
Submitted:
04 October 2023
Posted:
06 October 2023
You are already at the latest version
Abstract
Transcriptional regulation constitutes a key step in gene expression regulation. Myocyte enhancer factor 2C (MEF2C) is a transcription factor of MADS box family involved in the early development of several cell types including muscle cells. Over the last decade a novel layer of complexity modulating gene regulation is emerging as non-coding RNAs have been identified, impacting in both transcriptional and post-transcriptional regulation. microRNAs represent the most studied and abundantly expressed subtype of small non-coding RNAs and their functional role have been widely documented. On the other hand, our knowledge on the transcriptional and post-transcriptional regulatory mechanisms that drive microRNA expression is still incipient. We recently demonstrated that MEF2C is able to transactive the long, but not short, regulatory element upstream of miR-23a-miR-27a-miR-24-2 transcriptional start site. However, MEF2C over-expression and silencing, respectively, displayed distinct effects on each of the miR-23a-miR-27a-miR-24-2 mature cluster members, without affecting pri-miRNA expression levels, supporting thus additional MEF2C-driven regulatory mechanisms. Within this study we demonstrated a complex post-transcriptional regulatory mechanisms directed by MEF2C in the regulation of miR-23a-miR-27a-miR-24-2 cluster members, distinctly involving different domains of the MEF2C transcription factor and the physical interaction with pre-miRNAs and RNA interacting proteins such as Ksrp, HnRNPa3 and Ddx17.
Keywords:
Mef2c
; microRNAs
; RNPs
Introduction
Transcriptional regulation constitutes a key step in gene expression regulation. Multiple types of transcription factors have been identified from flies to humans, regulating multiple developmental, homeostatic and pathological processes [1,2,3]. In this context, a core of transcription factors has been identified to play essential roles in myogenesis, such as SRF, NKX2.5, GATA4 and MEF2C [4]. Myocyte enhancer factor 2C (MEF2C) is a transcription factor of MADS box family involved in the early development of several cell types, including neural, immune, cartilaginous and endothelial cells, yet the main role of MEF2C is exerted by regulating muscle development (i.e., skeletal, cardiac and smooth) [5,6,7,8,9,10,11]. MEF2C deficient mice are embryonic lethal displaying complex cardiovascular defects as the early heart tube does not undergoes looping morphogenesis and thus the future right ventricle is absent [12,13]. Importantly, MEF2C also plays a pivotal role in cardiac pathological conditions such as cardiac hypertrophy [13] and it represents an essential cornerstone for cardiac reprogramming [15,16].
Multiple studies have reported the essential role of MEF2C regulating gene expression in different biological contexts, including cardiac [17,18,19,20,21], skeletal [22,23,24] and smooth muscle [25] cells. Transcriptional activity of MEF2C is dependent of the carboxyl terminal, a process that is sumoylation-dependent [26], whereas the MADS and MEF2 domains mediate DNA binding, dimerization and co-factor interactions [27,28,29]. Adjacent to the MEF2 domain is the HJURP-C (Holliday junction recognition protein C-termial) domain, followed by two transcriptional activation domains (TAD1 and TAD2), which are responsible for transcriptional activation [29]. Although different MEF2C isoforms have been reported, their functional role remains rather elusive [30].
Over the last decade a novel layer of complexity modulating gene regulation is emerging as non-coding RNAs have been identified, impacting in both transcriptional and post-transcriptional regulation. Non-coding RNAs are broadly classified according to their transcript length into small non-coding RNAs (<200 nt) and long non-coding RNAs (>200 nt) [31]. Among the small non-coding RNAs, microRNAs represent the most studied and abundantly expressed subtype. MicroRNAs are nuclearly encoded and transcribed into microRNA precursor molecules by RNA polymerase II. In certain genomic localization, microRNAs are clustered in such a way that the primary transcript contains multiple microRNA precursors, leading to a pri-mRNA precursor. Pri-miRNAs are then processed by nucleases such as Drosha and Dgcr8 to generate distinct pre-miRNA molecules, that are subsequently exported by exportin-5/Ran protein complex to the cytoplasm [32]. Within the cytoplasm the pre-miRNAs are processed into a mature microRNAs duplex by Dicer RNAse and loaded into the RISC complex. Within the RISC complex, one of the strands of the double-stranded microRNA molecule is subsequently degraded and the remaining one conforms the mature microRNA that is now able to scan RNA molecules for sequence homology of its seed sequence leading to post-transcriptional RNA target cleavage, translation repression and/or RNA deadenylation. Thus, final output consequence, in most cases, is a decrease on the miRNA/protein target abundance [33]. Importantly, there are emerging evidences that certain microRNAs can also modulate transcriptional regulation, by exerting their function within the nucleus, impacting thus on alternative splicing and RNA and microRNA transcriptional regulation [34].
microRNAs are highly conserved during evolution, ranging from C. elegans to humans. microRNAs display temporal and spatial differential expression during embryonic and adulthood, contributing thus to both embryonic development and tissue homeostasis [35]. Impaired expression and/or function of microRNAs thus have been recently reported to lead to pathological conditions [36,37,38,39,40]. In this context, several microRNAs have been reported to be regulated by MEF2C [41] and similarly several microRNAs also modulate MEF2C expression in different biological contexts [42,43,44,45,46,47,48].
The functional role of microRNAs during cardiac development and diseases have been extensively documented [36,37,38,39,49]. Within this context, genetic deletion of miR-1-2 [50,51] and miR-126 [52,53], respectively, have revealed the essential role of these miRNAs in cardiac and vascular embryonic development. Importantly, several clustered microRNAs such as miR-19-72 [54,55,56], miR-106a-363 [57], miR-106b-25 [58,59] and miR-23/27/24 [60,61,62], have been reported to play modulatory roles in distinct biological contexts, including the cardiovascular system [59,60,63,64,65]. While our current understanding of the detailed functional relevance of each of these microRNAs is progressively emerging, our knowledge on the transcriptional and post-transcriptional regulatory mechanisms that drive microRNA expression is still incipient.
Lee et al. [66] demonstrated that miR-23a-miR-27a-miR-24-2 cluster is transcribed as an RNA polymerase II-dependent primary transcript whose main transcriptional regulation is driven by a −600 bp upstream promoter. We subsequently reported the identification of upstream regulatory elements driving miR-23a-miR-27a-miR-24-2 transcriptional regulation in both cardiac and skeletal muscle cells [67]. Within this context, we demonstrated that MEF2C is able to transactive the long (-1830 to +1 nt) regulatory element but not the short (-776 to +1 nt) element, in accordance with the identification of MEF2 regulatory binding sites distribution. However, MEF2C over-expression and silencing, respectively, displayed distinct effects on each of the miR-23a-miR-27a-miR-24-2 cluster members, without affecting pri-miRNA expression levels in different cell types [67], supporting thus additional MEF2C-driven regulatory mechanisms. Within this study we report complex transcriptional and post-transcriptional regulatory mechanisms directed by MEF2C in the regulation of miR-23a-miR-27a-miR-24-2 cluster, distinctly involving different domains of the MEF2C transcription factor and the physical interaction with pre-miRNAs and RNA interacting proteins such as Ksrp, HnRNPa3 and Ddx17.
Results
We have previously characterized the transcriptional potential of the 1.8 Kb upstream sequences of the miR-23a-miR-27a-miR-24-2 cluster and reported that MEF2C is capable of transcriptionally activate these regulatory regions in HL1 atrial cardiomyocytes. Such transcriptional activation enhances thus the expression of the miR-23a-miR-27a-miR-24-2 pri-miRNA. However, we have also previously reported that MEF2C overexpression and inhibition, respectively, distinctly regulate the expression of each of the miR-23a-miR-27a-miR-24-2 mature cluster members, independently of its transcriptional potential. Importantly, modulation of miR-23a-miR-27a-miR-24-2 cluster members by MEF2C is tissue-specific. Therefore, our previous data suggest that MEF2C modulation of miR-23a-miR-27a-miR-24-2 cluster members is exerted by either direct or indirect post-transcriptional mechanisms. Since microRNAs have been recently reported to exert both cytoplasmic and well as nuclear function, we firstly explored the subcellular localization of the miR-23a-miR-27a-miR-24-2 mature cluster members. qRT-PCR of nuclear and cytoplasmic fractions revealed that all three members, i.e., miR-23a_3p, miR-27a_3p and miR-24_3p are similarly localized in both subcellular compartments in HL1 cardiomyocytes (Figure 1A), in contrast to miR-130a that is preferentially and significantly enhanced within the cytoplasm (Figure 1A), while Xist2 is enhanced in the nucleus (Figure 1A), serving as internal subcellular fractioning controls.
Subsequently we therefore tested whether MEF2C modulation of miR-23a-miR-27a-miR-24-2 cluster members is exerted by either direct or indirect post-transcriptional mechanisms. We first explored the plausible interaction between MEF2C with miR-23a-miR-27a-miR-24-2 cluster members by performing MEF2C protein pull-down assays (Supplementary Figure S1A). Our data demonstrated that MEF2C interacts with pre-miR-23a and pre-miR-27a but not to pre-miR-24-2 (Figure 1B). Importantly, MEF2C neither binds to the mature microRNAs, i.e., miR-23a_3p, miR-27a_3p and miR-24_3p (Figure 1C), nor to the miR-23a-miR-27a-miR-24-2 pri-miRNA (Figure 1D), demonstrating a direct post-transcriptional role of MEF2C governing the expression of the precursor forms of miR-23a_3p and miR-27a_3p but not miR-24_3p (Figure 1E).
We subsequently tested which part of the MEF2C transcription factor is exerting pre-miR-23a and pre-miR-27a modulation and whether it also affects the expression of the mature microRNA cluster members. For this purpose, we constructed two distinct MEF2C variants. The first one lacks the SRF-type DNA-binding and dimerization domain, the MADS_MEF2_like and the HJURP_C domain at the 5´end (MEF2C 5´del) (Supplementary Figure S1B). The second one lacks the 3´end (MEF2C 3´del) and thus maintained these domains (Supplementary Figure S1B). In addition, we also performed MEF2C overexpression and silencing studies. MEF2C over-expression and silencing were successfully achieved. In addition, the overexpression of MEF2C full-length, MEF2C 5´del, and MEF2C 3´del show similar levels (Supplementary Figure S1C). Furthermore, transactivation assays of the L regulatory element of the miR-23a-miR-27a-miR-24-2 locus was successfully achieved with MEF2C full-length and MEF2C 3´del constructs, but not with MEF2C 5´del, as expected, since the latter lacks DNA-binding and dimerization domains (Supplementary Figure S1D).
qRT-PCR analyses of precursor pre-miR-23a, pre-miR-27a and pre-miR-24-2 demonstrate that both MEF2C 5´del and MEF2C 3´del significantly increase the steady state levels of pre-miR-23a_3p, pre-miR-27a_3p but not of pre-miR-24_3p (Figure 2A) while over-expression of full length MEF2C and silencing resulted in minimal but significant downregulation of pre-miR-23a, premiR-27a but not of pre-miR-24-2 (Figure 2A). Thus, these data reinforced the previous observations by MEF2C pulldown demonstrating a regulatory role on pre-miR-23a, premiR-27a but not of pre-miR-24-2. qRT-PCR analyses of mature miR-23a-miR-27a-miR-24-2 cluster members demonstrate that MEF2C full-length over-expression significantly down-regulates miR-23a_3p, miR-27a_3p but not miR-24_3p expression (Figure 2B) while MEF2C silencing selectively downregulates only miR-27a_3p. Importantly, MEF2C 3´del and MEF2C 5´del significantly up-regulated all mature microRNA cluster members, i.e., miR-23a_3p, miR-27a_3p and miR-24_3p (Figure 2B).
Therefore, our data indicates that both 5´end and 3´end of MEF2C regions play inhibitory roles to modulate mature miR-23a-miR-27a-miR-24-2 cluster members. Moreover, mature miR-24_3p expression is modulated by both MEF2C 5´del and MEF2C 3´del but not its precursor form (pre-miR-24-2) (Figure 2C), suggesting a dual role for MEF2C regulating such post-transcriptional events (Figure 2A and 2B), yet it should be taken into account that miR-24-1 precursor, located in a distinct chromosomal locus, might also contribute to miR-24_3p expression levels.
Modulation of miR-23a-miR-27a-miR-24-2 cluster members by MEF2C can also be indirectly exerted by association with ribonucleic proteins (RNPs). We therefore tested the tissue-specific expression of distinct RNPs previously reported to interact with distinct microRNAs [66,67,68,69] in different cell types, showing differential regulation of mature miR-23a-miR-27a-miR-24-2 cluster members [65]. Therefore, we evaluated expression of seven distinct RNPs (Adar1, Ddx5, Ddx17, HnRNPa1, HnRNPa3, HnRNPa2b1, Ksrp) in 3T3 fibroblasts, HL1 atrial cardiomyocytes and Sol8 skeletal myoblasts.
Adar1, Ddx5 and HnRNPa1 displayed increased expression in 3T3 fibroblasts as compared to HL1 atrial cardiomyocytes and Sol8 skeletal myoblasts (Figure 3A). Ksrp displayed similarly enhanced expression in 3T3 fibroblasts and HL1 atrial cardiomyocytes as compared to Sol8 skeletal myoblasts (Figure 3A). HnRNPa2b1 and Ddx17 a similar expression pattern with enhanced expression in HL1 cardiomyocytes, while HhRNPa3 displayed the opposite pattern, i.e., decreased expression in HL1 atrial cardiomyocytes as compared to 3T3 fibroblasts and Sol8 skeletal myoblasts (Figure 3A). Overall, these data showed that the RNA constituents of all mentioned RNPs are expressed in these three distinct cell lines tested. However, the distinct RNPs showed a differential expression in these cell lines, supporting thus a plausible contribution of these RNPs regulating the distinct miR-23a-miR-27a-miR-24-2 cluster members by MEF2C in different cell types, as previously demonstrated [65].
We also tested whether these RNP transcripts are distinctly distributed within the subcellular compartments in HL1 cardiomyocytes. Our data revealed that Adar1 is highly enriched in the nuclear compartment whereas Ddx5, Ddx17 and Ksrp are prominently localized in the cytoplasm. On the other hand, HnRNPa1, HnRNPa3 and HnRNPa2b1 are similarly distributed within both nuclear and cytoplasmic compartments, in line with MEF2C mRNA distribution (Figure 3B).
We additionally tested if these RNPs are regulated by MEF2C. Over-expression of MEF2C full length resulted in up-regulation of Adar1 and down-regulation of HnRNPa3 and Ksrp, while Ddx5, Ddx17, HnRNPa1 and HnRNPa2b1 were not altered (Figure 3C). MEF2C silencing decreased Ddx17, HnRNPa3, HnRNPa2b1 and Ksrp while increased HnRNPa1. Adar1 and Ddx5 display no significant differences (Figure 3C). MEF2C 5´del significantly increased Adar1 and significantly downregulated Ddx5, Ddx17, HnRNPa1, HnRNPa3, HnRNPa2b1 and Ksrp (Figure 3C), while MEF2C 3´del significantly increased Adar1, HnRNPa1, HnRNPa3 while Ddx5, Ddx17 and Ksrp display significant downregulation (Figure 3C). HnRNPa2b1 displayed no significant differences after MEF2C 3´del over-expression (Figure 3C). In sum, our data demonstrate that these RNPs are distinctly modulated by MEF2C. Particularly, it is important to highlight that Ksrp is similarly down-regulated in all experimental conditions, Ddx17 is down-regulated in MEF2C 3´del and MEF2C 5´del and MEF2C siRNAs conditions, while HnRNPa3 is downregulated by MEF2C full-length, MEF2C siRNA and MEF2C 3´del. Furthermore, Ddx5, Ddx17 and Ksrp are downregulated by MEF2C 3´del and MEF2C 5´del over-expression (Figure 3D), supporting a plausible role of these RNPs, distinctly regulating miR-23a-miR-27a-miR-24-2 cluster members and/or its precursors.
We subsequently tested whether MEF2C can interact with these RNPs. MEF2C protein pulldown demonstrate that Ddx17, Ksrp and HnRNPa3 are interacting with MEF2C (Figure 4A), while no significant interaction was observed for HnRNPa1, HnRNPa2b1, Ddx5 and Adar1, respectively (Figure 4A).
Finally, we performed RNPs silencing assays in order to dissect if RNP inhibition can modulate miR-23a-miR-27a-miR-24-2 cluster members. Ddx17, HnRNPa1 and HnRNPa2b1 siRNA administration does not significantly modulate miR-23a_3p, miR-27a_3p and miR-24_3p pre-miRNAs, respectively, except for HnRNPa1 that significantly up-regulates pre-miR-27a and pre-miR-24-2 (Figure 4B and 4D). On the other hand, Ddx5, HnRNPa3 and Ksrp inhibition enhanced miR-23a_3p, miR-27a_3p, miR-24_3p pre-miRNAs, except pre-miR-24-2 after Ksrp inhibition (Figure 4B and 4D). For the mature miR-23a-miR-27a-miR-24-2 cluster members, Ddx17 and HnRNPa1 silencing significantly up-regulated while Ddx5 significantly downregulated all miR-23a-miR-27a-miR-24-2 cluster members, i.e., miR-23a_3p, miR-27a_3p and miR-24_3p (Figure 4C and 4D). On the other hand, HnRNPa2b1 inhibition exclusively upregulates miR-23a_3p, but not miR-27a_3p and miR-24_3p while HnRNPa3 silencing leads to downregulation of miR-23a_3p, upregulation of miR-27a_3p and no significant modulation of miR-24_3p (Figure 4C and 4D). Finally, Ksrp silencing leads to downregulation of all miR-23a-miR-27a-miR-24-2 cluster members, i.e., miR-23a_3p, miR-27a_3p and miR-24_3p (Figure 4C and 4D). Thus, these observations revealed that Ddx5, HnRNPa3 and Ksrp are essential primarily for pre-miR-23a and pre-miR-27a, and to a lesser extend for pre-miR-24-2 (only Ddx5 and HnRNPa3), supporting a key role modulating differential expression of the miR-23a-miR-27a-miR-24-2 cluster members. Similarly, HnRNPa2b1, HnRNPa3 and Ksrp silencing also distinct modulate mature miR-23a-miR-27a-miR-24-2 cluster members. In sum, these data illustrate that distinct RNPs can impact on differential pre-miRNA and mature miR-23a-miR-27a-miR-24-2 cluster members expression.
Discussion
Within the last decade, our understanding of the functional role of distinct microRNAs has greatly emerged, however our knowledge on the transcriptional and post-transcriptional regulatory mechanisms driving microRNA expression is still incipient. We previously demonstrated that MEF2C over-expression and silencing, respectively, displayed distinct effects on each of the mature miR-23a-miR-27a-miR-24-2 cluster members [67], supporting thus additional MEF2C-driven regulatory mechanisms. We provide herein evidences that MEF2C can directly bind to pre-miR23a and pre-miR-27a, but not to pre-miR-24-2. Importantly, MEF2C does not directly bind to either the pri-miRNA miR-23a-miR-27a-24-2 precursor nor to the mature miR-23a_3p, miR-27a_3p and miR-24_3p molecules. Furthermore, we also demonstrated that distinct MEF2C domains can differentially modulate both pre-miRNAs and microRNAs expression. While there are emerging evidences that distinct proteins can influence MEF2C expression levels leading to sumoylation and caspase cleavage [26,72], this is to the best of our knowledge the first proof that a transcription factor can influence microRNA biogenesis by directly interacting with pre-miRNA molecules.
On the other hand, ample evidences are reported on the key role of distinct ribonucleoproteins (RNPs) modulating microRNA expression [73,74,75,76,77,78]. Thus, to further support the plausible role of several of these RNPs in MEF2C-driven miR-23a-miR-27a-miR-24-2 expression, we analyzed the expression of seven distinct RNPs in three distinct cell types (fibroblasts, cardiomyocytes and skeletal muscle myoblasts) demonstrating that all of them are indeed expressed while displaying cell type enrichment, i.e., Ddx17 and HnRNPa2b1 are more abundantly expressed in cardiomyocytes, while Adar1, Ddx5 and HnRNPa3 are widely expressed in fibroblasts. Furthermore, we demonstrate that these RNPs displayed distinct subcellular distribution, i.e., Adar1 is primarily in the nucleus, Ddx5, Ddx17 and Ksrp are primarily in the cytoplasm while HnRNPa1, HnRNPa2b1 and HnRNPa3 are both nuclear and cytoplasmic, in line with previous reports [79,80,81,82,83,84]. Importantly, we firstly demonstrated that mature miR-23a-miR-27a-miR-24-2 cluster microRNA members are equally distributed in both nuclear and cytoplasmic subcellular compartments, support the notion that they might exert distinct functional roles, as recently reported [85,86,87] and thus can be distinctly regulated in the cytoplasm vs. the nucleus. Furthermore these data also support that distinct RNPs might be involved in differential and subcellular compartment-specific miR-23a-miR-27a-miR-24-2 cluster members expression.
Scarce evidences have been reported for transcription factors directly binding to RNPs [88], supporting a plausible role in post-transcriptional regulation. For MEF2C, only AUF1 binding have been reported, promoting skeletal muscle myogenesis [84]. Within this study we report for the first time that MEF2C can directly bind to Ddx17, HnRNPa3 and Ksrp mRNAs, respectively. Additionally, MEF2C indirectly regulates Adar1 and HnRNPa2b1 expression. Furthermore, we also demonstrate that distinct MEF2C domains differently contribute to RNPs expression. In this context, both 5´and 3´MEF2C ends can selectively inhibit Ddx5, Ddx17 and Ksrp expression, while enhance Adar1 expression. On the other hand, HnRNPa1, HnRNPa3 and HnRNPa2b1 are distinctly regulated by MEF2C 3´and 5´ends respectively. While additional studies are required to fully understand the molecular mechanisms directing MEF2C 3´and 5´ends modulation of these RNPs, our data support the notion that they might be transcriptionally regulated since the MEF2C 5´del construct, lacking transcriptional potential (Supplementary Figure S1D), primarily downregulates their expression while MEF2C 3´del constructs displays the opposite pattern. In sum our data demonstrate that MEF2C can directly and indirectly regulate distinct RNPs in cardiomyocytes, with potential impact on miR-23a-miR-27a-miR-24-2 cluster members expression.
As previously stated, a large body of evidences have been reported on RNPs modulating microRNA expression [76,77,78,79,80,81], yet their role in differential microRNAs cluster expression has only been reported for Adar1 [85,86,87,88] and Ksrp [94,95,96]. Several studies reported miR-27b regulating Ksrp expression in distinct biological settings [97,98,99], but no proofs have been reported for miR-27b to be regulated by Ksrp. miR-27b regulation by RNPs have only been reported for HnRNPa1 in colorectal cancer [100] and HnRNPa2b1 in preeclampsia [101]. Importantly no data have been reported for the involvement of these RNPs in the regulation of the miR-23a_3p or miR-24, as well as for any of the miR-23a-miR-27a-miR-24-2 cluster members, except for Ksrp regulating miR-23a_3p [102] and Adar1 regulating pre-miR-27a_3p to mature miR-27a_3p processing in cancer [103].
We provide herein evidences that silencing Krsp selectively upregulates pre-miR-23a and pre-miR-27a but not pre-miR-24-2 expression. Similarly, silencing HnRNPa1 leads to pre-miR-27a and pre-miR-24-2 downregulation without affecting pre-miR-23a expression. For the mature microRNAs, Ksrp and Ddx5 inhibition diminished all mature miR-23a-miR-27a-miR-24-2 cluster members, while selective inhibition of Ddx17 and HhRNPa1 enhanced all mature miR-23a-miR-27a-miR-24-2 cluster members. Curiously, HnRNPab1 silencing, selectively upregulates miR-23a_3p but not miR-27a_3p and miR-24_3p, while HnRNPa3 silencing upregulates miR-23a_3p and downregulates miR-27a_3p and miR-24_3p. It is important nonetheless in this context that miR-24 levels, might results for the amplification of the mature miR-24_3p from both pre-miR-24-1 and pre-miR-24-2 precursors, as previously mentioned. Importantly, MEF2C directly interacts with Ddx17, HnRNPa3 and Ksrp, and MEF2C silencing is essential for proper Ddx17, HnRNPa1, HnRNPa2b1, HnRNPa3 and Ksrp, also proving proofs of the differential role of the MEF2C C-terminal and N-terminal on this regulation.
Overall, these data demonstrate a complex and pivotal role of distinct RNPs regulating miR-23a-miR-27a-miR-24-2 cluster members, and support the notion that distinct RNPs, particularly HnRNPa1 and Ksrp plays a pivotal role in regulating differential expression of miR-23a-miR-27a-miR-24-2 cluster members by acting selectively on distinct premiRNAs. Surprisingly, selective inhibition of mature miR-23a-miR-27a-miR-24-2 cluster members by RNPs silencing is observed only for HnRNPa2b1 and HnRNPa3, but they do not recapitulate the effects provided by MEF2C silencing, supporting the notion that combinatorial rather than single MEF2C-driven RNPs modulation is occurring. Furthermore, it is important to highlight in this context, that all mature miR-23a-miR-27a-miR-24-2 cluster members are similarly expressed in both subcellular nuclear and cytoplasmic compartment as well as several RNPs. Notably, siRNA silencing would only be affecting those events occurring in the cytoplasm and therefore, inhibition might only be partial. The causal relationship between such distinct subcellular compartment localization deserves further analyses and might provide novel insights on the precise molecular mechanisms controlling differential expression of the mature microRNAs of genomic clustered microRNAs.
In summary, we provide herein evidence of the complex post-transcriptional regulatory mechanism exerted by MEF2C on the regulation of miR-23a-miR-27a-miR-24-2 cluster members. MEF2C can directly and selectively bind to pre-miR-23a_3p and pre-miR-27a_3p but not to pre-miR-24-2. MEF2C can directly bind to distinct RNPs, such as Ddx7, HhRNPa3 and Ksrp, and indirectly regulate the expression of other RNPs such as Adar1 and HnRNPa2b1. Importantly, such regulation is distinctly exerted by the MEF2C amino- and carboxy-terminals. Silencing of MEF2C-binding RNP Ksrp selectively regulates pre-miR-23a and pre-miR-27a, but not pre-miR-24-2, supporting the notion of a direct implication of this pathway on the differential expression of miR-23a-miR-27a-miR-24-2 cluster members, yet a combinatorial action of distinct RNPs seems to be required to fully achieved the final miR-23a-miR-27a-miR-24-2 cluster members expression of the mature microRNAs.
Materials & Methods
MEF2C pulldown assays
For immunoprecipitation of endogenous MEF2C, HL1 cells were lysed with PEB buffer for 10 min on ice and centrifuged at 10 000 g for 15 min at 4°C. The supernatants were incubated with protein A Sepharose beads (Abcam) coated with 15 μg of antibody that recognized MEF2C (Cell Signalling) or control IgG (Abcam) for 2 h at 4°C, respectively. The corresponding beads were washed with NT2 buffer (50 mM Tris–HCl [Ph 7.5], 150 mM NaCl, 1 mM MgCl2, 0.05% NP-40). Protein complexes were incubated with 20 units of DNase I (15 min at 37°C). In this step, an aliquot from each reaction was isolated for Western blot validation. Subsequently they were further incubated with 0.1% SDS/0.5 mg/ml Proteinase K (30 min at 55°C) to remove DNA and proteins, respectively. The RNA isolated from the IP materials were further assessed by RT-qRT-PCR analysis.
Nuclear/cytoplasmic distribution
Cytoplasmic and nuclear RNA fractions from HL1 cardiomyocytes were isolated with Cytoplasmic & Nuclear RNA Purification Kit (Norgen, Belmont, CA, USA) following the manufacturer’s instructions. After RNA isolation, RT-qRT-PCR analysis for nuclear enriched Xist2 mRNA marker and cytoplasmic Gapdh mRNA marker were performed to validate enrichment on each subcellular fractions. RT-qRT-PCR analysis of distinct microRNAs, Xist2 and RNPs was subsequently performed as detailed above.
Generation of MEF2C 3´delection and 5´delection constructs
pcDNA MEF2C plasmids were used to generation two distinct constructs, with 3´and 5´deletions, respectively [104]. MEF2C 3´deletion (MEF2C 3´del) was constructed by deletion the 3´ fragment ranging from nucleotide 1112 of the MEF2C full length (PstI restriction site) to the 3´end of the mouse MEF2C transcript (NM_001170537.2) (Supplementary Figure S1B). Thus, this construct deleted the last 288 amino acids of the MEF2C protein (NP_001164008), maintaining thus the SRF-type DNA-binding and dimerization domain (1-59 aa), the MADS_MEF2_like domain (2-78 aa) and the HJURP_C domain (110-156 aa).
MEF2C 5´deletion (MEF2C 5´del) was constructed by deletion of the 5´fragment spanning from nucleotide 1 until nucleotide 1522 of the MEF2C full length (ScaI restriction site), i.e., mouse MEF2C transcript (NM_001170537.2) (Supplementary Figure S1B). Thus, this construct deleted the first 314 amino acids of the MEF2C protein (NP_001164008), thus deleting the SRF-type DNA-binding and dimerization domain (1-59 aa), the MADS_MEF2_like domain (2-78 aa) and the HJURP_C domain (110-156 aa).
Plasmid transfections
siRNA transfections
HL1 cardiomyocytes (6 × 105 cells per well) were transfected with siRNA-MEF2C, siRNA-Adar1, siRNA-HnRNPa3, siRNA-Ksrp, siRNA-Ddx5, siRNA-Ddx17, siRNA-HnRNPa1 and siRNA-HnRNPa2b1 (Sigma, Aldrich, Munich, Germany), respectively, as previously described [100,101,102]. siRNA sequences are provided in Supplementary Table S1.
RNA isolation and retrotranscription
Total RNA was isolated using Trizol (Roche) according to manufacturer’s guidelines and DNase treated using RNase-Free DNase (Roche) for 15 min at room temperature. In all cases, at least three distinct pooled samples were used to perform the corresponding RT-qRT-PCR experiments.
qRT-PCR analyses (mRNA)
First strand cDNA was synthesized at 50°C for 1 h using 1 μg of RNA, oligo-dT primers and Superscript III Reverse Transcriptase (Invitrogen) according to manufacturer’s guidelines. Negative controls to assess genomic contamination were performed for each sample, without reverse transcriptase, which resulted in all cases in no detectable amplification product. RT-qRT-PCR was performed in CFX384TM thermocycler (Bio-Rad) and SyBR Green detection system. Reactions were performed in 96-well plates with optical sealing tape (Cultek) in 20 μL total volume containing SYBR Green Mix (Finnzymes) and the corresponding cDNA. Two internal controls, mouse Gusb and Gapdh mRNAs, were used in parallel for each run and represented as previously described [105,106,107]. Amplification conditions were as follows: denaturalization step of 95 °C for 10 min, followed by 40 cycles of 95 °C for 30 s, 60 °C for 30 s, 72 °C for 30 s; with final elongation step of 72 °C for 10 min. All primers were designed to span exon-exon boundaries using online Primer3 software Primer3input (http://bioinfo.ut.ee/primer3-0.4.0/). Primer sequences are provided in Supplementary Table S1. Amplification bands of pri-miRNA and pre-miRNAs are illustrated in Supplementary Figure S1E, demonstrating a single transcript for pri-miRNA miR-23a-miR-27a_miR-24-2 and specific amplifications for each pre-miRNA, i.e., pre-miRNA-23a, pre-miR-27a and pre-miR-24-2, respectively. No amplifications were observed in PCR control reactions containing only water as the template. Each PCR reaction was performed at least three times to obtain representative averages. The Livak method was used to analyze the relative quantification RT-PCR data [108] and normalized in all cases taking as 100% the wild-type (control) value, as previously described [105,106].
qRT-PCR analyses (microRNA)
For microRNA expression analyses, 20 ng of total RNA was used for retro-transcription with Universal cDNA Synthesis Kit II (Exiqon) and the resulting cDNA was diluted 1/80, following manufacture´s guidelines. Real time PCR experiments were performed with 1 μL of cDNA, SsoFast EvaGreen mix and corresponding primer sets as described on Supplementary Table S1. All qRT-PCRs were performed using a CFX384TM thermocycler (Bio-Rad) following the manufacturer’s recommendations. The relative level of expression of each gene was calculated as described by Livak & Schmittgen [108] using Gapdh as internal control for mRNA expression analyses and 5S for microRNA expression analyses, respectively. Each PCR reaction was performed at least three times to obtain representative averages.
Statistical analyses
For statistical analyses of datasets, unpaired Student’s t-tests were used. Significance levels or P values are stated in each corresponding figure legend. P < 0.05 was considered statistically significant.
Supplementary Materials
Figure S1. Panel A. Western blot analsyes demonstrating Mef2c pulldown assay as compared to IgG negative control. Panel B. Schematic representation of the MEF2C constructs, i.e., MEF2C FL, MEF2C 3´del and MEF2C 5´del. Note that within MEF2C 3´del construct, the sequences from a Pst1 site to the 3´ of the MEF2C gene are deleted, preserving the Srf, MADS-MEF2 and HJURP_C domains. On the other hand, within MEF2C 5´del construct, the sequences from the 5´ of the MEF2C gene to the ScaI site are deleted, eliminating therefore the Srf, MADS-MEF2 and HJURP_C domains. Panel C. qRT-PCR analyses of Mef2c expression in HL1 atrial cardiomyocytes after transfection with the distinct MEF2C constructs and MEF2C siRNA. Observe that similar expression levels are obtained after transfection with MEF2C FL, MEF2C 5´del y MEF2C 3´del while MEF2C siRNA significantly diminished its expression. Panel D. Transactivation analyses of the L fragment of the miR-23a-miR-27a-miR-24-2 locus with MEF2C FL, MEF2C 5´del and MEF2C 3´ constructs. Observe that MEF2C FL and MEF2C 3´ constructs can transactive this fragment while MEF2C 5´del cannot transactive it as reported by luciferase assays. Panel E. RT-PCR analyses of pri-miRNA and pre-miRNAs amplification products as revealed in gel electrophoresis. pre-miR-23a, pre-miR-27a and pre-miR-24-2 amplification resulted in products below 100 bp, as expected, while pri-miRNA amplification resulted in an approximately 350 bp band, as expected. Figure S2. Schematic representation of the miR-23a-miR-27a-mir-24-2 locus with its genomic sequence, upon which the pre-miRNA precursor sequences is highlighted (pre-miR-23a in yellow, pre-miR-27a in green and pre-miR-24-2 in pink). Primer sequences for pri-miRNA amplification are highlighted in blue, spanning from pre-miR-23a to pre-miR-24-2 sequence. Primers for pre-miRNA amplification are underlined. Table S1. List of primer and siRNA sequences.
Acknowledgments
We would like to thank Jorge Perez Fernandez and Cristina Sanchez Fernandez for critical reading of the manuscript. This work is supported by grants of the Spanish Government to D.F and A.A. (PID2019-107492GB-100) and by grant-in-aid (CTS-446) of the Junta de Andalucia Regional Council.
References
- Wilkinson, A.C.; Göttgens, B. Transcriptional regulation of haematopoietic stem cells. Adv Exp Med Biol. 2013, 786, 187–212. [Google Scholar] [CrossRef] [PubMed]
- Bolte, C.; Whitsett, J.A.; Kalin, T.V.; Kalinichenko, V.V. Transcription Factors Regulating Embryonic Development of Pulmonary Vasculature. Adv Anat Embryol Cell Biol. 2018, 228, 1–20. [Google Scholar] [CrossRef]
- Dias, S.; Xu, W.; McGregor, S.; Kee, B. Transcriptional regulation of lymphocyte development. Curr. Opin. Genet. Dev. 2008, 18, 441–448. [Google Scholar] [CrossRef]
- Paige, S.L.; Plonowska, K.; Xu, A.; Wu, S.M. Molecular Regulation of Cardiomyocyte Differentiation. Circ. Res. 2015, 116, 341–353. [Google Scholar] [CrossRef] [PubMed]
- Nagel, S.; Venturini, L.; Meyer, C.; Kaufmann, M.; Scherr, M.; Drexler, H.G.; Macleod, R.A.F. Transcriptional deregulation of oncogenic myocyte enhancer factor 2C in T-cell acute lymphoblastic leukemia. Leuk. Lymphoma 2011, 52, 290–297. [Google Scholar] [CrossRef] [PubMed]
- Ma, K.; Chan, J.K.L.; Zhu, G.; Wu, Z. Myocyte Enhancer Factor 2 Acetylation by p300 Enhances Its DNA Binding Activity, Transcriptional Activity, and Myogenic Differentiation. Mol. Cell. Biol. 2005, 25, 3575–3582. [Google Scholar] [CrossRef] [PubMed]
- Sartorelli, V.; Huang, J.; Hamamori, Y.; Kedes, L. Molecular Mechanisms of Myogenic Coactivation by p300: Direct Interaction with the Activation Domain of MyoD and with the MADS Box of MEF2C. Mol. Cell. Biol. 1997, 17, 1010–1026. [Google Scholar] [CrossRef]
- Han, J.; Jiang, Y.; Li, Z.; Kravchenko, V.V.; Ulevitch, R.J. Activation of the transcription factor MEF2C by the MAP kinase p38 in inflammation. Nature 1997, 386, 296–299. [Google Scholar] [CrossRef]
- Liu, D.; Kang, J.S.; Derynck, R. TGF-β-activated Smad3 represses MEF2-dependent transcription in myogenic differentiation. EMBO J. 2004, 23, 1557–1566. [Google Scholar] [CrossRef]
- Aude-Garcia, C.; Collin-Faure, V.; Bausinger, H.; Hanau, D.; Rabilloud, T.; Lemercier, C. Dual roles for MEF2A and MEF2D during human macrophage terminal differentiation and c-Jun expression. Biochem. J. 2010, 430, 237–244. [Google Scholar] [CrossRef]
- Lyons, M.R.; Schwarz, C.M.; West, A.E. Members of the Myocyte Enhancer Factor 2 Transcription Factor Family Differentially RegulateBdnfTranscription in Response to Neuronal Depolarization. J. Neurosci. 2012, 32, 12780–12785. [Google Scholar] [CrossRef]
- Lin, Q.; Schwarz, J.; Bucana, C.; Olson, E.N. Control of Mouse Cardiac Morphogenesis and Myogenesis by Transcription Factor MEF2C. Science 1997, 276, 1404–1407. [Google Scholar] [CrossRef] [PubMed]
- Bi, W.; Drake, C.J.; Schwarz, J.J. The Transcription Factor MEF2C-Null Mouse Exhibits Complex Vascular Malformations and Reduced Cardiac Expression of Angiopoietin 1 and VEGF. Dev. Biol. 1999, 211, 255–267. [Google Scholar] [CrossRef]
- Kolodziejczyk, S.M.; Wang, L.; Balazsi, K.; DeRepentigny, Y.; Kothary, R.; Megeney, L.A. MEF2 is upregulated during cardiac hypertrophy and is required for normal post-natal growth of the myocardium. Curr. Biol. 1999, 9, 1203–1206. [Google Scholar] [CrossRef] [PubMed]
- Song, K.; Nam, Y.-J.; Luo, X.; Qi, X.; Tan, W.; Huang, G.N.; Acharya, A.; Smith, C.L.; Tallquist, M.D.; Neilson, E.G.; et al. Heart repair by reprogramming non-myocytes with cardiac transcription factors. Nature 2012, 485, 599–604. [Google Scholar] [CrossRef] [PubMed]
- Qian, L.; Huang, Y.; Spencer, C.I.; Foley, A.; Vedantham, V.; Liu, L.; Conway, S.J.; Fu, J.-D.; Srivastava, D. In vivo reprogramming of murine cardiac fibroblasts into induced cardiomyocytes. Nature 2012, 485, 593–598. [Google Scholar] [CrossRef]
- Voronova, A.; Al Madhoun, A.; Fischer, A.; Shelton, M.; Karamboulas, C.; Skerjanc, I.S. Gli2 and MEF2C activate each other's expression and function synergistically during cardiomyogenesis in vitro. Nucleic Acids Res. 2011, 40, 3329–3347. [Google Scholar] [CrossRef] [PubMed]
- Lockhart, M.M.; Wirrig, E.E.; Phelps, A.L.; Ghatnekar, A.V.; Barth, J.L.; Norris, R.A.; Wessels, A. Mef2c Regulates Transcription of the Extracellular Matrix Protein Cartilage Link Protein 1 in the Developing Murine Heart. PLOS ONE 2013, 8, e57073. [Google Scholar] [CrossRef]
- Vanpoucke, G.; Goossens, S.; De Craene, B.; Gilbert, B.; Van Roy, F.; Berx, G. GATA-4 and MEF2C transcription factors control the tissue-specific expression of the T-catenin gene CTNNA3. Nucleic Acids Res. 2004, 32, 4155–4165. [Google Scholar] [CrossRef]
- Di Lisi, R.; Millino, C.; Calabria, E.; Altruda, F.; Schiaffino, S.; Ausoni, S. Combinatorial cis-Acting Elements Control Tissue-specific Activation of the Cardiac Troponin I Gene in Vitro and in Vivo. J. Biol. Chem. 1998, 273, 25371–25380. [Google Scholar] [CrossRef]
- Zang, M.; Li, Y.; Xue, L.; Jia, H.; Jing, H. Cooperative activation of atrial naturetic peptide promoter by dHAND and MEF2C. J. Cell. Biochem. 2004, 93, 1255–1266. [Google Scholar] [CrossRef]
- Kuisk, I.R.; Li, H.; Tran, D.; Capetanaki, Y. A Single MEF2 Site Governs Desmin Transcription in Both Heart and Skeletal Muscle during Mouse Embryogenesis. Dev. Biol. 1996, 174, 1–13. [Google Scholar] [CrossRef]
- Chen, S.L.; Wang, S.-C.M.; Hosking, B.; Muscat, G.E.O. Subcellular Localization of the Steroid Receptor Coactivators (SRCs) and MEF2 in Muscle and Rhabdomyosarcoma Cells. Mol. Endocrinol. 2001, 15, 783–796. [Google Scholar] [CrossRef] [PubMed]
- Sartorelli, V.; Huang, J.; Hamamori, Y.; Kedes, L. Molecular Mechanisms of Myogenic Coactivation by p300: Direct Interaction with the Activation Domain of MyoD and with the MADS Box of MEF2C. Mol. Cell. Biol. 1997, 17, 1010–1026. [Google Scholar] [CrossRef]
- Pagiatakis, C.; Gordon, J.W.; Ehyai, S.; McDermott, J.C. A Novel RhoA/ROCK-CPI-17-MEF2C Signaling Pathway Regulates Vascular Smooth Muscle Cell Gene Expression. J. Biol. Chem. 2012, 287, 8361–8370. [Google Scholar] [CrossRef] [PubMed]
- Grégoire, S.; Yang, X.-J. Association with Class IIa Histone Deacetylases Upregulates the Sumoylation of MEF2 Transcription Factors. Mol. Cell. Biol. 2005, 25, 2273–2287. [Google Scholar] [CrossRef] [PubMed]
- Molkentin, J.D.; Black, B.L.; Martin, J.F.; Olson, E.N. Mutational Analysis of the DNA Binding, Dimerization, and Transcriptional Activation Domains of MEF2C. Mol. Cell. Biol. 1996, 16, 2627–2636. [Google Scholar] [CrossRef] [PubMed]
- Janson, C.; Chen, Y.; Li, Y.; Leifer, D. Functional regulatory regions of human transcription factor MEF2C. Mol. Brain Res. 2001, 97, 70–82. [Google Scholar] [CrossRef]
- Dong, C.; Yang, X.-Z.; Zhang, C.-Y.; Liu, Y.-Y.; Zhou, R.-B.; Cheng, Q.-D.; Yan, E.-K.; Yin, D.-C. Myocyte enhancer factor 2C and its directly-interacting proteins: A review. Prog. Biophys. Mol. Biol. 2017, 126, 22–30. [Google Scholar] [CrossRef]
- Infantino, V.; Convertini, P.; Menga, A.; Iacobazzi, V. MEF2C exon α: Role in gene activation and differentiation. Gene 2013, 531, 355–362. [Google Scholar] [CrossRef]
- Hombach S, Kretz M. Non-coding RNAs: Classification, Biology and Functioning. Adv Exp Med Biol. 2016, 937, 3–17. [CrossRef] [PubMed]
- Ha, M.; Kim, V.N. Regulation of microRNA biogenesis. Nat. Rev. Mol. Cell Biol. 2014, 15, 509–524. [Google Scholar] [CrossRef]
- Esteller, M. Non-coding RNAs in human disease. Nat. Rev. Genet. 2011, 12, 861–874. [Google Scholar] [CrossRef]
- Catalanotto, C.; Cogoni, C.; Zardo, G. MicroRNA in Control of Gene Expression: An Overview of Nuclear Functions. Int. J. Mol. Sci. 2016, 17, 1712. [Google Scholar] [CrossRef] [PubMed]
- Wojciechowska, A.; Braniewska, A.; Kozar-Kamińska, K. MicroRNA in cardiovascular biology and disease. Adv. Clin. Exp. Med. 2017, 26, 868–874. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Wang, D.-Z. microRNAs in cardiovascular development. J. Mol. Cell. Cardiol. 2012, 52, 949–957. [Google Scholar] [CrossRef]
- Beermann, J.; Piccoli, M.-T.; Viereck, J.; Thum, T.; Clézardin, P.; Coleman, R.; Puppo, M.; Ottewell, P.; Bonnelye, E.; Paycha, F.; et al. Non-coding RNAs in Development and Disease: Background, Mechanisms, and Therapeutic Approaches. Physiol. Rev. 2016, 96, 1297–1325. [Google Scholar] [CrossRef] [PubMed]
- Barwari, T.; Joshi, A.; Mayr, M. MicroRNAs in Cardiovascular Disease. J. Am. Coll. Cardiol. 2016, 68, 2577–2584. [Google Scholar] [CrossRef]
- Wong, L.L.; Wang, J.; Liew, O.W.; Richards, A.M.; Chen, Y.-T. MicroRNA and Heart Failure. Int. J. Mol. Sci. 2016, 17, 502. [Google Scholar] [CrossRef]
- Lin, S.; Gregory, R.I. MicroRNA biogenesis pathways in cancer. Nat. Rev. Cancer 2015, 15, 321–333. [Google Scholar] [CrossRef]
- Piasecka, A.; Sekrecki, M.; Szcześniak, M.W.; Sobczak, K. MEF2C shapes the microtranscriptome during differentiation of skeletal muscles. Sci. Rep. 2021, 11, 1–14. [Google Scholar] [CrossRef]
- Chen, H.-P.; Wen, J.; Tan, S.-R.; Kang, L.-M.; Zhu, G.-C. MiR-199a-3p inhibition facilitates cardiomyocyte differentiation of embryonic stem cell through promotion of MEF2C. J. Cell. Physiol. 2019, 234, 23315–23325. [Google Scholar] [CrossRef]
- Melnik, S.; Gabler, J.; Dreher, S.I.; Hecht, N.; Hofmann, N.; Großner, T.; Richter, W. MiR-218 affects hypertrophic differentiation of human mesenchymal stromal cells during chondrogenesis via targeting RUNX2, MEF2C, and COL10A1. Stem Cell Res. Ther. 2020, 11, 1–18. [Google Scholar] [CrossRef]
- Tan, Y.; Shen, L.; Gan, M.; Fan, Y.; Cheng, X.; Zheng, T.; Niu, L.; Chen, L.; Jiang, D.; Li, X.; et al. Downregulated miR-204 Promotes Skeletal Muscle Regeneration. BioMed Res. Int. 2020, 2020, 1–9. [Google Scholar] [CrossRef]
- Chinchilla, A.; Lozano, E.; Daimi, H.; Esteban, F.J.; Crist, C.; Aranega, A.E.; Franco, D. MicroRNA profiling during mouse ventricular maturation: a role for miR-27 modulating Mef2c expression. Cardiovasc. Res. 2010, 89, 98–108. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Sui, L.; Hong, X.; Yang, M.; Li, W. MiR-448 promotes vascular smooth muscle cell proliferation and migration in through directly targeting MEF2C. Environ. Sci. Pollut. Res. 2017, 24, 22294–22300. [Google Scholar] [CrossRef]
- Shen, L.; Chen, L.; Zhang, S.; Zhang, Y.; Wang, J.; Zhu, L. MicroRNA-23a reduces slow myosin heavy chain isoforms composition through myocyte enhancer factor 2C (MEF2C) and potentially influences meat quality. Meat Sci. 2016, 116, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Yelamanchili, S.V.; Chaudhuri, A.D.; Chen, L.-N.; Xiong, H.; Fox, H.S. MicroRNA-21 dysregulates the expression of MEF2C in neurons in monkey and human SIV/HIV neurological disease. Cell Death Dis. 2010, 1, e77–e77. [Google Scholar] [CrossRef]
- DeVeale, B.; Swindlehurst-Chan, J.; Blelloch, R. The roles of microRNAs in mouse development. Nat. Rev. Genet. 2021, 22, 307–323. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Ransom, J.F.; Li, A.; Vedantham, V.; von Drehle, M.; Muth, A.N.; Tsuchihashi, T.; McManus, M.T.; Schwartz, R.J.; Srivastava, D. Dysregulation of Cardiogenesis, Cardiac Conduction, and Cell Cycle in Mice Lacking miRNA-1-2. Cell 2007, 129, 303–317. [Google Scholar] [CrossRef]
- Wei, Y.; Peng, S.; Wu, M.; Sachidanandam, R.; Tu, Z.; Zhang, S.; Falce, C.; A Sobie, E.; Lebeche, D.; Zhao, Y. Multifaceted roles of miR-1s in repressing the fetal gene program in the heart. Cell Res. 2014, 24, 278–292. [Google Scholar] [CrossRef]
- Wang, S.; Aurora, A.B.; Johnson, B.A.; Qi, X.; McAnally, J.; Hill, J.A.; Richardson, J.A.; Bassel-Duby, R.; Olson, E.N. The Endothelial-Specific MicroRNA miR-126 Governs Vascular Integrity and Angiogenesis. Dev. Cell 2008, 15, 261–271. [Google Scholar] [CrossRef] [PubMed]
- Fish, J.E.; Santoro, M.M.; Morton, S.U.; Yu, S.; Yeh, R.-F.; Wythe, J.D.; Ivey, K.N.; Bruneau, B.G.; Stainier, D.Y.R.; Srivastava, D. miR-126 Regulates Angiogenic Signaling and Vascular Integrity. Dev. Cell 2008, 15, 272–284. [Google Scholar] [CrossRef] [PubMed]
- Kuo, G.; Wu, C.-Y.; Yang, H.-Y. MiR-17-92 cluster and immunity. J. Formos. Med Assoc. 2019, 118, 2–6. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Li, Y.; Qi, P.; Ma, Z. Biology of MiR-17-92 Cluster and Its Progress in Lung Cancer. Int. J. Med Sci. 2018, 15, 1443–1448. [Google Scholar] [CrossRef] [PubMed]
- Gu, H.; Liu, Z.; Zhou, L. Roles of miR-17-92 Cluster in Cardiovascular Development and Common Diseases. BioMed Res. Int. 2017, 2017, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.-H.; Ikeda, G.; Tada, Y.; von Bornstädt, D.; Santoso, M.R.; Wahlquist, C.; Rhee, S.; Jeon, Y.-J.; Yu, A.C.; O’brien, C.G.; et al. miR-106a–363 cluster in extracellular vesicles promotes endogenous myocardial repair via Notch3 pathway in ischemic heart injury. Basic Res. Cardiol. 2021, 116, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Tan, W. miR-106b-25/miR-17-92clusters: Polycistrons with oncogenic roles in hepatocellular carcinoma. World J. Gastroenterol. 2014, 20, 5962–72. [Google Scholar] [CrossRef]
- Khuu, C.; Utheim, T.P.; Sehic, A. The Three Paralogous MicroRNA Clusters in Development and Disease, miR-17-92, miR-106a-363, and miR-106b-25. Scientifica 2016, 2016, 1–10. [Google Scholar] [CrossRef]
- Ru, L.; Wang, X.-M.; Niu, J.-Q. The miR-23–27–24 cluster: an emerging target in NAFLD pathogenesis. Acta Pharmacol. Sin. 2021, 43, 1167–1179. [Google Scholar] [CrossRef]
- Rogler, C.E.; Matarlo, J.S.; Kosmyna, B.; Fulop, D.; Rogler, L.E. Knockdown of miR-23, miR-27, and miR-24 Alters Fetal Liver Development and Blocks Fibrosis in Mice. Gene Expr. 2017, 17, 99–114. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.; Wu, C.-J.; Yasuda, T.; Cruz, L.O.; Khan, A.A.; Lin, L.-L.; Nguyen, D.T.; Miller, M.; Lee, H.-M.; Kuo, M.-L.; et al. miR-23∼27∼24 clusters control effector T cell differentiation and function. J. Exp. Med. 2016, 213, 235–249. [Google Scholar] [CrossRef] [PubMed]
- Chiang, D.Y.; Kongchan, N.; Beavers, D.L.; Alsina, K.M.; Voigt, N.; Neilson, J.R.; Jakob, H.; Martin, J.F.; Dobrev, D.; Wehrens, X.H.; et al. Loss of MicroRNA-106b-25 Cluster Promotes Atrial Fibrillation by Enhancing Ryanodine Receptor Type-2 Expression and Calcium Release. Circ. Arrhythmia Electrophysiol. 2014, 7, 1214–1222. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Xue, H.; Jin, Q.-H.; Guo, J.; Chen, Y.-D. Increased expression of ryanodine receptor type-2 during atrial fibrillation by miR-106-25 cluster independent mechanism. Exp. Cell Res. 2018, 375, 113–117. [Google Scholar] [CrossRef] [PubMed]
- Bang, C.; Fiedler, J.; Thum, T. Cardiovascular Importance of the MicroRNA-23/27/24 Family. Microcirculation 2011, 19, 208–214. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Kim, M.; Han, J.; Yeom, K.-H.; Lee, S.; Baek, S.H.; Kim, V.N. MicroRNA genes are transcribed by RNA polymerase II. EMBO J. 2004, 23, 4051–4060. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Torres, F.; Aranega, A.E.; Franco, D. Identification of regulatory elements directing miR-23a–miR-27a–miR-24-2 transcriptional regulation in response to muscle hypertrophic stimuli. Biochim. et Biophys. Acta (BBA) - Gene Regul. Mech. 2014, 1839, 885–897. [Google Scholar] [CrossRef]
- Briata, P.; Chen, C.-Y.; Giovarelli, M.; Pasero, M.; Trabucchi, M.; Ramos, A.; Gherzi, R. KSRP, many functions for a single protein. Front. Biosci. 2011, 16, 1787–1796. [Google Scholar] [CrossRef]
- Gherzi, R.; Chen, C.; Trabucchi, M.; Ramos, A.; Briata, P. The role of KSRP in mRNA decay and microRNA precursor maturation. Wiley Interdiscip. Rev. RNA 2010, 1, 230–239. [Google Scholar] [CrossRef]
- Cho, C.J.; Myung, S.-J.; Chang, S. ADAR1 and MicroRNA; A Hidden Crosstalk in Cancer. Int. J. Mol. Sci. 2017, 18, 799. [Google Scholar] [CrossRef]
- Xing Z, Ma WK, Tran EJ. The DDX5/Dbp2 subfamily of DEAD-box RNA helicases. Wiley Interdiscip Rev RNA 2019, 10, e1519. [CrossRef]
- Okamoto, S.-I.; Li, Z.; Ju, C.; Schölzke, M.N.; Mathews, E.; Cui, J.; Salvesen, G.S.; Bossy-Wetzel, E.; Lipton, S.A. Dominant-interfering forms of MEF2 generated by caspase cleavage contribute to NMDA-induced neuronal apoptosis. Proc. Natl. Acad. Sci. 2002, 99, 3974–3979. [Google Scholar] [CrossRef] [PubMed]
- Bach-Elias, M.; Kokolo, M. P68 RNA Helicase (DDX5) Required for the Formation of Various Specific and Mature miRNA Active RISC Complexes. MicroRNA 2022, 11, 36–44. [Google Scholar] [CrossRef]
- Dardenne, E.; Espinoza, M.P.; Fattet, L.; Germann, S.; Lambert, M.-P.; Neil, H.; Zonta, E.; Mortada, H.; Gratadou, L.; Deygas, M.; et al. RNA Helicases DDX5 and DDX17 Dynamically Orchestrate Transcription, miRNA, and Splicing Programs in Cell Differentiation. Cell Rep. 2014, 7, 1900–1913. [Google Scholar] [CrossRef] [PubMed]
- Lambert, M.-P.; Terrone, S.; Giraud, G.; Benoit-Pilven, C.; Cluet, D.; Combaret, V.; Mortreux, F.; Auboeuf, D.; Bourgeois, C.F. The RNA helicase DDX17 controls the transcriptional activity of REST and the expression of proneural microRNAs in neuronal differentiation. Nucleic Acids Res. 2018, 46, 7686–7700. [Google Scholar] [CrossRef] [PubMed]
- Motiño, O.; Francés, D.E.; Mayoral, R.; Castro-Sánchez, L.; Fernández-Velasco, M.; Boscá, L.; García-Monzón, C.; Brea, R.; Casado, M.; Agra, N.; et al. Regulation of MicroRNA 183 by Cyclooxygenase 2 in Liver Is DEAD-Box Helicase p68 (DDX5) Dependent: Role in Insulin Signaling. Mol. Cell. Biol. 2015, 35, 2554–2567. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Chendrimada, T.P.; Wang, Q.; Higuchi, M.; Seeburg, P.H.; Shiekhattar, R.; Nishikura, K. Modulation of microRNA processing and expression through RNA editing by ADAR deaminases. Nat. Struct. Mol. Biol. 2006, 13, 13–21. [Google Scholar] [CrossRef]
- Janknecht, R. Multi-talented DEAD-box proteins and potential tumor promoters: p68 RNA helicase (DDX5) and its paralog, p72 RNA helicase (DDX17). . 2010, 2, 223–34. [Google Scholar]
- Wong, S.K.; Lazinski, D.W. Replicating hepatitis delta virus RNA is edited in the nucleus by the small form of ADAR1. Proc. Natl. Acad. Sci. 2002, 99, 15118–15123. [Google Scholar] [CrossRef]
- Bahn, J.H.; Ahn, J.; Lin, X.; Zhang, Q.; Lee, J.-H.; Civelek, M.; Xiao, X. Genomic analysis of ADAR1 binding and its involvement in multiple RNA processing pathways. Nat. Commun. 2015, 6, 1–13. [Google Scholar] [CrossRef]
- Sakurai, M.; Shiromoto, Y.; Ota, H.; Song, C.; Kossenkov, A.V.; Wickramasinghe, J.; Showe, L.C.; Skordalakes, E.; Tang, H.-Y.; Speicher, D.W.; et al. ADAR1 controls apoptosis of stressed cells by inhibiting Staufen1-mediated mRNA decay. Nat. Struct. Mol. Biol. 2017, 24, 534–543. [Google Scholar] [CrossRef]
- Chou, C.-F.; Lin, W.-J.; Lin, C.-C.; Luber, C.A.; Godbout, R.; Mann, M.; Chen, C.-Y. DEAD Box Protein DDX1 Regulates Cytoplasmic Localization of KSRP. PLOS ONE 2013, 8, e73752. [Google Scholar] [CrossRef] [PubMed]
- Giovarelli, M.; Bucci, G.; Ramos, A.; Bordo, D.; Wilusz, C.J.; Chen, C.-Y.; Puppo, M.; Briata, P.; Gherzi, R. H19 long noncoding RNA controls the mRNA decay promoting function of KSRP. Proc. Natl. Acad. Sci. 2014, 111, 201415098–E5028. [Google Scholar] [CrossRef] [PubMed]
- Hall, M.P.; Huang, S.; Black, D.L.; Pollock, C.; Daily, K.; Nguyen, V.T.; Wang, C.; Lewandowska, M.A.; Bensaude, O.; Doxsey, M.E.S.J. Differentiation-induced Colocalization of the KH-type Splicing Regulatory Protein with Polypyrimidine Tract Binding Protein and the c-srcPre-mRNA. Mol. Biol. Cell 2004, 15, 774–786. [Google Scholar] [CrossRef] [PubMed]
- Stavast, C.J.; Erkeland, S.J. The Non-Canonical Aspects of MicroRNAs: Many Roads to Gene Regulation. Cells 2019, 8, 1465. [Google Scholar] [CrossRef] [PubMed]
- Santovito, D.; Weber, C. Non-canonical features of microRNAs: paradigms emerging from cardiovascular disease. Nat. Rev. Cardiol. 2022, 19, 620–638. [Google Scholar] [CrossRef] [PubMed]
- Santovito, D.; Egea, V.; Bidzhekov, K.; Natarelli, L.; Mourão, A.; Blanchet, X.; Wichapong, K.; Aslani, M.; Brunßen, C.; Horckmans, M.; et al. Noncanonical inhibition of caspase-3 by a nuclear microRNA confers endothelial protection by autophagy in atherosclerosis. Sci. Transl. Med. 2020, 12, eaaz2294. [Google Scholar] [CrossRef]
- Penrad-Mobayed, M.; Perrin, C.; L’hôte, D.; Contremoulins, V.; Lepesant, J.-A.; Boizet-Bonhoure, B.; Poulat, F.; Baudin, X.; Veitia, R.A. A role for SOX9 in post-transcriptional processes: insights from the amphibian oocyte. Sci. Rep. 2018, 8, 7191. [Google Scholar] [CrossRef]
- Panda, A.C.; Abdelmohsen, K.; Yoon, J.-H.; Martindale, J.L.; Yang, X.; Curtis, J.; Mercken, E.M.; Chenette, D.M.; Zhang, Y.; Schneider, R.J.; et al. RNA-Binding Protein AUF1 Promotes Myogenesis by Regulating MEF2C Expression Levels. Mol. Cell. Biol. 2014, 34, 3106–3119. [Google Scholar] [CrossRef]
- Tomaselli, S.; Bonamassa, B.; Alisi, A.; Nobili, V.; Locatelli, F.; Gallo, A. ADAR Enzyme and miRNA Story: A Nucleotide that Can Make the Difference. Int. J. Mol. Sci. 2013, 14, 22796–22816. [Google Scholar] [CrossRef]
- Chawla, G.; Sokol, N.S. ADAR mediates differential expression of polycistronic microRNAs. Nucleic Acids Res. 2014, 42, 5245–5255. [Google Scholar] [CrossRef]
- Chen, T.; Xiang, J.-F.; Zhu, S.; Chen, S.; Yin, Q.-F.; Zhang, X.-O.; Zhang, J.; Feng, H.; Dong, R.; Li, X.-J.; et al. ADAR1 is required for differentiation and neural induction by regulating microRNA processing in a catalytically independent manner. Cell Res. 2015, 25, 459–476. [Google Scholar] [CrossRef]
- Widmark, A.; Sagredo, E.A.; Karlström, V.; Behm, M.; Biryukova, I.; Friedländer, M.R.; Daniel, C.; Öhman, M. ADAR1- and ADAR2-mediated regulation of maturation and targeting of miR-376b to modulate GABA neurotransmitter catabolism. J. Biol. Chem. 2022, 298, 101682. [Google Scholar] [CrossRef] [PubMed]
- Briata, P.; Chen, C.-Y.; Giovarelli, M.; Pasero, M.; Trabucchi, M.; Ramos, A.; Gherzi, R. KSRP, many functions for a single protein. Front. Biosci. 2011, 16, 1787–1796. [Google Scholar] [CrossRef] [PubMed]
- Gherzi, R.; Chen, C.-Y.; Ramos, A.; Briata, P. KSRP Controls Pleiotropic Cellular Functions. Semin. Cell Dev. Biol. 2014, 34, 2–8. [Google Scholar] [CrossRef]
- Briata, P.; Chen, C.-Y.; Ramos, A.; Gherzi, R. Functional and molecular insights into KSRP function in mRNA decay. Biochim. et Biophys. Acta (BBA) - Gene Regul. Mech. 2013, 1829, 689–694. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Li, M.; Zhao, X.; Wang, H.; Zhu, J.; Wang, C.; Zhou, M.; Dong, H.; Zhou, R. Upregulation of KSRP by miR-27b attenuates schistosomiasis-induced hepatic fibrosis by targeting TGF-β1. FASEB J. 2020, 34, 4120–4133. [Google Scholar] [CrossRef] [PubMed]
- Zhou, R.; Gong, A.-Y.; Eischeid, A.N.; Chen, X.-M. miR-27b Targets KSRP to Coordinate TLR4-Mediated Epithelial Defense against Cryptosporidium parvum Infection. PLOS Pathog. 2012, 8, e1002702. [Google Scholar] [CrossRef] [PubMed]
- Xia, Z.; Lu, Y.; Li, X.; Mao, T.; Chen, X.-M.; Zhou, R. Upregulation of KSRP by miR-27b provides IFN-γ-induced post-transcriptional regulation of CX3CL1 in liver epithelial cells. Sci. Rep. 2015, 5, 17590–17590. [Google Scholar] [CrossRef]
- Dou, R.; Liu, K.; Yang, C.; Zheng, J.; Shi, D.; Lin, X.; Wei, C.; Zhang, C.; Fang, Y.; Huang, S.; et al. EMT-cancer cells-derived exosomal miR-27b-3p promotes circulating tumour cells-mediated metastasis by modulating vascular permeability in colorectal cancer. Clin. Transl. Med. 2021, 11, e595. [Google Scholar] [CrossRef]
- Gusar, V.; Timofeeva, A.; Chagovets, V.; Kan, N.; Vysokikh, M.; Marey, M.; Karapetyan, A.; Baev, O.; Sukhikh, G. Diagnostic Potential of Exosomal HypoxamiRs in the Context of Hypoxia–Sumoylation–HypoxamiRs in Early Onset Preeclampsia at the Preclinical Stage. Life 2022, 12, 101. [Google Scholar] [CrossRef] [PubMed]
- Chien, M.-H.; Lee, W.-J.; Yang, Y.-C.; Li, Y.-L.; Chen, B.-R.; Cheng, T.-Y.; Yang, P.-W.; Wang, M.-Y.; Jan, Y.-H.; Lin, Y.-K.; et al. KSRP suppresses cell invasion and metastasis through miR-23a-mediated EGR3 mRNA degradation in non-small cell lung cancer. Biochim. et Biophys. Acta (BBA) - Gene Regul. Mech. 2017, 1860, 1013–1024. [Google Scholar] [CrossRef] [PubMed]
- Qi, L.; Song, Y.; Chan, T.H.M.; Yang, H.; Lin, C.H.; Tay, D.J.T.; Hong, H.; Tang, S.J.; Tan, K.T.; Huang, X.X.; et al. An RNA editing/dsRNA binding-independent gene regulatory mechanism of ADARs and its clinical implication in cancer. Nucleic Acids Res. 2017, 45, 10436–10451. [Google Scholar] [CrossRef] [PubMed]
- Martin, J.F.; Schwarz, J.J.; Olson, E.N. Myocyte enhancer factor (MEF) 2C: a tissue-restricted member of the MEF-2 family of transcription factors. Proc. Natl. Acad. Sci. 1993, 90, 5282–5286. [Google Scholar] [CrossRef]
- Lozano-Velasco, E.; Hernández-Torres, F.; Daimi, H.; Serra, S.A.; Herraiz, A.; Hove-Madsen, L.; Aránega, A.; Franco, D. Pitx2 impairs calcium handling in a dose-dependent manner by modulating Wnt signalling. Cardiovasc. Res. 2016, 109, 55–66. [Google Scholar] [CrossRef]
- Lozano-Velasco, E.; Wangensteen, R.; Quesada, A.; Garcia-Padilla, C.; Osorio, J.A.; Ruiz-Torres, M.D.; Aranega, A.; Franco, D. Hyperthyroidism, but not hypertension, impairs PITX2 expression leading to Wnt-microRNA-ion channel remodeling. PLOS ONE 2017, 12, e0188473. [Google Scholar] [CrossRef]
- Domínguez, J.N.; Lodde, V.; Munk, R.; Abdelmohsen, K.; Gorospe, M.; Ginel, A.; Aránega, A.E.; Franco, D.; García-Padilla, C.; Jiménez-Sábado, V.; et al. Identification of atrial-enriched lncRNA Walras linked to cardiomyocyte cytoarchitecture and atrial fibrillation. FASEB J. 2021, 36, e22051–e22051. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
Figure 1.
Panel A) qRT-PCR analyses of the nuclear and cytoplasmic distribution of miR-23a_3p, miR-27a_3p, miR-24_3p mature microRNAs in HL1 cardiomyocytes. Note that all three microRNAs are similarly expressed in the nucleus and cytoplasm in contrast to miR-130a that is primarily cytoplasmic and the long non-coding RNA Xist2 that is preferentially nuclear. Panel B) qRT-PCR analyses of Mef2c pulldown assays for pre-miR-23a, pre-miR-27a and pre-miR-24, respectively. Note that increased levels are observed for pre-miR-23a, and pre-miR-27a but not for pre-miR-24. Panel C) qRT-PCR analyses of Mef2c pulldown assays for mature miR-23a_3p, miR-27a_3p and miR-24_3p, respectively. Note that none of the mature microRNAs are increased after Mef2c pull-down assays. Panel D) qRT-PCR analyses of Mef2c pulldown assays for pri-miR-23-miR-27a-miR-24-2. Panel E) Schematic representation of the Mef2c association to the miR-23a-miR-27a-miR-24-2 clustered microRNAs. All data are normalized to Gapdh for mRNA expression analyses and to 5S for microRNA expression analyses. *p<0.05, **p<0.01, *** p<0.001.
Figure 1.
Panel A) qRT-PCR analyses of the nuclear and cytoplasmic distribution of miR-23a_3p, miR-27a_3p, miR-24_3p mature microRNAs in HL1 cardiomyocytes. Note that all three microRNAs are similarly expressed in the nucleus and cytoplasm in contrast to miR-130a that is primarily cytoplasmic and the long non-coding RNA Xist2 that is preferentially nuclear. Panel B) qRT-PCR analyses of Mef2c pulldown assays for pre-miR-23a, pre-miR-27a and pre-miR-24, respectively. Note that increased levels are observed for pre-miR-23a, and pre-miR-27a but not for pre-miR-24. Panel C) qRT-PCR analyses of Mef2c pulldown assays for mature miR-23a_3p, miR-27a_3p and miR-24_3p, respectively. Note that none of the mature microRNAs are increased after Mef2c pull-down assays. Panel D) qRT-PCR analyses of Mef2c pulldown assays for pri-miR-23-miR-27a-miR-24-2. Panel E) Schematic representation of the Mef2c association to the miR-23a-miR-27a-miR-24-2 clustered microRNAs. All data are normalized to Gapdh for mRNA expression analyses and to 5S for microRNA expression analyses. *p<0.05, **p<0.01, *** p<0.001.
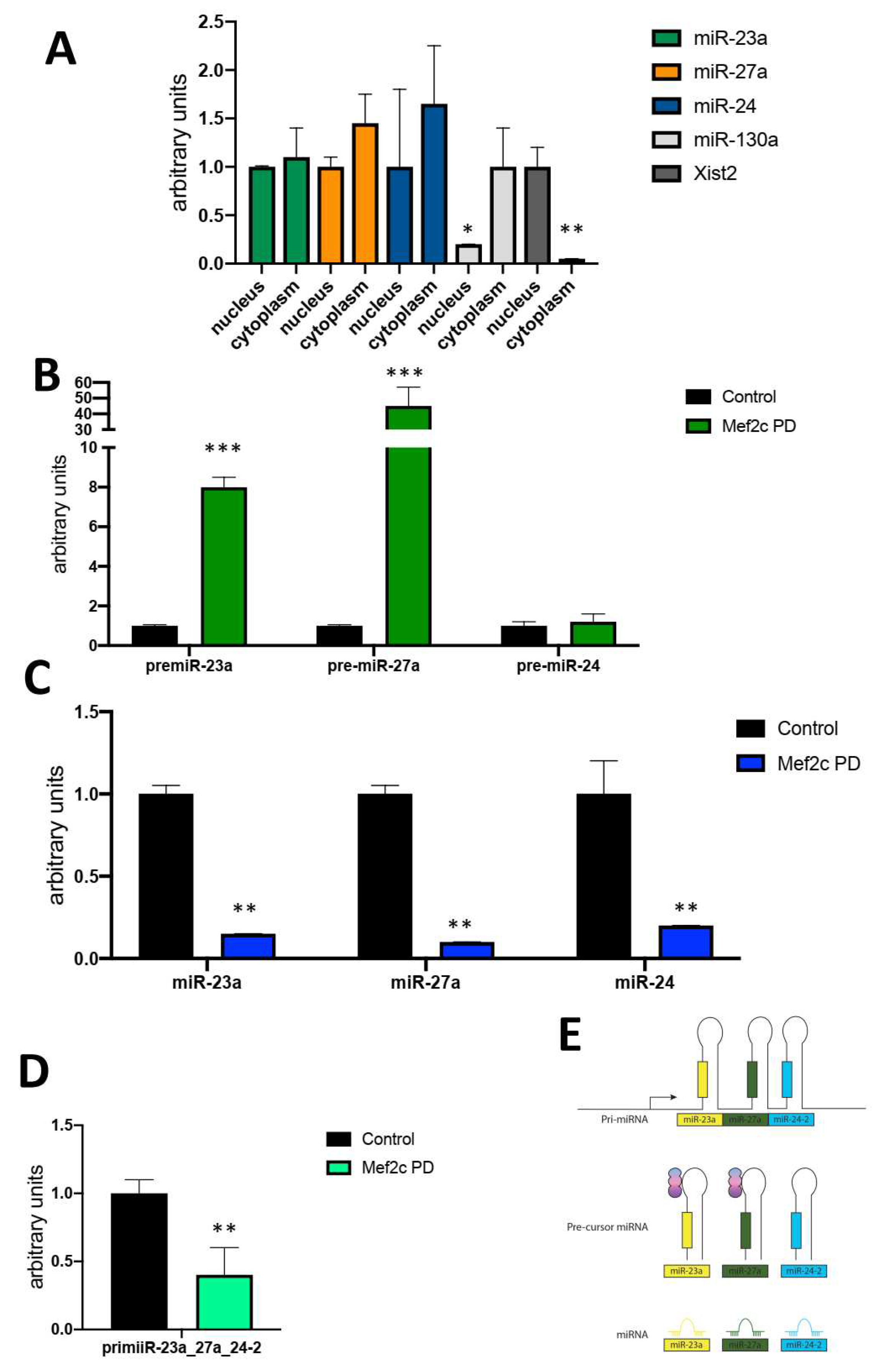
Figure 2.
Panel A) qRT-PCR analyses of pre-miR-23a, pre-miR-27a and pre-miR-24 expression after over-expression of Mef2c full-length (FL), Mef2c 5´del, Mef2c 3´del and Mef2c siRNA in HL1 cardiomyocytes, respectively. Note increased levels are observed for pre-miR-23a, and pre-miR-27a but not for pre-miR-24 after Mef2c 5´del and Mef2c 3´del overexpression, while Mef2c FL and Mef2c siRNA significantly decreased them. Panel B) qRT-PCR analyses of mature miR-23a_3p, miR-27a_3p and miR-24_3p expression after over-expression of Mef2c full-length (FL), Mef2c 5´del, Mef2c 3´del and Mef2c siRNA in HL1 cardiomyocytes, respectively. Note increased levels are observed for all mature microRNAs after Mef2c 5´del and Mef2c 3´del overexpression, while Mef2c FL significantly decreased miR-23a_3p and miR-27a_3p but not miR-24, while Mef2c siRNA only decreased miR-27a_3p. Panel C) Schematic representation of the Mef2c 5´del and Mef2c 3´del regulation to the miR-23a-miR-27a-miR-24-2 clustered microRNAs. All data are normalized to Gapdh for mRNA expression analyses and to 5S for microRNA expression analyses. *p<0.05, **p<0.01, *** p<0.001, **** p<0.0001.
Figure 2.
Panel A) qRT-PCR analyses of pre-miR-23a, pre-miR-27a and pre-miR-24 expression after over-expression of Mef2c full-length (FL), Mef2c 5´del, Mef2c 3´del and Mef2c siRNA in HL1 cardiomyocytes, respectively. Note increased levels are observed for pre-miR-23a, and pre-miR-27a but not for pre-miR-24 after Mef2c 5´del and Mef2c 3´del overexpression, while Mef2c FL and Mef2c siRNA significantly decreased them. Panel B) qRT-PCR analyses of mature miR-23a_3p, miR-27a_3p and miR-24_3p expression after over-expression of Mef2c full-length (FL), Mef2c 5´del, Mef2c 3´del and Mef2c siRNA in HL1 cardiomyocytes, respectively. Note increased levels are observed for all mature microRNAs after Mef2c 5´del and Mef2c 3´del overexpression, while Mef2c FL significantly decreased miR-23a_3p and miR-27a_3p but not miR-24, while Mef2c siRNA only decreased miR-27a_3p. Panel C) Schematic representation of the Mef2c 5´del and Mef2c 3´del regulation to the miR-23a-miR-27a-miR-24-2 clustered microRNAs. All data are normalized to Gapdh for mRNA expression analyses and to 5S for microRNA expression analyses. *p<0.05, **p<0.01, *** p<0.001, **** p<0.0001.
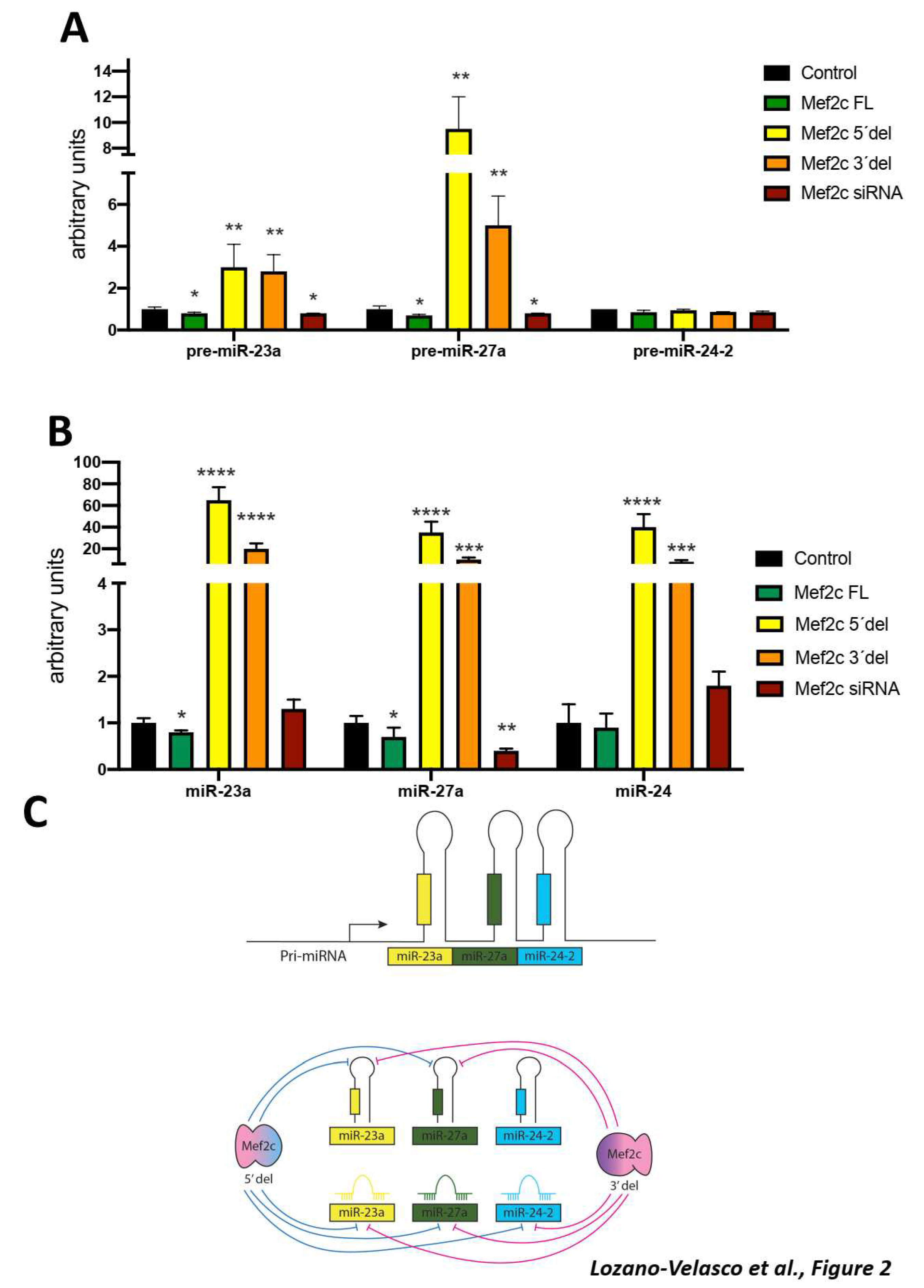
Figure 3.
Panel A) qRT-PCR analyses of distinct RNPs (Adar1, Ddx5, Ddx17, HnRNPa1, HnRNPa2b1, HnRNPa3, Ksrp) in three distinct cells lines, 3T3 fibroblasts, HL1 cardiomyocytes and Sol8 skeletal muscle myoblasts. Observe that these RNPs display distinct expression levels on each of the tested cell line. Panel B) qRT-PCR analyses of the nuclear and cytoplasmic distribution of these RNPs in HL1 cardiomyocytes. Note that Adar1 is preferentially expressed in the nucleus while Ddx5, Ddx17 and Ksrp are preferentially expressed in the cytoplasm. Panel C) qRT-PCR analyses of RNPs expression after over-expression of Mef2c full-length (FL), Mef2c 5´del, Mef2c 3´del and Mef2c siRNA in HL1 cardiomyocytes, respectively. Note that these RNPs are distinctly regulated by each of the Mef2c construct analysed. Panel D) Schematic representation of the Mef2c 5´del and Mef2c 3´del regulation of the RNPs. All data are normalized to Gapdh expression. *p<0.05, **p<0.01, *** p<0.001.
Figure 3.
Panel A) qRT-PCR analyses of distinct RNPs (Adar1, Ddx5, Ddx17, HnRNPa1, HnRNPa2b1, HnRNPa3, Ksrp) in three distinct cells lines, 3T3 fibroblasts, HL1 cardiomyocytes and Sol8 skeletal muscle myoblasts. Observe that these RNPs display distinct expression levels on each of the tested cell line. Panel B) qRT-PCR analyses of the nuclear and cytoplasmic distribution of these RNPs in HL1 cardiomyocytes. Note that Adar1 is preferentially expressed in the nucleus while Ddx5, Ddx17 and Ksrp are preferentially expressed in the cytoplasm. Panel C) qRT-PCR analyses of RNPs expression after over-expression of Mef2c full-length (FL), Mef2c 5´del, Mef2c 3´del and Mef2c siRNA in HL1 cardiomyocytes, respectively. Note that these RNPs are distinctly regulated by each of the Mef2c construct analysed. Panel D) Schematic representation of the Mef2c 5´del and Mef2c 3´del regulation of the RNPs. All data are normalized to Gapdh expression. *p<0.05, **p<0.01, *** p<0.001.
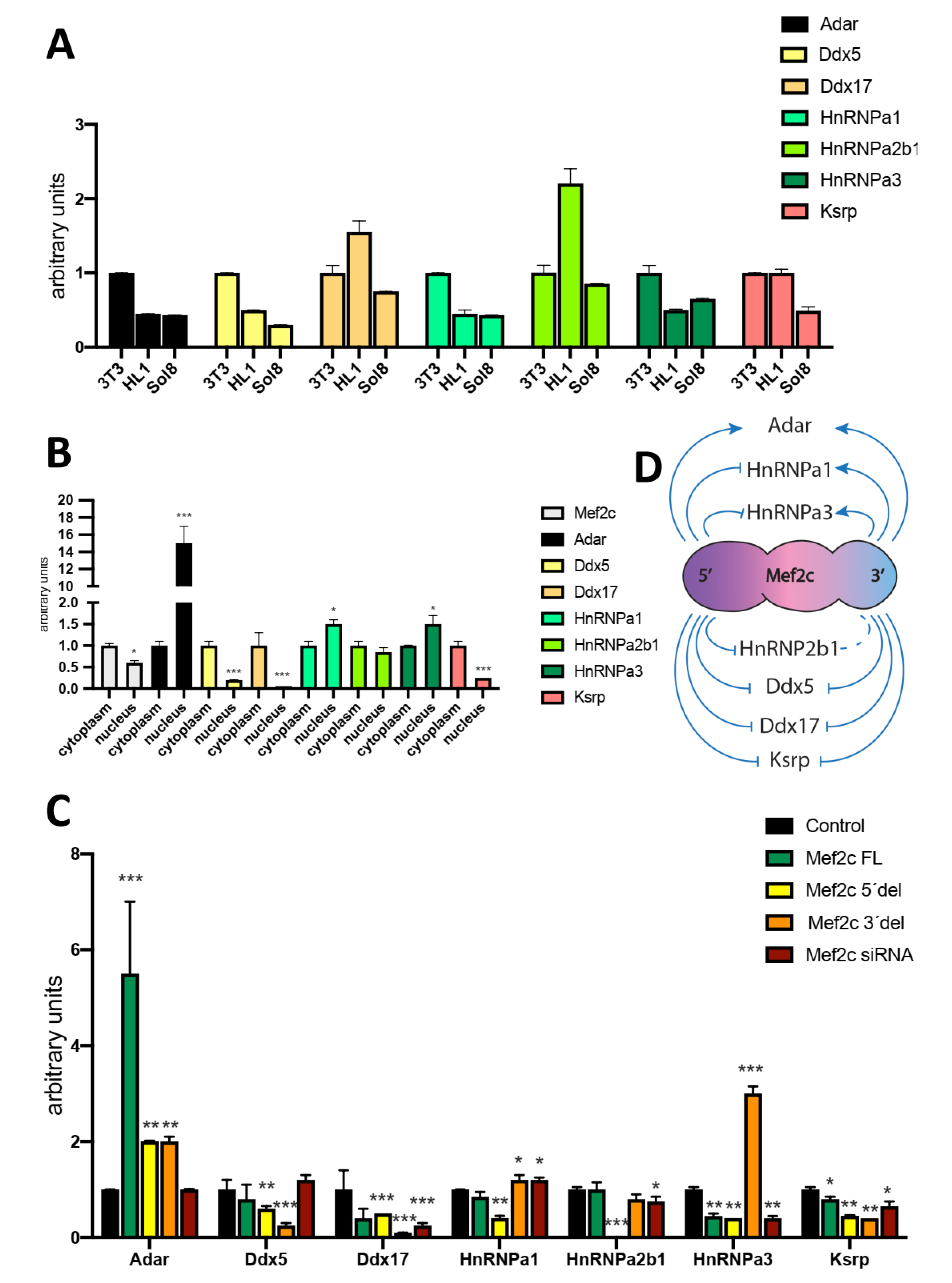
Figure 4.
Panel A) qRT-PCR analyses of Mef2c pulldown assays for RNPs in HL1 cardiomyocytes. Note that increased levels are observed for Ddx17, HnRNPa3 and Ksrp are observed. Panel B) qRT-PCR analyses of pre-miR-23a, pre-miR-27a and pre-miR-24-2 expression after silencing each of the RNP previously tested in HL1 cardiomyocytes, respectively. Note that silencing Ddx5 and HnRNPa3 enhanced expression of all pre-microRNAs while silencing Ksrp only up-regulated pre-miR-23a and pre-miR-27a but not pre-miR-24-2. Panel C) qRT-PCR analyses of mature miR-23a_3p, miR-27a_3p and miR-24_3p expression after silencing each of the RNP previously tested in HL1 cardiomyocytes, respectively. Note that silencing Ddx5 and Ksrp decreased expression of all pre-microRNAs while silencing HnRNPa2b1 selectively up-regulated only pre-miR-23a. Panel D) Schematic representation of the effects of RNP silencing on miR-23a-miR-27a-miR-24-2 pre-miRNAs and mature microRNAs expression, respectively. All data are normalized to Gapdh for mRNA expression analyses and to 5S for microRNA expression analyses. *p<0.05, **p<0.01, *** p<0.001.
Figure 4.
Panel A) qRT-PCR analyses of Mef2c pulldown assays for RNPs in HL1 cardiomyocytes. Note that increased levels are observed for Ddx17, HnRNPa3 and Ksrp are observed. Panel B) qRT-PCR analyses of pre-miR-23a, pre-miR-27a and pre-miR-24-2 expression after silencing each of the RNP previously tested in HL1 cardiomyocytes, respectively. Note that silencing Ddx5 and HnRNPa3 enhanced expression of all pre-microRNAs while silencing Ksrp only up-regulated pre-miR-23a and pre-miR-27a but not pre-miR-24-2. Panel C) qRT-PCR analyses of mature miR-23a_3p, miR-27a_3p and miR-24_3p expression after silencing each of the RNP previously tested in HL1 cardiomyocytes, respectively. Note that silencing Ddx5 and Ksrp decreased expression of all pre-microRNAs while silencing HnRNPa2b1 selectively up-regulated only pre-miR-23a. Panel D) Schematic representation of the effects of RNP silencing on miR-23a-miR-27a-miR-24-2 pre-miRNAs and mature microRNAs expression, respectively. All data are normalized to Gapdh for mRNA expression analyses and to 5S for microRNA expression analyses. *p<0.05, **p<0.01, *** p<0.001.
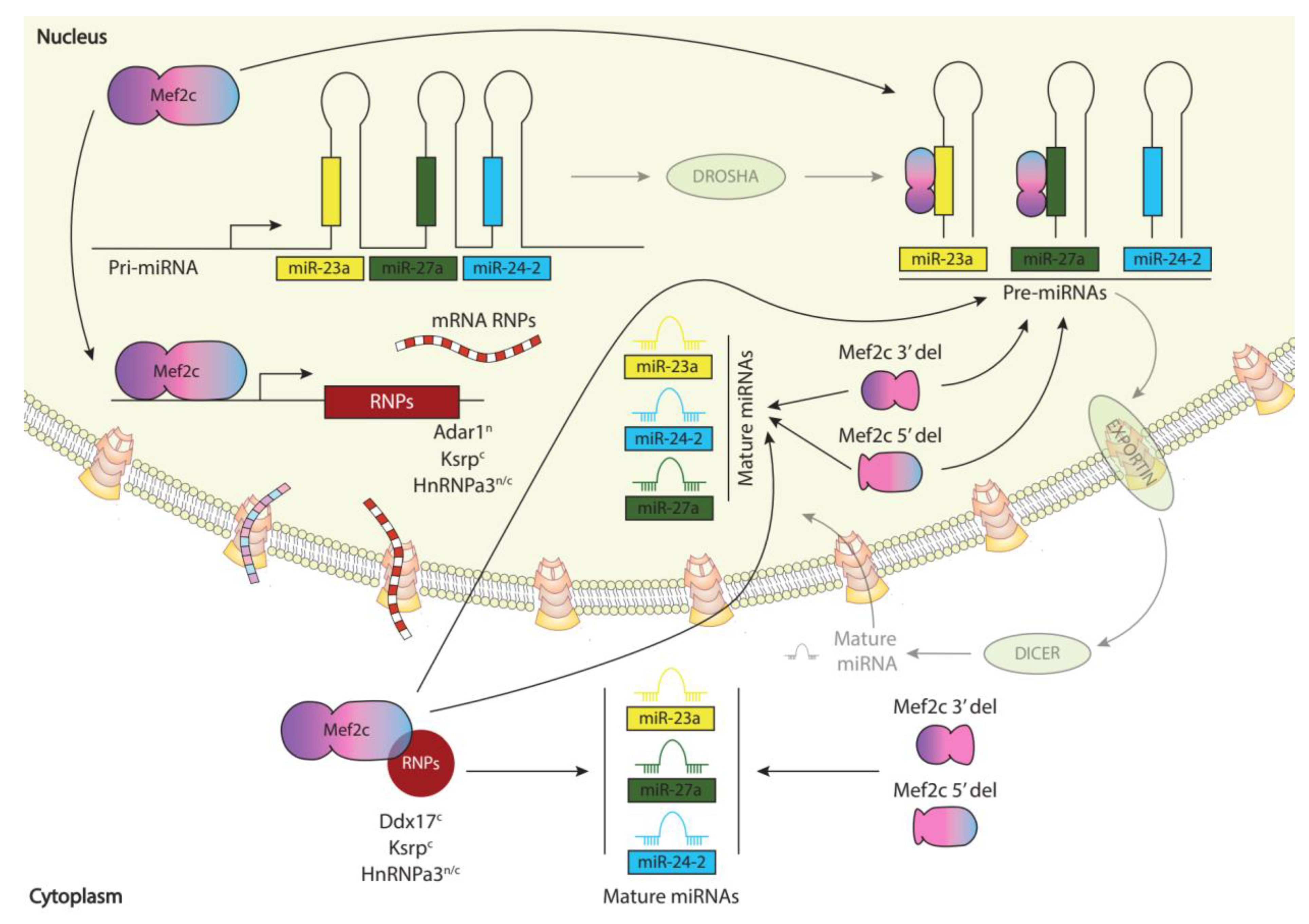
Figure 5.
Schematic representation of Mef2c post-transcriptional regulation of miR-23a-miR-27a-miR-24-2 clustered pre-miRNAs and microRNAs, respectively.
Figure 5.
Schematic representation of Mef2c post-transcriptional regulation of miR-23a-miR-27a-miR-24-2 clustered pre-miRNAs and microRNAs, respectively.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



