyanobacterial isolates from Cheffia reservoir. Isolated strains were identified using morphological features and phylogenetic analysis. Furthermore, toxin-related genes were localized using specific primers.
2. Materials and Methods
2.1. Collection of cyanobacterial samples and isolation
Cyanobacteria sample was collected from Cheffia reservoir (36°36'33.5"N 8°02'34.8"E, North-East of Algeria) using the plankton net (20 µm mesh size) on October 21, 2018. Cyanobacteria isolation was done by enrichment of cyanobacterial environmental samples in liquid culture medium, then plating by streaking each sample on 1.5% agar culture medium. Plates were observed regularly under an inverted microscope Leica® DMi8, and isolated strains were transferred to new plates and also maintained on liquid medium. Cyanobacteria were isolated, grown, and maintained in BG11 medium [19] under a 14:10 h light/dark cycle at 19 ± 1 °C. Cycloheximide (Sigma) at 100 μg mL−1 was added to the culture medium in the first isolation steps to inhibit the growth of eukaryotic microorganisms [20].
2.2. Morphological characterization
Morphological studies of isolated strains were carried out under an Olympus BX51 light microscope at 100x, 400x, and 1000x magnifications. In addition, photographs were taken with a Leica® DFC550 digital camera, and measurements were determined for each strain (n = 100 cells) at 1000 x magnification using Leica®Lasx software. Strains were classified according to the morphological criteria of Komárek & Anagnostidis [21,22].
2.3. DNA Extraction, Amplification (PCR) and Sequencing
Fresh cyanobacterial cultures at the exponential growth phase were harvested by centrifugation (Micro STAR 17R, VWR) at 12.000 g for 5 min and stored at −20 °C. Genomic DNA (gDNA) was extracted using the Purelink Genomic DNA Mini Kit (Invitrogen, San Diego, CA) according to the manufacturer’s instructions for Gram-negative bacteria. The integrity and quality of DNA was checked on 1.0 % agarose gel and gDNA was stored at −20 °C. The 16S rRNA gene was amplified using the primers CYA359F (GGGGAATYTTCCGCAATGGG), CYA781R (GACTACTGGGGTATCTAATCCCATT) [23], 27F1 (5'- AGA GTT TGA TCC TGG CTC AG-3') and 1494Rc (5'-TAC GGC TAC CTT GTT ACG AC-3') [24]. The PCRs reactions were performed with a Biometra TProfessional gradient thermocycler (Biometra, Göttingen, Germany), in a final volume of 20 μl following the methodologies described previously by Nübel et al. [23] and Neilan et al. [24]. The reaction mixture contained 4 μl 5× Green GoTaq® Flexi buffer (Promega), 2 μl MgCl2 (25 mM), 2 μl of each primer (10 μM), 1 μl of deoxynucleoside triphosphate (dNTP, 10 μM) mix (Promega), bovine serum albumin (BSA), 0.1 μl of GoTaq® DNA polymerase (Promega) and 1 μl of template DNA. PCR products were examined by electrophoresis in 1.0% agarose gels stained with SYBR® Safe DNA Gel Stain (Thermo Fisher Scientific) and then purified with a NucleoSpin® Gel and PCR Clean-up Kit (Macherey-Nagel, Düren, Germany) according to the manufacturer’s instructions. The obtained products were sequenced bidirectionally by Sanger sequencing by GATC Biotech (Germany).
For the obtained sequences with low DNA quality, purified PCR products were cloned into the pCR™2.1-TOPO® vector (Invitrogen TOPO® TA Cloning Kit) and then transformed into Escherichia coli OneShot® TOP10 cells (Invitrogen) following the instructions of the manufacturers. After the white-blue selection on ampicillin 1.5% agarose plates with Luria Bertani (LB) medium, colonies were transferred into fresh liquid LB medium with 100 μg ml–1 of ampicillin and cultured overnight at 37 °C with shaking at 2000 rpm. The Plasmid DNA was isolated using GenElute™ Plasmid Miniprep Kit (Sigma-Aldrich, St. Louis, MO, USA) according to the manufacturer’s instructions. The plasmids were sequenced bidirectionally, using the following primers: 27F1, CYA359F, CYA781R, and 1494Rc, by Sanger sequencing at GATC Biotech (Germany).
2.4. Sequence analysis
Raw forward and reverse sequences were assembled using Geneious (version 8.1.8) then deposited in the GenBank database (http://www.ncbi.nlm.nih.gov/) under the accession numbers OM401262 to OM401274. The obtained sequences were compared to previously published sequences in NCBI (National Center for Biotechnology Information; accessible via http://www.ncbi.nlm.nih.gov/) using the BLASTN algorithm. Based on the resulting blast analysis, a phylogenetic tree was constructed using the “One Click” mode of the online software Phylogeny.fr platform (http://www.phylogeny.fr/simple_phylogeny.cgi, [25]).
2.5. PCR amplification for toxins production
PCR was used to amplify toxin-synthesis-related genes encoding microcystin (mcyA), the amino transferase (AMT) of the microcystin and nodularin synthetases complexes (mcyE), saxitoxin (sxtA, sxtG, and sxtI), anatoxin (anaC), and cylindrospermopsin (cyrJ) by using specific primer sets (Table 1) and PCR programs described previously [26,27,28,29,30,31]. The PCR reactions were performed in a total volume of 18 µl using the GoTaq® DNA polymerase (Promega, Madison, WI, USA) 1 X PCR buffer, 2.5 mM MgCl2, 250 µM of dNTP mix, 10 pmol of each primer, 0.5 U of Taq DNA polymerase and 1 µl of genomic DNA. The PCRs were run either on a Veriti thermal cycler (Applied Biosystems, Foster City, CA, USA) or a Biometra TProfessional gradient thermocycler (Biometra, Göttingen, Germany). Obtained PCR products were examined by electrophoresis in 1.0% agarose gels stained with SYBR® Safe DNA Gel Stain (Thermo Fisher Scientific) and photographed under UV transillumination. Microcystis aeruginosa LEGE 91339 was used as a positive control for the mcyA and mcyE genes, Aphanizomenon gracile LMECYA 040 for the sxtA, sxtG and sxtI genes, Cylindrospermopsis raciborskii LEGE 97047 and Anabaena sp. LEGE X-002 for the cyrJ and anaF genes, respectively. These strains belong to the culture collection of the Blue Biotechnology and Ecotoxicology Laboratory (Blue Biotechnology and Ecotoxicology Culture Collection, LEGE-CC) of CIIMAR (Porto, Portugal).
3. Results
3.1. Cyanobacterial Isolation and Morphological Identification
In this work thirteen cyanobacteria isolates were obtained. Following the morphological features of Komárek & Anagnostidis [21,22], the isolated strains were identified as Geitlerinema (G.) amphibium (4 strains), Lyngbya (L.) nigra (1 strain), L. stagnina (1 strain), L. cincinnata (1 strain), Microcoleus sp. (1 strain), Pseudanabaena (P.) rosea (1 strain), Aphanothece sp. (2 strains), Microcystis (M.) flos-aquae (1 strain) and Microcystis sp. (1 strain). Overall, 69% of the isolates were identified morphologically at the species level, and 31% were identified only at the genus level. The morphological features of isolates are illustrated in Table 2 and Table 3 and Figure 1 and Figure 2 represent their photomicrographs.
3.2. Phylogenetic study
BLAST analysis showed high sequence similarities ranging from 97.26 to 99.91% between the 16S rRNA gene sequences of cyanobacterial isolates from Cheffia reservoir and 27 strain sequences from Chroococcales, Oscillatoriales and Synechococcales orders accessible in GenBank (Table 4).
The phylogenetic tree obtained with the studied strains is showed in Figure 3. The tree included five main clusters. Cluster A comprised sequences of Microcoleus sp. and Lyngbya spp.. Sub-cluster I included Microcoleus sp. CR8 isolate and Microcoleus HTT-U-KK5 strain from GenBank,. Sub-cluster II contained Lyngbya isolates with sequence from the same genera: L. martensiana H3b/33 and two other sequences from Oscillatoria UIC 10045 and Phormidium cf. irriguum CCALA 759. Cluster B consists only of the Microcystis flos-aquae CR13 isolated in this work with sequences from the same species, M. flos-aquae CHAB541, and those of Synechocystis SAG 45.90 and Sphaerovacum brasiliense CCIBt3094 from GenBank. Cluster C is represented by G. amphibium strains with the strain G. amphibium and Limnothrix, Jaaginema, Pseudanabaena, and Anagnostidinema sequences. Cluster D included the two strains of the picocyanobacterial genera Aphanothece and the isolate identified morphologically as Microcystis sp. grouped with sequences belonging to Cyanobacterium, Cyanobium, Synechococcus, Aphanothece, Cyanodictyon, Aphanocapsa, and Microcystis from GenBank. Finally, cluster E included the Psedanabaena isolate and sequences in the GenBank database of the species P. limnetica Lim1, L. redekei CCAP 1459/29 and Arthronema gygaxiana UTCC 393. Finally, cluster D included the two strains of the picocyanobacterial genera Aphanothece and the isolate identified morphologically as Microcystis sp. grouped with sequences belonging to Cyanobacterium, Cyanobium, Synechococcus, Aphanothece, Cyanodictyon, Aphanocapsa, and Microcystis sequences obtained from GenBank. Cluster E included the Psedanabaena isolate and sequences in the GenBank database of the species P. limnetica Lim1, L. redekei CCAP 1459/29, and Arthronema gygaxiana UTCC 3.
3.3. PCR amplification of toxin-encoding genes
The molecular screening of the genes involved on the biosynthesis of microcystin was positive to the mcyE specific microcystin/nodularin gene for Aphanothece sp. CR11 strain, as showed in Figure 4. However, none of the isolated strains was tested positive for anatoxin (anaC), saxitoxin (sxtA, sxtG, and sxtI) or cylindrospermopsin (cyrJ) genes.
4. Discussion
In order to characterize cyanobacteria isolates from Cheffia reservoir, morphological, molecular and phylogenetic analyses were conducted. The morphological identification up to the species level was successful for some strains. More importantly, the strains from the genera Geitlerinema, Lyngbya and Microcystis showed substantial phenotypic plasticity. All the isolated Geitlerinema strains have identical morphological characteristics and are similar to G. amphibium described by Bittencourt-Oliveira et al. [33]. However, our isolates vary from these organisms; their cell length was less than these organisms, which are 2.2–7 µm long, and their cell width was relatively wider than these organisms, which are 1.0–2.2 µm wide. The cells of isolated strains belonging to the species P. rosea, L. nigra, L. cincinnata, and L. stagnina exhibited a larger width than those described by Komárek & Anagnostidis [21], which are 1.61–2.67, 12.82–17.84, 13.46–19.66 and 13.00–19.36 µm, respectively. As well, the M. flos-aquae strain had cells with a larger diameter than the M. flos-aquae described by Komárek & Anagnostidis [22], which is 3.5–4.8 µm. The genus Microcystis has previously shown significant morphological flexibility [34,35,36]. This morphological plasticity has been related to several environmental or cultivation factors.
Our findings also showed that morphological and phylogenetic classifications might be incompatible. This is the case of the strain Microcystis sp. CR 12, assigned to the genus Microcystis based on the morphological description of this genus in Bergey's Manual of Determinative Bacteriology [37]. The genus Microcystis is characterized by the following features: coccoid cells with aerotops and a tendency to form colonies delimited by mucilage. Differences between molecular and morphological descriptions have been extensively reported [38,39,40,41]. Moreover, it has also been demonstrated that GenBank contains erroneously classified species. Strains in culture collections have also been misnamed, and they may be found in GenBank and other culture collections under various names [38]. Moreover, with the numerous revisions to cyanobacteria taxonomy, there is no simple way to update existing databases [42], generating even more difficulty for cyanobacteria assignment [27,39]. Nonetheless, the number of 16S rRNA gene sequences now accessible in global databases for some species is still low, and additional systematic efforts will be necessary to clarify existing taxonomic ambiguities [27].
The isolated Lyngbya strains were morphologically assigned to 3 distinct species, although phylogenetically related to one species, L. martensiana H3b/33. The reliability of 16S rRNA gene sequences to identify Lyngbya strains at the species level and lower has been contested [43]. Engene et al. [44] suggested only using 16S rRNA gene sequences to identify Lyngbya strains at the genus level. Furthermore, Fathalli et al. [45] confirmed that even when closely related species are morphologically different, it may not be possible to differentiate them properly by the 16S rRNA gene. The 16S rRNA gene is less effective for phylogenetic investigations of closely related organisms due to its conserved character and lower evolutionary rate fluctuation than protein-encoding genes [46].
Three distinct clades were obtained from the Oscillatoriales strains, confirming the polyphyletic origin of this order [47]. The obtained Microcystis morphospecies were mixed in the phylogenetic tree and cannot be identified as monophyletic organisms, which also confirms the polyphyly of the Chroococcales [14,48].
Our study represents the first report of toxic genotype from the genus Aphanothece in this ecosystem. Picocyanobacteria MCs producers have been isolated from the Tabocas reservoir in the Caruaru city, where the first human poisoning episode was reported [49]. MCs are the most dangerous group of cyanotoxins; they inhibit serine and threonine phosphatases−PP1 and PP2A, causing significant hepatotoxicity and acting as a carcinogen [50].
Some studies have also shown that strains of picocyanobacteria can produce MCs in culture, but no study has explored the presence of toxicity genes [51,52]. The picocyanobacteria group can also produce blooms responsible for the significant loss of benthic wildlife [52,53,54,55]. The most studied picocyanobacteria blooms have been observed in the northern Mediterranean at the Comacchio lagoons (North-West Adriatic coast) [56]. These blooms led to the death of bottom flora and benthic fauna and the loss of valuable resources of fish (eel and mullet) and mollusks [57]. Śliwińska-Wilczewska et al. [52] and Felpeto et al. [58] suggest that picocyanobacteria blooms are a new phenomenon that requires comprehensive studies. Although the impacts of climate change on picocyanobacterial blooms are diverse, most current evidence suggests that this process increases the amplitude and frequency of these events [52,59]. So far, little is known regarding the toxicity of picocyanobacteria, whereas the number of reports about their prevalence in ecosystems is growing. Therefore, the problem of picocyanobacteria toxicity requires more researchers’ attention and interest [52].
5. Conclusions
The results highlighted that identifying cyanobacteria isolates by combining morphological and molecular methods is still challenging. Furthermore, for the first time in this reservoir, a toxic genotype from picocyanobacteria was described; this group of cyanobacteria is often overlooked, although its toxicity is of great importance. Thus, water treatment methods in this ecosystem must consider the presence of picocyanobacteria strains. In addition, it is vitally important to conduct in-depth research on the function of picocyanobacteria in aquatic ecosystems.
Author Contributions
Conceptualization, L.B. and V.V.; Methodology, L.B., J.M, G.S.H and V.V; Formal Analysis, L.B., J.M, G.S.H and V.V; Investigation, L.B. and V.V; Resources, V.V.; Writing – Original Draft Preparation, L.B.; Writing – Review & Editing, J.M. and V.V; Supervision, V.V.; Funding Acquisition, V.V..
Funding
Lamia BENREDJEM is thankful to the Algerian Ministry of High Education and Scientific Research for the fellowship funding. We want to thank the Cheffia Dam personnel, National Agency of Dams and Transfers (ANBT) for their excellent assistance during the sampling. CIIMAR acknowledges the projects FCT UIDB/04423/2020 and UIDP/04423/2020 and the WP9- Portuguese Blue Biobank under the Blue Economy Pact - Project Nº. C644915664-00000026 co-funded by PRR, The Portuguese Republic and the European Union.
Acknowledgments
The authors are grateful to Pedro N. Leão (Interdisciplinary Centre of Marine and Environmental Research, CIIMAR/CIMAR) for kindly providing the Invitrogen TOPO® TA Cloning Kit.
Conflicts of Interest
The authors declare no conflict of interest
References
- Svirčev, Z.; Chen, L.; Sántha, K.; Drobac Backović, D.; Šušak, S.; Vulin, A.; Palanački Malešević, T.; Codd, G.A.; Meriluoto, J. A Review and Assessment of Cyanobacterial Toxins as Cardiovascular Health Hazards. Arch Toxicol 2022, 96, 2829–2863. [Google Scholar] [CrossRef]
- Dittmann, E.; Fewer, D.P.; Neilan, B.A. Cyanobacterial Toxins: Biosynthetic Routes and Evolutionary Roots. FEMS Microbiol Rev 2013, 37, 23–43. [Google Scholar] [CrossRef]
- Jochimsen, L.M.; Armichael, A.W.C.; Ardo, E.M.C.; Usan, S.; Ookson, T.C.; Hristianne, C.; Olmes, E.M.H.; Ernadete, M.B.; Ntunes, D.C.A.; De M Elo, J.A.; et al. Liver Failure and Death after Exposure to Microcystins at a Hemodialysis Center in Brazil. N Engl J Med 1998, 338, 873–878. [Google Scholar] [CrossRef]
- Chorus, I.; Welker, M. Toxic Cyanobacteria in Water, 2nd edition; CRC Press, 2021. [Google Scholar]
- Castenholz, R.W. Species Usage, Concept, and Evolution in the Cyanobacteria (Blue-Green Algae). J Phycol 1992, 28, 737–745. [Google Scholar] [CrossRef]
- Palinska, K.A.; Liesack, W.; Rhiel, E.; Krumbein, W.E. Phenotype Variability of Identical Genotypes: The Need for a Combined Approach in Cyanobacterial Taxonomy Demonstrated on Merismopedia-like Isolates. Arch Microbiol 1996, 166, 224–233. [Google Scholar] [CrossRef]
- Komárek, J. A Polyphasic Approach for the Taxonomy of Cyanobacteria: Principles and Applications. Eur J Phycol 2016, 51, 346–353. [Google Scholar] [CrossRef]
- El Semary, N.A. Modern Methods for Detection and Elimination of Microcystins Toxins Produced by Cyanobacteria: Mini-Review. Journal of Applied Sciences 2010, 10, 1662–1666. [Google Scholar] [CrossRef]
- Yuan, J.; Yoon, K.-J. Overview of PCR Methods Applied for the Identification of Freshwater Toxigenic Cyanobacteria. In; 2021.
- Benredjem, L.; Berredjem, H.; Abdi, A.; Casero, M.C.; Quesada, A.; Fosso, B.; Marzano, M.; Pesole, G.; Azevedo, J.; Vasconcelos, V. Morphological, Molecular, and Biochemical Study of Cyanobacteria from a Eutrophic Algerian Reservoir (Cheffia). Environmental Science and Pollution Research 2022, 29, 27624–27635. [Google Scholar] [CrossRef]
- Saoudi, A.; Brient, L.; Boucetta, S.; Ouzrout, R.; Bormans, M.; Bensouilah, M. Management of Toxic Cyanobacteria for Drinking Water Production of Ain Zada Dam. Environ Monit Assess 2017, 189. [Google Scholar] [CrossRef]
- Touati, H.; Guellati, F.Z.; Arif, S.; Bensouilah, M. Cyanobacteria Dynamics in a Mediterranean Reservoir of the North East of Algeria: Vertical and Seasonal Variability. Journal of Ecological Engineering 2019, 20, 93–107. [Google Scholar] [CrossRef]
- Bryant, D.A. A Brief History of Cyanobacterial Research: Past, Present, and Future Prospects. In The Cell Biology of Cyanobacteria; Flores, E., Herrero, A., Eds.; Caister Academic Press: Norfolk, UK, 2014; pp. 1–1. [Google Scholar]
- Komárek, J.; Kaštovský, J.; Mareš, J.; Johansen, J.R. Taxonomic Classification of Cyanoprokaryotes (Cyanobacterial Genera) 2014, Using a Polyphasic Approach. Preslia 2014, 86, 295–335. [Google Scholar]
- Genuário, D.B.; Vieira Vaz, M.G.M.; Hentschke, G.S.; Sant ’ Anna, C.L.; Fiore, M.F. Halotia Gen. Nov., a Phylogenetically and Physiologically Coherent Cyanobacterial Genus Isolated from Marine Coastal Environments. Int J Syst Evol Microbiol 2015, 65, 633–675. [Google Scholar] [CrossRef]
- Hauer, T.; Bohunická, M. Reassessment of the Cyanobacterial Family Microchaetaceae and Establishment of New Families Tolypothrichaceae and Godleyaceae 1. J Phycol 2014, 50, 1089–1100. [Google Scholar] [CrossRef]
- Mai, T.; Johansen, J.R.; Pietrasiak, N.; Bohunická, M.; Martin, M.P. Revision of the Synechococcales (Cyanobacteria) through Recognition of Four Families Including Oculatellaceae Fam. Nov. and Trichocoleaceae Fam. Nov. and Six New Genera Containing 14 Species. Phytotaxa 2018, 365, 1–59. [Google Scholar] [CrossRef]
- Raabová, L.; Kovacik, L.; Elster, J.; Strunecký, O. Review of the Genus Phormidesmis (Cyanobacteria) Based on Environmental, Morphological, and Molecular Data with Description of a New Genus Leptodesmis. Phytotaxa 2019, 395, 1–16. [Google Scholar] [CrossRef]
- Rippka, E.; Deruelles, J.; Waterbury, N.B. Generic Assignments, Strain Histories and Properties of Pure Cultures of Cyanobacteria. Journal of General MicrobioZogy 1979, 111, 1–61. [Google Scholar] [CrossRef]
- Rippka, R. Isolation and Purification of Cyanobacteria. Methods Enzymol 1988, 167, 3–27. [Google Scholar]
- Komárek, J.; Anagnostidis, K. Cyanoprokaryota 2. Teil/ 2nd Part: Oscillatoriales; Büdel, B., Krienitz, L., Gärtner, G., Schagerl, M., Eds.; Elsevier/Spektrum: Heidelberg, Germany, 2005; Vol. 759. [Google Scholar]
- Komárek, J.; Anagnostidis, K. Cyanoprokaryota 1. Teil: Chroococcales; Ettl, H., Gärtner, G., Heynig, H., Mollenhauer, D., Eds.; Spektrum Akademischer Verlag, 1999. [Google Scholar]
- Nu¨bel, U.; Nu¨bel, N.; Garcia-Pichel, F.; Muyzer, G. PCR Primers To Amplify 16S RRNA Genes from Cyanobacteria. Appl Environ Microbiol 1997, 63, 3327–3332. [Google Scholar] [CrossRef]
- Neilan, B.A.; Jacobs, D.; Dot, T. Del; Blackall, L.L.; Hawkins, P.R.; Cox, P.T.; Goodman4, A.E. RRNA Sequences and Evolutionary Relationships among Toxic and Nontoxic Cyanobacteria of the Genus Microcystis. Int J Syst Bacteriol 1997, 693–697. [Google Scholar] [CrossRef]
- Dereeper, A.; Guignon, V.; Blanc, G.; Audic, S.; Buffet, S.; Chevenet, F.; Dufayard, J.F.; Guindon, S.; Lefort, V.; Lescot, M.; et al. Phylogeny. Fr: Robust Phylogenetic Analysis for the Non-Specialist. Nucleic Acids Res 2008, 36. [Google Scholar] [CrossRef]
- Jungblut, A.D.; Neilan, B.A. Molecular Identification and Evolution of the Cyclic Peptide Hepatotoxins, Microcystin and Nodularin, Synthetase Genes in Three Orders of Cyanobacteria. Arch Microbiol 2006, 185, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Lopes, V.R.; Ramos, V.; Martins, A.; Sousa, M.; Welker, M.; Antunes, A.; Vasconcelos, V.M. Phylogenetic, Chemical and Morphological Diversity of Cyanobacteria from Portuguese Temperate Estuaries. Mar Environ Res 2012, 73, 7–16. [Google Scholar] [CrossRef]
- Mihali, T.K.; Kellmann, R.; Muenchhoff, J.; Barrow, K.D.; Neilan, B.A. Characterization of the Gene Cluster Responsible for Cylindrospermopsin Biosynthesis. Appl Environ Microbiol 2008, 74, 716–722. [Google Scholar] [CrossRef]
- Rantala-Ylinen, A.; Känä, S.; Wang, H.; Rouhiainen, L.; Wahlsten, M.; Rizzi, E.; Berg, K.; Gugger, M.; Sivonen, K. Anatoxin-a Synthetase Gene Cluster of the Cyanobacterium Anabaena sp. Strain 37 and Molecular Methods to Detect Potential Producers. Appl Environ Microbiol 2011, 77, 7271–7278. [Google Scholar] [CrossRef] [PubMed]
- Saker, M.L.; Welker, M.; Vasconcelos, V.M. Multiplex PCR for the Detection of Toxigenic Cyanobacteria in Dietary Supplements Produced for Human Consumption. Appl Microbiol Biotechnol 2007, 73, 1136–1142. [Google Scholar] [CrossRef] [PubMed]
- Savela, H.; Spoof, L.; Perälä, N.; Preede, M.; Lamminmäki, U.; Nybom, S.; Häggqvist, K.; Meriluoto, J.; Vehniäinen, M. Detection of Cyanobacterial Sxt Genes and Paralytic Shellfish Toxins in Freshwater Lakes and Brackish Waters on Åland Islands, Finland. Harmful Algae 2015, 46, 1–10. [Google Scholar] [CrossRef]
- Hisbergues, M.; Christiansen, G.; Rouhiainen, L.; Sivonen, K.; Börner, T. PCR-Based Identification of Microcystin-Producing Genotypes of Different Cyanobacterial Genera. Arch Microbiol 2003, 180, 402–410. [Google Scholar] [CrossRef]
- Do Carmo Bittencourt-Oliveira, M.; Do Nascimento Moura, A.; De Oliveira, M.C.; Sidnei Massola, N. Geitlerinema Species (Oscillatoriales, Cyanobacteria) Revealed by Cellular Morphology, Ultrastructure, and DNA Sequencing. J Phycol 2009, 45, 716–725. [Google Scholar] [CrossRef]
- Bittencourt-Oliveira, M. do C.; Cabral De Oliveira, M.; Bolch, C.J.S. GENETIC Variability of Brazilian Strains of the Microcystis aeruginosa Complex (Cyanobacteria/Cyanophyceae) using the Phycocyanin Intergenic Spacer and Flanking Regions (Cpc BA) 1. J. Phycol 2001, 37, 810–818. [Google Scholar] [CrossRef]
- Haande, S.; Ballot, A.; Rohrlack, T.; Fastner, J.; Wiedner, C.; Edvardsen, B. Diversity of Microcystis aeruginosa Isolates (Chroococcales, Cyanobacteria) from East-African Water Bodies. Arch Microbiol 2007, 188, 15–25. [Google Scholar] [CrossRef]
- Otsuka, S.; Suda, S.; Li, R.; Matsumoto, S.; Watanabe, M.M. Morphological Variability of Colonies of Microcystis Morphospecies in Culture. J. Gen. Appl. Microbiol 2000, 46, 39–50. [Google Scholar] [CrossRef]
- Holt, J.G.; Krieg, N.R.; Sneath, P.H.A.; Staley, J.T.; Williams, S.T. Group 11. Oxygenic Phototrophic Bacteria. In Bergey’s Manual of Determinative Bacteriology; Hensyl, W.R., Ed.; Baltimore: Williams & Wilkins, 1994; pp. 377–425. [Google Scholar]
- Komárek, J. Recent Changes (2008) in Cyanobacteria Taxonomy Based on a Combination of Molecular Background with Phenotype and Ecological Consequences (Genus and Species Concept). Hydrobiologia 2010, 639, 245–259. [Google Scholar] [CrossRef]
- Lee, E.; Ryan, U.M.; Monis, P.; McGregor, G.B.; Bath, A.; Gordon, C.; Paparini, A. Polyphasic Identification of Cyanobacterial Isolates from Australia. Water Res 2014, 59, 248–261. [Google Scholar] [CrossRef] [PubMed]
- Litvaitis, M.K. A Molecular Test of Cyanobacterial Phylogeny: Inferences from Constraint Analyses. Hydrobiologia 2002, 468, 135–145. [Google Scholar] [CrossRef]
- Willame, R.; Boutte, C.; Grubisic, S.; Wilmotte, A.; Komárek, J.; Hoffmann, L. Morphological and Molecular Characterization of Planktonic Cyanobacteria from Belgium and Luxembourg. J Phycol 2006, 42, 1312–1332. [Google Scholar] [CrossRef]
- Komárek, J. Cyanobacterial Taxonomy: Current Problems and Prospects for the Integration of Traditional and Molecular Approaches. Algae 2006, 21, 349–375. [Google Scholar] [CrossRef]
- Fox, G.E.; Wisotzkey, J.D.; Jurtshuk, P.J. How Close Is Close: 16S RRNA Sequence Identity May Not Be Sufficient to Guarantee Species Identity. Int J Syst Bacteriol 1992, 42, 166–170. [Google Scholar] [CrossRef]
- Engene, N.; Cameron Coates, R.; Gerwick, W.H. 16S Rrna Gene Heterogeneity in the Filamentous Marine Cyanobacterial Genus Lyngbya. J Phycol 2010, 46, 591–601. [Google Scholar] [CrossRef]
- Fathalli, A.; Ben Rejeb Jenhani, A.; Moreira, C.; Welker, M.; Romdhane, M.; Antunes, A.; Vasconcelos, V. Molecular and Phylogenetic Characterization of Potentially Toxic Cyanobacteria in Tunisian Freshwaters. Syst Appl Microbiol 2011, 34, 303–310. [Google Scholar] [CrossRef]
- Premanandh, J.; Priya, B.; Teneva, I.; Dzhambazov, B.; Prabaharan, D.; Uma, L. Molecular Characterization of Marine Cyanobacteria from the Indian Subcontinent Deduced from Sequence Analysis of the Phycocyanin Operon (CpcB-IGS-CpcA) and 16S-23S ITS Region. The Journal of Microbiology 2006, 44, 607–616. [Google Scholar]
- Strunecky, O.; Pavlovna Ivanova, A.; Mares, J. An Updated Classification of Cyanobacterial Orders and Families Based on Phylogenomic and Polyphasic Analysis. J Phycol 2023, 59, 12–51. [Google Scholar] [CrossRef]
- Wanigatunge, R.P.; Magana-Arachchi, D.N.; Chandrasekharan, N. V.; Kulasooriya, S.A. Genetic Diversity and Molecular Phylogeny of Cyanobacteria from Sri Lanka Based on 16S RRNA Gene. Environmental Engineering Research 2014, 19, 317–329. [Google Scholar] [CrossRef]
- Domingos, P.; Rubim, T.K.; Molica, R.J.R.; Azevedo, S.M.F.O.; Carmichael, W.W. First Report of Microcystin Production by Picoplanktonic Cyanobacteria Isolated from a Northeast Brazilian Drinking Water Supply. Environ Toxicol 1999, 14, 3135. [Google Scholar] [CrossRef]
- Žegura, B. An Overview of the Mechanisms of Microcystin-LR Genotoxicity and Potential Carcinogenicity. Mini-Reviews in Medicinal Chemistry 2016, 16, 1042–1062. [Google Scholar] [CrossRef] [PubMed]
- Jakubowska, N.; Szeląg-Wasielewska, E. Toxic Picoplanktonic Cyanobacteria - Review. Mar Drugs 2015, 13, 1497–1518. [Google Scholar] [CrossRef] [PubMed]
- Śliwińska-Wilczewska, S.; Maculewicz, J.; Felpeto, A.B.; Latała, A. Allelopathic and Bloom-Forming Picocyanobacteria in a Changing World. Toxins (Basel) 2018, 10. [Google Scholar] [CrossRef]
- Schmidt, K.; Birchill, A.J.; Atkinson, A.; Brewin, R.J.W.; Clark, J.R.; Hickman, A.E.; Johns, D.G.; Lohan, M.C.; Milne, A.; Pardo, S.; et al. Increasing Picocyanobacteria Success in Shelf Waters Contributes to Long-Term Food Web Degradation. Glob Chang Biol 2020, 26, 5574–5587. [Google Scholar] [CrossRef]
- Mazur-Marzec, H.; Sutryk, K.; Kobos, J.; Hebel, A.; Hohlfeld, N.; Błaszczyk, A.; Toruńska, A.; Kaczkowska, M.J.; Łysiak-Pastuszak, E.; Kraśniewski, W.; et al. Occurrence of Cyanobacteria and Cyanotoxin in the Southern Baltic Proper. Filamentous Cyanobacteria versus Single-Celled Picocyanobacteria. Hydrobiologia 2013, 701, 235–252. [Google Scholar] [CrossRef]
- Stockner, J.; Callieri, C.; Cronberg, G. Picoplankton and Other Non-Bloom-Forming Cyanobacteria in Lakes. In The Ecology of Cyanobacteria: Their Diversity in Time and Space; Whitton, B.A., Potts, M., Eds.; Kluwer Academic Publishers: Dordrecht, Holland, 2000; pp. 195–231. [Google Scholar]
- Sorokin, Y.I.; Zakuskina, O.Y. Features of the Comacchio Ecosystem Transformed during Persistent Bloom of Picocyanobacteria. J Oceanogr 2010, 373–387. [Google Scholar] [CrossRef]
- Munari, C.; Rossi, R.; Mistri, M. Temporal Trends in Macrobenthos Community Structure and Redundancy in a Shallow Coastal Lagoon (Valli Di Comacchio, Northern Adriatic Sea). Hydrobiologia 2005, 550, 95–104. [Google Scholar] [CrossRef]
- Felpeto, A.B.; Śliwińska-Wilczewska, S.; Klin, M.; Konarzewska, Z.; Vasconcelos, V. Temperature-Dependent Impacts of Allelopathy on Growth, Pigment, and Lipid Content between a Subpolar Strain of Synechocystis sp. CCBA MA-01 and Coexisting Microalgae. Hydrobiologia 2019. [Google Scholar] [CrossRef]
- Zua, J.A.; Legrand, C.; Farnelid, H. Seasonal Dynamics in Picocyanobacterial Abundance and Clade Composition at Coastal and Offshore Stations in the Baltic Sea. Sci Rep 2022, 12, 14330. [Google Scholar] [CrossRef]
Figure 1.
Light microscope images of the filamentous isolated strain from Cheffia reservoir. a. G. amphibium CR1, b. L. nigra CR5, c. L. stagnina CR6, d. L. cincinnata CR7, e. Microcoleus sp. CR8, f. P. rosea CR9.
Figure 1.
Light microscope images of the filamentous isolated strain from Cheffia reservoir. a. G. amphibium CR1, b. L. nigra CR5, c. L. stagnina CR6, d. L. cincinnata CR7, e. Microcoleus sp. CR8, f. P. rosea CR9.
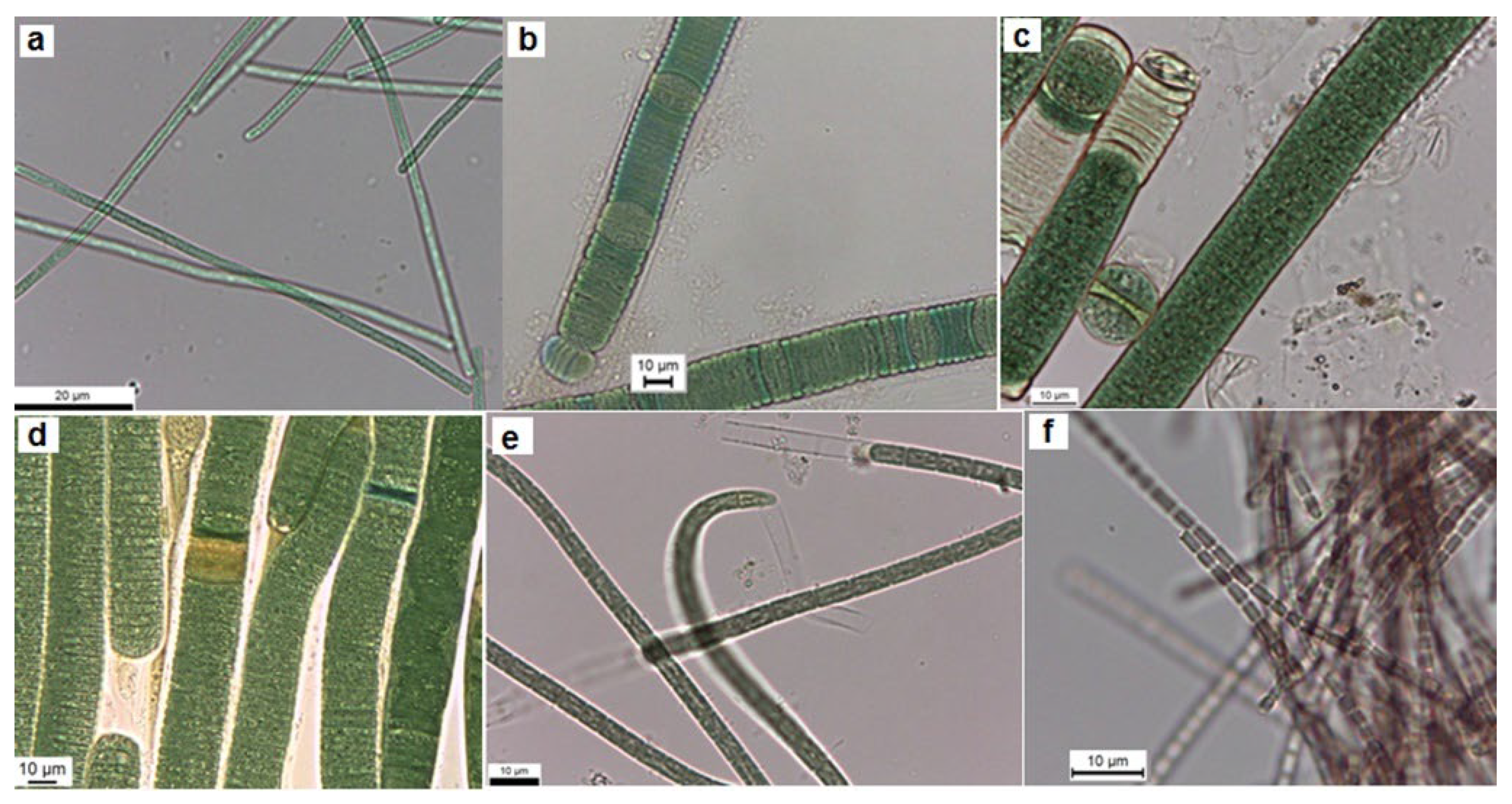
Figure 2.
Light microscope images of the colonial isolated strains from Cheffia reservoir. a. Aphanothece sp. CR10, b. Aphanothce sp. CR11, c. Microcystis sp. CR12, d. M. flos-aquae CR13.
Figure 2.
Light microscope images of the colonial isolated strains from Cheffia reservoir. a. Aphanothece sp. CR10, b. Aphanothce sp. CR11, c. Microcystis sp. CR12, d. M. flos-aquae CR13.
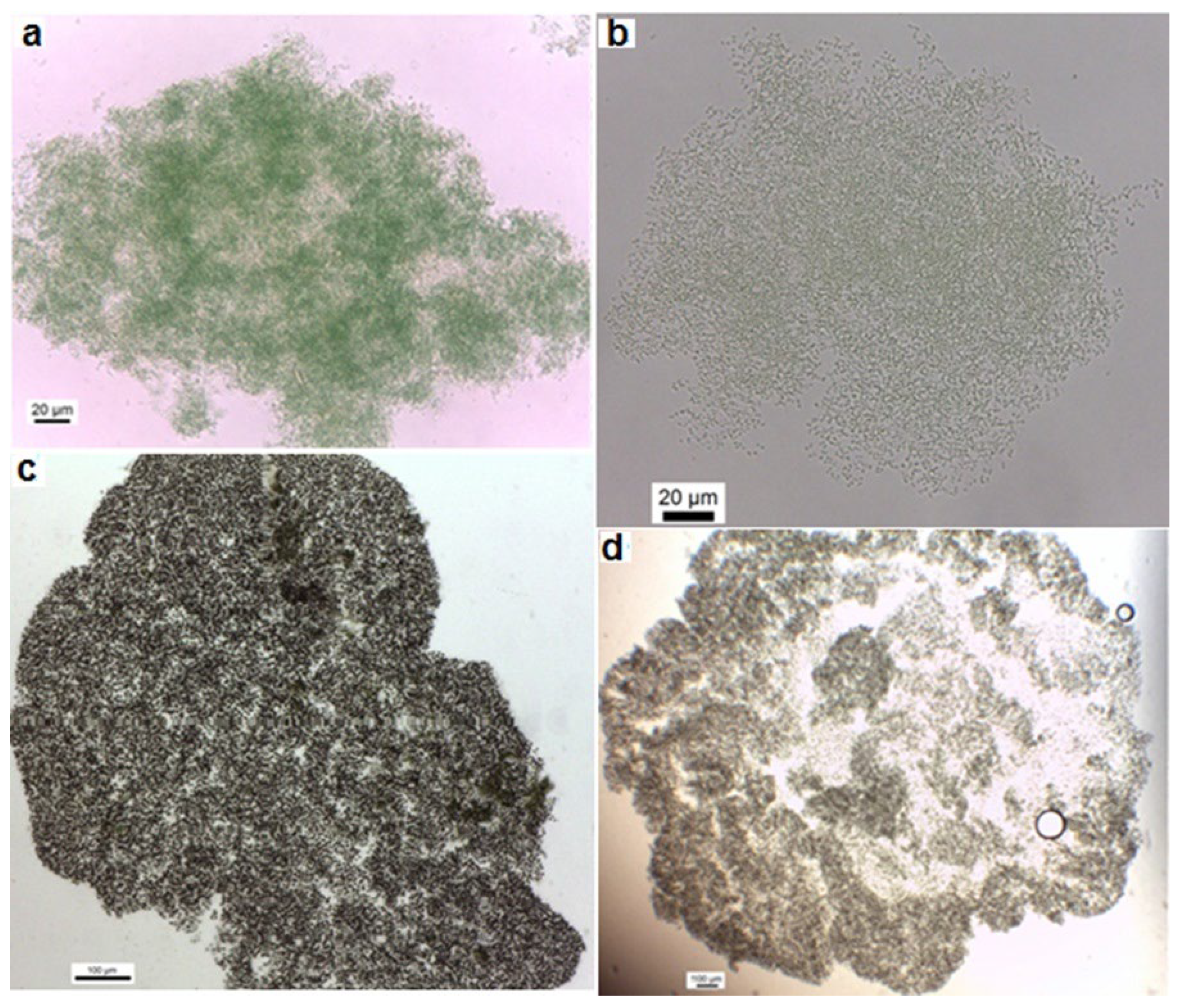
Figure 3.
Phylogenetic relationships of Cheffia reservoir cyanobacteria isolates with Genbank cyanobacteria strains based on 16S rRNA sequences. Escherichia coli (J01859) was used as outgroup. Isolates from this study are marked in pink.
Figure 3.
Phylogenetic relationships of Cheffia reservoir cyanobacteria isolates with Genbank cyanobacteria strains based on 16S rRNA sequences. Escherichia coli (J01859) was used as outgroup. Isolates from this study are marked in pink.
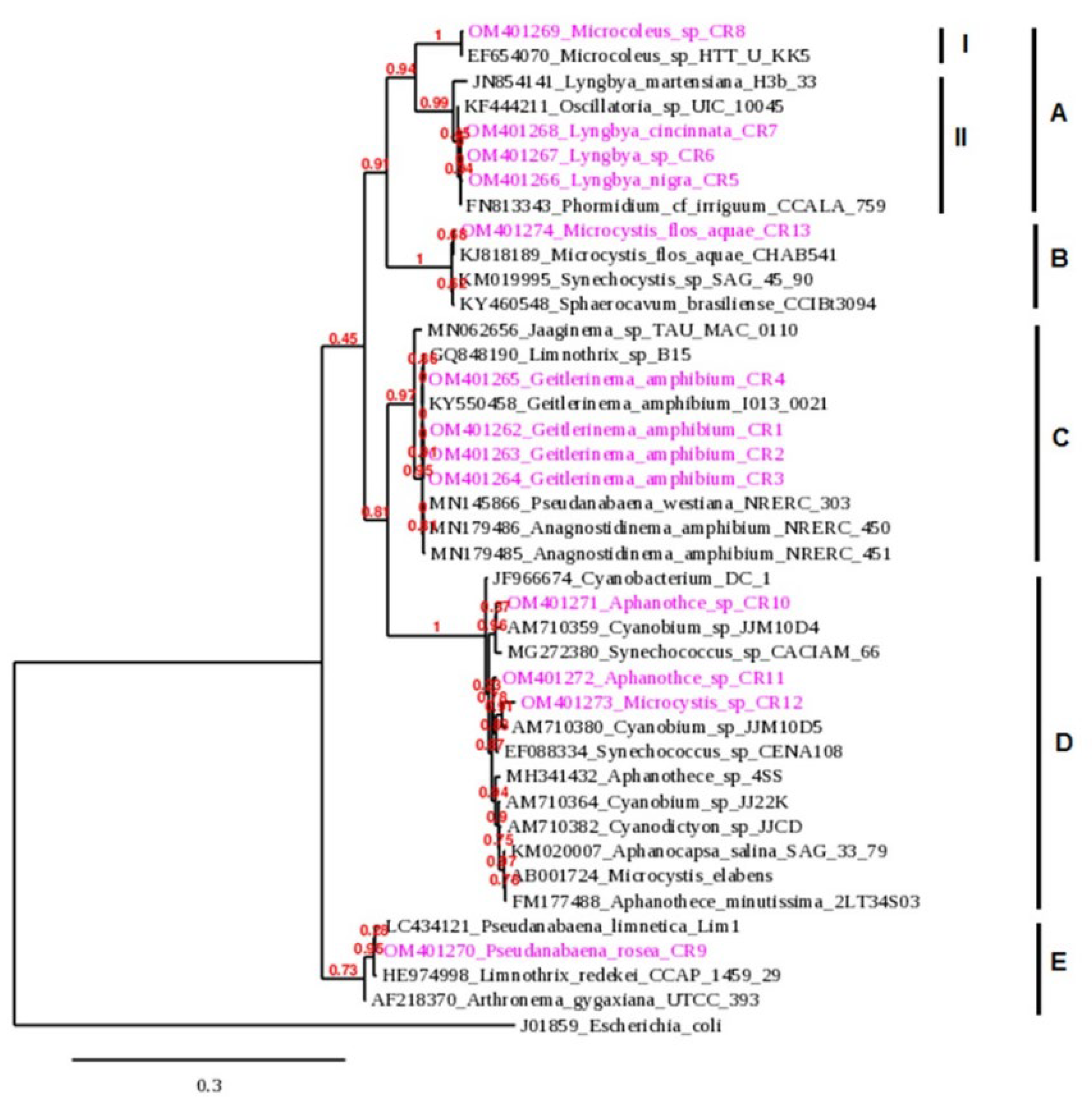
Figure 4.
Gel electrophoresis of the mcyE gene fragments obtained from isolated strains from Cheffia reservoir. M: Marker (DNA ladder 1 kb), + C: positive control, M. aeruginosa LEGE 91339; Lane 1: G. amphibium CR1; Lane 2: G. amphibium CR2; Lane 3: G. amphibium CR3; Lane 4: G. amphibium CR4; Lane 5: L. nigra CR5; Lane 6: L. stagnina CR6; Lane 7: L. cincinnata CR7; Lane 8: Microcoleus sp. CR8; Lane 9: P. rosea CR9; Lane 10: Aphanothce sp. CR10; Lane 11: Aphanothce sp. CR11; Lane 12: Microcystis sp. CR12; Lane 13: M. flos-aquae CR13; - C: negative control.
Figure 4.
Gel electrophoresis of the mcyE gene fragments obtained from isolated strains from Cheffia reservoir. M: Marker (DNA ladder 1 kb), + C: positive control, M. aeruginosa LEGE 91339; Lane 1: G. amphibium CR1; Lane 2: G. amphibium CR2; Lane 3: G. amphibium CR3; Lane 4: G. amphibium CR4; Lane 5: L. nigra CR5; Lane 6: L. stagnina CR6; Lane 7: L. cincinnata CR7; Lane 8: Microcoleus sp. CR8; Lane 9: P. rosea CR9; Lane 10: Aphanothce sp. CR10; Lane 11: Aphanothce sp. CR11; Lane 12: Microcystis sp. CR12; Lane 13: M. flos-aquae CR13; - C: negative control.


Table 1.
Primer sets used for PCR.
| Target | Primer pair | Sequence (5’−3’) | Size (bp) | Reference |
|---|---|---|---|---|
| mcyA | mcyA-Cd1F mcyA-Cd1R |
AAAATTAAAAGCCGTATCAAA AAAAGTGTTTTATTAGCGGCTCAT |
297 | [32] |
| mcyE | HEPF HEPR |
TTTGGGGTTAACTTTTTTGGGCATAGTC AATTCTTGAGGCTGTAAATCGGGTTT |
472 | [26] |
| sxtA | sxtA855F sxtA1480R |
GACTCGGCTTGTTGCTTCCCC GCCAAACTCGCAACAGGAGAAGG |
648 | [31] |
| sxtG | sxtG432F sxtG928R |
AATGGCAGATCGCAACCGCTAT ACATTCAACCCTGCCCATTCACT |
519 | [31] |
| sxtI | sxtI 682F sxtI 877R |
GGATCTCAAAGAAGATGGCA GCCAAACGCAGTACCACTT |
200 | [27] |
| cyrJ | cynsulF cylnamR | ACTTCTCTCCTTTCCCTATC GAGTGAAAATGCGTAGAACTTG |
584 | [28] |
| anaC | anaC-genF anaC-genR |
TCTGGTATTCAGTCCCCTCTAT CCCAATAGCCTGTCATCAA |
366 | [29] |
Table 2.
Morphological characteristics of the filamentous cyanobacterial strains isolated from Cheffia reservoir. L, length (lm); W, wide (µm).
Table 2.
Morphological characteristics of the filamentous cyanobacterial strains isolated from Cheffia reservoir. L, length (lm); W, wide (µm).
| Isolates | Trichome description | Cell description | Dimensions (µm) |
|
G. amphibium CR1; CR2; CR3; CR4 |
Bright blue-green trichomes, motile, without sheaths, more less straight, not constricted at cross walls | Cells cylindrical | L= 1.44–4.73 W= 1.38–2.92 |
| L. nigra CR5 | Straight or slightly curved trichomes, constricted at the cross-walls, with calyptras, isopolar with sheaths. Sheaths thin and firm open at the end, colorless or bluish | Cells short, discoid, with granular content and aerotopes | L= 1.95–5.15 W= 12.82–17.84 |
| L. stagnina CR6 | Trichomes isopolar. Sheaths thin and firm, distinct, colorless, open at the ends. Filaments constricted at cross-walls and motile by gliding | Cells grayish blue-green, distinctly shorter than wide | L= 1.9–5.2 W= 13.00–19.36 |
| L. cincinnata CR7 | Cylindrical, slightly waved trichomes, constrictions at cross wall, isopolar. Sheaths firm, thick, colorless, lamellated and opened | Cells discoid, cell content is granulated, aerotops are rare | L= 2.09–4.62 W= 13.46–19.66 |
| Microcoleus sp. CR8 | Wavy or screw-like coiled trichomes, constricted at cross walls, motile, isopolar and delimited by firm and thickened sheaths, homogenous, open at the ends, hyaline | Cells mostly isodiametric or slightly longer than wide with aerotopes. Apical cells are conical without calyptras | L= 3.09–6.02 W= 3.08–5.17 |
| P. rosea CR9 | Straight or slightly curved trichomes, reddish violet, without sheath, with constrictions at the cross walls | Cells cylindrical without aerotops | L= 1.67–3.89 W= 1.61–2.67 |
Table 3.
Morphological characteristics of the colonial cyanobacterial strains isolated from Cheffia reservoir.
Table 3.
Morphological characteristics of the colonial cyanobacterial strains isolated from Cheffia reservoir.
| Isolates | Colony description | Mucilage | Cell description | Cell diameter (µm) |
| Aphanothece sp. CR10 | Irregular, microscopic to macroscopic colonies, blue-green | Thin, colorless mucilage and distinct at the margin | Cells mainly spherical or oval, with gas vesicle, bright blue-green content, and delimited with individual envelopes. Envelopes are firm, colored in dark blue-green |
1.07–1.98 |
| Aphanothece sp. CR11 | Spherical colonies, rough in outline, microscopic, free-living, forming macroscopic granular agglomerations | Mucilage colorless and homogeneous, delimited at the margin, and follows the irregular outline of the colony, not diffluent, without a refractive outline | Cells spherical, pale or bright blue-green, slightly distant from one another, having fine granulation, without gas vesicles, and enveloped by thin individual layer | 0.90–1.36 |
|
Microcystis sp. CR12 |
Colonies macroscopic, lenticular, slightly elongate, three-dimensional, agglomerated in macroscopic, free-floating, gelatinous, blue-green masses | Narrow, colorless mucilage, distinctly delimited along cell agglomerations and forming refractive outline | Cells spherical, densely aggregated, with individual envelopes, content is homogeneous, olive green or brownish with aerotopes | 4.02–5.97 |
| M. flos-aquae CR13 | Spherical colonies, with irregular margin, microscopic to macroscopic, free-floating, compact, or clathrate, with densely irregularly arranged cells gathered in small agglomerations | Mucilage colorless, slightly distant from cell clusters, and delimited by slightly refractive outline | Cells spherical or hemispherical after division, with individual thick envelopes. Cell content appears granular, olive green, or brownish, with aerotopes |
3.98–5.77 |
Table 4.
16S rRNA gene-sequence-based identity (%) between cyanobacterial isolates from Cheffia reservoir and their closest match available in Genbank (NCBI).
Table 4.
16S rRNA gene-sequence-based identity (%) between cyanobacterial isolates from Cheffia reservoir and their closest match available in Genbank (NCBI).
| Isolates | Closest Match (Accession Number) | Query coverage (%) | Percent identity (%) |
|---|---|---|---|
| G. amphibium CR1 | Geitlerinema amphibium I013-0021 (KY550458) | 82 | 99.39 |
| Anagnostidinema amphibium NRERC-450 (MN179486) | 98 | 99.13 | |
| Pseudanabaena westiana NRERC-303 (MN145866) | 98 | 99.13 | |
| Limnothrix sp. B15 (GQ848190) | 98 | 99.13 | |
| G. amphibium CR2 | Geitlerinema amphibium I013-0021 (KY550458) | 83 | 99.74 |
| Pseudanabaena westiana NRERC-303 (MN145866) | 99 | 99.56 | |
| Limnothrix sp. B15 (GQ848190) | 100 | 99.56 | |
| Anagnostidinema amphibium NRERC-451 (MN179485) | 100 | 99.49 | |
| Jaaginema sp. TAU-MAC 0110 (MN062656) | 96 | 97.59 | |
| G. amphibium CR3 | Geitlerinema amphibium I013-0021 (KY550458) | 85 | 99.91 |
| Anagnostidinema amphibium NRERC-450 (MN179486) | 100 | 99.70 | |
| Pseudanabaena westiana NRERC-303 (MN145866) | 100 | 99.70 | |
| Limnothrix sp. B15 (GQ848190) | 100 | 99.70 | |
| Jaaginema sp. TAU-MAC 0110 (MN062656) | 97 | 97.71 | |
| G. amphibium CR4 | Geitlerinema amphibium I013-0021 (KY550458) | 85 | 99.91 |
| Limnothrix sp. B15 (GQ848190) | 100 | 99.85 | |
| Anagnostidinema amphibium NRERC-450 (MN179486) | 100 | 99.70 | |
| Pseudanabaena westiana NRERC-303 (MN145866) | 100 | 99.70 | |
| Jaaginema sp. TAU-MAC 0110 (MN062656) | 98 | 97.73 | |
| L. nigra CR5 | Phormidium cf. irriguum CCALA 759 (FN813343) | 99 | 99.85 |
| Oscillatoria sp. UIC 10045 (KF444211) | 93 | 99.35 | |
| Lyngbya martensiana H3b/33 (JN854141) | 95 | 97.78 | |
| L. stagnina CR6 | Phormidium cf. irriguum CCALA 759 (FN813343) | 97 | 99.85 |
| Oscillatoria sp. UIC 10045 (KF444211) | 90 | 99.28 | |
| Lyngbya martensiana H3b/33 (JN854141) | 92 | 97.73 | |
| L. cincinnata CR7 | Phormidium cf. irriguum CCALA 759 (FN813343) | 97 | 99.63 |
| Oscillatoria sp. UIC 10045 (KF444211) | 91 | 99.04 | |
| Lyngbya martensiana H3b/33 (JN854141) | 93 | 97.50 | |
| Microcoleus sp. CR8 | Microcoleus sp. HTT-U-KK5 (EF654070) | 99 | 99.40 |
| P. rosea CR9 | Pseudanabaena limnetica Lim1 (LC434121) | 100 | 99.27 |
| Limnothrix redekei CCAP 1459/29 (HE974998) | 100 | 99.05 | |
| Arthronema gygaxiana UTCC 393 (AF218370) | 100 | 98.39 | |
| Aphanothece sp. CR10 | Synechococcus sp. CACIAM 66 (MG272380) | 100 | 99.11 |
| Cyanobium sp. JJM10D4 (AM710359) | 100 | 98.96 | |
| Aphanothece minutissima 2LT34S03 (FM177488) | 100 | 97.41 | |
| Microcystis elabens (AB001724) | 100 | 97.26 | |
| Aphanothece sp. 4SS (MH341432) | 86 | 97.79 | |
| Aphanothce sp. CR11 | Synechococcus sp. CENA108 (EF088334) | 100 | 98.96 |
| Cyanobium sp. JJ22K (AM710364) | 100 | 98.30 | |
| Cyanobacterium DC-1 (JF966674) | 100 | 98.30 | |
| Cyanodictyon sp. JJCD (AM710382) | 100 | 98.22 | |
| Aphanocapsa salina SAG 33.79 (KM020007) | 100 | 98.07 | |
| Aphanothece sp. 4SS (MH341432) | 87 | 97.89 | |
| Microcystis sp. CR12 | Synechococcus sp. CENA108 (EF088334) | 100 | 98.08 |
| Cyanobium sp. JJM10D5 (AM710380) | 100 | 98.00 | |
| M. flos-aquae CR13 | Microcystis flos-aquae CHAB541 (KJ818189) | 100 | 99.77 |
| Sphaerocavum brasiliense CCIBt3094 (KY460548) | 100 | 99.32 | |
| Synechocystis sp. SAG 45.90 (KM019995) | 100 | 99.47 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



