
Submitted:
04 October 2023
Posted:
06 October 2023
You are already at the latest version
Abstract
The in vitro production and cryopreservation of mammalian embryos generate reactive oxygen species (ROS) due to conditions of the system that can overcome their antioxidant protection. Resveratrol is an antioxidant used in in vitro systems improving blastocyst rates, but its effect on antioxidant enzymes such as glutathione (GSH) in embryos produced by in vitro fertilization (IVF) after vitrification has not been reported. The objective of this study was to evaluate the effect of resveratrol in the in vitro maturation medium (IVM) of sheep oocytes (Ovis aries) on the levels of ROS and GSH in embryos produced by IVF subjected to vitrification. Resveratrol was added at 0µM, 0.25µM, 0.5µM and 1µM during oocyte in vitro maturation (IVM). Matured oocytes were fertilized with thawed ram sperm. Embryos were cultured in sequential media until blastocysts, then were vitrified for 24 h and after heating they were stained with DCFH-DA (2',7'-dichlorodihydrofluorescein diacetate) to determine the presence of ROS and with Cell Tracker Blue® for the presence of GSH. The quantitative values of ROS and GSH were obtained through the Image J image processor. The results showed that resveratrol increases GSH and decreases ROS production (p<0.05). It is concluded that its use in sheep oocytes during IVM has a beneficial effect on embryos produced by IVF subjected to vitrification, protecting them from oxidative damage by promoting the synthesis of antioxidants.
Keywords:
Resveratrol
; ROS
; GSH
; vitrification
1. Introduction
The in vitro production of embryos is a biotechnology that impacts livestock systems by promoting genetic improvement. The cryopreservation of gametes and embryos has been widely developed worldwide in recent years [1], however, intrinsic processes can cause damage to cells and reduce embryo viability. One of the main factors detected is the increase in reactive oxygen species (ROS) [2], derived from an imbalance with the endogenous embryonic antioxidants.
To solve the damage, antioxidants such as resveratrol have been added to the in vitro culture media. Martínez et al. [3], observed that adding resveratrol at 0.25 and 0.5 μM to sheep oocytes (Ovis aries) during the in vitro maturation (IVM) improved their quality and promoted the compaction of morulae produced by somatic cell nuclear transfer (TNCS). Likewise, it is known that the supplementation of resveratrol (0.25 and 0.5 μM) in the IVM medium of mature cow and goat oocytes reduces intracellular ROS levels, increases glutathione (GSH) concentrations, stimulates embryonic development and gene expression [4]. In sheep, Zabihi et al. [5] did not find differences in the IVM rate but found significant differences in the blastocyst rate at concentrations of 0.25 and 0.5 μM of resveratrol, the quality of these being better at 0.5 μM.
In pigs, it is known that the addition of resveratrol during IVM and/or vitrification modulates the apoptotic process, improving the cryotolerance of vitrified porcine oocytes, while in bovine oocytes it reduces ROS levels, increases blastocyst rates and the number of embryonic cells. Particularly for bovine embryos, supplementation with low concentrations of resveratrol in the medium for in vitro development reduces ROS production and increases beta-oxidation levels, leading to higher survival rates after vitrification [6]. In the sow, Kwak et al. [7] used resveratrol at 0.1, 0.5 and 2.0 μM, during IVM of oocytes to evaluate its effect on ROS and GSH levels, on gene expression in mature oocytes, cumulus cells and blastocysts derived from the IVF. Observing that 2.0 μM of resveratrol improves the developmental potential of embryos, decreases ROS, and increases GSH But authors did not prove its effect on vitrified embryos.
Abdul Rahman et al. [8], say that “glutathione constitutes the major non-protein sulphydryl compound in mammalian cells, which confers protection against oxidative damage”. Working with mice, authors directly added 0.01 mM of exogenous L-glutathione enzyme to the in vitro development culture media, showing that these improved intracellular GSH contents and vitrification outcomes in preimplantation embryos as observed through embryo morphology and preimplantation development. They also observed that this supplementation reduced the ROS levels in these embryos.
García-Martínez et al. [9], proved that glutathione ethyl ester supplemented to the IVM media protects bovine oocytes against oxidative stress induced by subsequent vitrification/warming, by preserving the mitochondrial distribution pattern, diminishing the cytoplasmic and mitochondrial ROS contents, and making embryo development like non-vitrified oocytes.
Other antioxidants have been proved in porcine, where Xiang et al. [10] found that when adding 1.5µM of astaxanthin to the culture media the developmental competence of parthenogenetic zygotes improved (51.7% blastocysts). This concentration also improved the blastocyst formation rate of vitrified cloned zygotes (23.3% blastocysts).
In bovine, Giraldo [11] determined the damage in vitrified embryos due to malondialdehyde (MDA) produced during the oxidation of phospholipids. By measuring MDA concentrations in media subjected to vitrification and post-vitrification with and without embryos, he found that in the first case it produced 35.26% more MDA compared to the second. He also observed that, at a higher concentration of cryoprotectants (DMSO-dimethyl sulfoxide + DMF-dimethylformin 20% each), the concentrations of MDA, ROS and thiobarbituric acid (TBAR) decreased.
Given that resveratrol shows an antioxidant effect that favors the IVM of oocytes, as well as the quality of embryos produced from them, in this study we evaluated its effect on the production of ROS, GSH and nuclei number of embryos produced by IVF after devitrification in sheep (Ovis aries).
2. Materials and Methods
Oocyte collection
Ovaries were collected from domestic sheep (Ovis aries) slaughtered at a local slaughterhouse and transported to the laboratory within 2 h at 30-35 °C in saline solution (0.9% NaCl) with 1% antibiotics (10,000 IU/mL of penicillin, 10 mg/mL of streptomycin sulfate and 25 μg/mL of amphotericin) (Antibac-Antifun 100x, In Vitro, SA). Once in the laboratory, the ovaries were washed and the cumulus-oocyte complexes (COC) were collected by puncturing the ovarian follicles (2 to 5 mm diameter) with a 10 mL syringe and a 20 G needle, containing TCM- 199® with HEPES (25 mM) with heparin (100 IU/mL) [3]. The COC were selected based on the ASEBIR [12] criteria.
In vitro maturation (IVM) of oocytes
IVM was carried out in TCM-199® supplemented with cysteine (0.57 mM), D-glucose (3.05 mM), polyvinyl alcohol (0.1%), sodium pyruvate (0.91 mM), 10% Fetal Bovine Serum (FBS, Microlab), 10 ng/mL of epidermal growth factor (EGF), 0.1 IU/mL-1 of recombinant follicle-stimulating hormone (FSH, Gonal-F Merck), 5 IU mL-1 of equine chorionic gonadotropin (eCG, Gonaforte Parfarm S.A.) and antibiotics (0.6%) (Antibac-Antifun 100x, In Vitro, SA). The IVM medium was added with the different concentrations of resveratrol. The COC were distributed in groups of 25 to 30 in cells of four-cell boxes (Nunc), in 400 μL of IVM medium from each treatment covered with mineral oil, culturing them for 24 h at 38.5°C, 5% CO2 and humidity at saturation. After IVM, 10% of the COC from each treatment were stripped from the cumulus cells (CC) in 0.5 mg/mL-1 of hyaluronidase, to determine the rate of in vitro maturation (IVM- rate of oocytes in Metaphase II or MII) [3] (Martínez et al., 2019) by observing the expansion of the CC and the presence of the first polar body [12], the rest was used for IVF.
In vitro fertilization (IVF) and in vitro development (IVD) of embryos
The COC showing expanded CC were washed in 50 µL of BO-IVF™ in vitro fertilization (IVF) medium (IVF Bioscience) covered with mineral oil and tempered at 38.5°C. Then 30 oocytes were distributed in four-cell culture dishes, in 100 µL of the IVF medium [13]. Straws with semen from fertile rams were thawed at 37°C before use. Post-thawing sperm motility was evaluated. “Swim up” was performed in Eppendorf tubes [2] in the IVM medium recovered from the oocytes that were matured from the control group [14]. Ten µL were taken from the upper part of the tubes with the motile sperm fraction (0.5 x 106 mL-1 sperm) and added to the IVF medium containing the oocytes from each treatment, incubating for 24 h under the same conditions [2]. Fertilized oocytes (zygotes) were washed twice in 50 µL of Cleavage medium (Cook Medical®) for the in vitro development (IVD) and transferred to a new culture dish by placing 25 zygotes per 50 µL of IVD medium covered with mineral oil. They were incubated for four days under the same conditions. On day 4 of culture, the embryos were transferred to another dish with 50 µL of Blastocyst medium (Cook Medical®), the IVD rate (early blastocysts) and embryonic morphology were evaluated in each treatment [2]. The embryos remained in Blastocyst medium (Cook Medical®) for an additional 24 h. The next day (day 5 of culture), early blastocysts were prepared for vitrification.
Vitrification and devitrification of embryos
Following Bhat et al. [15], the early blastocysts of each experimental group were vitrified using the Open Pulled Straw (OPS) technique [16]. Which consisted of washing the blastocysts in 800 µL of Holding medium (TCM-199 without HEPES supplemented with 10% FBS). They were then sequentially equilibrated first in 600 µL of Holding medium supplemented with 7.5% of ethylene glycol and 7.5% of DMSO for 3 min. Then in 600 µL of Holding medium with 0.342 g/mL of sucrose, 16.5% of ethylene glycol and 16.5% of DMSO for 20 sec. Immediately afterwards, 2 µL drops were formed with the embryos, collected by capillarity in an OPS, these were then placed inside straws (0.25 mL) that were heat sealed and subsequently immersed in the liquid nitrogen tank for 24 hours.
For devitrification, the OPS were thawed using the three-step technique [17]. First, each straw was removed from the nitrogen tank, the sealed end was cut, and the OPS was removed from the inside of the straw. The tip of the straw was introduced into 400 µL of Holding medium supplemented with 50% sucrose so that by gravity the embryos could go down, leaving them for 5 min. Embryos were then transferred to 1000 µL of Holding medium supplemented with 25% sucrose for 5 min. Finally, the embryos were washed in 400 µL of Holding medium without sucrose for 5 min. The devitrified early blastocysts were cultured again for 24 h in Blastocyst medium (Cook Medical®) under the same conditions until they reached the late blastocyst and hatched blastocyst stages [18].
Quantification of ROS and GSH in devitrified embryos
A sample of 10% of the devitrified early blastocysts from each group was taken, following the methodology of Martínez et al. [3]. At the end of post-vitrification IVD (day 7 of culture), 10 µM of DCFH-DA (2',7'-dichlorodihydrofluorescein diacetate) and 10 µM of the Cell Tracker Blue® stain were added to the medium containing the embryos, the culture dish was covered with aluminum foil and incubated for 30 minutes under the conditions described. Subsequently, the embryos were washed 3 times in 100 µL of DPBS (Dulbeco Phosphate Buffered Saline) and mounted on slides, covering them with coverslips and sealing them with nail polish. They were evaluated in an epifluorescence microscope with a 460 nm UV filter to evaluate ROS and 405 nm to evaluate GSH. Photographs were taken. The higher the fluorescence intensity, the presence of higher levels of ROS or GSH in the embryos was qualitatively considered. Fluorescence intensity was evaluated in an Image J image processor (version 1.53; Wayne Rasband, National Institute of Health, USA), to obtain semiquantitative data.
DAPI staining of nuclei
Prior to vitrification and after it, following the methodology of Vazquez-Avendaño et al. [19] with some modifications, on day 7 of culture the number of nuclei present in the blastocysts was determined. To do this, embryos of each group were washed for 2 min in 100 µL of DPBS (In vitro, S.A.), were fixed for 24 h in 400 µL of 4% paraformaldehyde, in the refrigerator. Subsequently, they were deposited on a slide in a 2 µL drop, removing the excess of the fixative. Ten 10 µL of a 2.5% solution of 4',6'-diamidino-2-phenylindole (DAPI) in DPBS was added to the slide and covered with a coverslip. A 405 nm UV filter was used to evaluate them. Once the images of each group were obtained, the nuclei of each embryo were counted.
Statistical analysis
The response variables analyzed were rate of in vitro maturation (IVM) of oocytes, embryonic segmentation, in vitro development (IVD), blastocysts with post-vitrification development capacity, presence of ROS, GSH and embryonic quality. These variables were analyzed considering the control group and the experimental groups (with the different concentrations of resveratrol) using Student's T test for paired samples. A one-way ANOVA was used considering the intragroup response variables, followed by the Duncan test, using the SAS Visual Analytical 2020.1 program, at a significance of p<0.05.
3. Results
3.1. Oocyte collection
Eight hundred and fifteen ovaries were collected from domestic sheep (O. aries), with an average aspiration rate of 3 oocytes/ovary. The IVM rate of oocytes treated with different concentrations of resveratrol was analyzed, IVF was verified by the embryonic segmentation rate and the IVD rate.
3.2. In vitro maturation (IVM) of oocytes, in vitro fertilization (IVF) and in vitro development (IVD) of embryos
The IVM rate was similar in oocytes treated with 0.5 and 1 µM of resveratrol and was higher compared to oocytes treated with 0 and 0.25 µM (p≤0.05). The IVD, determined by the embryonic cleavage rate, was also similar for 0.5 and 1 µM of resveratrol and higher with respect to 0 and 0.25 µM (p≤0.05). The blastocyst rate, although it did not show statistical differences between groups (p≥0.05), it was observed that 0.25 µM of resveratrol reduces the rate of late blastocysts (Table 1).
3.3. Vitrification and devitrification of embryos
3.4. ROS and GSH levels in devitrified embryos
The qualitative evaluation of the presence of Reactive Oxygen Species (ROS) showed no differences in late blastocysts between groups, and glutathione (GSH) was present in all groups treated with resveratrol (Figure 2).
The semiquantitative analysis (Image J) determined that ROS levels are reduced with the different treatments (0.25, 0.5 and 1 µM), particularly with 0.5 µM (p≤0.05) of resveratrol; while GSH levels increased at 0.25 and 0.5 µM of resveratrol with respect to the control group, being greater at 0.5 µM (p≤0.05) (Table 3 and Figure 3).
3.5. DAPI staining of nuclei
3.5.1. Presence of nuclei in early blastocysts prior to vitrification
Prior to vitrification, blastocysts showed significant differences in the nuclei present between treatments, particularly for 0.5 and 1 µM of resveratrol, with respect to the control group (p≤0.05). Significant differences were also observed between 0.25 vs. 0.5 µM and between 0.5 vs. 1 µM (p≤0.05), with 1 µM being the treatment in which the embryos showed a greater number of nuclei, but not for 0.5 µM of resveratrol (Figure 4 and Table 4).
3.5.2. Presence of nuclei in late blastocysts after devitrification
At 48 h after devitrification, the quantification of nuclei in the blastocysts showed significant differences between 0.25 and 1 µM of resveratrol (p≤0.05), with the number of nuclei again being greater at 1 µM). However, in all treated groups the number of nuclei present in devitrified embryos was lower than that in non-vitrified embryos (Table 5).
4. Discussion
Effect of resveratrol on IVM, DIV and blastocyst production
It is known that the maturation rate influences the success of IVF and subsequent embryonic development. Other factors that influence this are culture media, changes in pH, osmolarity, temperature, exposure to light, concentration of CO2 and oxygen, humidity, manipulation of cells and cryopreservation methods [20,21]. If the conditions are not adequate, they will generate stress to the oocyte by affecting the cytoplasm and cell membrane, causing changes in nucleic acids, lipids, and proteins [22,23].
This oxidative stress is caused by reactive oxygen species (ROS) produced by cellular metabolism that can damage and/or kill oocytes and embryos, through a decrease in ATP, blockage in embryonic development, alterations in methylation of DNA and histone modification [20,24,25].
Reactive oxygen species are produced naturally during cellular metabolism. However, during embryonic development, especially in the transition from morula to early blastocyst, an increase in ROS occurs and, consequently, at this stage the blastocyst is very susceptible to oxidative stress, especially when endogenous antioxidants (enzymatic and non-enzymatic) do not achieve balance. This is why there is a need to add antioxidants to the culture media [4]. As the embryo progresses in development, cellular metabolism increases and cells are more susceptible to oxidative stress, specifically in the mitochondrial respiratory chain. Complexes I and II produce superoxide and nitrile radicals that affect mitochondrial proteins and the function of different metabolic enzymes in the electron transport chain [26]. It has been reported that mitochondrial DNA (mtDNA) is more sensitive to oxidative stress than nuclear DNA [27], possibly because mtDNA lacks histones, which protect against such damage. Furthermore, it does not have an efficient repair system and is located near the inner mitochondrial membrane, the site of greatest ROS production [28,29]. Additionally, oxidative damage to mtDNA can induce mutations and alter mitochondrial function and integrity [30]. This oxidation-reduction imbalance, in humans, induces degenerative mitochondrial diseases such as Alzheimer's, Parkinson's, and amyotrophic lateral sclerosis [31,32].
In an in vivo system, cells are protected from these effects by the enzymatic and non-enzymatic antioxidants present, for example, oocytes are protected by ovarian follicle fluid [33]. In contrast, in in vitro systems, antioxidants are added [34] such as α-tocopherol, vitamins, proteins, antioxidant enzymes, thiolic compounds, metal chelators, vitamin E and its derivatives, cysteamine, ascorbic acid and extracts of plants with antioxidant properties [34,35]. At physiological concentrations, ROS are essential for cell metabolism and signaling, while their overproduction will generate alterations [36,37].
Resveratrol is an antioxidant used in in vitro oocyte culture media, because promotes IVM rates. At 0.25 and 0.5 µM, resveratrol increases cleavage and blastocyst rates [4,7]. In the present study, oocytes treated with 0.5 and 1 μM resveratrol improved IVM, embryonic cleavage rate, and blastocyst production. It is known that the addition of this antioxidant during IVM favors the cytoplasmic and nuclear maturation of the oocyte. This effect could be related to its ability to increase the expression and nuclear translocation of transcription factors that prevent oxidative stress in the oocyte [3]. It could also be because it increases the concentration of intracellular reduced glutathione (GSH) and favors the concentrations of the enzyme glutathione peroxidase, essential for the embryonic oxidant balance [4]. This allows a decrease in ROS production, as has been observed in pigs, cattle, sheep, and goats.
Effect of resveratrol on embryo vitrification and devitrification
Vitrification subdues the cell to significant thermal and oxidative stress. In the present study, ethylene glycol was used as a cryoprotectant, since its efficiency in the vitrification of ovine and bovine embryos has been reported [16,38]. It has properties such as high penetration speed and low toxicity, in addition to avoiding removing the cryoprotectant prior to embryo transfer; that is, it does not interfere with embryo implantation in embryo recipient females [39].
On the other hand, the Open Pulled Straw (OPS) system allowed embryonic survival after vitrification (64%) at the highest concentration of resveratrol (1 µM), although it was lower than that reported by González [18], who obtained data greater than 70%.
Embryos are exposed to oxidative stress during vitrification, affecting rates of IVM, IVD and early blastocysts when compared to in vivo systems [4,5]. However, in the present study resveratrol supplemented during IVM reduced ROS levels in blastocysts after vitrification. Giaretta et al. [40] and Salzano et al. [41] reported that this antioxidant, when added to the culture medium has a positive effect on development and cryotolerance of blastocysts.
Effect of resveratrol on ROS and GSH levels in devitrified embryos
In the present study the vitrification and devitrification methods to which sheep embryos produced by IVF were subjected, allowed a reduction in ROS levels, which was minimal at 0.5µM, and an increase in GSH levels, reaching the highest levels at this concentration. Greater embryonic survival was achieved after devitrification, at 0.5 µM and 1 µM of resveratrol (p<0.05). This demonstrates that resveratrol, used in the IVM of oocytes, reduces embryonic oxidative stress by promoting the synthesis of endogenous antioxidants such as GSH.
Glutathione is the main non-protein sulfide component in mammalian cells and is recognized for protecting the cell from oxidative damage, regulating the intracellular redox balance [42]. It may also be important in biological processes such as DNA and protein synthesis, and cell proliferation during embryonic development [43]. In cattle, it has been considered an important marker of oocyte viability and quality [44]. Additionally, GSH synthesis during in vitro maturation is associated with the formation of the male pronucleus after fertilization [45,46] and development early embryonic [43].
During embryonic development, GSH is important in DNA and protein synthesis, and cell proliferation [44]. In cattle, it has been considered an important biochemical marker of oocyte viability and quality [43,46].
Effect of resveratrol on embryo quality
Hernández et al. [47], reported a low number of nuclei (44.2±9.9) present in parthenogenetic blastocysts of O. aries and in interspecies cloned blastocysts of O. aries and O. canadensis mexicana (46.7±8.1 nuclei). Lorenzo-Torres et al. [48], reported 122 nuclei in good quality blastocysts produced by IVF. These results are lower than those obtained in the present study, but they are similar to those reported by Agata et al. [49], Mastrorocco et al. [50] and Nadri et al. [51]. Before vitrification, resveratrol at 1µM showed a greater number of nuclei in early blastocysts. After devitrification, although the number of nuclei decreased, the highest value was also at 1µM of resveratrol. In both cases, 1 µM of resveratrol overcome the number of nuclei of control group and the other treatments.
It is shown that resveratrol not only favored the IVM rate of sheep oocytes but had a positive effect on the production of higher quality embryos, conferred cryotolerance capacity during the vitrification process.
5. Conclusions
Resveratrol supplemented at 0.5 µM and 1 µM during IVM of Ovis aries oocytes increased the rate of maturation, improved the development of sheep embryos to the early blastocyst stage from the fourth day of IVD, and favors embryonic cryotolerance to vitrification reducing ROS levels, an effect inversely proportional to GSH levels. But, at 0.25 µM it has a negative effect on IVM, IVD, blastocyst production rate and embryonic survival rate after vitrification. While at 1 µM it favors a greater number of nuclei present in sheep embryos produced by IVF, this indicates that its quality improves before and after vitrification.
6. Patents
Part of the methodology applied in this study was based on the patent No. 394003.
Author Contributions
A.C.G.G., D.A.A.G., J.L.R.S., J.R.V.A. and M.C.N.M. performed the experiments; D.A.A.G., J.P.R.U., C.C.R., J.R.V.A., J.E.H.P. and M.C.N.M. drafted the manuscript; D.A.A.G. analyzed the data. M.C.N.M., J.P.R.U. and D.A.A.G. designed the study; M.C.N.M and D.A.A.G. gave the laboratory equipments, reactives, media and fees facilities for the experiments. All authors have read and agreed to the published version of the manuscript
Funding
This research received no external funding.
Institutional Review Board Statement
Ethical review and approval were waived for this study due institutional codes of ethics were not affected, as sheep ovaries were collected from slaughterhouses and ram semen was provided in frozen straws by the University gene bank.
Data Availability Statement
Data are available from first author Andrea Carolina González Garzón (andrea.gonzalez.garzon@unillanos.edu) upon request.
Acknowledgments
The authors would like to thank the Consejo Nacional de Humanidades, Ciencias y Tecnologías (CONAHCyT) for the scholarship number 1168492 of the main author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Parra-Cortés R. I., Valderrama-Rodas M. & Martínez-Correal G. (2021) Situación actual y perspectivas de la ganadería de bovinos Criollos en América Latina. Archivos Latinoamericanos de Producción Animal, 2(3-4). [CrossRef]
- Rodríguez Suástegui J. L., Romo García S. D., Casas Hernández E., Hernández Pichardo J. E. (2017). Desarrollo de mórulas de ovino en medio simple o secuencial: Relación entre evaluación morfológica y viabilidad embrionaria. Revista de Salud Animal, 39(1), 9-18.
- Martínez Ibarra J.L., Espinoza Mendoza E.A., Rangel Santos R., Ambriz-García D.A., Navarro-Maldonado M.C. (2019). Effect of resveratrol on the in vitro maturation of ovine (Ovis aries) oocytes and the subsequent development of handmade cloned embryos. Veterinaria México, 5(4). [CrossRef]
- Mukherjee, A., Malik, H., Saha, A. P., Dubey, A., Singhal, D. K., Boateng, S., Saugandhika, S., Kumar, S., De, S., Guha, S. K., & Malakar, D. (2014). Resveratrol treatment during goat oocytes maturation enhances developmental competence of parthenogenetic and hand-made cloned blastocysts by modulating intracellular glutathione level and embryonic gene expression. Journal of Assisted Reproduction and Genetics, 31(2), 229–239. [CrossRef]
- Zabihi, A., Shabankareh, H. K., Hajarian, H., & Foroutanifar, S. (2021). In vitro maturation medium supplementation with resveratrol improves cumulus cell expansion and developmental competence of Sanjabi sheep oocytes. Livestock Science, 243. [CrossRef]
- Madrid Gaviria, S., López Herrera, A., Urrego, R., Restrepo Betancur, G., & Echeverri Zuluaga, J. J. (2019). Effect of resveratrol on vitrified in vitro produced bovine embryos: Recovering the initial quality. Cryobiology, 89, 42–50. [CrossRef]
- Kwak, S. S., Cheong, S. A., Jeon, Y., Lee, E., Choi, K. C., Jeung, E. B., & Hyun, S. H. (2012). The effects of resveratrol on porcine oocyte in vitro maturation and subsequent embryonic development after parthenogenetic activation and in vitro fertilization. Theriogenology, 78(1), 86–101. [CrossRef]
- Abdul Rahman, N.-S., Mohamed Noor Khan, N.-A., Eshak, Z., Sarbandi, M.-S., Mohammad Kamal, A.-A., Abd Malek, M., Abdullah, F., Abdullah, M.A. & Othman, F. (2022). Exogenous L-Glutathione improves vitrification outcomes in murine preimplantation embryos. Antioxidants, 11, 2100: 1-21. https:// doi.org/10.3390/antiox11112100.
- García-Martínez, T., Vendrell-Flotats, M., Martínez-Rodero, I., Ordóñez-León, E. A., Álvarez-Rodríguez, M., López-Béjar, M., Yeste, M. & Mogas, T. (2020). Glutathione Ethyl Ester Protects In Vitro-Maturing Bovine Oocytes against Oxidative Stress Induced by Subsequent Vitrification/Warming. International Journal of Molecular Sciences, 21, 7547: 1-22. [CrossRef]
- Xiang, D., Jia, B., Zhang, B., Liang, J., Hong, Q., Wei, H., & Wu, G. (2022). Astaxanthin supplementation improves the subsequent developmental competence of vitrified porcine zygotes. Frontiers in Veterinary Science, 9:871289. [CrossRef]
- Giraldo G., J. J. (2011). Efecto del crioprotector dimetilformamida sobre la viabilidad de embriones bovinos producidos in vitro. Tesis Maestría. Universidad Nacional de Colombia, 96 pages. [CrossRef]
- Asociación para el Estudio de la Biología de la Reproducción (ASEBIR). (2015). Criterios ASEBIR de valoración morfológica de oocitos, embriones tempranos y blastocistos humanos. Cuadernos de Embriología Clínica. Madrid, 3er Edición: 9–75.
- Filipiak Y., Larocca C., Martínez M. (2017). Comportamiento del Semen Bovino Sexado Congelado-Descongelado en Fertilización in vitro (FIV) Capacitado Mediante BO en dos Concentraciones versus Percoll. International Journal of Morphology, 35(4), 1337-1341.
- Salgado-Cruz E., y Lopera-Vásquez R. (2020). Aspectos esenciales sobre las técnicas de fertilización in vitro en bovinos”. Revista de Investigaciones Veterinarias del Perú, 31(3), e17138. [CrossRef]
- Bhat, M. H., Sharma, V., Khan, F. A., Naykoo, N. A., Yaqoob, S. H., Vajta, G., Khan, H. M., Fazili, M. R., Ganai, N. A., & Shah, R. A. (2015). Open pulled straw vitrification and slow freezing of sheep IVF embryos using different cryoprotectants. Reproduction, Fertility, and Development, 27(8), 1175–1180. [CrossRef]
- Vajta G. (2000). Vitrification of the oocytes and embryos of domestic animals. Animal Reproduction Science, 60-61, 357–364. [CrossRef]
- Sosa F. & Hansen P. J. (2019) Protocol for embryo vitrification using open pulled straws. Procedures for In Vitro Production of Bovine Embryos - University of Florida.
- González Mendoza, D. F. (2020). Efecto del protocolo de vitrificación y sistemas de empaque sobre la tasa de supervivencia de embriones ovinos obtenidos in vivo. Tesis Maestría. Universidad Nacional de Colombia.
- Vazquez-Avendaño, J. R., Hernández-Martínez, S., Hernández-Pichardo, J. E., Rivera-Rebolledo, J. A., Ambríz-García, D. A., & Navarro-Maldonado, M. del C. (2017). Efecto del uso de medio secuencial humano en la producción de blastocistos de hembra Ovis canadensis mexicana por clonación manual. Acta Zoológica Mexicana (n.s.), 33(2), 328–338. [CrossRef]
- Guerin, P., El Mouatassim, S. & Menezo, Y. (2001). Oxidative stress y protección contra especies reactivas de oxígeno en el embrión preimplantacional y su entorno. Human Reproduction Update, 7: 175–189.
- Méndez, M. S., Argudo, D. E., Soria, M. E., Galarza, L. R., & Perea, F. P. (2020). Efecto de la adición de melatonina en el medio de maduración y/o vitrificación de ovocitos sobre la producción in vitro de embriones bovinos. Revista de Investigaciones Veterinarias del Perú, 31(1), e17557. Epub 31 de marzo de 2020. [CrossRef]
- Miclea I., Pacala N., Hettig A., Zahan M. & Miclea V. (2012). Las com binaciones de alfa tocoferol y ácido ascórbico influyen en la maduración de los ovocitos de oveja. Artículos Científicos Ciencias Animales y Biotecnologías, 45: 310–313.
- Sollecito, N.V., Pereira, E. C. M., Grázia, J. G. V., Neves, B. P., Couto, B. V. R., Andrade, V. B., Miranda, M. S., Silva, J. K. R. & Borges, A. M. (2019). La actividad antioxidante del extracto oleoso obtenido de Lippia origanoides mejora la calidad de los embriones bovinos producidos in vitro. Arquivo Brasileiro de Medicina Veterinária e Zootecnia, 71: 723–731.
- Brykczynska, U., Hisano, M., Erkek, S., Ramos, L., Oakeley, E. J., Roloff, T. C.; et al. (2010). Repressive and active histone methylation mark distinct promoters in human and mouse spermatozoa. Nature Structural and Molecular Biology. 17(6): 679–687.
- De Castro, L. S., De Assis, P. M., Siquiera, A. F. P., Hamilton, T. R. S., Mendes, C. M., Losano, J. D. A.; et al. (2015). Sperm oxidative stress is detrimental to embryo development: A dose dependent study model and a new and more sensitive oxidative status evaluation. Oxidative Medicine and Cellular Longevity. 2016: 1–12. [CrossRef]
- Selivanov, V. A., Votyakova, T. V., Pivtoraiko, V. N., Zeak, J., Sukhomlin, T., Trucco, M.; et al. (2011). Reactive oxygen species production by forward and reverse electron fluxes in the mitochondrial respiratory chain. Beard DA, editor. PLoS Computational Biology, 31, 7(3): e1001115.
- Yakes, F. M. & Van Houten, B. (1997). Mitochondrial DNA damage is more extensive and persists longer than nuclear DNA damage in human cells following oxidative stress. Proceedings of the National Academy of Sciences of the United States of America, 21, 94(2): 514–519.
- Finkel, T, & Holbrook, N. J. (2000). Oxidants, oxidative stress and the biology of ageing. Nature; 9, 408(6809): 239–247.
- Chinnery, P. F., Elliott, H. R., Hudson, G., Samuels, D. C. & Relton, C. L. (2012). Epigenetics, epidemiology and mitochondrial DNA diseases. International Journal of Epidemiology 41(1): 177–187.
- Han, Y. & Chen, J. Z. (2013). Oxidative stress induces mitochondrial DNA damage and cytotoxicity through independent mechanisms in human cancer cells. Biomedical Research International, ID 825065, 8 pages. [CrossRef]
- Guo, C., Sun, L., Chen, X. & Zhang, D. (2013). Oxidative stress, mitochondrial damage and neurodegenerative diseases. Neural Regeneration Research, 8(21): 2003–2014.
- Kirkinezos, I. G. & Moraes, C. T. (2001). Reactive oxygen species and mitochondrial diseases. Seminars in Cell and Developmental Biology, 12(6): 449–457.
- Combelles, C. M., Gupta, S., Agarwal, A. (2009). ¿Podría el estrés oxidativo influir en la maduración in vitro de los ovocitos? Reproductive Biomed Online, 18: 864–880.
- Asiye, I. S., Ozen, B. O., Alper, B. (2018). Maduración in vitro de ovocitos bovinos: Efectos beneficiosos de la cisteamina. Journal of Dairy, Veterinary & Animal Research, 7: 64–65.
- Ileana, M. N. P., Andrea, H., Marius., Z., Vasile, M. (2012). Alfa-tocoferol y las combinaciones de ácido ascórbico influyen en la maduración de los ovocitos de oveja. Animal Science Biotechnology, 45: 310–313.
- Sunderam, S., Kissin, D. M., Crawford, S. B., Folger, S. G., Jamieson, D. J., Warner, L. & Barfield, W. D. (2015). Vigilancia de la tecnología reproductiva asistida. Estados Unidos, 2012. Informe semanal de morbilidad y mortalidad: Resúmenes de vigilancia 64: 1–29.
- Lees, J. G., Gardner, D. K., Harvey, A. J. (2017). Pluripotent stem cell metabolism and mitochondria: Beyond ATP. Stem Cells International, 2017, ID 2874283, 17 pages . [CrossRef]
- Vargas Reyes, J. N. (2013). Efecto de las técnicas de congelación lenta y vitrificación con etilenglicol sobre la calidad poscriopreservación de embriones bovinos producidos in vitro. https://ciencia.lasalle.edu.co/maest_ciencias_veterinarias/10.
- Youngs, C. R. (2011) Cryopreservation of Preimplantation Embryos of Cattle, Sheep, and Goats. Biology. [CrossRef]
- Giaretta, E., Spinaci, M., Bucci, D., Tamanini, C. & Galeati, G. (2013). Effects of resveratrol on vitrified porcine oocytes. Oxidative Medicine and Cellular Longevity, 2013, ID 920257: 7 pages.
- Salzano, A., Albero, G., Zullo, G., Neglia, G., Abdel-Wahab, A., Bifulco, G., Zicarelli, L., & Gasparrini, B. (2014). Effect of resveratrol supplementation during culture on the quality and cryotolerance of bovine in vitro produced embryos. Animal Reproduction Science, 151(3-4), 91–96. [CrossRef]
- Battin, E. E., & Brumaghim, J. L. (2009). Antioxidant activity of sulfur and selenium: A review of reactive oxygen species scavenging, glutathione peroxidase, and metal-binding antioxidant mechanisms. Cell Biochemistry and Biophysics, 55(1), 1–23. [CrossRef]
- De Matos, D. G., Gasparrini, B., Pasqualini, S. R., & Thompson, J. G. (2002). Effect of glutathione synthesis stimulation during in vitro maturation of ovine oocytes on embryo development and intracellular peroxide content. Theriogenology, 57(5), 1443–1451. [CrossRef]
- Zuelke, K. A., Jeffay, S. C., Zucker, R. M. & Perreault, S. D. (2003). Glutathione (GSH) concentrations vary with the cell cycle in maturing hamster oocytes, zygotes, and pre-implantation stage embryos. Molecular Reproduction and Development, 64(1): 106–112.
- De Matos, D. G., Furnus, C. C., Moses, D. F., Martinez, A. G., & Matkovic, M. (1996). Stimulation of glutathione synthesis of in vitro matured bovine oocytes and its effect on embryo development and freezability. Molecular Reproduction and Development, 45(4), 451–457. [CrossRef]
- De Matos, D. G., Furnus, C. C., & Moses, D. F. (1997). Glutathione synthesis during in vitro maturation of bovine oocytes: Role of cumulus cells. Biology of Reproduction, 57(6), 1420–1425. [CrossRef]
- Hernández Martínez S., Hernández Pichardo J.E., Vazquez Avendaño J.R., Ambríz García D.A. & Navarro Maldonado M.C. (2020) Developmental dynamics of cloned Mexican bighorn sheep embryos using morphological quality standards. Veterinary Medicine and Science, 6: 382–392. [CrossRef]
- Lorenzo-Torres, A., Rangel-Santos, R., Ruíz-Flores, A. & Ambríz-García, D. (2022). In vitro embryo production from ewes at different physiological stages. Journal of Veterinary Sciences, 23(6): e87. [CrossRef]
- Agata Anzalone, D., Palazzese, L., Czernik, M., Sabatucci, A., Valbonetti, L., Capra, E., & Loi, P. (2021) Controlled spermatozoa–oocyte interaction improves embryo quality in sheep. Scientifc Reports, 11: 22629. [CrossRef]
- Mastrorocco, A., Cacopardo, L.; Lamanna, D., Temerario, L., Brunetti, G., Carluccio, A., Robbe, D. & Dell’Aquila, M. E. (2021). Bioengineering approaches to improve in vitro performance of prepubertal lamb oocytes. Cells, 10: 1458. [CrossRef]
- Nadri, T., Towhidi, A., Zeinoaldini, S., Riazi, G., Sharafi, M., Zhandi, M., Kastelic, J., & Gholami, D. (2022). Supplementation of freezing medium with encapsulated or free glutathione during cryopreservation of bull sperm. Reproduction in Domestic Animals = Zuchthygiene, 57(5), 515–523. [CrossRef]
Figure 1.
Late blastocysts produced by IVF from oocytes with resveratrol during IVM, after devitrification. A) 0 µM resveratrol (late expanded blastocysts); B) 0.25 µM resveratrol; C) 0.5 µM resveratrol (expanded blastocysts); D) 1 µM resveratrol (hatched blastocysts). A; B; D: Magnification 10X. C. Magnification 20X.
Figure 1.
Late blastocysts produced by IVF from oocytes with resveratrol during IVM, after devitrification. A) 0 µM resveratrol (late expanded blastocysts); B) 0.25 µM resveratrol; C) 0.5 µM resveratrol (expanded blastocysts); D) 1 µM resveratrol (hatched blastocysts). A; B; D: Magnification 10X. C. Magnification 20X.
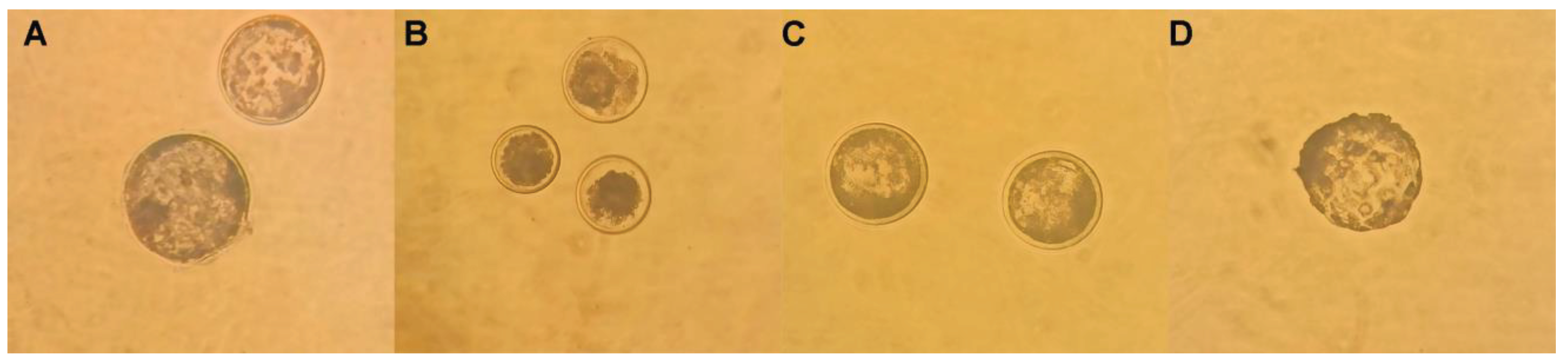
Figure 2.
Qualitative evaluation of the presence of ROS and GSH in late blastocysts produced by IVF from oocytes treated with resveratrol during IVM, after devitrification. A. 0 µM; B: 0.25 µM; C: 0.5 µM and D: 1 µM resveratrol. Green images DCFH staining for ROS. Epifluorescence microscope (460 nm UV filter). Blue images Cell Tracker Blue® staining for GSH. Epifluorescence microscope (405nm UV filter). Magnification 10X.
Figure 2.
Qualitative evaluation of the presence of ROS and GSH in late blastocysts produced by IVF from oocytes treated with resveratrol during IVM, after devitrification. A. 0 µM; B: 0.25 µM; C: 0.5 µM and D: 1 µM resveratrol. Green images DCFH staining for ROS. Epifluorescence microscope (460 nm UV filter). Blue images Cell Tracker Blue® staining for GSH. Epifluorescence microscope (405nm UV filter). Magnification 10X.
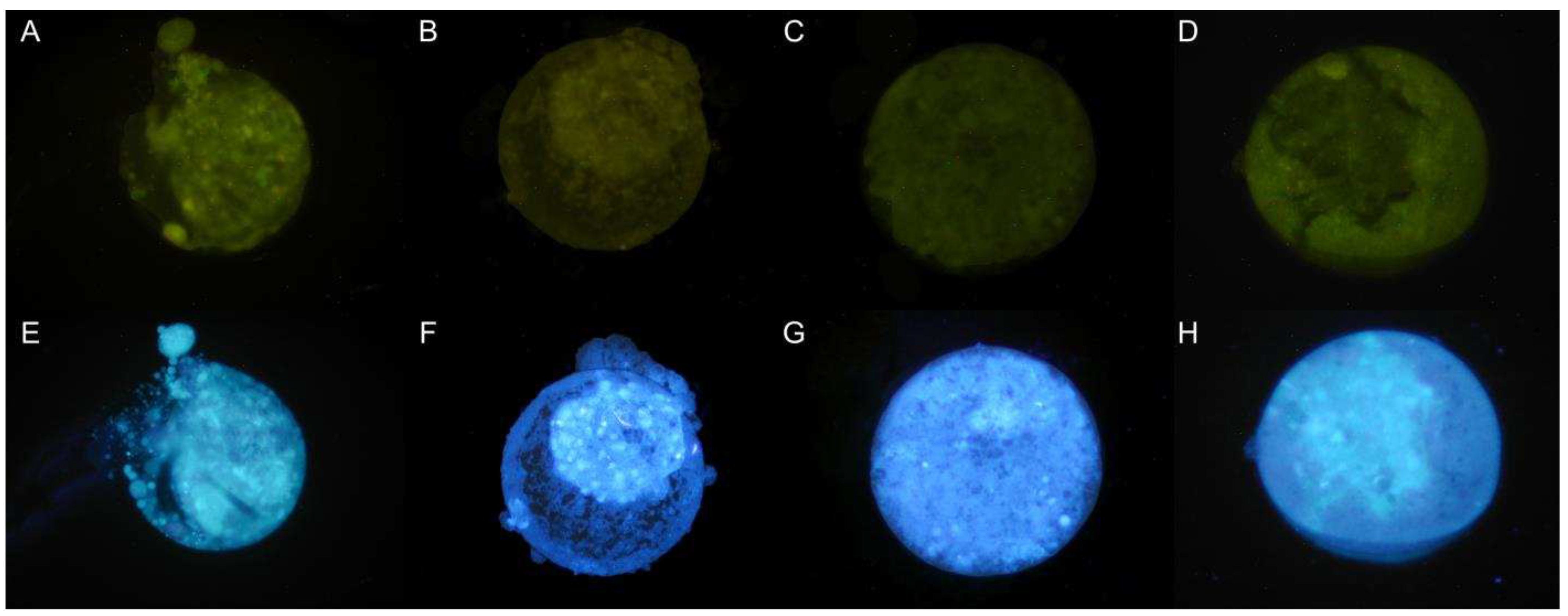
Figure 3.
Fluorescence levels (Image J) of Reactive Oxygen Species (ROS) and glutathione (GSH) in sheep blastocysts produced by IVF from oocytes treated with resveratrol, after devitrification. Literals between bars show significant differences (p≤0.05).
Figure 3.
Fluorescence levels (Image J) of Reactive Oxygen Species (ROS) and glutathione (GSH) in sheep blastocysts produced by IVF from oocytes treated with resveratrol, after devitrification. Literals between bars show significant differences (p≤0.05).
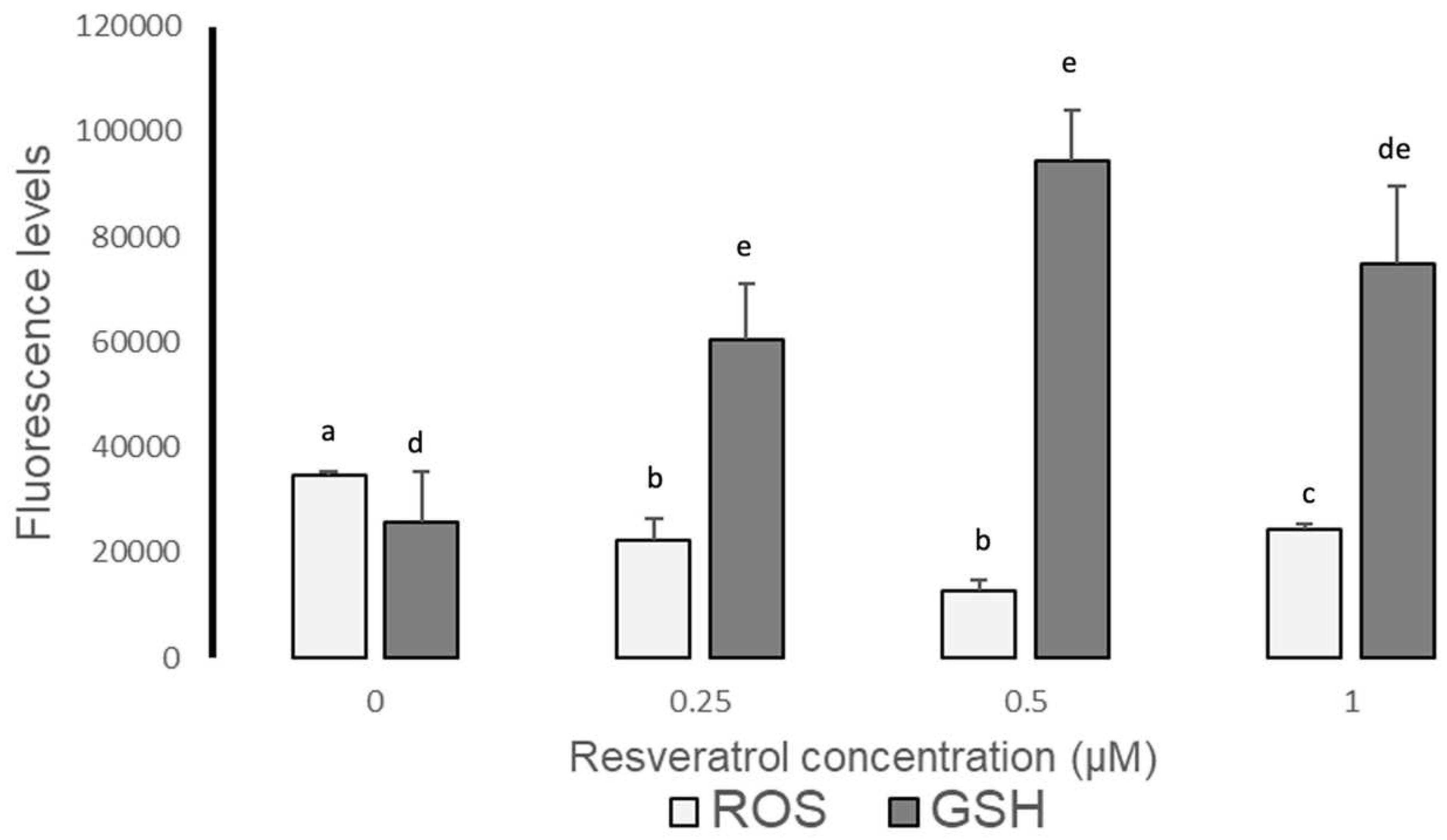
Figure 4.
Presence of nuclei in early blastocysts prior to vitrification, produced by IVF from oocytes treated with resveratrol during IVM. Images A. 0 µM; B. 0.25 µM; C. 0.5 µM and D. 1 µM resveratrol. DAPI stain. Epifluorescence microscope (405 nm UV filter). 20X magnification.
Figure 4.
Presence of nuclei in early blastocysts prior to vitrification, produced by IVF from oocytes treated with resveratrol during IVM. Images A. 0 µM; B. 0.25 µM; C. 0.5 µM and D. 1 µM resveratrol. DAPI stain. Epifluorescence microscope (405 nm UV filter). 20X magnification.
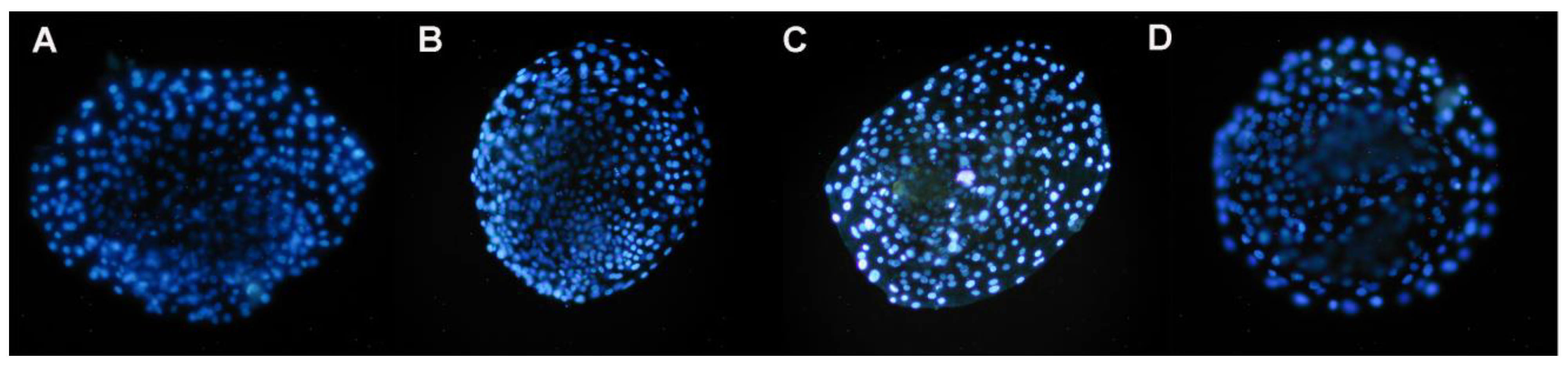

Table 1.
Rate of IVM, IVD and O. aries blastocysts from oocytes treated with resveratrol.
| RESVERATROL | 0 µM | 0.25 µM | 0.5 µM | 1 µM |
|---|---|---|---|---|
| IVM | 75 ± 6.5a | 74 ± 8.5a | 81 ± 6.1b | 81 ± 6.5b |
| CLEAVAGE (IVD) | 63.1 ± 5.4a | 60 ± 4.6a | 68.6 ± 4.7b | 69.2 ± 4.7b |
| EARLY BLASTOCYSTS | 8.4 ± 3.4a | 13.5 ± 4.1a | 16.2 ± 6a | 16.8 ± 6.5a |
| LATE BLASTOCYSTS | 30.2 ± 3.9a | 25.6 ± 3.1a | 30.2 ± 3.5a | 31.4 ± 4.5a |
Different literals a-b show significant differences in rows at p≤0.05 (Mean ± SE). Considering an n in MIV=2,204 oocytes, DIV n=2,204 oocytes and embryos and blastocysts n=1,100 embryos.
Table 2.
Rate of late blastocysts produced by IVF from oocytes treated with resveratrol during IVM, after devitrification.
Table 2.
Rate of late blastocysts produced by IVF from oocytes treated with resveratrol during IVM, after devitrification.
| Resveratrol | ||||
|---|---|---|---|---|
| Late Blastocysts | 0 µM | 0.25 µM | 0.5 µM | 1 µM |
| Mean ± SE | 58 ± 20.9a | 41 ± 23.6a | 59 ± 7.3a | 64 ± 13.8a |
Same literal in a row shows no statistical differences (p≥0.05).
Table 3.
Semiquantitative analysis (Image J) of fluorescence levels of ROS and GSH in late blastocysts produced by IVF from oocytes treated with resveratrol during IVM, after devitrification.
Table 3.
Semiquantitative analysis (Image J) of fluorescence levels of ROS and GSH in late blastocysts produced by IVF from oocytes treated with resveratrol during IVM, after devitrification.
| Variables | Concentración de resveratrol | |||
|---|---|---|---|---|
| Fluorescence Intensity Units | 0 µM | 0.25 µM | 0.5 µM | 1 µM |
| ROS (Mean ± SE) | 34.722 ± 768.4a | 22.229 ± 4,314.2bc | 12.929 ± 1,789.3b | 24.447 ± 1,104.8c |
| GSH (Mean ± SE) | 25.733 ± 9,637a | 60.478 ± 10,586b | 94.446 ± 9,589b | 74.759 ± 14,800ab |
a,b,c, Literals between lines show significant differences (p≤0.05).
Table 4.
Number of nuclei present in early blastocysts prior to vitrification, produced by IVF from oocytes treated with resveratrol during IVM.
Table 4.
Number of nuclei present in early blastocysts prior to vitrification, produced by IVF from oocytes treated with resveratrol during IVM.
| Variable | Resveratrol concentration | |||
|---|---|---|---|---|
| Number of nuclei | 0 µM | 0.25 µM | 0.5 µM | 1 µM |
| Mean ± SE | 267±28.8a | 260±33.2ac | 185±11.7bc | 354±21.5d |
Literals a-b-c-d: between lines show significant differences between treatments (p≤0.05). Day 7 of culture.
Table 5.
Number of nuclei present in blastocysts produced by IVF from oocytes treated with resveratrol during IVM, after devitrification.
Table 5.
Number of nuclei present in blastocysts produced by IVF from oocytes treated with resveratrol during IVM, after devitrification.
| Variable | Resveratrol concentration | |||
|---|---|---|---|---|
| Number of nuclei | 0 µM | 0.25 µM | 0.5 µM | 1 µM |
| Mean ± SD | 109±4.3a | 90±12.3a | 147±21b | 150±19.4b |
Literals a-b: between rows show significant differences between treatments (p≤0.05). Forty-eight hours after devitrification.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



