Submitted:
08 October 2023
Posted:
09 October 2023
You are already at the latest version
Abstract
Iris laevigata Fisch. is a perennial aquatic plant with strong cold resistance and showy flowers. However, the flowering period of a single flower is only two days, greatly limiting its potential use in landscaping and the cutting-flower industry. In addition, I. laevigata is often challenged with various abiotic stresses including high salinity and drought in its native habitat. Thus, breeding novel cultivars with prolonged flowering time and higher resistance to abiotic stress is of high importance. In this study, we first performed genome-wide identification of WRKYs, key transcription factors in modulating flowering time and abiotic stress responses. We overexpressed IlWRKY22 in Arabidopsis thaliana and found the flowering time was delayed in the transgenic plants. Molecular characterization further revealed that IlWRKY22 promotes the expression of CO and GA20OX genes in the photoperiod and gibberellin pathways, respectively, and inhibits the expression of SPL3 in the aging pathway. We also created transgenic Nicotiana tabacum overexpressing IlWRKY22, which showed significantly improved resistance to both salt and drought compared to control plants. Specifically, the photosynthetic rate, maximum potential quantum efficiency of Photosystem II (Fv/Fm), and chlorophyll content were higher in the transgenic plants, which was accompanied with higher antioxidant enzyme activity and lower levels of reactive oxygen species. Thus, our study revealed a unique dual function of IlWRKY22, an excellent candidate gene for breeding novel I. laevigata cultivars of desirable traits.
Keywords:
Iris laevigata
; WRKY transcription factor
; flowering time regulation
; salt stress
; drought stress
1. Introduction
A key event in the life cycle of angiosperms is the formation of flowers, which marks the transitions of plants from vegetative growth to reproductive growth [1]. It not only directly affects the growth, development, and yield of crop plants, but also has a great impact on horticultural plants as the flower is the most important ornamental part in landscaping [2]. Flowering is controlled by a very large and fine-tuned regulatory network, which integrates various exogenous and endogenous factors to determine the most appropriate flowering time [3,4,5].
On the other hand, plants are also challenged with multiple types of abiotic stress and must respond to survive. Among them, high salinity and drought are the main abiotic stresses that affect plant growth and development and even cause death [6]. Under high salinity, ions from the soil accumulate in the cytoplasm and thus disrupt the osmotic regulation. It further induces secondary oxidative stress, leading to the inhibition of photosynthesis and metabolism and ultimately a deduction in plant yield [7]. Similar to high salinity, drought causes osmotic stress and secondary stress including oxidative stress, damage to cellular components (e.g., membrane lipids, proteins, and nucleic acids), and metabolic dysfunction [8]. To cope with these abiotic stresses, evolution has equipped plants with a series of complex regulatory networks for survival [9].
The two seemingly unrelated processes of flowering and response to abiotic stress have one thing in common: both are under tight control of transcriptional factors (TFs). Indeed, the critical roles of TFs in flowering have been extensively studied [10]. In fact, some of the most important proteins in flowering pathways are TFs or transcriptional regulators including Constans (CO) in the photoperiod pathway and flowering locus C (FLC), a MADS-box TF that represses flowering. Similarly, TFs play a critical role in the responses to abiotic stresses [11]. By activating or inhibiting the expression of target genes, they regulate plant metabolism and adaptability to abiotic stress, leading to an increased interest in using TFs to combat environmental stresses in the face of global warming [12,13].
Interestingly, TFs may regulate multiple independent biological processes because they can interact with distinct sets of substrates. The WRKY family is a good example that controls a diverse range of physiological events via a complex gene network [14,15]. For instance, WRKY regulates plant tolerance to salt, drought, low temperature, high temperature, and other stresses as well as flowering by interacting with abscisic acid (ABA), jasmonic acid (JA), and ROS. WRKYs are named after the highly conserved WRKYGQK domain at the N-terminus [16]. The C terminus contains a CX4-7CX22 or 23HXH/C zinc finger that is important for protein interactions and DNA binding [17,18]. Based on the number of WRKY domains and the type of zinc finger structure, WRKYs are grouped into group Ⅰ (two WRKY domains and a C2H2 type zinc finger structure), group II (one WRKY domain and a C2H2 type zinc finger structure), and group III (one WRKY domain and a C2HC type zinc finger structure) [19]. Advances in sequencing have led to the genome-wide identification of WRKYs in many plant species including 74 members in Arabidopsis, 59 in grape, 127 in apple, 102 in rice, and 119 in corn [20].
Iris laevigata Fisch is native in Northeast China with strong cold resistance and beautiful flowers of unique shape and bright color[21]. Such desirable traits make I. laevigata a good candidate for both landscaping and industrial use as a cutting flower. However, the potential is dampened because of the short flowering period with only 2 days for a single flower and 20 days for the population[21]. I. laevigata is also sensitive to abiotic stress[22]. Thus, extending the flowering period and increasing the resistance to abiotic stress are two central goals in the field of breeding I. laevigata of novel properties[23,24]. One prerequisite to achieve these goals is to identify key genes controlling the target traits, which has been propelled tremendously by the advances in genome sequencing [25,26,27].
However, mechanistic studies at the molecular levels in Iris lag far behind compared with other plant species due to the lack of genome information. Our group pioneered the first transcriptomics studies in I. laevigata[21]. The dataset also allowed us to perform genome-wide identification of TFs. In this study, we report the identification of 68 WRKYs in I. laevigata, which are distributed to the three groups that regulate multiple biological processes. We further experimentally studied the involvement of IlWRKY22 in the content of both flowering and abiotic stress response. WRKY22 has been shown to play an important role in the defense against aphids[28], bacteria[29], low temperature[30], flowering[31] and senescence[32]. Our data demonstrated that IlWRKY22 is a negative regulator in flowering and a positive regulator in resistance to salt and drought. Thus, IlWRKY22 is a multifaceted TF that can be tweaked to improve multiple traits.
2. Results
2.1. Genome-Wide Identification of WRKY Genes in I. laevigata
We performed genome-wide mining of WRKY sequences in I. lavigata and found a total of 68 putative members (Figure 1). Phylogenetic analysis of the coded proteins showed that they are distributed to groups Ⅰ (n = 22), Ⅱa (n = 5), Ⅱb (n = 7), Ⅱc (n = 1), Ⅱd (n = 13), Ⅱe (n = 9), and Ⅲ (n = 11). By contrast, the 59 WRKY members in Arabidopsis thaliana showed a different pattern of distribution among the groups: groups Ⅰ (n = 11), Ⅱa (n = 2), Ⅱb (n = 7), Ⅱc (n = 16), Ⅱd (n = 6), Ⅱe (n = 6), and Ⅲ (n = 11). Given the distinctive roles of each group, this may indicate functional divergence of WRKY genes in I. lavigata.
2.2. Cloning of IlWRKY22 and Subcellular localization
We next cloned WRKY22 using I. lavigata cDNA with PCR. The sequence of IlWRKY22 was 840 bp, encoding a protein of 279 amino acids with a molecular weight of 30.6 kDa. The theoretical isoelectric point of the predicted protein was 5.02, with a total of 44 negatively (Asp and Glu) and 33 positively (Arg and Lys) charged residues, respectively. The instability index (II) of IlWRKY22 was 50.16, suggesting it was an unstable protein. In addition, IlWRKY22 protein was predicted to be hydrophilic due to the presence of more hydrophilic amino acid residues than hydrophobic ones. Sequence alignment showed that it contains the highly conserved WRKYGQK domain (Figure 2A) and it is most similar to WRKY22 from Asparagus officinalis (Figure 2B) with the C2H2 zinc finger[33], a typical feature of the group IIe of the WRKY family.
The IlWRKY22 protein was predicted to be in the nucleus. To verify this in vivo, we transiently expressed an IlWRKY22-GFP fusion protein in tobacco leaves. We found the fusion protein is exclusively expressed in the nucleus of epidermal cells, while fluorescence signals can be seen throughout the entire cell for the GFP-only as a negative control (Figure 2C). Thus, this data supported the prediction and agreed with the putative TF function of IlWRKY22.
2.3. Overexpressing IlWRKY22 in both Arabidopsis and tobacco
To study the in-planta function of IlWRKY22, we cloned the open reading frame (ORF) of IlWRKY22 into pCAMBIA1300. The resulting pCAMBIA1300: IlWRKY22-GFP construct was transformed into Agrobacterium, which was further used to generate transgenic Arabidopsis (via the flower dip method) and tobacco plants (via the leaf disk method). We obtained 14 positive transgenic Arabidopsis lines and 10 tobacco lines, respectively, via Hygromycin B selection and PCR verification (Figure S1). These plants were allowed to self-cross for two consecutive generations, and randomly selected T3 lines of Arabidopsis and tobacco were used for further experiments. RT-PCR showed much-enhanced expression levels of WRKY22 in these transgenic lines of both Arabidopsis (designated as OE1, OE4, and OE9) and tobacco (designated as OE2, OE3, and OE5) compared to the corresponding WT plants (Figure S2), three lines with high expression level were selected for follow-up experiment.
2.4. Overexpressing IlWRKY22 delays flowering in Arabidopsis thaliana
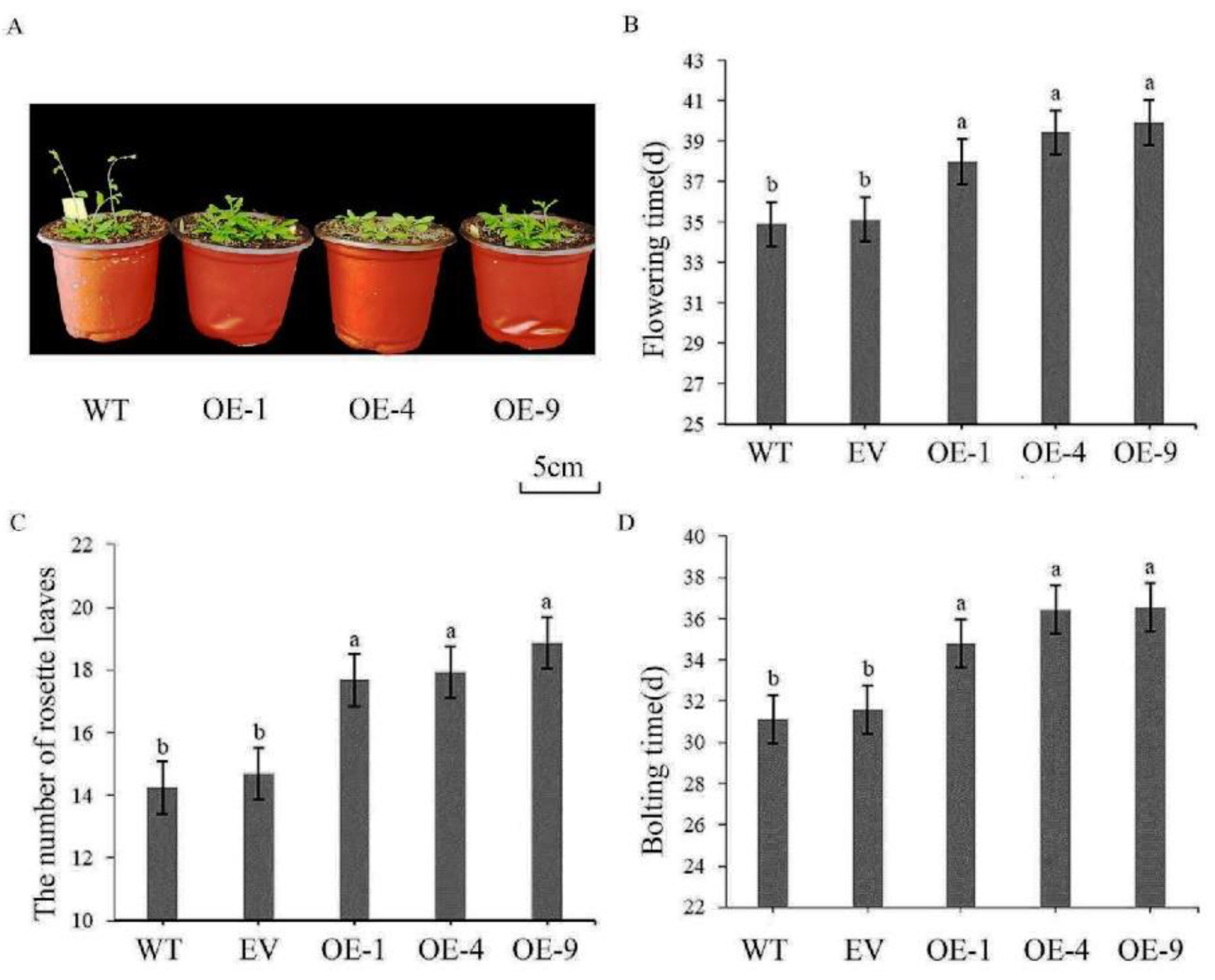
To investigate the impact of overexpressing IlWRKY22 on flowering, we compared the flowering statistics of WT and transgenic plants harboring IlWRKY22 or the empty vector (EV) under short-day conditions (Figure 4A). In WT, the bolting time, flowering time, and the number of rosette leaves were 31.0 d, 34.9 d, and 14.3, respectively. The EV plants showed very comparable numbers of 31.6 d, 35.1 d, and 14.8, respectively. Thus, the GV1300 vector had a negligible impact on the growth and development of Arabidopsis. By contrast, overexpressing IlWRKY22 significantly prolonged the bolting time to 35.8 d and the flowering time to 39.1 d with the number of rosette leaves of 18.1 (Figure 4B-D).
Figure 3.
A Phenotypic observation of transgenic Arabidopsis overexpressing IlWRKY22. B Flowering time. C The number of rosette leaves. D Bolting time. Bar graph data are Mean ± SD, marked with different lowercase letters to indicate significant differences (p < 0.05).
Figure 3.
A Phenotypic observation of transgenic Arabidopsis overexpressing IlWRKY22. B Flowering time. C The number of rosette leaves. D Bolting time. Bar graph data are Mean ± SD, marked with different lowercase letters to indicate significant differences (p < 0.05).
2.5. Overexpression of IlWRKY22 Modulates the Expression of Flowering Time Genes
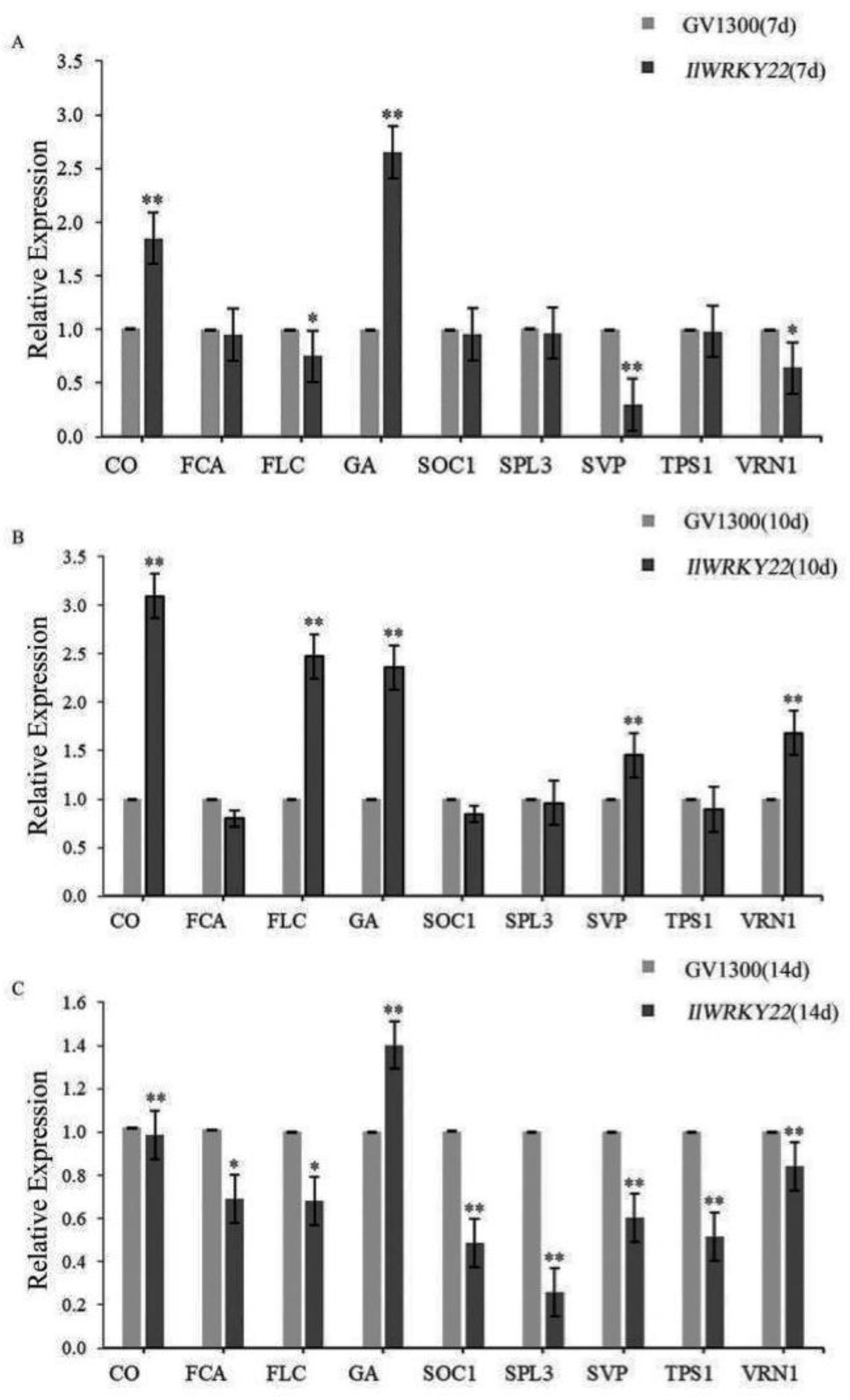
To understand the molecular mechanisms by which IlWRKY22 inhibits flowering, we quantified the mRNA levels of key genes in flowering pathways. We found significant differential expression levels of genes in the photoperiod pathway, gibberellin pathway, and temperature pathway including SVP, FLC, and VRN1 between the OE and EV plants in 7-day plants (Figure 4A). This suggested that IlWRKY22 may regulate these pathways in the early flowering transition. On day 10, the OE plants showed an increase in FLC (Figure 4B), a major flowering suppressor that can integrate the signals from SVP and VRN1 to inhibit the expression of FT. Such an up-regulation of FLC was in line with the delayed flowering phenotype. Among all the genes examined, only GA20OX was up-regulated in the OE plants (1.4-fold higher) compared to the EV plants at day 14, while all other genes showed lower expression levels in the OE plants (Figure 4C). Notably, SPL3 showed 80% down-regulation in the OE plants. This indicated that the OE plants were still at the early stage of flowering transition on day 14.
Figure 4.
Expression of flowering-related genes in 7- (A), 10- (B), and 14- (C) day-old seedlings. *: p < 0.05; **: p < 0.01.
Figure 4.
Expression of flowering-related genes in 7- (A), 10- (B), and 14- (C) day-old seedlings. *: p < 0.05; **: p < 0.01.
2.6. Overexpressing IlWRKY22 in Nicotiana tabacum enhances resistance to abiotic stress
IlWRKY22 is a multifaceted gene with many orthogonal functions during distinct biological processes. Thus, we next explored its role in abiotic stress[34]. Three lines of transgenic plants harboring IlWRKY22 (OE2, OE3, and OE5) were used, and both the WT plants and plants harboring pCAMBIA1300 (EV) were used as controls. All five lines showed similar overall phenotypes under normal conditions (Figure 5). Salt treatment, however, led to a more severe inhibition of growth in control plants compared to the EV plants. After 7 days post-treatment (dpt), the leaf growth rate of WT and EV lines was lower than that of the transgenic lines. At 14 dpt, transgenic lines were significantly taller than the control plants, which showed signs of yellowing and wilting of leaves. Under drought stress, WT and EV plants showed obvious wilting at 7 dpt and significantly lower plant height at 14 dpt compared to the transgenic lines.
2.7. Impact of overexpressing IlWRKY22 on photosynthesis under abiotic stress
The finding of the positive impact of IlWRKY22 on plant growth under abiotic stress prompted us to examine the photosynthesis performance between the transgenic and control plants. We quantified key parameters including the net photosynthetic rate, total chlorophyll content, chloroplast photosystem II photochemical maximum absorption (Fv/Fm), stomatal conductance, intercellular CO2 concentration, and transpiration rate. We found that all lines showed similar values at the beginning of the salt treatment. At both 7 and 14 dpt, the transgenic lines showed higher net photosynthetic rate, total chlorophyll content, and Fv/Fm, as well as lower stomatal conductance, intercellular CO2 content, and transpiration compared to the control plants(Figure 6A-F). For drought treatment, we found an overall decrease in the net photosynthetic rate, Fv/Fm, stomatal conductance, intercellular CO2 content, and transpiration rate in both the control and transgenic plants as the stress progressed. While the net photosynthetic rate, total chlorophyll content, and Fv/Fm were higher in the transgenic plants at both 7 and 14 dpt, stomatal conductance, intercellular CO2 content, and transpiration rate showed an opposite trend(Figure 6G-L). Thus, IlWRKY22 enhances abiotic resistance in transgenic tobacco by up-regulating the total chlorophyll content and reducing the stomatal opening.
2.8. The involvement of ROS regulation in IlWRKY22-enhanced abiotic stress resistance
Abiotic stress can lead to the production of reactive oxygen species (ROS) in plants, which oxidize lipids and generate malondialdehyde (MDA). On the other hand, ROS and their derivatives can be counterbalanced by the antioxidant system including superoxide dismutase (SOD), catalase (CAT), and peroxidase (POD). To study the involvement of the ROS system in IlWRKY22-enhanced abiotic stress resistance, we quantified the levels of MDA, O2-, and H2O2, as well the enzymatic activity of SOD, CAT, and POD in the leaf. We found increasing levels of MDA, O2-, and H2O2 in all plants as the salt/drought stress progressed (Figure 7A-C, G-I), indicating abiotic stress-induced oxidative stress in plants. Under both conditions, the OE plants showed significantly lower levels of MDA, O2-, and H2O2 at both 7 and 14 dpt compared to the control plants (p < 0.05). The NBT and DAB staining also visually showed less O2- and H2O2 accumulation in the OE plants compared to the WT and EV plants (Figure 7M-N). The lower ROS levels in the OE plants were accompanied with higher levels of SOD, CAT, and POD compared to the control plants (Figure 7D-F, J-L). This suggested that compared with the control, plants overexpressing IlWRKY22 had a stronger ability to scavenge ROS under salt and drought stress.
2.9. The IlWRKY22 on expression levels of stress-related genes NtCAT, NtHAK1, NtPMA4, NtPOD, NtSOD, and NtSOS1 under abiotic stress
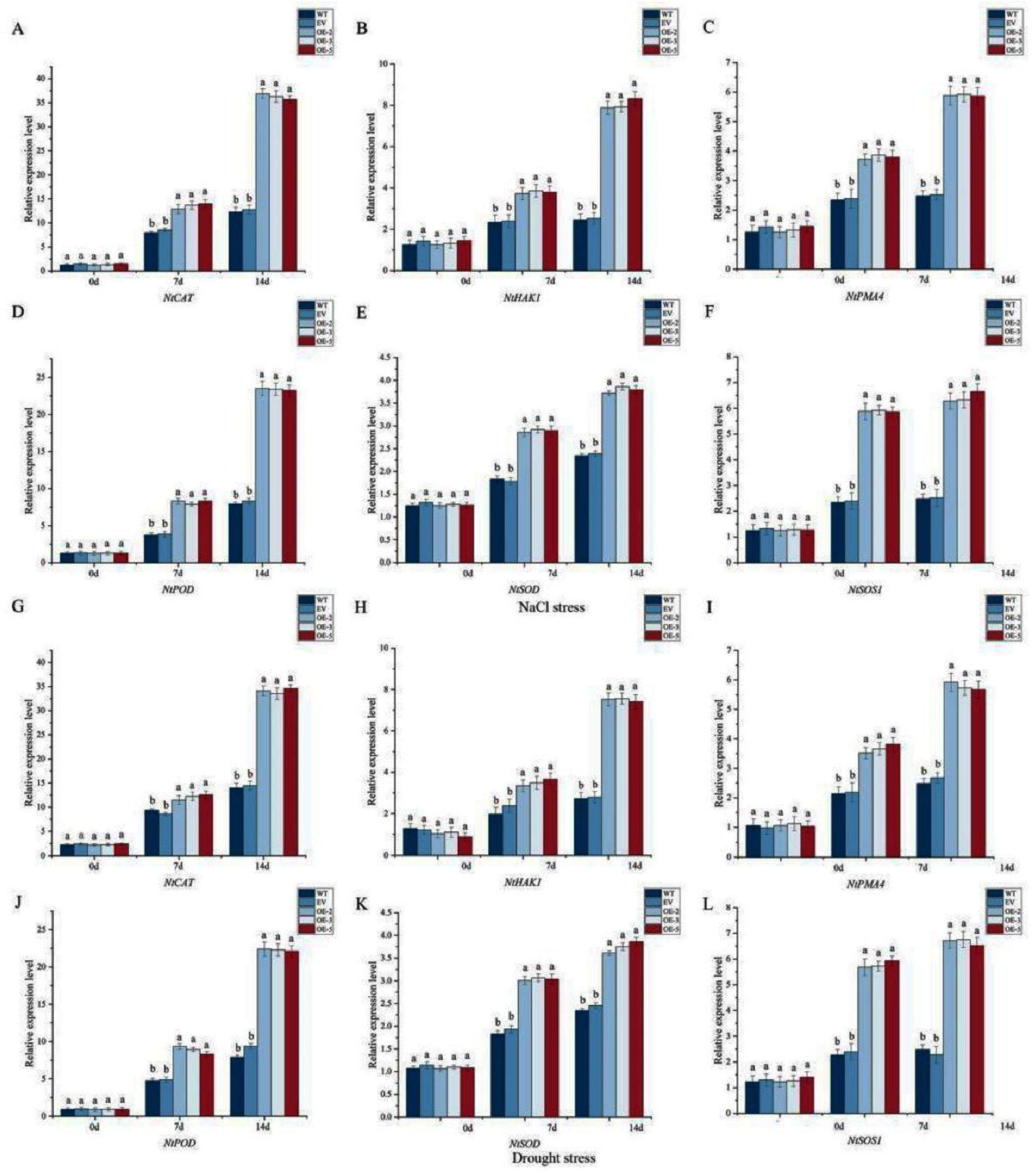
To further verify the role of IlWRKY22 in abiotic stress tolerance, we quantified the relative expression levels of stress-related genes. We found that the levels of NtSOD, NtPOD, NtCAT, NtHAK1, NtPMA4, and NtSOS1 were significantly up-regulated under abiotic stress regardless the genetic background (Figure 8). However, OE lines showed the highest degree of increase at both 7 and 14 dpt compared to that of the control plants. This indicated that IlWRKY22 could directly or indirectly regulate the expression of downstream stress response genes.
3. Discussion
Using our I. laevigata transcriptome data from the flower, we identified a total of 68 WRKYs and classified them into three groups: 22 in Group I, 35 in Group II, and 11 in Group III. These three groups of WRKYs may participate in various biological processes or pathways based on previous functional studies in multiple plant species (Table S2). Group I WRKYs are involved in plant disease resistance, abiotic stress (e.g., salt, cold, and heat stress) responses, senescence, and developmental processes (e.g., pollen development and seed coat development). Group II genes are mainly involved in defense responses to bacterial and fungal infection, abiotic stresses (salt, cadmium, drought stress) response, senescence, and developmental processes (e.g., lignin synthesis, pollen development, and flowering). They also participate in auxin transport and jasmonic acid signaling pathways. The biological function of Group III is also diverse, including defense against bacteria and/or fungi, abiotic stress (e.g., ozone, temperature, and drought), senescence, development of lateral roots and leaves, and interaction with plant hormone pathways (salicylic acid, abscisic acid, brassinosteroids, ethylene hormone, and jasmonic acid). The functional redundancy of these WRKYs indicates their key roles in responding to both external and internal stimuli [35]. It can also be reflected by the presence of redundant sequences in many plant species including Xanthoceras sorbifolium[36] and tomato[37], possibly due to duplication during plant evolution. Indeed, several sub-genome duplication events may have led to the formation of a large WRKY family in most modern eukaryotes[38].
Due to the large number of WRKYs in I. laevigata, we selected IlWRKY22 for functional studies because it is less explored compared to other WRKYs. We first focused on its role in flowering and found that overexpressing IlWRKY22 delays flowering in Arabidopsis. This provides another line of evidence to support the key roles of WRKYs in controlling flowering time. For example, heterologous expression of the cotton GbWRKY1 gene in Arabidopsis promotes flowering by regulating the transcription of SOC1[39]. Similarly, an up-regulation of BcWRKY22 in Chinese cabbage by low temperature promotes bolting and flowering via increasing BcSOC1[40]. In addition, a WRKY11 mutant in rice causes a late flowering phenotype [41]. Because flowering is a complex process that is sensitive to multiple environmental factors[42], such data collectively indicates that WRKYs can sense and integrate perturbations in the environment and initiate dynamic regulation of flowering [43].
Corresponding to the multiple environmental aspects affecting flowering, plants evolved multiple flowering transition pathways including the de-repression of CDF on CO and FT by the photoperiod pathway, the de-repression of FLC on FT in the vernalization, temperature, and autonomous pathways, as well as the de-repression of SOC1 by degrading DELLA proteins in the GA pathway[44]. To delineate the exact mechanisms in which IlWRKY22 was involved, we quantified the expression of 10 genes in multiple pathways (Figure 4). Our data showed a significant up-regulation of CO and GA20OX, indicating that the primary networks downstream of IlWRKY22 were the photoperiod pathway and the GA pathway. This in turn resulted in altered metabolism of other players for flowering control including SPL3. A lower-level expression of SPL3 in the transgenic plants may further lead to a down-regulation of SOC1, which ultimately causes a delay in flowering. Interestingly, SPL3 is a major effector in the aging pathway as overexpressing or knocking-out darktreated AtWRKY22 led to accelerated and delayed senescence phenotypes, respectively, in Arabidopsis[45]. Thus, it raises the possibility that IlWRKY22 regulates flowering transition via the aging pathway and that IlWRKY22 is the converging point of multiple pathways including flowering and senescence.
In addition to flowering control, our study further revealed a role of IlWRKY22 in responding to abiotic stress. Compared to controls, plants overexpressing IlWRKY22 showed much-improved growth under both salt and drought stress (Figure 5). This is in line with the finding that WRKY22 is significantly up-regulated by drought stress[46]. The enhanced resistance to abiotic stress can be attributed to up-regulation in photosynthesis (higher chlorophyll content, net photosynthesis, and Fv/Fm) and water retention capacity (lower stomatal conductance, intercellular CO2, and transpiration rate, Figure 6). Photosynthesis is an important indicator of the ability of plants to respond to abiotic stress[47]. Under high-salt and drought conditions, plants lose water due to high osmotic pressure, leading to stomatal closing, disrupted chloroplast structures, decreased activity of chlorophyll enzymes, and eventually inhibition of photosynthesis[48,49,50]. Our data suggested that the protective role of IlWRKY22 involved multiple processes that can alleviate the adverse impact of abiotic stress.
Although the exact molecular mechanisms by which IlWRKY22 protects plants against abiotic stress are not clear, one possible mechanism is via ROS regulation. This is evidenced by the observation that the transgenic plants overexpressing IlWRKY22 accumulate less , H2O2, and MDA compared to that of the control plants (Figure 7). The generation of ROS and secondary damage to the cell membrane including membrane lipid peroxidation under environmental challenges have been well-documented in plants[51,52]. By lowering the level of oxidative stress, IlWRKY22 may protect plants from excessive damage to the cellular membrane under stress. In addition, numerous studies have shown the key role of antioxidant enzymes including SOD, PDA, and CAT in counterbalancing ROS in plants[52,53]. Consistent with these studies, we found that the enzyme activities of SOD, CAT, and POD were increased in IlWRKY22-overexpressing plants under stress (Figure 7). In summary, both a decrease in ROS accumulation and an increase in antioxidant enzyme activity led to a lower level of oxidative stress under salt and/or drought treatment in transgenic plants, which contributed to a better photosynthetic capacity and enhanced resistance under unfavorable conditions.
4. Materials and methods
4.1. Plant materials
I. lavigata was planted in the nursery of the School of Landscape Architecture, Northeast Forestry University. Growth conditions were the same as described before [21].Nicotiana tabacum seeds were disinfected with 75% alcohol (LIRCON) for 1 minute and rinsed with water three times. Seeds were then disinfected with 2% NaClO (XILONG SCIENTIFIC) for 10 minutes and rinsed with water five times. Seeds were cultured in MS (Hope Bio-Technology) at 25°C with a photoperiod of 14 hours light/10 hours dark. Plants of 4-6 true leaves were used for transformation.
Seeds of Arabidopsis thaliana were surface-sterilized in 75% alcohol (LIRCON) for 1 minute and rinsed with water. Seeds were then disinfected in 0.8% NaClO (XILONG SCIENTIFIC) for 10 min and rinsed with water. Sterilized seeds were sowed on 1/2 MS (Hope Bio-Technology), and vernalized at 4°C in the dark for 2 days, followed by cultivation at a greenhouse. After 7-9 days, seedlings were transferred into a potting mix (peat soil/vermiculite/perlite of 5/3/2) and allowed to grow for 3 weeks before transformation. Growth conditions included 16 h light/8 h dark and 20-22°C.
4.2. Gene Sequence Identification and Phylogenetic Analysis
Putative WRKY sequences were identified from the transcriptome data of I. laevigata [54] with the online annotation tool from BGI (https://www.bgi.com/home). The open reading frames of these sequences were determined by the ORF Finder tool (https://www.ncbi.nlm.nih.gov/orffinder/), and the conserved domains were identified by CD Search tool (https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi). Putative sequences were then subject to BLAST (https://blast.ncbi.nlm.nih.gov/Blast.cgi), and then a final list of WRKY in I. laevigata were obtained. Phylogenetic analysis was performed in MEGA X using WRKY sequences from both I. laevigata and Arabidopsis (obtained from https://phytozome-next.jgi.doe.gov/). Sequence alignment was performed using Clustal W. A Phylogenetic tree was built using the neighbor-joining (NJ) method with 1000 bootstraps, and then annotated using iTOL (https://itol.embl.de/).
4.3. Gene cloning and sequence analysis
Total flower RNA was extracted using a kit (KANGWEISHIJI), and the integrity was checked by electrophoresis. Frist-strand cNA was synthesized using the Primer Script TM RT kit (TakaRa). IlWRKY22 (accession number: ON399552) was cloned using KOD-plus-neo (Toyobo) with primers (Ruibo Kexing) listed in Table S1. Both the PCR product and the pCAMBIA1300 vector were digested with SalLI and BamHI, which were then used to create pCAMBIA1300-IlWRKY70-GFP using ClonExpressII (Vazyme). The recombinant vector was transformed into E. coli DH5α (WEIDI) for sequencing. Positive plasmids were transferred into Agrobacterium tumefaciens GV3101 (WEIDI) for subsequent infection experiments. Bioinformatics analysis of IlWRKY22 was performed as described[21]. BLASTp was used to search for homologous sequences.
4.4. Subcellular localization
The subcellular localization of WRKY22 was determined as described[55]. Briefly, bacteria GV3101 (WEIDI) harboring pCAMBIA1300-Pro35S:IlWRKY22-GFP were cultured and collected. After resuspending in 10 mMMaCl2 to an OD600 of 1.5, acetosyringone was added to a final concentration of 200 µM. The bacterial solution was activated for 3 hours in the dark, and then injected to the lower side of N. benthamiana leaves using a syringe. Expression of the fusion protein WRKY22-GFP was visualized under a microscope in the transformed epidermal cells[21].
4.5. Plant transformation
Transgenic Arabidopsis plants were created using a method reported before[56]. Seeds of transformed plants were selected on 1/2 MS medium supplemented with 25 mg/L Hyg. After 7-9 days, T1 seedlings that survived on the screening medium were transferred to a potting mix (peat soil/vermiculite/perlite of 3/2/1) and then cultured at 20°C with a photoperiod of 16 h/8 h (light/dark). Transgenic plants were then screened by PCR, the primers (Ruibo Kexing) listed in Table S1. Homozygous lines were obtained by three consecutive generations of screening. IlWRKY22 in the leaf was determined by qRT-PCR using ACT2 (AT3G18780) as the internal reference, the primers (Ruibo Kexing) listed in Table S1. Three lines with high expression levels were selected.
Agrobacterium-mediated transformation of tobacco leaf disk and screening of the resulting transgenic plants were performed according to a previously published method [57]. Genomic DNA was extracted using a modified CTAB method. The copy number of IlWRKY22 was determined by the segregation ratio, and two homozygous transgenic lines with a single copy of IlWRKY22 were used for subsequent physiological experiments.
4.6. Determination of flowering phenotypes and gene expression
4.7. Abiotic stress treatment
Transgenic plants harboring IlWRKY22 or the empty vector, as well as the wild-type plants were used for abiotic stress treatment. Plants were first grown in a potting mix (peat soil/perlite/vermiculite of 3/1/1) at 28°C, a relative humidity of 75%, and with a photoperiod of 16 h/8 h (light/dark) for 30 days. Plants with similar overall fitness were then subjected to abiotic stress treatment. For salt treatment, plants were watered with 100 ml of 300 mM NaCl every two days. For drought, plants were withheld from watering. Leaves were sampled at 0, 5 and 10 days after the treatment.
4.8. Photosynthetic parameters in tobacco s under Abiotic stress
A Li-6400 portable photosynthesis instrument was used to quantify various photosynthetic parameters including net photosynthetic rate, stomatal conductance, intercellular CO2 concentration and transpiration rate. Three individual plants were randomly selected from each line (transgenic, empty vector control, and wild-type plants), and the 3rd and 4th leaf were used for measurement[63]. The Li-6400 instrument was calibrated before taking measurements and data was recorded after the reading was stable. Five measurements were taken for each leaf. The total chlorophyll content was determined as described, using the acetone extraction method[63].
4.9. Other physiological measurements of abiotic-treated tobacco plants
Commercially available kits from Grace Biotechnology were used for the quantification of catalase (catalog number G0105F, and the same hereafter), hydrogen peroxide (G0112F), and O2- (G0116F). Malondialdehyde (MDA), superoxide dismutase (SOD) activity, and peroxidase (POD) activity were determined according to a previous report[57]. Briefly, 0.5 g of leaves were homogenized in 5 mL of pre-cooled 50 mM PBS, pH 7.8 in a pre-cooled mortar. After centrifugation at 3000 rpm, 4°C, the supernatant was used. p-Nitro-Blue tetrazolium chloride (NBT) and 3,3′-diaminobenzidine tetrahydrochloride (DAB) staining were performed as described[57].
4.10. Real-time RT-PCR analyses
The expression patterns of IlWRKY22 in different parts of I. laevigata were measured from roots, tubers and leaves of normal growth, and the response of IlWRKY22 in leaves of Iris laevigata 0, 1, 3, 6, 12 and 24 hours after salt and drought stress were measured. Tobacco leaves with positive overexpression of IlWRKY22 at 30 days of growth will be used to measure the expression of IlWRKY22. The expression levels of stress response genes (NtCAT, NtHAK1, NtPMA4, NtPOD, NtSOD, and NtSOS1) were measured in WT, EV and 3 overexpressed IlWRKY22 lines of tobacco leaves under 0, 7, and 14 days of NaCl and drought stress. The method of total RNA extraction and reverse transcription to cDNA is the same as the method of total RNA extraction and reverse transcription above. Actin genes are the internal reference genes of I. laevigata and tobacco, respectively. Real-Time RT-PCR was performed by SYBR Green I method. The specific experimental procedures were followed according to the instructions of UltraSYBR Mixture (CWBIO). The results were calculated as 2 -△△CT, the primers (Ruibo Kexing) listed in Table S1.
4.11. Statistical Analyses
Statistical analyses were performed in SPSS (version 26), and the least significant difference method was used to test the significance at an alpha level of 0.05. Origin (version 2021) was used for plotting.
5. Conclusions
We conclude that IlWRKY22 has the dual function of flowering time control and regulation in abiotic stress. Overexpressing IlWRKY22 in Arabidopsis causes a delay in flowering via modulation of the expression of CO, GA20OX, SPL3, and SOC1. In addition, overexpression of IlWRKY22 in tobacco enhances the tolerance to salt and drought treatment by positively regulating the antioxidant system and photosynthesis. Thus, IlWRKY22 is a good candidate for breeding Iris of prolonged flowering time and enhanced capacity under abiotic stress.
Supplementary Materials
Supplemental Figure 1. A Positive validation gel plot of IlWRKY22 transgenic T0 generation plants, M is Maker of length 2000, H2O is negative control, P is pCAMBIA1300: IlWRKY22-GFP plasmid positive control, WT is WT Arabidopsis, and 1-14 is T0 transgenic Arabidopsis. B Positive validation gel plot of IlWRKY22 transgenic T0 generation plants, M is Maker of length 2000, H2O is negative control, P is pCAMBIA1300: IlWRKY22-GFP plasmid positive control, WT is WT tobacco, and 1-14 is T0 transgenic tobacco. Supplemental Figure 2. A Relative expression of IlWRKY22 in fourteen T3 transgenic Arabidopsis lines. B Relative expression of IlWRKY22 in ten T3 transgenic tobacco lines.
Author Contributions
This work was supported by Lijuan Fan, Zhaoqian Niu, Guiling Liu, Gongfa Shi, Ziyi Song, Qianqian Yang, Sheng Zhou, Ling Wang.
Funding
This study was supported by the Fundamental Research Funds for the Central Universities (2572023CT18) and the Natural Fund Project of Heilongjiang Province (LH2021C017).
Data Availability Statement
Data from this study are available from the corresponding author upon reasonable request.
Conflicts of Interest
We declare no conflict of interest.
References
- Wellmer, F.; Riechmann, J.L. Gene networks controlling the initiation of flower development. Trends Genet. 2010, 26, 519-527. [CrossRef]
- Rehman, S.; Bahadur, S.; Xia, W. An overview of floral regulatory genes in annual and perennial plants. Gene 2023, 885, 147699. [CrossRef]
- He, Y.; Chen, T.; Zeng, X. Genetic and Epigenetic Understanding of the Seasonal Timing of Flowering. Plant Commun 2020, 1, 100008. [CrossRef]
- Wang, F.; Li, S.; Kong, F.; Lin, X.; Lu, S. Altered regulation of flowering expands growth ranges and maximizes yields in major crops. Front. Plant Sci. 2023, 14, 1094411. [CrossRef]
- Kozlov, K.; Singh, A.; Berger, J.; Bishop-von, W.E.; Kahraman, A.; Aydogan, A.; Cook, D.; Nuzhdin, S.; Samsonova, M. Non-linear regression models for time to flowering in wild chickpea combine genetic and climatic factors. BMC Plant Biol. 2019, 19, 94. [CrossRef]
- Sun, Y.; Zhou, J.; Guo, J. Advances in the knowledge of adaptive mechanisms mediating abiotic stress responses in Camellia sinensis. Front Biosci (Landmark Ed) 2021, 26, 1714-1722. [CrossRef]
- Retraction: Exogenous application of moringa leaf extract improves growth, biochemical attributes, and productivity of late-sown quinoa. PLoS One 2022, 17, e272392.
- Zhu, J.K. Abiotic Stress Signaling and Responses in Plants. Cell 2016, 167, 313-324. [CrossRef]
- Su Wei; Xiao Liuying; Sun Guangwen; Liu Houcheng; Song Shiwei; Chen Riyuan.Functional analysis of transcription factor BcWRKY22 in low temperature-induced bolting and flowering of Brassica campestris.Molecular Plant Breeding. 2020, 18, 3862-3870.
- Shah, S.; Weinholdt, C.; Jedrusik, N.; Molina, C.; Zou, J.; Grosse, I.; Schiessl, S.; Jung, C.; Emrani, N. Whole-transcriptome analysis reveals genetic factors underlying flowering time regulation in rapeseed (Brassica napus L.). Plant Cell Environ. 2018, 41. [CrossRef]
- Golldack, D.; Luking, I.; Yang, O. Plant tolerance to drought and salinity: stress regulating transcription factors and their functional significance in the cellular transcriptional network. Plant Cell Rep. 2011, 30, 1383-1391. [CrossRef]
- Fedoroff, N.V.; Battisti, D.S.; Beachy, R.N.; Cooper, P.J.; Fischhoff, D.A.; Hodges, C.N.; Knauf, V.C.; Lobell, D.; Mazur, B.J.; Molden, D. et al. Radically rethinking agriculture for the 21st century. Science 2010, 327, 833-834. [CrossRef]
- Xia, Y.; Huang, G.; Zhu, Y. Sustainable plant disease control: biotic information flow and behavior manipulation. Sci. China Life Sci. 2019, 62, 1710-1713. [CrossRef]
- Phukan, U.J.; Jeena, G.S.; Shukla, R.K. WRKY Transcription Factors: Molecular Regulation and Stress Responses in Plants. Front. Plant Sci. 2016, 7, 760. [CrossRef]
- Khoso, M.A.; Hussain, A.; Ritonga, F.N.; Ali, Q.; Channa, M.M.; Alshegaihi, R.M.; Meng, Q.; Ali, M.; Zaman, W.; Brohi, R.D. et al. WRKY transcription factors (TFs): Molecular switches to regulate drought, temperature, and salinity stresses in plants. Front. Plant Sci. 2022, 13, 1039329. [CrossRef]
- Agarwal, P.; Reddy, M.P.; Chikara, J. WRKY: its structure, evolutionary relationship, DNA-binding selectivity, role in stress tolerance and development of plants. Mol. Biol. Rep. 2011, 38, 3883-3896. [CrossRef]
- Khoso, M.A.; Hussain, A.; Ritonga, F.N.; Ali, Q.; Channa, M.M.; Alshegaihi, R.M.; Meng, Q.; Ali, M.; Zaman, W.; Brohi, R.D. et al. WRKY transcription factors (TFs): Molecular switches to regulate drought, temperature, and salinity stresses in plants. Front. Plant Sci. 2022, 13, 1039329. [CrossRef]
- Jiang, J.; Ma, S.; Ye, N.; Jiang, M.; Cao, J.; Zhang, J. WRKY transcription factors in plant responses to stresses. J. Integr. Plant Biol. 2017, 59, 86-101. [CrossRef]
- Chen, X.; Li, C.; Wang, H.; Guo, Z. WRKY transcription factors: evolution, binding, and action. Phytopathology Research 2019, 1. [CrossRef]
- Bakshi, M.; Oelmuller, R. WRKY transcription factors: Jack of many trades in plants. Plant Signal Behav 2014, 9, e27700. [CrossRef]
- Liu, G.; Li, F.; Shi, G.; Wang, L.; Wang, L.; Fan, L. Identification of MADS-Box Transcription Factors in Iris laevigata and Functional Assessment of IlSEP3 and IlSVP during Flowering. Int. J. Mol. Sci. 2022, 23. [CrossRef]
- Yang, J.; Yu, S.; Shi, G.F.; Yan, L.; Lv, R.T.; Ma, Z.; Wang, L. Comparative analysis of R2R3-MYB transcription factors in the flower of Iris laevigata identifies a novel gene regulating tobacco cold tolerance. Plant Biol (Stuttg) 2022, 24, 1066-1075. [CrossRef]
- Chen, K.; Wang, Y.; Zhang, R.; Zhang, H.; Gao, C. CRISPR/Cas Genome Editing and Precision Plant Breeding in Agriculture. Annu. Rev. Plant Biol. 2019, 70, 667-697. [CrossRef]
- Kourelis, J.; van der Hoorn, R.; Sueldo, D.J. Decoy Engineering: The Next Step in Resistance Breeding. Trends Plant Sci. 2016, 21, 371-373. [CrossRef]
- Marks, R.A.; Hotaling, S.; Frandsen, P.B.; VanBuren, R. Representation and participation across 20 years of plant genome sequencing. Nat Plants 2021, 7, 1571-1578. [CrossRef]
- Michael, T.P.; Alba, R. The tomato genome fleshed out. Nat. Biotechnol. 2012, 30, 765-767. [CrossRef]
- The tomato genome sequence provides insights into fleshy fruit evolution. Nature 2012, 485, 635-641. [CrossRef]
- Coustham, V.; Li, P.; Strange, A.; Lister, C.; Song, J.; Dean, C. Quantitative modulation of polycomb silencing underlies natural variation in vernalization. Science 2012, 337, 584-587. [CrossRef]
- Zhang, X.; Bernatavichute, Y.V.; Cokus, S.; Pellegrini, M.; Jacobsen, S.E. Genome-wide analysis of mono-, di- and trimethylation of histone H3 lysine 4 in Arabidopsis thaliana. Genome Biol 2009, 10, R62. [CrossRef]
- He, Y. Control of the transition to flowering by chromatin modifications. Mol. Plant 2009, 2, 554-564. [CrossRef]
- Srikanth, A.; Schmid, M. Regulation of flowering time: all roads lead to Rome. Cell. Mol. Life Sci. 2011, 68, 2013-2037. [CrossRef]
- Eriksson, S.; Bohlenius, H.; Moritz, T.; Nilsson, O. GA4 is the active gibberellin in the regulation of LEAFY transcription and Arabidopsis floral initiation. Plant Cell 2006, 18, 2172-2181. [CrossRef]
- Khoso, M.A.; Hussain, A.; Ritonga, F.N.; Ali, Q.; Channa, M.M.; Alshegaihi, R.M.; Meng, Q.; Ali, M.; Zaman, W.; Brohi, R.D. et al. WRKY transcription factors (TFs): Molecular switches to regulate drought, temperature, and salinity stresses in plants. Front. Plant Sci. 2022, 13, 1039329. [CrossRef]
- Ahmad, I.; Zhu, G.; Zhou, G.; Liu, J.; Younas, M.U.; Zhu, Y. Melatonin Role in Plant Growth and Physiology under Abiotic Stress. Int. J. Mol. Sci. 2023, 24. [CrossRef]
- Chen, H.; Li, X.; Li, F.; Li, D.; Dong, Y.; Fan, Y. Bioinformatics Analysis of WRKY Family Genes in Erianthus fulvus Ness. Genes (Basel) 2022, 13. [CrossRef]
- Liu Zhi.Identification of WRKY transcription factor family in Xanthoceras sorbifolia and analysis of response patterns to abiotic stresses. Northeast Forestry University. 2020, 16.
- Liu Yuxuan. Identification and expression analysis of WRKY gene family in tomato. Shenyang Agricultural University. 2020, 13.
- Zhang, Y.; Wang, L. The WRKY transcription factor superfamily: its origin in eukaryotes and expansion in plants. BMC Evol. Biol. 2005, 5, 1. [CrossRef]
- Luo, X.; Li, C.; He, X.; Zhang, X.; Zhu, L. ABA signaling is negatively regulated by GbWRKY1 through JAZ1 and ABI1 to affect salt and drought tolerance. Plant Cell Rep. 2020, 39, 181-194. [CrossRef]
- Wang, Y.; Huang, X.; Huang, X.; Su, W.; Hao, Y.; Liu, H.; Chen, R.; Song, S. BcSOC1 Promotes Bolting and Stem Elongation in Flowering Chinese Cabbage. Int. J. Mol. Sci. 2022, 23. [CrossRef]
- Cai, Y.; Chen, X.; Xie, K.; Xing, Q.; Wu, Y.; Li, J.; Du C; Sun, Z.; Guo, Z. Dlf1, a WRKY transcription factor, is involved in the control of flowering time and plant height in rice. PLoS One 2014, 9, e102529. [CrossRef]
- Amasino, R.M.; Michaels, S.D. The timing of flowering. Plant Physiol. 2010, 154, 516-520. [CrossRef]
- Wang, H.; Chen, W.; Xu, Z.; Chen, M.; Yu, D. Functions of WRKYs in plant growth and development. Trends Plant Sci. 2023, 28, 630-645. [CrossRef]
- Liu, Y.; Yang, J.; Yang, M. [Pathways of flowering regulation in plants]. Sheng Wu Gong Cheng Xue Bao 2015, 31, 1553-1566.
- Zhou, X.; Jiang, Y.; Yu, D. WRKY22 transcription factor mediates dark-induced leaf senescence in Arabidopsis. Mol. Cells 2011, 31, 303-313. [CrossRef]
- Balti, I.; Benny, J.; Perrone, A.; Caruso, T.; Abdallah, D.; Salhi-Hannachi, A.; Martinelli, F. Identification of conserved genes linked to responses to abiotic stresses in leaves among different plant species. Funct. Plant Biol. 2020, 48, 54-71.
- Anuradha, M.; Sivaraju, K.; Krishnamurthy, V. Effect of waterlogging on physiological characteristics, yield and quality of flue-cured tobacco. Indian Journal of Plant Physiology 2013, 18, 67-70. [CrossRef]
- Elshoky, H.A.; Yotsova, E.; Farghali, M.A.; Farroh, K.Y.; El-Sayed, K.; Elzorkany, H.E.; Rashkov, G.; Dobrikova, A.; Borisova, P.; Stefanov, M. et al. Impact of foliar spray of zinc oxide nanoparticles on the photosynthesis of Pisum sativum L. under salt stress. Plant Physiol Biochem 2021, 167, 607-618. [CrossRef]
- Razi, K.; Muneer, S. Drought stress-induced physiological mechanisms, signaling pathways and molecular response of chloroplasts in common vegetable crops. Crit. Rev. Biotechnol. 2021, 41, 669-691. [CrossRef]
- Zahra, N.; Al, H.M.; Hafeez, M.B.; Rehman, A.; Wahid, A.; Siddique, K.; Farooq, M. Regulation of photosynthesis under salt stress and associated tolerance mechanisms. Plant Physiol Biochem 2022, 178, 55-69. [CrossRef]
- Skopelitis, D.S.; Paranychianakis, N.V.; Paschalidis, K.A.; Pliakonis, E.D.; Delis, I.D.; Yakoumakis, D.I.; Kouvarakis, A.; Papadakis, A.K.; Stephanou, E.G.; Roubelakis-Angelakis, K.A. Abiotic stress generates ROS that signal expression of anionic glutamate dehydrogenases to form glutamate for proline synthesis in tobacco and grapevine. Plant Cell 2006, 18, 2767-2781. [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol Biochem 2010, 48, 909-930. [CrossRef]
- Jaleel, C.A.; Riadh, K.; Gopi, R.; Manivannan, P.; Inès, J.; Al-Juburi, H.J.; Chang-Xing, Z.; Hong-Bo, S.; Panneerselvam, R. Antioxidant defense responses: physiological plasticity in higher plants under abiotic constraints. Acta Physiol. Plant. 2009, 31, 427-436. [CrossRef]
- Yang, J.; Yu, S.; Shi, G.F.; Yan, L.; Lv, R.T.; Ma, Z.; Wang, L. Comparative analysis of R2R3-MYB transcription factors in the flower of Iris laevigata identifies a novel gene regulating tobacco cold tolerance. Plant Biol (Stuttg) 2022, 24, 1066-1075. [CrossRef]
- Collings, D.A. Subcellular localization of transiently expressed fluorescent fusion proteins. Methods Mol Biol 2013, 1069, 227-258. [CrossRef]
- Wang, H.; Liu, Z.; Xie, J.; Li, J.; Zhang, J.; Yu, J.; Hu, L.; Zhang, G. The CaALAD Gene From Pepper (Capsicum annuum L.) Confers Chilling Stress Tolerance in Transgenic Arabidopsis Plants. Front. Plant Sci. 2022, 13, 884990. [CrossRef]
- Wang, Y.; Cao, S.; Guan, C.; Kong, X.; Wang, Y.; Cui, Y.; Liu, B.; Zhou, Y.; Zhang, Y. Overexpressing the NAC transcription factor LpNAC13 from Lilium pumilum in tobacco negatively regulates the drought response and positively regulates the salt response. Plant Physiol Biochem 2020, 149, 96-110. [CrossRef]
- Zhao, X.Y.; Liu, M.S.; Li, J.R.; Guan, C.M.; Zhang, X.S. The wheat TaGI1, involved in photoperiodic flowering, encodes an Arabidopsis GI ortholog. Plant Mol. Biol. 2005, 58, 53-64. [CrossRef]
- Sanagi, M.; Aoyama, S.; Kubo, A.; Lu, Y.; Sato, Y.; Ito, S.; Abe, M.; Mitsuda, N.; Ohme-Takagi, M.; Kiba, T. et al. Low nitrogen conditions accelerate flowering by modulating the phosphorylation state of FLOWERING BHLH4 in Arabidopsis. Proc Natl Acad Sci U S A 2021, 118. [CrossRef]
- Fukazawa, J.; Ohashi, Y.; Takahashi, R.; Nakai, K.; Takahashi, Y. DELLA degradation by gibberellin promotes flowering via GAF1-TPR-dependent repression of floral repressors in Arabidopsis. Plant Cell 2021, 33, 2258-2272. [CrossRef]
- Tayengwa, R.; Sharma, K.P.; Pierce, C.F.; Werner, B.E.; Neff, M.M. Overexpression of AtAHL20 causes delayed flowering in Arabidopsis via repression of FT expression. BMC Plant Biol. 2020, 20, 559. [CrossRef]
- Chen, Y.; Zhang, L.; Zhang, H.; Chen, L.; Yu, D. ERF1 delays flowering through direct inhibition of FLOWERING LOCUS T expression in Arabidopsis. J. Integr. Plant Biol. 2021, 63, 1712-1723. [CrossRef]
- Wang, Y.; Cui, Y.; Liu, B.; Wang, Y.; Sun, S.; Wang, J.; Tan, M.; Yan, H.; Zhang, Y. Lilium pumilum stress-responsive NAC transcription factor LpNAC17 enhances salt stress tolerance in tobacco. Front. Plant Sci. 2022, 13. [CrossRef]
Figure 1.
Phylogenetic analysis of WRKYs from I. laevigata (in red) and Arabidopsis (in black).
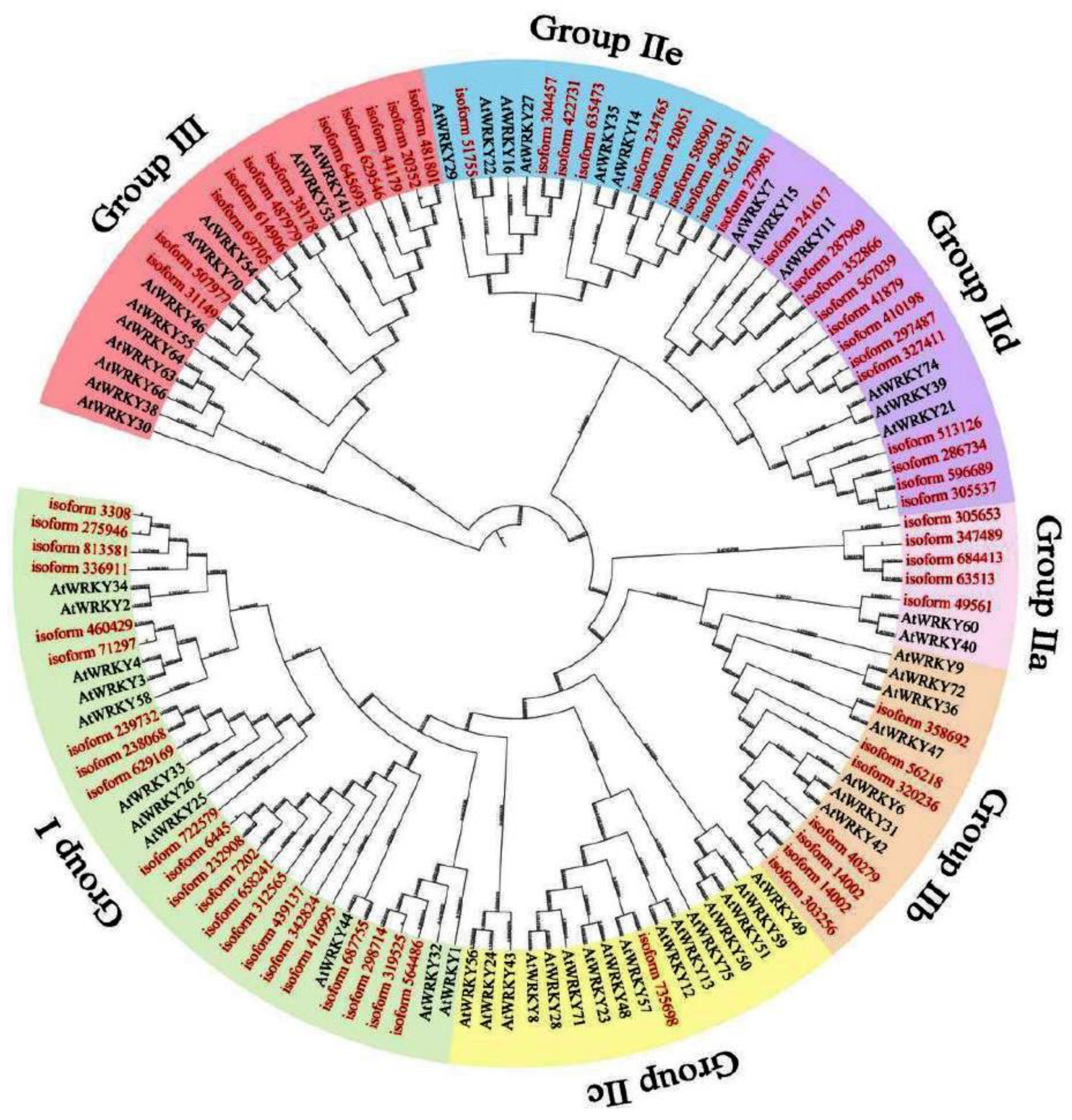
Figure 2.
Sequence phylogenetic analysis and subcellular localization of IlWRKY22. A IlWRKY22 was aligned with its homologs from Zingiber officinale (XP 042466882.1), Zingiber officinale (XP 042374628.1), Carex littledalei (KAF3333198.1), Lolium rigidum (XP 047076847.1), Lolium perenne (XP 0512272831), Dioscorea cavenensis subsp.rotundata (XP 039124149.1), Dendrobium catenatum (XP 020684026.1), Apostasia shenzhenica (PKA59051.1), Gastrodia elata (UUG47650.1), Nymphaea colorata (XP 031504405.1), Nvmphaea thermarum (KAF3792846.1), Cinnamomum micranthum f. kanehirae (RWR80315.1), Cocos nucifera (KAG1367745.1), Elaeis guineensis (XP 010924759.1), Phoenix dactvlifera (XP 0087924072), Gossypium raimondii (XP 0124900221), Durio zibethinus (XP 022757461.1), Cajanus cajan (XP 020220494), and Sesamum indicum (XP 0110929901). The red box was the highly conserved WRKYGQK domain. B A phylogenetic tree of IlWRKY22. C Subcellular localization of IlWRKY22. From left to right: Green fluorescence of GFP, cells under bright field, and cells under superposition of GFP and bright field. Scale bar was 20 μm.
Figure 2.
Sequence phylogenetic analysis and subcellular localization of IlWRKY22. A IlWRKY22 was aligned with its homologs from Zingiber officinale (XP 042466882.1), Zingiber officinale (XP 042374628.1), Carex littledalei (KAF3333198.1), Lolium rigidum (XP 047076847.1), Lolium perenne (XP 0512272831), Dioscorea cavenensis subsp.rotundata (XP 039124149.1), Dendrobium catenatum (XP 020684026.1), Apostasia shenzhenica (PKA59051.1), Gastrodia elata (UUG47650.1), Nymphaea colorata (XP 031504405.1), Nvmphaea thermarum (KAF3792846.1), Cinnamomum micranthum f. kanehirae (RWR80315.1), Cocos nucifera (KAG1367745.1), Elaeis guineensis (XP 010924759.1), Phoenix dactvlifera (XP 0087924072), Gossypium raimondii (XP 0124900221), Durio zibethinus (XP 022757461.1), Cajanus cajan (XP 020220494), and Sesamum indicum (XP 0110929901). The red box was the highly conserved WRKYGQK domain. B A phylogenetic tree of IlWRKY22. C Subcellular localization of IlWRKY22. From left to right: Green fluorescence of GFP, cells under bright field, and cells under superposition of GFP and bright field. Scale bar was 20 μm.
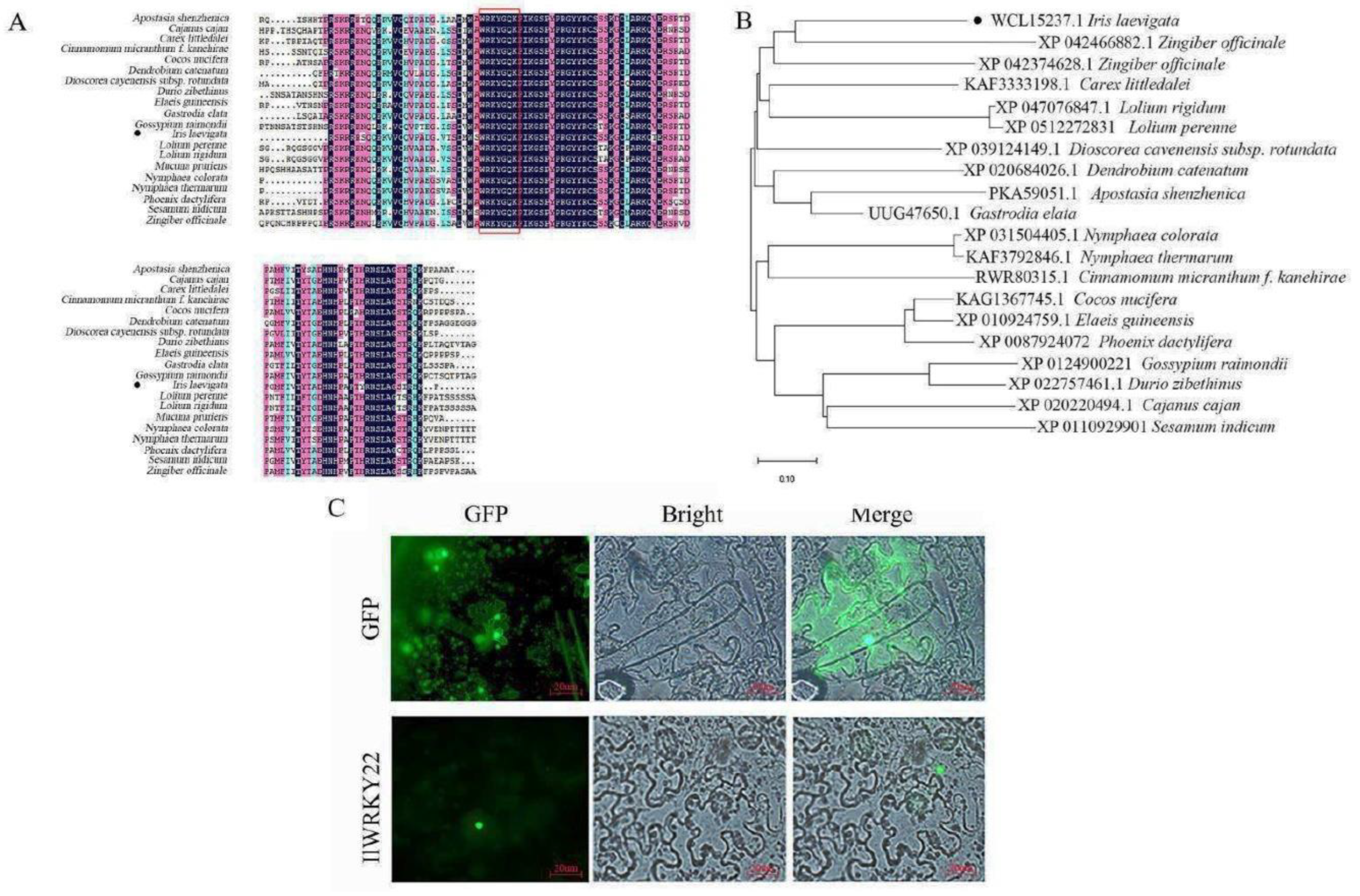
Figure 5.
Overall plant phenotypes under abiotic stress at 0, 7 and 14 days post treatment. A Top view and elevation of the growth state of WT, EV, and OE lines of tobacco at 0, 7, and 14 days in the normal growth state. B Top view and elevation of the growth status of WT, EV, and OE lines of tobacco at 0, 7, and 14 days under natural drought stress. C Top view and elevation of the growth status of WT, EV, and OE lines of tobacco at 0, 7, and 14 days under NaCl stress at 300mM. The scale bar is 20 cm.
Figure 5.
Overall plant phenotypes under abiotic stress at 0, 7 and 14 days post treatment. A Top view and elevation of the growth state of WT, EV, and OE lines of tobacco at 0, 7, and 14 days in the normal growth state. B Top view and elevation of the growth status of WT, EV, and OE lines of tobacco at 0, 7, and 14 days under natural drought stress. C Top view and elevation of the growth status of WT, EV, and OE lines of tobacco at 0, 7, and 14 days under NaCl stress at 300mM. The scale bar is 20 cm.

Figure 6.
Impact of overexpressing IlWRKY22 on photosynthetic and transpiration rate indexes under abiotic stress. Net photosynthetic rate, total chlorophyll content, Fv/Fm, stomatal conductance, intercellular CO2 content, and transpiration rate at 0, 7, and 14 days under 300 mM NaCl (A-F) or drought (G-L) were shown. Different letters above the bars indicated significant differences (p < 0.05).
Figure 6.
Impact of overexpressing IlWRKY22 on photosynthetic and transpiration rate indexes under abiotic stress. Net photosynthetic rate, total chlorophyll content, Fv/Fm, stomatal conductance, intercellular CO2 content, and transpiration rate at 0, 7, and 14 days under 300 mM NaCl (A-F) or drought (G-L) were shown. Different letters above the bars indicated significant differences (p < 0.05).
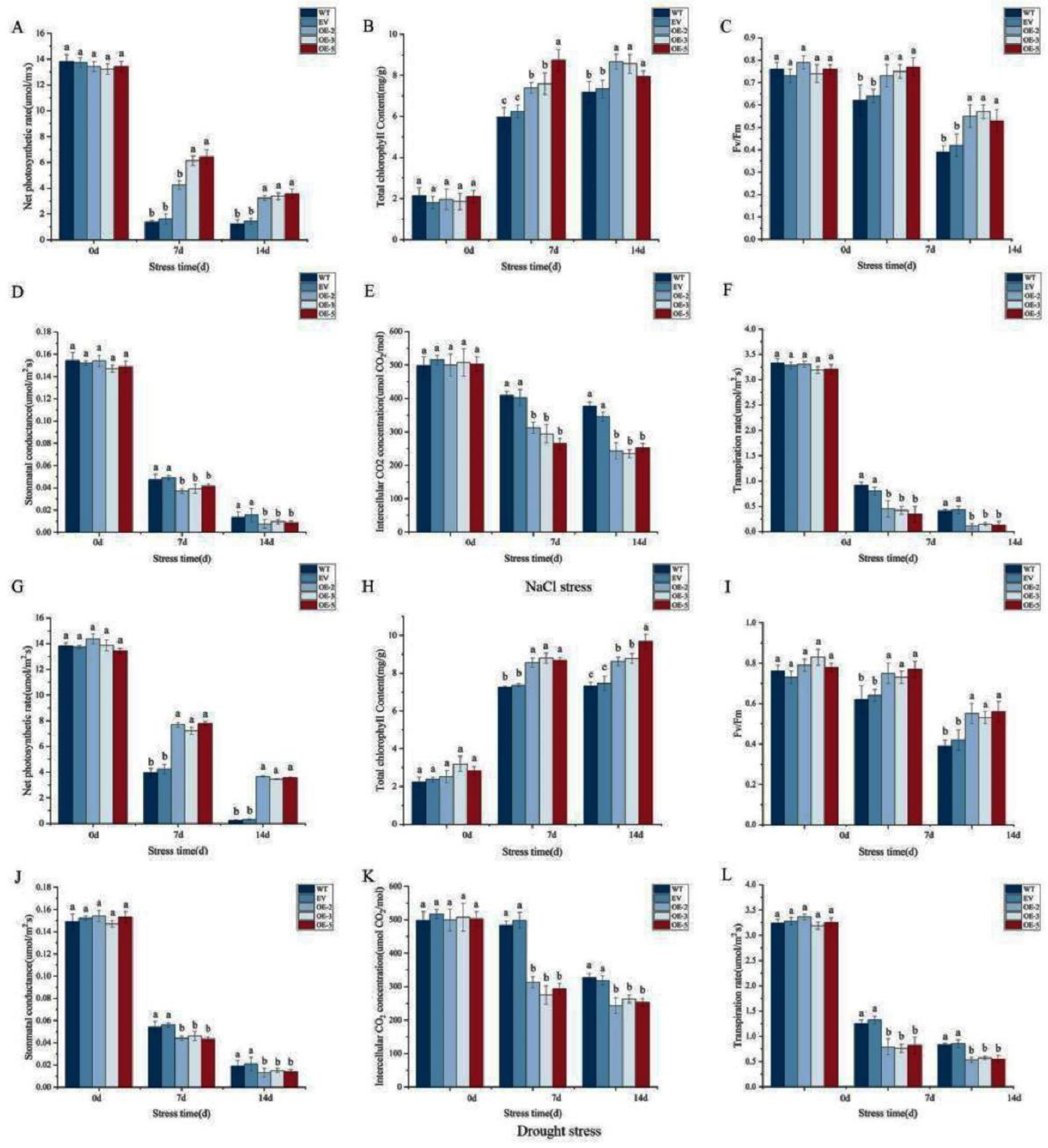
Figure 7.
Involvement of ROS regulation. MDA, O2-, H2O2 and the activities of SOD, CAT and POD, as well as leaf disc staining at 0, 7, and 14 days under 300 mM NaCl (A-F, and M) or drought (G-L, and N) were shown. Different letters above the bars indicated significant differences (p < 0.05).
Figure 7.
Involvement of ROS regulation. MDA, O2-, H2O2 and the activities of SOD, CAT and POD, as well as leaf disc staining at 0, 7, and 14 days under 300 mM NaCl (A-F, and M) or drought (G-L, and N) were shown. Different letters above the bars indicated significant differences (p < 0.05).

Figure 8.
Expression of abiotic stress-responsive genes. The expression levels of NtCAT, NtHAK1, NtPMA4, NtPOD, NtSOD, and NtSOS1 at 0, 7, and 14 days under 300 mM NaCl (A-F) or drought (G-L) were shown. Different letters above the bars indicated significant differences (p < 0.05).
Figure 8.
Expression of abiotic stress-responsive genes. The expression levels of NtCAT, NtHAK1, NtPMA4, NtPOD, NtSOD, and NtSOS1 at 0, 7, and 14 days under 300 mM NaCl (A-F) or drought (G-L) were shown. Different letters above the bars indicated significant differences (p < 0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



