Submitted:
08 October 2023
Posted:
09 October 2023
You are already at the latest version
Abstract
Reactive oxygen species (ROS) are a major risk factor for the development of oxidation and inflammation. Ryegrass (pasture), a natural dietary fiber source, acts as an antioxidant with well-known anti-inflammatory properties. The current study aimed to examine the alkaline phosphatase (ALP)-dependent Kelch-like-ECH-associated protein 1- Nuclear factor erythroid 2-related factor 2 (Keap1-Nrf2) signaling pathway that facilitates the anti-oxidative and anti-inflammatory properties of pasture and its protective effects on ileal permeability, systemic inflammation, nutrient absorption, liver dysfunction, growth performance, intestinal organ development, and metabolic syndrome in commercial diet-challenged geese. The lipopolysaccharides (LPS), ALP, ROS, and tight junction proteins were determined from the ileum tissues, and the antioxidant enzymes, immunoglobulins, and metabolic syndrome were determined from liver tissues via Enzyme-linked immunosorbent assay (ELISA) kits. While the Nrf2 and LPS/ROS/Keap1-induced inflammatory cytokines were determined by the quantitative reverse transcription polymerase chain reaction (RT-qPCR) method. The intestinal morphology was examined by Hematoxylin and Eosin (H&E) staining to determine nutrient absorption in ileal tissues. Pasture-dependent ALP activity significantly ameliorated commercial diet-induced LPS and ROS-facilitated ileal barrier dysfunction and systemic inflammation. The pasture supplementation induced nutrient absorption by influencing intestinal organ development. Our biochemical assays showed that the activation of Nrf2 significantly increased antioxidant enzymes and immunoglobulins, and decreased Keap1 and malondialdehyde (MDA) concentrations in the liver tissues after pasture supplementation. The correlation analysis of ALP, LPS, ROS, and Keap1-Nrf2 pathway-dependent antioxidant enzymes with other host markers showed that pasture supplementation induced intestinal barrier function via reducing LPS/ROS-induced intestinal permeability and systemic inflammation in the ileum and liver tissues. Pasture intake might attenuate LPS/ROS-induced gut permeability, systemic inflammation, and metabolic syndrome and augment gut barrier functions, nutrient absorption, immunity, and growth performance, possibly by its Keap1-Nrf2 pathway-dependent antioxidant properties.
Keywords:
Geese
; Pasture grazing
; Intestinal alkaline phosphatase
; Lipopolysaccharides
; Systemic inflammation
; Keap1-Nrf2
1. Introduction
The poultry’s health appears to be determined by intestinal homeostasis, which is altered by oxidative stress caused by either heat stress or feed stress [1]. Geese, ducks, broilers, quails, layers, pigeons, and turkeys are continuously exposed to lipopolysaccharides (LPS) through a variety of sources, including feed, water, and tiny dust particles, all of which are known to contain small levels of LPS. However, the diverse gram-negative bacterial community found in the intestines is the main natural source of LPS [2]. Gram-negative bacteria such as Escherichia coli [3], Cyanobacteria [4], and Bacterodales [5] produce LPS as a part of their outer membranes. Then the LPS undergoes ROS activation-induced systemic inflammation in geese [3]. Which further caused leaky gut-induced intestinal fluid loss, gut permeability, nutrient absorption, and diarrhea in geese and broilers [6].
In LPS-challenged animals, LPS activates ROS production and inflammatory responses, which are the main disease-causing elements in modern animal life [7]. Therefore, finding new modifiable feeding strategies that may encounter endogenous LPS-induced disease, causing oxidative stress and inflammation, can be helpful. Different feeding strategies, such as pectin [8], dietary fiber and threonine [9], and phytochemicals [10], have been applied to deal with LPS in different animal models.
The oral administration of intestinal alkaline phosphatase and different dietary fiber sources such as galactooligosaccharides, glucomannon, and ryegrass has been used to stimulate endogenous ALP activation in rats and geese [3,11]. Intestinal ALP is endogenously secreted from the gut epithelium and is said to regulate gut homeostasis. Loss of ALP activity is correlated with enhanced gut permeability, bacterial translocation, and systemic inflammation [12]. Intestinal ALP is believed to play a vital role in detoxifying LPS, flagellin, and CpG-DNA [13]. However, the studies supporting endogenous ALP regulation in geese to detoxify LPS-induced ROS production, gut permeability, nutrient absorption, and liver inflammation need to be understand.
In this situation, discovering a unique component with anti-oxidative and anti-inflammatory characteristics can be helpful. In light of this, ryegrass, a major source of dietary fiber, possesses a variety of intriguing biological features, such as antioxidant, anti-inflammatory, and antimicrobial properties [3,14].
Geese are herbivores, and due to their unique habit of using fresh grass, ryegrass was believed to improve their health and growth performance. Considering the earlier defined beneficial properties of ryegrass, we aimed to discover its defensive impacts against commercial diet (low dietary fiber source)-induced LPS/ROS-facilitated ileal and liver dysfunction. Furthermore, we investigated ryegrass-dependent ALP-targeted Keap1-Nrf2 signaling pathway-induced antioxidant enzymes, anti-inflammation, immunoglobulin activation, growth performance, intestinal organ development, and metabolic profiles in commercial diet-challenged geese.
2. Materials and Methods
2.1. Ethical approval
All animal experiments were approved by the Research Bioethics Committee of Henan Agricultural University (approval HENAU-2021).
2.2. Experimental animals
A total of 180, 25-day-old Wanfu geese were purchased from Henan Daidai Goose Agriculture and Animal Husbandry Development Co., Ltd. (Zhumadian, China). Geese with similar weight 693.6 g (±3.32) were evenly divided into two groups: (1) in-house feeding group (IHF, n = 90) and (2) artificial pasture grazing group (AGF, n = 90). Each of the six replicas of each group contained 15 geese. All geese have unrestricted access to fresh water and food. From 25 to 45 days, IHF geese received grower feed, and from 46 to 90 days, finisher feed (Table S1). The artificial pasture gazing system was developed in the form of a ryegrass supplement.
2.3. Sample collection
At the ages of 45d, 60d, and 90d, the geese were slaughtered using the Muslim method. Individual body weight gain and average feed consumption were assessed on a weekly basis. A simple method for measuring pasture intake was used by Cartoni et al. [15]. Livers were etched with phosphate-buffered saline, preserved in liquid nitrogen, and then stored at –80°C for further examination.
2.4. Determination of ALP, LPS, and ROS levels
Alkaline phosphatase, lipopolysaccharide, and reactive oxygen species were measured from ileal tissues. All kits were purchased from Shanghai Enzyme Link Biotechnology Co., Ltd. (Shanghai, China), and the experimental procedures were performed according to the manufacturer's instructions.
2.5. Measurement of gut permeability
The concentration of tight junction proteins is a widely recognized indicator called intestinal permeability. To this end, the concentrations of the tight junction proteins ZO-1, occludin, and claudin from ileal tissues were determined in this study. Kits were purchased from Shanghai Enzyme Link Biotechnology Co., Ltd. (Shanghai, China), and all experimental procedures were performed according to the manufacturer's instructions.
2.6. Determination of Antioxidant activity
Heme oxygenase 1 (HO-1) and glutathione reductase (GSR) in liver tissues were detected by ELISA kits (Shanghai Meilian Biotechnology Co., Ltd. Shanghai, China). Malondialdehyde (MDA), glutathione peroxidase (GSH-Px), total antioxidant capacity (T-AOC), total superoxide dismutase (T-SOD), and catalase (CAT) were determined from liver tissues using diagnostic kits (Nanjing Jiancheng Bioengineering Institute, Nanjing, Jiangsu, P.R. China) according to the manufacturer's instructions.
2.7. Measurement of metabolic (plasma lipid) profiles
Liver total cholesterol (T-CHO), low-density lipoprotein cholesterol (LDL-C), high-density lipoprotein cholesterol (HDL-C), triglycerides (TG), and blood urea nitrogen (BUN) were assessed using kits from Nanjing Jiancheng Bioengineering Company (Nanjing, Jiangsu, China) following the manufacturer's instructions.
2.8. Hematoxylin and Eosin (H&E) staining
First, the ileum tissues were fixed with 10% paraformaldehyde and stained with Harris’ hematoxylin solution at a temperature of 60-70°C for 6 h, and then rinsed in tap water until the water was colorless. Next, the tissues were separated twice with 10% acetic acid and 85% ethanol in water for 2 h and 10 h, respectively, and the tissues were rinsed with tap water again. In the bluing step, the tissues were soaked in saturated lithium carbonate solution for 12 h, then rinsed with tap water, and finally stained with eosin Y ethanol solution for 48 h.
2.9. RNA extraction and cDNA synthesis for RT-qPCR analysis
Total RNA from liver and ileal tissues (approximately 50 to 100 mg) was extracted using 1 mL of Trizol (MagZol reagent (#R4801-02; Magen Biotechnology, Guangzhou, Guangdong, China)) according to the manufacturer's instructions. The concentration and purity of total RNA were assessed using a NanoDrop 2000 UV-Vis spectrophotometer (Thermo Scientific, Wilmington, USA). Then, RNA was reverse transcribed into cDNA using HiScript® III RT SuperMix for qPCR (+gDNA wiper) (Vazyme, Nanjing, Jiangsu, P.R. China) according to the manufacturer's instructions. The content and purity of total cDNA were assessed using a NanoDrop 2000 UV-Vis spectrophotometer (Thermo Scientific, Wilmington, USA). The cDNA samples were subsequently amplified by real-time quantitative polymerase chain reaction using the ChamQ Universal SYBR qPCR Master Mix from Vazyme Biotechnology (Nanjing, Jiangsu, China). Gene-specific primers for each gene were generated using Primer3Web version 4.1.0 (Table S2). PCR was performed using ChamQ Universal SYBR qPCR Master Mix from Vazyme Biotechnology (Nanjing, Jiangsu, China) on a C1000 Touch PCR Thermal Cycler (BIO-RAD Laboratories, Shanghai, China) as follows: 40 cycles of 95°C for 15s and 60°C for 30s. Each measurement will be performed three times. Messenger ribonucleic acid (mRNA) expression of target genes relative to beta-actin (β-actin) was estimated using 2-ΔΔCT approach.
2.10. Statistical analysis
Raw data are presented as mean ± standard deviation. Statistical analysis was performed using SPSS 20.0 software (=D3 SPSS, Inc., 2009, Chicago, IL, USA www.spss.com). Differences between the two groups were found by Student's t-test, and P < 0.05 was considered statistically significant. Origin 2021 was used to make graphs, and Pearson correlation analysis was performed by OECloud tool (https://cloud.oebiotech.cn.) to evaluate the relationship between host markers.
3. Results
3.1. A commercial diet-dependent decline in ALP activity caused gut permeability and systemic inflammation
To examine whether ALP plays an important role in geese’s health, we determine the protein level of ALP from ileum tissues at different sampling stages, such as 45d, 60d, and 90d. We observed a significant reduction in ALP level in pasture-lacking geese from 45d to 90d (Figure 1A). As ALP is considered to be closely linked to regulating gut barrier functions, we determined the markers that are usually involved in deteriorating them. The increased concentration of ALP agonists such as LPS and ROS was associated with low dietary fiber source (commercial diet) intake in geese with age (Figures 1B and C). Furthermore, the tight junction proteins for commercial diet-fed geese and artificial pasture grazing geese were measured from the ileum tissues. Again, the decline in ALP was observed to be involved in reducing ZO-1, occludin, and claudin in pasture-deficient geese (Figures 1D-F). Intestinal hyperpermeability is considered to augment systemic inflammation. Thus, the pro-inflammatory cytokines were measured from the ileal tissues of geese. The systemic inflammatory-related markers were increased in 90-day-old geese compared with younger geese. Herein, the pasture supplementing geese showed lower mRNA levels of these markers than those of pasture-deficient geese (Figures 1G-K).
3.2. Commercial diet caused deterioration of nutrient absorption
After confirming ALP-deficient ileal dysbiosis in IHF geese, we sought to investigate intestinal morphology by H&E staining to determine nutrient absorption in ileal tissues. The effect of two feeding systems on the histomorphology of ileal tissues has been described in Figure 2 and Table 1. The highest villus height, villus width, villus surface area, and distance between villi were observed in pasture intake geese compared with commercial diet feeding geese at 45d, 60d, and 90d. The crypt depth and villus height to crypt depth ratio were improved in IHF geese despite in AGF geese. Furthermore, we determined the membrane thickness of ileal tissues in all sections (Figures 3 A and B). The relative thickness of the muscular tonic was higher in AGF geese than that of IHF geese (Table 2). Similarly, the relative thickness of the muscularis mucosa of AGF geese was greater than that of IHF geese from 45 days to 90 days (Figure 3B). These results suggest that pasture supplementation was effective in improving nutrient absorption by increasing villus surface area and the overall thickness of the wall of the ileum.
3.3. Commercial diet-dependent gut permeability is paralleled by liver inflammation
To find out the effects of diet-related IAP alterations in geese, we compared the mRNA levels of inflammatory markers from two feeding groups of geese. For liver tissues, qRT-PCR was applied. The mRNA levels of pro-inflammatory mediators such as iNOS and COX2 were increased in IHF geese compared with AGF geese at 45d, 60d, and 90d (Figure 4A and B). Furthermore, we determined the mRNA levels of inflammatory cytokines, including TNF-α, IL-6, and IL-1β from liver tissues (Figures 4C-E). Surprisingly, we found the pasture to be effective in decreasing them in geese.
3.4. Long-term pasture intake causes redox signaling pathway activation
In our previous research work, we proved that high dietary fiber source-dependent ROS production was beneficial in activating the Nrf2 signaling pathway by dissociating Nrf2 from Keap1 in the cecal tissues of meat geese [3]. Herein, we chose the liver tissues and hypothesized that pasture supplementation could regulate redox signaling Keap1-Nrf2 pathway-dependent antibodies to impede liver oxidation. In our study, the mRNA expression levels of Nrf2 were increased in pasture supplementing geese instead of Keap1 (Figures 5A and B). Further, we examined the antioxidants such as HO-1, GSH-PX, GSR, CAT, T-SOD, and T-AOC, which were increased in response to Nrf2 activation in AGF geese at 45d, 60d, and 90d (Figures 5C-H). To determine, whether these antioxidants could mimic the oxidation in the liver of geese, we measured the ROS mediator such as MDA from liver tissues. Surprisingly, the protein level of MDA was decreased by pasture supplementation in geese (Figure 5I). It has been described that the regulation of Nrf2 can cause changes in apoptosis and autoantibody production [16]. Therefore, we hypothesized that the activation of the Nrf2 signaling pathway could induce IgA, IgG, and IgM production in response to oxidative stress in live geese. Again, we found the pasture supplementation to be involved in augmenting these antibodies in geese compared with pasture-lacking geese at 45d, 60d, and 90d (Figures 5J-L).
3.5. Long-term pasture intake caused improved growth performance, intestinal organ development, and the metabolic profile of geese
The growth performance of the geese is shown in Figures 6A and B and Table 3. Geese (n = 190) at 25 days of age were randomly distributed to groups as follows: in-house feeding geese (IHF) and artificial pasture grazing geese (AGF). Assessment of geese growth performance during the grower phase (45d) and finisher phase (60d and 90d) by unpaired student T-test (Figure 6 and Table 3) showed that there was a significant difference in average daily feed intake (ADFI) (P = 1.62E-20, 7.42E-23, and 2.61E-26) and average daily gain (ADG) (P = 1.33E-14, 5.60E-16, and 4.99E-22, respectively). Moreover, the estimated pasture intake (66.09±1.03, 67.98±1.75, and 93.47±1.23 g DM/d, for 45d, 60d, and 90d, respectively) was observed to be helpful in increasing ADG during the finisher phase at 90d (82.3±0.09 g/d). Briefly, average body weight (ABW) and feed conversion ratio (FCR) were significantly improved during the grower phase (P = 2.48E-06 and 5.73E-21) and the finisher phase (P = 1.44E-07 and 0.000411, and 1.43E-17 and 1.11E-16, respectively).
The relative length of different sections of intestine at different growth stages, such as 45d, 60d, and 90d, are represented in Table 4. The relative lengths of the rectum, cecum, ileum, jejunum, duodenum, large intestine, small intestine, and total intestine of pasture supplementing geese were significantly greater than those of commercial diet-fed geese. Furthermore, the artificial pasture grazing system was seen to be effective in averting metabolic syndrome in geese with a significantly increased lipid profile of the liver and in reducing urea nitrogen levels (Figure 7).
3.6. Integrated analysis of host markers
Herein, the findings of our study demonstrate the Pearson correlation of ALP-dependent Keap1-Nrf2 signaling pathway-induced antioxidants with LPS/ROS-induced gut barrier dysfunctions, systemic inflammation, growth performance, and metabolic syndrome in Figure 8. As shown in Figures 8A-C, the ALP, Nrf2, HO-1, GSH-PX, GSR, CAT, T-SOD, and T-AOC were significantly positively correlated with HDL-C, tight junction proteins (ZO-1, occludin, and claudin), immunoglobulins (IgA, IgG, and IgM), the relative lengths of the rectum, cecum, ileum, jejunum, duodenum, large intestine, small intestine, and total intestine, and F:G at 45d, 60d, and 90d of sample collection. LPS, ROS, MDA, and Keap1 were significantly positively correlated with inflammatory cytokines (IL-1β, COX2, TNF-α, IL-6, and iNOS), metabolic syndrome (LDL-C, T-CHO, TG, and BUN), ABW, and ADG at 45d, 60d, and 90d of sample collection. In summary, the correlation analysis showed that the pasture intake significantly induced ALP and Nrf2-dependent antioxidant enzymes (except MDA) that strongly regulate gut barrier functions, immunity, intestinal organ development, HDL-C, and F:G and attenuate endotoxemia (LPS), oxidative stress, metabolic syndrome, and systemic inflammation in geese.
4. Discussion
Chronic low-grade inflammation in liver tissues may result from low dietary fiber diet-induced gut bacterial LPS-mediated ROS production [3]. Finding safe and effective ways to prevent the development of chronic low-grade inflammation is immediately required. The modern poultry feed formulation is built on grains with lower dietary fiber content, which may result in improved growth performance with compromised digestive improvements [17]. According to Huyghebaert et al. [18], these advancements are known to be harmful for visceral growth and to increase the risk of low immunity, intestinal infections, and metabolic illnesses. However, the relationship between shifts in dietary fiber contents and reducing the incidence of chronic low-grade inflammation by activating the ALP-dependent redox signaling mechanism in liver tissues remains to be explored in geese. Therefore, the present study demonstrates for the first time that a long-term pasture grazing system can ameliorate commercial diet-induced LPS/ROS-mediated gut permeability, intestinal and liver systemic inflammation, and metabolic syndrome by improving the ALP-induced redox signaling pathway.
The consumption of a diet high in fat, carbohydrates, calories, and protein may cause LPS-induced ROS synthesis [19,20]. Which may cause intestinal mucosal impairment and intestinal permeability [21]. It has been suggested that the decline in ALP activity with aging is paralleled by increased gut permeability and systemic inflammation in humans and mice [22]. However, the role of dietary fiber in inducing the ALP-dependent decline in gut permeability needs to be explored in geese. In our study, we first explained the impacts of gut permeability markers such as LPS and ROS in response to artificial pasture grazing (high dietary fiber source) and commercial diet feeding (low dietary fiber source) systems. Then we hypothesized that the decline in ALP activity (dependent on feeding systems) may augment ileal barrier functions in geese. Herein, the results of our study are inconsistent with those of Ali, et al., [3], in which pasture was effective in augmenting ALP activity in geese. It is well known that ALP dephosphorylates the adherence of LPS and ROS [23]. Here, we observed that pasture supplementation not only induces ALP activity but also reduces the harmful effects of LPS and ROS in pasture-supplementing geese [24].
Considering the key role of the intestinal barrier in maintaining the intestinal environment, disturbance of this barrier during low dietary fiber intake is related to a variety of chronic-related diseases, such as IBD, metabolic syndrome, Alzheimer’s disease, and osteoarthritis [25]. However, the exact role of the intestinal barrier during dietary fiber intake has been poorly studied. A few studies in geese and pigs have reported an increase in gut permeability with low dietary fiber intake [9,26]. In tissues from dietary fiber-deficient geese, a decrease in ZO-1, occluding, and claudin was found [3]. In in vitro studies, the upregulation of ALP in Caco-2 and T84 cells significantly increased the mRNA levels of ZO-1 and ZO-2 and ameliorated the LPS-induced intestinal permeability [27]. In this study, we hypothesized that pasture supplementation-dependent ALP activity could prevent gut hyperpermeability in geese. Interestingly, the loss of ALP activity in pasture-lacking geese was linked with increased ileal permeability and reduced protein levels of ZO-1, occluding, and claudin, while pasture supplementation resulted in increased ileal barrier functions and reduced endotoxin (LPS) and ROS concentrations in geese at 45d, 60d, and 90d of age. The possibility of restoring a fully operational intestinal epithelial barrier through pasture supplementation is a very appealing and nutritionally therapeutic intervention.
The dietary fibers that enhance the villus height/crypt depth ratio might also enhance the absorptive ability of the small intestinal epithelium, and vice versa [28]. In our study, pasture supplementation significantly increased the villus height, villus width, surface area, and distance between villi of the ileum of geese. However, the crypt depth and villus height to crypt depth ratio were decreased in AGF geese compared with IHF geese. It is said that the short villi may cause reduced absorption because of two circumstances. First, shortening may occur due to the loss of intestinal surface area, and second, the cells that are destroyed might be the mature cells. As nutrient absorption is essential for the osmotic absorption of water, while water absorption might be impaired because of compromised nutrient absorption. Additionally, the enhanced crypt depth is directly connected to enhanced water secretion. This is one of the primary mechanisms through which toxin-synthesizing bacteria such as Campylobacter jejuni [29], Salmonella [30], Brachyspira intermedia [31], and E. coli induce hypersecretory diarrhea. Furthermore, some authors have observed that multiple sources of dietary fiber may have a trophic impact on the small intestinal villi. As an example, in rats, the feeding of pectin (25 g/kg) for 14 days, in contrast to chicken, considerably improved the villus height and crypt depth ratio [32].
LPS has been considered to be closely linked to causing inflammation and several acute and chronic diseases [33]. Whereas ALP has the ability to dephosphorylate the toxic effects of LPS [34]. Pasture supplementation in geese led to significantly increased ALP and has the capacity to decrease endogenous LPS and ROS in ileal tissues. In addition to a decreased amount of LPS and ROS in AGF geese, we found significantly decreased systemic levels of pro-inflammatory cytokines such as iNOS, COX2, IL-6, IL-1β, and TNF-α. The levels of these cytokines have been correlated with inflammation in the ileum and liver tissues. In addition, these cytokines have been shown to exert harmful impacts on gut permeability by inducing gastrointestinal mucosal abnormalities [35,36]. We confirmed a significant decrease in iNOS, COX2, IL-6, IL-1β, and TNF-α in AGF geese, and we found them to be involved in ileal permeability and ileal and liver inflammation in geese lacking pasture supplementation.
In response to LPS/ROS-induced ileal permeability and inflammation, an immune response is activated by the redox-signaling Keap1-Nrf2 pathway. Upon ROS-induced oxidative stress, Keap1 dissociates from Nrf2 and activates Nrf2-targeted antioxidant enzymes [37]. Following the above studies, the ileal ALP-dependent Nrf2 and its targeted enzymes such as HO-1, GSH-PX, GSR, CAT, T-SOD, and T-AOC were augmented in pasture supplementing geese at 45d, 60d, and 90d. Malondialdehyde (MDA) is considered a weapon for oxidative stress that causes liver disorders and generates liver fibrosis, non-lkohalic fatty liver disease, hepatitis C, etc., [38]. Furthermore, MDA deteriorated the quality of broiler meat by destroying muscle mitochondrial protein and lipid [39]. Luckily, we found that the pasture supplementation was effective in reducing DMA levels in liver tissues, thus protecting the geese from fatty liver diseases. It is said that the endogenous regulation of Nrf2-targeted enzymes can stimulate adaptive immunity through immunoglobulin production [16]. Herein, with increasing antioxidant enzyme levels, the IgA, IgG, and IgM protein levels were enhanced in pasture supplementing geese, suggesting that Nrf2 may be involved in regulating adaptive immunity in geese.
Geese like to eat grass, and due to their distinctive behavior of using high-fiber feeds, ryegrass was considered to promote growth performance, intestinal organ development, and the metabolic profile of the liver. The chemical composition of feed was differentiated on the basis of crude protein (%), crude fat (%), ash (%), dry matter (%), moisture (%), neutral detergent fiber (%), acid detergent fiber (%), calcium (%), and phosphorous (%). The overall feed intake (commercial diet plus grass (g DM/d)) of AGF geese was significantly higher compared with IHF geese. However, the average body weight of IHF geese was significantly greater than that of AGF geese at 45d, 60d, and 90d. It’s interesting to note that the average daily gain (g/d) of AGF geese was significantly improved at 90d compared with IHF geese. It might be due to pasture intake that impedes the harmful impacts of oxidative stress (i.e., caused by heat stress or feed stress) in geese. Furthermore, the feed to gain ratio (or feed conversion ratio) was significantly improved in AGF geese compared with those of IHF geese at 45d, 60d, and 90d. Following the reports of Ling et al., [40] and J. Chen et al., [41], the dietary fiber present in the ryegrass was sufficient to improve ADFI, ADG, and F:G in geese.
Intestinal organ development describes nutrient digestibility and might be dependent on the F:G of the whole body. Therefore, in this study, the development of intestinal organs was measured, which included the rectum, cecum, ileum, jejunum, duodenum, large intestine, small intestine, and total intestine. In the current study, pasture supplementation had significant effects on the relative lengths of the above-mentioned intestinal organs, which were supported by reports that a high dietary fiber diet mainly modified the relative lengths of the intestines of adult genders, geese, and broilers [40,42,43].
Liver biochemical indexes are considered important in determining the metabolism and health status of geese. The concentrations of HDL-C, LDL-C, and T-CHO reflect the levels of lipid metabolism. In a few studies, the 11% accumulation of crude fiber showed improved blood lipid metabolism in broilers [40]. In the present study, a sufficient amount of pasture supplementation was effective in reducing T-CHO and LDL-C and inducing HDL-C in the liver tissues of geese. Additionally, triglycerides are stored in the liver, where they are confined in cytoplasmic lipid droplets. Its higher quantity in the liver causes NAFLD, dyslipidemia, obesity, and type 2 diabetes [44]. Fortunately, our results were different from those of studies in which pasture supplementation restricted the higher accumulation of TG in the liver of geese. Urea nitrogen is a waste product of protein that is taken in the form of food. It is stored in the liver and then travels through the blood to the kidneys, where it is then filtered out of the blood. Urea nitrogen describes how well the kidneys are working. Few studies regarding dietary fiber describe the functions of blood urea nitrogen [45]. Following the previous studies, the dietary fiber present in the pasture was sufficient to control the working of the kidneys by ameliorating excess BUN production in the liver of geese [46].
Lastly, we described the Pearson correlation analysis of ALP-dependent Keap1-Nrf2 signaling pathway-induced antioxidants with LPS/ROS-induced gut barrier dysfunctions, systemic inflammation, growth performance, and metabolic syndrome. The results of this correlation analysis showed that the pasture intake significantly induced ALP and Nrf2-dependent antioxidant enzymes (except MDA) that strongly regulate gut barrier functions, immunity, intestinal organ development, HDL-C, and F:G and attenuate endotoxemia (LPS), oxidative stress, metabolic syndrome, and systemic inflammation in geese.
5. Conclusions
In conclusion, artificial pasture grazing system-induced alkaline phosphatase production seems to restore intestinal health by targeting LPS/ROS-induced intestinal barrier dysfunction, systemic inflammation, nutrient absorption, and metabolic syndrome. The mechanisms behind its action may be related to Keap1-Nrf2 signaling pathway-induced antioxidant enzymes, immunoglobulins, growth performance, and lipid metabolism. The beneficial effects of artificial pasture grazing systems as an alternative source of dietary fiber may help to understand gut barrier function-related diseases in animals.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Acknowledgments
This work was supported by grants from the Modern Agroindustry Technology Research System of China (CARS-34) and the Science and Technology Innovation Team of Henan Province High Quality Forage and Animal Health (No.22IRTSTHN022).
Conflicts of Interest
No potential conflicts of interest were disclosed.
References
- Obianwuna, U.E.; Agbai Kalu, N.; Wang, J.; Zhang, H.; Qi, G.; Qiu, K.; Wu, S. Recent Trends on Mitigative Effect of Probiotics on Oxidative-Stress-Induced Gut Dysfunction in Broilers under Necrotic Enteritis Challenge: A Review. Antioxidants (Basel, Switzerland) 2023, 12. [Google Scholar] [CrossRef]
- Romano, K.P.; Hung, D.T. Targeting LPS biosynthesis and transport in gram-negative bacteria in the era of multi-drug resistance. Biochimica et biophysica acta. Molecular cell research 2023, 1870, 119407. [Google Scholar] [CrossRef]
- Ali, Q.; Ma, S.; Farooq, U.; Niu, J.; Li, F.; Li, D.; Wang, Z.; Sun, H.; Cui, Y.; Shi, Y. Pasture intake protects against commercial diet-induced lipopolysaccharide production facilitated by gut microbiota through activating intestinal alkaline phosphatase enzyme in meat geese. Frontiers in immunology 2022, 13, 1041070. [Google Scholar] [CrossRef] [PubMed]
- Volk, A.; Lee, J. Cyanobacterial blooms: A player in the freshwater environmental resistome with public health relevance? Environmental research 2023, 216, 114612. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Ali, A.; Kapardar, R.K.; Dar, G.M.; Nimisha; Apurva; Sharma, A. K.; Verma, R.; Sattar, R.S.A.; Ahmad, E.; et al. Implication of gut microbes and its metabolites in colorectal cancer. Journal of cancer research and clinical oncology 2023, 149, 441–465. [Google Scholar] [CrossRef] [PubMed]
- Pham, V.H.; Abbas, W.; Huang, J.; Guo, F.; Zhang, K.; Kong, L.; Zhen, W.; Guo, Y.; Wang, Z. Dietary coated essential oil and organic acid mixture supplementation improves health of broilers infected with avian pathogenic Escherichia coli. Animal nutrition (Zhongguo xu mu shou yi xue hui) 2023, 12, 245–262. [Google Scholar] [CrossRef] [PubMed]
- Moradi Vastegani, S.; Hajipour, S.; Sarkaki, A.; Basir, Z.; Parisa Navabi, S.; Farbood, Y.; Khoshnam, S.E. Curcumin mitigates lipopolysaccharide-induced anxiety/depression-like behaviors, blood-brain barrier dysfunction and brain edema by decreasing cerebral oxidative stress in male rats. Neuroscience letters 2022, 782, 136697. [Google Scholar] [CrossRef] [PubMed]
- Sahasrabudhe, N.M.; Beukema, M.; Tian, L.; Troost, B.; Scholte, J.; Bruininx, E.; Bruggeman, G.; van den Berg, M.; Scheurink, A.; Schols, H.A.; et al. Dietary Fiber Pectin Directly Blocks Toll-Like Receptor 2-1 and Prevents Doxorubicin-Induced Ileitis. Frontiers in immunology 2018, 9, 383. [Google Scholar] [CrossRef] [PubMed]
- Wellington, M.O.; Hamonic, K.; Krone, J.E.C.; Htoo, J.K.; Van Kessel, A.G.; Columbus, D.A. Effect of dietary fiber and threonine content on intestinal barrier function in pigs challenged with either systemic E. coli lipopolysaccharide or enteric Salmonella Typhimurium. Journal of animal science and biotechnology 2020, 11, 38. [Google Scholar] [CrossRef]
- Chauhan, A.; Islam, A.U.; Prakash, H.; Singh, S. Phytochemicals targeting NF-κB signaling: Potential anti-cancer interventions. Journal of pharmaceutical analysis 2022, 12, 394–405. [Google Scholar] [CrossRef]
- Okazaki, Y.; Katayama, T. Glucomannan consumption elevates colonic alkaline phosphatase activity by up-regulating the expression of IAP-I, which is associated with increased production of protective factors for gut epithelial homeostasis in high-fat diet-fed rats. Nutrition research (New York, N.Y.) 2017, 43, 43–50. [Google Scholar] [CrossRef]
- Fawley, J.; Gourlay, D.M. Intestinal alkaline phosphatase: a summary of its role in clinical disease. The Journal of surgical research 2016, 202, 225–234. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.S.; Gehr, T.W.; Ghosh, S. Curcumin and chronic kidney disease (CKD): major mode of action through stimulating endogenous intestinal alkaline phosphatase. Molecules (Basel, Switzerland) 2014, 19, 20139–20156. [Google Scholar] [CrossRef]
- Escobar-Correas, S.; Broadbent, J.A.; Andraszek, A.; Stockwell, S.; Howitt, C.A.; Juhász, A.; Colgrave, M.L. Perennial Ryegrass Contains Gluten-Like Proteins That Could Contaminate Cereal Crops. Frontiers in nutrition 2021, 8, 708122. [Google Scholar] [CrossRef] [PubMed]
- Cartoni Mancinelli, A.; Mattioli, S.; Dal Bosco, A.; Piottoli, L.; Ranucci, D.; Branciari, R.; Cotozzolo, E.; Castellini, C. Rearing Romagnola geese in vineyard: Pasture and antioxidant intake, performance, carcass and meat quality. Italian Journal of Animal Science 2019, 18, 372–380. [Google Scholar] [CrossRef]
- Barati, M.T.; Caster, D.J. The potential of Nrf2 activation as a therapeutic target in systemic lupus erythematosus. Metabolites 2022, 12, 151. [Google Scholar] [CrossRef] [PubMed]
- Salvatore, S.; Battigaglia, M.S.; Murone, E.; Dozio, E.; Pensabene, L.; Agosti, M. Dietary Fibers in Healthy Children and in Pediatric Gastrointestinal Disorders: A Practical Guide. Nutrients 2023, 15. [Google Scholar] [CrossRef] [PubMed]
- Huyghebaert, G.; Ducatelle, R.; Van Immerseel, F. An update on alternatives to antimicrobial growth promoters for broilers. Veterinary journal (London, England : 1997) 2011, 187, 182–188. [Google Scholar] [CrossRef]
- Lu, Y.C.; Yeh, W.C.; Ohashi, P.S. LPS/TLR4 signal transduction pathway. Cytokine 2008, 42, 145–151. [Google Scholar] [CrossRef]
- Cui, J.; Chen, Y.; Wang, H.Y.; Wang, R.F. Mechanisms and pathways of innate immune activation and regulation in health and cancer. Human vaccines & immunotherapeutics 2014, 10, 3270–3285. [Google Scholar] [CrossRef]
- Guo, S.; Al-Sadi, R.; Said, H.M.; Ma, T.Y. Lipopolysaccharide causes an increase in intestinal tight junction permeability in vitro and in vivo by inducing enterocyte membrane expression and localization of TLR-4 and CD14. The American journal of pathology 2013, 182, 375–387. [Google Scholar] [CrossRef] [PubMed]
- Kühn, F.; Adiliaghdam, F.; Cavallaro, P.M.; Hamarneh, S.R.; Tsurumi, A.; Hoda, R.S.; Munoz, A.R.; Dhole, Y.; Ramirez, J.M.; Liu, E.; et al. Intestinal alkaline phosphatase targets the gut barrier to prevent aging. JCI insight 2020, 5. [Google Scholar] [CrossRef] [PubMed]
- Koyama, I.; Matsunaga, T.; Harada, T.; Hokari, S.; Komoda, T. Alkaline phosphatases reduce toxicity of lipopolysaccharides in vivo and in vitro through dephosphorylation. Clinical Biochemistry 2002, 35, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.H.; Lim, B.S.; Lee, Y.K.; Yang, H.C. Effects of hydrogen peroxide (H2O2) on alkaline phosphatase activity and matrix mineralization of odontoblast and osteoblast cell lines. Cell biology and toxicology 2006, 22, 39–46. [Google Scholar] [CrossRef]
- Hills, R.D., Jr.; Pontefract, B.A.; Mishcon, H.R.; Black, C.A.; Sutton, S.C.; Theberge, C.R. Gut Microbiome: Profound Implications for Diet and Disease. Nutrients 2019, 11. [Google Scholar] [CrossRef] [PubMed]
- Xi, Y.; Huang, Y.; Li, Y.; Huang, Y.; Yan, J.; Shi, Z. The effects of dietary protein and fiber levels on growth performance, gout occurrence, intestinal microbial communities, and immunoregulation in the gut-kidney axis of goslings. Poultry science 2022, 101, 101780. [Google Scholar] [CrossRef]
- Liu, W.; Hu, D.; Huo, H.; Zhang, W.; Adiliaghdam, F.; Morrison, S.; Ramirez, J.M.; Gul, S.S.; Hamarneh, S.R.; Hodin, R.A. Intestinal Alkaline Phosphatase Regulates Tight Junction Protein Levels. Journal of the American College of Surgeons 2016, 222, 1009–1017. [Google Scholar] [CrossRef]
- Ali, Q.; Ma, S.; La, S.; Guo, Z.; Liu, B.; Gao, Z.; Farooq, U.; Wang, Z.; Zhu, X.; Cui, Y.; et al. Microbial short-chain fatty acids: a bridge between dietary fibers and poultry gut health - A review. Animal bioscience 2022, 35, 1461–1478. [Google Scholar] [CrossRef]
- Alemka, A.; Whelan, S.; Gough, R.; Clyne, M.; Gallagher, M.E.; Carrington, S.D.; Bourke, B. Purified chicken intestinal mucin attenuates Campylobacter jejuni pathogenicity in vitro. Journal of medical microbiology 2010, 59, 898–903. [Google Scholar] [CrossRef]
- Pourabedin, M.; Chen, Q.; Yang, M.; Zhao, X. Mannan-and xylooligosaccharides modulate caecal microbiota and expression of inflammatory-related cytokines and reduce caecal Salmonella Enteritidis colonisation in young chickens. FEMS microbiology ecology 2017, 93, fiw226. [Google Scholar] [CrossRef]
- Mappley, L.J.; La Ragione, R.M.; Woodward, M.J. Brachyspira and its role in avian intestinal spirochaetosis. Veterinary microbiology 2014, 168, 245–260. [Google Scholar] [CrossRef] [PubMed]
- McNabney, S.M.; Henagan, T.M. Short chain fatty acids in the colon and peripheral tissues: a focus on butyrate, colon cancer, obesity and insulin resistance. Nutrients 2017, 9, 1348. [Google Scholar] [CrossRef]
- Guerville, M.; Boudry, G. Gastrointestinal and hepatic mechanisms limiting entry and dissemination of lipopolysaccharide into the systemic circulation. American journal of physiology. Gastrointestinal and liver physiology 2016, 311, G1–g15. [Google Scholar] [CrossRef] [PubMed]
- Estaki, M.; DeCoffe, D.; Gibson, D.L. Interplay between intestinal alkaline phosphatase, diet, gut microbes and immunity. World journal of gastroenterology 2014, 20, 15650–15656. [Google Scholar] [CrossRef] [PubMed]
- Simmons, D.L.; Botting, R.M.; Hla, T. Cyclooxygenase isozymes: the biology of prostaglandin synthesis and inhibition. Pharmacological reviews 2004, 56, 387–437. [Google Scholar] [CrossRef]
- Varadhan, R.; Yao, W.; Matteini, A.; Beamer, B.A.; Xue, Q.L.; Yang, H.; Manwani, B.; Reiner, A.; Jenny, N.; Parekh, N.; et al. Simple biologically informed inflammatory index of two serum cytokines predicts 10 year all-cause mortality in older adults. The journals of gerontology. Series A, Biological sciences and medical sciences 2014, 69, 165–173. [Google Scholar] [CrossRef]
- Horie, Y.; Suzuki, T.; Inoue, J.; Iso, T.; Wells, G.; Moore, T.W.; Mizushima, T.; Dinkova-Kostova, A.T.; Kasai, T.; Kamei, T.; et al. Molecular basis for the disruption of Keap1–Nrf2 interaction via Hinge & Latch mechanism. Communications Biology 2021, 4, 576. [Google Scholar] [CrossRef]
- Arroyave-Ospina, J.C.; Wu, Z.; Geng, Y.; Moshage, H. Role of Oxidative Stress in the Pathogenesis of Non-Alcoholic Fatty Liver Disease: Implications for Prevention and Therapy. Antioxidants (Basel, Switzerland) 2021, 10. [Google Scholar] [CrossRef]
- Chen, Z.; Xing, T.; Li, J.; Zhang, L.; Jiang, Y.; Gao, F. Oxidative stress impairs the meat quality of broiler by damaging mitochondrial function, affecting calcium metabolism and leading to ferroptosis. Animal bioscience 2022, 35, 1616–1627. [Google Scholar] [CrossRef]
- Ling, J.; GAO, Y.-y.; Hui, Y.; WANG, W.-c.; LIN, Z.-p.; YANG, H.-y.; HUANG, S.-b.; Lin, Y. Effects of dietary fiber and grit on performance, gastrointestinal tract development, lipometabolism, and grit retention of goslings. Journal of Integrative Agriculture 2014, 13, 2731–2740. [Google Scholar]
- Chen, J.; Weng, K.; Liu, J.; Gu, W.; Luo, S.; Zheng, M.; Cao, Z.; Zhang, Y.; Zhang, Y.; Chen, G. Effect of different free-range systems on the growth performance, carcass traits, and meat quality of Yangzhou geese. Animal Biotechnology 2022, 1–7. [Google Scholar] [CrossRef] [PubMed]
- He, C.; Wu, H.; Lv, Y.; You, H.; Zha, L.; Li, Q.; Huang, Y.; Tian, J.; Chen, Q.; Shen, Y. Gastrointestinal development and microbiota responses of geese to honeycomb flavonoids supplementation. Frontiers in Veterinary Science 2021, 8, 739237. [Google Scholar] [CrossRef] [PubMed]
- Zheng, M.; Mao, P.; Tian, X.; Meng, L. Effects of grazing mixed-grass pastures on growth performance, immune responses, and intestinal microbiota in free-range Beijing-you chickens. Poultry science 2021, 100, 1049–1058. [Google Scholar] [CrossRef] [PubMed]
- Tomizawa, M.; Kawanabe, Y.; Shinozaki, F.; Sato, S.; Motoyoshi, Y.; Sugiyama, T.; Yamamoto, S.; Sueishi, M. Triglyceride is strongly associated with nonalcoholic fatty liver disease among markers of hyperlipidemia and diabetes. Biomedical reports 2014, 2, 633–636. [Google Scholar] [CrossRef]
- Yang, H.L.; Feng, P.; Xu, Y.; Hou, Y.Y.; Ojo, O.; Wang, X.H. The Role of Dietary Fiber Supplementation in Regulating Uremic Toxins in Patients With Chronic Kidney Disease: A Meta-Analysis of Randomized Controlled Trials. Journal of renal nutrition : the official journal of the Council on Renal Nutrition of the National Kidney Foundation 2021, 31, 438–447. [Google Scholar] [CrossRef]
- Yu, J.; Yang, Z.; Yang, H.; Wang, Z. Effects of cottonseed meal on growth performance, liver redox status, and serum biochemical parameters in goslings at 1 to 28 days of age. BMC Veterinary Research 2022, 18, 347. [Google Scholar] [CrossRef]
Figure 1.
A commercial diet-dependent decline in ALP activity caused gut permeability and systemic inflammation. (A) ALP protein level in ileal tissues, (B) LPS protein level in ileal tissues, (C) ROS concentration from ileal tissues, (D-F) and ileal tight junction proteins ZO-1, occludin, and claudin, measured by ELISA kits. (G) COX2 mRNA level in ileal tissues, (H) iNOS mRNA level in ileal tissues, (I) IL-6 mRNA level in ileal tissues, (J) IL-1β mRNA level in ileal tissues, and (K) TNF-α mRNA level in ileal tissues, normalized by β-actin and measured by qPCR. In-house feeding system (IHF) and artificial pasture grazing system (AGF). Data with different superscript letters are significantly different (P < 0.05) according to the unpaired student T-Test. The asterisks symbol indicates significant differences *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001.
Figure 1.
A commercial diet-dependent decline in ALP activity caused gut permeability and systemic inflammation. (A) ALP protein level in ileal tissues, (B) LPS protein level in ileal tissues, (C) ROS concentration from ileal tissues, (D-F) and ileal tight junction proteins ZO-1, occludin, and claudin, measured by ELISA kits. (G) COX2 mRNA level in ileal tissues, (H) iNOS mRNA level in ileal tissues, (I) IL-6 mRNA level in ileal tissues, (J) IL-1β mRNA level in ileal tissues, and (K) TNF-α mRNA level in ileal tissues, normalized by β-actin and measured by qPCR. In-house feeding system (IHF) and artificial pasture grazing system (AGF). Data with different superscript letters are significantly different (P < 0.05) according to the unpaired student T-Test. The asterisks symbol indicates significant differences *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001.
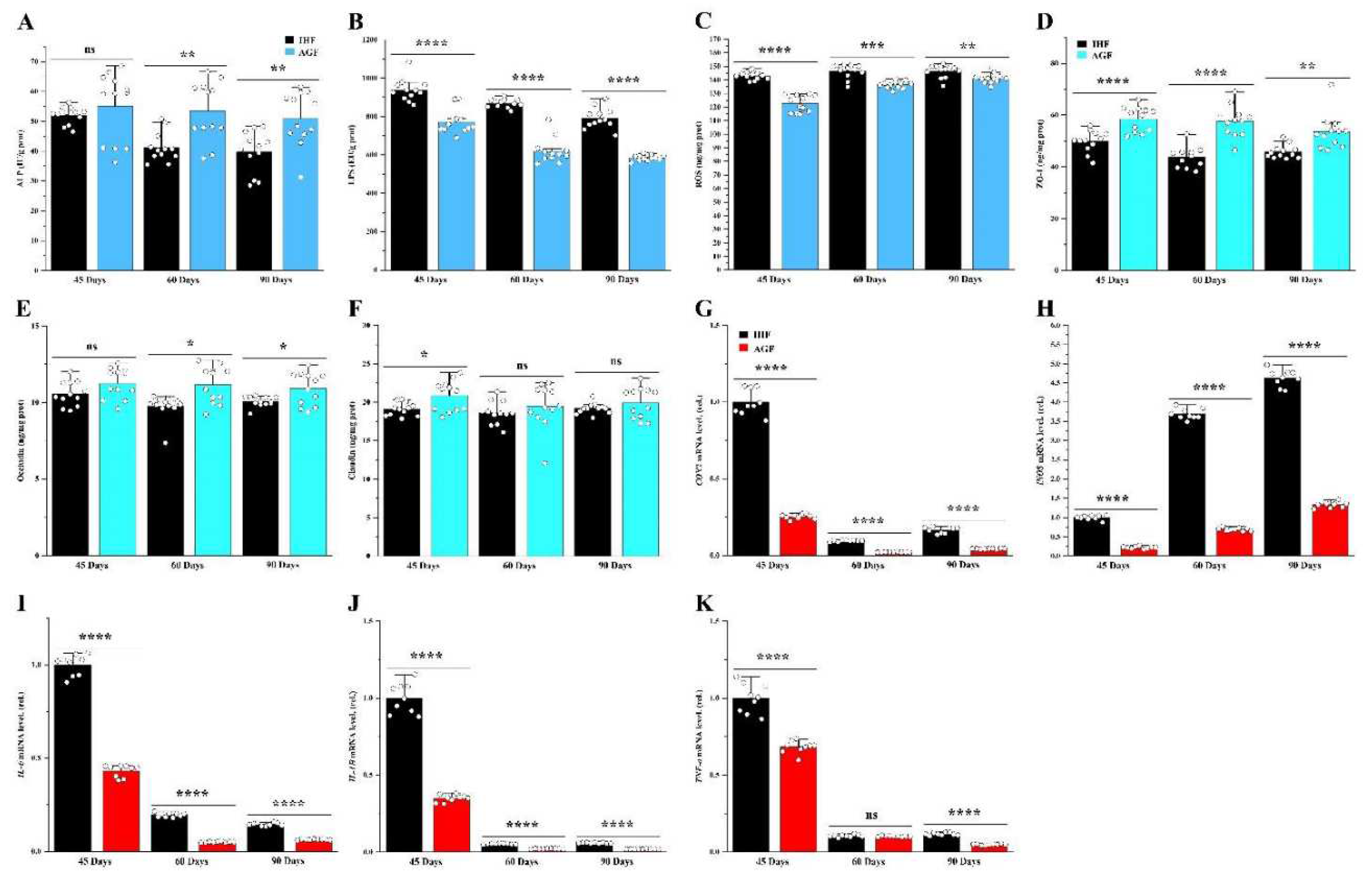
Figure 2.
Commercial diet cause deterioration of nutrient absorption. VH – villus height; VW – villus width; DBV – distance between two villi; CD – crypt depth. In-house feeding system (IHF) and artificial pasture grazing system (AGF).
Figure 2.
Commercial diet cause deterioration of nutrient absorption. VH – villus height; VW – villus width; DBV – distance between two villi; CD – crypt depth. In-house feeding system (IHF) and artificial pasture grazing system (AGF).

Figure 3.
Commercial diet cause deterioration of nutrient absorption by effecting thickness of ileal muscular tonic and muscularis mucosa (50µm). (A) Light micrograph of the wall of ileum of geese, hematoxylin and eosin: 1 – outer layer of muscular tonic; 2 – inner layer of muscular tonic; 3 – outer layer of lamina muscularis mucosa (LMM); 4 – ganglion of submucosal nerve plexus; 5 – inner layer of lamina muscularis mucosa (LMM); 6 – crypts; 7 – pillars of unstriated muscle cells (between crypts). (B) Comparison of the cecal membrane thickness of geese with different feeding systems (50µm). In-house feeding system (IHF) and artificial pasture grazing system (AGF).
Figure 3.
Commercial diet cause deterioration of nutrient absorption by effecting thickness of ileal muscular tonic and muscularis mucosa (50µm). (A) Light micrograph of the wall of ileum of geese, hematoxylin and eosin: 1 – outer layer of muscular tonic; 2 – inner layer of muscular tonic; 3 – outer layer of lamina muscularis mucosa (LMM); 4 – ganglion of submucosal nerve plexus; 5 – inner layer of lamina muscularis mucosa (LMM); 6 – crypts; 7 – pillars of unstriated muscle cells (between crypts). (B) Comparison of the cecal membrane thickness of geese with different feeding systems (50µm). In-house feeding system (IHF) and artificial pasture grazing system (AGF).
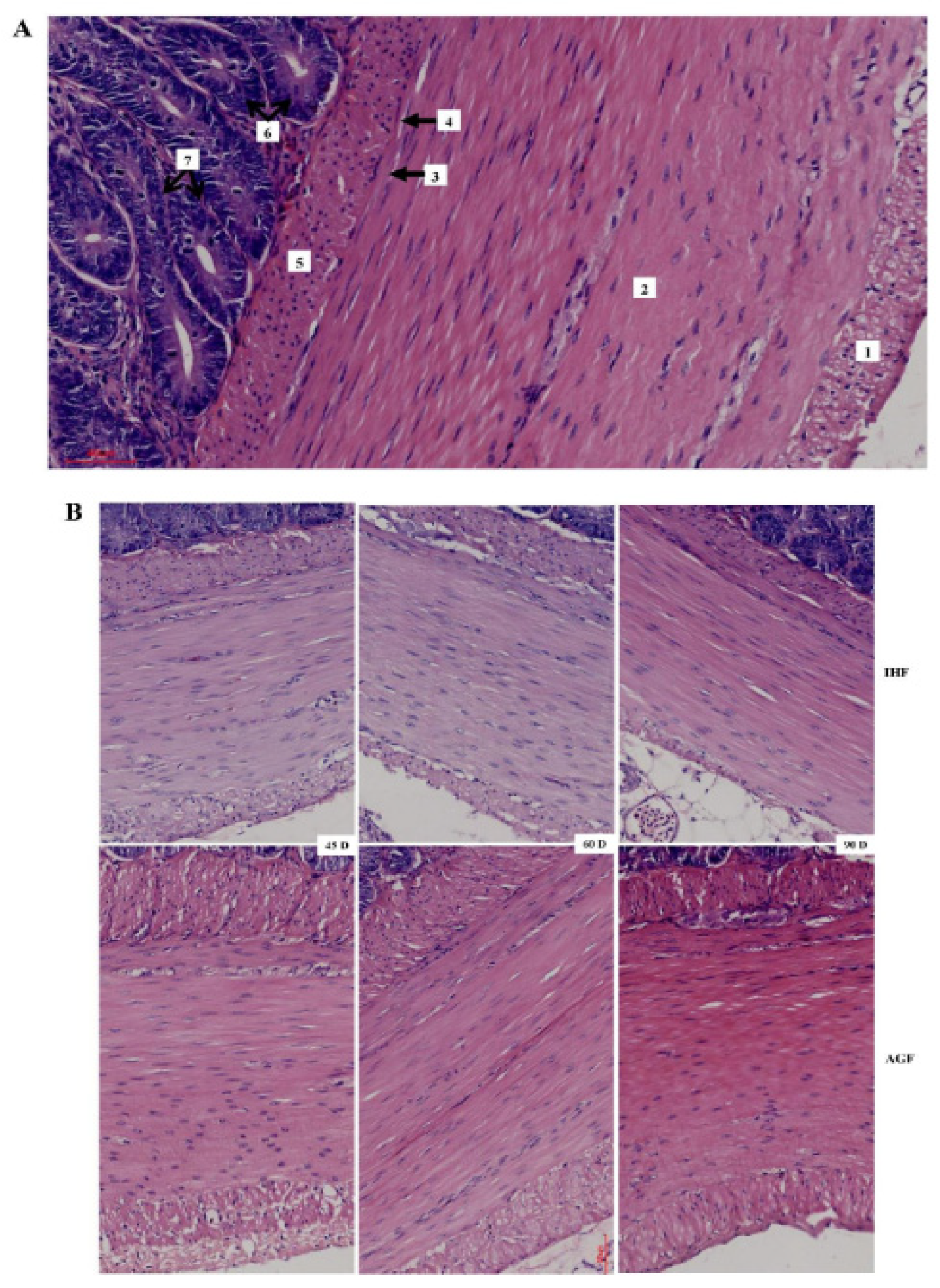
Figure 4.
Commercial diet-dependent gut permeability is paralleled by liver inflammation. (A)COX2 mRNA level in liver tissues, (B) iNOS mRNA level in liver tissues, (C) IL-6 mRNA level in liver tissues, (D) IL-1β mRNA level in liver tissues, and (E) TNF-α mRNA level in liver tissues, normalized by β-actin and measured by qPCR. In-house feeding system (IHF) and artificial pasture grazing system (AGF). Data with different superscript letters are significantly different (P < 0.05) according to the unpaired student T-Test. The asterisks symbol indicates significant differences *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001.
Figure 4.
Commercial diet-dependent gut permeability is paralleled by liver inflammation. (A)COX2 mRNA level in liver tissues, (B) iNOS mRNA level in liver tissues, (C) IL-6 mRNA level in liver tissues, (D) IL-1β mRNA level in liver tissues, and (E) TNF-α mRNA level in liver tissues, normalized by β-actin and measured by qPCR. In-house feeding system (IHF) and artificial pasture grazing system (AGF). Data with different superscript letters are significantly different (P < 0.05) according to the unpaired student T-Test. The asterisks symbol indicates significant differences *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001.
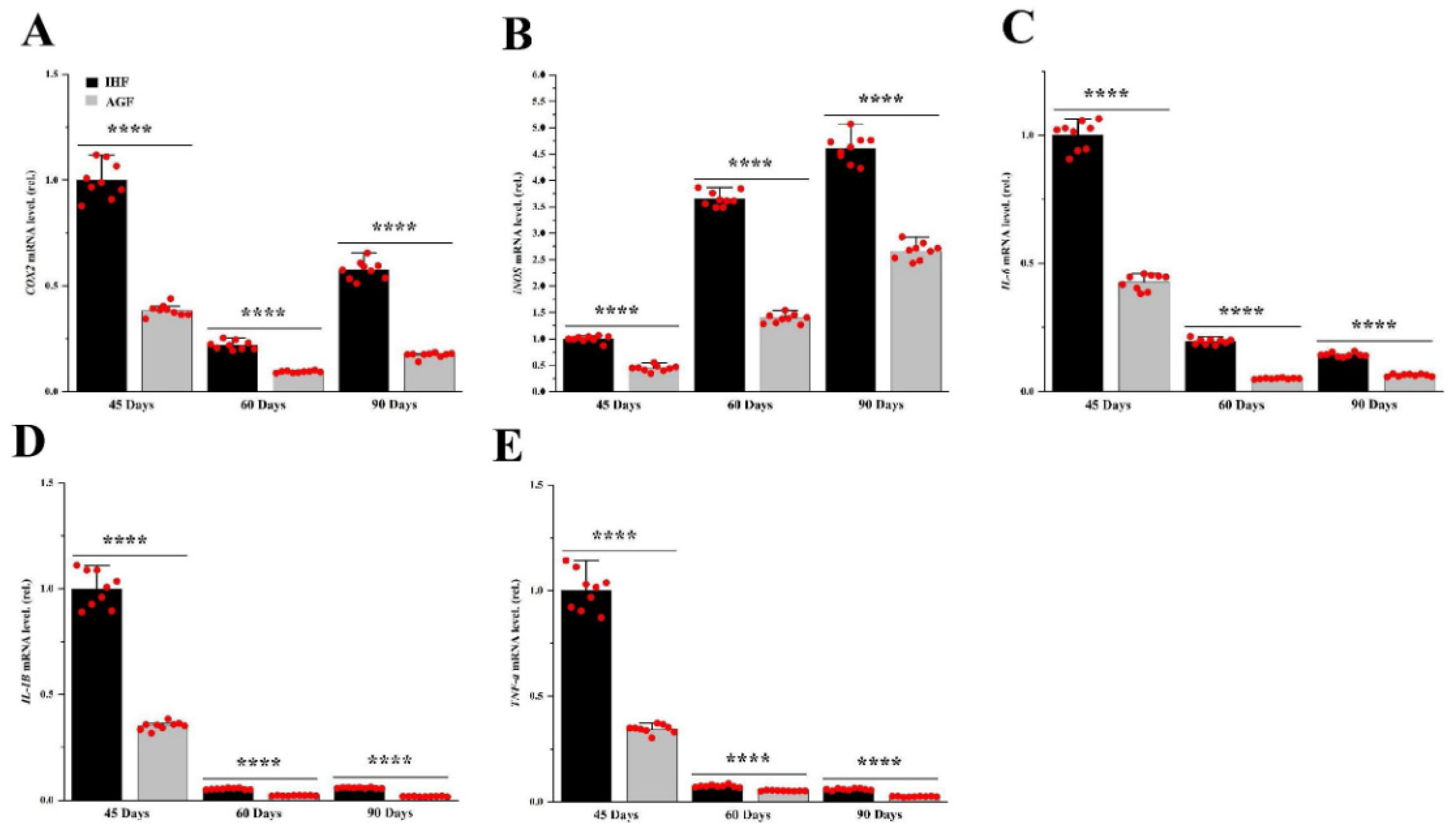
Figure 5.
Long-term pasture intake causes redox signaling pathway activation. (A)Keap1 mRNA level in liver tissues and (B) Nrf2 mRNA level in liver tissues, normalized by β-actin and measured by qPCR. (C) HO-1 protein level in liver tissues, (D) GSH-PX protein level in liver tissues, (E) GSR protein level in liver tissues, (F) CAT protein level in liver tissues, (G) T-SOD protein level in liver tissues, (H) T-AOC protein level in liver tissues, (I) MDA protein level in liver tissues, (J) IgA protein level in liver tissues, (K) IgG protein level in liver tissues, and (L) IgM protein level in liver tissues, measured by ELISA kits. In-house feeding system (IHF) and artificial pasture grazing system (AGF). Data with different superscript letters are significantly different (P < 0.05) according to the unpaired student T-Test. The asterisks symbol indicates significant differences *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001.
Figure 5.
Long-term pasture intake causes redox signaling pathway activation. (A)Keap1 mRNA level in liver tissues and (B) Nrf2 mRNA level in liver tissues, normalized by β-actin and measured by qPCR. (C) HO-1 protein level in liver tissues, (D) GSH-PX protein level in liver tissues, (E) GSR protein level in liver tissues, (F) CAT protein level in liver tissues, (G) T-SOD protein level in liver tissues, (H) T-AOC protein level in liver tissues, (I) MDA protein level in liver tissues, (J) IgA protein level in liver tissues, (K) IgG protein level in liver tissues, and (L) IgM protein level in liver tissues, measured by ELISA kits. In-house feeding system (IHF) and artificial pasture grazing system (AGF). Data with different superscript letters are significantly different (P < 0.05) according to the unpaired student T-Test. The asterisks symbol indicates significant differences *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001.
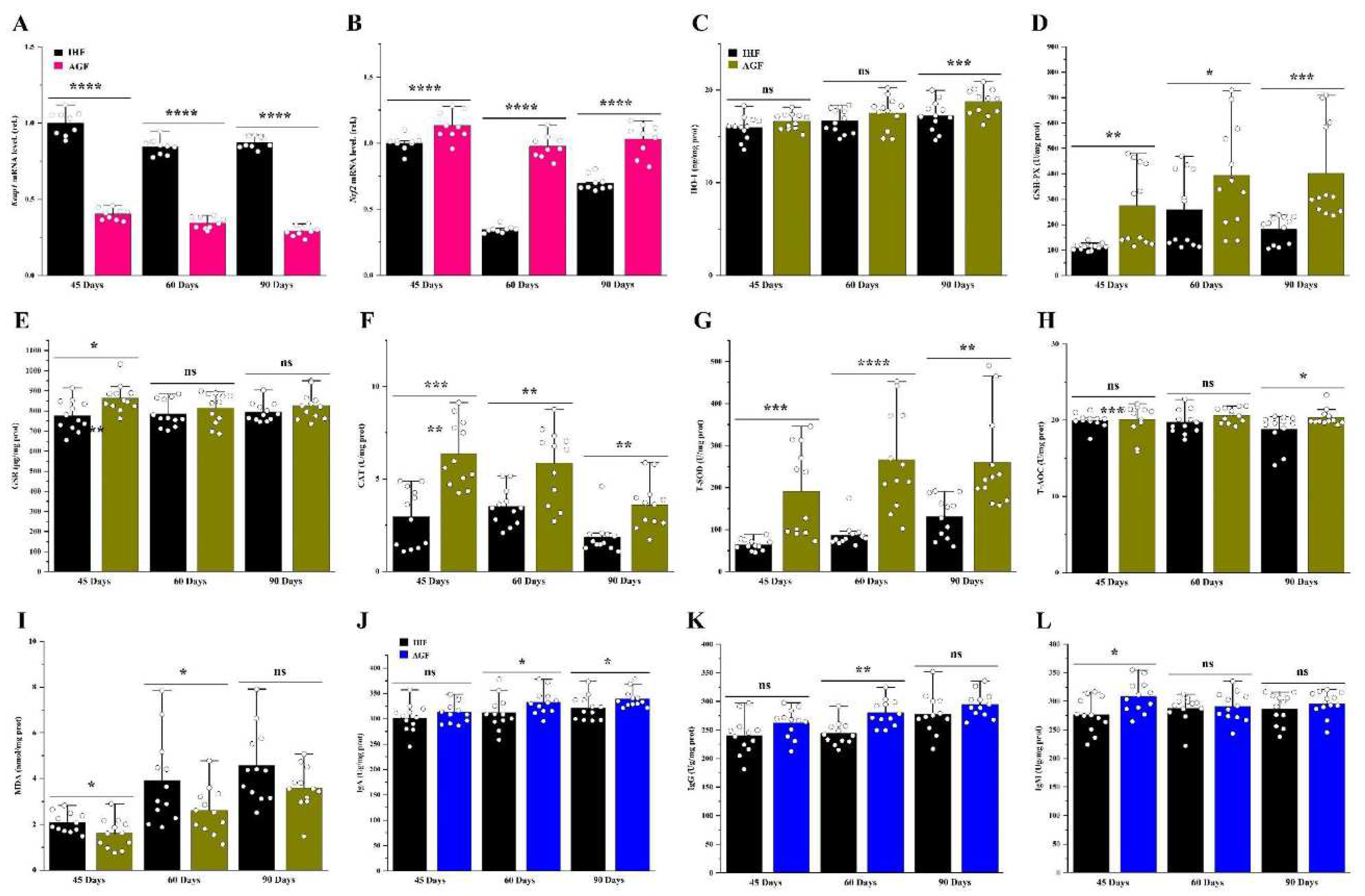
Figure 6.
Long-term high dietary fiber diet caused improved growth performance of geese. (A) ADFI and ADG measurement on 45d, 60d, and 90d, and (B) ABW and FCR measurement on 45d, 60d, and 90d. In-house feeding system (IHF) and artificial pasture grazing system (AGF). Data with different superscript letters are significantly different (P < 0.05) according to the unpaired student T-Test. The asterisks symbol indicates significant differences *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001.
Figure 6.
Long-term high dietary fiber diet caused improved growth performance of geese. (A) ADFI and ADG measurement on 45d, 60d, and 90d, and (B) ABW and FCR measurement on 45d, 60d, and 90d. In-house feeding system (IHF) and artificial pasture grazing system (AGF). Data with different superscript letters are significantly different (P < 0.05) according to the unpaired student T-Test. The asterisks symbol indicates significant differences *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001.
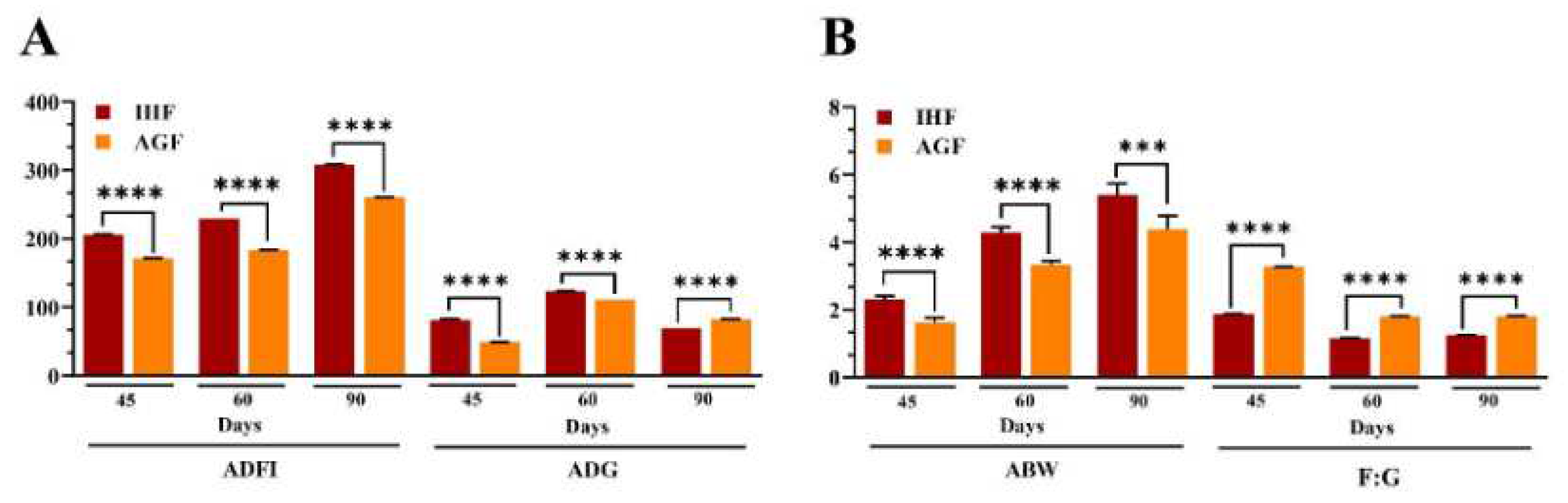
Figure 7.
Long-term high dietary fiber diet caused improved metabolic profile of geese. (A) HDL-C protein level in liver tissues, (B) LDL-C protein level in liver tissues, (C) TG protein level in liver tissues, (D) T-CHO protein level in liver tissues, and (E) BUN protein level in liver tissues, measured by ELISA kits. In-house feeding system (IHF) and artificial pasture grazing system (AGF). Data with different superscript letters are significantly different (P < 0.05) according to the unpaired student T-Test. The asterisks symbol indicates significant differences *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001.
Figure 7.
Long-term high dietary fiber diet caused improved metabolic profile of geese. (A) HDL-C protein level in liver tissues, (B) LDL-C protein level in liver tissues, (C) TG protein level in liver tissues, (D) T-CHO protein level in liver tissues, and (E) BUN protein level in liver tissues, measured by ELISA kits. In-house feeding system (IHF) and artificial pasture grazing system (AGF). Data with different superscript letters are significantly different (P < 0.05) according to the unpaired student T-Test. The asterisks symbol indicates significant differences *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001.
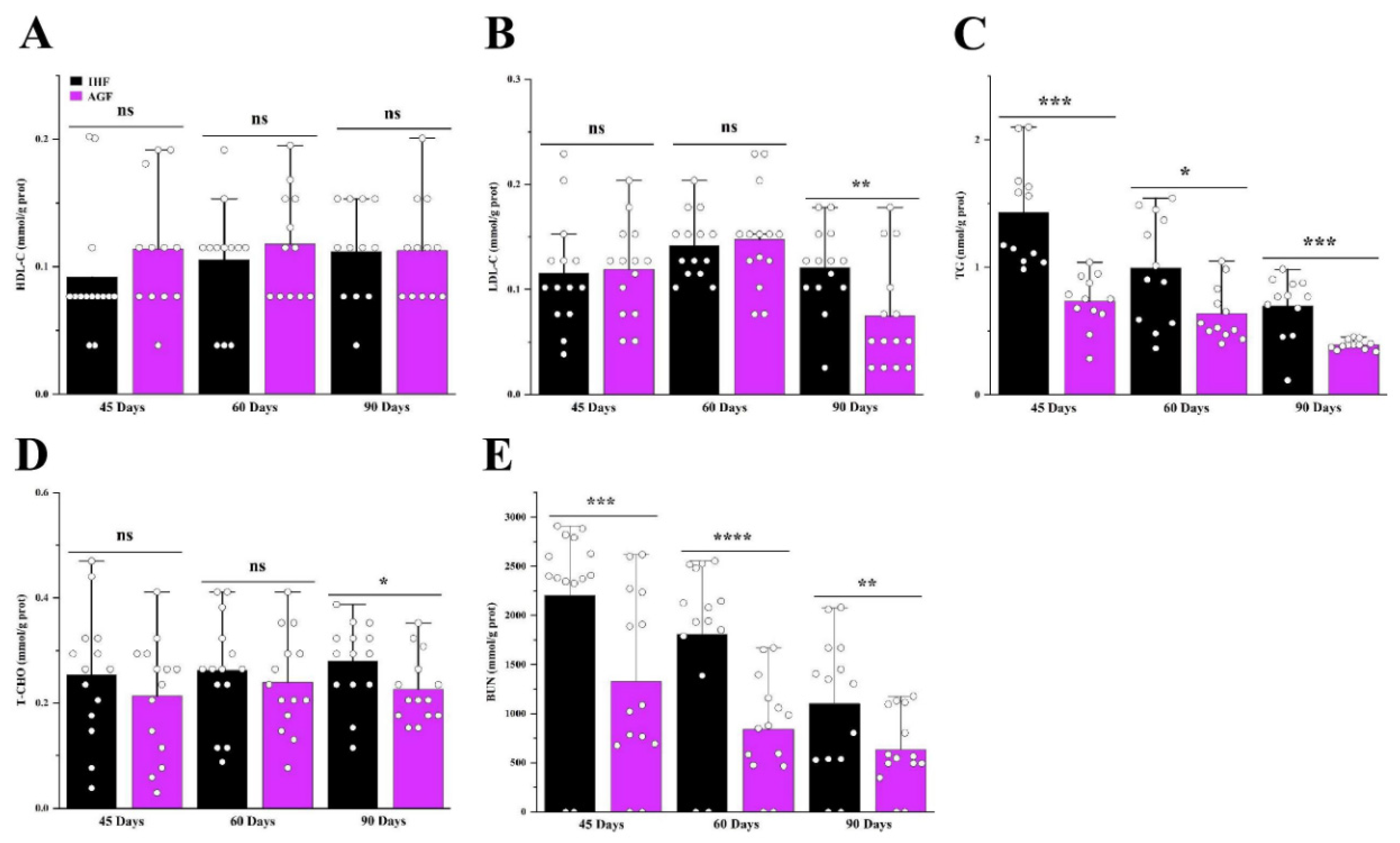
Figure 8.
Pearson correlation analysis of ALP-dependent Keap1-Nrf2 signaling pathway-induced antioxidants with LPS/ROS-induced gut barrier dysfunctions, systemic inflammation, growth performance, and metabolic syndrome at 45d (A), 60d (B), and (C) 90d. In the correlation heatmaps, the significant correlations were represented as (r > 0.52 or r < -0.52, p < 0.01). The size of circle and its color intensity are proportional to the correlation values.
Figure 8.
Pearson correlation analysis of ALP-dependent Keap1-Nrf2 signaling pathway-induced antioxidants with LPS/ROS-induced gut barrier dysfunctions, systemic inflammation, growth performance, and metabolic syndrome at 45d (A), 60d (B), and (C) 90d. In the correlation heatmaps, the significant correlations were represented as (r > 0.52 or r < -0.52, p < 0.01). The size of circle and its color intensity are proportional to the correlation values.
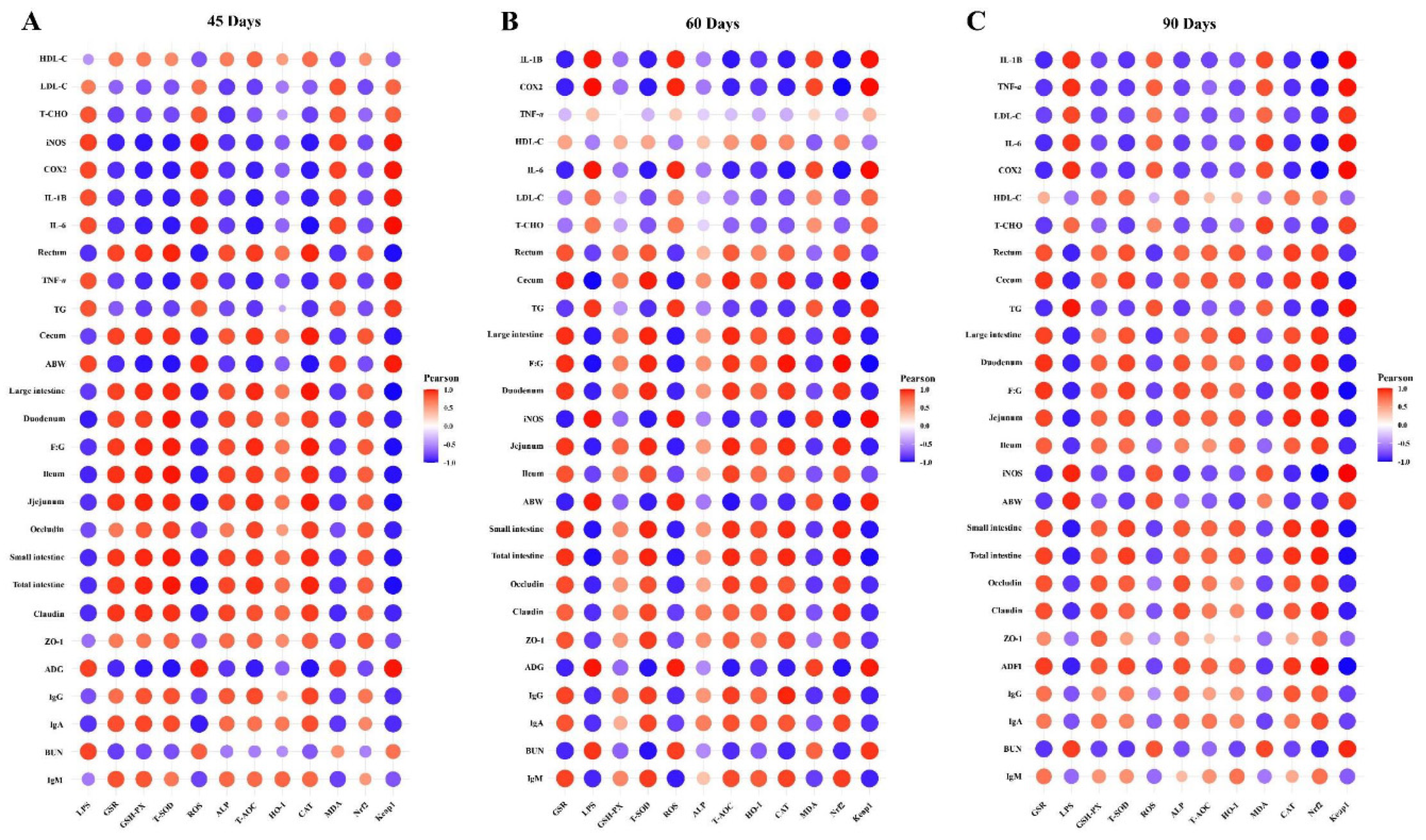

Table 1.
Commercial diet cause deterioration of nutrient absorption. Data expressed as mean ± SEM.
| Parameters | 45 D | 60 D | 90 D | ||||||
|---|---|---|---|---|---|---|---|---|---|
| IHF | AGF | P>0.05 | IHF | AGF | P>0.05 | IHF | AGF | P>0.05 | |
| Inner layer (um) | 51.82±5.76 | 59.5±6.13 | 0.02 | 54.51±5.77 | 80±8.62 | 6.4378E-05 | 48.6±7.3 | 70.29±5.69 | 9.4035E-05 |
| Outer layer (um) | 10.32±2.22 | 14.36±3.51 | 0.02 | 10.97±2.25 | 19.25±3.63 | 0.0004 | 8.53±1.18 | 15.48±2.13 | 6.1937E-05 |
| Total (um) | 62.14±5 | 73.86±9.04 | 0.010 | 65.48±3.93 | 99.26±9.26 | 0.000005 | 57.13±7.57 | 85.77±6.8 | 0.00002 |
| Relative thickness of muscular tonic (um) | 26.98±3.13 | 45.6±6.55 | 0.00005 | 15.34±1.33 | 29.81±2.44 | 0.0000001 | 10.69±1.91 | 19.76±2.71 | 0.00003 |
| Inner layer (um) | 8.89±1.82 | 14.25±3.66 | 0.00464366 | 8.5±1.11 | 21.37±3.68 | 4.7427E-06 | 6.53±1.27 | 11.68±3.4 | 0.003 |
| Outer layer (um) | 1.07±0.17 | 1.58±0.21 | 0.0004 | 1.16±0.25 | 1.61±0.31 | 0.01 | 0.99±0.15 | 1.68±0.33 | 0.0004 |
| Total (um) | 9.95±1.94 | 15.83±3.49 | 0.002 | 9.65±1.01 | 22.97±3.64 | 3.0064E-06 | 7.52±1.32 | 13.35±3.48 | 0.002 |
| Relative thickness of muscularis mucosa (um) | 4.33±0.99 | 9.76±2.21 | 0.0001 | 2.25±0.19 | 6.89±0.99 | 0.0000003 | 1.41±0.32 | 3.13±1.05 | 0.002 |
Table 2.
Commercial diet cause deterioration of nutrient absorption by effecting thickness of ileal muscular tonic and muscularis mucosa (50µm). In-house feeding system (IHF) and artificial pasture grazing system (AGF). Data expressed as mean ± SEM.
Table 2.
Commercial diet cause deterioration of nutrient absorption by effecting thickness of ileal muscular tonic and muscularis mucosa (50µm). In-house feeding system (IHF) and artificial pasture grazing system (AGF). Data expressed as mean ± SEM.
| Parameters | 45 D | 60 D | 90 D | ||||||
|---|---|---|---|---|---|---|---|---|---|
| IHF | AGF | P>0.05 | IHF | AGF | P>0.05 | IHF | AGF | P>0.05 | |
| Inner layer (um) | 51.82±5.76 | 59.5±6.13 | 0.02 | 54.51±5.77 | 80±8.62 | 6.4378E-05 | 48.6±7.3 | 70.29±5.69 | 9.4035E-05 |
| Outer layer (um) | 10.32±2.22 | 14.36±3.51 | 0.02 | 10.97±2.25 | 19.25±3.63 | 0.0004 | 8.53±1.18 | 15.48±2.13 | 6.1937E-05 |
| Total (um) | 62.14±5 | 73.86±9.04 | 0.010 | 65.48±3.93 | 99.26±9.26 | 0.000005 | 57.13±7.57 | 85.77±6.8 | 0.00002 |
| Relative thickness of muscular tonic (um) | 26.98±3.13 | 45.6±6.55 | 0.00005 | 15.34±1.33 | 29.81±2.44 | 0.0000001 | 10.69±1.91 | 19.76±2.71 | 0.00003 |
| Inner layer (um) | 8.89±1.82 | 14.25±3.66 | 0.00464366 | 8.5±1.11 | 21.37±3.68 | 4.7427E-06 | 6.53±1.27 | 11.68±3.4 | 0.003 |
| Outer layer (um) | 1.07±0.17 | 1.58±0.21 | 0.0004 | 1.16±0.25 | 1.61±0.31 | 0.01 | 0.99±0.15 | 1.68±0.33 | 0.0004 |
| Total (um) | 9.95±1.94 | 15.83±3.49 | 0.002 | 9.65±1.01 | 22.97±3.64 | 3.0064E-06 | 7.52±1.32 | 13.35±3.48 | 0.002 |
| Relative thickness of muscularis mucosa (um) | 4.33±0.99 | 9.76±2.21 | 0.0001 | 2.25±0.19 | 6.89±0.99 | 0.0000003 | 1.41±0.32 | 3.13±1.05 | 0.002 |
Table 3.
Long-term pasture intake caused improved growth performance of geese. Data expressed as mean ± SEM.
Table 3.
Long-term pasture intake caused improved growth performance of geese. Data expressed as mean ± SEM.
| Age, d | Parameters | IHF | AGF* | P-value |
|---|---|---|---|---|
| 45 d | ADFI (g/d) | 206.3±0.27 | 171.37±0.23 | 1.62E-20 |
| 60 d | 229.21±0.01 | 183.3±0.27 | 7.42E-23 | |
| 90 d | 308.3±0.09 | 260.4±0.09 | 2.61E-26 | |
| 45 d | ABW (kg) | 2.31±0.13 | 1.63±0.14 | 2.48E-06 |
| 60 d | 4.28±0.16 | 3.33±0.11 | 1.44E-07 | |
| 90 d | 5.39±0.34 | 4.38±0.4 | 0.000411 | |
| 45 d | ADG (g/d) | 81.5±1.26 | 49.06±0.12 | 1.33E-14 |
| 60 d | 123.15±0.35 | 110.88±0.02 | 5.60E-16 | |
| 90 d | 69.38±0.02 | 82.3±0.09 | 4.99E-22 | |
| 45 d | F:G | 1.87±0.01 | 3.27±0.01 | 5.73E-21 |
| 60 d | 1.16±0.01 | 1.8±0.01 | 1.43E-17 | |
| 90 d | 1.25±0 | 1.81±0.01 | 1.11E-16 |
*AGF, the average grass intake (g DM/d) for 45, 60, and 90 days was 66.09±1.03, 67.98±1.75, and 93.47±1.23, respectively. P-value**, is determined on the base of commercial diet fed to both groups (excluding grass intake).
Table 4.
Long-term pasture intake caused improved intestinal organ development of geese (cm/Kg. Data expressed as mean ± SEM.
Table 4.
Long-term pasture intake caused improved intestinal organ development of geese (cm/Kg. Data expressed as mean ± SEM.
| Age, d | Parameters | IHF | AGF | P-value |
|---|---|---|---|---|
| 45 d | Rectum (cm/Kg) | 7.04±0.6 | 8.9±0.84 | 0.001 |
| 60 d | 3.94±0.53 | 5.22±0.46 | 0.001 | |
| 90 d | 2.99±0.3 | 3.99±0.37 | 0.000 | |
| 45 d | Cecum (cm/Kg) | 10.02±0.63 | 13.87±1.1 | 0.001 |
| 60 d | 5.08±0.36 | 7.93±0.35 | 2.6984E-07 | |
| 90 d | 3.94±0.17 | 6.19±0.3 | 3.54706E-05 | |
| 45 d | Ileum (cm/Kg) | 39.9±4.16 | 47.78±4.24 | 0.004 |
| 60 d | 23.12±1.43 | 29.57±0.56 | 6.31188E-07 | |
| 90 d | 16.42±1 | 24.31±1.75 | 1.1555E-06 | |
| 45 d | Jejunum (cm/Kg) | 42.13±3.18 | 49.34±8.1 | 0.03 |
| 60 d | 23.2±0.73 | 29.49±1.7 | 4.15458E-06 | |
| 90 d | 17.23±1.41 | 22.95±1.13 | 7.64026E-06 | |
| 45 d | Duodenum (cm/Kg) | 20.04±1.1 | 27.26±2.11 | 1.10792E-05 |
| 60 d | 13.98±2.42 | 16.29±1.21 | 0.03 | |
| 90 d | 9.36±1.71 | 12.18±1.77 | 0.01 | |
| 45 d | Large intestine (cm/Kg) | 17.06±0.9 | 22.77±1.69 | 1.29825E-05 |
| 60 d | 9.02±0.67 | 13.15±0.68 | 4.78312E-07 | |
| 90 d | 6.94±0.44 | 10.18±0.58 | 2.02335E-06 | |
| 45 d | Small intestine (cm/Kg) | 102.07±5.98 | 124.38±12.34 | 0.001 |
| 60 d | 60.31±3.55 | 75.35±1.99 | 1.96589E-06 | |
| 90 d | 43.01±1.44 | 59.43±3.06 | 1.56872E-07 | |
| 45 d | Total intestine (cm/Kg) | 119.13±6.26 | 147.15±13.93 | 0.001 |
| 60 d | 69.32±4.12 | 88.5±2.41 | 4.66565E-06 | |
| 90 d | 49.94±1.74 | 69.62±3.51 | 1.15652E-07 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



