
Submitted:
07 October 2023
Posted:
10 October 2023
You are already at the latest version
Abstract
Extracellular vesicles (EVs), including exosomes, microvesicles, and other lipid vesicles derived from cells, play a pivotal role in intercellular communication by transferring information between cells. EVs secreted by progenitor and stem cells have been associated with the therapeutic effects observed in cell-based therapies, and they also contribute to tissue regeneration following injury, such as in orthopedic surgery cases. This review briefly explores the involvement of EVs in tissue repair and nerve regeneration, their potential as drug carriers, and their significance in stem cell research and cell-based therapies. It underscores the importance for bioengineers to comprehend and manipulate EV activity to optimize the efficacy of tissue engineering and regenerative therapies.
Keywords:
nerve regeneration
; non-coding RNAs
; exosome
; traumatic brain injury
; blood-brain barrier
1. Introduction
Extracellular vesicles (EVs) are defined as heterogeneous vesicles known to be evolutionarily conserved, originating from endosomes or the plasma membrane and released by cells [1, 2]. They play active roles in both normal physiology and pathophysiology, owing to the significance of intercellular communication in prokaryotes and eukaryotic cells, which has driven their evolution [3]. EVs emerged as structures that facilitate this communication [4]. Today, it is known that the structures that provide encoded cellular functions are EVs. EVs were first thought to be procoagulant platelet-derived particles in plasma in 1946. Later, as a result of the study carried out in 1967 by Wolf, these structures began to be called platelet powder [5]. In those years, he thought that these structures carried just cell residues that provide coagulation activity. However, it was later realized that they had much more functional duties than carrying the cell debris. EVs have transmembrane proteins, cytosolic proteins, and small RNA content consisting of the lipid bilayer and are defined as bioactive vesicles that are secreted from cells [6-10]. Generally, six main types of EVs have been defined according to their release mechanism and size. These are, nanovesicles [11], exosomes [12], microvesicles [13], apoptotic bodies [14], large oncosomes [15], and giant plasma membrane vesicles [16] (Figure 1.)
Among these EVs, the most studied sub-types are exosomes and microvesicles [17]. Exosomes and microvesicles have different properties according to their cargo contents. Although exosomes and microvesicles are similar in size, their release pathways are completely different. Exosomes are produced by plasma membrane fusion of multivesicular endosomes (MVEs) followed by the release of intraluminal vesicles (ILVs), while mcrovesicles are secreted by outward vesiculation of the plasma membrane [18]. While various pathways exist for EV generation, the exact mechanism is still largely unknown. Cells can communicate with neighboring cells or distant cells by the secretion of EVs and sub-types. All EVs have surface molecules that allow them to be targeted to recipient cells [19]. After the EV binds to the target cell via receptor-ligand interaction, intercellular signaling is induced or it can enter the cell by endocytosis or phagocytosis. Thus, it triggers many changes to the physiological state of the target cell [20].
1.1. Exosome Biogenesis.
Exosomes are bioactive vesicles secreted by cells, ranging in size from 40 to 200 nm, and taking an active role in intercellular cargo and communication, constituting the important subgroup of EVs. Exosomes, formed due to the differentiation of the endosome, originate from the cell membrane [17, 18]. Their nanoscale components play a sophisticated role in mRNA regulation and are associated with many physiological and pathological functions [19]. Subsequent studies have shown that exosomes have functions as a mediator of cell-to-cell communication. They can easily interact with neighboring cells to facilitate the transfer of active molecules [20]. While exosomes are made up of specially ordered proteins, lipids, and nucleic acids, depending on its cell-type. In addition to approximately 4400 proteins, 194 lipids, 1639 mRNAs, and 764 miRNAs have been in the exosome content. The specific components from different cell types can also be identified. These different components reveal both the complex structure and potential functional diversity of exosomes [21, 22].
The biogenesis of exosomes is quite complicated, and begins with the activation of cell-specific receptors, then, continues the organized regulation of signaling pathways to initiate the cellular process [23]. There are currently two kinds of pathways that play a vital role in exosome biogenesis. The first one is dependent pathways “endosomal sequence complex transport (ESCRT)” and the second one is independent pathways. These pathways cannot be independent from each other [24, 25]. These pathways may work either synergistically or different exosome subpopulations may be dependent on different mechanisms [26]. Exosome biogenesis mainly; originates from the endosomal system. This process begins with the formation of the early endosome with inward budding of the cell membrane followed by a second inward budding of the endosome membrane. With the second inward budding, intraluminal vesicles (ILVs) occur in the late endosome. Late endosomes containing ILV are also described as circumscribed multivesicular bodies (MVBs) [27, 28]. After this step, MVBs either fuse with the lysosome for degradation or follow the endocytic pathway for exosome formation [29]. During the endocytic process, plasma membrane surface proteins are embedded in the inner membrane of endocytic vesicles [30]. Endocytic vesicles with plasma membrane surface proteins line the endoplasmic reticulum membrane and are then processed in the ILV in the Golgi complex. As the final step of the process, ILVs are taken into the extracellular space via the exocytotic pathway, where MVBs fuse with the plasma membrane and are released as exosomes.
1.1. Microvesicles and Their Biogenesis.
Microvesicles, which belong to the class of extracellular vesicles (EVs), are larger in size compared to exosomes and represent a less studied subgroup of EVs. They typically range in size from 100 to 1000 nm [31, 32].
The biogenesis of microvesicles differs significantly from that of exosomes. While exosomes are formed through inward budding of the endosome, microvesicles can be released by outward budding from the plasma membrane [13].
Regarding the biogenesis of microvesicles, cytoskeletal reorganization and associated molecules play a crucial role. One such molecule is Ca2+. A substantial increase in Ca2+ levels trigger the activation of calpain, which breaks down cytoskeletal proteins. This, along with the modulation of flippase, floppase, and scramblase, allows for the remodelling of membrane asymmetry, facilitating the outward budding of microvesicles [33, 34]. Actin, in coordination with myosin, induces a myosin-based contraction of the cytoskeleton following the cytoskeletal changes [35]. Through a mechanism involving ATP-dependent contraction, the coordinated action of actin and myosin leads to the separation of microvesicles from the plasma membrane [36, 37]. Although the biogenesis of microvesicles involves more intricate processes, information on the intrinsic signals driving their formation remains limited. Microvesicles, also known as ectosomes, carry a diverse cargo, including cytosolic proteins, nucleic acids, metabolites, and plasma membrane proteins [38].
2. Stem Cell-Based EVs
Since the pioneering discovery of stem cells by Till and McCulloch in 1961, their application in both scientific and clinical contexts has gained significant momentum in recent years, attracting growing interest [39, 40]. Stem cells are unspecialized cells that possess the remarkable ability to self-renew through division and differentiate into various cell types. They are crucial for basic and clinical research as they can proliferate to yield enough cells and differentiate into desired cell types based on their genetic potential. Moreover, they can integrate into the recipient's tissue after transplantation [41].
According to the stem cell hypothesis, the microenvironment plays a pivotal role in maintaining and directing stem cells. The preservation, differentiation, and repair of stem cells are intricately linked to the presence of this microenvironment, known as the stem cell niche [42]. The stem cell niche consists of stem cells, supportive cells, and scaffolds, collectively contributing to the regulation of stem cell population and processes of determination, differentiation, and repair [43].
Advancements in stem cell technology have opened exciting prospects in regenerative and translational medicine. Stem cells can be broadly categorized into two classes: embryonic stem cells (ESCs) and mesenchymal stem cells (MSCs) [42, 44]. ESCs, being pluripotent, have the potential to differentiate into cells from any of the three embryonic germ layers [45]. However, the use of embryonic stem cells is associated with certain challenges, including ethical concerns and the risk of teratoma formation. As an alternative, the utilization of programmable induced pluripotent stem cells shows great promise [46]. In contrast to embryonic stem cells, mesenchymal stem cells (MSCs) have more advantages despite their limited differentiation capacity [42, 46, 47]. There are several reasons why the use of MSCs is advantageous compared to other treatments:
Availability: MSCs can be obtained from various sources, such as bone marrow, adipose tissue, and umbilical cord blood, making them more readily accessible compared to other stem cell types [48].
Reduced immunogenicity: MSCs exhibit low immunogenicity, meaning they are less likely to elicit an immune response when transplanted into a recipient. This characteristic makes MSCs a suitable choice for allogeneic transplantation, where cells from a donor are used for therapy [49].
Immunomodulatory properties: MSCs possess immunomodulatory capabilities, allowing them to regulate immune responses and reduce inflammation. This feature is particularly beneficial in conditions where excessive immune responses contribute to tissue damage [50].
Tissue regeneration and repair: MSCs can promote tissue regeneration and repair through various mechanisms, including the secretion of growth factors, cytokines, and extracellular vesicles. These factors can stimulate endogenous repair processes and modulate the surrounding microenvironment [51].
Safety profile: MSCs have shown a favorable safety profile in preclinical and clinical studies, with a low incidence of adverse effects. This makes them a promising therapeutic option for a wide range of diseases and conditions [18].
Non-tumorigenic nature: Unlike embryonic stem cells, MSCs have a lower risk of forming teratomas or other tumors. This characteristic enhances their safety profile and reduces concerns associated with tumorigenicity [52].
Overall, the advantages of MSCs, including their availability, immunomodulatory properties, regenerative potential, safety profile, and reduced risk of tumorigenicity, make them a favorable choice for various therapeutic applications [47-49].
I. MSCs offer easy accessibility and can be isolated from various tissues.
II. MSCs possess multipotent capabilities, allowing them to differentiate into multiple cell types.
III. MSCs can be efficiently prepared according to clinical practice-specific standards within a reasonable timeframe.
IV. MSCs can be stored for future therapeutic use, enabling repeated treatments.
V. Allogeneic MSCs have been used in clinical trials without significant adverse reactions. VI. The safety and efficacy of MSCs have been extensively documented in numerous clinical studies [47].
To overcome the limitations of stem cells and facilitate cell-free therapies, there has been an increasing focus on studying stem cell-derived extracellular vesicles (EVs). It is well-known that stem cells actively employ EVs in cell communication within their microenvironment. Among the various types of stem cells, mesenchymal stem cells (MSCs) have received particular attention due to their wide range of cell sources, including bone marrow (BM-MSCs), adipose tissue (AD-MSCs), and umbilical cord blood (UC-MSCs) [53]. The intercellular interactions mediated by stem cell EVs have been demonstrated to play a crucial role in disease treatment. The bioactive molecules carried by EVs exert their effects on target cells through several mechanisms, including:
Direct stimulation of target cells via surface-bound ligands.
Transfer of activated receptors to recipient cells.
Epigenetic reprogramming of recipient cells through the delivery of functional proteins, lipids, and non-coding RNAs [54].
Exosomes can engage in communication with both nearby and distant cells. Numerous studies have investigated the therapeutic efficacy of stem cell EVs in various disease models, as summarized in Table 1.
2.1. Stem Cell-Based EVs in Peripheral Nerve Regeneration
The therapeutic potential of stem cells and their underlying mechanisms largely depend on the conduction pathways and timing of transplantation, particularly in the context of peripheral nerve injuries, which have been shown to be one of the most effective methods [70]. Numerous studies have demonstrated that systemic administration of stem cells in experimental stroke models reduces post-stroke brain damage, enhances neurological recovery, and activates neurodegenerative processes [71]. Study by Doeppner et al, 2015 showed that the therapeutic efficacy of extracellular vesicles (EVs) in peripheral nerve damage has been found to be comparable to that of stem cells when applied systemically [72].
Research investigating the effects of mesenchymal stem cell-derived exosomes has shown a reduction in inflammation and a contribution to neuronal regeneration in rats with spinal cord injury [73-75]. Ma et al, 2019 reported that umbilical cord MSC-derived EVs effectively promote functional recovery and nerve regeneration in a rat model of sciatic nerve injury, suggesting a promising clinical approach for peripheral nerve repair [7]. Indeed, researchers have suggested that the clinical application of MSC-derived EVs may be more appropriate than the application of stem cells.
Similar studies obviously pull out the role of stem cell derived EVs which can facilitate the creation of a suitable micro-environment for nerve regeneration by activating the secretion of neurotrophic and nerve growth factors, thereby this promotes the regeneration of the damaged neuron [76-78]. Today, clinical studies of stem cell derived EVs are expanding and their functions in neural regeneration is considered not only a promising tool, but also less expensive therapy [79].
2.1. Stem Cell-Based EVs in Central Nervous System Regeneration
Studies investigating the therapeutic efficacy of stem cell-derived extracellular vesicles (EVs) have focused on their potential for central nervous system (CNS) regeneration. While the field is still relatively new, these studies have provided valuable insights, although the complete mechanism of regeneration remains to be fully elucidated.
Traumatic brain injury (TBI) is a common event that can have wide-ranging effects on various systems. Exosomes derived from human adipose MSCs (AdMSC-EXs) have demonstrated significant benefits in mouse models of TBI. These exosomes have been shown to increase neuron count, reduce inflammation, and improve sensory and cognitive function more effectively than AdMSCs alone [80]. Besides, there are some recent studies about CNS axon regeneration ophthalmologic animal models by using exosomes [81-84].
Another study investigating MSC-derived exosomes has focused on Alzheimer's disease. It has been suggested that exosomes derived from adipose stem cells express high levels of neprilysin (NEP), an enzyme involved in amyloid β (Aβ) degradation. This finding indicates a potential therapeutic role for these exosomes in reducing Aβ levels in the brain and positively influencing the disease course [75, 85]
In a study by Mendes-Pinheiro et al., the injection of MSCs secretome in a rat model of Parkinson's disease resulted in improvements in animal behavior compared to the 6-OHDA group. The secretome was found to contain neurotrophic and neuroprotective molecules, such as vascular endothelial growth factor (VEGF), brain-derived neurotrophic factor (BDNF), and glial cell line-derived neurotrophic factor (GDNF), which showed beneficial effects on dopaminergic neurons [86].
In another study by Chen et al. 2019, It was reported by in vitro experiments of SH-SY5Y cell line, that the proliferation of human umbilical cord exosomes could be stimulated with 6-hydroxydopamine (6-OHDA). This contributed to the proliferation of rats with Parkinson's model in the substantia nigra by decreasing dopaminergic neuron loss and apoptosis, and the rise in dopamine levels in the striatum [87].
In a study reported in 2012, researchers reported that adipose tissue MSCs (AT-MSCs) and MSCs from Wharton's jelly (WJ-MSCs) can secrete neuro-regulatory/trophic factors that can increase the metabolic viability of hippocampal neurons in vitro [88]. In near future, EVs will have vital role by crossing the blood-brain barrier by the aid of suitable subtypes. Especially, they will be able to induce neurological recovery through address delivery of cargo proteins and non-coding RNAs in EV-subtypes which can serve treatment of central nervous system diseases [89].
3. Conclusions
There is currently a growing interest in clinical trials investigating the therapeutic effect of MSCs in a wide variety of target systems and their potential applications. While the ethical concerns and risk of teratoma associated with stem cells still impact clinical studies, the use of stem cell derived EVs shows promise in overcoming these limitations. EVs possess similar therapeutic capabilities as stem cells but without many of the drawbacks associated with cell-based therapies. These unique properties position EVs as a valuable therapeutic approach. In the future, EVs may not only be utilized for therapeutic purposes but also for diagnostic applications as well. With their ability to regulate gene expression, facilitate tissue regeneration, and exert anti-inflammatory effects, EVs have the potential to become highly effective therapeutic agents. Moreover, by modifying EVs, their therapeutic efficacy can be further enhanced through targeted delivery using nanocarrier systems. Stem cell derived EVs have the advantage of spreading therapeutic properties both locally and systemically, making them a safer alternative to cellular therapy and transplantation surgery. As research in this field deepens in the coming years, stem cell EVs are expected to offer significant advancements in the field of regenerative medicine.
However, several challenges need to be addressed for the successful use of stem cell-derived extracellular vesicles (SC-EVs). These challenges include:
Standardization: It is crucial to establish standardized protocols for the isolation and characterization of EVs to ensure consistent quality and reproducibility of therapeutic preparations.
Storage Optimization: Optimal storage conditions for SC-EVs must be determined to maintain their stability and therapeutic efficacy during storage and transportation.
Sterilization: Ensuring the sterility of EVs without compromising their content and structural integrity is important for safe clinical applications.
Scalable Production: Developing large-scale production techniques is necessary to meet the increasing demand for SC-EVs in clinical settings.
Once these challenges are overcome, future research can focus on the development of bioengineered MSCs that have enhanced properties for targeted delivery of various therapeutic molecules and increased yield of EVs. We anticipate that there will be significant advancements in this field with numerous new studies emerging in the coming years.
Author Contributions
B.Y.; writing. E.C.M.; writing and figure preparation; ZA; review and editing. B.B-N.; writing—review and editing, A.S.; supervision, review and editing, funding acquisition. All authors have read and agreed to the published version of the manuscript.” Please turn to the CRediT taxonomy for the term explanation.
Acknowledgments
B.Y. acknowledges funding from TUSEB Project Number:24212, AS would like to thank SRMRC 2-NIHR funding and E.C.M would like to thank the University of Birmingham, College of Engineering and Physical Science for a 6-month fellowship funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Akers, J.C., D. Gonda, R. Kim, B.S. Carter, and C.C. Chen, Biogenesis of extracellular vesicles (EV): exosomes, microvesicles, retrovirus-like vesicles, and apoptotic bodies. Journal of neuro-oncology 2013, 113, 1–11. [CrossRef]
- Yáñez-Mó, M. P. R.-M. Siljander, Z. Andreu, A. Bedina Zavec, F.E. Borràs, E.I. Buzas, K. Buzas, E. Casal, F. Cappello, and J. Carvalho, Biological properties of extracellular vesicles and their physiological functions. Journal of extracellular vesicles 2015, 4, 27066. [Google Scholar]
- Regev-Rudzki, N., D. W. Wilson, T.G. Carvalho, X. Sisquella, B.M. Coleman, M. Rug, D. Bursac, F. Angrisano, M. Gee, and A.F. Hill, Cell-cell communication between malaria-infected red blood cells via exosome-like vesicles. Cell 2013, 153, 1120–1133. [Google Scholar] [CrossRef]
- Waters, C.M. and B.L. Bassler, Quorum sensing: cell-to-cell communication in bacteria. Annu. Rev. Cell Dev. Biol. 2005, 21, 319–346. [Google Scholar] [CrossRef]
- Wolf, P. The nature and significance of platelet products in human plasma. British journal of haematology 1967, 13, 269–288. [Google Scholar] [CrossRef] [PubMed]
- Desrochers, L.M., M. A. Antonyak, and R.A. Cerione, Extracellular vesicles: satellites of information transfer in cancer and stem cell biology. Developmental cell 2016, 37, 301–309. [Google Scholar] [CrossRef]
- Ma, P., Y. Pan, W. Li, C. Sun, J. Liu, T. Xu, and Y. Shu, Extracellular vesicles-mediated noncoding RNAs transfer in cancer. Journal of hematology & oncology 2017, 10, 1–11. [Google Scholar]
- Kim, S.-M. and H.-S. Kim, Engineering of extracellular vesicles as drug delivery vehicles. Stem cell investigation 2017, 4. [Google Scholar]
- AYGAN, B., M. KAYA, E. CANSEVER MUTLU, and İ. KÜÇÜK, THE ROLE OF EXOSOMES IN DISEASES AND THEIR USE FOR DIAGNOSIS AND THERAPEUTIC PURPOSE. Beykent Üniversitesi Fen ve Mühendislik Bilimleri Dergisi 2021, 14.
- Thomaidou, A.C. M. Goulielmaki, A. Tsintarakis, P. Zoumpourlis, M. Toya, I. Christodoulou, and V. Zoumpourlis, miRNA-Guided Regulation of Mesenchymal Stem Cells Derived from the Umbilical Cord: Paving the Way for Stem-Cell Based Regeneration and Therapy. International Journal of Molecular Sciences 2023, 24, 9189. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q., X. Zhuang, J. Mu, Z.-B. Deng, H. Jiang, L. Zhang, X. Xiang, B. Wang, J. Yan, and D. Miller, Corrigendum: Delivery of therapeutic agents by nanoparticles made of grapefruit-derived lipids. Nature Communications 2016, 7.
- Harding, C.V., J.E. Heuser, and P.D. Stahl, Exosomes: looking back three decades and into the future. 2013.
- D'Souza-Schorey, C. and J.W. Clancy, Tumor-derived microvesicles: shedding light on novel microenvironment modulators and prospective cancer biomarkers. Genes & development 2012, 26, 1287–1299. [Google Scholar]
- Battistelli, M. and E. Falcieri, Apoptotic bodies: particular extracellular vesicles involved in intercellular communication. Biology 2020, 9, 21. [Google Scholar] [CrossRef]
- Di Vizio, D., J. Kim, M.H. Hager, M. Morello, W. Yang, C.J. Lafargue, L.D. True, M.A. Rubin, R.M. Adam, and R. Beroukhim, Oncosome formation in prostate cancer: association with a region of frequent chromosomal deletion in metastatic disease. Cancer research 2009, 69, 5601–5609. [CrossRef]
- Levental, K.R. and I. Levental, Giant plasma membrane vesicles: models for understanding membrane organization. Current topics in membranes 2015, 75, 25–57. [Google Scholar]
- Ingato, D., J.U. Lee, S.J. Sim, and Y.J. Kwon, Good things come in small packages: Overcoming challenges to harness extracellular vesicles for therapeutic delivery. Journal of Controlled Release 2016, 241, 174–185. [CrossRef]
- Mutlu, E.C., Ö. Kaya, M. Wood, I. Mager, K.Ç. Topkara, Ç. Çamsarı, A. Birinci Yildirim, A. Çetinkaya, D. Acarel, and J. Odabaşı Bağcı, Efficient doxorubicin loading to isolated dexosomes of immature JAWSII cells: Formulated and characterized as the bionanomaterial. Materials 2020, 13, 3344.
- Raposo, G. and W. Stoorvogel, Extracellular vesicles: exosomes, microvesicles, and friends. Journal of Cell Biology 2013, 200, 373–383. [Google Scholar] [CrossRef]
- Tkach, M. and C. Théry, Communication by extracellular vesicles: where we are and where we need to go. Cell 2016, 164, 1226–1232. [Google Scholar] [CrossRef]
- Mathivanan, S., C.J. Fahner, G.E. Reid, and R.J. Simpson, ExoCarta 2012: database of exosomal proteins, RNA and lipids. Nucleic acids research 2012, 40, D1241–D1244. [CrossRef]
- Kim, D.-K., B. Kang, O.Y. Kim, D.-s. Choi, J. Lee, S.R. Kim, G. Go, Y.J. Yoon, J.H. Kim, and S.C. Jang, EVpedia: an integrated database of high-throughput data for systemic analyses of extracellular vesicles. Journal of extracellular vesicles 2013, 2, 20384. [CrossRef]
- Keller, S., M.P. Sanderson, A. Stoeck, and P. Altevogt, Exosomes: from biogenesis and secretion to biological function. Immunology letters 2006, 107, 102–108. [CrossRef] [PubMed]
- Stuffers, S., C. Sem Wegner, H. Stenmark, and A. Brech, Multivesicular endosome biogenesis in the absence of ESCRTs. Traffic 2009, 10, 925–937. [CrossRef] [PubMed]
- Henne, W.M., N.J. Buchkovich, and S.D. Emr, The ESCRT pathway. Developmental cell 2011, 21, 77–91.
- Maas, S.L., X.O. Breakefield, and A.M. Weaver, Extracellular vesicles: unique intercellular delivery vehicles. Trends in cell biology 2017, 27, 172–188. [CrossRef] [PubMed]
- Ailawadi, S., X. Wang, H. Gu, and G.-C. Fan, Pathologic function and therapeutic potential of exosomes in cardiovascular disease. Biochimica et Biophysica Acta (BBA) - Molecular Basis of Disease 2015, 1852, 1–11. [CrossRef] [PubMed]
- Barile, L., T. Moccetti, E. Marbán, and G. Vassalli, Roles of exosomes in cardioprotection. European heart journal 2017, 38, 1372–1379.
- Schorey, J.S., Y. Cheng, P.P. Singh, and V.L. Smith, Exosomes and other extracellular vesicles in host–pathogen interactions. EMBO reports 2015, 16, 24–43. [CrossRef]
- Villarroya-Beltri, C., F. Baixauli, C. Gutiérrez-Vázquez, F. Sánchez-Madrid, and M. Mittelbrunn. Sorting it out: regulation of exosome loading. in Seminars in cancer biology. 2014. Elsevier.
- György, B., T.G. Szabó, M. Pásztói, Z. Pál, P. Misják, B. Aradi, V. László, E. Pállinger, E. Pap, and A. Kittel, Membrane vesicles, current state-of-the-art: emerging role of extracellular vesicles. Cellular and molecular life sciences 2011, 68, 2667–2688. [CrossRef]
- Bebawy, M., A. Roseblade, F. Luk, T. Rawling, A. Ung, and G.E. Grau, Cell-derived microparticles: new targets in the therapeutic management of disease. Journal of Pharmacy & Pharmaceutical Sciences 2013, 16, 238–253.
- Piccin, A., W.G. Murphy, and O.P. Smith, Circulating microparticles: pathophysiology and clinical implications. Blood reviews 2007, 21, 157–171. [CrossRef]
- Al-Nedawi, K., B. Meehan, J. Micallef, V. Lhotak, L. May, A. Guha, and J. Rak, Intercellular transfer of the oncogenic receptor EGFRvIII by microvesicles derived from tumour cells. Nature cell biology 2008, 10, 619–624. [CrossRef] [PubMed]
- Sedgwick, A.E., J.W. Clancy, M. Olivia Balmert, and C. D’Souza-Schorey, Extracellular microvesicles and invadopodia mediate non-overlapping modes of tumor cell invasion. Scientific reports 2015, 5, 1–14.
- Blanton, H., J. Jaccard, J. Klick, B. Mellers, G. Mitchell, and P.E. Tetlock, Transparency should trump trust: Rejoinder to McConnell and Leibold (2009) and Ziegert and Hanges (2009). 2009.
- McConnell, R.E., J.N. Higginbotham, D.A. Shifrin Jr, D.L. Tabb, R.J. Coffey, and M.J. Tyska, The enterocyte microvillus is a vesicle-generating organelle. Journal of Cell Biology 2009, 185, 1285–1298.
- Cocucci, E. and J. Meldolesi, Ectosomes and exosomes: shedding the confusion between extracellular vesicles. Trends in cell biology 2015, 25, 364–372. [Google Scholar] [CrossRef]
- Till, J. and E. McCulloch, A direct measurement of the radiation sensitivity of normal mouse bone marrow cells. Radiation research 2011, 175, 145–149. [Google Scholar] [CrossRef]
- Till, J. and E. McCulloch, A direct measurement of the radiation sensitivity of normal bone marrow cells. Radiat Res 1961, 14, 1419–1430. [Google Scholar] [CrossRef]
- Allahverdiyev, A., Somatik ve Kök Hücre Kültür Sistemlerinin Temel İlkeleri. İstanbul: Nobel Tıp Kitabevi 2018, 1.
- Schofield, R. The relationship between the spleen colony-forming cell and the haemopoietic stem cell. Blood cells 1978, 4, 7–25. [Google Scholar] [PubMed]
- Can, A., Kök Hücre: biyolojisi, türleri ve tedavide kullanımları. Akademisyen Tıp Kitabevleri, Ankara 2014.
- Nawaz, M., F. Fatima, K.C. Vallabhaneni, P. Penfornis, H. Valadi, K. Ekström, S. Kholia, J.D. Whitt, J.D. Fernandes, and R. Pochampally, Extracellular vesicles: evolving factors in stem cell biology. Stem cells international 2016, 2016.
- Takahashi, K., K. Tanabe, M. Ohnuki, M. Narita, T. Ichisaka, K. Tomoda, and S. Yamanaka, Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell 2007, 131, 861–872. [CrossRef]
- Alvarez, C.V., M. Garcia-Lavandeira, M. Garcia-Rendueles, E. Diaz-Rodriguez, A.R. Garcia-Rendueles, S. Perez-Romero, T.V. Vila, J.S. Rodrigues, P.V. Lear, and S.B. Bravo, Defining stem cell types: understanding the therapeutic potential of ESCs, ASCs, and iPS cells. Journal of molecular endocrinology 2012, 49, R89–R111.
- Golchin, A. and T.Z. Farahany, Biological products: cellular therapy and FDA approved products. Stem cell reviews and reports 2019, 15, 166–175. [Google Scholar] [CrossRef]
- Wang, Z.-g., Z.-y. He, S. Liang, Q. Yang, P. Cheng, and A.-m. Chen, Comprehensive proteomic analysis of exosomes derived from human bone marrow, adipose tissue, and umbilical cord mesenchymal stem cells. Stem Cell Research & Therapy 2020, 11, 1–11.
- Chen, W., L. Lv, N. Chen, and E. Cui, Immunogenicity of mesenchymal stromal/stem cells. Scandinavian Journal of Immunology 2023, 97, e13267. [CrossRef]
- Yi T, Song SU. Immunomodulatory properties of mesenchymal stem cells and their therapeutic applications. Arch Pharm Res. 2012, 35, 213–221. [CrossRef]
- Tianyi Wu, Yang Liu, Bin Wang and Gang Li, The roles of mesenchymal stem cells in tissue repair and disease modification. Current Stem Cell Research and Therapy 2014, 9, 424–431. [CrossRef]
- Karnas, E., P. Dudek, and E.K. Zuba-Surma, Stem cell-derived extracellular vesicles as new tools in regenerative medicine-Immunomodulatory role and future perspectives. Frontiers in Immunology 2023, 14, 1120175. [CrossRef]
- Squillaro, T., G. Peluso, and U. Galderisi, Clinical trials with mesenchymal stem cells: an update. Cell transplantation 2016, 25, 829–848. [CrossRef]
- Khalyfa, A. and D. Gozal, Exosomal miRNAs as potential biomarkers of cardiovascular risk in children. Journal of translational medicine 2014, 12, 1–12. [Google Scholar] [CrossRef]
- Li, T., Y. Yan, B. Wang, H. Qian, X. Zhang, L. Shen, M. Wang, Y. Zhou, W. Zhu, and W. Li, Exosomes derived from human umbilical cord mesenchymal stem cells alleviate liver fibrosis. Stem cells and development 2013, 22, 845–854. [CrossRef] [PubMed]
- Takahara, K., M. Ii, T. Inamoto, T. Nakagawa, N. Ibuki, Y. Yoshikawa, T. Tsujino, T. Uchimoto, K. Saito, and T. Takai, microRNA-145 mediates the inhibitory effect of adipose tissue-derived stromal cells on prostate cancer. Stem cells and development 2016, 25, 1290–1298. [CrossRef]
- Bai, L., H. Shao, H. Wang, Z. Zhang, C. Su, L. Dong, B. Yu, X. Chen, X. Li, and X. Zhang, Effects of mesenchymal stem cell-derived exosomes on experimental autoimmune uveitis. Scientific reports 2017, 7, 4323. [CrossRef]
- Potter, D.R., B.Y. Miyazawa, S.L. Gibb, X. Deng, P.P. Togaratti, R.H. Croze, A.K. Srivastava, A. Trivedi, M. Matthay, and J.B. Holcomb, Mesenchymal stem cell-derived extracellular vesicles attenuate pulmonary vascular permeability and lung injury induced by hemorrhagic shock and trauma. The journal of trauma and acute care surgery 2018, 84, 245.
- Zhang, Y., Y. Pan, Y. Liu, X. Li, L. Tang, M. Duan, J. Li, and G. Zhang, Exosomes derived from human umbilical cord blood mesenchymal stem cells stimulate regenerative wound healing via transforming growth factor-β receptor inhibition. Stem cell research & therapy 2021, 12, 1–14.
- Yuan, X., D. Li, X. Chen, C. Han, L. Xu, T. Huang, Z. Dong, and M. Zhang, Extracellular vesicles from human-induced pluripotent stem cell-derived mesenchymal stromal cells (hiPSC-MSCs) protect against renal ischemia/reperfusion injury via delivering specificity protein (SP1) and transcriptional activating of sphingosine kinase 1 and inhibiting necroptosis. Cell death & disease 2017, 8, 3200.
- Haga, H., I.K. Yan, K. Takahashi, A. Matsuda, and T. Patel, Extracellular vesicles from bone marrow-derived mesenchymal stem cells improve survival from lethal hepatic failure in mice. Stem cells translational medicine 2017, 6, 1262–1272. [CrossRef] [PubMed]
- Lee, M., T. Liu, W. Im, and M. Kim, Exosomes from adipose-derived stem cells ameliorate phenotype of Huntington's disease in vitro model. European Journal of Neuroscience 2016, 44, 2114–2119. [CrossRef] [PubMed]
- Bian, S., L. Zhang, L. Duan, X. Wang, Y. Min, and H. Yu, Extracellular vesicles derived from human bone marrow mesenchymal stem cells promote angiogenesis in a rat myocardial infarction model. Journal of molecular medicine 2014, 92, 387–397. [CrossRef]
- Xu, K., D. Ma, G. Zhang, J. Gao, Y. Su, S. Liu, Y. Liu, J. Han, M. Tian, and C. Wei, Human umbilical cord mesenchymal stem cell-derived small extracellular vesicles ameliorate collagen-induced arthritis via immunomodulatory T lymphocytes. Molecular Immunology 2021, 135, 36–44. [CrossRef] [PubMed]
- Keshtkar, S., M. Kaviani, F.S. Sarvestani, M.H. Ghahremani, M.H. Aghdaei, I.H. Al-Abdullah, and N. Azarpira, Exosomes derived from human mesenchymal stem cells preserve mouse islet survival and insulin secretion function. EXCLI journal 2020, 19, 1064.
- Yan, Y., W. Jiang, Y. Tan, S. Zou, H. Zhang, F. Mao, A. Gong, H. Qian, and W. Xu, hucMSC exosome-derived GPX1 is required for the recovery of hepatic oxidant injury. Molecular Therapy 2017, 25, 465–479. [CrossRef]
- Sheykhhasan, M., N. Kalhor, A. Sheikholeslami, M. Dolati, E. Amini, and H. Fazaeli, Exosomes of mesenchymal stem cells as a proper vehicle for transfecting miR-145 into the breast cancer cell line and its effect on metastasis. BioMed Research International 2021, 2021.
- Guo, Q., J. Yan, T. Song, C. Zhong, J. Kuang, Y. Mo, J. Tan, D. Li, Z. Sui, and K. Cai, microRNA-130b-3p contained in MSC-derived EVs promotes lung cancer progression by regulating the FOXO3/NFE2L2/TXNRD1 axis. Molecular Therapy-Oncolytics 2021, 20, 132–146. [CrossRef] [PubMed]
- Sharma, A., R. Kulkarni, H. Sane, N. Awad, A. Bopardikar, A. Joshi, S. Baweja, M. Joshi, C. Vishwanathan, and N. Gokulchandran, Phase 1 clinical trial for intravenous administration of mesenchymal stem cells derived from umbilical cord and placenta in patients with moderate COVID-19 virus pneumonia: results of stage 1 of the study. American Journal of Stem Cells 2022, 11, 37.
- Doeppner, T.R., B. Kaltwasser, M. Teli, E. Bretschneider, M. Bähr, and D.M. Hermann, Effects of acute versus post-acute systemic delivery of neural progenitor cells on neurological recovery and brain remodeling after focal cerebral ischemia in mice. Cell death & disease 2014, 5, e1386–e1386.
- Han, Y., D. Seyfried, Y. Meng, D. Yang, L. Schultz, M. Chopp, and D. Seyfried, Multipotent mesenchymal stromal cell–derived exosomes improve functional recovery after experimental intracerebral hemorrhage in the rat. Journal of neurosurgery 2018, 131, 290–300.
- Doeppner, T.R., J. Herz, A. Görgens, J. Schlechter, A.-K. Ludwig, S. Radtke, K. de Miroschedji, P.A. Horn, B. Giebel, and D.M. Hermann, Extracellular vesicles improve post-stroke neuroregeneration and prevent postischemic immunosuppression. Stem cells translational medicine 2015, 4, 1131–1143. [CrossRef]
- Han, D., C. Wu, Q. Xiong, L. Zhou, and Y. Tian, Anti-inflammatory mechanism of bone marrow mesenchymal stem cell transplantation in rat model of spinal cord injury. Cell biochemistry and biophysics 2015, 71, 1341–1347. [CrossRef]
- de Rivero Vaccari, J.P., F. Brand III, S. Adamczak, S.W. Lee, J. Perez-Barcena, M.Y. Wang, M.R. Bullock, W.D. Dietrich, and R.W. Keane, Exosome-mediated inflammasome signaling after central nervous system injury. Journal of neurochemistry 2016, 136, 39–48.
- Vilaça-Faria, H., A.J. Salgado, and F.G. Teixeira, Mesenchymal stem cells-derived exosomes: a new possible therapeutic strategy for Parkinson’s disease? Cells 2019, 8, 118. [CrossRef]
- Fan, C.-G., Q. -J. Zhang, F.-W. Tang, Z.-B. Han, G.-S. Wang, and Z.-C. Han, Human umbilical cord blood cells express neurotrophic factors. Neuroscience letters 2005, 380, 322–325. [Google Scholar] [CrossRef]
- Caplan, A.I. and J.E. Dennis, Mesenchymal stem cells as trophic mediators. Journal of cellular biochemistry 2006, 98, 1076–1084. [Google Scholar] [CrossRef]
- Zhou, Y., H. Xu, W. Xu, B. Wang, H. Wu, Y. Tao, B. Zhang, M. Wang, F. Mao, and Y. Yan, Exosomes released by human umbilical cord mesenchymal stem cells protect against cisplatin-induced renal oxidative stress and apoptosis in vivo and in vitro. Stem cell research & therapy 2013, 4, 1–13. [Google Scholar]
- De Miroschedji, K.N.M. , Human platelet lysate derived extracellular vesicles for use in medicine. 2019, Google Patents.
- Patel, N.A., L. D. Moss, J.-Y. Lee, N. Tajiri, S. Acosta, C. Hudson, S. Parag, D.R. Cooper, C.V. Borlongan, and P.C. Bickford, Long noncoding RNA MALAT1 in exosomes drives regenerative function and modulates inflammation-linked networks following traumatic brain injury. Journal of neuroinflammation 2018, 15, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Yu, B., X. -R. Li, and X.-M. Zhang, Mesenchymal stem cell-derived extracellular vesicles as a new therapeutic strategy for ocular diseases. World journal of stem cells 2020, 12, 178. [Google Scholar] [CrossRef] [PubMed]
- Mead, B., E. Cullather, N. Nakaya, Y. Niu, C. Kole, Z. Ahmed, and S. Tomarev, Viral delivery of multiple miRNAs promotes retinal ganglion cell survival and functional preservation after optic nerve crush injury. Experimental eye research 2020, 197, 108071. [Google Scholar] [CrossRef] [PubMed]
- Mead, B., X. Chamling, D.J. Zack, Z. Ahmed, and S. Tomarev, TNFα-mediated priming of mesenchymal stem cells enhances their neuroprotective effect on retinal ganglion cells. Investigative ophthalmology & visual science 2020, 61, 6–6. [Google Scholar]
- Mead, B., A. Logan, M. Berry, W. Leadbeater, and B.A. Scheven, Paracrine-mediated neuroprotection and neuritogenesis of axotomised retinal ganglion cells by human dental pulp stem cells: comparison with human bone marrow and adipose-derived mesenchymal stem cells. PloS one 2014, 9, e109305. [Google Scholar] [CrossRef] [PubMed]
- Katsuda, T., R. Tsuchiya, N. Kosaka, Y. Yoshioka, K. Takagaki, K. Oki, F. Takeshita, Y. Sakai, M. Kuroda, and T. Ochiya, Human adipose tissue-derived mesenchymal stem cells secrete functional neprilysin-bound exosomes. Scientific reports 2013, 3, 1197. [Google Scholar] [CrossRef]
- Mendes-Pinheiro, B., S. I. Anjo, B. Manadas, J.D. Da Silva, A. Marote, L.A. Behie, F.G. Teixeira, and A.J. Salgado, Bone marrow mesenchymal stem cells' secretome exerts neuroprotective effects in a Parkinson's disease rat model. Frontiers in bioengineering and biotechnology 2019, 7, 294. [Google Scholar] [CrossRef]
- Chen, H.-X., F. -C. Liang, P. Gu, B.-L. Xu, H.-J. Xu, W.-T. Wang, J.-Y. Hou, D.-X. Xie, X.-Q. Chai, and S.-J. An, Exosomes derived from mesenchymal stem cells repair a Parkinson’s disease model by inducing autophagy. Cell death & disease 2020, 11, 288. [Google Scholar]
- Ribeiro, C.A., J. S. Fraga, M. Grãos, N.M. Neves, R.L. Reis, J.M. Gimble, N. Sousa, and A.J. Salgado, The secretome of stem cells isolated from the adipose tissue and Wharton jelly acts differently on central nervous system derived cell populations. Stem cell research & therapy 2012, 3, 1–7. [Google Scholar]
- Li, Y., Y. Tang, and G.-Y. Yang, Therapeutic application of exosomes in ischaemic stroke. Stroke and Vascular Neurology 2021, 6. [Google Scholar]
Figure 1.
Some release studies of nanovesicles [11], schematic representation of exosomes [12] and apoptotic bodies [14].
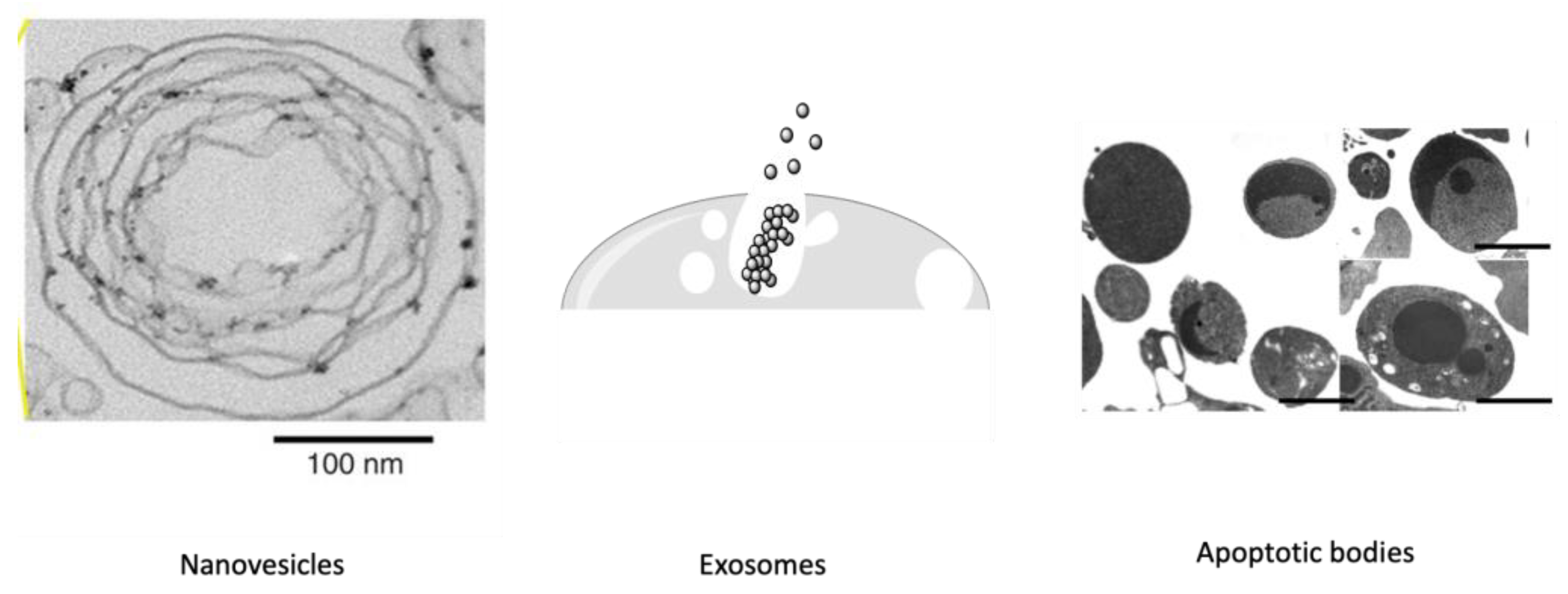

Table 1.
Use of different stem cell-derived EVs in preclinical studies.
| INVESTİGATED DİSEASE | EV SOURCES | Used EVs | THERAPEUTİC MECHANİSM | Ref. |
|---|---|---|---|---|
| Liver Fibrosis | Human Umbilical Cord MSC | Exosome | By inhibiting the epithelial-to-mesenchymal transition (EMT) by increasing E-cadherin-positive cells and decreasing N-cadherin- and vimentin-positive cells | Li et al, 2013[55] |
| Prostate Cancer | Human Adipose-derived stromal cell MSC | Exosome | BclxL activity decreased by miR-145 decreased proliferation and increased apoptosis | Takahara et al, 2016 [56] |
| Autoimmune Uveoretinitis | Human Umbilical Cord MSCs | Exosome | By inhibiting the migration of inflammatory cells | Bai et al, 2017 [57] |
| Acute Lung Injury | Human Bone Marrow-Derived MSCs | Exosomes and Microvesicles | Regulate immunity and reduce Pulmonary capillary permeability by Delivery of Angiopoietin-1 mRNA | Potter et al, 2018 [58] |
| Wound Healing | Human Umbilical Cord MSCSs | Exosome | Reduces scar formation and myofibroblast accumulation by transfer of microRNAs and suppression of TGF-β | Zhang et al, 2021 [59] |
| Kidney Injury | Induced Pluripotent Stem Cell iPSC-MSCs | Exosomes and Microvesicles | By transporting the specificity protein (SP1) to renal tubular epithelial cells, it increases the expression of sphingosine kinase 1 and inhibits necroptosis | Yuan et al, 2017 [60] |
| Osteoporosis | Induced Pluripotent Stem Cell iPSC-MSCs | Exosome | Protection against bone loss by activation of the PI3K/Akt signaling pathway | Haga et al, 2017 [61] |
| Huntington's Disease | Human Adipose-derived stromal cell MSCs | Exosome | Reducing mHtt aggregate level, improving abnormal apoptotic protein level by regulating PGC-1, phospho-CREB | Lee et al, 2016 [62] |
| Myocardial Infarction | Human Bone Marrow-Derived MSCs | Exosomes and Microvesicles | Protecting heart tissue from ischemic damage by promoting neoangiogenesis | Bian et al, 2014 [63] |
| Rheumatoid Arthritis | Human Umbilical Cord MSCs | Exosomes and Microvesicles | Reducing joint inflammation, synovial hyperplasia, and cartilage destruction by decreasing T cell proliferation | Xu et al, 2021 [64] |
| Diabetes-Type 1 | Human Wharton's Jelly-Derived MSCs | Exosome | It increases the insulin content and improves normoglycemia with the increase in VEGF expression | Keshthar et al, 2020 [65] |
| Diabetes-Type 2 | Human Umbilical Cord MSCs | Exosome | It restores phosphorylation (tyrosine domain) of insulin receptor substrate 1 and protein kinase B, promotes expression and membrane translocation of glucose transporter 4 in muscle, and increases glycogen storage in the liver to maintain glucose homeostasis, thereby inhibiting β-cell apoptosis. | Yan et al, 2018 [66] |
| Breast Cancer | Human Adipose-derived stromal cell MSCs | Exosome | Overexpression of miR-145 leads to the downregulation of ROCK1, which inhibits cell proliferation and suppresses metastasis. | Sheykhhasan et al, 2021 [67] |
| Lung Cancer | Human Umbilical Cord MSCs | Exosomes and Microvesicles | miR-130b-3p directly targeted FOXO3, and FOXO3 upregulated Keap1 expression to downregulate NFE2L2, thereby inhibiting TXNRD1. FOXO3 overexpression or silencing of NFE2L2 or TXNRD1 decreased lung cancer cell proliferation, invasion, and migration but increased apoptosis. | Guo et al, 2021 [68] |
| Gastric Cancer | Human Bone Marrow-Derived MSCs | Exosome | Inhibition of gastric cancer cell migration and invasion by miR-221 transfection | Kim et al, 2017 [8] |
| Covid-19 | Human Umbilical Cord MSCs | Exosome | A clinical trial of the treatment of chronic cough is still ongoing | Sharma et al, 2022 [69] |
* Tables may have a footer.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



