
Submitted:
08 October 2023
Posted:
08 October 2023
You are already at the latest version
Abstract
Intergenerational Justice entitles the maximum retention of Earth’s Biodiversity. The United Nations COP 15 committed to the protection of 30% of Earth’s terrestrial environments to provide Biospheric Sustainability. However, proactive amphibian sustainability needs support from amphibian Reproduction Technologies, Biobanking, and Conservation Breeding Programs [RBCs]. These technologies include the reliable practice of stimulation of reproduction and the collection of sperm and oocytes, refrigerated and cryopreserved sperm storage, in vitro fertilization. Emerging technologies promise the perpetuation of species solely from biobanked biomaterials. However, although exemplary national programs for amphibian RBCs are now established in the USA and Australia, these potentials are not fully realised resulting in unacceptable biodiversity loss. To fully realise these potentials an independent and democratic global organisation is proposed to, 1) focus on the bioregions with the highest amphibian biodiversity, 2) maximise regional inclusion in international initiatives, and 3) provide for maximum community engagement and financial resources. We present the first overall multidisciplinary review of amphibian RBCs, including their geopolitical, cultural, and scientific background, along with their development and application to economically, efficiently, and reliably, perpetuate amphibian biodiversity. We provide genetic models, the potential for diploid cell cryopreservation to perpetuate species, along with significant new information on methodologies and techniques, and standardise nomenclature for specificity in publications and media consistency.
Keywords:
COP 15
; biobanking
; amphibian
; bioregionalism
; intergenerational justice
; ART
; multilateralism
; de-extinction
; artificial intelligence
; effective altruism
1. Introduction
A rapid and escalating series of declines and extinctions in amphibian populations resulting from widespread anthropogenic damage to global ecosystems demands proactive and innovative strategies for the perpetuation of amphibian biodiversity. The 2022 United Nations COP 15, titled "Ecological Civilization: Building a Shared Future for All Life on Earth", offers a promising blueprint for Biospheric Sustainability. This plan focuses on conserving, protecting, restoring, and sustainably managing biodiversity, with the ambitious goal of safeguarding 30% of Earth's terrestrial environments [1]. However, it's essential to recognize that habitat protection alone cannot fully achieve Biospheric Sustainability to ensure the Earth's biosphere continues to function in support all forms of life. Unfortunately, even within COP 15’s initiatives, protected ecosystems will undergo profound changes over the coming decades, leading to inevitable species population declines and extinctions in the wild [2].
Therefore, the full realization of proactive amphibian sustainability needs the vital and transformative technology of RBCs; Reproduction Technologies, Biobanking, and Conservation Breeding Programs [CBPs; 3–12]. RBCs can safely, securely, and economically, perpetuate Endangered and Critically Endangered species, and can produce individuals for repopulation, augmentation, or supplementation programs [8,12–16; Table 1]. However, the complete potential of RBCs lies in the perpetuation of species genetic diversity in Biobanks through restoring individuals from cryopreserved cell lines [7,10,17,18, section 5.6.], particularly of those otherwise neglected species with no ecological, cultural, or research significance [19,20; section 4.2. Range, Scope, and Networking].
Proactive amphibian sustainability recognises that most amphibian biodiversity exists in developing countries undergoing rapid cultural and economic changes and that the global framework for RBCs includes a biogeographic and biopolitical onus to develop regionally representative Biobanks, especially in developing countries with the highest amphibian biodiversity (section 3.1. Biogeography, Species Richness, and Habitat Loss) Achieving proactive amphibian sustainability necessitates seamless integration with local communities and collaboration with regional, national, and international programs. This includes active public influence and engagement along with a sense of ownership and entitlement [1,20,21,22,23,24,25,26,27,28,29,30]. Two fundamental principles underpin this approach: 1) The increasing assertion of developing countries in a multi-lateral geopolitical landscape, fueled by growing cultural independence, technical and educational expertise, and economic sovereignty. These factors contribute to embracing and driving proactive measures for amphibian conservation; and 2) The growing significance of non-governmental organizations (NGOs) in collaborative partnerships with individuals, the scientific community, businesses, and government entities. This multi-stakeholder engagement fosters a united effort to preserve amphibian biodiversity and promote sustainable practices.
The IUCN Conservation Planning Specialist Group, One Plan Approach to Conservation, includes all species populations regardless of their location through integrating in situ with ex situ approaches [24], including Biobanked individuals, as metapopulations [31], and defining them as “the total population of a species both in situ and ex situ including viable biomaterials in Biobanks capable of restoring individuals”. This concept within the framework of a redefinition of ‘nature’ as Metanature [Figure 1, section 1.1] satisfies the inclusivity of all biodiversity in an Anthropogenic Biosphere.
Amphibians provide the optimal vertebrate taxon to implement and exemplify ex situ management through RBCs, due to biological, technical, cultural, economic, and political factors. Cultural, economic, and political factors include their public presence and popularity and amenability to regional community programs including ecosystem protection [15,21,22,27]. Biological and technical factors include their highest proportion of threatened vertebrate species, insentience, small size, ease of care and reproduction, high fecundity, and proven amenability for RBCs including Biobanking of their genetic diversity [3,4,5,6,7,8,9,10,11,12,13,14,15,16,17,18].
Unfortunately, current initiatives have not satisfied the need for proactive amphibian sustainability, particularly in regions with the highest amphibian biodiversity [2,3]. Technologies for these initiatives are well developed [3,4,5,6,7,8,9,10,11,12,13]; however, the full potential of RBCs for species perpetuation hinges on the further development of a surprisingly neglected key technologies: the ability to restore amphibian species solely from biobanked somatic cells [18; section 5.15. Advanced Reproduction Technologies (aARTs)]. This neglect is even more remarkable when considering the extraordinary public support and financing for emerging global de-extinction initiatives to restore long-gone species from poorly preserved biomaterial [32]. To unlock the full capabilities of RBCs for it is crucial to prioritize and advance research in somatic cell biotechnologies. By addressing this gap, we can harness the tremendous potential of RBCs and fulfill our commitment to perpetuating amphibian biodiversity.
In this review we present the first comprehensive multidisciplinary synthesis of the state of the art and their geopolitical, cultural, and scientific backgrounds, to economically, efficiently, and reliably, perpetuate amphibian biodiversity. We emphasise the need for maximum community engagement and the critical role amphibian RBCs can play in broader biobanking and sustainability initiatives. We develop genetic models and highlight the potential for cell cryopreservation and cloning to perpetuate species through the first detailed review of amphibian cloning in the English literature, along with significant new information on methodologies and techniques. We present these subjects as section 2. Biopolitical, Biogeographical, and Biological Background; 3, Targeting Threats to Amphibians, 4. Conservation Breeding Programs; 5, Genetic Diversity and Management; 6. RBC Technologies; and 7. Biobanking – Economic, Political, Cultural Factors. We conclude with a road ahead to foster RBCs for Proactive Amphibian Sustainability through a democratic globally representative organization to, 1) focus on the bioregions with the highest amphibian biodiversity, 2) maximise regional inclusion in international initiatives, and 3) provide for maximum community engagement and financial resources.
Although we focus on amphibians many principles discussed in this review are applicable to other taxa, particularly biobanking as generic to all animalia. We primarily reference contemporary reviews published since 2015, and provide appendixes to expand core themes, along with a standardized nomenclature to enhance specificity and uniformity in publications, reports, and management plans. We follow the Associated Press conventions for the use of acronyms and address misconceptions in the literature.
2. Biopolitical, Biogeographical, and Biological Background
To fully maximize the potential of Amphibian Sustainability it is imperative to have a clear understanding of its political, cultural, and economic context, where empowerment toward Biospheric Sustainability must be fostered globally, and especially in economically challenged biodiversity-rich developing countries [1,24,26,27,28,29,30]. Otherwise, the future for both biodiversity and humanity is bleak [33,34,35]. Global incentives are growing for sustainability to address these existential challenges, and increasingly looking forward to a supportive moral framework of social and intergenerational justice. We contextualise these challenges through a background summary (Appendix A.1. The Anthropocene and Biospheric Sustainability).
2.1. Redefining Nature
An impediment toward Amphibian Sustainability, and the realisation of biotechnologies such as RBCs to perpetuate species, is the definition of Nature as "The phenomena of the physical world collectively, including plants, animals, the landscape, and other features and products of the earth, as opposed to humans or human creations" [36]. The future of Biospheric Sustainability requires a redefining of nature, through challenging the opposition between nature and humanity in Judeo/Christian theology [37,38], including the embracing life on or increasingly beyond Earth [39,40,41].
To remove the conceptual barriers that hamper the full potential of proactive amphibian sustainability we define the empowering concept of ‘Metanature’ as the ‘Collective phenomena of the physical world, including its components, – landscapes, plants, animals, humans, and human creations. These phenomena are recognized as interdependent, functional, and moral entities, within a Biosphere extending into space’ (Figure 1). In Figure 1. we also include a futuristic potential for species perpetuation in Genomic Restoration Databases we define as ‘Databases containing comprehensive genomic information for reviving or restoring living organisms.’.
2.2. The Anthropocene and Proactive Amphibian Sustainability
Amphibians are essential for ecosystem function [42] and enrich our lives at physical, emotional, cognitive, social, and spiritual levels. They support ecotourism, offer companionship as pets [43,44,45], and serve as valuable subjects for research [10,11]. These benefits not only enhance our current existence, but potentially extend to space colonization and terraforming [39,40]. Recent models indicate that declining trophic complexity, network connectivity, and community resilience, in natural ecosystems could lead to future amphibian rates double that of the previously predicted alarming levels [46,47]. Amphibians necessitate specific microhabitats, adequate food sources, and secure refuges, making them highly susceptible to even minor ecological disturbances. Furthermore, the rapid decline of insect populations, a primary food source for amphibians and for ecosystem stability, exacerbates these threats [48,49].
Most amphibian species inhabit developing countries, many of which grapple with a legacy of instability, poverty, and ongoing habitat destruction often without regard to environmental consequences [50; Appendix A.1]. Additionally, novel pathogens pose severe threats to entire suites of species with no immediate solutions in sight [51]. Therefore, the increased rates of decline and extinction of amphibians in the wild are formidable and urgent challenges [47,52,53]. The survival of many amphibian species in the wild depends on identifying threats and their amelioration supported by transformational economic and social changes [54,55]. These initiatives in tandem with amphibian RBCs can secure amphibians' irreplaceable contributions in the long-term to our ecosystems, culture, and overall quality of life [Appendix A.1].
3. Targeting Threats to Amphibians
The prioritisation of initiatives for proactive amphibian sustainability is dependent on species threats in the wild, their conservation status, and the effective targeting of RBCs.
3.1. Biogeography, Species Richness, and Habitat Loss
The highest diversity of anurans is found in tropical and subtropical regions, particularly on islands and mountains where varied habitats or isolating mechanisms promote speciation. Major biogeographical areas with high anuran diversity include Southeast Asia, Africa, SE USA, and Central and South America [56,57], with most new species discoveries in the tropical regions of South America, Asia, Indo-Pacific, and Africa [58]. The ten top countries in order of amphibian species richness are Brazil 1,022, Colombia 771, Ecuador 626, Peru 572, China 461, India 434, Mexico 396, regional Papua New Guinea 395, Indonesia 365, and Venezuela 363. Amphibian RBCs have mostly been developed in Australia the 15th country with 245 species, the Russian Federation the 69th country with 33 species, and the USA the 12th with 313 species [52].
Salamanders and caecilians are more restricted in their global distribution than anurans. Among the 10 salamander families, nine are predominately temperate, including those families generally placed near the base of salamander phylogenies with a center of diversity in Southeast North America. However, the highly threatened Plethodontidae are mainly tropical and includes 228 of the 555 currently described salamander species [59,60,61]. Half of these species are in the Mesoamerican Highlands of Mexico and South and Central America, with many new species being discovered in Brazil [62]. The distribution of the basal salamander family Cryptobranchoidea is mainly in Northeast Asia with a few species in Eastern North America [63]. Caecilians are mostly found in Central and South America, the South Asian tropics, and Eastern and Western Africa, but inexplicably not in Central Africa. The conservation status of many caecilian species is unknown due to their subterranean habitats [61,63,64].
Habitat loss is the main driver of amphibian declines, affecting ~65% of all amphibian species and up to ~90% of threatened species [52], with ~70% of all amphibian species in tropical regions being the most threatened [50]. The COP15 mandate to protect 30% of the terrestrial area offers hope for some threatened ecosystems and species, but including degraded habitats will not significantly protect many threatened amphibians [1]. Protecting biodiversity requires discrete targeting, where 30% of unprotected amphibian phylogenetic diversity, comprising 80% of salamander, and 65% of caecilian and anuran phylogenetic diversity could be protected by targeting only 2% of the global terrestrial area [65]. Overall amphibian species diversity also exists in a small percentage of terrestrial areas. For example, the small country of Ecuador is home to 8% (626) of species of which almost half are endemic [66], and islands with high anuran endemicity include Melanesia with 15% of species in 0.7% of the global terrestrial area [67], and Madagascar with 8% of species in 0.4% of the global terrestrial area [68]. Unfortunately, many amphibian species depend on mountain habitats with a high altitudinal range [69,70] and projected global heating to a likely 2.5°C will result in the extinction of many of these species [34].
3.2. Pathogens and Parasites
A major threat to amphibians is the skin pathogen, Batrachochytrium dendrobatidis (Bd), which spread globally in the early to mid-20th century, causing amphibian declines and extinctions. Bd now infects ~7% of species in ~70% of surveyed countries [71]. A congener is confined to Eurasia but if introduced would threaten the high salamander biodiversity of the Americas [59]. Inter-regional transmission of these pathogens can occur through keratinous skin, including on crayfish, nematode worms [72], fish [73,74], and migratory bird’s feet [75]. Parasites from four Phyla also infect amphibians with their species host range increasing with global warming [63,76,77]. The key to safeguarding amphibians amidst emerging infectious diseases lies in factoring in pathogens when developing conservation plans rather than focusing on the impossibility of total control [15,79].

3.3. Data Bases for Endangerment Status
Databases integration is essential to provide contemporary and accurate information on endangerment status [60,79,80, section 3.3. Data Bases for Endangerment Status]. A little recognised but highly significant threat is insufficient funding for the IUCN Red List and associated biological knowledge to support modelling that provides valuable insights toward species conservation [60,79,80,81,82]. More than half of the amphibian species on the IUCN Red List have either no assessment or outdated assessments [83], particularly concerning RBCs (section 3.3), and species in regions with high amphibian diversity such as developing countries, a need illustrated by the discovery since 2000 of ~150-200 new species per year [52]. Surprisingly, adequate annual funding of the IUCN and associated databases, representing a minuscule amount of less than 0.001% of global biospheric funding, would greatly help not only proactive amphibian sustainability, but also help save many thousands of species in other taxa.
3.4. Commercialisation and Trade
Commercialisation of any biological resources can yield both conservation benefits through increased engagement, and potential costs due to mismanagement or over-exploitation. The commercial amphibian trade of amphibian companion animals and exotic pets includes hundreds of individuals and tens of species [84,85]. However, it is impossible to determine if trade levels are sustainable due to the impossibility of tracking individual species using the World Customs Organization's international coding system [86], along with deficiencies in taxonomic data, including cryptic species and biological and threatened species data [87; section 4.3.1. CBP Founders and Data Bases]. Unfortunately, the provision of founders to institutional and private carers CBPs is conflated with other trade, resulting in excessive burdens, while criminal and irresponsible individuals and organisations profit [15,84]. These burdens to the conservation community particularly apply to onerous CITES prohibitions on trade involving Endangered and Critically Endangered species trade to the exclusion of private carers, including species that will be increasingly dependent on Private Carers for their survival [15].
4. Conservation Breeding Programs [CBPs]
Nomenclature; Convention - IUCN standard - Conservation Breeding Programs [CBPs]. Other: Captive Assurance Colonies, Captive Insurance Colonies. Ambiguous: Captive Breeding Colonies, Captive Colonies as these can include commercial breeding. Qualifiers: in situ - in range, ex situ – out of range. Redundancy; ‘survival’ in Captive Survival Assurance Colonies.
4.1. Preferred terms for field conservation
The amphibian conservation community uses a confusing plethora of terms for field conservation. We researched credible authoritative sources from different disciplines to rationalise a standard terminology for the field of RBCs [Table 1].

Table 1.
Preferred terms for use for RBCs and field projects.
| Preferred Term | Misnomers – alternatives | |
|---|---|---|
| Species Programs | ||
| Head Starting | The raising of eggs or embryos harvested from nature to advanced stages for release. | [88] |
| Repopulation | Repopulating a lost species in previously populated habitat. | Rehabilitation. Reintroduction. |
| Augmentation | The addition of individuals from captive-bred or stable, wild populations to support extant populations. | Supplementation |
| Translocation | Translocation is the transfer of a species from one location to another within their home range. | [89] |
| Relocation | Relocation is moving animals under immediate threat of extirpation from one location to another within their home range. | [89] |
| Assisted Migration | Moving populations to locations outside the historical species range, mainly in response to global heating. | [90] |
| Habitat Programs | ||
| Mitigation | Minimising damage and maximizing the eco-sustainability of environments. | [91] |
| Rehabilitation | Rehabilitation focuses on the reparation of ecosystem processes …. to repair the capacity of ecosystems to provide habitats for biota and eco-services. | [91] |
| Restoration | The aspirational target of restoring native ecosystems. | [92] |
4.2. Range, Scope, and Networking
Conservation Breeding Programs can be in- or out-range (in situ or ex situ), in private or institutional facilities, linked to community-based programs and habitat protection, or be partially or totally self-supporting through volunteers, ecotourism, and trade [15,24,93,94,95]. They offer an opportunity to guarantee species survival through long-term commitment and management strategies complementing their focus’ on perpetuation, repopulation, conservation research or education, or community engagement [10,11,43,44,45]. Institutional CBPs alone cannot support the current need for CBPs [3,96]. Nevertheless, the cascade of species declines toward extinction in the coming decades [96,97], but Private Carers and their NGOs can fill the gap [15]. Unfortunately, many species do not appeal to either institutional or private CBPs [19]; however, with targeted resource allocation toward RBCs these species could be saved until perpetuation solely in Biobanks [20].
Surprisingly, when cryopreserved sperm can minimise CBP size to a few living individuals, Canessa [98] suggested a conflict of choice for CBPs between a large captive population for a single species fully ensuring against extinction or smaller CBPs for several species. A similar sentiment was expressed by Tapley et al. [19], who suggested neglecting species to extinction that could not predictably be restored in nature due to habitat destruction, pathogens, or other causes, which will include many amphibians over the long term. Tapley et al. [19] was furthered by Griffiths [98] “When selecting species for captive programs, dilemmas may emerge between choosing species that have a good chance of surviving after reintroduction because their threats are reversible and those that are doomed to extinction in the wild as a result of irreversible threats”.
These dystopian neglects are particularly morally unacceptable for species whose habitat is irreplaceably lost and dependent on CBPs for their survival until RBCs enable their perpetuation in Biobanks for timely restoration in the future [4,9,10,17,18,20]. Furthermore, the accurate prediction of species’ future potential survival in nature is very difficult. For instance, some species are repopulating as they adapt to pathogens infested environments [99], assisted evolution or vaccination could enable pathogen-exposed species to survive [100], and rapidly evolving ecosystems including urban green environments could provide increased survivability through pathogen exclusion. We have defined the management of Triage 1, and particularly Triage 3, to include the perpetuation of all species, where RBCs can substantially reduce the size and cost of CBPs, and potentially perpetuate species solely in Biobanks (section 4.3.2. The Species Survival Triage and RBC Prioritisation).
Perhaps the greatest potential for CBPs is found in citizen conservation through private carers and their NGOs, a largely untapped and unsupported public resource that already maintains many threatened species and could support many more, along with local community engagement, habitat protection, or restoration [15]. The increasing support of private carers and their NGOs along with institutional RBCs focussed on species in immediate need, could economically perpetuate all currently AArk-recommended species, and as facilities grow the expected cascade of species over the next few decades [3,15,101].
The movement toward amphibian RBCs supporting genetic diversity through sperm storage is gaining momentum through assisted gene flow between natural populations and CBPs [103], citizen science driven field collection and cryopreservation, and sperm biobanking [104,105]. In a direct link between the climate catastrophe and the use of RBCs, following the unprecedented 2019/20 megafires in Australia sperm is being cryopreserved in the field from both burnt and unburnt sites along the East coast of Australia [106].
4.3. Species Prioritisation for RBCs
Having a logical framework for species prioritisation in RBCs is crucial, primarily based on local expertise as exemplified by Amphibian Ark (AArk) assessments [3]. The minimum effective population size in nature to recommend RBCs depends on the species’ autecology, demographics, and reproductive cycles including age of maturity and fecundity, and the area of potential habitat [107,108,109,110]. A common bias in the RBC literature toward the importance of institutional agendas is the generalisation of the term ‘Threatened Species’ that includes Vulnerable, Endangered, and Critically Endangered species. The survival of Vulnerable species can be secured through management practices such as habitat protection or replacement, which are primary conservation goals [3,29,95,111]. The full potential of RBCs prioritises Critically Endangered species assessed as under immediate threat of extinction, then RBCs of various impacts for Endangered species. Additionally, there exists an opportunity to biobank newly discovered species while simultaneously describing and vouchering specimens (section 6.4. Material Types and Vouchering).
4.3.1. CBP Founders and Data Bases
The semi-autonomous Amphibian Ark is the central authoritative global organisation supporting amphibian RBCs. The AArk portal lists the activities of CBPs globally and provides contemporary information through a newsletter [3]. Based on the regional expertise of biologists and field conservationists, assessments of species endangerment status lists approximately 400 Endangered or Critically Endangered species. Almost 100% of these species are also recommended for RBCs [112]. Other important perspectives toward the perpetuation of endangered species are the Royal Zoological Society of London, Evolutionarily Distinct and Globally Endangered (EDGE] species lists, where emphasis is placed on the representation of phylogenetic lineages [113,114] and recognition of Data Deficient species that are often rare and therefore liable to extinction [115].
At the time of this review, the IUCN Red List does not offer a rational and accurate list of Biobanking [Table 2], or Species Recommended for Community-Based Conservation Programs (CBPs) labelled as ‘ex situ conservation” [Table 3]. Instead, it primarily lists Least Concern species and no caecilians. Notably, there needs to be more consistency between AArk Biobanking [112] and IUCN lists [83]. A framework for evaluating the impact of the IUCN Red List of Threatened Species was published but the global RBC community needs informing of any moves toward its implementation [116].
Besides their ecological function, phylogenetic significance, or iconic status, species can be also targeted for RBCs for traits that also contribute to direct human psychological well-being. These traits are particularly significant for displays in zoos, with private carers as companion animals or as contributors to CBPs, and as components of urban biodiversity, for instance, attractively coloured or patterned amphibians, large amphibians, or amphibians of mythological significance [15,43].
4.3.2. The Species Survival Triage and RBC Prioritisation
The application of Amphibian Sustainability requires the most efficient and reliable direction of resources toward species. We recommend a triage of care based on the immediacy of need and potential support.
Triage 1. Vulnerable and Common Species - Survival in Nature - Species maintaining viable populations in nature in protected or unprotected habitats, with adequate effective reproducing population sizes. Field activities could include augmentation, repopulation, and population of rehabilitated, restored, or created ecological systems.
Triage 2. Endangered or Critically Endangered species with effective reproducing populations in nature that could be maintained or recovered. RBC priorities: genetic diversity maintained in CBPs and Biobanks, along with broodstock, in anticipation of progeny release to augment existing populations, or to repopulate rehabilitated, restored, or created ecological systems.
Triage 3. Species are unlikely to survive in nature for the foreseeable future due to irrecoverable habitat loss or other threats. RBCs are critical for these species and can include females without studbook management as display or pet species, with their generic diversity perpetuated through Biobanked cryopreserved sperm. Once technologies are developed for the recovery of females from viable cryopreserved diploid cells or tissues these species genetic diversity could be solely perpetuated in Biobanks. Cost is moderate with CBPs, and eventually low with Biobanking for perpetuity.
4.4. Citizen Conservation, Private Carers, and CBPs
“Ultimately, breeding space and staff is limited in zoos. With dedicated private keepers, we can expand our capacities while sharing and gaining knowledge — a win-win situation for breeding and conserving endangered species”. Anna Rauhhaus , 2023.
Citizen conservation turns citizens into practicing conservationists: raising awareness, motivating people to get directly involved, and bringing together different areas of expertise to make a significant contribution to biodiversity conservation. Citizen conservation unites private amphibian carers and professional conservationists in an inclusive and powerful societal approach to support proactive amphibian sustainability. Zoo-based CBPs for amphibians are limited in number and tend to focus on charismatic or iconic species [31], or their regional programs [8], however, private carers CBPs are only limited by their interests [15].

There are exciting possibilities for greater incorporation of the vast global potential of private carers, with their proven track record with a wide variety of anuran and salamander species. This could lead to conducting CBPs for threatened species at greatly reduced costs and building public and political support, both within and outside the species' native range. Private groups play a crucial role in ensuring the survival of threatened amphibians outside their natural habitat and foster strong relationships for in situ conservation efforts, for instance, the Responsible Herpetological Project [117]. Private carers have the facilities, time, passion, and knowledge to care for and reproduce threatened species [15, Table 4, Figure 2]. A vast opportunity for species perpetuation also exists where species in private carers’ collections as colour morphs or as domestic varieties can be used to re-establish overall species genetic diversity through Biobanked sperm [15,16]. However, CITES restricts species provision to NGOs and private carers outside their national boundaries, thus hampering initiatives toward private carers CBPs.

The success demonstrated by Private Carers in successfully reproducing numerous species of anurans and salamanders underscores their excellent standards of care and highlights their potential to make significant contributions to CBPs if provided with the opportunity [15, Table 4, Figure 2].

5.1. Genetic Management Guidelines
A widely suggested global target for captive breeding programs is to retain 90% of the source population heterozygosity (Ht/Ho) for 100 years. Concerning biobanking strategies, this may not necessarily equate to sampling every single individual in the living population, but rather capturing the gene pool of the genetically effective population size (Ne), as opposed to actual or census size (N). Ne is the size of a representative population that would give rise to the same variance of gene frequency, or rate of inbreeding as the actual population under consideration. The effective size is typically much smaller than the number of potentially breeding adults in populations, averaging an order of magnitude lower than census population sizes [126]. Unequal sex ratio, family size variance, and population size fluctuations are the major variables predicted to affect Ne. Selection of animals for biobanking must consider the variables that affect Ne for a given species or population, whilst seeking to make efficient use of resources to preserve maximum diversity through a minimum number of samples [138].
The ‘Amphibian Population Management Guidelines’ [110] detail the management of genetic diversity in CBPs. The guidelines provide target population sizes and minimum founder numbers for a range of scenarios to achieve maintenance of genetic diversity, albeit without considering the use of biobanking and particularly RBCs Howell et al. [13,14] quantified the potential cost and genetic benefits of Biobanking where supplementing an anuran CBP with frozen founder sperm through in vitro fertilisation every generation results in major cost reductions, minimised inbreeding and greatly reduced captive populations, that would exceed the genetic retention benchmark of 90% of source population heterozygosity, and even allow 95% or 99% heterozygosity targets to be realistically achieved. Modelling with two other anurans similarly showed that back-crossing with frozen founder sperm with every generation would minimise rates of inbreeding and reduce colony sizes and program costs [14].
A major challenge in CBPs is the loss of genetic variation through selection for domestication, leading to individuals being poorly to wild environments [131,140]. Cryopreserved gametes may represent a reservoir of rare but significant alleles that could help address this challenge [16,141]. A structured approach is needed to determine the degree to which cryopreserved sperm can contribute to maintaining or enhancing genetic diversity including 1) the number of founder animals, 2) the number of additional biobanked founders, and 3) the initial frequency of a given allele in the founder population [110]. The number of founders and initial allele frequency allows a calculation of the likelihood that that allele will persist into subsequent generations (Figure 3). More difficult to quantify is the significance of an allele to the future population adaptation to changing conditions. Rare alleles could be highly significant toward adaptation to changing environmental conditions, such as effective camouflage appropriate for specific vegetation cycles or pathogens [142,143]. These uncertainties have resulted in insufficient estimates for Biobanking needs (110, Table 5].
Whilst inbreeding depression and loss of diversity have been shown to contribute to extinction risk, these observations typically rely on the assumption of naturally outbreeding populations. Amphibian species have been challenging to profile in terms of population genetics. Furthermore, many populations are isolated and likely to be naturally inbreeding rather than outbreeding. The impacts of inbreeding depression and diversity loss on extinction risk in inbreeding amphibian populations have neither been examined experimentally nor modelled and are therefore difficult to predict accurately. Understanding how CBPs to support wild populations will affect genetic diversity and extinction requires timely, baseline genetic data, thorough characterisation, and documentation of living and biobank breeders, and prospective genetic modelling. The fundamental management issue is at what effective population size in nature should CBPs be implemented, and how does the relationship between a CBP’s genetic diversity and the recipient population size and genetic diversity affect management strategies such as assisted gene flow [130-131; section 5.1.2. Assisted Gene Flow].
5.1.1. Biobanking Genetic Diversity and CBPs
The relative approaches to using RBCs, dependent on species needs, can be considered relative to a complete program based on the perpetuation of a threatened species with imminent extinction in nature, and with only a few if any vouchered specimens in museums. Both Mahony and Clulow [145] and Kouba et al. [146] recommended biobanking of all Threatened species (Vulnerable, Endangered, and Critically Endangered species), in contradiction to AArk recommendations (section 4.3.1. CBP Founders and Data Bases). Amphibian Ark recommends that at least twenty pairs of animals (or groups of individuals) are collected as founder animals for CBPs, ideally unrelated, and successfully reproducing, access AAR Founder Tool [150]. Many more than this number may be needed to ensure that twenty pairs survive and successfully reproduce.
In any case, the maintenance of genetic diversity in CBPs without using cryopreserved sperm is inefficient and costly, and even with Amphibian Population Management Guidelines CBP populations lose genetic diversity with each generation [110]. The loss rate of genetic diversity in CBPs depends on the program period, the generation time, studbook management, the CBP population, and the loss rate of founders due to natural or stochastic mortality. Schad [110,150] recommends a minimum number of 20 founders to capture 97.5% heterozygosity. However, dependent on the generation time and lifespan, even with studbook pairing, to maintain only 97.5% heterozygosity in CBPs for periods of only 10-25 years requires CBPs with 10 to 25 each of males and females and CBP populations from 80 to 1,500 individuals [110,150]. Cryopreserved sperm enables male founder numbers to potentially be much higher, the number of generations is effectively one, and the indefinite perpetuation of the initial genetic diversity. Even if fertilisation rates with in vitro fertilisation are low, the high fecundity of subsequent adults in most species can produce numerous offspring [Figure 4]. Species could even be perpetuated through females of amphibian varieties in private collections CBPs to provide oocytes with genetic diversity opportunistically reestablished with cryopreserved sperm, [15,16].
In agreement with the AArk model [110,150] we propose a foundational population, at the time of rescue of a species in a CBPs with sperm Biobanking, of at least 20 live females and 20 live males, along with the maximum practical number of males Biobanked sperm, including sampling from the wild and male founders. For biologically unexplored species up to 12 males and 12 females are vouchered, sperm and male and female tissues Biobanked, and all individuals sampled for phylogenetic analysis. A bias toward females with a total founder population of 40, 20 adult females and 10 adult males, could be beneficial to the production of progeny if releases are a priority [149]. The rate of loss of genetic diversity in populations in the wild will not be the same as randomly predicted because of environmental selection against unfit individuals. A population size at which ≤ 100 of effectively reproducing adults in the wild is predicted to suffer a 10% loss in ‘fitness’ [148], through loss of genetic diversity over 30 years [151].
5.1.2. Assisted Gene Flow
Assisted Gene Flow involves the translocation of genes between CBPs or biobanks, and wild populations comprising a species metapopulation (core population) [31,129] to maintain or increase allelic diversity [genetic variation] [131]. Assisted gene flow can enhance fitness, reproductive capacity, and the evolutionary adaptability. However, it can also lead to outbreeding depression and loss of alleles [151], genetic incompatibility [108,139], risk of disease transmission, and consequently the recipient populations survival [131]. The beneficial or detrimental effects on populations of assisted gene flow depend on the multiplicity translocated of genes and alleles, the proportions of the introduced genotypes to recipient genotypes, environmental selection toward the introduced genotype, overall population size, and subsequent genetic drift [108,129,139].
In most cases, assisted gene flow toward subpopulations should utilise pooled genetic diversity from the core metapopulation, to minimise inbreeding depression and to avoid supplementing deleterious genes, rather than considering each sub-population as a genetic unit [152]. Environmental management, in conjunction with genetic management, could optimise the maintenance of these populations, particularly if their cultural significance encourages community participation and environmental management can be emphasised in public education [153].
In the case of isolated populations characterised by their unique genetic variation, assisted gene flow should be within the population, where foreign genes could result in male/female incompatibility, low fecundity, or even ecological mismatches with reproductive cues, and therefore reduce fecundity and survival. This limitation has been substantially evidenced in a cline of a small ground-nesting frog with little dispersal ability but may be found in other wide-ranging species with similar autecology’s and isolating mechanisms such as low dispersal [139].
Assisted gene flow with wild populations can yield benefits only if the translocated genomes comprise a high percentage of those of the wild population and are highly positively selected. Frankham et al. [151] demonstrated that, in the absence of inbreeding depression, assisted gene flow was beneficial only when the introduced genotypes had much higher fitness than local genotypes, and where fitness traits were controlled by a few genes of large effect. Significant genomic turnover only occurred when the selective advantage of the adaptive traits, and translocation fraction, were extremely high. When the adaptive trait was controlled by many loci of small effect, the benefits of assisted gene flow took over 10 generations to realize, however, the generation times of many amphibian species are comparatively short [154]. Assisted gene flow was harmful after only 10-20 generations in the presence of strong outbreeding depression, or through the introduction of deleterious genes. Various genetic traits are manifest throughout species ontogeny, where dependent on the amphibian species, fungal pathogens were shown to be more lethal to either the tadpole or the adult stage [155].
Assisted gene flow can be implemented at any life stage. However, to maximise natural selection and minimise the effects of genetic selection in RBCs, the most effective strategy is the introduction of genetically and allelically diverse egg masses or early larvae to wild populations. This approach could also be accomplished through on-site collecting oocytes from gravid females, followed immediately by in vitro fertilisation by stored or fresh sperm. In any case, more genetic and demographic monitoring of artificially produced populations is essential to inform adaptive management [156].
5.2. Genetically Unbiased Founders in CBPs and Biobanking
Sampling founders for RBCs, whether as live or Biobanked individuals, demands the selection of genetically unbiased individuals to ensure maximum heterozygosity and adaptability to a future of rapidly changing global environments [108,129,131,139,151,157]. Sperm sampling includes anthropocentric bias, terminal investment effects bias, and sperm yield bias. The selection of males for field sperm collection is preferably undertaken seasonally during major spawning events [158,159].
Anthropogenic bias can occur through targeting traits that do not represent maximum heterogeneity such as peripheral sub-populations [160,161], or more easily captured, more vocal, or larger males. Whereas, in most anurans preferred males are smaller than females, possibly as an evolutionary mechanism to maximise food resource allocation toward oocyte production [162], maximization heterozygosity can occur with smaller ‘sneaker" males [163] or through size-independent delayed partial clutch fertilisation [164].
Terminal investment bias has been shown in amphibians where fungal-infected males in two anuran families produced greater sperm numbers [165], or Myxosporidean infected males yield very high sperm numbers along with numerous spores [166]. Myxosporideans infect all anuran life stages [167] of three of five Australian anuran families [168], and in Rhinella a focus genus for RBCs [146,169].
Sperm yield bias in sperm numbers, concentration, and motility, is dependent on male mass and corresponding testes mass across metapopulations as dependent on environmental factors such as seasonal cycles, climate, and prey availability [170,171,172]. The relative effects of genetics and physiological adaptation on these responses are uncertain [173,174]. The effective sperm yield after recovery from cryopreservation may also be lower in inbred males, with reduced sperm quality, total motility, speed, and viability [173], and effects on post-release individuals [174].
5.3. Assisted Evolution.
Similar – Directed evolution, Artificial selection.
Assisted Evolution provides novel genotypes with increased survivability through introducing genes, modifying existing genes, or selecting for mutations [175]. Genetic engineering has yet to be effectively applied toward proactive amphibian sustainability, with the most promising genetic engineering application being increased immunity [176].
6. Reproduction and Biobanking Technologies
Similar: Assisted Reproductive Technologies [ART]; Genetic Resource Banking, Genome Resource Banking.
To improve readability, we use abbreviated terms for instance ‘sperm’ for ‘spermatozoa’, and ‘hormonal stimulation’ for ‘exogenous hormonal stimulation’. We use ‘oocytes’ for unfertilised ‘eggs’, Salamandroidea will be generically referred to as salamanders, with internally fertilising Salamandridae and the externally fertilising Cryptobranchoidea specifically referred to where appropriate. Convention – Reproduction Technologies [RTs]. Other: Assisted Reproductive Technologies [ART], Reproductive Technologies, Amphibian Reproductive Technologies. Convention – Biobanking. Other: Genome Resource Banking, Gene Banking, Genetic Resource Banking.
We present an overview of the development and current state of current amphibian reproduction technologies [177], and two methodological challenges for assessing sperm fertility. We provide the first review of amphibian cloning in English and standardised nomenclatures for RBCs.
6.1. History
The application of amphibian RBCs began in the late 20th Century, with hormonal stimulation in Russia in 1989, and sperm cryopreservation in Russia [12] and Australia [16] in 1997/1998 (Figure 5.; Appendix A.6, Table A2). A broad review of possibilities for RBCs to maintain genetic diversity was presented in 1999 [4]. Sperm-based RBCs remain the primary method for maintaining genetic diversity [12,16,141] and include more than 34 anurans and 5 salamanders [7,8,9], with assisted gene flow using sperm between wild populations and RBCs [105,125,146]. Cryopreserved anuran testicular sperm fertilised oocytes at the turn of the 21st Century in Russia [178,179] and in Australia [180], followed by urinal sperm in 2011 [181]. These techniques were then applied to salamanders [12,60,120], with several of these anurans and salamanders having subsequently reproduced [125]. The development and application of amphibian RBCs largely focused on sperm collection, its refrigerated or cryopreserved storage, and its use for in vitro fertilisation [12,16,141]. The rate of research has recently increased in correspondence with the establishment of national programs for amphibian RBCs in the USA and Australia. However, the full potential of RBCs needs the final development of heterocytoplasmic cloning from cryopreserved nuclei (Appendix A.5). Surprisingly even though heterocytoplasmic cloning with fresh nuclei was established in anurans as early as 1963, and in salamanders in 1971 (Table 7), heterocytoplasmic cloning from cryopreserved nuclei has received little recent attention [18].
6.2. Ethics
“The humanest possible treatment of experimental animals, far from being an obstacle, is actually a prerequisite for successful animal experiments.” [182] through 1) reduction - minimisation of animal numbers, 2) refinement - minimize stress and trauma, and 3) replacement through other means [183,184]. Sperm collection, refrigerated storage, cryopreservation, and oocyte collection and storage, and in vitro fertilisation, only need fundamental biological laboratory and animal handling experience. Techniques for diploid cell culture and restoration technologies such as cloning, and intracytoplasmic sperm injection [ICSI], require sophisticated laboratory facilities and technical expertise.
6.3. Assisted Mating, Spawning, and Reproductive Maturity
Most temperate anurans and salamanders show intermittent circannual sexual maturity, while most tropical species have continuous circannual sexual maturity [171,172]. Temperature primarily influences reproductive maturity with precipitation then triggering spawning [185]. Environmental simulation [12] and hormonal stimulation [12,141,186] inducing mating and spawning. Low temperature brumination in some cool temperate species increases mating and spawning responses to hormonal stimulation [12] and promotes gamete collection [181]. Different rearing temperatures between male and female axolotls optimised gamete maturity [122].
6.4. Material Types and Vouchering
Genetic diversity can be held as tissues, cells, viable nuclei, mitochondria, DNA, mRNA, and Genomic Restoration Databases [187], however, only cryopreserved cells and tissues can currently recover individuals [4,9,10,12,16,188]. Vouchered specimens support proactive amphibian sustainability by relating genotypic to phenotypic characteristics within ecological contexts [146,189,190,191,192] by providing information on the life, history, biology, longevity, pathology, and growth rates of amphibians [187,191]. This information contributes to the epidemiology of amphibian extinctions and declines from Chytrid [187,193] and assists with balancing conservation with economic development in highly amphibian biodiverse regions [190].
6.5. Life Stage and Sample Collection
The availability of different amphibian life stages and their ability to provide suitable cells, tissues, or whole organisms for RBCs guide sampling approaches [187]. Amphibians are highly fecund and oocytes or larvae from the wild, or from head-starting programs, can provide founders for CBPs and embryo stem cells for cloning [194]. As life stages progress toward maturity in the wild the possibility of pathogen transmission increases. On the other hand, survival also evidences environmental adaptation and genetic fitness [156]. Males of many temperate anurans mature in one year, with most over two years, and warm temperate anurans generally in less than one year [134] and as little as three months [195]. A range of different life stages of threatened species can also be sourced from zoos or private carer CBPs [102]. Kouba and Julien [105] generalised caution against using early life stages, although some species’ males maturation times are only six to eight months including founders from tadpoles in their Lithobates [Rana] servosus RBC.
6.6. Sperm Qualities - Activation, Motility, Speed, Vitality, and DNA integrity
Misnomers for sperm speed [velocity] - ‘forward progressive motility’, ‘backward progressive motility’. Vitality as viability. Mitochondrial sheaths or collars as ‘mitochondrial vesicles’
Amphibian sperm consists of a head consisting of a condensed nucleus surrounded by a plasma membrane. Mitochondria are in a sheath along the sperm head, in a collar at the base, or along the flagellum [196,197]. The misnomer ‘mitochondrial vesicle’ for these structures was casually used in 2003 [198], despite a lack of vesicular structure, and ‘mitochondrial vesicle’ being a formal scientific term for vesicles containing mitochondria found within cell organelles. It was further misrepresented as a genuine cell organelle, and as in 2003 [199] without reference to precedence is expert literature [196]. Following this misrepresented, the misnomer has unfortunately proliferated in the amphibian RBC to the stature of an article in 2022 titled “The Impact of Time and Environmental Factors on the Mitochondrial Vesicle and Subsequent Motility of Amphibian Sperm” [200], along with its generic misuse by the same research group [104]. The four basic metrics of sperm motility are the percentages of 1) immotile sperm, 2) activated sperm showing at least flagella or twitching movements, that extends to 3) swimming sperm, and 4) sperm speed as absolute or directional [201]. Computer-aided sperm assessment [CASA] provides accurate metrics of sperm swimming [201]. Biochemical metrics are plasma membrane integrity and DNA integrity [12]. Comparisons between reproduction technologies are confounded by different metrics, ambiguous terminology, variations between testicular or hormone-induced sperm, and oocyte quality, along with a variety of physical and chemical techniques [141,203,204,205).
6.7. Oocytes Collection and Storage
Oocytes can be collected from gravid females in the field or captivity [206]. Females are generally more recalcitrant to hormonal stimulation than males and often require a priming dose of hormones [12,180,186]. However, ultrasound has enabled better targeting of hormonal stimulation for both oocyte and sperm production [120,194,207]. Oocytes can be collected from the oviduct by abdominal massage, spawned into physiological saline, or post-mortem from ovarian excision [12,16,180]. The viability period of ovulated oocytes depends on the species natural spawning temperature, storage temperature, and the osmotic and ionic environment. Cool storage has extended the oocyte viability of tropical or sub-tropical species for hours [208], and with cool to cold-temperate species up to a week [12]. Accumulating evidence suggests that dry storage is optimal, but that isotonic solutions maintain fertility for hours in some species. In female/male compatibility studies, using oocytes immediately on expression through abdominal massage or canulation is recommended due to a rapid loss of fertility [139]. However, in general studies oocyte infertility is physiological and would not be expected to significantly affect the genetics of progeny [Appendix A.2. Oocytes Viability Period and Storage Environment).
6.8. Sperm Collection from Testes Macerates
Considerations when collecting sperm include sourcing through testes macerates or hormonal stimulation, donor’s stress and trauma, genetically unbiased collection, and avoiding pathogen dissemination [Table 6, section 3.2. Pathogens and Parasites). The maceration of the testes into physiological saline reliably provides large quantities of highly concentrated sperm [16,180,204,205,206], with euthanasia, dissection of testes, and maceration achieved in a few minutes [139]. Testicular sperm is convenient for testing outbreeding depression between multiple female/male compatibilities between sub-populations [139], where large quantities of sperm of consistent pH and osmolarity must tolerate refrigerated storage for ≤ 12 hr. without fertility loss. In contrast, hormonally induced sperm is subject to changes in motility, quality, and concentration [201]. The use of testicular sperm has been criticised by a misleading argument that it cannot be collected from threatened species [105,125]. Due to the high fecundity of most amphibian species (Figure 7), numerous surplus individuals of most threatened species are available [16,60,204). Therefore, euthanasia to collect testicular sperm is acceptable from: 1) males taken without threatening the population, 2) CBPs surplus males, and 3) males vouchered for taxonomy.
6.9. Sperm Collection through Hormonal Stimulation
Sperm can be collected using hormonal stimulation, then later optimised with metrics for success including yield, quality, and production peaks [201]. However, hormonal stimulation produces few if any low-quality sperm in some anurans [209] and salamanders [210]. Salamander sperm can be collected in pre-spermatophores as gelatinous fluid expelled by massaging the lower oviduct and cloaca [120], as spermic urine [210,211] semen [118], or deposited spermatophores [119]. The deposition of spermatophores does not require handling after hormone injection or topical application, however, sperm collection from spermatophores remains challenging [210], and cryoprotectants do not penetrate the outer shell of spermatophores [119].
Hormonal stimulation produced extremely low concentrations spermatozoa with none swimming, and no reproductive behaviour between pairs with the Endangered, Hamilton's frog [Leiopelma hamiltoni, 209], and whose three cogenerers are the only living representatives of a highly significant phylogenetically basal Anura family [Leiopelmatidae] [113,114]. A similar response is predicted with Archey’s frog, L. archeyi. the globally top priority amphibian species for conservation actions on the Zoological Society of London’s, Evolutionary Distinct and Globally Endangered program [113].
6.10. Sperm Donor Stress, Trauma, and Pathogens
Sperm collection from testicular macerates only requires euthanasia, while hormonally induced sperm generally requires injection and then prolonged stressful handling on multiple occasions over many hours. Hormone injections are unsuitable for small delicate salamander or anuran species due to stress and trauma [60]. However, has been recommended for salamanders in general [210]. These species include most plethodontids, which include many threatened species, and that comprise 65% of salamander species [52]. Alternative administration methods for hormones include topical or nasal application [60,201,212]. Hormone injection has been used successfully for larger salamanders [60,120,207], including for the commercial production of Chinese giant salamanders [Andrias clade; 60]. Sperm storage, even during cryopreservation, can harbour a wide range of pathogens [213]. The collection of testicular sperm should limit pathogens to internal parasites and viruses. However, hormonally stimulated spermic urine could also include contamination by bacteria, fungi, and external parasites [63].
Table 6.
Factors to consider when selecting testes macerates or the use of exogenous hormonal stimulation of spermiation to collect sperm.
Table 6.
Factors to consider when selecting testes macerates or the use of exogenous hormonal stimulation of spermiation to collect sperm.
| Collection Method | Stress | Trauma | Sperm Yield | Donor Size | Pathogens |
|---|---|---|---|---|---|
| Testes Macerates | Low. | Low. | Very High | All Species. | Endo-parasites, virus. |
| Spermic Urine Anurans |
High, dependent on collection period and technique. | Unacceptable for very small species. | Moderate - Very Low. | Unacceptable for very small species | Endo-parasites, exo-parasites, intestinal parasites, fungus, virus, bacteria. |
| Hormonal Stimulation Salamanders |
High, dependent on collection period and technique. | Unacceptable for small to very small species. | Moderate - Very Low. | Unacceptable for small to very small species | Endo-parasites, exo-parasites, intestinal parasites, fungus, virus, bacteria. |
6.11. Refrigerated Storage of Sperm and Testes
Refrigerated storage of anuran sperm enables acclimation to cryoprotectants and provides fertile sperm for several days to a week for delayed in vitro fertilisation, transport between facilities, or for later cryopreservation [12,16,105,146]. Salamander sperm has a much shorter viability period than anuran sperm with only a day before substantial declines occur [12,16]. Seasonality can impact sperm quality and its amenability for refrigerated storage [159]. Antibiotics and oxygenation can extend storage life [214].
6.12. Sperm Cryopreservation
Terminology. Recommended – formulation. Misnomer slang – cocktail.
Aqueous based formulations of cryoprotectants, termed cryodiluents, are mixed with testes macerates, or with spermic urine, to produce cryosuspensions for cryopreservation [16]. There is a natural variation in the amenability of different species of sperm to cryopreservation, but success is now reliably achieved with both amphibians and salamanders [12,16,60,141]. Experimental evidence shows that cryopreserved mammalian sperm will remain integral for more than 200 years even under the effects of space radiation [41](Appendix A.3).
6.13. Artificial Fertilisation
Mis-described as including only ‘in vitro’ fertilisation and not artificial insemination, along with ‘dry fertilisation’ [Silla and Kouba, 2022]
Artificial fertilisation can be potentially achieved in all amphibians, and in vitro fertilisation is commonly used for anurans and salamanders [16,210], with recent advances for salamanders to artificial insemination [123], with oviviparous caecilians requiring artificial insemination [64]. Many thousands of swimming sperm are needed for fertilisation, showing a high natural selective bias toward female sperm choice through favoured sperm traits to locate the oocyte and then to penetrate the gel and fertilise [12]. Our knowledge of artificial is limited by the number of amphibian species and families providing comparable metrics and techniques, of sperm concentration and quality, fertilisation environments including osmolarity and ionic formulation, and fertilisation period [16,60,210]. We provide a substantial background for improved comparative studies in Appendix A.4. Artificial Fertilisation.
6.14. Intracytoplasmic Sperm Injection
Even sperm with integral plasma membranes could be used for intracytoplasmic sperm injection. Surprisingly the utilisation of ICSI with immotile integral sperm [215,216] has received little attention. However, although Herbert [202 used ICSI to produce embryos these did not generate metamorphs due to technical failures.
6.15. Advanced Reproduction Technologies [aARTs]
The potential of aARTs to perpetuate amphibian biodiversity includes many parallels between amphibians and fishes [4,217,219]. The large oocytes and embryos and current technologies preclude the possibility of amphibian oocyte cryopreservation [7,17], where amphibian oocytes are far larger than mammalian oocytes and early embryos (0.075-0.2 mm diam.) where cryopreservation is common [17], and zebrafish embryos from 0.8 mm diam. where cryopreservation is advanced [218, Figure 7]. The most promising method for the perpetuation of the amphibian diploid genome is through cloning [4,18, see Table /, ibid], although ovarian stem cells could provide for the female diploid genome [17].
Cloning enables the restoration of the cloned species from the oocytes of donor species [17]. Amphibian cloning [somatic cell nuclear transfer [SCNT]] began as a laboratory technique using early gastrula nuclei for homocytoplasmic cloning in 1952 [220] and finally produced adult frogs in 1962 [221]. Heterocytoplasmic cloning is a critical RBC for the perpetuation of species solely in Biobanks. Surprisingly, recent reviews of RBCs, in 2022 and including cloning [17], have not included heterocytoplasmic cloning, which after 1961 has produced tadpoles, adults, and F1 offspring of anurans between species and subspecies in the same genus (for recent taxonomy see 234) , and salamanders between species since 1971 (Table 7, ibid]. Using the method derived from Nikitina (219), Kaurova et al. [18] found that homotransplantation of nuclei of cryopreserved Bufo bufo early embryonic cells only developed to the gastrula stage [for previously unpublished in English technique details sees section Appendix A.5. Cloning].
Overall, these studies (Table 7. ibid) show that: 1) species that produce inviable offspring are not suitable for cloning, 2) that close congeners are the most preferable for heterocytoplasmic cloning, 3) further research is needed to establish the optimum recipient species focusing on donor candidate species for RCBs and for cloning using cell lines, 4) that phylogeny should be included in RBC management plans to specify potential recipients for heterocytoplasmic cloning, 5) that early embryonic cells are most favourable for cloning, 6) that a complex suite of genotypic and phenotypic expression including reproductive potential can occur in clones and their offspring.
Table 7.
Heterocytoplasmic cloning in amphibians 1957-1962. * Viable hybrids. # viable hybrids with sterile males.
Table 7.
Heterocytoplasmic cloning in amphibians 1957-1962. * Viable hybrids. # viable hybrids with sterile males.
| Nucleus donors | Recipients | Results | Authors |
|---|---|---|---|
| Rana pipiens | R. catesbeina | Late blastula/early neurula | [220] |
| #R. brevipoda | R. nigromaculata | Died metamorphisis | [221] |
| R. pipiens | R. sylvatica | Late blastula/early neurula | [222] |
| R. nigromaculata | R. brevipoda | Adults | [223] |
| Xonopus llaevis | X. laevis | Adults | [224] |
| R. pipiens | R. palustris | Post-neurula | [225] |
| R. nigromaculata | R. brevipoda | Adults but poor reproduction | [226] |
| R. japonica | R. ornativentris | Adult frogs then F2 with female R. japonica | [227] |
| R. japonica | R. ornativentris | Adult frogs then F2 with female R. japonica | [228] |
|
R. nigromaculata R. temporaria |
R. brevipoda R. japonica |
F3. Reproductive capacity. % abnormality dependent on sex crosses. | [229] |
|
R. japonica R. temporaria |
R. temporaria R. japonica |
Adult frogs | [230] |
|
R. brevipoda R. plancyi R. brevipoda R. esculenta |
R. plancyi R. brevipoda R. esculenta R. brevipoda |
Adult frogs | [231] |
|
P. waltlii P. poireti |
P. poireti P. waltlii |
Adult salamanders | [232,234] |
7. Implementing RBCs – Economic, Political, Cultural Factors
A key component of amphibian RBCs is the provision of Biobanks to benefit amphibians and other threatened biodiversity [11]. RBCs as long-term strategies require: 1) personal, social, cultural, and political inclusion; 2) integrated partnerships between all facets of proactive amphibian sustainability; 3) infrastructure with minimum cost and maximum outcomes; and 4) financial, cultural, and political support [15,146]. A key ethical and practical issue with Biobanking is the ownership of Biobanked material, not only for entitlement and possession but how these relate to monetisation and financing (section 6.2, 6.5).
7.1. Monetisation of Proactive Amphibian Sustainability
Unfortunately, the monetisation status of RBCs has received minimal attention outside the RBC community regarding their potential for species perpetuation. The 2007 Amphibian Conservation Action Plan [116, Table 1.) was produced by 78 contributors; 69 from the collective West (USA 53, UK 7, other Western countries 9, developing countries 9). The plan directed only 1% toward reproduction technologies or biobanking and 11% toward CBPs or repopulation programs [107,108].
Table 8.
Proposed funding distribution in the 2007 Amphibian Conservation Action Plan [116). Budget costs in US$ millions. %GT = percentage of grand total.
Table 8.
Proposed funding distribution in the 2007 Amphibian Conservation Action Plan [116). Budget costs in US$ millions. %GT = percentage of grand total.
| Activity | Budget | % GT | Notes. |
|---|---|---|---|
| Distribution/Systematics | 407.2 | 67.7 | Biogeography 185.2 [26.8%], Biodiversity hotspots 176.4 (29.0%), Systematics 47.3 (7.9%). |
| Generic Threats | 118.0 | 19.6 | Pollution 63.8. (10.6%), Pathogens 37.3 (6.2%), Climate catastrophe 10.7 (1.8%), Over-harvest 6.3. (1.0%). |
| Conservation status | 2.7 | 4.5 | |
| Total | 527.9 | 87.7 | |
| RBCs | |||
| Reproduction Technologies and Biobanking | 6.3 | 1.1 | Bioresource Banking 5.1 (0.9%), Genome Resource Banking 1.0 (0.2%). |
| Captive programs | 60.2 | 10.0 | 100 facilities/species over 5 years |
| Repopulation/augmentation | 5.1 | 0.9 | |
| Total | 72.0 | 12.0 | |
| Grand Total (GT) | 601.7 |
The development of reproduction technologies and Biobanking was only granted 1.1% of the total, [241,242]. Ch 11. Bioresource Banking of somatic cells [241] directed finance toward San Diego Zoo Wildlife Alliance's Frozen Zoo [11, section 6.3.], with the establishment of 50 satellite facilities and cell culturing capabilities. Ch 7. Appendix A.1. Genome Resource Banking supported the development of sperm biobanking technologies by initiating research in North American zoos, and then mentoring trainees from developing countries (as range countries) to develop RBC programs. [242]. In 2007 amphibian sperm cryopreservation with fertilisation was only achieved in Australia [16] and Russia [12]. Surprisingly neither Ch. 11 nor 7 addressed the need for aARTs to restore individuals from diploid genomes. All the RBC technical goals listed in Appendix A.1. have been achieved in Australia, Russia, or the USA, by zoo or university-based programs. Unachieved objectives include Goal 8. the implementation and maintenance of an inventory and database for the management of cryopreserved samples [section 6.4], and Goal 9. increasing scientific capacity in developing countries.
Most generic threats to be funded by the 2007 ACAP are now highly prioritised in global policy, or well researched by academic programs, with the issues of over-harvest and pathogens [242,243,244,245,246] as ubiquitous threats largely to be addressed through national and international trade regulation (sections 2.2,2.4). However, in 2023 participants of the ninth World Congress of Herpetology and survey respondents produced a similar list of popular academic research priorities to the 2007 ACAP, including the climate crisis, pathogens, biodiversity, populations, and habitat, within a framework of polarised boundaries between anthropogenic activities and nature (section 1). Management was then included with Proactive Amphibian Sustainability through RBCs receiving a short mention. Fortunately, an increasing emphasis on evidence-based research in socioecological conservation was recognised [247]. However, a significant survey-based research paper with concerning community involvement and support of amphibian RBCs was not referenced [15, section 2.3,3.4]. In any case, implementing RBCs demands a much higher focus by the sustainability community than academic projects that overlap with globally prioritised programs addressing threats to sustainability. Conservation status with respect to RBCs has not been addressed by the IUCN (section 3.1.1). Nevertheless, the IUCN has multiple working groups for RBCs, and is in the process of publishing a transparent, inclusive, and un-monetised new ACAP.
7.2. Contemporary Cost Estimates of Amphibian RBCs
Costs and resources for RTs, Biobanking and CBPs will be country, facility, and economy specific. The use of cryopreserved sperm results in an ~20-fold reduction in costs for an institutional long-term CBP in a high-income Western industrialised economy [13,14]. Furthermore, the costs of private carers CBPs are estimated to be less than US$5,000 per species per year covered by private carers, with genetic diversity secured in Biobanks as the only cost. The cost of the biobanking component of RBCs is relatively minor. Basic biobanking facilities costs are estimated to range from approximately US$9,800 [13,14] to US$15,400 [8]. In the UK, liquid nitrogen costs US$1.50 a Liter and uses about 6L per tank. Tanks can typically hold 6000 samples. Biobanking costs are ~US$120- a sample [249]. The estimated cost to establish amphibian cell lines is US$15,000 (Matt Guille, pers. comm). Processing tissue for 10 vials including technical time, washing steps, and reagents costs ~US$90 [249].
7.3. Entitlement, Biopiracy, and Ownership Models
Global agreements provide responsible and sustainable biobanking practices that balance conservation interests, equitable access, benefit-sharing, research, and innovation. The United Nations Convention of Biodiversity [250], COP 15 [1], and the Nagoya Protocol for Access and Benefit-sharing [25] are international agreements for the equitable sharing of benefits from the utilization of genetic resources and safeguarding indigenous people’s traditional rights and stewardship of nature. Other ownership models include ‘Common Heritage’ where no entity can claim exclusive ownership and encourage Biobanking for the greater good of humanity, and ‘Private Ownership’ where all rights over Biobanked material and associated traditional knowledge are owned by investors.
7.4. Biobanking Facilities
Biobanking includes three categories, two held near or in CBPs ‘Research Banks’ for everyday use and ‘Active Management Banks’ for assisted gene flow. A third category, higher security ‘Perpetuity Banks’ holds samples mainly for species restoration [101]. Biobanking requires the establishment of storage facilities with ongoing financial support [8,188], with samples collected, processed, stored, and distributed, within conventional guidelines and standards [187,251], with the International Society for Biological and Environmental Repositories [ISBER] promoting effective management [252}. Perpetuity Biobanks now have few amphibian samples; however, the capacity is there.
The San Diego Zoo Wildlife Alliance's Frozen Zoo®, is one of the world’s largest and contains 10,000 samples, with 100 Endangered or Critically Endangered Mammalia, 62 Reptilia/Aves, and 85 Extinct Mammalia [253]. However, amphibians are poorly represented with 24 amphibian species in total with most of Least Concern, with only 11 threatened and 15 species with one sample [11, Table 9.].
The Chinese Academy of Science, Kunming Cell Bank, was established in 1986 and has now preserved 1455 cell lines from 298 animal species [4 insects, 24 fishes, 17 amphibians and reptiles, 25 birds, 228 mammals], along with 1200 tissue samples from over 200 animal species [254]. Nature’s SAFE was established in the UK in 2020 [249]. Working closely with Chester Zoo and the European Association of Zoos and Aquaria [EAZA] the EAZA Biobank is one of Europe’s largest living Biobanks dedicated toward saving highly endangered species from extinction by collecting, storing, and regenerating reproductive cells and cell lines. The EAZA Biobank collects and stores samples from 400 Member from zoos and aquariums located in 48 countries. Four dedicated institutions provide qualified staff and funds for infrastructure and equipment for long-term, high-quality storage facilities [Hubs] at Copenhagen, Antwerp, and Edinburgh Zoo, and in Berlin the Leibniz Institute for Zoo and Wildlife Research, that provide sample processing, facilitate sample transfer, and serve as sample backup facilities [255]. The Biodiversity Biobanks South Africa provide co-ordination across existing South African biodiversity Biobanks, aim to increase the range and quality of stored samples of South Africa’s exceptionally high levels of species diversity and endemism, and already houses a wealth of Biobank samples collected over more than 20 years [256].
7.5. Biobanking Databases
“A Biobank is only as good as the quality and extent of its recorded information and database that accompanies it” Rhiannon Lloyd, 2023.
Baseline information is essential for Biobanking to design an efficient global system for targeting sample collection across ecosystems and taxa. Data associated with biological samples are also vital for applications and research (257,258; section 5.2.2.). Collaborative data sharing and the standardisation in methodologies are critical, especially in ensuring that samples are correctly collected, processed, stored, and recorded. However, there is a lack of collated information between institutions’ Biobanked collections, and many attempts to obtain data on amphibians for this review were ignored, with this similarly being generally reported by those trying to advance this need. Groups including CryoArks [259] are working toward a global database. The IUCN Species Survival Commission, Animal Biobanking for Conservation Specialist Group, as reported in news services is working toward the collection, banking, sharing, and cataloguing of genetic resources, and has convened a series of virtual and in-person meetings. The group has a signature website with no details except the group leader, and has not yet presented a democratic, transparent, and accountable working structure with no meeting reports publicly available [260].
7.6. Financing Amphibian RBCs
“Finance for biodiversity and alignment of financial flows with nature to drive finances toward sustainable investments.” COP 15
The global economy will face annual losses of $2.7 trillion by 2030 if sustainability ecological tipping points are reached [261] and advancing RBCs should be a global financial priority for Endangered and Critically Endangered species. This priority particularly applies to amphibian as the most threatened terrestrial vertebrate taxon, with proven amenability to RBCS, and a substantive community support and ecosociological potential [244]. The loss of biodiversity is primarily driven by overconsumption in wealthier nations rooted in historical injustices and power imbalances. Consequently, there is growing global financial support for [262].
COP 15, Target 4, supports RBCs to assist gene flow to restore species genetic diversity and adaptive potential through the Kunming-Montreal Global Biodiversity Framework fund that offers financing toward developing countries [263). Implementation of the Nagoya Protocol incudes the Global Environment Facility [GEF] to support in-country research institutions, publicity and education, and project development [25]. There is a growing impetus to trade debt for environmental sustainability; RBC programs with assisted evolution to ameliorate coral bleaching [264]. Private sponsorship for Amphibian RBCs and Biobanks could also include many private philanthropic sources [266]. Furthermore, at a micro-economic level, local engagement with RBCs includes wealth creation through native entitlement, ecotourism, and industrial potentials including responsible trade and sale [15].
Even though not generically Perpetuity Biobanks, support and financing should be provided for nascent amphibian biobanking programs, that are proxies for Perpetuity Banks. For example, the USA National Amphibian Genome Bank [146], the Taronga Conservation Society Australia [248], Panamanian and other regional RBC facilities until integration with, or as, Perpetuity Banks can be formally established (section 6.4).
7.7. Media, Public and Political Presence
Branding of an industrial activity such as global Biobanking requires a marketing approach to terminology and lexicon including the avoidance of jargon [267]. Demographics of target audiences must be established, and effective marketing conducted, brand fidelity developed and fostered. Simple images and emotions with social and cultural context are more effective than statistics and abstract concepts in engaging support. Solution-orientated messaging offers personal well-being by reducing anxiety by offering direct opportunities and spurring helpful activities [268]. Deextinction provides a salient model for public engagement of species perpetuation through biobanking. Deextinction projects conflate species conservation and animal welfare, within a global cultural and ethical framework of Biospheric Sustainability, Intergenerational Justice, and Effective Altruism. For example, the vision of ‘Colossal Biosciences’ in recovering both keystone and iconic species has inspired institutional and private investors globally to provide hundreds of millions. Their vision is ‘healing Earth and its inhabitants, righting the wrongs of habitat loss, overhunting, poaching and the unnatural mistakes of humanity. Together, humanity and these not-forgotten species will wake up to a better tomorrow and enjoy another day in the sun’ [266].
7.8. Artificial Intelligence, Management, and the Future
For the benefit of RBCs, artificial intelligence can: 1) provide an overview species assessments and management recommendations, 2) strategise, coordinate, and synchronise organisational activities, 3) create written and visual lexicon for public presence, 4) garner support and fundraising, 5) empower and encourage participants, 6) standardise and collate disparate databases, and 7) optimise experimental design and publication [269,270]. Furthermore, artificial intelligence can seamlessly integrate our biological and cultural experiences into a comprehensive knowledge matrix to unlock the full potential of human consciousness [271]. Biobanks and genomic restoration databases can play a crucial role in this matrix by archiving biomaterials or information that can define cells, tissues, or individuals, and their behaviour [272]. An exciting potential for RBCs is their contribution to space colonisation and terraforming, by enabling the efficient and reliable transportation and restoration of viable biological materials [16,39,40]. The utilisation of RBCs in these environments will reciprocally provide knowledge contributing to Biospheric Sustainability on Earth.
7.9. Developing Cultural Influence
The future of amphibian sustainability depends on long-term social, cultural, and political support. This support relies on effectively merging our traditional, conventional, and emerging narratives and myths about humanity's place in the cosmos [247, section 1.). Biocentrism presents Earths biodiversity including humanity as unique [273], metamodernism avoids the fractures of the past and presents a holistic discourse that is both unifying and individualistically empowering [274], and metanature is an encompassing canvas on which to expands humanity’s potentials within a sustainable framework (section 1). When combined, these offer a broad philosophical and practical foundation for propelling humanity toward a sustainable future.
This theme manifests where traditional ecological knowledge as practiced by Indigenous people for millennia [275], or by amphibian private carers [15], conflates with RBCs to create functional fact-based narratives to support amphibian sustainability. These biocultural approaches, through the local community’s entitlements and cultural values, extend to other geopolitical entities to support social–ecological systems over the long term [276]. For instance, these benefits include research on culturally important species by indigenous peoples and ethnic groups in biodiverse regions that typically have disproportionately higher numbers of Data-Deficient species, many of which are now finally recognised as Endangered [277].
9. Conclusion and the Road Ahead
It is difficult to predict if or when biodiversity extinctions will plateau, but many more than the current 400 amphibian species will require RBCs in the immediate future. Through this review we have discussed the potential of Reproduction Technologies, Biobanking, and Conservation Breeding Programs (RBCs), to contribute to Biospheric Sustainability in preventing a potentially catastrophic loss of biodiversity as existential threats exponentially collapse global ecosystems. Heritage Biobanks must be placed as national treasures to manage genomic resources and preserve genetic diversity with long-term security as a core component of this incentive.
In tandem with global incentives, we have demonstrated that current amphibian RBCs offer proven, cost-effective, and reliable means to perpetuate amphibian species in need. With further technological development species can be perpetuated solely in biobanks. Amphibian RBCs also offer an opportunity to closely engage the global community from indigenous custodians, through private carers and CBPs, to major global initiatives for Biospheric sustainability. Unfortunately, this opportunity has not been fulfilled, and the few extant amphibian RBCs programs are largely incentives in wealthy Western countries. Outside these well-funded programs, funding toward developing counties is non-existent or gestural such as seed funding.
This review has shown that there is no contemporary independent, democratic, transparent, accountable, and inclusive umbrella organisation fully supporting the practical needs of RBCs for amphibians. Other potentials include extending RBCs beyond bio-technocrats to contributions by expertise in the critical field’s media, public relations, information technology, and artificial intelligence. An inclusive non-governmental organisation could fulfil global RBC goals through broadly partnering with the sustainability community, and especially a wider range of biopolitical and geopolitical entities, including stakeholders such as social groups and local communities.
A respected and powerful voice for the global presence of biobanking, RTs, and CBPs, in the multi-billion Biospheric Sustainability initiatives would include a voice toward financial support and official mandates. These resources provide solid career streams within a long-term global project framework and stimulating incentives that will attract a wide range of contributors to build projects by competing for influence and resources. This focus on amphibians, the most needy and amenable vertebrate taxon to RBCs, would offer a model to advance RBCs for other taxa.
Ethics
Approval for experiment in Appendix A.4. Artificial Fertilisation.
Acknowledgments
In this section, you can acknowledge any support given which is not covered by the author contribution or funding sections. This may include administrative and technical support, or donations in kind (e.g., materials used for experiments).
Conflicts of Interest
The authors declare no conflict of interest.” Authors must identify and declare any personal circumstances or interest that may be perceived as inappropriately influencing the representation or interpretation of reported research results.
Appendix A
Appendix A.1. The Anthropocene and Biospheric Sustainability
The Anthropocene epoch was defined by Crutzen and Stoermer [278] as human activities dominating biospheric ecology as described in Ellis and Ramankutty [279]. However, conventionally the Anthropocene began in the mid-20th century with the ‘Great Acceleration’ of industrial, technical, and scientific development, along with increasing consumerism, resource extraction, and massive pollution [280]. The beginning of the Anthropocene is more difficult to define from a biological perspective. For instance, European expansionism and colonialisation, punctuated in 1492 by the discovery of the New World [281,282], and of Australia in 1788 [283], promoted massive ecological modification in these three continents. However, the earliest major anthropogenic environmental influence was the megafaunal extinctions by paleolithic hunter-gatherers through predation and fires [284,285]. As noted by Wallace [1876], "We live in a zoologically impoverished world, from which all the hugest, and fiercest, and strangest forms have recently disappeared."
The crisis of Biospheric Sustainability is now a pressing existential issue for humanity and is reaching a catastrophic and irreversible stage for many major ecosystems [1,46]. To address this generic crisis, Biospheric Sustainability, the greatest biopolitical and economic transformation in history is being implemented [1]. UNEP COP 15 [1] prioritised five Anthropogenic contributors to biodiversity loss: 1) alteration of environments, 2) climate catastrophe, 3) pollution, 4) overconsumption, and 5) invasive species. Irrespective of these initiatives, Amphibian Biodiversity faces an immediate future of increasing rates of decline and extinction from these and other threats [286, section Part 2].
The burgeoning population along with fossil fuel use has resulted in the highest atmospheric CO2 levels for 3 million years with climate catastrophe predictions of droughts, fires, floods, human misery, and biodiversity loss [46]. Synergism between multiple climate and non-climate threats will cause massive species extinctions [2,35,47,287,288], with the climate catastrophe alone risking the extinction of 15% of species at the inevitable 1.5°C, 20% at the likely 2.0°C, and 40% at the disastrous 3.0°C [34]. Consequently, the amphibian conservation community must cooperate and collaborate to garner all available political, economic, and human resources, to provide Proactive Amphibian Sustainability including the timely development and application of RBCs.
Biospheric Sustainability requires advanced civilisations [1] that utilise resources to ensure equitable social, cultural, and economic well-being through Intergenerational Justice [289,290]. From the perspective of Western colonialism, civilizations are defined by a centralized state supported by a theocracy and social hierarchy, sophisticated transport, communication, and militarisation, and a morality encouraging hegemonic structures for dominance and exploitation [282]. These unsustainable civilisations eventually collapse through the centralisation of resources to elites, over-exploitation of resources, lowered individual opportunity and cultural capital, inefficient economies, cultural decay, and social unrest [291]. Biospheric Sustainability has been coined as a universal concept to avoid the collapse of our now global civilisation, albeit within a framework of respect for global cultures, theocracies, and political systems [55]. When applied these principles lead to cohesive, inclusive, and rewarding social structures, and resilient communities implementing prioritising resource use for the long-term well-being of future generations; an exemplar is Costa-Rica the “Living Eden” with 5% of amphibian biodiversity in 0.03% of Earth’s terrestrial area [292].
The loss of biodiversity through resource exploitation and pollution is primarily driven by overconsumption in wealthier nations and is rooted in historical injustices and power imbalances [262], exacerbated by colonialist and Darwinian cultural and racial bigotries [293]. The foundational characteristics of unbridled capitalist economies are antithetical to Biospheric Sustainability, where the means of production, under increasing plutocratic control, plugs profits into a cycle of resource extraction and labour, in an expanding sphere of environmental and cultural destruction [294].
Increasing consumerism in a growing human population, projected to reach 11 billion by 2100 is particularly challenging [295]. Regulated consumer marketing based on need rather than want, along with responsible social media in regulated culturally sensitive economies, can foster social inclusivity, knowledge-based decision-making, and effective political and environmental action [1]. Encouraging such practices can counter social isolation and empower individuals to be active participants in creating positive change. These initiatives can transform humanity towards a balanced and interconnected system that harmonizes economies, social interactions, and cultural values with nature. Embracing the principles of the Cochabamba People's Agreement [296] can lead to greater respect for life and a sustainable coexistence with our environment.
Positive environmental goals can be effectively achieved through the adoption of negative GDP growth models in overconsuming countries, coupled with sustainable GDP growth in developing countries [294,297]. The primary focus is on minimizing poverty and enhancing human well-being, happiness, and sustainability [294]. By implementing these cultural, political, and economic standards in a multi-polar world, we can strive towards both Biospheric Sustainability and Intergenerational Justice. By working together towards this transformative vision, we can create a more environmentally conscious and socially responsible society. While the path to Biospheric Sustainability faces a broad range of challenges [1], remaining optimistic and proactive by taking decisive actions and adopting sustainable practices, we can address this malaise and create a brighter future for generations to come [298,299].
Community-based conservation projects [300] offer a unique avenue for proactive amphibian sustainability that bridges regional and global communities. However, it's crucial to recognize that communities burdened by low incomes have limited participation in environmental sustainability initiatives. Therefore, proactive amphibian sustainability including RBCs should align with poverty-reduction programs that empower traditional stakeholders. In line with this perspective, de Lange et al. [300] advocate for inclusive and just transformations toward biospheric sustainability, supported by a conservation basic income for residents in critical conservation areas. This investment pales in comparison to the estimated US$44 trillion global economic production reliant on biospheric sustainability [301].
Entitlement of traditional owners is often essential for environmental protection [302,303], especially in the unique and specialised ecosystems supporting much amphibian biodiversity [292,304]. Many situations that displace traditional owners for habitat protection often have neo-colonial undertones that must be recognised and addressed [305,306]. Rooted in historical colonization, displacements are often driven by external economic and political interests. Foreign company’s interests prioritize resource extraction, without national investment toward development, leaving neo-colonized nations dependent on primary commodity exports [1]. Addressing these problems requires bolstering transparency and social responsibility in governance, to foster sustainable economic development, while acknowledging historical power imbalances and creating equitable trade relationships [1]. This concept extends to amphibian community-based projects where reciprocity between institutional and private contributors from affluent countries will produce optimal outcomes [15,307].
Appendix A.2. Oocytes Viability Period and Storage Environment
However, the selection process behind the immediate loss of oocyte fertility is uncertain, and therefore any genetic bias on progeny or confounding of experimental results. The generalised conventions of oocyte storage in ionic formulations as applied to amphibian conservation research [169,208] are based on standardised techniques for laboratory practice [308,309,310], where isotonic ionic formulations slow egg gel hydration, support sperm penetration toward the oocyte, with lowering to 2.5% of ionicity at fertilisation optimal for development [202]. However, the fertility of some anuran oocytes in water is maintained for up to an hour or more [311], and female sperm choice for greater progeny heterozygosity provides oocytes in spawns that resist initial fertilisation during spawning, leaving unfertilized but fertilizable oocytes with hydrated gel for later males [164]. Dry storage of cool-temperate anuran oocytes at 4°C in Eppendorf tubes, to prevent dehydration and to maintain ionic stasis, over five days maintained 70% fertility and 40% hatch rates, with some fertility and development extended to eight days [312]. Gagarinskiy et al [313] showed that a 6.5 atm gaseous mixture of carbon monoxide and oxygen doubled the refrigerated storage viability of the unfertilised oocytes of a cool temperate anuran, compared with controls when fertilised over eight days. However, from four to eight days of storage the hatch rate declined to a low level like that of controls. Therefore, the causes of oocyte loss of viability are ions or proteins diffusing from the gel that are needed for sperm motility or oocyte metabolism [314,315,316], or damage to the cell envelope [313].
Appendix A.3. Sperm Cryopreservation
Ever since the first successes at the turn of the century, then success with all trialled species, a proliferation of research with various species has been conducted toward optimal cryopreservation techniques. Browne et al. [16] through a literature review and canvassing the research community recommended a 5-10% [v/v] concentration of dimethyl formamide [DMFA] and 1-10% [w/v] sucrose with cryopreservation achieved through moderate cooling rates. Poo and Hinkson [173] found 5% DMFA with 10% trehalose with moderate cooling rates found similar recovery rates in four anurans from two families. Earlier studies showed some success with glycerol and saccharides at high concentrations [180,317]. However, Naranjo et al. [203] found high toxicity of 10% glycerol and favoured only 5% DMFA with low 2.5% sucrose concentrations, similarly very low saccharide concentrations of 1% were found most effective with glycerol ineffective when compared to DMS0 [204,205].
Other recent studies have surprisingly achieved higher fertilisation rates in cryopreserved sperm than expected through fresh sperm using relatively fast cooling rates [103; Appendix 4]. Moderate cooling rates over liquid nitrogen [16] can be achieved with simple Styrofoam apparatus [203,318]. The cooling rate depends on the height of the straws above the liquid nitrogen, the material of the apparatus and box, and how long the LN2 is in the container before sperm freezing. Controlled rate freezers enable reliable staged freezing [180,204,205,206]. Following Shishova et al [181] and Kaurova et al. [178] a recent study showed that very high 15% DMSO concentrations with1% saccharides, or high 12% DMFA with 10% saccharides, were equally successful with moderate cooling through slow immersion into LN2 vapour much superior to slower staged cooling (319). Success with very fast cooling through vitrification by direct immersion in liquid nitrogen has not been published for amphibian sperm (201).
Appendix A.4. Artificial Fertilisation
Dry fertilisation for amphibians mixes oocytes with testicular macerates in isotonic solutions where unactivated sperm are then left for ~10 min. before flooding with water both activates and dilutes sperm concentrations ~40:1 [16,208,320]. Activated suspensions may also be mixed with oocytes and then left for a fertilisation period from 15-30 min. before washing with water to eliminate harmful cryoprotectants [103,181]. However, the use of dry fertilisation was historically misrepresented in Silla and Kouba [321] through miss-referencing early studies in the USA that placed fresh active hormonally induced sperm on oocytes, rather than earlier studies of dry fertilisation of fishes in Russia [322].
To define the period limitations of the ‘dry Fertilisation’ technique, Rana temporaria sperm were mixed oocytes with immotile testicular sperm suspensions at 230-240 mosmol kg-1 and held in a 3 cm diam. Petri dish with wet filter paper to prevent oocyte dehydration. Control oocytes fertilised with the immediate addition of water showed good fertility. After 15, 30, 60, and 240 min. no oocyte cleavage had occurred in any of the other dishes, or after water was added to promote fertilisation. Presumably, drying of the sperm suspension resulted in final sperm infertility as R. temporaria oocytes are capable of remaining fertile for many hours without the addition of water [pers comm].
Sperm fertility is normally expressed as the percentage of oocytes fertilised in comparison to a ‘saturation’ concentration of fresh sperm. However, fertilisation curves of sperm concentrations over the fertilisation period have only been presented for a few species, and concentrations are often expressed as those from sperm suspensions irrespective of fertilisation technique and period.
Figure A1.
Rhinella (Bufo) arenarum [Bufonidae] fresh testicular sperm concentrations, the percentage in vitro fertilisation, and minutes post-application before washing with fresh water [333].
Figure A1.
Rhinella (Bufo) arenarum [Bufonidae] fresh testicular sperm concentrations, the percentage in vitro fertilisation, and minutes post-application before washing with fresh water [333].
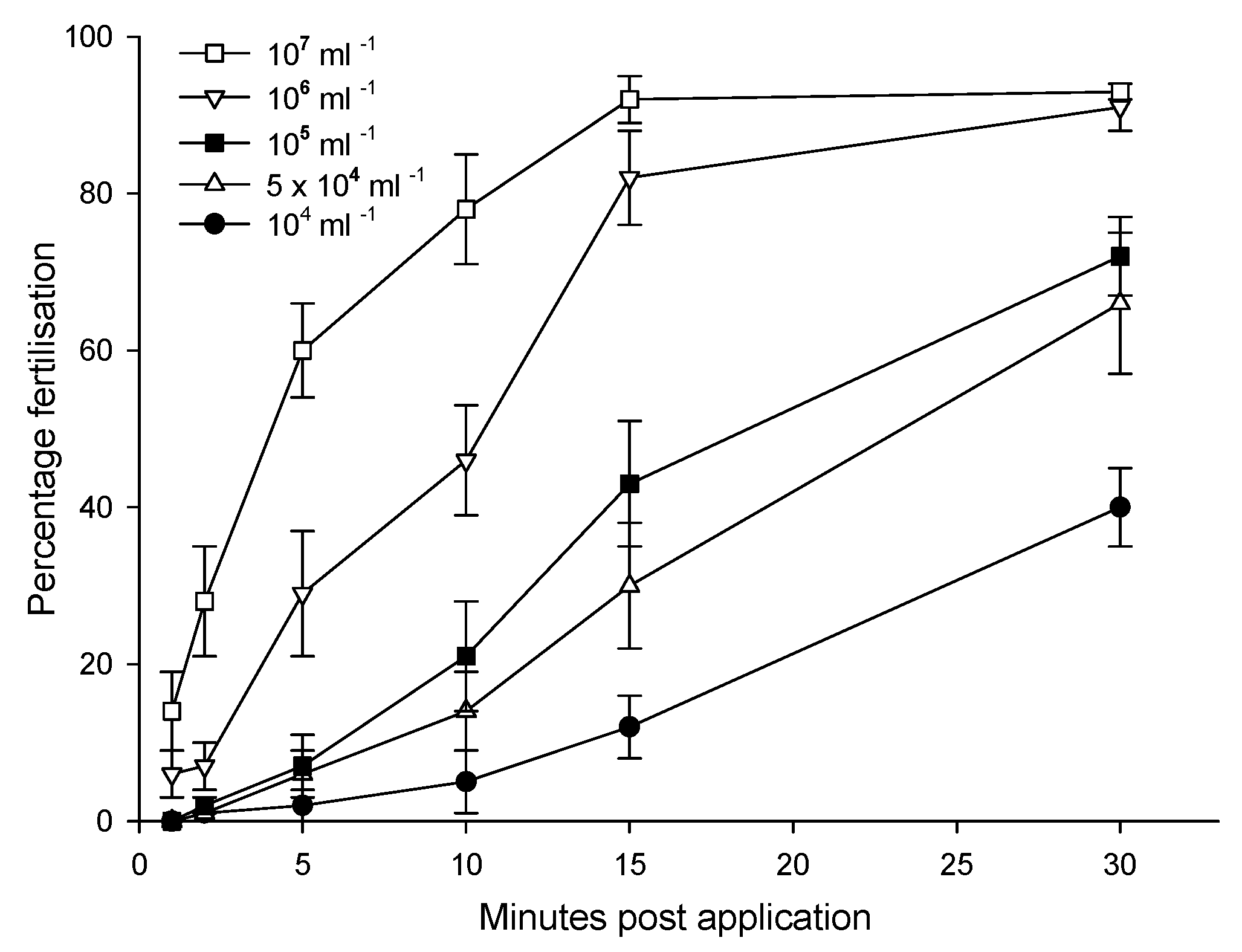
We found publications of sperm concentrations and fertilisation rates, with activated testicular suspensions of Bufo arenarum [333], Limnodynastes tasmaniensis [311], and Pseudophryne guentheri [334]; with dry fertilisation with Rhinella [Bufo] marinus [180], and between activated testicular suspensions and sperm in spermic urine of R.temporaria [181].
Cabada [333] showed saturated fertilisation after a 10 min. period of fresh activated Rhinella (Bufo) arenarum testicular sperm to oocytes at concentrations of ~107 ml-1 sperm and Holinger and Corton [334] with 105-7 ml-1 Xenopus laevis, and Browne et al. [180] with ~106 ml-1 Rh. marinus using ‘dry fertilisation’. The saturated fertilisation found with ~104 ml-1 of freshly activated L. tasmaniensis testicular sperm was after 30 min. [Figure 3., 311], and Silla with P. guentheri at 2.5 x 105 ml-1 after 60 min. [335] corresponded with those predicted in Cabada [333]
In one comparative fertilisation curve study spermic urine was more fertile than testicular sperm. Shishova et al. [181] with R. temporaria found saturated fertilisation with testicular sperm only reached 60% and then declined after a concentration of ~108 ml-1, whereas spermic urine only required one magnitude less of ~107 ml-1 to saturate. The fertilisation rate also declined far more slowly with spermic urine where after 15 min. a concentration of ~106 ml-1 provided ~45% fertilization while testicular sperm was ~5%. Fertilisation rates with Anaxyrus (Bufo) fowleri fresh spermic urine at a concentration of ~4.9 × 106 ml-1 sperm after 5 min. was ~85%, and after cryopreservation and freeze/thaw dilution at 2.4 x 105 ml-1 were ~20% [estimated from Figure 3, Burger et al. 2022]. These fertilisation rates are very exceptional at ~3x than those expected using fresh spermic urine [Burger et al. 2022] or fresh testicular sperm [333].
Appendix A.5. Cloning
Totipotent cells were obtained by dissociating early embryos of the common toad Bufo bufo at the early blastula stage in Niu-Twitty's medium without calcium. The use of a cryoprotective solution based on the Niu-Twitty medium with the addition of 0.5 - 1% BSA, 10% DMSO, and 10% sucrose for freezing totipotent toad cells made it possible to obtain the maximum number of intact cells. The preservation of the nuclei of cryopreserved cells was judged by the results of homotransplantation by injection into unfertilized oocytes whose own nuclei were inactivated by X-ray irradiation. The results of the visual assessment of the cell (in points) after cryopreservation in different media are presented Table 1. After the injection of nuclei of cryopreserved and control cells into enucleated eggs, the initial development of embryos up to the stage of late blastula was observed. Thus, the possibility of cryopreservation of amphibian totipotent germ cells with subsequent use of their nuclei for the reconstruction of eggs has been shown (Kaurova et al., 1998; Table 1).
Table A1.
The preservation of totipotent cells of the common toad Bufo bufo after cryopreservation in Niu-Twitty medium with different concentrations of DMSO and sucrose (visual assessment according to a 5-point System.
Table A1.
The preservation of totipotent cells of the common toad Bufo bufo after cryopreservation in Niu-Twitty medium with different concentrations of DMSO and sucrose (visual assessment according to a 5-point System.
| Assessment of cell preservation in points | ||||
|---|---|---|---|---|
| DMSO (%) | Sucrose (%) | |||
| 5 | 10 | 20 | 30 | |
| 5 | 3 | 3 - 4 | - | - |
| 10 | - | 5 | 3 - 4 | 3 |
| 20 | - | 4 | 4 | - |
Appendix A.6.
Table A2.
A list of milestone research achievements and practical applications of amphibian RBCs.
| Year | Milestone | Ref |
|---|---|---|
| 1986 | Release of hormonally stimulated reproduction of E salamander | 337 |
| 1989 | Hormonal stimulation of reproduction of many anurans and salamanders | 338 |
| 1996 | Cryopreservation of anuran sperm in pieces of testes with fertilisation | 178,179 |
| 1998 | Cryopreservation of totipotent cells and their use in reconstruction of enucleated eggs | 18 |
| 1998 | Cryopreservation of anuran sperm suspensions with fertilisation | 180 |
| 1998 | Saccharides and anuran sperm cryopreservation | 317 |
| Refrigerated storage of anuran sperm and oocytes | 208 | |
| 2003 | Cryopreservation of Cryptobranchidae (Caudata) sperm | 194 |
| 2006 | Novel use of high concentrations of LHRH-A for gamete collection in anurans | 169 |
| 2006 | Novel use of dopamine antagonists for gamete collection in anurans | 168 |
| 2010 | Hormonal stimulation and in vitro fertilisation in Salamandridae | 118 |
| 2011 | Cryopreservation of hormonally induced anuran sperm with fertilisation | 181 |
| 2011 | Cryopreservation of hormonally induced anuran sperm with dimethyl formamide | 181 |
| 2013 | Refrigerated storage of in situ (in carcasses) anuran sperm for 7 days | 339 |
| 2016 | Generation of a reproductively mature adult from cryopreserved Salamandridae sperm | 194 |
| 2018 | Refrigerated storage of anuran oocytes in the oviducts of live females | 312 |
| 2018 | Reproductively mature anurans from cryopreserved testicular sperm | 206 |
| 2019 | Cryopreservation of anuran sperm from nature and in vitro fertilisation | 125 |
| 2020 | Seasonality in hormonally stimulated sperm | 159 |
| 2021 | Reproductively mature CE anurans from cryopreserved testicular sperm | 205 |
| 2021 | Generation of a mature adult from cryopreserved Cryptobranchoidea sperm | 194 |
| 2021 | Cryopreservation of refrigerated sperm from carcasses | 339 |
| 2022 | Assisted gene flow between in situ and ex situ anurans | 102 |
| 2022 | Gentamicin increases refrigerated sperm storage | 214 |
| 2022 | Ultrasound optimisation of Salamandridae hormonal stimulation | 124 |
| 2023 | Low saccharide concentrations of 1-5% optimal for sperm cryopreservation | 203, 205 |
| 2023 | High pressure of carbon monoxide and oxygen for anuran oocyte refrigerated storage | 313 |
References
- UNEP. UN Biodiversity Conference (COP 15). December 7-19, 2022. Available online: https://www.unep.org/un-biodiversity-conference-cop-15 (accessed on 23 September 2023).
- Finn, C.; Gratarolla, F.; Pincheira-Donoso, D. More Losers than Winners: Investigating Anthropocene Defaunation through the Diversity of Population Trends. Biol. Rev. 2023, 98, 1732–1748. [Google Scholar] [CrossRef] [PubMed]
- AArk. Amphibian Ark. Available online: www.amphibianark.org/ (accessed on 27 September 2023).
- Clulow, J.; Mahony, M.; Browne, R.; Pomering, M.; Clark, A. Applications of Assisted Reproductive Technologies (ART) to Endangered Anuran Amphibians. In Declines and disappearances of Australian frogs, Campbell, A., Ed.; Environment Australia: Canberra, Australia, 1999; pp. 219–225. ISBN 0. [Google Scholar]
- Clulow, J.; Clulow, S. Cryopreservation and Other Assisted Reproductive Technologies for the Conservation of Threatened Amphibians and Reptiles: Bringing the ARTs up to Speed. Reprod. Fert. Dev. 2016, 28, 1116–1132. [Google Scholar] [CrossRef] [PubMed]
- Ananjeva, N.B.; Uteshev, V.K.; Orlov, N.L.; Ryabov, S.A.; Gakhova, E.N.; Kaurova, S.A. , Kramarova L.I., Shishova N.V., Browne R.K. Comparison of the Modern Reproductive Technologies for Amphibians and Reptiles. Russ. J. Herpetol. 2017, 24, 275–290. [Google Scholar] [CrossRef]
- Clulow, J.; Upton, R.; Trudeau, V.L.; Clulow, S. Amphibian Assisted Reproductive Technologies: Moving from Technology to Application. Adv. Exp. Med. Biol. 2019, 1200, 413–463. [Google Scholar] [CrossRef] [PubMed]
- Della-Tonga, G.; Howell, L.G.; Clulow, J.; Langhorne, C.J.; Marcec-Greaves, R.; Calatayud, N.E. Evaluating Amphibian Biobanking and Reproduction for Captive Breeding Programs According to the Amphibian Conservation Action Plan Objectives. Theriogenology 2020, 150, 412–431. [Google Scholar] [CrossRef] [PubMed]
- Strand, J.; Thomsen, H.; Jensen, J.B.; Marcussen, C.; Nicolajsen, T.B.; Skriver, M.B.; Søgaard, I.M.; Ezaz, T.; Purup, S.; Callesen, H.; et al. Biobanking in Amphibian and Reptilian Conservation and Management: Opportunities and Challenges. Conserv. Gen. Res. 2020, 12, 709–725. [Google Scholar] [CrossRef]
- Strand, J.; Fraser, B.; Houck, M.L.; Clulow, S. Culturing and Biobanking of Amphibian Cell Lines for Conservation Applications. In Reproductive Technologies and Biobanking for the Conservation of Amphibians, Silla, A.J., Kouba, A.J., Heatwole, H., Eds.; CRC Press: London, UK, 2022; ISBN 9781486313334. [Google Scholar]
- Mooney, A.; Ryder, O.A.; Houck, M.L.; Staerk, J.; Conde, D.A.; Buckley, Y.M. Maximizing the Potential for Living Cell Banks to Contribute to Global Conservation Priorities. Zoo Biol. 2023. [Google Scholar] [CrossRef] [PubMed]
- Uteshev, V.K.; Gakhova, E.N.; Kramarova, L.I.; Shishova, N.V.; Kaurova, S.A.; Kidova, E.A.; Kidov, A.A.; Browne, R.K. Russian Collaborative Development of Reproduction Technologies for the Sustainable Management of Amphibian Biodiversity. Asian Herpetol. Res. 2023, 14, 103–115. [Google Scholar] [CrossRef]
- Howell, L.G.; Mawson, P.R.; Frankham, R.; Rodger, J.C.; Upton, R.M.O.; Witt, R.W.; Calatayud, N.E.; Clulow, S.; Clulow, J. Integrating Biobanking could Produce Significant Cost Benefits and Minimise Inbreeding for Australian Amphibian Captive Breeding Program. Reprod. Fert. Dev. 2021, 33, 573–587. [Google Scholar] [CrossRef]
- Howell, L.G.; Frankham, R.; Rodger, J.C.; Witt, R.R.; Clulow, S.; Upton, R.M.O.; Clulow, J. Integrating Biobanking Minimises Inbreeding and Produces Significant Cost Benefits for a Threatened Frog Captive Breeding Programs. Conserv. Lett 2021, 14, 12776. [Google Scholar] [CrossRef]
- Browne, R.K.; Janzen, P.; Bagaturov, M.F.; van Houte, D.K. Amphibian Keeper Conservation Breeding Programs. J. Zoo. Res. 2018, 2, 29–46. [Google Scholar] [CrossRef]
- Browne, R.K.; Silla, A.J.; Upton, R.; Della-Togna, G.; Marcec-Greaves, R.; Shishova, N.V.; Uteshev, V.K.; Proano, B.; Perez, O.D.; Mansour, N.; et al. Sperm Collection and Storage for the Sustainable Management of Amphibian Biodiversity. Theriogenology 2019, 133, 187–200. [Google Scholar] [CrossRef] [PubMed]
- Clulow, J.; Upton, R.; Clulow, S. Cryopreservation of Amphibian Genomes: Targeting the Holy Grail, Cryopreservation of Maternal-haploid and Embryonic-diploid Genomes. In Reproductive Technologies and Biobanking for the Conservation of Amphibians, Silla, A.J., Kouba, A.J., Heatwole, H., Eds.; CRC Press; London, UK, 2022; pp. 147–165, ISBN 9781486313334.
- Kaurova, S.A.; Nikitina, L.A.; Uteshev, V.K.; Gakhova, E.N. Cryopreservation of Totipotent Embryo Cells and Their use in Reconstruction of Enucleated Eggs, In Proceedings of the 15th Working Meeting, Pushchino, Russian, 13–15, October 1998; pp. 206–208 (In Russian). (In Russian).
- Tapley, B.; Bradfield, B.; Michaels, C.; Bungard, M. Amphibians and Conservation Breeding Programmes: do all Threatened Amphibians Belong on the Ark? Biodivers. Conserv. 2015, 24, 2625–2645. [Google Scholar] [CrossRef]
- Wiedenfeld, D.A.; Alberts, A.C.; Angulo, A.; Bennett, E.L.; Byers, O.; Contreras-MacBeath, T.; Drummond, G.; da Fonseca, G.A.B.; Gascon, C.; Harrison, I. Conservation Resource Allocation, Small Population Resiliency, and the Fallacy of Conservation Triage. Conserv Biol 2021, 35, 1388–1395. [Google Scholar] [CrossRef]
- Runge, M.C. An Introduction to Adaptive Management for Threatened and Endangered Species. J. Fish Wildl. Manage. 2011, 2011, 2, 220–223. [Google Scholar] [CrossRef]
- Mc Cartney, A.M.; Head, M.A.; Tsosie, K.S.; Sterner, B.; Glass, J.R.; Paez, S.; Geary, J.; Hudson, M. Indigenous Peoples and Local Communities as Partners in the Sequencing of Global Eukaryotic Biodiversity. npj Biodiversity 2023, 2, 8:1–8:14. [Google Scholar] [CrossRef]
- GEF. The Global Environment Facility (GEF). Available online: https://www.thegef.org/ (accessed on 27 September 2023).
- Byers, O.; Lees, C.; Wilcken, J.; Schwitzer, C. The One Plan Approach: The Philosophy and Implementation of CBSG’s Approach to Integrated Species Conservation Planning. WAZA Mag. 2013, 14, 2–5. [Google Scholar]
- ABS. 2014. Available online: https://www.cbd.int/abs/ (accessed on 27 September 2023).
- Khoday, K. Decolonising the Environment: Third World Approaches to the Planetary Crisis. Indones. J. Int. Law, 2022, 19, 189–211. [Google Scholar] [CrossRef]
- Abiddin, N.Z.; Ibrahim, I.; Abdul Aziz, S.A. Non-Governmental Organisations (NGOs) and Their Part towards Sustainable Community Development. Sustainability 2022, 14, 4386:1–4386:13. [Google Scholar] [CrossRef]
- Adams, W.; Mulligan, M. Decolonising Nature: Strategies for Conservation in a Developing Era. 2002. Earthscan Publications Ltd; Sterling, VA, USA, ISBN 1-85383-749-0.
- Seddon, P.J.; Griffiths, C.J.; Soorae, P.S.; Armstrong, D.P. Reversing Defaunation: Restoring Species in a Changing World. Science 2014, 345, 406–412. [Google Scholar] [CrossRef]
- IUCN/SSC. 2014. Available online: www.iucn.org/about/work/programmes/species/publications/iucn_guidelines_and__policy__statements/ (accessed on 27 September 2023).
- Conde, D.A.; Colchero, F.; Gusset, M.; Pearce-Kelly, P.; Byers, O.; Flesness, N.; Browne, R.K.; Jones, O.R. Zoos through the Lens of the IUCN Red List: A Global Metapopulation Approach to Support Conservation Breeding Programs. PLoS One, 2013, 8, 80311:1–80311:9. [Google Scholar] [CrossRef] [PubMed]
- Woolly Mammoth De-extinction Project & Process | Colossal. Available online: https://colossal.com/mammoth/ (accessed on 27 September 2023).
- Bradshaw, C.J.A.; Ehrlich, P.R.; Beattie, A.; Ceballos, G.; Crist, E.; Diamond, J.; Dirzo, R.; Ehrlich, A.H.; Harte, J.; Harte, M.E.; Graham, P.; et al. Underestimating the Challenges of Avoiding a Ghastly Future. Front. Conserv. Sci. 2021, 1, e615419. [Google Scholar] [CrossRef]
- Hoegh-Guldberg, O.; Jacob, D.; Taylor, M.; Bindi, M.; Brown, S.; Camilloni, I.; Diedhiou, A.; Djalante, R.; Ebi, K.L.; Engelbrecht, F.; et al. Impacts of 1.5ºC Global Warming on Natural and Human Systems. In Global Warming of 1.5°C. An IPCC Special Report on the impacts of global warming of 1.5°C above pre-industrial levels and related global greenhouse gas emission pathways, in the context of strengthening the global response to the threat of climate change, sustainable development, and efforts to eradicate poverty; Cambridge University Press: Cambridge, UK, 2022; pp. 175–312. [Google Scholar] [CrossRef]
- Finn, C.; Gratarolla, F.; Pincheira-Donoso, D. More Losers than Winners: Investigating Anthropocene Defaunation through the Diversity of Population Trends. Biol. Rev. Camb. Philos. Soc. 2023, 98, 1732–1748. [Google Scholar] [CrossRef]
- Oxford Dictionary. ‘Nature, n’ Available online:. Available online: https://www.oed.com/search/dictionary/?scope=Entries&q=nature (accessed on 27 September 2023).
- Spencer, S. Judeo-Christian Religion's Impact on Humans' Attitudes Towards Their Environment, 2016, Swarthmore College Environmental Studies. Available online: http://fubini.swarthmore.edu/~ENVS2/sierra/Essay2.html (accessed on 27 September 2023).
- Lind, A.G.; Nobre, B. Ecology as a New Foundation for Natural Theology. Religions 2021, 12, 660:1–660:11. [Google Scholar] [CrossRef]
- Aglietti, G.S. Current Challenges and Opportunities for Space Technologies. Front. Space Technol. 2020, 1. [Google Scholar] [CrossRef]
- Johnson, A.R. Biodiversity Requirements for Self-Sustaining Space Colonies. Futures 2019, 110, 24–27. [Google Scholar] [CrossRef]
- Wakayama, S.; Ito, D.; Kamada, Y.; Shimazu, T.; Suzuki, T.; Nagamatsu, A.; Araki, R.; Ishikawa, T.; Kamimura, S.; Hirose, N.; et al. Evaluating the Long-term Effect of Space Radiation on the Reproductive Normality of Mammalian Sperm Preserved on the International Space Station. Sci. Adv. 2021, 7, eabg5554. [Google Scholar] [CrossRef]
- Valencia-Aguilar, A.; Cortés-Gómez, A.M.; Ruiz-Agudelo, C.A. Ecosystem Services Provided by Amphibians and Reptiles in Neotropical Ecosystems. Int. J. Biodivers. Sci. Ecosyst. Serv. Man. 2013, 9, 257–272. [Google Scholar] [CrossRef]
- Fisher, J.C., Dallimer, M., Irvine, K.N.; Aizlewood, S. G.; Austen, G. E.; Fish, R. D.; King, P. M.; Davies, Z. G. Human Well-being Responses to Species’ Traits. Nat. Sustain 2023. [CrossRef]
- Methorst, J.; Bonn, A.; Marselle, M.; Böhning-Gaese, K.; Rehdanz, K. Species Richness is Positively Related to Mental health – A Study for Germany. Landscape and Urban Planning 2021, 211, 104084:1–104084:11. [Google Scholar] [CrossRef]
- Greenwell, P.J.; Riley, L.M.; Lemos de Figueiredo, R.; Brereton, J.E.; Mooney, A.; Rose, P.E. The Societal Value of the Modern Zoo: A Commentary on How Zoos Can Positively Impact on Human Populations Locally and Globally. J. Zool. Bot. Gard. 2023, 4, 53–69. [Google Scholar] [CrossRef]
- Cowie, R.H.; Bouchet, P.; Fontaine, B. The Sixth Mass Extinction: fact, fiction or speculation? Biol. Reviews 2022, 97, 640–663. [Google Scholar] [CrossRef]
- Castellanos, E.; Lemos, M.F.; Astigarraga, L.; Chacón, N.; Cuvi, N.; Huggel, C.; Miranda, L.; Vale, M.M.; Ometto, J.P.; Peri, P.L.; et al. Central and South America. In Climate Change 2022: Impacts, Adaptation and Vulnerability. Contribution of Working Group II to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change, Pörtner, H.O., Roberts, D.C., Tignor, M., Poloczanska, E.S., Mintenbeck, K. Alegría, A. Craig, M., Langsdorf, S. Löschke, S., Möller, V., Okem, A., Rama, B., Eds.; Cambridge University Press: Cambridge, UK and New York, NY, USA, 2022; pp. 1689–1816. [Google Scholar] [CrossRef]
- Campbell, L.G.; Anderson, K.A.; Marcec-Greaves, R.; Cardoso, P.; Barton, P.S.; Birkhofer, K.; Chichorro, F.; Deacon, C.; Fartmann, T.; Fukushima, C.S.; Gaigher, R.; Habel, J.C.; Hallmann, C.A. Scientists' Warning to Humanity on Insect Extinctions. Biol. Conserv., 2020, 242, 108426:1–108426:1. [Google Scholar]
- Wagner, D.L.; Grames, E.M.; Forister, M.L.; Berenbaum, M.R.; Stopak, D. Insect Decline in the Anthropocene: Death by a Thousand Cuts. Proc. Nat.l Acad. Sci. USA 2023, 118, 2023989118. [Google Scholar] [CrossRef] [PubMed]
- Catenazzi, A. State of the World's Amphibians. Ann. Rev. Environ. Res. 2015, 40, 91–119. [Google Scholar] [CrossRef]
- Johnson, P.T.J. Amphibian diversity: Decimation by Disease. Proc. Natl. Acad. Sci. USA 2006, 103, 3011–3012. [Google Scholar] [CrossRef] [PubMed]
- AmphibiaWeb. Available online: https://amphibiaweb.org (accessed on 27 September 2023).
- Alroy, J. Current Extinction Rates of Reptiles and Amphibians. Proc. Natl. Acad. Sci. USA 2015, 112, 13003–13008. [Google Scholar] [CrossRef] [PubMed]
- Stokstad, E. Can a Dire Ecological Warning Lead to Action? Science 2019, 364, 517–518. [Google Scholar] [CrossRef]
- Danilov-Danil’yan, V.I., Reyf, I.E. Foundations of Sustainability in Nature and Society. In The Biosphere and Civilization: In the Throes of a Global Crisis; Springer, Cham, ZG; 2018; pp.199-205, ISBN 978-3-319-67193-2.
- Wiens, J.J. Global Patterns of Diversification and Species Richness in Amphibians. Am. Nat. 2007, 170 (Suppl 2), 86–106. [Google Scholar] [CrossRef]
- Bickford, D.; Poo, S.; Posa, M.R.C.; Gower, D.; Johnson, K.; Richardson, J.; Rosen, B.; Ruber, L.; Williams, S. Southeast Asian biodiversity crisis. In Biotic evolution and environmental change in Southeast Asia; Gower, A., Johnson, K., Richardson, J., Rosen, B., Rüber, L., S. Williams, S., Eds.; Cambridge University Press: UK, 2012; pp. 434–462. ISBN 9780511735882. [Google Scholar]
- Rivera-Correa, M.; Baldo, D.; Candioti, F.V.; Orrico, V.; Blackburn, D.C.; Castroviejo-Fisher, S.; Onn, C.K.; Gambale, P.; Gower, D.J.; Quah, E.; et al. Amphibians in Zootaxa: 20 years Documenting the Global Diversity of Frogs, Salamanders, and Caecilians. Zootaxa 2021, 4979, 057–069. [Google Scholar] [CrossRef]
- Womack, M.C.; Steigerwald, E.; Blackburn, D.C.; Cannatella, D.C.; Catenazzi, A.; Che, J.; Koo, M.S.; McGuire, J.A.; Ron, S.R.; Spencer, C.L.; et al. State of the Amphibia 2020: A Review of Five Years of Amphibian Research and Existing Resources. Ichthyol. Herpetol. 2022, 110, 638–661. [Google Scholar] [CrossRef]
- Browne, R.K.; Kaurova, S.A.; Vasudevan, K.; McGinnity, D.; Venu, G.; Gonzalez, M.; Uteshev, V.K.; Marcec-Greaves, R. Reproduction Technologies for the Sustainable Management of Caudata (salamander) and Gymnophiona (Caecilian) biodiversity. Reprod. Fert. Devel. 2022, 34, 479–497. [Google Scholar] [CrossRef]
- Venu, G.; Raju, N.G.; Wilkinson, M.; Browne, R.K.; Varadh, K.; Balakrishna, G.N.; Ramakrishna, S.; Venkatachalaiah, G. First records of the Long-headed Caecilian, Ichthyophis longicephalus Pillai, 1986 (Gymnophiona: Ichthyophiidae) from the States of Karnataka and Tamil Nadu, India with Comments on its Conservation Status. J. Anim. Divers. 2020, 2, 5–10. [Google Scholar] [CrossRef]
- Jaramillo, A.F.; De La Riva, I.; Guayasamin, J.M.; Chaparro, J.C.; Gagliardi-Urrutia, G.; Gutiérrez, R.C. , Brcko, I.; Vilà, C.; Castroviejo-Fisher, S. Vastly Underestimated Species Richness of Amazonian Salamanders (Plethodontidae: Bolitoglossa) and Implications about Plethodontid Diversification. Mol. Phylogenet. Evol. 2020, 149, 106841:1–106841:23. [Google Scholar] [CrossRef] [PubMed]
- Browne, R.K.; Wang, Z.; Okada, S.; McGinnity, D.; Luo, Q.; Taguchi, Y.; Kilpatrick, D.; Hardman, R.; Janzen, P.; Zhang, Z.; et al. The Sustainable Management of Giant Salamanders (Cryptobranchoidea). Review. Sustainability America, Belize, 2020.
- Gower, D. J.; Wilkinson, M. Conservation Biology of Caecilian Amphibians. Conserv. Biol. 2005, 19, 45–55. [Google Scholar] [CrossRef]
- Upton, J.; Gray, C.L.; Tapley, B.; Murray, K.A.; Gumbs, R. Poor Protection of Amphibian Evolutionary History Reveals Opportunities for Global Protected Areas. bioRxiv preprint 2020. [Google Scholar] [CrossRef]
- Ortega-Andrade, H. M.; Rodes Blanco, M.; Cisneros-Heredia, D.F.; Guerra Arévalo, N.; López de Vargas-Machuca, K.G.; Sánchez-Nivicela, J.C.; Yánez Muñoz, M. H. Red List Assessment of Amphibian Species of Ecuador: A Multidimensional Approach for their Conservation. PloSOne 2021, 16, e0251027:1–e0251027:28. [Google Scholar] [CrossRef]
- Oliver, P.M.; Bower, D.S.; McDonald, P.J.; Kraus, F.; Luedtke, J.; Neam, K.; Hobin, L.; Chauvenet, A.L.M.; Allison, A.; Arida, E.; et al. Melanesia Holds the World's most Diverse and Intact Insular Amphibian Fauna. Commun. Biol. 2022, 5, 1182:1–1182:10. [Google Scholar] [CrossRef]
- Andreone, F.; Carpenter, A.I.; Cox, N.; du Preez, L.; Freeman, K.; Furrer, S.; Garcia, G.; Glaw, F.; Glos, J.; Knox, D. The Challenge of Conserving Amphibian Megadiversity in Madagascar. PLOS Biol. 2008, 6, e118:0943–e118:0946. [Google Scholar] [CrossRef] [PubMed]
- Villacampa, J.; Whitworth, A.; Allen, L.; Malo, J.E. Altitudinal Differences in Alpha, Beta and Functional Diversity of an Amphibian Community in a Biodiversity Hotspot. Neotrop. Biodiver. 2019, 5(1), 60–68. [Google Scholar] [CrossRef]
- Khatiwada, J.R.; Zhao, T.; Chen, Y.; Wang, B.; Xei, F.; Cannatella, D.C.; Jiang, J. Amphibian Community Structure along Elevation Gradients in Eastern Nepal Himalaya. BMC Ecol. 2019, 19, 19:1–19:11. [Google Scholar] [CrossRef] [PubMed]
- Olson, D.H.; Ronnenberg, K.L.; Glidden, C.K.; Christiansen, K.R.; Blaustein, A.R. Global Patterns of the Fungal Pathogen Batrachochytrium dendrobatidis support Conservation Urgency. Front. Vet. Sci. 2021, 8, 685877:1–685877:20. [Google Scholar] [CrossRef]
- Kolby, J.E.; Daszak, P. The Emerging Amphibian Fungal Disease, Chytridiomycosis: A Key Example of the Global Phenomenon of Wildlife Emerging Infectious Diseases. Microbiol. Spectr. 2016, 4, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Green, D.E.; Dodd, C.K. Presence and Significance of Chytrid Fungus Batrachochytrium dendrobatidis and other Amphibian Pathogens at Warm-water Fish Hatcheries in Southeastern North America. Herpetol. Conserv. Biol. 2007, 2, 43–47. [Google Scholar]
- 74. Liew, N.; Mazon Moya, M.J.; Wierzbicki, C.J.; Hollinshead, M.; Dillon, M.J.; Thornton, C.R.; Ellison, A.; Cable, J.; Fisher, M.C.; Mostowy, S. Chytrid Fungus Infection in Zebrafish Demonstrates that the Pathogen can Parasitize Non-amphibian Vertebrate Hosts. Nat. Commun. 2017, 8, 15048:1–15048:10. [Google Scholar] [CrossRef]
- Burrowes, P.A.; De la Riva, I. Detection of the Amphibian Chytrid Fungus Batrachochytrium dendrobatidis in Museum Specimens of Andean Aquatic Birds: Implications for Pathogen Dispersal. J. Wildl. Dis. 2017, 53, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Gleason, F.H.; Chambouvet, A.; Sullivan, B.K.; Lilje, O.; Jodi, J.L.; Rowley, J.J.L. Multiple Zoosporic Parasites pose a Significant Threat to Amphibian Populations. Fungal Ecol. 2014, 11, 181–192. [Google Scholar] [CrossRef]
- Hernandez-Caballero, I.; Garcia-Longoria, L.; Gomez-Mestre, I.; Marzal, A. The Adaptive Host Manipulation Hypothesis: Parasites Modify the Behaviour, Morphology, and Physiology of Amphibians. Diversity, 2022, 14, 739:1–739:18. [Google Scholar] [CrossRef]
- Mendelson, J.R.; Whitfield, S.M.; Sredl, M.J. A Recovery Engine Strategy for Amphibian Conservation in the Context of Disease. Biol. Conserv. 2019, 236, 188–191. [Google Scholar] [CrossRef]
- Conde, D.A. , Staerk, J.; Colchero, F.; Vaupel, J.W. Data Gaps and Opportunities for Comparative and Conservation Biology. Conserv. Biol. 2019, 16, 9658–9664. [Google Scholar] [CrossRef]
- Tapley, B.; Michaels, C.J.; Gumbs, R.; Böhm, M.; Luedtke, J.; Pearce-Kelly, P.; Jodi, J.L.; Rowley, J.J.L. The Disparity Between Species Description and Conservation Assessment: A Case Study in Taxa with High Rates of Species Discovery. Biol. Conserv. 2018, 220, 209–214. [Google Scholar] [CrossRef]
- Streicher, J.W.; Sadler, R.; Loader, SP. Amphibian Taxonomy: Early 21st Century Case Studies. J. Nat. Hist. 2020, 54, 1–13. [Google Scholar] [CrossRef]
- Betts, J.; Young, R.P.; Hilton-Taylor, C.; Hoffmann, M.; Rodríguez, J.P.; Stuart, S.N.; Milner-Gulland, E.J. A Framework for Evaluating the Impact of the IUCN Red List of Threatened Species. Conserv. Biol. 2020, 34, 632–643. [Google Scholar] [CrossRef]
- IUCN 2023. The IUCN Red List of Threatened Species. Version 2022-2. Available online: https://www.iucnredlist.org (accessed on 27 September 2023).
- Garner, T.W.J.; Stephen, I.; Wombwell, E.; Fisher, M.C. The Amphibian Trade: Bans or Best Practice? EcoHealth 2009, 6, 148–151. [Google Scholar] [CrossRef] [PubMed]
- Kemp, T. S. Conservation and the future of amphibians, In Kemp, T. S., Amphibians: A Very Short Introduction; online edn, Oxford Academic, Oxford, USA, 2021; pp. 117–130, ISBN 9780198842989.
- Gerson, H. International Trade in Amphibians: a Customs Perspective. Alytes 2012, 29, 103–115. [Google Scholar]
- Schlaepfer, M.A.; Hoover, C.; Dodd Jr, C.K. Challenges in Evaluating the Impact of the Trade in Amphibians and Reptiles on Wild Populations. BioScience 2005, 55, 253–264. [Google Scholar] [CrossRef]
- Knapp C., R. , A., Lemm, J., Grant, T., Jackintell, L. Eds.; University of California Press: Berkeley, CO, USA, 2005; pp. 199–209, ISBN: 9780520238541.Iguanas. In Iguanas: Biology and Conservation. Alberts, A., Lemm, J., Grant, T., Jackintell, L. Eds.; University of California Press: Berkeley, CO, USA, 2005; University of California Press: Berkeley, CO, USA, 2005; pp. 199–209. ISBN 9780520238541. [Google Scholar]
- Langridge, J.; Sordello, R.; Reyjol, Y. Outcomes of Wildlife Translocations in Protected Areas: What is the Type and Extent of Existing Evidence? A Systematic Map Protocol. Environ. Evid. 2020, 9, 16:1–16:11. [Google Scholar] [CrossRef]
- LeDee, OA.; Handler, S.D.; Hoving, C.L.; Swanston, C.W.; Zuckerberg, B. Preparing Wildlife for Climate Change: How Far Have We Come? Journal of Wildlife Management 2021, 85, 7–16. [Google Scholar] [CrossRef]
- The SER International Primer on Ecological Restoration (Version 2: October, 2004). Society for Ecological Restoration International Science & Policy Working Group, Available online: https://cdn.ymaws.com/www.ser.org/resource/resmgr/custompages/publications/ser_publications/ser_primer.pdf.
- Galatowitsch, S.; Bohnen, J. Predicting Restoration Outcomes Based on Organizational and Ecological Factors. Restoration Ecol. 2020, 28, 1201–1212. [Google Scholar] [CrossRef]
- Byers, O.; Lees, C.; Wilken, J.; Schwitzer, C. The One Plan approach: The Philosophy and Implementation of CBSG's Approach to Integrated Species Conservation Planning. WAZA Mag. 2013, 14, 4–5. [Google Scholar]
- Griffiths, R.A.; Lissette Pavajeau, L. Captive Breeding, Reintroduction, and the Conservation of Amphibians. Conserv. Biol. 2008, 22, 852–861. [Google Scholar] [CrossRef]
- Mendelson III, J.R. Frogs in Glass Boxes: Responses of Zoos to Global Amphibian Extinctions. In The Ark and Beyond: The Evolution of Zoo and Aquarium Conservation; Minteer, B.A., Maienschein, J., Collins, J.P., Eds.; University of Chicago Press: Chicago, Ch, USA, 2018; pp. 298–313. ISBN 9780226538464. [Google Scholar]
- Canessa, S. No Conservation without Representation? Linked Decisions and Priority Setting in Amphibian ex situ programmes. Anim. Conserv. 2017, 20, 124–125. [Google Scholar] [CrossRef]
- Griffiths, R.A. Which Amphibians Should Qualify for the Ark? Anim. Conserv. 2017, 20, 120–121. [Google Scholar] [CrossRef]
- Scheele, B.C.; Guarino, F.; Osborne, W.; Hunter, D.A.; Skerratt, L.F.; Driscoll, D.A. Decline and Re-expansion of an Amphibian with High Prevalence of Chytrid Fungus. Biol. Conserv. 2014, 170, 86–91. [Google Scholar] [CrossRef]
- Barnett, K.M.; Civitello, D.J. Ecological and Evolutionary Challenges for Wildlife Vaccination. Trends Parasitol. 2020, 36, 970–978. [Google Scholar] [CrossRef]
- Ballou, J.D.; Lacy, R.C.; Traylor-Holzer, K.; Bauman, K.; Ivy, J.A.; Asa, C. Strategies for Establishing and Using Genome Resource Banks to Protect Genetic Diversity in Conservation Breeding Programs. Zoo Biol. 2022, 42, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Burger, I.; Julien, A.R.; Kouba, A.J.; Councell, K.R.; Barber, B.; Pacheco, C.; Kouba, C.K. Linking in situ and ex situ Populations of the Endangered Puerto Rican Crested Toad (Peltophryne lemur) through Genome Banking. Conserv. Sci. Pract. 2021, 3, e525:1–e525:12. [Google Scholar] [CrossRef]
- Burger, I.J.; Lampert, S.S.; Kouba, C.K.; Morin, D.J.; Kouba, A.J. Development of an Amphibian Sperm Biobanking Protocol for Genetic Management and Population Sustainability. Conserv. Physiol. 2022, 10, coac032:1–coac032:16. [Google Scholar] [CrossRef] [PubMed]
- Burger, I.J.; Chen, L.; Lampert, S.S.; Kouba, C.K.; Barber, D.; Smith, D.; Cobos, C.; Kouba, A.J. Applying Sperm Collection and Cryopreservation Protocols Developed in a Model Amphibian to Three Threatened Anuran Species Targeted for Biobanking Management. Biol. Conserv. 2023, 277, 109850:1–109850:6. [Google Scholar] [CrossRef]
- Kouba, C.K. ; Silla, A.J., Kouba, A.J., Heatwole, H., Eds.; CRC Press: London, UK, 2022; pp. 188-203, ISBN 9781032372075.Banks. In Reproductive Technologies and Biobanking for the Conservation of Amphibians; Silla, A.J., Kouba, A.J., Heatwole, H., Eds.; CRC Press: London, UK, 2022; CRC Press: London, UK, 2022; pp. 188–203. ISBN 9781032372075. [Google Scholar]
- Upton, R.; The University of Newcastle, Australia. Career Summary. Available online: https://www.newcastle.edu.au/profile/rose-upton (accessed on 23 September 2023).
- Ficetola, G.F.; Rondinini, C.; Bonardi, A.; Baisero, D; Padoa-Schioppa, E. Habitat Availability for Amphibians and Extinction Threat: a Global Analysis. Divers. Distrib. 2014, 21, 1–10. [Google Scholar] [CrossRef]
- Berger-Tal, O.; Blumstein, D.T.; Swaisgood, R.R. Conservation Translocations: a Review of Common Difficulties and Promising Directions. Anim. Conserv. 2020, 23, 121–131. [Google Scholar] [CrossRef]
- Ficetola, G. F. , Carlo R.; Bonardi A.; Katariya V.; Schioppa E. P.; Angulo A. An Evaluation of the Robustness of Global Amphibian Range Maps. J. Biogeogr. 2014, 41, 211–221. [Google Scholar] [CrossRef]
- Amphibian Population Management Guidelines. Created at an Amphibian Population Management Workshop; 2007 -11; San Diego, CA, USA. Schad, K., Ed.; Amphibian Ark, 2008, 31 p. Available online: https://www.amphibianark.org/wp-content/uploads/2018/08/AArk-Amphibian-Population-Management-Guidelines.pdf. 10 December.
- Zippel, K.; Johnson, K.; Agliardo, R.G.; Gibson, R.; McFadden, M.; Browne, R.; Martinez, C.M.; Townsend, E. The Amphibian ARK: A Global Community for Ex situ Conservation of Amphibians. Herpetol. Conserv. Biol. 2011, 6, 340–352. [Google Scholar]
- Conservation Needs Assessments. AArk. 2023. Available online: https://www.conservationneeds.org/default.aspx (accessed on 23 September 2023).
- EDGE. Evolutionary Distinct and Globally Endangered. Zoological Society of London. Available online: https://www.edgeofexistence.org/species/ (accessed on 23 September 2023).
- Gumbs, R.; Gray, C.L.; Bohm, M; Burfield, I. J.; Couchman, O.R.; Faith, D.P.; Forest, F.; Hoffmann, M.; Isaac, N.J.B.; Jetz, W. et al. The EDGE Protocol: Advancing the Prioritisation of Evolutionarily Distinct and Globally Endangered Species for Practical Conservation Action. PLoS Biol. 2023, 21, e3001991:1–e3001991:22. [Google Scholar] [CrossRef] [PubMed]
- Howard, S.D.; Bickford, D.P. Amphibians Over the Edge: Silent Extinction Risk of Data Deficient Species. Divers. Distrib. 2014, 20, 837–846. [Google Scholar] [CrossRef]
- Betts, J.; Young, R.P.; Hilton-Taylor, C.; Hoffmann, M.; Rodríguez, J.P.; Stuart, S.N. , Milner-Gulland, E.J. A Framework for Evaluating the Impact of the IUCN Red List of Threatened Species. Conserv. Biol. 2020, 34, 632–643. [Google Scholar] [CrossRef]
- Responsible Herpetological Project. Available online: https://responsibleherpetoculture.foundation/ (accessed on 27 September 2023).
- Mansour, N.; Lahnsteiner, F.; Patzner, R.A. Collection of Gametes from Live Axolotl, Ambystoma mexicanum, and Standardization of in vitro Fertilization. Theriogenology 2011, 75, 354–361. [Google Scholar] [CrossRef] [PubMed]
- Figiel, C. Cryopreservation of Sperm from the Axolotl Ambystoma mexicanum: Implications for Conservation. Herpetol. Conserv. Biol. 2013, 8, 748–855. [Google Scholar]
- Marcec, R.M. Development of Assisted Reproductive Technologies for Endangered North American Salamanders by Ruth Marie Marcec. Faculty of Mississippi State University, Degree of Doctor of Philosophy in Animal Physiology, 2016; Mississippi, MS, USA.
- Figiel Jr., C. R. Cold Storage of Sperm from the Axolotl, Ambystoma mexicanum. Herpetol. Conserv. Biol. 2022, 15, 367–371. [Google Scholar]
- Figiel Jr., C. R. Effects of Water Temperature on Gonads Growth in Ambystoma mexicanum Axolotl Salamanders. Animals 2023, 13, 874:1–874:8. [Google Scholar] [CrossRef]
- Taniguchi-Sugiura, Y.; Tanaka, E.M. Artificial Insemination in Axolotl. In Salamanders. Methods in Molecular Biology; Seifert, A.W., Currie, J.D., Eds.; Humana: New York, NY, USA, 2023; Volume 2562, pp. 417–423. ISBN 978-1-0716-2659-7. [Google Scholar]
- Marcec-Greaves, R.M.; Kouba, C.K.; Willard, S.T.; Kouba, A.J. Ovarian Ultrasound Analysis for Developing Temporal and Spatially Explicit Hormone Regimens for Induced Ovulation and Egg deposition in the Tiger Salamander (Ambystoma tigrinum). Theriogenology Wild 2023, 2, 100038:1–100038:8. [Google Scholar] [CrossRef]
- Lampert, S.S.; Burger, I.J.; Julien, A.R.; Gillis, A.B.; Kouba, A.J.; Barber, D.; Kouba, C.K. Sperm Cryopreservation as a Tool for Amphibian Conservation: Production of F2 Generation Offspring from Cryo-Produced F1 Progeny. Animals 2023, 13, 53. [Google Scholar] [CrossRef] [PubMed]
- Frankham, R. Genetics and extinction. Biol. Conserv. 2005, 126, 131–140. [Google Scholar] [CrossRef]
- AVISE, J. C. Phylogeography: The History and Formation of Species; JSTOR: Harvard University Press, 2000; ISBN 978-0-674-26870-8. [Google Scholar]
- Frankham, R.; Ballou, J.D.; Briscoe, D.A. Introduction to Conservation Genetics; Cambridge University Press, 2002, ISBN 9780511808999.
- Onley, I.R.; Moseby, K.E.; Austin, J.J. Genomic Approaches for Conservation Management in Australia under Climate Change. Life 2021, 11, 653. [Google Scholar] [CrossRef]
- Johnson, W.E., Koepfli, K. The Role of Genomics in Conservation and Reproductive Sciences. In: Holt, W., Brown, J., Comizzoli, P. (eds) Reproductive Sciences in Animal Conservation. Advances in Experimental Medicine and Biology, vol 753. Springer, New York, NY. 2014.
- Ralls, K.; Ballou, J.D.; Dudash, M.R.; Eldridge, M.D.B.; Fenster, C.B.; Lacy, R.C.; Sunnucks, P.; Frankham, R. Call for a Paradigm Shift in the Genetic Management of Fragmented Populations. Conserv. Lett. 2018, 11, 1–6. [Google Scholar] [CrossRef]
- Frankham, R. Do Island Populations have less Genetic Variation than Mainland Populations? Heredity 1997, 78, 311–327. [Google Scholar] [CrossRef]
- Edmands, S. Between a Rock and a Hard Place: Evaluating the Relative Risks of Inbreeding and Outbreeding for Conservation and Management. Molec. Ecol. 2007, 16, 463–475. [Google Scholar] [CrossRef] [PubMed]
- Richter, S.C.; Seigel, R.A. Annual Variation in the Population Ecology of the Endangered Gopher Frog, Rana sevosa Goin and Netting. Copeia 2002, 2002, 962–972. [Google Scholar] [CrossRef]
- Spielman, D.; Brook, B. W.; Frankham, R. Most Species are not Driven to Extinction before Genetic Factors Impact Them. Proceedings of the National Academy of Sciences U S A, 2004, 101, 15261–15264. [Google Scholar] [CrossRef]
- The black-footed ferret project. Revive and Restore. Available online: https://reviverestore.org/projects/black-footed-ferret/ (accessed on 23 September 2023).
- Willi, Y.; Hoffmann, A.A. Demographic Factors and Genetic Variation Influence Population Persistence under Environmental Change. J. Evol. Biol. 2009, 22, 124–133. [Google Scholar] [CrossRef]
- Johnston, L.A.; Lacy, R.C. Genome Resource Banking for Species Conservation: Selection of Sperm Donors. Cryobiology 1995, 32, 68–77. [Google Scholar] [CrossRef]
- Byrne, P.G.; Silla, A.J. Genetic Management of Threatened Amphibians: Using Artificial Fertilisation Technologies to Facilitate Genetic Rescue and Assisted Gene Flow. In Reproductive Technologies and Biobanking for the Conservation of Amphibians; Silla A.J., Kouba A.J., Heatwole H., Eds.; CSIRO, Melbourne, Australia, 2022; pp. 124–146, ISBN 9781032372075.
- Farquharson, K.A.; Hogg, C.J.; Grueber, C.E. A Meta-analysis of Birth-origin Effects on Reproduction in Diverse Captive Environments. Nat. Commun. 2018, 9, 1055:1–1055:10. [Google Scholar] [CrossRef]
- Anastas, Z.M.; Byrne, P.G.; O’Brien, J.K.; Hobbs, R.J.; Upton, R.; Silla, A.J. The Increasing Role of Short-Term Sperm Storage and Cryopreservation in Conserving Threatened Amphibian Species. Animals 2023, 13, 2094. [Google Scholar] [CrossRef]
- Beebee, T.J.C. Conservation Genetics of Amphibians. Heredity 2005, 95, 423–427. [Google Scholar] [CrossRef]
- Brennan, R.S.; Garrett, A.D.; Huber, K.E.; Hargarten, H.; Pespeni, M.H. Rare Genetic Variation and Balanced Polymorphisms are Important for Survival in Global Change Conditions. Proc. R. Soc. B. 2019, 286, 20190943:1–20190943:9. [Google Scholar] [CrossRef] [PubMed]
- Harnal, V.K.; Wildt, D.E.; Bird, D.M.; Monfort, S.L.; Ballou, J.D. Computer Simulations to Determine the Efficacy of Different Genome Resource Banking Strategies for Maintaining Genetic Diversity. Cryobiology 2002, 44, 122–131. [Google Scholar] [CrossRef] [PubMed]
- Mahony, M.J.; Clulow, J. Appendix 2. Cryopreservation and Reconstitution Technologies: A Proposal to Establish A Genome Resource Bank For Threatened Australian Amphibians. In Guidelines for minimising disease risks associated with captive breeding, raising and restocking programs for Australian frogs; Murray, K., Skerratt, L.F., Marantelli, G., Berger, L., Hunter, D., et al., Eds.; Canberra: Australian Government, Department of Sustainability, Environment, Water, Population and Communities, 2011, ID 1011-1151. [Google Scholar]
- Kouba, A.J. Genome Resource Banks as a Tool for Amphibian Conservation. In Reproductive Technologies and Biobanking for the Conservation of Amphibians. In Reproductive Technologies and Biobanking for the Conservation of Amphibians; Silla A.J., Kouba A.J., Heatwole H., Eds.; CSIRO, Melbourne, Australia, 2022; pp. 188–203, ISBN 9781032372075.
- Witzenberger, K.A.; Hochkirch, A. Ex situ Conservation Genetics: a Review of Molecular Studies on the Genetic Consequences of Captive Breeding Programs for Endangered Animal Species. Biodivers. Conserv. 2011, 20, 1843–1861. [Google Scholar] [CrossRef]
- Robert, A. Captive Breeding Genetics and Reintroduction Success. Biol. Conserv. 2009, 142, 2915–2922. [Google Scholar] [CrossRef]
- Karthikeyan, V.; Barkha, S.; Venu, G.; Gowri, M.; Lisa, G.; Kishor. G.B. Ex-situ Management of Amphibians in Indian Zoos. Central Zoo Authority, New Delhi, 2022; pp.102.
- AArk Online Founder Calculation Tool. Available online: https://www.amphibianark.org/tools/Founder%20calculation%20tool.htm (accessed on 23 September 2023).
- Frankham, R. , Ballou, J. D., Briscoe, D. A. Introduction to Conservation Genetics, 2nd ed.; Cambridge: Cambridge University Press, UK, 2010; p. 644. ISBN -0521702712. [Google Scholar]
- Albert, E.M.; Ferna´ndez-Beaskoetxea, S.; Godoy, J.A.; Tobler, U.; Schmidt, B.R.; Bosch, J. Genetic Management of an Amphibian Population after Chyridiomycosis Outbreak. Conserv. Genet. 2015, 16, 103–111. [Google Scholar] [CrossRef]
- Prieto-Benítez, S.; Morente-López, J.; Teso, M.L.R.; Lara-Romero, C.; García-Fernández, A.; Torres, E.; Iriondo, J.M. Evaluating Assisted Gene Flow in Marginal Populations of a High Mountain Species. Front. Ecol. Evol. 2021, 9, 638837. [Google Scholar] [CrossRef]
- Grummer, J.A.; Booker, T.R.; Matthey-Doret, R.; Nietlisbach, P.; Thomaz, A.T.; Whitlock, M.C. The Immediate Costs and Long-term Benefits of Assisted Gene Flow in Large Populations. Conserv. Biol. 2022, 36, 13911:1–13911:12. [Google Scholar] [CrossRef]
- Scheele, B.C.; Hunter, D.A.; Skerratt, L.F.; Brannelly, L.A.; Driscoll, D.A. Low Impact of Chytridiomycosis on Frog Recruitment Enables Persistence in Refuges Despite High Adult Mortality. Biol. Conserv. 2015, 182, 36–43. [Google Scholar] [CrossRef]
- Crates, R.; Stojanovic, D.; Heinsohn, R. The Phenotypic Costs of Captivity. Biol. Rev. Camb. Philos. Soc. 2023, 98, 434–449. [Google Scholar] [CrossRef] [PubMed]
- Stock, S.E.; Klop-Toker, K.; Wallace, S.; Kelly, O.; Callen, A.; Seeto, R.; Mahony, S.V.; Hayward, M.W.; Mahony, M.J. Uncovering Inbreeding, Small Populations, and Strong Genetic Isolation in an Australian Threatened Frog, Litoria littlejohni. Conserv. Genet. 2023. [Google Scholar] [CrossRef]
- Penhos, J.C.; Cornfield, J.; Cordero, F. Jr. Seasonal Variation of Spermiation Produced by Chorionic Gonadotropin in the Frog. Rev. Soc. Argent. Biol. 1954, 87, 77–87. [Google Scholar] [PubMed]
- Arregui, L.; Bóveda, P.; Gosálvez, J.; Kouba, A.J. Effect of Seasonality on Hormonally Induced Sperm in Epidalea calamita (Amphibia, Anura, Bufonidae) and its Refrigerated and Cryopreserved Storage. Aquaculture 2020, 529, e735677. [Google Scholar] [CrossRef]
- Keller, L. F.; Waller, D. M. Inbreeding Effects in Wild Populations. Trends Ecol. Evol. 2002, 17, 230–241. [Google Scholar] [CrossRef]
- Reed, D.H. , Frankham, R. Correlation Between Fitness and Genetic Diversity. Conserv. Biol. 2003, 17, 230–237. [Google Scholar] [CrossRef]
- Shine, R. Sexual Selection and Sexual Dimorphism in the Amphibia. Copeia 1979, 2, 297–306. [Google Scholar] [CrossRef]
- Taborsky, M.; Oliveira, R.F.; Brockmann, H.J. The Evolution of Alternative Reproductive Tactics: Concepts and Questions. In Alternative Reproductive Tactics: An Integrative Approach; Oliveira, R., Taborsky, M., Brockmann, H.J., Eds.; Cambridge University Press: Cambridge, UK, 2008; pp. 1–22. [Google Scholar] [CrossRef]
- Vieites, D.R.; Nieto-Román, S.; Barluenga, M.; Palanca, A.; Vences, M.; Meyer, A. Post-mating Clutch Piracy in an Amphibian. Nature 2004, 431, 305–308. [Google Scholar] [CrossRef]
- Brannelly, L.A.; Webb, R.; Skerratt, L.F.; Berger, L. Amphibians with Infectious Disease Increase their Reproductive Effort: Evidence for the Terminal Investment Hypothesis. Open Biol. 2016, 6, 150251:1–150251:9. [Google Scholar] [CrossRef] [PubMed]
- Browne, R.K.; Li, H.; Vaughan, M. Sexually Mediated Shedding of Myxobolus fallax Spores During Spermiation of Litoria fallax (anura). Dis. Aquat. Org. 2006, 72, 71–75. [Google Scholar] [CrossRef] [PubMed]
- Eiras, J.C. An Overview on the Myxosporean Parasites in Amphibians and Reptiles. Acta Parasitol. 2005, 50, 267–275. [Google Scholar]
- Browne, R. K.; Li, H., Seratt, J.; Kouba. A. Progesterone Improves the Number and Quality of Hormone Induced Fowler toad (Bufo fowleri) Oocytes. Reprod. Biol. Endocrinol 2006, 4, 1–7. [CrossRef]
- Browne, R.K.; Seratt, J.; Vance, C.; Kouba, A. Hormonal Priming, Induction of Ovulation and In-vitro Fertilization of the Endangered Wyoming Toad (Bufo baxteri). Reprod. Biol. Endocrinol 2006, 4, 34. [Google Scholar] [CrossRef]
- Rastogi, R.K. Seasonal Cycle in Anuran (Amphibia) Testis: The Endocrine and Environmental Controls. Bollettino di zoologia 1976, 43, 151–172. [Google Scholar] [CrossRef]
- Rastogi, R.K.; Tammaro, L.; Di Meglio, M.; Lela, L.; Di Matteo, L.; Chieffi, G. Circannual Testicular Rhythm in the Green Frog, Rana esculenta. Bollettino di zoologia 1981, 48, 97–105. [Google Scholar] [CrossRef]
- Poo, S.; Hinkson, K.M.; Stege, E. Sperm Output and Body Condition are Maintained Independently of Hibernation in an Endangered Temperate Amphibian. Reprod. Fert. Dev. 2019, 31, 796–804. [Google Scholar] [CrossRef]
- Poo, S.; Hinkson, K.M. Applying Cryopreservation to Anuran Conservation Biology. Conserv. Sci. Pract. 2019, 1, 91:1–91:10. [Google Scholar] [CrossRef]
- Poo, S.; Bogisich, A.; Mack, M.; Lynn, B.K.; Devan-Song, A. Post-release Comparisons of Amphibian Growth reveal Challenges with Sperm Cryopreservation as a Conservation Tool. Conserv. Sci. Pract. 2022, 4, 572:1–572:9. [Google Scholar] [CrossRef]
- Rangel, G. 2015. Available online: https://sitn.hms.harvard.edu/flash/2015/from-corgis-to-corn-a-brief-look-at-the-long-history-of-gmo-technology/ (accessed on 27 September 2023).
- Eskew, E.A.; Shock, B.C.; LaDouceur, E.E.B.; Keel, K.; Miller, M.R.; Foley, J.E.; Todd, B.D. Gene Expression Differs in Susceptible and Resistant Amphibians Exposed to Batrachochytrium dendrobatidis. R. Soc. Open Sci. 2018, 5, 170910. [Google Scholar] [CrossRef]
- Amphibian Ark and the IUCN SSC ART and Biobanking Working Group webinars. Available online: https://www.amphibianark.org/art-videos/ (accessed on 23 September 2023).
- Kaurova S., A.; Chekurova N., R.; Melnikova E., V.; Uteshev V., K.; Gakhova E., N. Cryopreservation of Frog Rana temporaria Sperm Without Loss of Fertilizing Capacity. In Proceedings of the 14th Working Meeting, Pushchino, 13–15 May 1996; 1996; Gakhova, E.N., Karnaukhov, V.N., Eds.; Pushchino Press: Pushchino, Russian; pp. 106–108. (In Russian). [Google Scholar]
- Kaurova S., A.; Uteshev V., K.; Chekurova N., R.; Gakhova E., N. Cryopreservation of Testis of Frog Rana temporaria. Infusionsther Transfusionsmed 1997, 24, 379. [Google Scholar]
- Browne, R. K.; Clulow, J.; Mahony, M.; Clark, A. Successful Recovery of Motility and Fertility of Cryopreserved Cane Toad (Bufo marinus) sperm. Cryobiology 1998, 37, 339–345. [Google Scholar] [CrossRef]
- Shishova, N. R. , Utishev, V. K., Kaurova, S. A., Browne, R. K., Gakhova, E. N. Cryopreservation of Hormonally Induced Sperm for the Conservation of Threatened Amphibians with Rana temporaria as a Model Species. Theriogenology 2011, 75, 220–232. [Google Scholar] [CrossRef]
- Russell, W.M.S.; Burch, R.L. The Principles of Humane Experimental Technique; Universities Federation for Animal Welfare: Wheathampstead, UK, 1959; ISBN 0-900767-78-2. [Google Scholar]
- Tannenbaum, J.; Bennett, B.T. Russell and Burch's 3Rs Then and Now: The Need for Clarity in Definition and Purpose. J. Am. Assoc. Lab. Anim. Sci. 2015, 54, 120–132. [Google Scholar]
- NC3R. National Center for the Replacement, Reduction, and Refinement of Animals in Research. Available online: https://www.nc3rs.org.uk/who-we-are/3rs (accessed on 23 September 2023).
- Ulloa, J.S.; Aubin, T.; Llusia, D.; Courtois, É.A.; Fouquet, A.; Gaucher, P.; Pavoine, S.; Sueur, J. Explosive Breeding in Tropical Anurans: Environmental Triggers, Community Composition and Acoustic Structure. BMC Ecol. 2019, 19, 28:1–28:17. [Google Scholar] [CrossRef]
- Trudeau, V.L.; Raven, B.H.; Pahuja, H.K.; Narayan, E. J 2022. Chapter 4. Hormonal Control of Amphibian Reproduction. In Reproductive Technologies and Biobanking for the Conservation of Amphibians, Silla, A.J., Kouba, A.J., Heatwole, H., Eds.; CRC Press: London, UK, 2022; ISBN 9781032372075. [Google Scholar]
- Zimkus, B.M.; Hassapakis, C.L.; Houck, M.L. Integrating Current Methods for the Preservation of Amphibian Genetic Resources and Viable Tissues to Achieve Best Practices for Species Conservation. Amphib. Reptile. Conserv. 2018, 12, 1–27. [Google Scholar]
- Bolton, R.L.; Mooney, A.; Pettit, M.T.; Bolton, A.E.; Morgan, L.; Drake, G.J.; Appeltant, R.; Walker, S.L.; Gillis, J.D.; Hvilsom, C. Resurrecting Biodiversity: Advanced Assisted Reproductive Technologies and Biobanking. Reprod. Fert. 2022, 3, R121–R146. [Google Scholar] [CrossRef]
- Garcia, N.C.; Robinson, W.D. Current and forthcoming Approaches for Benchmarking Genetic and Genomic Diversity. Front. Ecol. Evol. 2021, 9, 622603:1–622603:15. [Google Scholar] [CrossRef]
- Krell, F.T.; Wheeler, Q.D. Specimen Collection: Plan for the Future. Science 2014, 344, 815–816. [Google Scholar] [CrossRef]
- Rocha, L.A.; Aleixo, A.; Allen, G.; Almeda, F.; Baldwin, C.C.; Barclay, M.V.L.; Bates, M.; Bauer, A.M.; Benzoni, F.; Berns; C.M. et al. Specimen Collection: An Essential Tool. Science 2014, 344, 814–815. [CrossRef]
- Li, Y.; Hopkins, A.J.M.; Davis, R.A. Going, Going, Gone The Diminishing Capacity of Museum Specimen Collections to Address Global Change Research: A Case Study on Urban Reptiles. Animals 2023, 13, 1078:1–1078:13. [Google Scholar] [CrossRef]
- O'Hanlon, S.J.; Rieux, A.; Farrer, R.A.; Rosa, G.M.; Waldman, B.; Bataille, A.; Kosch, T.A.; Murray, K.A.; Brankovics, B.; Fumagalli, M.; et al. Recent Asian Origin of Chytrid Fungi causing Global Amphibian Declines. Science 2018, 360, 621–627. [Google Scholar] [CrossRef]
- McGinnity, D.; Reinsch, S.R.; Schwartz, H.; Trudeau, V.; Browne, R.K. Semen and Egg Collection, Sperm Cryopreservation, and in vitro Fertilisation with Threatened North American Giant Salamanders (Cryptobranchus alleganiensis). Reprod. Fertil. Dev. 2021, 34, 470–477. [Google Scholar] [CrossRef]
- Hamer, A.J.; Mahony, M.J. 2007. Life History of an Endangered Amphibian Challenges the Declining Species Paradigm. Australian J. Zool. 2007, 55, 79–88. [Google Scholar] [CrossRef]
- Lee, M.S.Y.; Jamieson, B.G.M. The Ultrastructure of the Spermatozoa of Bufonid and Hylid Frogs (Anure, Amphibia): Implications for Phylogeny and Fertilisation Biology. Zool. Scripta 1993, 22, 309–323. [Google Scholar] [CrossRef]
- Scheltinga, D.M.; Jamieson, B.G.M. Spermatogenesis and the Mature Spermatozoon: Form, Function and Phylogenetic Implications, In Reproductive biology and phylogeny of Anura; Jamieson, G.M., Ed.; Science Publishers Inc.: NH, USA. 2013; Volume 2, pp. 119–252. ISBN 1-57808-288-9. [Google Scholar]
- Roth, T.L.; Obringer, A.R. Reproductive Research and the Worldwide Amphibian Extinction Crisis. In Reproductive sciences and integrated conservation, Holt, W.V., Pickard A.R., Rodger J.C., Wildt D.E. Eds.; Cambridge University Press: Cambridge, UK, 2003; pp. 359–374. ISBN 0-521. [Google Scholar]
- Kouba, A.J.; Vance, C.K.; Frommeyer, M.A.; Roth, T.L. Structural and Functional Aspects of Bufo americanus Spermatozoa: Effects of Inactivation and Reactivation. J. Exp. Zool. 2003, 295, 172–182. [Google Scholar] [CrossRef]
- Chen, D.M.; Moore, M.G.; Willis, E.L.; Kouba, A.J.; Kouba, C.K. The Impact of Time and Environmental Factors on the Mitochondrial Vesicle and Subsequent Motility of Amphibian Sperm. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2022, 268, 11119:1–11119:8. [Google Scholar] [CrossRef]
- Silla, A.J.; Langhorne, C.J. Protocols for Hormonally Induced Sperm. In Reproductive Technologies and Biobanking for the Conservation of Amphibians, Silla, A.J., Kouba, A.J., Heatwole, H., Eds.; CSIRO Publishing: Melbourne, Australia, 2022; pp. 106–123. ISBN 9781486313341. [Google Scholar]
- Herbert, D. Studies of Assisted Reproduction in the Spotted Grass Frog Limnodynastes tasmaniensis: Ovulation, Early Development and Microinjection (ICSI), Masters by Research thesis; The University of Newcastle, Australia, 2004.
- Naranjo, R.E.; Naydenova, E.; Proaño-Bolaños, C.; Vizuete, K.; Debut, A.; Arias, M.T.; Coloma, L.A. Development of Assisted Reproductive Technologies for the Conservation of Atelopus sp. (spumarius complex). Cryobiology, 2022, 105, 20–31. [Google Scholar] [CrossRef]
- Upton, R.; Clulow, S.; Calatayud, N.E.; Colyvas, K.; Seeto, R.G.Y.; Wong, L.A.M.; Mahony, M.J.; Clulow, J. Generation of Reproductively Mature Offspring from the Endangered Green and Golden Bell Frog Litoriaaurea using Cryopreserved Spermatozoa. Reprod. Fertil. Dev. 2021, 33, 562–572. [Google Scholar] [CrossRef]
- Upton, R.; Clulow, S.; Colyvas, K.; Mahony, M.; Clulow, J. Paradigm Shift in Frog Sperm Cryopreservation: Reduced Role for Non-penetrating Cryoprotectants. Reproduction 2023, 165, 583–592. [Google Scholar] [CrossRef]
- Upton, R.; Clulow, S.; Mahony, M.J.; Clulow, J. Generation of a Sexually Mature Individual of the Eastern Dwarf Tree Frog Litoriafallax, from Cryopreserved Testicular Macerates: Proof of Capacity of Cryopreserved Sperm Derived Offspring to Complete Development. Conserv. Physiol. 2018, 6, coy043:1–coy043:5. [Google Scholar] [CrossRef]
- Graham, K.M’; Langhorne, C.J.; Vance, C.K.; Willard, S,T.; Kouba, A.J. Ultrasound imaging improves hormone therapy strategies for induction of ovulation and in vitro fertilization in the endangered dusky gopher frog (Lithobates sevosa). Conserv. Physiol. 2018, 6, coy020. [CrossRef]
- Browne, R.K.; Clulow, J.; Mahony, M. Short-term Storage of Cane Toad (Bufo marinus) Gametes. Reproduction 2001, 121, 167–173. [Google Scholar] [CrossRef]
- Germano, J.M.; Cree, A.; Molinia, F.C; Arregui, L.; Bishop, P.J. Hormone Treatment does not Reliably Induce Spermiation or Mating in Hamilton’s frog from the Archaic Leiopelmatid Lineage. Reprod. Fert. Devel. 2021, 34, 447–452. [Google Scholar] [CrossRef]
- Guy, E.L.; Gillis, A.B.; Kouba, A.J.; Barber, D.; Poole, V.; Marcec-Greaves, R.M.; Kouba, C.K. Sperm Collection and Cryopreservation for Threatened Newt Species. Cryobiology 2020, 94, 80–88. [Google Scholar] [CrossRef]
- Uteshev, V. K. , Kaurova, S. A., Shishova, N. V., Stolyarov, S. D., Browne, R. K., Gakhova, E. N. In vitro Fertilisation with Hormonally Induced Sperm and Eggs from Sharp-ribbed Newts Pleurodeles waltl. Russ. J. Herpetol. 2015, 22, 35–40. [Google Scholar] [CrossRef]
- Campbell, L.G.; Marcec-Greaves, R. Topical Application of Hormone Gonadotropin-releasing Hormone (GnRH-A) Stimulates Reproduction in the Endangered Texas Blind Salamander (Eurycea rathbuni). Conser. Sci. Pract. 2022, 4, e609:1–e609:8. [Google Scholar] [CrossRef]
- Jaiswal, A.N.; Vagga, A. Cryopreservation: A Review Article. Cureus 2022, 14, e31564. [Google Scholar] [CrossRef]
- Kaurova, S.A.; Browne, R.K.; Uteshev, V.K. Antibiotics for the Refrigerated Storage at 4 C of Hormonally Induced European Common Frog (Rana temporaria) Spermatozoa. Theriogenology Wild 2022, 1, 100009:1–100009:8. [Google Scholar] [CrossRef]
- Ishibashi, S.; Kroll, K.L.; Amaya, E. Generation of Transgenic Xenopus laevis: III. Sperm Nuclear Transplantation. CSH Protoc. 2007, 18, pdb.prot4840. [Google Scholar] [CrossRef]
- Miyamoto, K. , Simpson, D., Gurdon, J. B. Manipulation and in vitro Maturation of Xenopus laevis Oocytes, followed by Intracytoplasmic Sperm Injection, to Study Embryonic Development. J. Vis. Exp. 2015, 96, e52496:1–e52496:7. [Google Scholar] [CrossRef]
- de Siqueira-Silva, D.H.; Saito, T.; dos Santos-Silva, A.P.; da Silva Costa, R.; Psenicka, M.; Yasui, G.S. Biotechnology Applied to Fish Reproduction: Tools for Conservation. Fish Physiol. Biochem. 2018, 44, 1469–1485. [Google Scholar] [CrossRef]
- Khosla, K.; Kangas, J.; Liu, Y.; Zhan, L.; Daly, J.; Hagedorn, M.; Bischof, J. Cryopreservation and Laser Nanowarming of Zebrafish Embryos Followed by Hatching and Spawning. Adv. Biosyst. 2020, 4, e2000138. [Google Scholar] [CrossRef] [PubMed]
- Nikitina, L.A. Nuclear Transplantation in Fish and Amphibians. Physiol. Gen. Biol. Rev. 1997, 13, 35–72. (In Russian) [Google Scholar]
- Briggs, R.; King, T.J. Transplantation of Living Nuclei from Blastula Cells into Enucleated Frogs’ Eggs. Proc. Natl. Acad. Sci. USA 1952, 38, 455–463. [Google Scholar] [CrossRef] [PubMed]
- Sambuichi, H. The Roles of the Nucleus and the Cytoplasm in Development. I. An Interspecific Hybrid Frog, Developed from a Combination of Rana nigromaculata nigromaculata Cytoplasm and a Diploid Nucleus of Rana nigromaculata brevipoda. J. Sci. Hiroshima-Univ., Ser. B Div. 1957, 1, 33–41. [Google Scholar]
- Moore, J.A. Transplanation of Nuclei Between Rana pipiens and Rana sylvatica. Exp. Cell. Res. 1958, 14, 532–540. [Google Scholar] [CrossRef] [PubMed]
- Sambuichi, H. The Roles of the Nucleus and the Cytoplasm in Development. III. Diploid Nucleocytoplasmic Hybrids, Derived from Rana nigromaculata brevipoda Cytoplasm and Rana nigromaculata nigromaculata Nuclei. J. Sci. Hiroshima Univ. 1961, 20, 1–5. [Google Scholar]
- Gurdon, J.B. Adult Frogs Derived from the Nuclei of Single Somatic Cells. Dev. Biol. 1962, 4, 256–273. [Google Scholar] [CrossRef]
- Hennan, S. Nucleocytoplasmic Hybrids Between Rana pipiens and Rana palustris. I. Analysis of the Developmental Properties of the Nuclei by Means of Nuclear Transplantation. Dev. Biol. 1963, 11, 243–267. [Google Scholar] [CrossRef]
- Kawamura, T.; Nishioka, M. Reciprocal Diploid Nucleocytoplasmic Hybrids between Two Species of Japanese Pond Frogs and their Offspring. J. Sci. Hiroshima Univ., Ser. B., Div. 1, Zoology 1963, 21, 65–84. [Google Scholar]
- Kawamura, T.; Nishioka, M. Nucleo-cytoplasmic Hybrid Frog between Two Species of Japanese Brown Frogs and their Offspring. J. Sci. Hiroshima Univ., Ser. B., Div. 1, Zoology 1963, 21, 107–134. [Google Scholar]
- Kawamura, T.; Nishioka, M. Viability and Abnormalities of the Offspring of Nucleo-cytoplasmic Hybrids between Rana japonica and Rana ornativentris. Sci. Rep. Lab. Amphibian Biol. Hiroshima Univ. 1972, 1, 95–209. [Google Scholar]
- Nishioka, M. Abnormalities of the Offspring of Nucleo-cytoplasmic Hybrids between Rana nigromaculata and Rana brevipoda. Sci. Rep. Lab. Amphibian Biol. Hiroshima Univ. 1972, 1, 1–94. [Google Scholar]
- Nishioka, M. Nucleo-cytoplasmic Hybrids between Rana japonica and Rana temporaria temporaria. Sci. Rep. Lab. Amphibian Biol. Hiroshima Univ. 1, 211–243.
- Nishioka, M. Nucleo-cytoplasmic Hybrids between Rana brevipoda and Rana plancyi chosenica. Sci. Rep. Lab. Amphibian Biol. Hiroshima Univ. 1, 259–275.
- Gallien, L.; Aimar, C. A New Mode of Gemellarity, Effected by Nuclear Graft in Urodele Amphibia of the Genus Pleurodeles. C. R. Acad. Hebd. Seances Acad. Sci. D. 1971, 272, 3348–3351. [Google Scholar] [PubMed]
- Gallien, C.L.; Aimar, C.; Guillet, F. Nucleocytoplasmic Interactions During Ontogenesis in Individuals Obtained by Intra- and Interspecific Nuclear Transplantation in the Genus Pleurodeles (Urodale Amphibian. Dev. Biol. 1973, 33, 154–170. [Google Scholar] [CrossRef]
- Mezhzherin, S.; Morozov-Leonov, S.; Nekrasova, O.; Rostovskaya, O. Geographic Peculiarities of Structure and Hemicloning Reproduction of Pelophylax esculentus Water Frog Complex (Anura, Ranidae) Populations in the East European Plain within Ukraine. Amphibia-Reptilia 2023, 44, 263–275. [Google Scholar] [CrossRef]
- Reznick, D.; Travis, J. The Empirical Study of Adaptation in Natural Populations. Adaptation, Ch. 8 1996, 243–289.
- Rudin-Bitterli, T.S.; Evans, J.P.; Mitchell, N.J. Geographic Variation in Adult and Embryonic Desiccation Tolerance in a Terrestrial Breeding Frog. Evolution 2020, 74, 1186–1199. [Google Scholar] [CrossRef] [PubMed]
- Antwis, R.E.; Browne, R.K. Ultraviolet Radiation and Vitamin D3 in Amphibian Health, Behaviour, Diet and Conservation. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2009, 154, 184–190. [Google Scholar] [CrossRef] [PubMed]
- Verschooren, E.; Brown, R.K.; Vercammen, F.; Pereboom, J. Ultraviolet B Radiation (UV-B) and the Growth and Skeletal Development of the Amazonian Milk Frog (Trachycephalus resinifictrix) from Metamorphosis. J. Physiol. Pathophysiol. 2011, 2, 34–42. [Google Scholar]
- Baines, F.M.; Chattell, J.; Dale, J.; Garrick, D.; Gill, I.; Goetz, M.; Skelton, T.; Swatman, M. How much UVB does my reptile need? The UV-Tool, a guide to the selection of UV lighting for reptiles and amphibians in captivity. J. Zoo Aquarium Res. 2016, 4, 42–63. [Google Scholar]
- Tapley, B.; Rendle, M.; Baines, F.M.; Goetz, M.; Bradfield, K.S.; Rood, D.; Lopez, J.; Garcia, G.; Routh, A. Meeting Ultraviolet B Radiation Requirements of Amphibians in Captivity: a Case Study with Mountain Chicken Frogs (Leptodactylus fallax) and General Recommendations for Pre-release Health Screening. Zoo Biol. 2015, 34, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Ryder. O.A. Chapter 11. Bioresource Banking Efforts in Support of Amphibian Conservation. In Amphibian Conservation Action Plan. Gascon, C., Collins, J.P., Moore, R.D., Church, D.R., McKay, J., Mendelson JR III, E. Eds.; IUCN/SSC Amphibian Specialist Group: Gland, Switzerland and Cambridge, UK, 2007; ISBN 978-2-8317-1008-2. [Google Scholar]
- Pukazhenthi, B.S.; Pelican, K. ; Gascon, C., Collins, J.P., Moore, R.D., Church, D.R., McKay, J., Mendelson JR III, E. Eds.; IUCN/SSC Amphibian Specialist Group: Gland, Switzerland and Cambridge, UK, 2007; pp.38−39, ISBN 978-2-8317-1008-2.Banking. In Amphibian Conservation Action Plan; Gascon, C., Collins, J.P., Moore, R.D., Church, D.R., McKay, J., Mendelson JR III, E., Eds.; IUCN/SSC Amphibian Specialist Group: Gland, Switzerland and Cambridge, UK, 2007; IUCN/SSC Amphibian Specialist Group: Gland, Switzerland and Cambridge, UK, 2007; ISBN 978-2-8317-1008-2. [Google Scholar]
- Ghose, S.L.; Yap, T.A.; Byrne, A.Q.; Sulaeman, H.; Rosenblum, E.B.; Chan-Alvarado, A.; Chaukulkar, S.; Greenbaum, E.; Koo, M.S.; Kouete, M.T.; et al. Continent-wide Recent Emergence of a Global Pathogen in African Amphibians. Front. Conserv. Sci 2023, 4, 1069490. [Google Scholar] [CrossRef]
- Gray, M.J.; Carter, E.D.; Piovia-Scott, J.J.; Cusaac, P.W.; Peterson, A.C.; Whetstone, R.D.; Hertz, A.; Muniz-Torres, A.Y.; Bletz, M.C.; Woodhams, D.C. Broad Host Susceptibility of North American Amphibian Species to Batrachochytrium salamandrivorans Suggests High Invasion Potential and Biodiversity Risk. Nat. Commun 2023, 14, 3270. [Google Scholar] [CrossRef]
- Scheele, B.C.; Hollanders, M.; Hoffmann, E.P.; Newell, D.A.; Lindenmayer, D.B.; McFadden, M.; Gilbert, D.J.; Grogan, L.F. Conservation Translocations for Amphibian Species Threatened by Chytrid Fungus: A Review, Conceptual Framework, and Recommendations. Conserv. Sci. Pract. 2021, 3, e524:1–e524:15. [Google Scholar] [CrossRef]
- Scheele, B.C.; Pasmans, F.; Skerratt, L.F.; Berger, L.; Martel, A.; Beukema, W.; Acevedo, A.A.; Burrowes, P.A.; Carvalho, T.; Catenazzi, A.; et al. Amphibian Fungal Panzootic causes Catastrophic and Ongoing Loss of Biodiversity. Science 2019, 363, 1459–1463. [Google Scholar] [CrossRef]
- Grant, E.H.; Amburgey, S.M.; Gratwicke, B.; Chaves, V.A.; Belasen, A.M.; Bickford, D.; Bruhl, C.A.; Calatayud, N.E.; Clemann, N.; Clulow, S. Amphibian Conservation in the Anthropocene. Biol. Conserv. 2023, 236, 543–547. [Google Scholar] [CrossRef]
- Taronga Conservation Society Australia. Available online: https://taronga.org.au/conservation-and-science/current-research/frog-conservation-biobanking (accessed on 27 September 2023).
- Nature’s SAFE. Saving animals from extinction and halt biodiversity loss. Available online: https://www.natures-safe.com/ (accessed on 27 September 2023).
- United Nations Convention on Biodiversity. Available online: https://www.un.org/en/observances/biological-diversity-day/convention (accessed on 27 September 2023).
- da Cruz, R.E.; da Silva Olivera, H.G.; Salvarani, F.M. Wild Animals Biobanks: Literature Review. Res. Soc. Dev. 2022, 11, e48411831268:1–e48411831268:9. [Google Scholar] [CrossRef]
- International Society for Biological and Environmental Repositories [ISBER]. Available online: https://www.isber.org/page/BPR (accessed on 21 September 2023).
- San Diego Zoo Wildlife Alliance's Frozen Zoo. Available online: https://science.sandiegozoo.org/resources/frozen-zoo%C2%AE (accessed on 21 September 2023).
- Chinese Academy of Science Kunming cell bank. Available online: http://english.kiz.cas.cn/gre/skl_Facility/ (accessed on 21 September 2023).
- Hvilsom, C.; Mekarska, A; de Man, D. Play it Cool! How EAZA’s Collaborative Banking of Samples Helps Improve Population Management and Conservation. World Association of Zoos and Aquariums (WAZA) News 2022, 3, 20–22. [Google Scholar]
- Biodiversity Biobanks South Africa. Available online: https://bbsa.org.za/ (accessed on 21 September 2023).
- Conde, D.A. , Staerk, J.; Colchero, F.; da Silva, R.; Scholey, J.; Baden, H,M.; Jouvet L.; Syed, H.; Jongejans, E.; Meiri. S.; Gaillard, J. et al. Data Gaps and Opportunities for Comparative and Conservation Biology. Conserv. Biol. 2013, 27, 1135–1141. [Google Scholar] [CrossRef]
- Ryder, O.A. , Onuma, M. Viable Cell Culture Banking for Biodiversity Characterization and Conservation. Annu. Rev. Anim. Biosci. 2018, 6, 83–98. [Google Scholar] [CrossRef] [PubMed]
- CryoArks. Animal Biobanking for Research and Conservation. Available online: https://www.cryoarks.org/ (accessed on 21 September 2023).
- ABC, SG. IUCN Species Survival Commission, Animal Biobanking for Conservation Specialist Group. 2023. Available online: https://www.iucn.org/our-union/commissions/group/iucn-ssc-animal-biobanking-conservation-specialist-group (accessed on 27 September 2023).
- Johnson, J.; A; Ruta, G; Baldos, U; Cervigni, R; Chonabayashi, S; Corong, E; Gavryliuk, O; Gerber, J; Hertel, T; Nootenboom, C. et al. 2021. Available online: http://hdl.handle.net/10986/35882 (accessed on 27 September 2023).
- Roe, D.; Holland, E.; Nisi, N. Mitchell, T.; Tasnim, T. Loss and Damage Finance should apply to Biodiversity Loss. Nat. Ecol. Evol. 2023, 7, 1336–1338. [Google Scholar] [CrossRef] [PubMed]
- Kunming-Montreal Global Biodiversity Framework. Targets and Guidance Notes. Available online: https://www.cbd.int/gbf/targets/ (accessed on 27 September 2023).
- Groves, C.R.; Game, E.T.; Anderson, M.G.; Cross, M.; Enquist, C.; Ferdaña, Z.; Girvetz, E.; Gondor, A.; Hall, K.R.; Higgins, J. Incorporating Climate Change into Systematic Conservation Planning. Biodivers. Conserv. 2012, 21, 1651–1671. [Google Scholar] [CrossRef]
- GIFT, 2023. Available online: https://global-inst.com/. Why Act Now: The World in 2050. Jeffrey Sachs, Jane Goodall, Obiora Ike, Brian Wong. Available online: https://www.youtube.com/watch?v=KEp8XGvT8MY (accessed on 21 September 2023).
- Guenot, M. Billion-dollar Startup Plans to bring the Dodo back from the Dead, and claims it can also Revive the Woolly Mammoth. Available online: https://www.businessinsider.com/scientists-revive-mammoth-dodo-colossal-biosciences-2023-2 (accessed on 12 February 2023).
- Martinez, A.; Mammola, S. SSpecialized Terminology Reduces the Number of Citations of Scientific Papers. Proc. R. Soc. B, Biol. Sci. 2021, 288. [Google Scholar] [CrossRef]
- Litvin, S.W. ,Kar, G.H. Individualism/collectivism as a Moderating Factor to the Self-image Congruity Concept. J. Vacat. Mark. 2004, 10, 23–32. [Google Scholar] [CrossRef]
- Wang, H.; Fu, T.; Du, Y.; Gao, W.; Huang, K.; Liu, Z.; Chandak, P.; Liu, S.; Katwyk, P.V.; Deac, A. Scientific Discovery in the Age of Artificial Intelligence. Nature 2023, 620, 47–60. [Google Scholar] [CrossRef]
- Goralski, M.A; Tan, T.K. Artificial Intelligence and Sustainable Development. Inter. J. Manag. Educ., 2020, 18, 100330. [Google Scholar] [CrossRef]
- Bhardwaj, A.; Kishore, S.; Pandey, D.K. Artificial Intelligence in Biological Sciences. Life (Basel) 2022, 12, 1430. [Google Scholar] [CrossRef]
- Powell, D.M.; Meyer, T.G. , Duncan, M. 2023. By Bits and Pieces: The Contributions of Zoos and Aquariums to Science and Society via Biomaterials. J. Zool. Bot. Gard. 2023, 4, 277–287. [Google Scholar] [CrossRef]
- Biocentrism. Science Direct, 2023. Available online: https://www.sciencedirect.com/topics/social-sciences/biocentrism (accessed on 21 September 2023).
- Pipere, A.; Mārtinsone, K. Metamodernism and Social Sciences: Scoping the Future. Soc. Sci. 2022, 11, 457:1–457:20. [Google Scholar] [CrossRef]
- Haq, S.M.; Pieroni, A.; Bussmann, R.W.; Abd-ElGawad, A.M.; El-Ansary, H.O. Integrating Traditional Ecological Knowledge into Habitat Restoration: Implications for Meeting Forest Restoration Challenges. J. Ethnobiol. Ethnomed. 2023, 19, 33:1–33:19. [Google Scholar] [CrossRef]
- Folke, C.; Biggs, R.; Norström, A.V.; Reyers, B.; Rockström, J. Social-ecological Resilience and Biosphere-based Sustainability Science. Ecol. Soc. 2016, 21, 41. [Google Scholar] [CrossRef]
- Reyes-García, V.; Cámara-Leret, R.; Halpern, B.S.; O’Hara, C.; Renard, D.; Zafra-Calvo, N.; Sandra Díaz. Biocultural Vulnerability Exposes Threats of Culturally Important Species. PNAS 2023, 120, e2217303120. [CrossRef]
- Crutzen, P.J.; Stoemer, E.F. The “Anthropocene”. The International Geosphere–Biosphere Programme (IGBP): A Study of Global Change of the International Council for Science (ICSU) Newsletter. Global Change Newsletters 2000, 41, 17–18. [Google Scholar]
- Ellis, E.C.; Ramankutty, N. Putting People in the Map: Anthropogenic Biomes of the World. Front. Ecol. Environ. 2008, 6, 439–447. [Google Scholar] [CrossRef]
- Ellis, E.C. The Great Acceleration, Anthropocene: A Very Short Introduction (Very Short Introductions); Oxford University Press: New York, NY, USA, 2018; p. 208. ISBN -13. [Google Scholar]
- Sobecki, S. 'New World Discovery', Oxford Handbook Topics in Literature (online edn, Oxford Academic, 16 Dec. 2013), Available online:. Available online: doi.org/10.1093/oxfordhb/9780199935338.013.141 (accessed on 29 September 2023).
- Arnason, J.P. Civilisation, Culture and Power: Reflections on Norbert Elias’s Genealogy of the West. In Civilising Processes and Modernity – A Debate. Palgrave Studies on Norbert Elias; Bogner, A., Mennell, S., Eds.; Civilisations, Palgrave Macmillan: Cham, ZG, 2022; ISBN 978-3-030-80378-0. [Google Scholar]
- Boon, P.I. The Environmental History of Australian Rivers: a Neglected Field of Opportunity? Mar. Freshw. 2018, 71, 1–45. [Google Scholar] [CrossRef]
- Turvey, S.T. Chapter 2. In the shadow of the megafauna: prehistoric mammal and bird extinctions across the Holocene. In Holocene Extinctions; Turvey, S.T., Ed.; Oxford University Press: New York, NY, USA, 2009; ISBN 9780199535095. [Google Scholar]
- Dantas, V.L.; Pausas, J.G. The Legacy of the Extinct Neotropical Megafauna on Plants and Biomes. Nat. Commun. 2022, 13, 129:1–129:13. [Google Scholar] [CrossRef] [PubMed]
- Cornford. R.; Spooner, F.; McRae, L.; Purvis, A.; Freeman, R. Ongoing Over-exploitation and Delayed Responses Change Highlight the Urgency for Action to Promote Vertebrate Recoveries by 2030. Proc. Biol Sci. 2023, 290, 20230464:1–20230464:9. [Google Scholar] [CrossRef]
- Bateman, I.; Balmford, A. Current Conservation Policies Risk Accelerating Biodiversity Loss. Nature 2023, 618, 671–674. [Google Scholar] [CrossRef] [PubMed]
- Boehm, S.; Schumer, C.; 10 Big Findings from the 2023. IPCC Sixth Assessment Report on Climate Change. 2023. Available online: https://www.wri.org/insights/2023-ipcc-ar6-synthesis-report-climate-change-findings (accessed on 20 September 2023).
- Barry, B. Sustainability and Intergenerational Justice. Theoria, 1997, 89, 43–65. [Google Scholar] [CrossRef]
- Spanning, R. Youth in the Anthropocene: Questions of Intergenerational Justice and Learning in a More-Than-Human World. In Youth Cultures in a Globalized World.; Knapp, G., Krall, H. Eds.; Springer, Cham, ZG, 2021; pp 113–133, ISBN 3030651762.
- Brozović, D. Societal collapse: A literature review. Futures 2023, 145, 103075:1–103075:24. [Google Scholar] [CrossRef]
- Quesada-Román, A.; Pérez-Umaña, D. State of the Art of Geodiversity, Geoconservation, and Geotourism in Costa Rica. Geosciences 2020, 10, 211:1–211:17. [Google Scholar] [CrossRef]
- Oluniyi, O. Darwin Comes to Africa: Social Darwinism and British Imperialism in Northern Nigeria.; Discovery Institute press: Seattle, WA, USA, 2023; ISBN 978-1637120231. [Google Scholar]
- Moore, J.W. Anthropocene or Capitalocene? Nature, history, and the Crisis of Capitalism. In Anthropocene or Capitalocene? Nature, history, and the Crisis of Capitalism; Moore, J.W., Ed.; PM Press: Oakland, CA, 2016; pp. 1–13. ISBN 978-1629631486. [Google Scholar]
- Lidicker, W.Z. Jr. A Scientist’s Warning to Hhumanity on Human Population Growth. Global Ecol. Conserv. 2020, 24, e01232:1–e01232:19. [Google Scholar] [CrossRef]
- CPA. 2010. World People’s Conference on Climate Change and the Rights of Mother Earth. April 22nd, Cochabamba, Bolivia. Cochabamba People’s Agreement. 22nd April, 2010. Available online: https://pwccc.wordpress.com/2010/04/24/peoples-agreement/ (accessed on 27 September 2023).
- Saito, K. Marx in the Anthropocene: Towards the idea of degrowth communism. Cambridge University Press: Cambridge, UK, 2023; 300p., ISBN 978-1108844154.
- Sachs, J.; Goodall, J.; Ike, O.; Brian Wong, B.; GIFT. Why Act Now: The World in 2050. 2023. Available online: https://www.youtube.com/watch?v=KEp8XGvT8MY (accessed on 22 September, 2023).
- Graeber, D.; Wengrow, D. The Dawn of Everything: a New History of Humanity. Farrar, Straus and Giroux: New York, NY, USA, 2021; ISBN 978-0374157357.
- de Lange, E.; Sze, J.S.; Allan, J.; Atkinson, S.; Booth, H.; Fletcher, R.; Khanyari, M.; Saif, O. A Global Conservation Basic Income to Safeguard Biodiversity. Nat. Sustain. 2023, 6, 1016–1023. [Google Scholar] [CrossRef]
- GBI. Global Basic Income Foundation. 2023. Available online: https://www.globalincome.org/English/Global-Basic-Income.html (accessed on 24 September 2023).
- Domínguez, L.; Luoma, C. Decolonising Conservation Policy: How Colonial Land and Conservation Ideologies Persist and Perpetuate Indigenous Injustices at the Expense of the Environment. Land 2020, 9, 65:1–65:22. [Google Scholar] [CrossRef]
- Hof, C.; Araujo, M.B.; Jetz, W; Rahbek, C. Additive threats from pathogens, climate and land-use change for global amphibian diversity. Nature 2012, 480, 516–519. [Google Scholar] [CrossRef]
- Ceballos, G.; Ehrlich. P.R.; Barnosky, A.D.; García, A.; Pringle, R.M.; Palmer, T.M. Accelerated Modern Human–induced Species Losses: Entering the Sixth Mass Extinction. Sci. Adv 2015, 1, e140025. [CrossRef]
- Inukai, I. Terminal Colonialism and its Impact on Developing Economic Growth: a Case of Ghana. J. Afr. Stud. 1981, 20, 1–14. [Google Scholar]
- Willcock, S.; Cooper, G.S.; Addy, J. Dearing, J.A. Earlier Collapse of Anthropocene Ecosystems Driven by Multiple Faster and Noisier Drivers. Nat. Sustain 2023. [CrossRef]
- Ziegler, T. Cologne Zoo Aquariums 50-year anniversary – The Road to a Conservation Center. WAZA News 2022, 3, 15–19. [Google Scholar]
- Katagiri, C. On the Fertilizability of the Frog Egg I. J. Fac. Sci. Hokkaido Univ., Ser. VI. Zool. 1961, 14, 607–613. [Google Scholar]
- Wolf, DP.; Hendrick, J.L. A Molecular Approach to Fertilisation. II. Viability and Artificial Fertilisation of Xenopus laevis Gametes. Dev. Biol. 1971, 25, 348–359. [Google Scholar] [CrossRef]
- Hollinger, TG.; Corton, G.L. Artificial Fertilization of Gametes from the South African Clawed Frog, Xenopus laevis. Gamete Res. 1980, 3, 45–57. [Google Scholar] [CrossRef]
- Edwards, D.L.; Mahony, M.J.; Clulow, J. Effect of Sperm Concentration, Medium Osmolality and Oocyte Storage on Artificial Fertilisation Success in a Myobatrachid Frog (Limnodynastes tasmaniensis). Reprod. Fertil. Dev. 2004, 16, 347–354. [Google Scholar] [CrossRef]
- Uteshev, V.K.; Gakhova, E.N.; Kramarova, L.I.; Shishova, N.V.; Kaurova, S.A.; Browne, R.K. Refrigerated Storage of European Common Frog Rana temporaria Oocytes. Cryobiology 2018, 83, 56–59. [Google Scholar] [CrossRef]
- Gagarinskiy, E.; Uteshev, V.; Fesenko, E.Jr. Prolonged hypothermic storage of oocytes of the European Common Frog Rana temporaria in a Gas Mixture of Oxygen and Carbon Monoxide. PLoS ONE 2023, 18, e0288370:1–e0288370:10. [Google Scholar] [CrossRef]
- Olson, J.H.; Chandler, D.E. Xenopus laevis Egg Jelly contains Small Proteins that are Essential to Fertilization. Dev. Biol. 1999, 210, 401–410. [Google Scholar] [CrossRef]
- Omata, S. Relative Roles of Jelly Layers in Successful Fertilization of Bufo japonicus. J. Exp. Zool. 1993, 265, 329–335. [Google Scholar] [CrossRef]
- Ukita, M.; Itoh, T.; Watanabe, T.; Watanabe, A.; Onitake, K. Substances for the Initiation of Sperm Motility in Egg-jelly of the Japanese Newt, Cynops pyrrhogaster. Zool. Sci. 1999, 16, 793–802. [Google Scholar] [CrossRef]
- Browne, R.K.; Clulow, J.; Mahony, M. The effect of saccharides on the post-thaw recovery of Cane Toad (Bufo marinus) Spermatozoa. Cryo-Lett. 2002, 23, 121–128. [Google Scholar]
- Liu, M.; Yuan, S.; Zhao, Z.; Liu, M.; Lin, A.; Gong, Q. Non-programmable Cryopreservation of Sperm from Industrial-farmed Murray Cod (Maccullochella p peelii). Aquaculture 2021, 541, 736811:1–736811:8. [Google Scholar] [CrossRef]
- Hobbs, R.J.; Upton, R.; Calatayud, N.E.; Silla, A.J.; Daly, J.; McFadden, M.S.; O’Brien, J.K. Cryopreservation Cooling Rate Impacts Post-thaw Sperm Motility and Survival in Litoria booroolongensis. Animals 2023, 13, 3014:1–3014:17. [Google Scholar] [CrossRef]
- Dabagyan, N.V.; Sleptsova, L.A. Artificial insemination. In Common Frog Rana temporaria. Objects of Developmental Biology.; Detlaf, T.A., Ed.; Nauka: Moscow, RU, 1975. [Google Scholar]
- Silla, A.J.; Kouba, A.J. Chapter 1. In tegrating Reproductive Technologies into the Conservation Toolbox for the Recovery of Amphibian Species. In Reproductive Technologies and Biobanking for the Conservation of Amphibians; Silla, A.J., Kouba, A.J., H Heatwole, H., Eds.; CRC Press: London, UK, 2022; pp. 1–9. ISBN 9781032372075. [Google Scholar]
- Browne, R.K. Book Review of ‘Reproductive Technologies and Biobanking for the Conservation of Amphibians’ by Silla, A. J., Kouba, A.J., Heatwole, H. (eds.); CSIRO Publishing: Victoria, Australia, 2022. 238 pp.; Herpetol. Rev. 2023, 54, 332–335. [Google Scholar]
- Cabada, M.O. Sperm Concentration and Fertilisation Rate in Bufo arenarum (Amphibia: Anura). J. Exp. Biol. 1975, 62, 481–486. [Google Scholar] [CrossRef] [PubMed]
- Hollinger, T.G.; Corton, G.L. Artificial Fertilization of Gametes from the South African Clawed Frog, Xenopus laevis. Gamete Res. 1980, 3, 45–57. [Google Scholar] [CrossRef]
- Silla, A.J. Artificial Fertilisation in a Terrestrial Toadlet (Pseudophryne guentheri): Effect of Medium Osmolarity, Sperm Concentration and Gamete Storage. Reprod. Fert. Devel. 2013, 25, 1134–1141. [Google Scholar] [CrossRef]
- Burger, I.J.; Lampert, S.S.; Kouba, C.K.; Morin, D.J.; Kouba, A.J. Development of an Amphibian Sperm Biobanking Protocol for Genetic Management and Population Sustainability. Conserv. Physiol. 2022, 10, coac032. [Google Scholar] [CrossRef]
- Serbinova, I. A.; Tuniyev, B.S. 1986. Keeping, captive breeding and reintroduction of northern banded newt (Triturus vittatus). In Sokolov V. E., Syroechkovskiy E. E. (Eds.), First all-Union Meeting on Problems of Zooculture, , Moscow. Moscow, Russ: Academy of Sciences of the USSR, 147–150. (In Russian). 2–4 December.
- Goncharov, B.F.; Shubravy, O.I.; Serbinova, I.A.; Uteshev, V.K. The USSR programme for breeding amphibians, including rare and endangered species. Int. Zoo Yearbook 1986, 28, 10–21. [Google Scholar] [CrossRef]
- Kaurova S., A. , Uteshev V. K., Gapeyev A. B., Shishova N. V., Gakhova E. N., Browne R. K., Kramarova L. I. Cryopreservation of spermatozoa obtained postmortem from the European common frog Rana temporaria. Reprod. Fert. Dev., 2021, 33, 588–595. [Google Scholar]
Figure 1.
The concepts of Metanature, Biospheric Sustainability, and RBCs, in tandem can perpetuate genetic diversity to provide for human wellbeing and intergenerational justice for the foreseeable future.
Figure 1.
The concepts of Metanature, Biospheric Sustainability, and RBCs, in tandem can perpetuate genetic diversity to provide for human wellbeing and intergenerational justice for the foreseeable future.
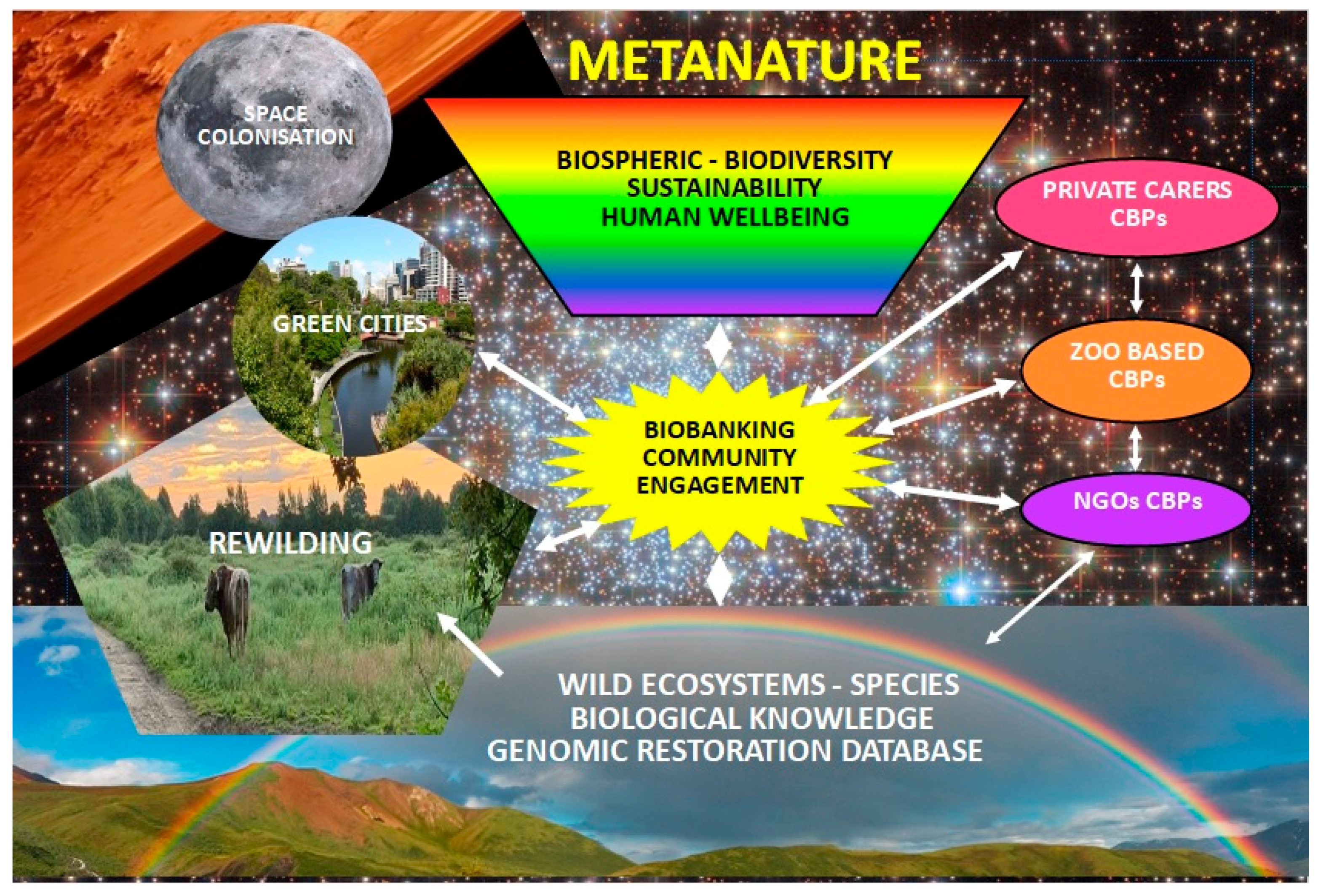
Figure 2.
(a) Critically Endangered San Martin Fringe-Limbed Treefrog, Ecnomiohyla valancifer. (b) Critically Endangered demonic poison frog, Minyobates steyermarki. Images courtesy Peter Janzen.
Figure 2.
(a) Critically Endangered San Martin Fringe-Limbed Treefrog, Ecnomiohyla valancifer. (b) Critically Endangered demonic poison frog, Minyobates steyermarki. Images courtesy Peter Janzen.

Figure 3.
(a) The proportional heterozygosity compared to founder numbers. (b) The probability that alleles will persist into the next generation is based on the initial allelic frequency in the founder the population and the number of founders.
Figure 3.
(a) The proportional heterozygosity compared to founder numbers. (b) The probability that alleles will persist into the next generation is based on the initial allelic frequency in the founder the population and the number of founders.
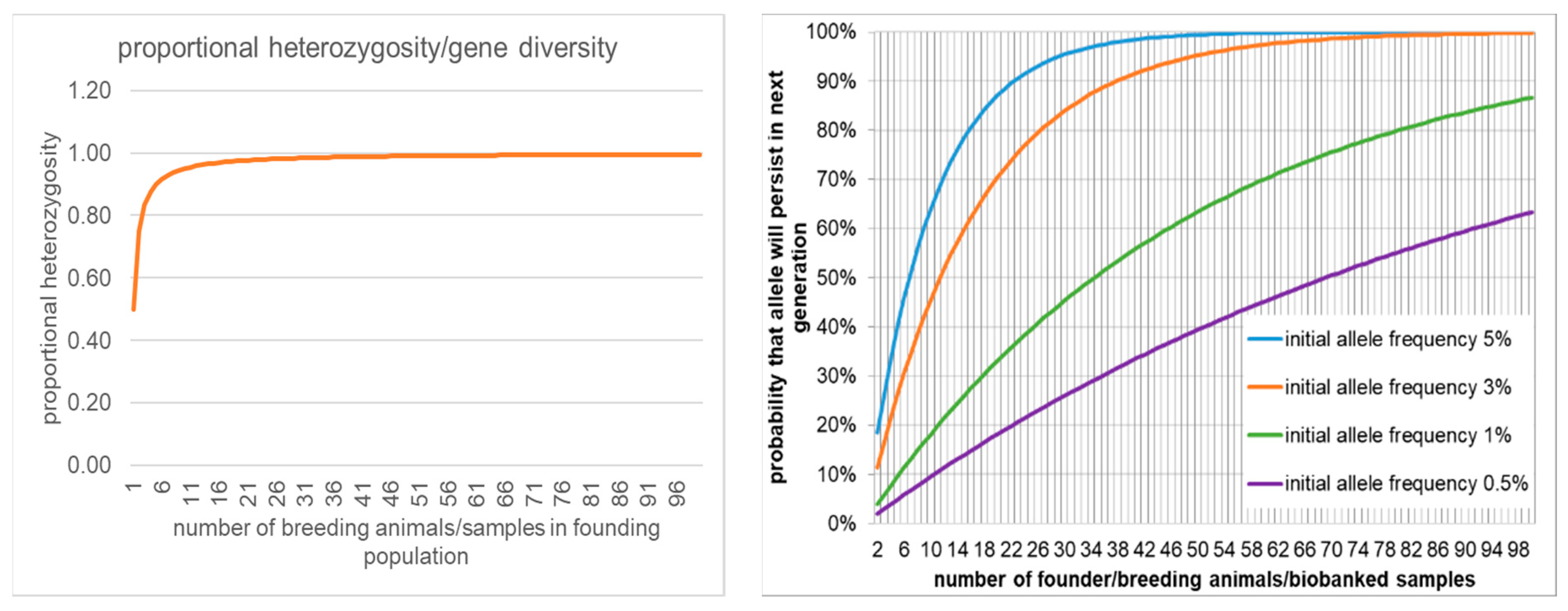
| Endangerment | CE | EN | VU | NT | LC |
|---|---|---|---|---|---|
| Anurans | 9 / 8, 89% | 16 / 16, 100% | 11 / 11, 100% | 15 / 11, 100% | 21 / 11, 100% |
| Salamanders | 6 / 6, 100% | 7 / 6, 86% | 10 / 11, 100% | 13 / 12, 92% | 44 / 42, 95% |
Figure 4.
Anuran oocyte numbers. Left. the lower 90% (698 of 791 species) spawn up to 2,100 oocytes, whereas the final 10% spawn up to 20,000 oocytes; mainly in Bufonid and large Ranid species. Right – the lower 50% of species mainly spawn < 70 oocytes, with many spawning <36 oocytes, but some up 350 oocytes.
Figure 4.
Anuran oocyte numbers. Left. the lower 90% (698 of 791 species) spawn up to 2,100 oocytes, whereas the final 10% spawn up to 20,000 oocytes; mainly in Bufonid and large Ranid species. Right – the lower 50% of species mainly spawn < 70 oocytes, with many spawning <36 oocytes, but some up 350 oocytes.
Figure 5.
A timeline of some major milestones in the development and application of amphibian RBCs from1980 to 2023. For details see Appendix A.6.
Figure 5.
A timeline of some major milestones in the development and application of amphibian RBCs from1980 to 2023. For details see Appendix A.6.
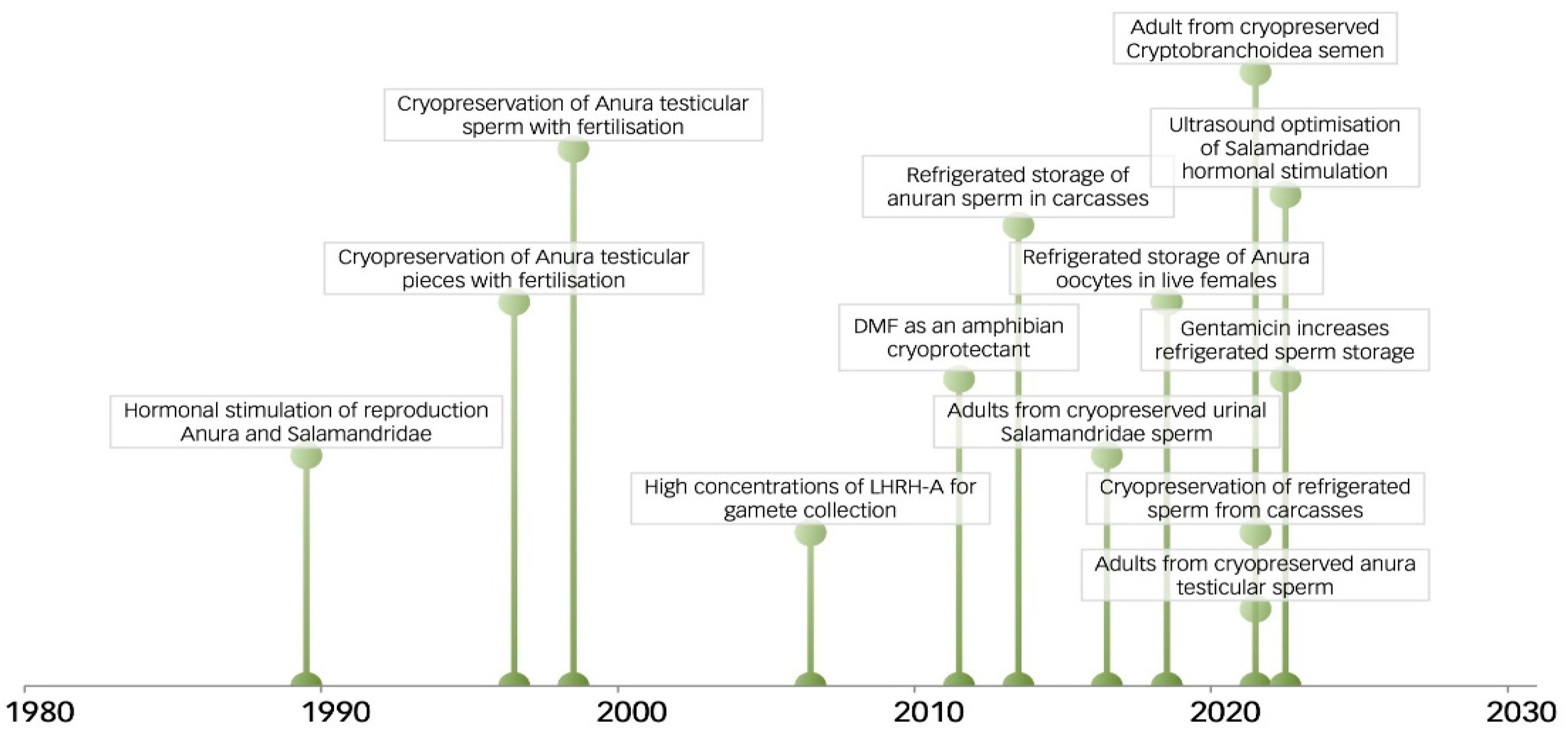
Figure 6.
Hamilton’s frog [Leiopelma hamiltoni]. Image by Phil Bishop. Attribution ShareAlike 2.5. https://en.wikipedia.org/wiki/Hamilton%27s_frog#/media/File:Leiopelma_hamiltoni02.jpg Phil Bishop, CC BY-SA 2.5 <https://creativecommons.org/licenses/by-sa/2.5>, via Wikimedia Commons.
Figure 6.
Hamilton’s frog [Leiopelma hamiltoni]. Image by Phil Bishop. Attribution ShareAlike 2.5. https://en.wikipedia.org/wiki/Hamilton%27s_frog#/media/File:Leiopelma_hamiltoni02.jpg Phil Bishop, CC BY-SA 2.5 <https://creativecommons.org/licenses/by-sa/2.5>, via Wikimedia Commons.

Figure 7.
Anuran oocyte volumes (mm3). (a) 805 species with oocyte diameters to 4.9 mm and volumes to 31.3 mm3. (b) 428 of these 805 species from the first two bars in the left figure with oocyte diameters ≤ 1.8 mm and volumes ≤ 5.0 mm3. The only successful cryopreservation of mammalian embryos 0.0002 to 0.004 mm3, or zebra fish embryos 0.3 mm3 are found in the size range of the first bars of both figures.
Figure 7.
Anuran oocyte volumes (mm3). (a) 805 species with oocyte diameters to 4.9 mm and volumes to 31.3 mm3. (b) 428 of these 805 species from the first two bars in the left figure with oocyte diameters ≤ 1.8 mm and volumes ≤ 5.0 mm3. The only successful cryopreservation of mammalian embryos 0.0002 to 0.004 mm3, or zebra fish embryos 0.3 mm3 are found in the size range of the first bars of both figures.
Table 2.
IUCN Red List recommendations for species in total for 423 Threatened Species for 3.4.2. Genome Resource Banking. 8th August 2023. [IUCN, 2023]. In brackets = IUCN Red List species number of species in IUCN Red List Category. Left = the number recommended for Genome Resource Banking. Right the % of species recommended for Genome Resource Banking. IUCN Red List Categories: LC = Least Concern, VU = Vulnerable, EN = Endangered, CE = Critically Endangered., DD = Data Deficient.
Table 2.
IUCN Red List recommendations for species in total for 423 Threatened Species for 3.4.2. Genome Resource Banking. 8th August 2023. [IUCN, 2023]. In brackets = IUCN Red List species number of species in IUCN Red List Category. Left = the number recommended for Genome Resource Banking. Right the % of species recommended for Genome Resource Banking. IUCN Red List Categories: LC = Least Concern, VU = Vulnerable, EN = Endangered, CE = Critically Endangered., DD = Data Deficient.
| LC | VU | EN | CE | DD | |
|---|---|---|---|---|---|
| Anura | 326 (3027) 10.7% | 42 (625) 6.7% | 32 (964) 3.3% | 23 (591) 4.0% | 1,000 |
| Salamanders | 13 (189) 6.9% | 17 (111) 1.5% | 17 (169) 10.1% | 8 (129) 6.2% | 48 |
| Caecilians | 0 (75) 0% | 0 (4) 0% | 0 (11) 0% | 0 (3) 0% | 97 |
| Total | 339 (3,291) 10.3% | 59 (740) 8.0% | 49 (1,144) 4.3% | 31 (723) 4.3% | 0 (1145) 0% |
Table 3.
IUCN Red List recommendations for 793 species in total for ex-situ Conservation - Captive breeding/artificial propagation. 8th August 2023 [IUCN, 2023]. In brackets = IUCN Red List species number of species in IUCN Red List Category. Left = the number recommended for Captive breeding/artificial propagation. Right the % of species recommended for Captive breeding/artificial propagation. IUCN Red List Categories: NT = Near Threatened, LC = Least Concern VU = Vulnerable, EN = Endangered, CE = Critically Endangered, DD = Data Deficient.
Table 3.
IUCN Red List recommendations for 793 species in total for ex-situ Conservation - Captive breeding/artificial propagation. 8th August 2023 [IUCN, 2023]. In brackets = IUCN Red List species number of species in IUCN Red List Category. Left = the number recommended for Captive breeding/artificial propagation. Right the % of species recommended for Captive breeding/artificial propagation. IUCN Red List Categories: NT = Near Threatened, LC = Least Concern VU = Vulnerable, EN = Endangered, CE = Critically Endangered, DD = Data Deficient.
| NT | LC | VU | EN | CE | DD | |
|---|---|---|---|---|---|---|
| Anura | 34 (341) | 294 (3,027) 10.7% | 42 (625) 6.7% | 64 (964) 3.3% | 217 (591) 4.0% | 209 |
| Salamanders | 0 (65) | 2 (189) 6.9% | 5 (111) 1.5% | 7 (169) 10.1% | 11 (129) 6.2% | 48 |
| Caecilians | na | 0 (75) 0% | 0 (4) 0% | 0 (11) 0% | 0 (3) 0% | 97 |
| Total | 34 (406) | 296 (3,291) 10.3% | 47 (740) 8.0% | 71 (1,144) 4.3% | 228 (723) 4.3% | na |
Table 4.
Species surveyed in a limited canvassing of private carers collections in 1998; their IUCN Red List status, number cared for / number reproduced and percentage reproduced. CE = Critically Endangered, EN = Endangered, VU = Vulnerable, NT = Near Threatened, LC = Least Concern. [from 15, Appendix A.1.].
Table 4.
Species surveyed in a limited canvassing of private carers collections in 1998; their IUCN Red List status, number cared for / number reproduced and percentage reproduced. CE = Critically Endangered, EN = Endangered, VU = Vulnerable, NT = Near Threatened, LC = Least Concern. [from 15, Appendix A.1.].
| Endangerment | CE | EN | VU | NT | LC |
|---|---|---|---|---|---|
| Anurans | 9 / 8, 89% | 16 / 16, 100% | 11 / 11, 100% | 15 / 11, 100% | 21 / 11, 100% |
| Salamanders | 6 / 6, 100% | 7 / 6, 86% | 10 / 11, 100% | 13 / 12, 92% | 44 / 42, 95% |
Table 5.
Examples of various estimates of necessary numbers of biobanked individuals for different established or recommended programs.
Table 5.
Examples of various estimates of necessary numbers of biobanked individuals for different established or recommended programs.
| Population | Males | Females | Application | Reference |
|---|---|---|---|---|
| Metapopulation? | 10 | [144] | ||
| Metapopulation | 5 | 5 | 27 Australian Threatened species. | [145] |
| Metapopulation | 20+ | 20+ | Conservation Breeding Programs | [110] |
| Fragmented/sub pop. | 10 | Every 4 years for CBPs depending on a species mean kinship | [146] | |
| Metapopulation | 15 unspecified sex | For CBP to prevent loss of genetic diversity and inbreeding depression. | [105,147,148] | |
| Metapopulation. Geographic clusters | 20 | 10 | Different geographic clusters | [149] |
Table 9.
IUCN Red List species number recommendations for Red List Category in brackets. Left of brackets the number and right of brackets, the % Biobanked in the San Diego Zoo Wildlife Alliance Frozen Zoo® living cell collection (as of April 2019). In brackets = IUCN Red List species number of species in IUCN Red List Category [83]. % species = % of amphibian species of all species in each category. % Biobanked = % of amphibian species as Biobanked species in each category. IUCN Red List Categories; VU = Vulnerable, EN = Endangered, CE = Critically Endangered, EX = Extinct in Wild.
Table 9.
IUCN Red List species number recommendations for Red List Category in brackets. Left of brackets the number and right of brackets, the % Biobanked in the San Diego Zoo Wildlife Alliance Frozen Zoo® living cell collection (as of April 2019). In brackets = IUCN Red List species number of species in IUCN Red List Category [83]. % species = % of amphibian species of all species in each category. % Biobanked = % of amphibian species as Biobanked species in each category. IUCN Red List Categories; VU = Vulnerable, EN = Endangered, CE = Critically Endangered, EX = Extinct in Wild.
| Class | VU | EN | CE | EW | EX |
|---|---|---|---|---|---|
| Amphibia | 2 (740) 0.2% | 4 (1,144) 0.3% | 3 (723) 0.4% | 1 (2) 50% | 0 (36/100*) 0% |
| Reptilia/Aves | 58 (1379) 4.2% | 27 (1,197) 2.3% | 32 (666) 4.8% | 3 (7) 60% | 0 (191) 0% |
| Mammalia | 80 (557) 14.4% | 66 (550) 12.0% | 34 (233) 14.6% | 2 (2) 100% | 85 (1) 1.2% |
| % species / % Biobanked | 27.6% / 1.4% | 40.7% / 4.1% | 44.5% / 4.3% | 9.0% / 10% | 34.0%*, 0% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



