Submitted:
08 October 2023
Posted:
09 October 2023
You are already at the latest version
Abstract
Trichoderma spp. are widely used in plant disease control as biocontrol agents. Inducing plant disease resistance is an important biocontrol mechanism of Trichoderma. It is of great significance to explore the factors involved in the surface recognition and interaction of Trichoderma with plants. Hydrophobin is a small molecular protein rich in cysteine in higher filamentous fungi and has been found to participate in the morphogenesis and interaction of fungi with other organisms. In this study, the type II hydrophobic protein-coding gene Thfb6 from T. harzianum TH33 was cloned and heterologously expressed in Escherichia coli BL21 (DE3). The purified recombinant protein Thfb6 induced a hypersensitive response in tobacco leaves, a burst of reactive oxygen species, and an accumulation of callose, phenolic substances, and lignin in tobacco, indicating that Thfb6 can induce an early defense reaction in tobacco. Thfb6 also enhanced the systemic resistance of tobacco against tobacco mosaic virus. The rate of lesion number reduction by treatment with Thfb6 could be up to 36.02%. Thfb6 increased the activity of tobacco defense-related enzymes, including phenylalanine ammonia-lyase (PAL), polyphenol oxidase, and peroxidase, which were all more than three times the control after 3–5 days post treatment of Thfb6. The transcription of resistance-related genes, including the PAL gene, non-expresser of pathogenesis-related gene1 (NPR1), and basic pathogenesis-related gene 1 (PR1-b), were upregulated after Thfb6 treatment. The relative expression levels of the PAL and NPR1 genes were highest on the third and fourth days after Thfb6 treatment, at 5.5 times and 8.3 times that of the control, respectively. The relative expression level of PR1 on the sixth day after Thfb6 treatment was 5.3 times that of the control. These results indicate that the improvement of defense-related enzyme activity and the induced expression of disease-resistance-related genes were the main mechanisms by which Thfb6 induced systemic resistance in tobacco.
Keywords:
hydrophobin
; elicitor
; Trichoderma
; induced systemic resistance
; tobacco
1. Introduction
Trichoderma is an important biocontrol fungus [1]. The antagonistic capacity of Trichoderma has been widely studied, and the mechanisms associated with it include competition for space and nutrients, antibiosis, and mycoparasitism [2]. Trichoderma colonizes and interacts with plants in the rhizosphere. Certain molecules, so-called elicitors, produced and released by Trichoderma, play an important role in the signal exchange between fungi and plants [3]. As early as 1997, Bigirimana et al. demonstrated for the first time that Trichoderma can cause induced systemic resistance (ISR) in plants, and subsequent studies have found that Trichoderma can produce a variety of elicitors to induce a defense response in plants [1,4]. The reported elicitors from Trichoderma belong to the Cerato-platanin family (such as Sm1, epl1, and Swollenin), glycoside hydrolases (such as Thph1, ThPG1, and Eix), and hydrophobic proteins [5,6,7,8].
Hydrophobins are a family of small proteins found uniquely in filamentous fungi that can self-assemble to form protein films at the interfaces between fungal cell walls and hydrophobic surfaces or at water–air interfaces and thus participate in a variety of developmental processes, including the formation of aerial hypha and fruiting body, sporulation, and differentiation of fungal infection structures [9,10,11,12,13,14]. Because of the amphiphilic properties of hydrophobin, it is widely studied as a promising drug delivery carrier in the biomedical field [15]. In the agricultural field, hydrophobin has been found to play a role in the interaction of fungi with other organisms [16]. We previously reported that T. harzianum strain TH33 can control Fusarium wilt and induce disease resistance in cucumbers [17,18]. There are six class II hydrophobin-coding genes in the TH33 genome with different expression patterns at different stages of fungal development, suggesting that they have different functions [19,20]. In this study, we reported that hydrophobin Thfb6 from TH33 is an elicitor that can cause a hypersensitive response (HR) response and ISR in tobacco and improve resistance to tobacco mosaic virus (TMV) infection.
2. Materials and Methods
2.1. Strains and plant
Wild-type T. harzianum TH33 was originally isolated from soil samples in the Bejing region as described previously [18]. The fungus was maintained on potato dextrose agar (PDA) medium at 25°C in the dark [21]. Escherichia coli competent cells were purchased from TaKaRa Biotechnology (Dalian, China) and grown in LB medium at 37°C. Tobacco cells were provided by the Protein Pesticide Group in IPPCAAS. Tobacco BY2 suspension cells were cultured in Murashige and Skoog media at 25℃ with shaking at 150rpm in the dark according to Wang’s method [22]. Tobacco plants (Nicotiana tabacum cv. Samsun NN) were cultured in 24–26°C greenhouse with 16 h of light and 8 h of darkness per day.
2.2. Gene cloning of Thfb6
Total RNA of T. harzianum TH33 was extracted using an E.Z.N.A.TM Fungal RNA Kit (Omega Bio-tek, Norcross, GA, USA), and cDNA was synthesized using a TransScriptTM Two-Step RT-PCR Super Mix Kit (TransGen Biotech, Beijing, China) with oligo dT as primers. Based on the Thfb6 sequence in the TH33 genome (GenBank number: PRJNA272949) [21], a pair of gene-specific primers was designed to amplify the entire coding sequence of Thfb6. The forward primer was 5′-TGCGCTTCGCCACCATCACCG-3′, and the reverse primer was 5′-ATGAAGTTCTCTGCCATCGCTC-3′. The PCR product was cloned into pMD 18-T vector (TaKaRa Biotechnology, Dalian, China), and then transformed into E. coli DH5a. the recombinant plasmid was extracted, and the Thfb6 cDNA sequence was verified using DNA sequencing.
2.3. Expression and purification of Thfb6 protein
A Thfb6 expression vector was constructed by inserting a Thfb6 designaling peptide cDNA gene fragment into a pET-28a vector with a His6 tag (Novagen, USA) via In-Fusion cloning [23]. Thfb6 designaling peptide cDNA gene fragments from the genomic cDNA of TH33 were amplified using PCR. The forward primer contained the HindIII restriction site (underlined) and a 14 bp homologous sequence of the pET-28a vector at the 5′ end: 5′-CGAGCTCCGTCGACAAGCTTTCACTCAGGAGCACCAGTTCGATATC-3′. The reverse primer contained the XhoI restriction site (underlined) and a 15 bp homologous sequence of pET-28a at the 5′ end: 5′- GTGGTGGTGGTGGTGCTCGAGTTACTGGGGAAGGGCATCCTGGCAC -3′. PCR was performed under the following conditions: 94°C for 2 min and 35 cycles of 94°C for 20 s, 58°C for 20 s, 72°C for 20 s; and 72°C for 5 min. The PCR products were isolated by electrophoresis in 1.5% agarose gel and then ligated with a HindIII/XhoI linearized pET-28a vector using an In-Fusion® HD Cloning Kit (TAKARA) (TaKaRa Biotechnology, Dalian, China), generating Thfb6 expression vector pET-Thfb6, which was then transformed into E. coli BL21 (DE3)-competent cells to obtain prokaryotic expression strain E. coli BL21 (DE3)-Thfb6.
The recombinant Thfb6 was expressed in E. coli BL21 (DE3)-Thfb6 induced for 4 h by 0.5 mM IPTG (Sigma, USA) at 37℃. The cells were collected and resuspended in 20 mM Tris-HCl buffer (pH8.0) and disrupted with an ultrasonic disruptor. The supernatant was concentrated through centrifugation at 15,000 g for 30 min. The recombinant Thfb6 in the supernatant was purified using a ProteinIso ® Ni-NTA Resin affinity column (TransGen biotech) and dissolved in 20 mM Tris-HCl buffer (pH 8.0). Protein molecular markers were used to estimate the sizes of the purified proteins (Fermentas, Hanover, USA). The protein concentrations were measured using a BCA™ Protein Assay Kit (Pierce, Rockford, IL, USA).
2.4. Hypersensitive response assays
The HR-inducing activity of Thfb6 was assayed in 8-week-old tobacco plants (Nicotiana tabacum cv. Samsun NN). A volume of 50 μL Thfb6 proteins was infiltrated into tobacco leaves at a concentration of 20 µM with a syringe without a needle, and 50 μL of 20 mM Tris-HCl buffer was used as a negative control. At 24 h post-treatment (hpt), HR symptoms were observed according to a previously described method [24]. Hydrogen peroxide (H2O2) in leaves was detected by 3,3’-diaminobenzidine (DAB) staining. The leaves were first incubated in DAB-HCl (1mg /mL, pH 3.8) solution (Sigma, St. Louis, MO, USA) in darkness for 2 hours, then incubated in boiled ethanol for 10min to remove chlorophyll, and the treated leaves were subsequently stored in 70% glycerol. H2O2 accumulation was observed under an Olympus Stereoscope SZX-9 (Olympus America Inc., Melville, NY, USA) [25].
2.5. Analysis of callose deposition
Callose deposition in leaves was observed according to Chen’s method [26]. 8-month-old tobacco leaves were treated with 20 μM Thfb6, and 24 h later, the leaves were fixed with glutaraldehyde and removed chlorophyll with ethanol. the transparent leaves were then stained with 0.1% (w/v) aniline blue (pH 12.0) for 4 h. The stained leaves were examined for callose deposition under fluorescence microscope (AXiO Imager M1, DAPI filter, Carl Zeiss, Jena, Germany).
2.6. Detection of phenolic and lignin content
The phenolic content in tobacco suspension cells induced by Thfb6 was detected as previously described [26]. 20 µM Thfb6 was added to tobacco suspension cells and incubated for 108 h. then the cells were examined with the fluorescence microscope (AXiO Imager M1, DAPI filter) for phenolics accumulation which showed blue fluorescence. 20 mM Tris-HCl buffer (pH 8.0) was used as a negative control. To evaluate the lignin content in tobacco suspension cells, lignin in the cell wall was converted to lignin thioglycolate and dissolved in 1 mL of 1 N NaOH. The lignin content was expressed as the absorption measured at 280 nm using 1 N NaOH as the blank [22].
2.7. Bioassay for Thfb6-induced disease resistance in tobacco TMV resistance assays
Infection of tobacco plants by TMV was carried out using established methods [25]. The fourth and fifth leaves of tobacco were infiltrated with 50 µL (20 µM) Thfb6 protein solution, and 20 mM Tris-HCl buffer was used as a negative control. TMV inoculation was performed by gently rubbing the upper three leaves with 200 µL of the viral suspension (12 µg/mL) containing carborundum 3 days post treatment (dpt) with Thfb6. The number and size of TMV lesions were recorded on the third day post-TMV inoculation. The percentage of lesion inhibition was used to represent the resistance of tobacco to TMV, which was calculated as follows: Inhibition (%) = [(number (size) of lesions on control leaves − number (size) of lesions on protein-treated leaves)/number (size) of lesions on control leaves] × 100%. All experiments were repeated 3 times with 10 plants per replicate.
2.8. Determination of peroxidase, phenylalanine ammonia-lyase, and polyphenol oxidase activities
The fourth leaves of 8-week-old tobacco plants were treated with 50 µL (20 µM) Thfb6 protein solution, and 20 mM Tris-HCl was used as a negative control. The upper three leaves of treated leaves were harvested at different times (0, 1, 3, 5, and 7 d) after treatment and were immediately frozen in liquid nitrogen and stored at −80°C. For detection of enzyme activities, 2 g leaf samples were homogenized using a mortar and pestle with 50 mM sodium acetate buffer solution (pH 5.0). The homogenized solution was then centrifuged at 14,000 g for 15 min at 4 ℃. The supernatant was collected as crude enzyme solution. The activities of peroxidase (POD), phenylalanine ammonia-lyase (PAL), and polyphenol oxidase (PPO) in the leaves were determined using a corresponding enzyme activity detection kit (Suzhou Keming Biotechnology Co., Ltd. China); the operation was performed according to the manufacturer’s instructions.
2.9. RT-qPCR analysis of defense-related genes induced by Thfb6
Real-time quantitative PCR (RT-qPCR) was used to detect the expression of tobacco defense-related genes induced by Thfb6, including disease-course-related protein gene PR-1b, the systemic resistance marker gene non-expresser of pathogenesis-related gene1 (NPR1), and the defense response key enzyme gene PAL. The fourth leaves of 8-week-old tobacco plants were treated with 50 µL (20 µM) Thfb6 protein solution, and 20 mM Tris-HCl was used as negative control. The upper three leaves of three treated plants were pooled at 1–5 dpt. The total RNA of the leaf tissue was isolated, and the cDNA were produced and was used as template in RT-qPCR reactions. Beacon Designer 8 software was used to design PCR primers of PR-1b, NPR1 and PAL, ß-actin gene was used as an internal control (Table 1). PCR was performed under the following conditions: 94°C for 2-5 min, followed by 30-35 cycles at 94°C for 30 s, 55-56°C for 30 s, and 72°C for 30 s, and a final step at 72°C for 5-10 min. Each treatment was repeated three times.
2.10. Statistical analysis
All experiments and data presented here involved at least three repeats. The statistical analysis was performed by Microsoft Excel 2007. The data are presented as means and standard deviations. One way ANOVA with P< 0.05 was used to determine the statistical difference.
3. Results
3.1. cDNA cloning of Thfb6 and expression in E. coli
The full-length cDNA fragment coding Thfb6 was cloned from the constructed cDNA library of T. harzianum TH33 [19]. The Thfb6 gene (Genbank: OR419735.1) included an open reading frame of 309 bp encoding a protein of 102 amino acids with a theoretical molecular weight of 10.56 kDa. The cDNA sequence contained a conserved domain named hydrophobin_2 superfamily (pfam06766) and coded a type II hydrophobin, which showed 100% identity to hydrophobin 4 (hfb4) (GenBank: OPB37525.1) with unknown function from T. guizhouense, 53, 73% identity to hydrophobin type 4 (HFB4) coding gene (GenBank: OL361837.1) from T. viride TV154, and 43.59% identity to Thhyd1 coding gene (GenBank: KT258897) from T. harzianum T28, which has been identified to be a elicitor that can cause ISR in maize [27]. Analysis of the Thfb6 amino acid sequence with the SignaIP 5.0 Server (https://services.healthtech.dtu.dk/services/SignalP-5.0/) indicated that it contained a 21-amino acid signal peptide, suggesting that Thfb6 is a secreted protein. His-tagged signal peptide deleted Thfb6 was expressed in E. coli BL21 (DE3) and purified. The purified recombinant protein showed a single band with a molecular mass of 9.4 kDa on SDS-PAGE (Figure 1A).
3.2. Thfb6 triggers defense responses in tobacco
After treatment with Thfb6 for 24h, there were clearly defined necrotic lesions in leaves of tobacco at the infiltration sites (Figure 1B), while the control treatment did not show an HR response (Figure 1C). The tobacco leaves treated with Thfb6 stained with DAB showed an obvious accumulation of brick-like substances (Figure 1E), indicating that Thfb6 can induce tobacco cells to produce reactive oxygen species (ROS) (mainly H2O2), which combined with DAB to produce a reddish-brown substance, while the negative control leaves treated with buffer were dark gray (Figure 1D). When the Thfb6-treated tobacco leaves were dyed with aniline blue after chlorophyll removal, dot fluorescence was observed under a fluorescence microscope (Figure 2A), indicating that Thfb6 caused callose accumulation in the tobacco leaves. The leaves treated with control buffer (Tris-HCl) showed no change in callose deposition (Figure 2B). The pH of the Thfb6-treated tobacco cell suspensions increased significantly compared with that of the control within 20 min after treatment (Figure 2C), which showed that Thfb6 induced extracellular ion flow changes in tobacco suspension cells, as indicated by extracellular alkalization. Phenolic compounds are one of the main secondary metabolites produced in the plant defense response. Autofluorescence of tobacco cell walls was observed in Thfb6-treated cells (Figure 3A), showing deposition of wall-bound phenolic compounds, while no accumulation of phenols was observed in control cells (Figure 3B). This was confirmed by a significant increase in the lignin content of the incubated cells relative to the control (Figure 3C). The above results indicate that Thfb6 can trigger early defense events in tobacco as an elicitor.
3.3. Induction of TMV resistance in tobacco by Thfb6
To determine whether Thfb6 improved the resistance of tobacco to TMV, tobacco leaves were treated with Thfb6 and then inoculated with TMV. The number and size of TMV lesions on tobacco leaves were recorded (Table 2). The results showed that Thfb6 treatment could reduce the occurrence of TMV disease of tobacco. After 3 days of TMV inoculation, the TMV lesion number and lesion diameter of Thfb6-treated leaves were reduced by 36.02% and 41.57%, respectively, compared with control.
3.4. Thfb6 increased the activities of defensive enzymes PAL, POD and PPO in tobacco
Enzyme activities related to plant defense ability, including PAL, PPO, and POD, were detected for 7 consecutive days after Thfb6 treatment. The activity of all three enzymes increased due to Thfb6 (Figure 4). Among them, PAL activity was stimulated and peaked at 5 dpt and was approximately 5.71 times that of the control (Figure 4A). Both PPO and POD activities peaked at 3 dpt and were 4.87 and 3.07 times higher than those of the control, respectively (Figure 4B,C).
3.5. Expression of resistance-related genes induced by Thfb6
RT-qPCR was used to detect the expression of three resistance-related genes in tobacco after Thfb6 treatment. As shown in Figure 5, the expression of the PAL gene increased after Thfb6 treatment, reaching the highest value at 3 dpt, and the relative expression level was 5.5 times that of the control. The expression level of the NPR1 was significantly increased at 3 dpt and reached the highest level at 4 dpt. The relative expression level of NPR1 was 8.3 times higher than that of the control group and showed a downward trend at 5 dpt. The expression level of PR1-b continued to increase until 5 dpt with Thfb6, and the relative expression level was 5.3 times that of the control at 6 dpt. The results showed that Thfb6 treatment induced the increase of defense-related enzyme activities in tobacco leaves, suggesting that it could improve the disease resistance of tobacco.
4. Discussion
There are multiple hydrophobin-coding genes in fungal genomes with low sequence similarity, while only a few of them have been studied. For example, Aspergillus contains six to eight hydrophobic protein genes that have little amino acid sequence homology, except for eight cysteines [28], and different functions [29]. Some hydrophobins of pathogenic fungi are closely related to their pathogenicity, such as VdHP1, a hydrophobin that plays an important role in the development, adaptability, and virulence of Verticillium dahlia [30]. Regarding beneficial plant fungi, such as Trichoderma, some hydrophobins are involved in interactions with pathogens and plants and induce systemic resistance in plants as elicitors [31,32,33]. For example, T. virens mutant that overexpress the hydrophobin coding gene tvhydii1 showed greater resistance to R. solani AG2 than the wild strain, while colonization ability in plant roots of tvhydii1 deletion mutant was reduced [33]. hyd1 from T. harzianum T28 can interact with the ubiquitin 1-like (UBL) protein in maize roots and cause ISR of maize to leaf spot disease [27]. Exploring the function of hydrophobins of Trichoderma and clarifying the interaction mechanism between Trichoderma and plants are of great significance for the application of elicitors to reduce disease occurrence. In this study, we identified a hydrophobin Thfb6 from T. harzianum TH33 as an elicitor that can cause ISR in tobacco. Thfb6 has 100% similarity hfb4 from T. guizhouense TUCIM. The study of Thfb6 in his study will help us to further understand the function of hfb4 in TUCIM. Thfb6 has 43.59% similarity to the hydrophobin elicitor Thhyd1 from T. harzianum T28, illustrating that they may have different interaction mechanisms with plants.
Trichoderma can produce various types of elicitors that have different interaction mechanisms with plants and can cause ISR through different signaling pathways in plants. Many signaling pathways, including SA, jasmonic acid (JA), ethylene (ET), and abscisic acid (ABA), participate in the process of ISR in plants induced by Trichoderma [34,35]. For example, T. gamesii IMO5 and B21 cause ISR in maize through the JA and SA signaling pathways, respectively, improving the resistance of maize to Fusarium [36]. T. viridis causes maize ISR through the JA signaling pathway to improve resistance to pathogens [37]. T. harzianum T-78 induces the expression of genes related to the JA, SA, ET, and ABA signaling pathways in tomatoes and improves the resistance of tomatoes to gray mold [38]. T. harzianum T382 causes ISR in Arabidopsis thaliana through the phenylpropane pathway and increases resistance to gray mold [39]. However, there are few studies on the elicitors of Trichoderma in these interaction systems. Hydrophobin Thhyd1 from T. harzianum T28 has been reported to induce the resistance of maize to Botrytis cinerea primarily associated with brassinosteroid signaling, JA/ET signaling is also involved in this response [27].
Tobacco and its suspended cells are easy to prepare and are commonly used as experimental materials to study induced defense response [25]. Some fungal elicitors have been identified that can trigger broad-spectrum defense responses in plants, including cell wall reinforcement, biosynthesis of antimicrobial compounds, generation of ROS, and expression of defense genes, accompanied by the change of extracellular pH value, reflecting the ion flow changes inside and outside the cell to restrict pathogen infection [40,41]. ROS are synergistic and universal regulatory elements in the plant defense system. ROS have antibacterial effects and act as a signaling molecules to induce and participate in multiple signaling pathways in plant by interacting with a variety of molecules including lipids, Ca2+, NO and mitogen-activated protein kinase (MAPK), etc. [42]. The phenylpropanoid pathway plays a crucial role in plant defense [43], and the PAL is one of the core gene of the phenylpropanoid pathway [44]. Here, we demonstrated an oxidative burst occurred accompanied with an increase in extracellular pH of tobacco after Thfb6 treatment. the expression of the PAL gene in tobacco was induced, and PAL enzyme activity was enhanced. Moreover, the expression of the SA signaling pathway marker gene PR1-b and the systemic resistance marker gene NPR1 was simultaneously induced. The above results indicated that Thfb6 initiates the phenylpropanoid metabolic pathway and may cause ISR in tobacco through the SA signaling pathway. However, the molecular mechanisms involved in Thfb6-mediated signal transduction requires further study.
The PAL enzyme is a key enzyme in the phenylpropanoid pathway responsible for the synthesis of defense-related compounds such as lignin, phytolexin and flavonoids. PAL activity is often used as one of the important indices for plant resistance [45]. POD is involved in the polymerization of monolignols into lignin and the reinforcement of the cell wall, which increases tissue hardness to prevent pathogen entry [46]. PPO can oxidize phenolic substances to quinones, which are more toxic to pathogenic microorganisms. The activities of PAL, POD, and PPO in tobacco leaves treated with Thfb6 were significantly increased in this study. It is speculated that an increase in defense enzyme activity is one of the mechanisms by which Thfb6 induces resistance to TMV in tobacco.
Synthesis and accumulation of secondary metabolites such as callose, phenol substances and lignin play an important role in plant defense responses against biotic and abiotic stress, and were commonly used for identification of elicitor inducing ISR of plants [47]. Here, callose, phenolics, and lignin were induced in tobacco by Thfb6 treatment. The relationship between these secondary metabolites and plant resistance signaling molecules is unclear and requires further study.
5. Conclusions
We identified a type II hydrophobic protein elicitor, Thfb6, from biocontrol strain T. harzianum TH33. Thfb6 induced ROS burst in tobacco leaves and an accumulation of callose, phenols, and lignin in tobacco cells; increased the extracellular pH; and improved the resistance of tobacco plants to TMV. Further studies showed that Thfb6 increased the activity of tobacco defense-related enzymes, including PAL, PPO, and POD, and upregulated the expression of disease resistance-related genes, including PAL, NPR1, and PR1-b. It is speculated that Thfb6 may cause ISR in tobacco through the phenylpropanoid metabolic pathway and the SA signaling pathway. Next, we will explore the molecular mechanism of ISR by Thfb6 by studying the interacting proteins of Thfb6 in tobacco using transcriptomic and proteomic sequencing and by applying Thfb6 in the prevention and control of other plant diseases.
Author Contributions
Conceptualization, X.J. and M.L.; investigation, X.J. and M.L.; methodology, Q.C., J.M.; validation, J.L., P.H., Q.C., J.M. and Y.T.; data curation, J.L., P.H. and Y.T.; writing—original draft preparation, J.L., P.H. and M.L.; writing—review and editing, M.L.; project administration, M.L.; funding acquisition, M.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (Grant No. 31371983) and the National Key R&D Program of China (2017YFD0200900).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Acknowledgments
We thank LetPub (https://www.letpu b.com) for its linguistic assistance during the preparation of this manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Harman, G. E. Overview of mechanisms and uses of Trichoderma spp. Phytopathology 2006, 96, 190–194. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, P. K.; Horwitz, B. A.; Kenerley, C. M. Secondary metabolism in Trichoderma a genomic perspective. Microbiology 2012, 158, 35–45. [Google Scholar] [CrossRef]
- Mishra, D. S.; Prajapati, C. R.; Gupta, A. K.; Sharma, S. D. Relative bio-efficacy and shelf-life of mycelial fragments, conidia and chlamydospores of Trichoderma harzianum. Int. J. Plant Res. 2012, 25, 225–232. [Google Scholar]
- Zhang, H.; Ji, S.; Guo, R.; Zhou, C.; Wang, Y.; Fan, H.; Liu, Z. Hydrophobin HFBII-4 from Trichoderma asperellum induces antifungal resistance in poplar. Braz. J. Microbiol. 2019, 50, 603–612. [Google Scholar] [CrossRef] [PubMed]
- Ramírez, C. A.; Kloepper, J. W. Plant growth promotion by Bacillus amyloliquefaciens FZB45 depend on inoculum rate and P-related soil properties. Biol. Fert. Soils 2010, 46, 835–844. [Google Scholar] [CrossRef]
- Salas-Marina, M. A.; Silva-Flores, M. A.; Uresti-Rivera, E. E.; Castro-Longoria, E.; Herrera-Estrella, A.; Casas-Flore, S. Colonization of Arabidopsis roots by Trichoderma atroviride promotes growth and enhances systemic disease resistance through jasmonic acid/ethylene and salicylic acid pathways. Eur. J. Plant Pathol. 2011, 131, 15–26. [Google Scholar] [CrossRef]
- Cai, F.; Yu, G.; Wang, P.; Wei, Z.; Fu, L.; Shen, Q.; Chen, W. Harzianolide, a novel plant growth regulator and systemic resistance elicitor from Trichoderma harzianum. Plant Physiol. Bioch. 2013, 73, 106–113. [Google Scholar] [CrossRef]
- Contreras-Cornejo, H. A.; Macías-Rodríguez, L.; Del-Val, E.; Larsen, J. Ecological functions of Trichoderma spp. and their secondary metabolites in the rhizosphere: interactions with plants. FEMS Microbiol. Ecol. 2016, 92, fiw036–fiw036. [Google Scholar] [CrossRef]
- Wessels, J. G. H. Hydrophobins: proteins that change the nature of the fungal surface. Adv. Microb. Physiol. 1997, 38, 1–45. [Google Scholar]
- Zhang, S.; Xia, Y.X.; Kim, B.; Keyhani, N. O. Two Hydrophobins are involved in fungal spore coat rodlet layer assembly and each play distinct roles in surface interactions, development and pathogenesis in the entomopathogenic fungus, Beauveria bassiana. Mol. Microbiol. 2011, 80, 811–826. [Google Scholar] [CrossRef]
- Whiteford, J. R.; Lacroix, H.; Talbot, N. J.; Spanu, P. D. Stage-specific cellular localisation of two hydrophobins during plant infection by the pathogenic fungus Cladosporium fulvum. Fungal Genet. Biol. 2004, 41, 624–634. [Google Scholar] [CrossRef] [PubMed]
- Han, A. B. W.; Karin, S. Applications of hydrophobins: current state and perspectives. Appl. Microbiol. Biot. 2015, 99, 1587–1597. [Google Scholar]
- Klimes, A.; Dobinson, K. F. A hydrophobin gene, vdh1, is involved in microsclerotial development and spore viability in the plant pathogen Verticillium dahliae. Fungal Genet. Biol. 2006, 43, 283–294. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Ahn, I. P.; Rho, H. S.; Lee, Y. H. Mph1, a Magnaporthe grisea hydrophobin gene, is required for fungal development and plant colonization. Mol. Microbiol. 2005, 57, 1224–1237. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Yan, J.; Chen, Y. The progress of hydrophobin-based drug delivery system. China Biotechnology 2023, 43, 15–25. [Google Scholar]
- Xia, Y.; Talbot, N. J. Investigating the cell biology of plant infection by the rice blast fungus Magnaporthe oryzae. Curr. Opin. Microbiol. 2016, 34, 147–153. [Google Scholar]
- Jie, D.; Jie, M.; Pei, H.; Ying, T.; Yao, L.; Xiliang, J.; Mei, L. A Gα3 subunit Thga3 positively regulates conidiation, mycoparasitism, chitinase activity and hydrophobicity of Trichoderma harzianum. AMB Express 2020, 10, 221. [Google Scholar]
- Jie, M.; Lirong, W.; Xiliang, J.; Beilei, W.; Mei, L. Functions of the C2H2 transcription factor gene thmea1 in Trichoderma harzianum under copper stress based on transcriptome analysis. BioMed. Res. Int. 2018, 2018, 1–10. [Google Scholar] [CrossRef]
- Yang, X.; Li, M.; Zhang, L.; Pang, L.; Sun, q.; Jiang, X. Transcriptiome analysis of Trichoderma harzianum Th-33 in chlamydospore formation. Chinese J. Biol. Control 2015, 31, 85–89. [Google Scholar]
- Xinhong, P.; Beilei, W.; Shuaihu, Z.; Mei, L.; Xiliang, J. Transcriptome dynamics underlying chlamydospore formation in Trichoderma virens GV29-8. Front. Microbiol. 2021, 12, 654855. [Google Scholar]
- Sun, Q.; Jiang, X.; Pang, L.; Wang, L.; Li, M. Functions of thga1 gene in Trichoderma harzianum based on transcriptome analysis. BioMed. Res. Int. 2016, 2016, 8329513. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Yang, X.; Zeng, H.; Liu, H.; Zhou, T.; Tan, B.; Yuan, J.; Guo, L.; Qiu, D. The purification and characterization of a novel hypersensitive-like response-inducing elicitor from Verticillium dahliae that induces resistance responses in tobacco. Appl. Microbiol. Biotechnol. 2012, 93, 191–201. [Google Scholar] [CrossRef]
- Li, X.; Li, Y.; Chen, S.; Wang, J. Construction of stable infectious full-length and eGFP-tagged cDNA clones of Mirabilis crinkle mosaic virus via In-Fusion cloning. Virus Res. 2020, 286, 198039–198039. [Google Scholar] [CrossRef] [PubMed]
- D’Silva, I.; Heath, M.C. Purification and characterization of two novel hypersensitive response-inducing specific elicitors produced by the cowpea rust fungus. J. Biol. Chem. 1997, 272, 3924–3927. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Zeng, H.; Liu, Z.; Yang, X.; Guo, L.; Qiu, D. Mutational analysis of the Verticillium dahliae protein elicitor PevD1 identifies distinctive regions responsible for hypersensitive response and systemic acquired resistance in tobacco. Microbiol. Res. 2014, 169, 476–482. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Zeng, H.; Qiu, D.; Guo, L.; Yang, X.; Shi, H.; Zhou, T. Purification and characterization of a novel hypersensitive response-inducing elicitor from Magnaporthe oryzae that triggers defense response in rice. PLoS One 2012, 7, e37654. [Google Scholar] [CrossRef]
- Yu, C.; Dou, K.; Wang, S.; Wu, Q.; Ni, M.; Zhang, T.; Lu, Z.; Tang, J.; Chen, J. Elicitor hydrophobin hyd1 interacts with ubiquilin1-like to induce maize systemic resistance. J. Integr. Plant Biol. 2019, 62, 509–526. [Google Scholar] [CrossRef]
- Jensen, B. G.; Andersen, M. R.; Pedersen, M. H.; Frisvad, J. C.; Søndergaard, Ib. Hydrophobins from Aspergillus species cannot be clearly divided into two classes. BMC Res. Notes 2010, 3, 344. [Google Scholar] [CrossRef]
- Tanaka, T.; Terauchi, Y.; Yoshimi, A.; Abe, K. Aspergillus hydrophobins: Physicochemical properties, biochemical properties, and functions in solid polymer degradation. Microorganisms 2022, 10, 1498. [Google Scholar] [CrossRef]
- Zhang, X.; Zhao, L.; Liu, S.; Zhou, J.; Wu, Y.; Feng, Z.; Zhang, Y.; Zhu, H.; Wei, F.; Feng, H. Identification and functional analysis of a novel hydrophobic protein VdHP1 from Verticillium dahlia. Microbiol. Spectr. 2022, 10, e0247821–e0247821. [Google Scholar] [CrossRef]
- Moscatiello, R.; Sello, S.; Ruocco, M.; Barbulova, A.; Cortese, E.; Nigris, S.; Baldan, B.; Chiurazzi, M.; Mariani, P.; Lorito, M.; Navazio, L. The hydrophobin HYTLO1 secreted by the biocontrol fungus Trichoderma longibrachiatum triggers a NAADP-mediated calcium signaling pathway in Lotus japonicus. Int. J. Mol. Sci. 2018, 19, e2596. [Google Scholar] [CrossRef] [PubMed]
- Viterbo, A.; Chet, I. TasHyd1, a new hydrophobin gene from the biocontrol agent T. asperellum, is involved in plant root colonization. Mol. Plant Pathol. 2006, 7, 249–258. [Google Scholar] [CrossRef] [PubMed]
- Guzmán-Guzmán, P.; Alemán-Duarte, M. I.; Delaye, L.; Herrera-Estrella, A.; Olmedo-Monfil, V. Identification of effector-like proteins in Trichoderma spp. and role of a hydrophobin in the plant-fungus interaction and mycoparasitism. BMC Genet. 2017, 18, 16. [Google Scholar] [CrossRef] [PubMed]
- Shoresh, M.; Yedidia, I.; Chet, I. Involvement of jasmonic acid/ethylene signaling pathway in the systemic resistance induced in cucumber by Trichoderma asperellum T203. Phytopathology 2005, 95, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Szczech, M.; Nawrocka, J.; Felczyński, K.; Małolepsza, U.; Sobolewski, J.; Kowalska, B.; Maciorowski, R.; Jas, K.; Kancelista, A. Trichoderma atroviride, TRS25 isolate reduces downy mildew and induces systemic defense responses in cucumber in field conditions. Sci. Hortic. 2017, 224, 17–26. [Google Scholar] [CrossRef]
- Stefania, G.; Roberta, P.; Stefano, C. Selected isolates of Trichoderma gamsii induce different pathways of systemic resistance in maize upon Fusarium verticillioides challenge. Microbiol. Res. 2020, 233, 126406. [Google Scholar]
- Wang, K. D.; Borrego, E. J.; Kenerley, C. M.; Kolomiets, M. V. Oxylipins other than jasmonic acid are xylem-resident signals regulating systemic resistance induced by Trichoderma virens in Maize. Plant Cell 2020, 32, 166–185. [Google Scholar] [CrossRef]
- Martínez-Medina, A.; Fernández, I.; Sánchez-Guzmán, M. J.; Jung, S. C.; Pascual, J. A.; Pozo, M. J. Deciphering the hormonal signaling network behind the systemic resistance induced by Trichoderma harzianum in tomato. Front. Plant Sci. 2013, 4, 206. [Google Scholar] [CrossRef]
- Mathys, J.; De, Cremer, K. ; Timmermans, P.; Van, Kerckhove, S.; Lievens, B.; Vanhaecke, M.; Cammue, B. P. A.; De, Coninck, B. Genome-wide characterization of ISR induced in Arabidopsis thaliana by Trichoderma hamatum T382 against Botrytis cinerea infection. Front. Plant Sci. 2012, 3, 108. [Google Scholar] [CrossRef]
- Ojalvo, I.; Rokem, J. S.; Navon, G.; Goldberg, I. 31P NMR study of elicitor treated Ohaeeolus vulgaris cell suspension culture. Plant Physiol. 1987, 85, 716–719. [Google Scholar] [CrossRef]
- Pachten A, Barz W. Elicitor-stimulated oxidative burst and extracellular pH changes in protoplast suspensions prepared from cultured chickpea (Cicer arietinum L.) cells. J. Plant Physiol. 1999, 155, 795–797. [Google Scholar] [CrossRef]
- Bóka, K.; Orbán, N. New aspect of H2O2 signaling. Plant Signal Behav. 2007, 2, 498–500. [Google Scholar] [CrossRef] [PubMed]
- Yadav, V.; Wang, Z.; Wei, C.; Amo, A.; Ahmed, B.; Yang, X.; Zhang, X. Phenylpropanoid pathway engineering: an emerging approach towards plant defense. Pathogens 2020, 9, 312. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Zhu, L.; Tu, L.; Liu, L.; Yuan, D.; Jin, L.; Long, L.; Zhang, X. Lignin metabolism has a central role in the resistance of cotton to the wilt fungus Verticillium dahliae as revealed by RNA-Seq dependent transcriptional analysis and histochemistry. J. Exp. Bot. 2011, 62, 5607–5621. [Google Scholar] [CrossRef]
- Huang, J.; Gu, M.; Lai, Z.; Fan, B.; Shi, K.; Zhou, Y.H.; Yu, J.Q.; Chen, Z. Functional analysis of the Arabidopsis PAL gene family in plant growth, development, and response to environmental stress. Plant Physiol. 2010, 153, 1526–1538. [Google Scholar] [CrossRef]
- Marjamaa, K.; Kukkola, E. M.; Fagerstedt, K. V. The role of xylem class III peroxidases in lignification. J. Exp. Bot. 2009, 60, 367–376. [Google Scholar] [CrossRef]
- Chappell, J. Biochemistry and molecular biology of the isoprenoid biosynthetic pathway in plants. Annu. Rev. Plant Physiol. Mol. Biol. 1995, 46, 521–547. [Google Scholar] [CrossRef]
Figure 1.
Purified recombinant Thfb6 protein triggered hypersensitive response (HR) and reactive oxygen species (ROS) burst in tobacco leaves. (A) SDS-PAGE analysis of Thfb6 expression and purification. M: Protein molecular weight markers; lane 1: The cell-free extracts of E. coli; lane 2: soluble His-tagged Thfb6 protein; and lane 3, purified His-tagged Thfb6. (B) Tobacco leaves treated with 20 mM Tris-HCl buffer; (C) tobacco leaves treated with Thfb6 solution showing necrotic lesions at the injection site. The infiltration area in dotted circle of about 0.8 square centimeters. (D) control leaves treated with 20 mM Tris-HCl buffer; (E) Thfb6-treated leaves stained with DAB showing an obvious accumulation of brick-like substances. Scale bar =50 μm.
Figure 1.
Purified recombinant Thfb6 protein triggered hypersensitive response (HR) and reactive oxygen species (ROS) burst in tobacco leaves. (A) SDS-PAGE analysis of Thfb6 expression and purification. M: Protein molecular weight markers; lane 1: The cell-free extracts of E. coli; lane 2: soluble His-tagged Thfb6 protein; and lane 3, purified His-tagged Thfb6. (B) Tobacco leaves treated with 20 mM Tris-HCl buffer; (C) tobacco leaves treated with Thfb6 solution showing necrotic lesions at the injection site. The infiltration area in dotted circle of about 0.8 square centimeters. (D) control leaves treated with 20 mM Tris-HCl buffer; (E) Thfb6-treated leaves stained with DAB showing an obvious accumulation of brick-like substances. Scale bar =50 μm.
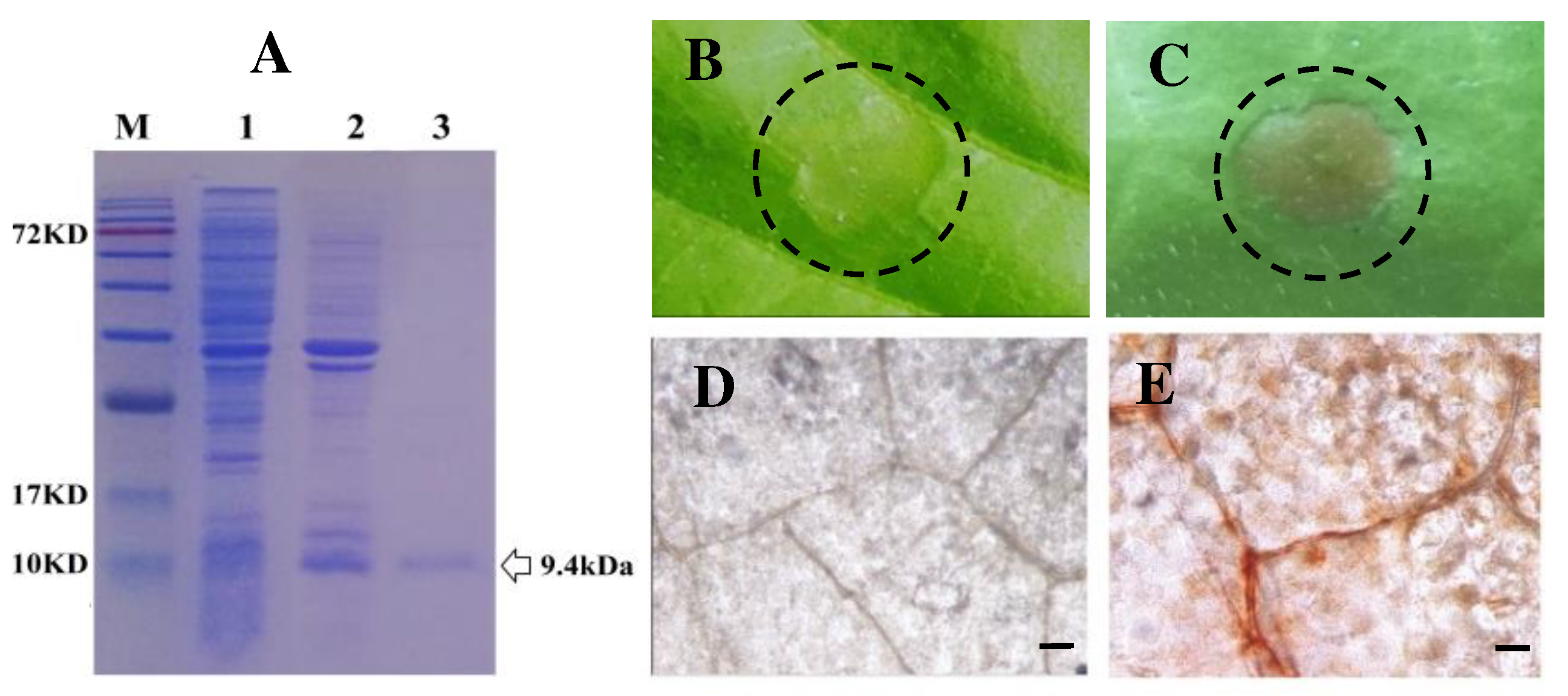
Figure 2.
Thfb6 induced callose accumulation in tobacco leaves and alkalization of extracellular medium of tobacco cells. (A) dot fluorescence produced by callose accumulation in Thfb6-treated tobacco leaves; (B) Tris-HCl buffer-treated tobacco leaves.Scale bar =50 μm; (C) pH variation of the tobacco cell suspensions. The error bars represent 6 SD of the mean.
Figure 2.
Thfb6 induced callose accumulation in tobacco leaves and alkalization of extracellular medium of tobacco cells. (A) dot fluorescence produced by callose accumulation in Thfb6-treated tobacco leaves; (B) Tris-HCl buffer-treated tobacco leaves.Scale bar =50 μm; (C) pH variation of the tobacco cell suspensions. The error bars represent 6 SD of the mean.
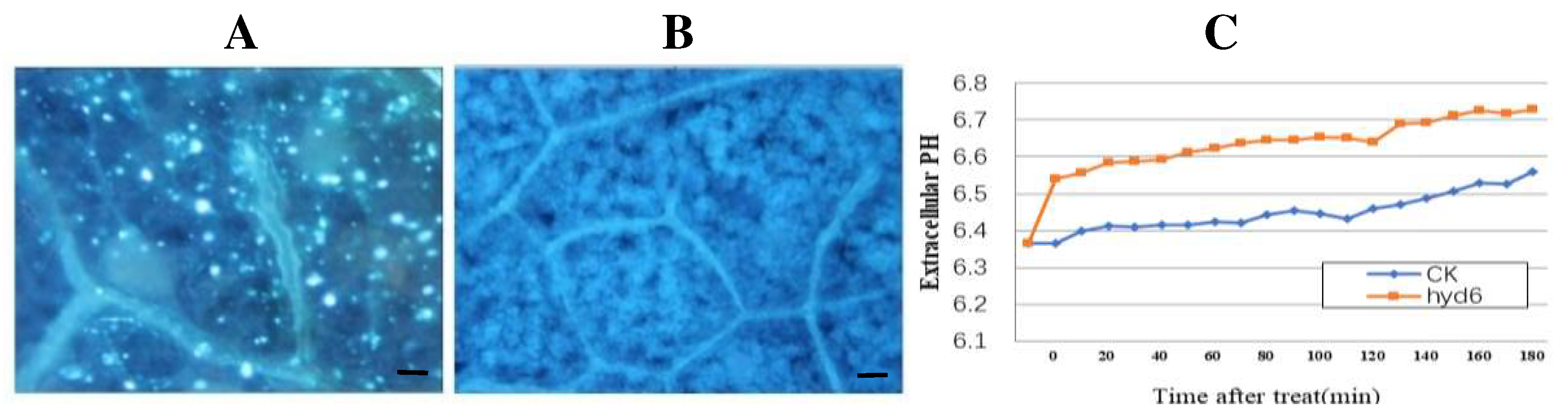
Figure 3.
Phenolics and lignin deposition induced by Thfb6 in tobacco cells. (A) 20 mM Tris-HCl buffer-treated tobacco cells as control; (B) Autofluorescence in Thfb6-treated in tobacco cells. Scale bar =2 μm; (C) lignin contents in Thfb6-treated and control tobacco cells suspensions. The asterisk denotes a statistical difference (p < 0.05) from control.
Figure 3.
Phenolics and lignin deposition induced by Thfb6 in tobacco cells. (A) 20 mM Tris-HCl buffer-treated tobacco cells as control; (B) Autofluorescence in Thfb6-treated in tobacco cells. Scale bar =2 μm; (C) lignin contents in Thfb6-treated and control tobacco cells suspensions. The asterisk denotes a statistical difference (p < 0.05) from control.
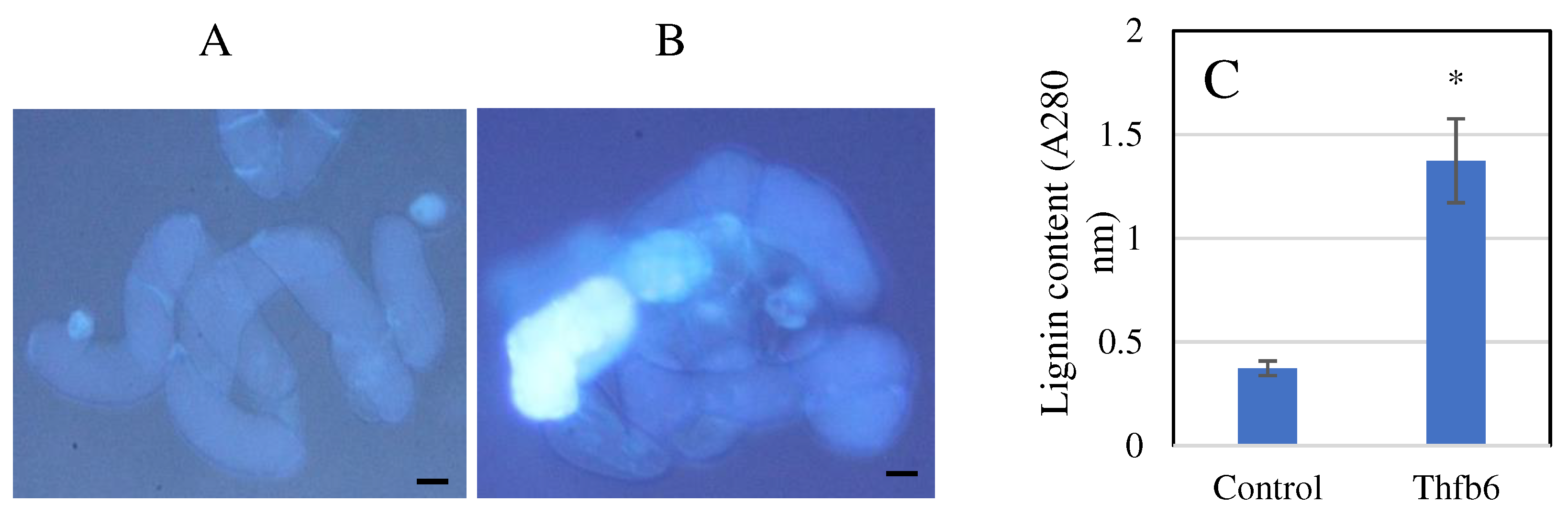
Figure 4.
Activity curves of phenylalanine ammonia-lyase (PAL), polyphenol oxidase (PPO), and peroxidase (POD) in tobacco leaves treated with Thfb6.
Figure 4.
Activity curves of phenylalanine ammonia-lyase (PAL), polyphenol oxidase (PPO), and peroxidase (POD) in tobacco leaves treated with Thfb6.
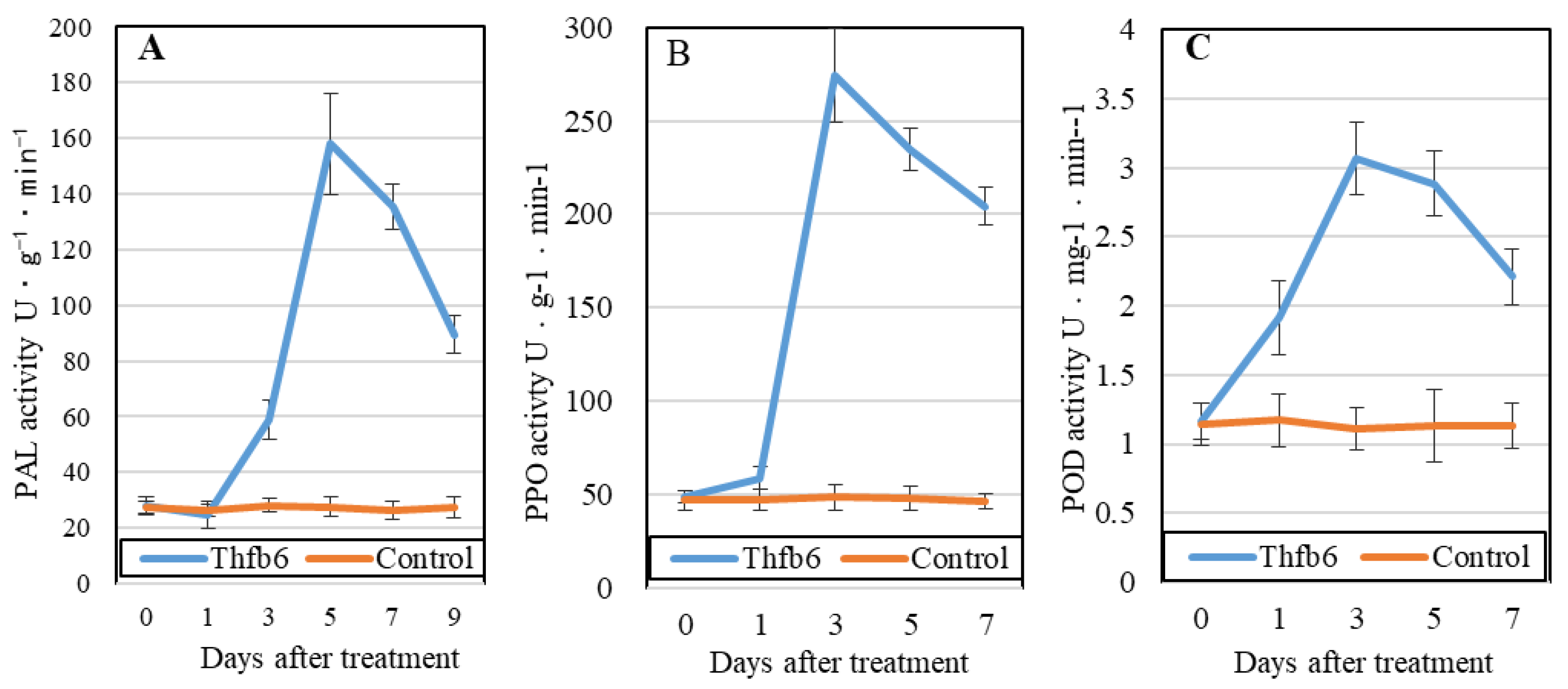
Figure 5.
Expression of resistance genes of tobacco induced by Thfb6.
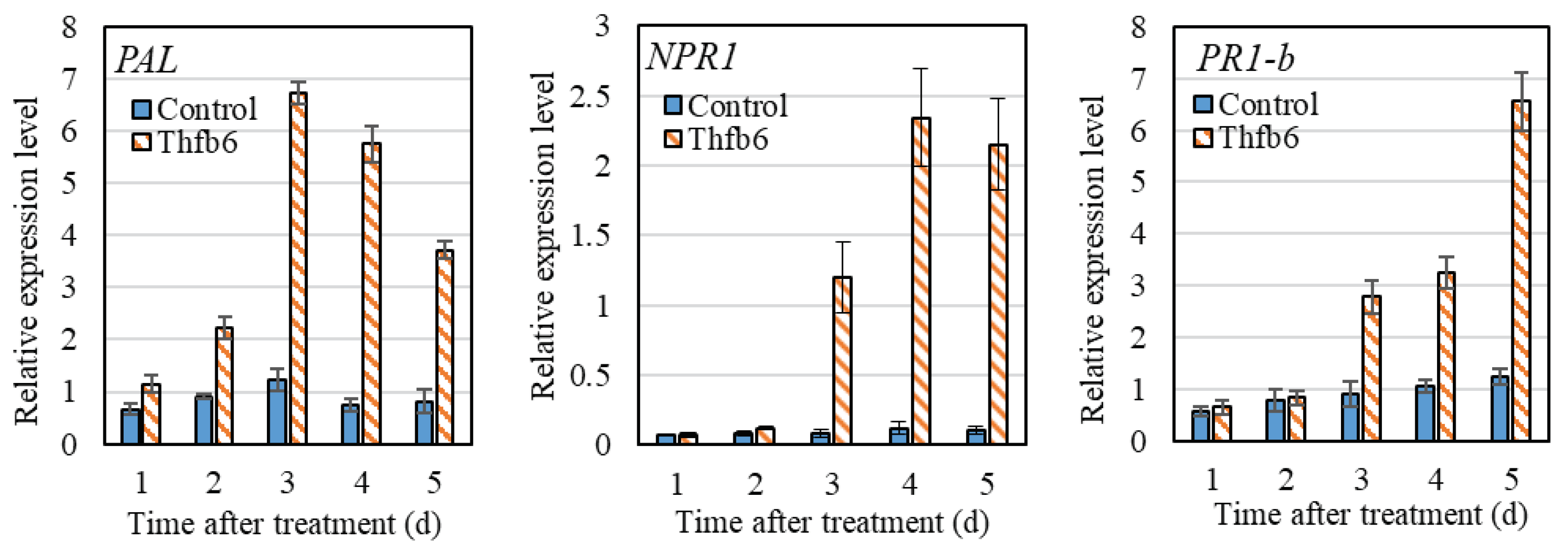

Table 1.
Primers used in the experiments. Primers designed to detect the expression of defense-related genes in tobacco leaves.
Table 1.
Primers used in the experiments. Primers designed to detect the expression of defense-related genes in tobacco leaves.
| Gene | Sequences 5′–3′ | Tm | GenBank accession no. |
|---|---|---|---|
| ß-Actin | Forward:TGTTATGGTTGGTATGGGTC Reverse: TCAGTAAGTAGCACAGGATG |
56°C | AB158612.1 |
| PAL | Forward:ACTCAGTGAACGACAACCC Reverse: TCAGATTAGATGGCAACCC |
56°C | AB289452.1 |
| PR-1b | Forward:TTTATGACGGCTGCTAAGG Reverse: CCTATGACAT TACCTGGAGG |
57°C | X12486.1 |
| NPR1 | Forward:CTGTATCTCTTGCTATGGCG Reverse: AATCTACTGTTGTCCTCTGT |
56°C | DQ837218.1 |
Table 2.
Thfb6 treatment reduced the occurrence and size of TMV lesions in tobacco leaves.
| dtp-TMV | Number of lesions | Diameter of lesions (mm) | ||||
|---|---|---|---|---|---|---|
| Control | Thfb6 | Inhibition (%) | Control | Thfb6 | Inhibition (%) | |
| 1 | 10.30 ± 2.10 | 7.10 ± 1.36 | 31.07 | 0.84 ± 0.10 | 0.66 ± 0.13 | 21.56* |
| 3 | 23.60 ± 4.12 | 15.10 ± 2.92 | 36.02** | 2.11 ± 0.15 | 1.23 ± 0.09 | 41.57** |
| 5 | 50.60 ± 5.72 | 36.83 ± 3.83 | 27.21* | 3.91 ± 0.23 | 2.30 ± 0.14 | 41.23** |
Data are representative of 10 plants per replicate. Asterisks denote statistically significant differences,* and ** indicate P < 0.05 and 0.01 respectively. dtp-TMV: days post TMV inoculation.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



