
Submitted:
09 October 2023
Posted:
11 October 2023
You are already at the latest version
Abstract
β3-Adrenoceptors mediate several functions in rodents that could be beneficial for the treatment of obesity and type 2 diabetes. This includes promotion of insulin release from the pancreas, of cellular glucose uptake, of lipolysis, and of thermogenesis in brown adipose tissue. In combination, they lead to a reduction of body weight in several rodent models including ob/ob mice and Zucker diabetic fatty rats. These findings stimulated drug development programs in various pharmaceutical companies, and at least nine β3-adrenoceptor agonists have been tested in clinical trials. However, all of these projects were discontinued due to lack of clinically relevant changes of body weight. Following a concise historical account of discoveries leading to such drug development programs we discuss species differences that explain why β3-adrenoceptors are not a meaningful drug target for the treatment of obesity and type 2 diabetes in humans.
Keywords:
β3-adrenoceptor
; obesity
; type 2 diabetes
; species difference
; insulin release
; glucose uptake
; thermogenesis
1. Introduction
Obesity is generally defined as a body mass index exceeding 30 kg/m2. Its prevalence has markedly increased in past decades irrespective of ethnicity, gender, socioeconomic status, or geographical location; affecting one third of the global population, it is now considered to be a pandemic [1]. Obesity increases the risk for multiple conditions, most importantly type 2 diabetes (T2DM). Other conditions occurring more frequently in obese subjects include lower urinary symptoms such as the overactive bladder syndrome [2], which is the only established and approved indication for β3-adrenoceptor (AR) agonists [3].
Behavioral modification is the first line management of obesity but often is insufficiently effective, leading to a need for medical or even surgical treatment. Agonists of glucagon-like peptide 1 receptors such as semaglutide have recently proven to be effective pharmacological treatments of obesity, but not all obese subjects are responsive to this drug class and even in responders their use can be limited by tolerability issues [4]. Therefore, a continuing medical need exists for alternative pharmacological treatment options. This manuscript discusses how the idea of targeting β3-AR for the treatment of obesity and T2DM developed and which evidence supports and argues against the validity of this concept.
In principle, a reduction of body weight and/or improvement of hyperglycemia can be achieved by three main means. A reduction of caloric intake (dieting) is a domain of behavioral modification but the recently introduced glucagon-like peptide 1 receptor agonists also use this mechanism [4,5]. However, there is no evidence linking β3-AR to control of food intake. A second approach is to increase energy expenditure leading to a negative energy balance if caloric intake does not increase to a similar extent. This can be attained by exercise (not covered here) and increased lipolysis and thermogenesis. More recently, an enhanced renal excretion of glucose has emerged as an additional mechanism to increase energy expenditure; this mechanism is used by sodium glucose transporter 2 inhibitors [6]. Various members of the latter drug class including canagliflozin, dapagliflozin, and empagliflozin have been found to decrease body weight in T2DM patients; however, with an average weight loss of < 2 kg, these effects are too limited to be considered as anti-obesity drugs [7]. While limited evidence in rodents points to a role of β3-AR in the regulation of renal function [8], there is no evidence linking them to renal glucose excretion. A third option linked to glycemic control but not necessarily to body weight is modulation of insulin release from pancreatic β cells and/or cellular glucose uptake.
Brown adipose tissue (BAT) activation has been considered a corner stone in improving metabolic health because it is a metabolic sink for glucose and free fatty acids (FAA) and correlates to improved glucose and insulin sensitivity [9,10,11,12]. Therefore, it was suggested to target BAT by β3-AR agonists such as mirabegron as a potential treatment for metabolic dysfunction and obesity [12,13,14].
Following a concise review of historical aspects leading to the initiation of research and development programs in the β3-AR field, this manuscript will summarize limited data on modulation of insulin release and/or cellular glucose uptake and then mainly focus on their effects on lipolysis and thermogenesis. In line with their proposed mechanisms of action in T2DM, β3-AR agonists did not improve hyperglycemia in rodent models of type 1 diabetes (T1DM), e.g., as induced by streptozotocin or alloxan [15]; therefore, data related to T1DM will not be covered here systematically.
2. Historical aspects
Soon after Lands had proposed a subdivision of β-AR into the subtypes β1 and β2 [16], emerging evidence supported that some responses to β-adrenergic agonists such as isoprenaline were not mediated by either of these two subtypes. Notably, this included evidence that an atypical receptor subtype, i.e., different from β1- and β2-AR, may mediate lipolysis in rodents [17,18,19]. Other physiological responses attributed to an atypical β-AR included inhibition of intestinal motility in rats [17,20] and relaxation of human urinary bladder [21]. The latter has meanwhile led to β3-AR agonists being a guideline recommended drug class for the treatment of the overactive bladder syndrome [3].
The initial research and development programs on β3-AR agonists as potential treatments of obesity and T2DM were stimulated by lipolysis and other functional studies [22]. The field gained traction upon the cloning of the human β3-AR [23] and its homologs of rats [24] and mice [25]. Of note, the latter study found that the rank order of potency of agonists to stimulate cAMP formation in transfected CHO cells correlated well with that to induce lipolysis in rat brown adipocytes. Moreover, the mRNA expression of the newly cloned rat receptor was reduced by 71% in obese Zucker rats as compared to lean animals. The interpretation of the findings from rodents was limited by the fact that the human and the rodent β3-AR genes differ in multiple ways. They differ in the presence and location of introns [26,27,28] and in their 5’ flanking regions and regulatory sequences [29] leading to possible differences in their transcriptional control.
2.1. Insight from gene polymorphism association studies
Soon after the cloning of the human β3-AR gene, it was reported that a Trp64Arg polymorphism in the coding region of the human β3-AR gene was associated with an increased capacity for weight gain [30] and an earlier onset of T2DM, particularly in Pima Indians [31]. While similar findings were obtained by some investigators [32,33,34,35], others did not confirm such associations particularly in Caucasian populations [36,37,38]. Others reported an association of the polymorphism with some but not other parameters related to metabolism and obesity [39] or that the polymorphism was associated with responses to some but not other β3-AR agonists [33].
A major review of these and other studies, mostly on cardiovascular phenotypes, concluded that the available evidence was insufficient to establish a link between the Trp64Arg polymorphism and obesity or diabetes but indicated a trend for the Arg64 allele as being a possible risk factor [40]. This conclusion was also supported by lipolysis studies adipocytes from genotyped obese and normal weight subjects upon stimulation with the atypical agonist CGP 12,177 [32] or similar studies involving subjects with multiple ethnicities and isoprenaline as the agonist [33,36]. Such a link would imply that the 64Arg variant of the receptor is hypofunctional. However, dedicated in vitro studies based on site-directed mutagenesis did not consistently find the 64Arg variant to be hypofunctional [41]. Importantly, the β3-AR locus has not shown up as a trait for obesity or diabetes in any of the genome-wide association studies. Evidence for an association of β3-AR gene polymorphisms and lower urinary tract function has also remained inconclusive [41].
2.2. Insight from β3-AR knock-out studies
Independent groups have generated β3-AR knock-out mice that exhibited several metabolism related phenotypes. One set of knock-out mice exhibited a 34% and 131% increase in total body fat in males and females, respectively [42]. The other knock-out line, reporting on male mice only, had a 42% increase in total body fat [43], implying a sex difference in the role of β3-ARs in metabolic control. The latter line also had an attenuated a respiratory rate response to the β3-AR agonist CL 316,243 in white adipose tissues (WAT) and BAT, whereas those to the β1- and β2-AR agonists dobutamine and terbutaline, respectively, were preserved [44]. Surprisingly at the time, the response to CGP 12,177 was attenuated in WAT but not BAT of the knock-out mice. This may be explained by more recent findings that CGP 12,177 is a partial agonist at β3-ARs, an orthosteric antagonist at the β1- and β2-AR, and an agonist at a non-orthosteric site of the β1-AR [45].
2.3. Early research and development programs
A detailed account of the early research and development work on β3-AR agonists as potential treatments of obesity and T2DM has been provided [22]. A starting point had been observations that ephedrine [46,47,48] and other sympathomimetic agents including various β-AR agonists increased thermogenesis and caused weight loss in the genetically obese ob/ob mice [49,50].
While the initial studies suggested that the weight loss response involves a β-AR, the challenge became to find compounds that mimic this effect but do not cause hypertension, tachycardia, tremor, or hypokalemia, side effects known to be mediated by β1- and/or β2-ARs [51]. As some of these effects apparently exhibit non-linear receptor-effector coupling (receptor reserve, also known as spare receptors) [52], a high degree of selectivity for the β3-AR relative to the other subtypes is required to avoid such adverse effects [53].
The apparently first research program on β3-ARs as a drug target for the treatment of obesity and T2DM was launched at Beecham Pharmaceuticals (now a part of GSK). It led to the discovery of atypical β-AR agonists including BRL 28,410, BRL 35,135 and BRL 37,344, which were effective on lipolysis but lacked the above side effects attributed to β1- and/or β2-ARs [54]. The compounds identified at Beecham Pharmaceuticals turned out to be agonists at the cloned β3-AR, although their selectivity for this subtype has meanwhile been challenged, particularly for the human β3-AR [55]. Subsequently, multiple companies filed patents disclosing β3-AR agonists and their potential use in the treatment of obesity and T2DM (see section 8).
Taken together, the knock-out mouse studies supported a role for the β3-AR in the regulation of total body fat in rodents. In contrast, human gene polymorphism studies remained inconclusive, and the β3-AR gene locus did not point to a role of this receptor in obesity and diabetes in genome-wide association studies. With hindsight, these findings challenge the wisdom behind obesity and T2DM drug discovery programs targeting β3-AR.
3. Insulin release and cellular glucose uptake
A physiological function of insulin is stimulation of cellular glucose uptake mostly into adipocytes, e.g., after a meal. While this helps to acutely maintain euglycemia, it can chronically lead to excessive lipid storage, i.e., obesity. Therefore, several studies have addressed the effects of β3-AR agonists on insulin release and circulating insulin levels, and on the interaction of insulin and β3-ARs in the control of cellular glucose uptake.
3.1. Insulin release
An early study reported that BRL 26,830 increased plasma insulin concentrations in fasted rats and improved glucose disposal after a glucose load in non-diabetic rats and mice [15]. Subsequent studies by others reported dose-dependent increases of blood insulin and glucagon concentrations along with a lowering of blood glucose in fasted mice; propranolol at 10-50 mg/kg attenuated these responses whereas metoprolol and ICI 118,551 at 50 mg/kg mimicked the propranolol response only partly [56]. No increase of blood insulin concentrations was observed in mice with streptozotocin-induced T1DM, implying that these effects required intact pancreatic β cells. Within the same report, BRL 26,830 also increased blood insulin and glucagon in dogs; while glucose levels were not affected, those of FFA increased markedly, pointing to a site of action not in the pancreas but rather in adipose tissue (AT) in canines.
In cultured rat pancreatic islet cells, neither BRL 26,830 nor the related compound BRL 28,410 stimulated insulin release in the presence of 2.8 or 5.6 mM glucose. The same group also studied the β3-AR agonist CL 316,243 in in situ perfused mouse pancreas [57]. The agonist concentration-dependently stimulated insulin secretion, which as partially inhibited by propranolol and ICI 118,551, but not by metoprolol; based on an only incomplete inhibition by even high concentrations of propranolol (200 µM, expected to saturate β1- and β2-ARs), the investigators proposed that a major part of this effect was mediated by β3-ARs.
Experiments in the rat insulinoma cell line RIN 1040-38 (a model for pancreatic β cells) found β3-AR expression and increased insulin release in the presence of BRL 37,344 and CL 316,243 [58]. However, this release had two characteristics that complicate interpretation of the data. Firstly, both agonists had bell-shaped concentration-response curves with maximal effects at 1-10 nM but a lack of effect at 100-100 nM. Second, the effect was transient with a peak after 30 min and return to control levels after 60 min. If the cells were transfected with the human wild-type β3-AR, the bell-shaped concentration-response curve remained; upon transfection with the 64Arg variant of the β3-AR, responses to CL 316,243 were not detected, again pointing to this variant being hypofunctional.
CL 316,243 increased pancreatic islet blood flow and plasma insulin concentration in rats while not affecting overall pancreatic blood flow [59]. This was prevented by a high dose of bupranolol (general β-AR antagonist also inhibiting β3-ARs) but not by nadolol (not inhibiting β3-ARs), implying involvement of a β3-AR. Based on these findings, the authors proposed that insulin release by a β3-AR agonist may occur at least partly secondary to vasodilation of microvessels in the islet of Langerhans. Of note, while vasodilation is typically attributed to β2-ARs, it can occur via β3-ARs in some vascular beds [60].
A study with three selective β3-AR agonists for 14 days in db/db mice, including solabegron that has been tested clinically in overactive bladder patients [61], reported dose-dependent reductions of plasma insulin concentrations [62], which may be secondary to overall metabolic improvements as shown by concomitant reductions of glucose and HbA1c.
The involvement of β3-AR in insulin release, food intake, and oxygen consumption induced by CL 316,243 in rat WAT was confirmed by experiments in genetically modified mouse models with either transgenic expression of the β3-AR in AT or in β3-AR knockout mice [63].
Taken together these data indicate that β3-AR agonists can promote insulin release from the pancreas of rats and mice upon acute administration, but the underlying cellular and molecular mechanisms remain unclear and may be indirect, i.e., secondary to vasodilation. Moreover, these acute effects were not observed in dogs and, at least in hyperglycemic mice, turn into the opposite upon chronic administration.
These studies in combination with the expression of β3-AR mRNA and protein in the human pancreas, specifically in the islets of Langerhans [58], have prompted limited investigations in human subjects. Eight healthy subjects received single oral doses of BRL 35,135 (8 mg) or salbutamol (8 mg) after pre-treatment with placebo, bisoprolol (5 mg) or nadolol (20 mg) [64]. Both agonists lowered serum potassium concentrations, a known β2-AR response, and increased serum glucose, insulin, and lactate. All three metabolic responses were blocked by nadolol but not bisoprolol, indicating that they occurred via β2-AR Interestingly, BRL 35,135 but not salbutamol increased serum FFA and glycerol concentrations (similar to findings in dogs [56]), but that also appeared to be a β2-AR effect. A recent study administered a high dose of mirabegron (100 mg q.d.) to 14 healthy women of various ethnicities for a period of 4 weeks [13]. Insulin responses in a glucose tolerance test were similar prior to and after 27 days of treatment. Taken together these limited data do not support a relevant β3-AR-mediated insulin release in humans, which is similar to dogs but contrasts findings in rats and mice.
3.2. Cellular glucose uptake
Despite the controversial data concerning the role of β3-AR in mediating insulin release, several lines of evidence have indicated the possible involvement of β3-AR in enhancing glucose tolerance and uptake. El Hadri et al. attributed this phenomenon to the complex interaction between feeding/fasting status and the expression of β3-AR in AT [65]. Nevertheless, insulin stimulated glucose transport in rat adipocytes with a rank order implying a β3-AR involvement [66,67,68,69]. Several studies supported this claim. For instance, using CL 316,243 in transgenic mouse model with either β3-AR specific expression in AT or with β3-AR knockout mice, suggested that insulin release, food intake and oxygen consumption induced by CL 316,243 is mediated by β3-AR in WAT [63]. Moreover, CL 316,243 seemed to increase glucose uptake in a tissue dependent manner, and variations were observed in different models of diabetes. CL 316,243 was effective in improving glucose uptake in BAT but not in inguinal WAT (iWAT) in T2DM mice. The opposite was observed in T1DM animals [70]. One study included different organs to examine the effect of BRL 37,344 in male Sprague Dawley rats. As expected, BRL 37,344 improved glucose uptake in skeletal muscle, heart and diaphragm, in addition to BAT and WAT [71]. Other than that, a one week treatment with CL 316,243 (1mg/kg/day) induced glucose uptake in Wistar rat WAT but not in guinea pigs, alongside upregulation of GLUT4 mRNA expression in scWAT and BAT of treated rats [72]. These data indicate a variation of β3-AR role and response in glucose uptake across species.
Functional evidence unravelling β3-AR induced glucose uptake in humans is limited, however, it is speculated that β3-AR agonists such as mirabegron improve glucose tolerance and uptake [73]. One study indicated that in obese subjects, mirabegron improved glucose tolerance and insulin sensitivity [74]. Another clinical trial indicated that a supratherapeutic dose of 200 mg of mirabegron in healthy subjects stimulated BAT glucose uptake and resting metabolic rate [75]. Other researchers proposed that glucose uptake in BAT of both in human subjects and in vitro is modulated by GLUT4 and uncoupling protein 1 (UCP1) is diurnal bound [76]. On another note, cold induced BAT activation increased glucose uptake in the supraclavicular and paraspinal regions [77], but it remains unclear whether β3-AR are involved in this effect.
In conclusion, β3-AR agonists consistently seemed to improve glucose uptake in various organs in human and animal models. This effect was more prominent in BAT and was mostly associated with a better metabolic status. However, given the limited presence of BAT in adult humans, it remains unclear how much this contributes to systemic glucose handling. Yet, more functional studies must be done, especially in humans to further support these conclusions.
4. Lipolysis, adipose tissue remodeling and obesity
AT is unlike other organs in the body. It has peculiar morphology, physiology and function that differ based on the location and the shade of the fat pad [78]. WAT mainly consists of unilocular adipocytes with lower mitochondrial and UCP1 expression and a large single lipid droplet; it chiefly serves as energy reservoir in addition to secretory function of various adipokines and hormones [79,80,81]. Plus, it counts for almost 80% of total body fat, that is primarily found in subcutaneous and visceral pools [82]. On the other hand, BAT is a more heterogenous pool of adipocytes; the majority are of multilocular morphology with higher mitochondrial and UCP1 levels and small lipid droplets, hence the brown color [78,83]. BAT is largely used in thermogenic responses, using UCP1 to dissipate energy as heat rather than ATP [84,85,86]. Accordingly, BAT plays an important physiological role in rodents and hibernating animals but less so in other groups of mammals including humans. While it has long been assumed that adult humans lack BAT, more recent studies including those using fluorodeoxyglucose positron emission tomography have revealed the presence of at least some BAT in adult humans [87]. These were located differently than in rodents, i.e., mostly in the supraclavicular and neck regions. However, these new findings do not affect the more general concept that BAT plays only a minor role in adult humans as the prevalence of BAT does not exceed some tens of percent. This may be too little to mediate robust systemic lipolytic responses.
Human brown adipocytes have been described to be comparable to murine beige adipocytes rather than brown, which are distinguished by having intermediate characteristics of white and brown adipocytes (unilocular and multilocular, respectively) [88]. Beige AT expresses specific genes such as transcription factor Tbx1, fatty acid transporter Slc27a1, as well as CD40 and CD137 [88]. Beige (also known as brite) adipocytes are the latest to be identified among the different shades of adipocytes; they are multilocular with fewer mitochondrial and UCP1 count than brown, but with more lipid droplets than white adipocytes [88,89]. Hence, the term beiging or browning refers to the transformation of WAT into a pool with beige characteristics in response to wide range of stimuli, increasing its thermogenic capacity, which has been associated with numerous positive metabolic outcomes and a target to treat these anomalies [90].
Adrenergic activation is known to be a positive modulator of metabolism, adipose physiology and activity in both human and animal models [75,91]. Still, divergence in adrenergic response and expression in AT have been documented across species [92,93]. Henceforth, in this section we will dwell further on the role of β3-AR and AT across species.
4.1. β3-Adrenoceptor expression in adipose tissue
Numerous reports have described the presence of β3-AR in rodent AT. This includes rat [24,66,94,95,96,97] and mouse WAT [98,99,100] and rat [24,95] and mouse BAT [98,99,100,101]. Among the two splice variants of murine β3-AR, the β3b-AR dominates in WAT, whereas the β3a-AR does so in BAT [99]. Of note, β3-AR mRNA expression in rodent AT markedly exceeds that of β1- and β2-AR [24]. β3-AR mRNA was also found abundantly in cell lines derived from murine WAT, e.g., 3T3-F422A cells [67,102,103].
In contrast, the data on β3-AR expression at the mRNA level in human AT are not fully conclusive. Some investigators found it in human WAT [104,105,106], in infant [105] and adult cervical and inter- and suprascapular BAT [107,108,109]; however, other reports did not confirm this [110,111]. Of note, most of these studies only reported qualitatively and lacked comparison to other subtypes. The technically most advanced study in the field comparing expression of all three subtypes in more than 30 human tissues reported that expression in AT was much lower than for the other two subtypes and below the detection limit (β1 2.29, β2 12.60, β3 0.19 fragments per kilobase of transcript length per million of mapped reads) [112] (Table 1). Thus, β3-AR mRNA is abundantly detected in rat and mouse WAT and BAT, whereas its detection in human BAT is inconsistent. Accordingly, β3-AR are the most abundantly expressed β-AR subtype in rodent AT [24] but the least abundantly expressed subtype in human AT [112]. Interestingly, Riis-Vestergaard et al. suggested based on using both CL 316,243 and mirabegron that sympathetic activation of human BAT is mediated by β1-AR activation and not β3-AR [113].
4.2. Lipolysis and thermogenesis
4.2.1. Non-primate animals studies
Lipolysis studies in rodents were instrumental in postulating that a third β-AR subtype must exist [22]. Thus, initial studies from the Zaagsma group and others found that the rank order of potency of various agonist and antagonist to stimulate and inhibit rat AT lipolytic responses, respectively, did not match those at the β1- and β2-AR [17,18,19,54,114,115,116,117,118,119]. However, studies specifically performed in WAT mainly found an involvement of β1-AR [119]. Studies published after the cloning of the human [23], rat [24] and mouse β3-AR [25] confirmed the involvement of β3-AR in lipolytic responses in rats [120,121,122,123,124,125,126,127,128]. Thus, lipolytic responses in rats have a strong β3-AR involvement, although some of these studies have detected a β1-AR contribution. Interestingly, the lipolytic response isoprenaline or CL 316,243 in rat AT in the absence of insulin did not depend on the presence of the enzyme adenosine deaminase, whereas the stimulation in the presence of insulin was markedly attenuated by presence of the enzyme, with even stronger inhibition by the enzyme in the combined presence of insulin and glucose [68].
The initial report on the cloning of the murine β3-AR described that the potency of various agonists to stimulate cAMP formation in transfected CHO cells correlated well with that to induce lipolysis in rat brown adipocytes [25]. Subsequent studies confirmed the involvement of β3-AR in lipolytic responses in mice [44,129]. Studies in dogs also found that lipolytic responses are largely mediated by β3-AR [130,131].
4.2.2. Human and primate studies
The role of β3-AR in lipolysis in humans and primates has been investigated extensively. An early study compared WAT from rat, dog, marmoset (Callithrix jacchus), baboon (Papio papio), macacque (Macaca fascicularis), and humans [132]. Isoprenaline was similarly potent in all six species. The intrinsic activity (expressed as fraction of maximum isoprenaline response) was about 1 in all species for noradrenaline. It also was about 1 for the β1-AR agonist dobutamine in rat and dog, about 0.9 in marmoset, and about 0.7 for baboon, macaque, and human. In contrast, it was about 1 for BRL 37,344 in rat and dog, about 0.6 for marmoset, and about 0.1 or less in baboon, macaque, and human. Other β3-AR agonists including CGP 12,177, CL-316,243, D7114, and SR 58,611 also had low potency and/or low efficacy in the latter three species. Antagonist experiments also supported the view that the lipolytic effects in baboon, macaque and human primarily involved β1- and β2-AR. Studies in isolated subcutaneous adipocytes from rhesus monkey reported that the β3-AR agonist L-750,355 concentration-dependently stimulated lipolysis; while its potency at the cloned monkey β3-AR was 28 nM, the lipolytic response did not reach an identifiable maximum even at 10 µM and was ≤20% of the isoprenaline response in concentrations close to its EC50 at the cloned receptor [133].
Several studies have explored the role of β3-AR in the regulation of lipolysis in humans, and the early studies have been reviewed previously [134]. One of the first experiments found that the activity of stereoisomers of β-AR antagonists could not be explained by involvement of β1/β2-AR in rats, but no such contradiction was found in human AT [19]. Later work from the same group found that BRL 37,344 stimulated lipolysis in human AT with much lower potency than in rat AT [118,126]. BRL 37,344 caused lipolysis in omental and subcutaneous white adipocytes by stimulating β2-adrenoceptors; lipolytic effects of CGP 12,177 may occur at least partly by a receptor distinct from β1-and β3-adrenoceptors [104]. Additionally, alprenolol, an antagonist with low affinity for β3-adrenoceptors, blocked the lipolytic response to isoprenaline in human but not rodent subcutaneous WAT [124], indicating that the human response is not mediated by β3-ARs. Others reported that the lipolytic effect of noradrenaline in human and monkey AT involved β1- and/or β2-AR with no evidence for an involvement of β3-AR [132]. Others found that lipolysis responses to CGP 12,177 were antagonized by bupranolol but only poorly by β1- and β2-AR antagonists; moreover, CGP 12,177 promoted lipolysis more effectively in human omental than in subcutaneous AT [135]. Thermogenic responses to an infusion of isoprenaline were attenuated by atenolol, indicating that they occurred by β1-AR stimulation. Several β3-AR agonists were tested in a follow-up study in human omental adipocytes: While some of them had lipolytic effects (BRL 37,344, CGP 12,177, CL 316,243, SM 11044 with some being only partial agonists), others did not (ICI D7114, SR 58611A, ZD 2079) [136], indicating that a lipolytic response in human AT may be limited to certain compounds and is not a universal response to β3-AR stimulation. Others also found that lipolytic responses in human WAT were only inconsistently achieved with β3-AR agonists [137].
Excitingly, these findings were negated by Cero et al. using mirabegron on human derived brown adipocytes. Mirabegron stimulated lipolysis and thermogenesis, while silencing β3-AR in BAT blocked these processes [109]. Moreover, a mutation in β3-AR gene reduces lipolysis in human WAT in response to L-755,507 [138]. Furthermore, a dose of 100 mg of mirabegron was enough to induce thermogenesis in supraclavicular skin in humans, without inciting off-target binding in comparison to higher doses [14]. These contradicted lines of evidence further the debate concerning the involvement of β3-adrenoceptors in human lipolysis.
On a different note, thermogenesis is a hallmark of BAT activity, that is chiefly achieved by sympathetic activation and generally associated with positive metabolic outcomes [46,50,83,84,108,109,139]. In healthy adult subjects, paracervical and supraclavicular brown adipose tissues were biopsied; these pools had 1000 times higher UCP1 expression than the adjacent WAT, with higher glucose uptake after cold exposure [85]. Interestingly, caffeine intake was suggested to ignite BAT thermogenesis in humans since it induces overall positive metabolic effect in obese and non-obese subjects, including thermogenesis, lipolysis, improved glucose tolerance and insulin [59,140,141]. Consequently, it was implied that caffeine might intercede β3-AR activation in BAT [142]. Since thermogenesis can be achieved by cold induction and other safe compounds such as caffeine, the use of β3-AR agonists to activate this process might not be a feasible option.
4.2.3. In vitro studies
Brown adipocytes, sourced from adult cynomolgus monkeys, were the focal point of a study that delved into the roles of β-AR in triggering lipolysis and thermogenesis. This investigation, employing agonists for the three β-AR subtypes (β1, β2, and β3), namely denopamine, procaterol, and CGP12177A, unveiled their shared responsibility in these metabolic processes [143]. Another study on immortalized brown adipocytes showed the additive role of β-AR subtypes in increasing cellular cAMP formation, evidenced by the activation of adenylyl cyclase by noradrenaline after these cells being incubated with CL 316,243. Hence, this activation was achieved by multiple β-AR subtypes, not only by β3-AR. Moreover, all β-AR agonists induced UCP1 expression, and the maximal rate was obtained by isoproterenol (100microM) [144]. Still, primary cell culture of mouse BAT showed that β3-AR activation using BRL 37,344 was the most effective one in inducing UCP1 synthesis [145]. Also, heat production from brown adipocytes of older rats (40 weeks old) and obese ones was lower compared to their younger littermates (12 weeks), which was activated equally by β3-AR agonist BRL 37,344 compared to non-selective agonist isoproterenol which exerted a significant lower response in obese rats [146]. Furthermore, mirabegron stimulated UCP1 expression in mouse brown adipocytes and 3T3-L1 cells [147].
Most recently, immortalized human brown adipocytes were studied using not only subtype-selective agonists but also receptor knock-down experiments to determine the β-AR subtype involved in lipolysis and UCP1 expression [113]. Lipolysis and UCP1 expression were stimulated by isoprenaline and dobutamine but not by procaterol, CL 316,243, or mirabegron. Similarly, knock-down of β1-AR attenuated the isoprenaline-induced UCP1 expression. These data strongly support the idea that β1-AR and not β3-AR are largely responsible for the modulation of human BAT.
In conclusion, a major contribution of β3-AR to lipolytic responses has consistently been shown in rats, mice, and dogs. On the other hand, a role of β3-AR has in humans has been reported only inconsistently and equivocally. Additionally, where detected, β3-AR often played a smaller role than β1- and/or β2-AR. Henceforth, studying β3-AR involvement in murine models can be considered irrelevant to human physiology.
4.3. Adipose tissue remodeling
AT is a dynamic endocrine organ that exhibits physiological changes in response to a range of stimuli, both positive and negative [148,149]. These alterations mainly involve variations in the adipokines profile, cell heterogeneity particularly within brown and beige pools, and thermogenic capacity that is mainly driven by UCP1. This process is referred to as adipose tissue remodeling [148,149,150,151,152].
Sympathetic activation is essential to maintain healthy function and plasticity of AT, inducing positive remodeling [109,153,154,155]. Cold has been used as a natural sympathetic activator of BAT [77,156,157]. Pharmacological interventions have been used to modulate and activate AT as well. For example, mirabegron was found to improve subcutaneous WAT (scWAT) dysfunction and to induce positive adipose remodeling indicated by increasing UCP1 expression and lipolysis in obese and insulin resistant human subject [74].
Notwithstanding, it has been debated whether the upper hand in sympathetic activation of AT is mediated by β3-AR or not across species. As shown in a recent study by Blondin et al., adrenergic mediated thermogenesis and lipolysis of BAT is driven by different adrenergic receptors. In humans it was found to be mediated through β2-AR activation rather than β3-AR, in rodents the opposite was documented. Not only the activation but rather the expression of AR was different, human BAT had a higher expression of β2-AR as well[93].
As stated before, adipose tissue, particularly BAT, exhibits considerable disparities between humans and rodents, encompassing differences in localization, physiology, morphology, and function. Importantly, BAT mass is notably lower in humans compared to rodents [86,158]. Different β-ARs subtypes are active and predominant as well. β3-AR is believed to be the key modulator of positive AT remodeling in rodents[79].
In fact, treatment with CL 316,243 for one week was found to induce AT remodeling in Wistar rats [72]. It also doubled the metabolic rate and raised body temperature, induced WAT browning and activation in wild type C57Bl/6 mice [159]. Indeed, CL 316,243 infusion in Wistar rats fed on high fat diet, found to increase FFA uptake and lipolysis in BAT as well [160]. In high fat fed mice, mirabegron treatment (2mg/kg of body weight) for 3 weeks seemed to lower body weight and adiposity. It reduced brown adipocytes’ size in the interscapular pool while increasing UCP1 expression , increasing beiging in the inguinal depot, and improved insulin sensitivity and glucose tolerance compared to the vehicle treated mice [147].Moreover, knockout of β3-AR impaired lipolysis in mice [158], upregulation of β1-AR mRNA in BAT and WAT, and increased adiposity [101]. These findings emphasize the role of β3-AR in rodent AT in inducing metabolic activity. These positive metabolic alterations in BAT were produced by caffeine intake in obese and non-obese rodents [141,161], giving off more convenient options in inducing positive adipose remodeling than β3-AR agonists.
The wide gap in results concerning AAT remodeling and β3-AR across the species, and in vivo vs. in vitro make translational research less valid using different β3-AR agonists in targeting AT dysfunction.
5. Obesity
Being a metabolic sink for glucose and FFA, BAT activation has been considered a corner stone in improving metabolic health, since it is correlated to improved glucose and insulin sensitivity [9,10,11,12,162]. Studies with ephedrine [46,47,48] and other sympathomimetic agents including various β-AR agonists had reported increased thermogenesis and weight loss in the genetically obese ob/ob mice [49,50]. Therefore, it was suggested that targeting β3-AR in BAT pharmacologically has therapeutic potential for metabolic dysfunction and obesity [12,13,14,22,163].
5.1. Non- primate animal studies
Conclusive evidence regarding the β3-AR/AT axis in developing obesity and metabolic derangements has been presented in rodents. For instance, in lean and obese Zucker diabetic fatty rats (a model of type 2 diabetes), CL 316,243 infusion revealed an anti-obesity and anti-diabetes effect by improving glucose tolerance, insulin sensitivity, thermogenesis, mitochondrial biogenesis, WAT, BAT and skeletal muscle glucose uptake and reducing plasma FFA levels [164]. Administration of CL 316243 in rats fed on high fat diet, seemed to increase energy expenditure, UCP1 in BAT, and prevented WAT hyperplasia [165]. Moreover, treating the ob/ob mouse model of obesity with BRL 37,344 seemed to rescue from metabolic dysfunction, by improving systemic levels of glucose, FFA and insulin. Importantly, β3-AR agonists reduced fat mass without affecting lean body mass in multiple studies in ob/ob mice [22]. Interestingly, chronic infusion of 1 mg/d of BRL 37,344 for 20 days increased UCP1 mRNA expression in BAT compared to acute intervention for 1 day [166]. BRL 35,135 also caused weight loss in ob/ob mice and in Zucker diabetic fatty rats.
On the transgenic level, UCP-DTA transgenic mice with toxigene-mediated ablation of BAT fed on western diet were significantly obese on week 12 of feeding, which was not stimulated by hyperphagia compared to the wild type. They also presented deleterious metabolic anomalies such as insulin resistance, glucose intolerance and hyperlipidemia. Moreover, these mice seemed to have lower rates of GLUT4 and β3-AR mRNA, and increased expression of tumor necrosis factor-α compared to the control littermates [167]. Similar findings were reported by Lowell et al. using the same transgenic mouse model in the absence of western diet. Moreover, using CL 316,243 exerted 50% lower thermogenic activity compared to the wild type [168]. Another model of β3-AR knockout mice, showed that after 8 weeks of high fat diet, the mice had glucose intolerance with hyperlipidemia, increased adiposity of WAT with inflammatory markers compared to the wild type [158].
5.2. Human and primate studies
In non-human primates, prepubertal baboons, different β3-AR agonists including SR 58,611A, BRL 37,344, CGP 12,177, and CL 316,243, did not seem to exert metabolic activity in WAT and BAT; furthermore, β3-AR mRNA was not found to be abundant in these fat pads as well, which was also not correlated to UCP1 expression [169].
Body mass index and body compositions were found to be strongly associate with lower BAT activity in morbid obese individuals as well as cold induced thermogenesis [170]. As a matter of fact, a study on BAT+ and BAT- men with similar age, body mass index and adiposity showed that after prolonged cold exposure, insulin sensitivity, glucose regulation and resting metabolic rate were all increased in BAT+ subjects only [171]. Another study evaluated BAT activity by integrated positron-emission tomography and computer tomography scanning with 18F-fluorodeoxyglucose in 14 overweight and obese compared to 10 lean men: BAT activity was found during cold exposure but not under thermoneutral conditions, and it was lower in obese than in lean subjects [156].
Noteworthy, comparing cold induced thermogenesis with mirabegron treatment in lean and obese subjects from both sexes, showed that upon 10 days of placing ice pack on one thigh for 30 minutes was enough to induce beiging in subcutaneous AT in both legs. Whereas, 10 weeks of daily 50 mg of mirabegron showed signs of beiging indicated by UCP1 expression from the same pool. In addition to that, no sex differences were found in beiging of scWAT in both arms of intervention [157]. Intriguingly, ephedrine did not seem to induce thermogenesis in intrascapular fat in healthy subjects, although it induced oxygen intake, flood flow to the area and skin temperature, in addition to lipolysis. These results indicate the possible increased temperature due to increased blood flow induced by sympathetic activation rather than thermogenesis, since brown adipocytes were not detected from the biopsies [172].
Importantly, it was suggested that mutations of β3-AR gene, especially Trp64Arg mutation, were associated with obesity and metabolic dysfunction in humans [33,37,173,174,175,176]. Contradicted data were reported, where scientists found no association between of β3-AR gene mutation and obesity or adiposity[177,178,179]. As such, results from two cohorts; the Québec Family Study and the Swedish Obese Subjects, concluded that Trp64Arg mutation in β3-AR gene was not associated with cardiometabolic insults, obesity, adiposity, or body composition change over the period of 12 years in the former. Similar outcomes were recorded in the latter, as this mutation wasn’t associated with any weight gain over time and no difference was found in this gene between the obese and non-obese subjects [178].
The current body of evidence concerning the involvement of β3-AR in obesity remains inconclusive, leaving uncertainty about whether mutations in β3-AR contribute to the pathogenesis of obesity or aid in its management by activating it. This ambiguity casts doubt on the efficacy and adequacy of targeting β3-AR in AT as a therapeutic approach for tackling obesity.
6. Clinical development programs
Based on the promising results in rodents and on some of the gene polymorphism studies, several pharmaceutical companies have launched clinical development programs for their β3-AR agonists in obese and/or diabetic patients. These have been reviewed by Larson [180], who had played an active part in the program by Merck & Co.
The Beecham program on BRL 26,830 apparently was the first to enter clinical development and has been tested in at least four trials. A short-term infusion of BRL 26,830 in obese subjects increased insulin sensitivity [181]. Chronic administration studies, published in abstract form only, found an increase in placebo-adjusted energy expenditure across three studies [180]. However, the overall effects on body weight were unimpressive, and they were accompanied by tremor [182], a typical adverse effect from β2-AR stimulation [183]. Therefore, this program was discontinued, and another with the backup compound BRL 35,135 was initiated. Administration of single oral doses of BRL 35,135 or of salbutamol in the absence and presence of antagonists were studied in healthy volunteers [64]. Decreases of serum potassium and increases of glucose, insulin, and lactate were apparently mediated by β2-AR; an increase in FFA occurred with BRL 35,135 but not with salbutamol; while both agonists increased basal metabolic rate, this was a β2 response for salbutamol and possibly included a β3 component for BRL 35,135. The induction of a thermogenic response to BRL 35,135 in non-obese men was confirmed in another study [184], but this did not determine the β-AR subtype being involved. Treatment of obese subjects for 10 days improved insulin sensitivity but did not lower body weight [185]. No subsequent studies were reported, indicating that the program was discontinued because the overall effects on body weight were discouraging.
CL 316,243 has good selectivity for β3- relative to β1- and β2-AR but its efficacy is only about 60% of that of isoprenaline [186]. Despite several studies showing beneficial effects in rats and in immortalized human brown adipocytes [187], it improved insulin effects with only moderate effects on energy expenditure in an 8-week, placebo-controlled clinical trials in healthy lean men [188].
After the phenylethanolamine RO 16-8714 had shown beneficial effects in rodents [94,189], its infusion in humans increased energy expenditure but also heart rate [190,191]. No follow-up studies were reported. ICI D-7114 has been studied in a 14-day, double-blind, randomized trial in obese patients but the results on energy expenditure and body weight also did not support further investigation [192]. Similarly, TAK-677 [193] (0.1 and 0.5 mg b.i.d. for 29 days) resulted in a statistically significant but small increase in energy expenditure relative to placebo in obese patients but did not affect body weight or fasting levels of glucose, insulin or FFA [194].
L-796,568 has been administered in single doses of 250 and 1000 mg to healthy overweight men in a placebo-controlled, 3-way cross-over trial [195]. The 1000 mg dose increased energy expenditure by about 8%, which was accompanied by an increase in plasma glycerol and FFA. While heart rate and diastolic blood pressure remained unchanged, systolic blood pressure increased by about 12 mm Hg. It was also tested in a 28-day placebo-controlled study with daily doses of 375 mg in non-diabetic overweight men [196]. The two groups did not differ in energy expenditure at study end, and glucose tolerance was not altered either. No additional clinical studies were reported.
Mirabegron [197] and solabegron [61] have been in phase II studies in obese/T2DM patients but no clinical outcomes were disclosed by Astellas and GSK, respectively; neither compound advanced to phase III studies in an obesity or T2DM indication. However, based on clinical availability of mirabegron for the overactive bladder syndrome indication [198], academic investigators have performed additional clinical studies. A supra-therapeutic single dose of mirabegron (200 mg) increased BAT metabolic activity in healthy male subjects as assessed by fluorodeoxyglucose in positron emission tomography [199]. In a follow-up study with single doses of 50 and 200 mg mirabegron, the effect on BAT metabolic activity increased more than dose-proportionally by the greater dose [108].
Based on the accumulated clinical evidence it has been concluded that β3-AR agonists yielded “a statistically significant elevation in total energy expenditure but this did not translate into a biologically meaningful negative energy balance”, i.e., a measurable loss of body weight [180].
7. Why do rodent and primate studies differ?
While the above data show that β3-AR are a promising target for the treatment of obesity, and perhaps T2DM, in rodents, the data in humans and non-human primates are less conclusive and generally show quantitatively much smaller if any effect (Table 2, Figure 1). Accordingly, clinical development programs of β3-AR agonists by multiple pharmaceutical companies yielded small effects on thermogenesis that failed to translate into clinically relevant effects on body weights and led to the discontinuation of such programs. The above data indicate why a drug target promising in rodents was insufficiently responsive in humans.
Firstly, the relative lack of efficacy in obese or diabetic humans is not due poor efficacy of drug candidates at the human receptor: Compounds that failed in clinical studies in obese and/or diabetic patients including mirabegron have been highly successful in other indications such as the overactive bladder syndrome. Secondly, a key factor in such species differences is the abundance of BAT which is high in rodents but sparse in adult humans. While some maneuvers can promote the beiging of AT in humans, the presence of BAT or beige AT relative to body weight appears limited in humans. Thirdly, β3-AR agonists promote pancreatic insulin release in rodents but have limited effects in humans. Fourthly, the expression of β3-AR in human AT appears to be considerably lower in humans than in rodents. While the lipolysis response in rodent WAT has a considerable β3-AR component, that in human WAT is largely carried by β1-AR. Accordingly, the lipolytic and thermogenic responses in humans upon systemic administration are much smaller than in rodents. All of these factors in combination lead to a pronounced body weight reduction in rodents but not in men.
8. Conclusion and future perspectives
Rodent studies generally support the role of β3-AR as a target for the treatment of obesity and diabetes, whereas human studies mostly do not. This includes studies at the genetic level (gene knock-out studies in mice vs. genome-wide association studies in humans; see section 2.3). Despite commercial efforts in various pharmaceutical companies (see sections 6 and 9), no β3-AR agonist has become a clinically available drug for the treatment of obesity and/or T2DM. To the contrary, at least nine β3-AR agonists have been tested clinically, some even in phase II studies, but failed to provide efficacy signals of sufficient strength to merit further development as anti-obesity/anti-diabetic drugs. In contrast, multiple β3-AR agonists have shown efficacy in patients with overactive bladder syndrome [61] and two of them, mirabegron [198] and vibegron [200] have become approved and guideline recommended treatments for this condition. In some cases such as mirabegron had insufficient effects sizes in the obesity/T2DM indication but became approved drugs in the overactive bladder indication, indicating that the disappointing clinical data in the obesity/T2DM indication were not due to testing of an ineffective compound. Clinical development efforts in obesity and/or T2DM largely failed because of a sparse present of BAT in adult humans, differences in β3-AR expression, and other factors (see section 6). As obesity is a market of huge potential commercial interest, it is telling that presently no major pharmaceutical company appears to be active in this space, including those who had active programs and discontinued them.
Some academic efforts have focused on inducing BAT or at least ‘beiging’ of WAT but it remains to be seen whether this will lead to a more successful use of β3-AR agonists as weight-lowering/anti-diabetic treatment. We personally are skeptical about these efforts for three main reasons: Firstly, clinical studies using β3-AR with various chemical structures have mostly found stimulation of thermogenesis in acute studies (single administration), whereas the limited chronic studies (multiple weeks) typically did not confirm this. This points to a possible role of desensitization of β3-AR [201]. Second, while β3-AR agonists generally are well tolerated, at least for mirabegron warnings on cardiovascular effects in a small fraction of patients have been issued [112]. It has been speculated that such adverse effects are related to phenylethanolamine backbone, which is present in mirabegron and several other β3-AR agonists and may cause indirect sympathomimetic activity [202]. Moreover, at least mirabegron has additional off-target effects such as antagonism at α1-adrenoceptors [203,204,205]. Such off-target effects were neither reported nor are expected for the β3-AR with other chemical backbones.
Third and most importantly, the key question is not whether β3-AR agonists will cause some weight loss relative to placebo; to make a professional clinical development program commercially viable they must match the success glucagon-like peptide 1 receptors such as semaglutide. Of note, compounds combining such agonism with that at receptors for glucose-dependent insulinotropic polypeptide, e.g., tirzepatide, apparently cause even greater weight loss [5]. We consider it highly unlikely that β3-AR agonists even with substantial beiging will ever reach such efficacy.
9. Patents
Following the lead of Beecham Pharmaceuticals (now a part of GSK) and its original drug discovery programs of β3-AR agonists as potential treatments of obesity [54], numerous pharmaceutical companies have filed patents seeking exclusivity for their β3-AR agonists as potential treatment of obesity and/or diabetes as reviewed elsewhere [206]. These patents cover thousands of compounds of various chemical structures [207]. They include patents from companies such as American Home Products, American Cyanamid Co., Asahi Kasei Pharma Corporation, Astellas Pharma, Bayer AG, Boehringer Ingelheim International, Bristol-Myers Squibb Co., Dainippon Pharmaceutical Co., Eli Lilly & Co., Fujisawa Pharmaceutical Co., Glaxo Group Ltd., Glenmark Pharmaceuticals Ltd., Imperial Chemical Industries Plc, Kaneka Corporation, Kissei Pharmaceutical, Merck & Co., Pfizer Inc., Sanofi SA, Smithkline Beecham Plc, Sumitomo Pharma, Tokyo Tanabe Co. Ltd., and Toyko Shinyaku Co. Ltd.; for details see [206]. Several of the compounds covered in those patents have entered clinical development (see section 6), but none has proceeded beyond phase II or even been approved in the obesity/T2DM therapeutic field.
Author Contributions
H.D. and M.C.M. jointly provided the conceptualization, performed the literature analysis, and drafted the manuscript.
Funding
This research received no external funding. Research on the role of β3-ARs in control of urinary bladder function in obese and diabetic animals in the author’s lab is funded by Deutsche Forschungsgemeinschaft (Mi 294/10-1).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
M.C.M is author of some patent applications in the β3-AR field, but none of them led to an awarded patent. Therefore, the authors declare no conflict of interest related to this manuscript.
References
- Blüher, M. Obesity: global epidemiology and pathogenesis. Nat. Rev. Endocrinol. 2019, 15, 288-298. [CrossRef]
- Zhang, L.; Zhu, L.; Xu, T.; Lang, J.; Li, Z.; Gong, J.; Liu, Q.; Liu, X. A population-based survey of the prevalence, potential risk factors, and symptom-specific bother of lower urinary tract symptoms in adult Chinese women. Eur. Urol. 2015, 68, 97-112. [CrossRef]
- Michel, M.C.; Cardozo, L.; Chermansky, C.J.; Cruz, F.; Igawa, Y.; Lee, K.S.; Sahai, A.; Wein, A.J.; Andersson, K.E. Current and emerging pharmacological targets and treatments of urinary incontinence and related disorders. Pharmacol. Rev. 2023, 75, 554-674. [CrossRef]
- Bergmann, N.C.; Davies, M.J.; Lingvay, I.; Knop, F.K. Semaglutide for the treatment of overweight and obesity: A review. Diabetes Obes. Metab. 2023, 25, 18-35. [CrossRef]
- Prillaman, M. Four key questions on the new wave of anti-obesity drugs. Nature 2023, 620, 28-30. [CrossRef]
- Michel, M.C.; Mayoux, E.; Vallon, V. A comprehensive review of the pharmacodynamics of the SGLT2 inhibitor empagliflozin in animals and humans. Naunyn Schmiedebergs Arch. Pharmacol. 2015, 388, 801-816. [CrossRef]
- Zhang, Y.-J.; Han, S.-L.; Sun, X.-F.; Wang, S.-X.; Wang, H.-Y.; Liu, X.; Chen, L.; Xia, L. Efficacy and safety of empagliflozin for type 2 diabetes mellitus: Meta-analysis of randomized controlled trials. Medicine 2018, 97, e12843-e12843. [CrossRef]
- Procino, G.; Carmosino, M.; Milano, S.; Dal Monte, M.; Schena, G.; Mastrodonato, M.; Gerbino, A.; Bagnoli, P.; Svelto, M. b3 adrenergic receptor in the kidney may be a new player in sympathetic regulation of renal function. Kidney Int. 2016, 90, 555-567. [CrossRef]
- Chen, K.Y.; Brychta, R.J.; Sater, Z.A.; Cassimatis, T.M.; Cero, C.; Fletcher, L.A.; Israni, N.S.; Johnson, J.W.; Lea, H.J.; Linderman, J.D. Opportunities and challenges in the therapeutic activation of human energy expenditure and thermogenesis to manage obesity. Journal of biological chemistry 2020, 295, 1926-1942.
- Darcy, J.; Tseng, Y.-H. ComBATing aging—does increased brown adipose tissue activity confer longevity? Geroscience 2019, 41, 285-296.
- Aldiss, P.; Betts, J.; Sale, C.; Pope, M.; Budge, H.; Symonds, M.E. Exercise-induced ‘browning’of adipose tissues. Metabolism 2018, 81, 63-70.
- Bel, J.S.; Tai, T.; Khaper, N.; Lees, S.J. Mirabegron: The most promising adipose tissue beiging agent. Physiological Reports 2021, 9, e14779.
- O’Mara, A.E.; Johnson, J.W.; Linderman, J.D.; Brychta, R.J.; McGehee, S.; Fletcher, L.A.; Fink, Y.A.; Kapuria, D.; Cassimatis, T.M.; Kelsey, N.; et al. Chronic mirabegron treatment increases human brown fat, HDL cholesterol, and insulin sensitivity. The Journal of Clinical Investigation 2020, 130, 2209-2219. [CrossRef]
- Loh, R.K.; Formosa, M.F.; La Gerche, A.; Reutens, A.T.; Kingwell, B.A.; Carey, A.L. Acute metabolic and cardiovascular effects of mirabegron in healthy individuals. Diabetes, Obesity and Metabolism 2019, 21, 276-284.
- Cawthorne, M.A.; Carroll, M.J.; Levy, A.L.; Lister, C.A.; Sennitt, M.V.; Smith, S.A.; Young, P. Effects of novel beta-adrenoceptor agonists on carbhohydrate metabolism: relevance for the treatment of non-insulin-dependent diabetes. Int. J. Obes. 1984, 8 Suppl. 1, 93-102.
- Lands, A.M.; Arnold, A.; McAuliff, J.P.; Luduena, F.P.; Brown, T.G. Differentiation of receptor systems activated by sympathetic amines. Nature 1967, 214, 597-598.
- Furchgott, R.F. The classification of adrenoceptors (adrenergic receptors). An evaluation from the standpoint of receptor theory. In Catecholamines, Blaschko, H., Muecholl, E., Eds.; Springer Verlag: New York, 1972; pp. 283-335.
- Harms, H.H.; Zaagsma, J.; van der Wal, B. Beta-adrenoceptor studies. III. on the beta-adrenoceptors in rat adipose tissue. Eur. J. Pharmacol. 1974, 25, 87-97. [CrossRef]
- Harms, H.H. Stereochemical aspects of beta-adrenoceptor antagonist-receptor interactions in adipocytes. Differentiation of beta-adrenoceptors in human and rat adipocytes. Life Sci. 1976, 19, 1447-1452. [CrossRef]
- Bianchetti, A.; Manara, L. In vitro inhibition of intestinal motility by phenylethanolaminetetralines: evidence of atypical ß-adrenoceptors in rat colon. Br. J. Pharmacol. 1990, 100, 831-839.
- Nergardh, A.; Boreus, L.O.; Naglo, A.S. Characterization of the adrenergic beta-receptor in the urinary bladder of man and cat. Acta Pharmacol. Toxicol. (Copenh.) 1977, 40, 14-21.
- Arch, J.R.S. Perspectives from ß3-adrenoceptor agonists on pharmacology, physiology and obesity drug discovery. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2008, 378, 225-240. [CrossRef]
- Emorine, L.J.; Marullo, S.; Briden-sutren, M.M.; Patey, G.; Tate, K.; Delavier-Klutchko, C.; Strosberg, A.D. Molecular characterization of the human ß3-adrenergic receptor. Science 1989, 245, 1118-1121.
- Muzzin, P.; Revelli, J.P.; Kuhne, F.; Gocayne, J.D.; McCombie, W.R.; Venter, J.C.; Giacobino, J.P.; Fraser, C.M. An adipose tissue-specific ß-adrenergic receptor. Molecular cloning and down-regulation in obesity. J. Biol. Chem. 1991, 266, 24053-24058.
- Nahmias, C.; Blin, N.; Elalouf, J.M.; Mattei, M.G.; Strosberg, A.D.; Emorine, L.J. Molecular characterization of the mouse ß3-adrenergic receptor: relationship with the atypical receptor of adipocytes. EMBO J. 1991, 10, 3721-3727.
- Granneman, J.G.; Lahners, K.N.; Rao, D.D. Rodent and human ß3-adrenergic receptor genes contain an intron within the protein-coding block. Mol. Pharmacol. 1992, 42, 964-970.
- Granneman, J.G.; Lahners, K.N.; Chaudhry, A. Characterization of the human ß3-adrenergic receptor gene. Mol. Pharmacol. 1993, 44, 264-270.
- Van Spronsen, A.; Nahmias, C.; Krief, S.; Briend-Sutren, M.-M.; Strosberg, A.D.; Emorine, L.J. The promoter and intron/exon structure of the human and mouse β3-adrenergic-receptor genes. Eur. J. Biochem. 1993, 213, 1117-1124. [CrossRef]
- Brown, J.A.; Machida, C.A. The 5′ flanking region of the rat β3-adrenergic receptor gene: divergence with the human gene and implications for species-specific gene expression. DNA Seq. 1994, 4, 319-324. [CrossRef]
- Clement, K.; Vaisse, C.; Manning, B.S.J.; Basdevant, A.; Guy-Grand, B.; Riuz, J.; Silver, K.D.; Shuldiner, A.R.; Froguel, P.; Strosberg, A.D. Genetic variation in the ß3-adrenergic receptor and an increased capacity to gain weight in patients with morbid obesity. N. Engl. J. Med. 1995, 333, 352-354. [CrossRef]
- Walton, J.; Silver, K.; Bogardus, C.; Knowler, W.C.; Celi, F.S.; Austin, S.; Manning, B.; Strosberg, A.D.; Stern, M.P.; Raben, N.; et al. Time of onset of non-insulin-dependent diabetes mellitus and genetic variation in the ß3-adrenergic receptor gene. N. Engl. J. Med. 1995, 333, 343-347. [CrossRef]
- Li, L.S.; Lönnqvist, F.; Luthman, H.; Arner, P. Phenotypic characterization of the Trp64Arg polymorphism in the beta 3-adrenergic receptor gene in normal weight and obese subjects. Diabetologia 1996, 39, 857-860. [CrossRef]
- Umekawa, T.; Yoshida, T.; Sakane, N.; Kogure, A.; Kondo, M.; Honjyo, H. Trp64Arg Mutation of ß3-adrenoceptor gene deteriorates lipolysis by ß3-adrenoceptor agonist in human omental adipocytes. Diabetes 1999, 48, 117-120.
- Kimura, K.; Sasaki, N.; Asano, A.; Mizukami, J.; Kayahashi, S.; Kawada, T.; Fushiki, T.; Morimatsu, M.; Yoshida, T.; Saito, M. Mutated human ß3-adrenergic receptor (Trp64Arg) lowers the response to ß3-adrenergic agonists in transfected 3T3-L1 preadipocytes. Horm. Metab. Res. 2000, 32, 91-96.
- Kadowaki, H.; Yasuda, K.; Iwamoto, K.; Otabe, S.; Shimokawa, K.; Silver, K.; Walston, J.; Yoshinaga, H.; Kosaka, K.; Yamada, N.; et al. A mutation in the ß3-adrenergic receptor gene is associated with obesity and hyperinsulinemia in Japanese subjects. Biochem. Biophys. Res. Commun. 1995, 215, 555-560. [CrossRef]
- Snitker, S.; Odeleye, O.E.; Hellmér, J.; Boschmann, M.; Monroe, M.B.; Shuldiner, A.R.; Ravussin, E. No effect of the Trp64Arg beta 3-adrenoceptor variant on in vivo lipolysis in subcutaneous adipose tissue. Diabetologia 1997, 40, 838-842. [CrossRef]
- Büettner, R.; Schäffler, A.; Arndt, H.; Rogler, G.; Nusser, J.; Zietz, B.; Enger, I.; Hügl, S.; Cuk, A.; Schölmerich, J.; Palitzsch, K.D. The Trp64Arg polymorphism of the ß3-adrenergic receptor gene is not associated with obesity or type 2 diabetes mellitus in a large population-based Caucasian cohort. J. Clin. Endocrinol. Metab. 1998, 83, 2892-2897. [CrossRef]
- Janssen, J.A.M.J.L.; Koper, J.W.; Stolk, R.P.; Englaro, P.; Uitterlinden, A.G.; Huang, Q.; van Leeuwen, J.P.T.M.; Blum, W.F.; Attanasio, A.M.F.; Pols, H.A.P.; et al. Lack of associations between serum leptin, a polymorphism in the gene for the beta 3-adrenergic receptor and glucose tolerance in the Dutch population. Clin. Endocrinol. (Oxf.) 1998, 49, 229-234. [CrossRef]
- Hoffstedt, J.; Poirier, O.; Thörne, A.; Lönnqvist, F.; Herrmann, S.M.; Cambien, F.; Arner, P. Polymorphism of the human ß3-adrenoceptor gene forms a well-conserved haplotype that is associated with moderate obesity and altered receptor function. Diabetes 1999, 48, 203-205. [CrossRef]
- Ahles, A.; Engelhardt, S. Polymorphic variants of adrenoceptors: physiology, pharmacology and role in disease. Pharmacol. Rev. 2014, 66, 598-637. [CrossRef]
- Michel, M.C. Are β3-adrenoceptor gene polymorphisms relevant for urology? Neurourol. Urodyn. 2023, 42, 33-39. [CrossRef]
- Susulic, V.S.; Frederich, R.C.; Lawitt, J.; Tozzo, E.; Kahn, B.B.; Harper, M.E.; Himms-Hagen, J.; Flier, J.S.; Lowell, B.B. Targeted disruption of the ß3-adrenergic receptor gene. J. Biol. Chem. 1995, 270, 29483-29492. [CrossRef]
- Revelli, J.P.; Preitner, F.; Samec, S.; Muniesa, P.; Kuehne, F.; Boss, O.; Vassalli, J.D.; Dulloo, A.; Seydoux, J.; Giacobino, J.P.; et al. Targeted gene disruption reveals a leptin-independent role for the mouse ß3-adrenoceptor in the regulation of body composition. J. Clin. Invest. 1997, 100, 1098-1106.
- Preitner, F.; Muzzin, P.; Revelli, J.P.; Seydoux, J.; Galitzky, J.; Berlan, M.; Lafontan, M.; Giacobino, J.P. Metabolic response to various ß-adrenoceptor agonists in ß3-adrenoceptor knock-out mice: evidence for a new ß-adrenergic receptor in brown adipose tissue. Br. J. Pharmacol. 1998, 124, 1684-1688. [CrossRef]
- Kaumann, A.J.; Molenaar, P. The low affinity site of the ß1-adrenoceptor and its relevance to cardiovascular pharmacology. Pharmacol. Ther. 2008, 118, 303-336.
- Massoudi, M.; Evans, E.; Miller, D.S. Thermogenic drugs for the treatment of obesity: screening using obese rats and mice. Ann. Nutr. Metab. 1983, 27, 26-37. [CrossRef]
- Yen, T.T.; McKee, M.M.; Bemis, K.G. Ephedrine reduces weight of viable yellow obese mice (Avy/a). Life Sci. 1981, 28, 119-128. [CrossRef]
- Arch, J.R.; Ainsworth, A.T.; Cawthorne, M.A. Thermogenic and anorectic effects of ephedrine and congeners in mice and rats. Life Sci. 1982, 30, 1817-1826. [CrossRef]
- Arch, J.R. The contribution of increased thermogenesis to the effect of anorectic drugs on body composition in mice. Am. J. Clin. Nutr. 1981, 34, 2763-2769. [CrossRef]
- Dulloo, A.G.; Miller, D.S. Thermogenic drugs for the treatment of obesity: sympathetic stimulants in animal models. Br. J. Nutr. 1984, 52, 179-196. [CrossRef]
- Arch, J.R.S. Challenges in β3-adrenoceptor agonist drug development. Ther. Adv. Endocrinol. Metab. 2011, 2, 59-64. [CrossRef]
- Brown, L.; Deighton, N.M.; Bals, S.; Söhlmann, W.; Zerkowski, H.R.; Michel, M.C.; Brodde, O.E. Spare receptors for ß-adrenoceptor-mediated positive inotropic effects of catecholamines in the human heart. J. Cardiovasc. Pharmacol. 1992, 19, 222-232.
- Arch, J.R.S. ß3-Adrenoceptor agonists: potential, pitfalls and progress. Eur. J. Pharmacol. 2002, 440, 99-107.
- Arch, J.R.; Ainsworth, A.T.; Cawthorne, M.A.; Piercy, V.; Sennitt, M.V.; Thody, V.E.; Wilson, C.; Wilson, S. Atypical beta-adrenoceptor on brown adipocytes as target for anti-obesity drugs. Nature 1984, 309, 163-165.
- Cernecka, H.; Sand, C.; Michel, M.C. The odd sibling: features of ß3-adrenoceptor pharmacology. Mol. Pharmacol. 2014, 86, 479-484. [CrossRef]
- Yoshida, T. The antidiabetic ß3-adrenoceptor agonist BRL 26830A works by release of endogenous insulin. Am. J. Clin. Nutr. 1992, 55, 237S-241S.
- Yoshida, T.; Yoshioka, K.; Hiraoka, N.; Umekawa, T.; Sakane, N.; Kondo, N. Effects of CL 316,243, a novel ß3-adrenoceptor agonist, on inuslin secretion in perfused mouse pancreas. Endocr. J. 1994, 41, 671-675.
- Perfetti, R.; Hui, H.; Chamie, K.; Binder, S.; Seibert, M.; McLenithan, J.; Silver, K.; Walston, J.D. Pancreatic ß-cells expressing the Arg64 variant of the ß3-adrenergic receptor exhibit abnormal insulin secretory activity. J. Mol. Endocrinol. 2001, 27, 133-144. [CrossRef]
- Atef, N.; Lafontan, M.; Double, A.; Helary, C.; Ktorza, A.; Penicaud, L. A specific ß3-adrenoceptor agonist induces pancreatic islet blood flow and insulin secretion in rats. Eur. J. Pharmacol. 1996, 298, 287-292.
- Guimaraes, S.; Moura, D. Vascular adrenoceptors: an update. Pharmacol. Rev. 2001, 53, 319-356.
- Ohlstein, E.H.; von Keitz, A.; Michel, M.C. A multicenter, double-blind, randomized, placebo controlled trial of the ß 3 -adrenoceptor agonist solabegron for overactive bladder. Eur. Urol. 2012, 62, 834-840. [CrossRef]
- Uehling, D.E.; Shearer, B.G.; Donaldson, K.H.; Chao, E.Y.; Deaton, D.N.; Adkison, K.K.; Brown, K.K.; Cariello, N.F.; Faison, W.L.; Lancaster, M.E.; et al. Biarylaniline phenethanolamines as potent and selective ß3 adrenergic receptor agonists. J. Med. Chem. 2006, 49, 2758-2771. [CrossRef]
- Grujic, D.; Susulic, V.S.; Harper, M.-E.; Himms-Hagen, J.; Cunningham, B.A.; Corkey, B.E.; Lowell, B.B. β3-adrenergic receptors on white and brown adipocytes mediate β3-selective agonist-induced effects on energy expenditure, insulin secretion, and food intake: a study using transgenic and gene knockout mice. Journal of Biological Chemistry 1997, 272, 17686-17693.
- Wheeldon, N.M.; McDevitt, D.G.; McFarlane, L.C.; Lipworth, B.J. ß-Adrenoceptor subtypes mediating the metabolic effects of BRL 35135 in man. Clin. Sci. 1994, 86, 331-337. [CrossRef]
- El Hadri, K.; Charon, C.; Pairault, J.; Hauquel-De Mouzon, S.; Quignard-Boulange, A. Down-regulation of β3-adrenergic receptor expression in rat adipose tissue during the fasted/fed transition: evidence for a role of insulin. Biochem. J. 1997, 323, 359-364. [CrossRef]
- Carpene, C.; Chalaux, E.; Lizarbe, M.; Estrada, A.; Mora, C.; Palacin, M.; Zorzano, A.; Lafontan, M.; Testar, X. ß3-Adrenergic receptors are responsible for the adrenergic inhibition of insulin-stimulated glucose transport in rat adipocytes. Biochem. J. 1993, 296, 99-105.
- Feve, B.; Pietri-Rouxel, F.; El Hadri, K.; Drumare, M.F.; Strosberg, A.D. Long term phorbol ester treatment down-regulates the ß3-adrenergic receptor in 3T3-F4424 adipocytes. J. Biol. Chem. 1995, 270, 10952-10959. [CrossRef]
- Gokmen-Polar, Y.; Coronel, E.C.; Bahouth, S.W.; Fain, J.N. Insulin sensitizes ß-agonist and forskolin-stimulated lipolysis to inhibition by 2’,5’-dideoxyadenosine. Am. J. Physiol. 1996, 270, C562-C569.
- Shimizu, Y.; Kielar, D.; Minokoshi, Y.; Shimazu, T. Noradrenaline increases glucose transport into brown adipocytes in culture by a mechanism different from that of insulin. Biochem. J. 1996, 314, 485-490.
- Kubo, N.; Kawahara, M.; Okamatsu-Ogura, Y.; Miyazaki, Y.; Otsuka, R.; Fukuchi, K. Evaluation of glucose uptake and uncoupling protein 1 activity in adipose tissue of diabetic mice upon β-adrenergic stimulation. Molecular Imaging and Biology 2019, 21, 249-256.
- Abe, H.; Minokoshi, Y.; Shimazu, T. Effect of a β3-adrenergic agonist, BRL35135A, on glucose uptake in rat skeletal muscle in vivo and in vitro. Journal of endocrinology 1993, 139, 479-486.
- Duffaut, C.; Bour, S.; Prévot, D.; Marti, L.; Testar, X.; Zorzano, A.; Carpéné, C. Prolonged treatment with the beta3-adrenergic agonist CL 316243 induces adipose tissue remodeling in rat but not in guinea pig: 2) modulation of glucose uptake and monoamine oxidase activity. Journal of physiology and biochemistry 2006, 62, 101-111.
- Dehvari, N.; da Silva Junior, E.D.; Bengtsson, T.; Hutchinson, D.S. Mirabegron: potential off target effects and uses beyond the bladder. British journal of pharmacology 2018, 175, 4072-4082.
- Finlin, B.S.; Memetimin, H.; Zhu, B.; Confides, A.L.; Vekaria, H.J.; El Khouli, R.H.; Johnson, Z.R.; Westgate, P.M.; Chen, J.; Morris, A.J. The β3-adrenergic receptor agonist mirabegron improves glucose homeostasis in obese humans. The Journal of clinical investigation 2020, 130, 2319-2331.
- Cypess, A.M.; Weiner, L.S.; Roberts-Toler, C.; Elía, E.F.; Kessler, S.H.; Kahn, P.A.; English, J.; Chatman, K.; Trauger, S.A.; Doria, A. Activation of human brown adipose tissue by a β3-adrenergic receptor agonist. Cell metabolism 2015, 21, 33-38.
- Lee, P.; Bova, R.; Schofield, L.; Bryant, W.; Dieckmann, W.; Slattery, A.; Govendir, M.A.; Emmett, L.; Greenfield, J.R. Brown adipose tissue exhibits a glucose-responsive thermogenic biorhythm in humans. Cell metabolism 2016, 23, 602-609.
- Saito, M.; Okamatsu-Ogura, Y.; Matsushita, M.; Watanabe, K.; Yoneshiro, T.; Nio-Kobayashi, J.; Iwanaga, T.; Miyagawa, M.; Kameya, T.; Nakada, K. High incidence of metabolically active brown adipose tissue in healthy adult humans: effects of cold exposure and adiposity. Diabetes 2009, 58, 1526-1531.
- Peirce, V.; Carobbio, S.; Vidal-Puig, A. The different shades of fat. Nature 2014, 510, 76-83.
- Granneman, J.G.; Li, P.; Zhu, Z.; Lu, Y. Metabolic and cellular plasticity in white adipose tissue I: effects of β3-adrenergic receptor activation. American Journal of Physiology-Endocrinology and Metabolism 2005, 289, E608-E616.
- Trayhurn, P.; Beattie, J.H. Physiological role of adipose tissue: white adipose tissue as an endocrine and secretory organ. Proceedings of the Nutrition Society 2001, 60, 329-339.
- Proença, A.R.; Sertié, R.A.L.; Oliveira, A.; Campaaa, A.; Caminhotto, R.; Chimin, P.; Lima, F.B. New concepts in white adipose tissue physiology. Brazilian journal of medical and biological research 2014, 47, 192-205.
- Wronska, A.; Kmiec, Z. Structural and biochemical characteristics of various white adipose tissue depots. Acta physiologica 2012, 205, 194-208.
- Klingenspor, M.; Bast, A.; Bolze, F.; Li, Y.; Maurer, S.; Schweizer, S.; Willershäuser, M.; Fromme, T. Brown adipose tissue. Adipose tissue biology 2017, 91-147.
- Cannon, B.; Nedergaard, J. Brown adipose tissue: function and physiological significance. Physiological reviews 2004.
- Virtanen, K.A.; Lidell, M.E.; Orava, J.; Heglind, M.; Westergren, R.; Niemi, T.; Taittonen, M.; Laine, J.; Savisto, N.-J.; Enerbäck, S. Functional brown adipose tissue in healthy adults. New England Journal of Medicine 2009, 360, 1518-1525.
- Pinckard, K.M.; Stanford, K.I. The heartwarming effect of brown adipose tissue. Molecular Pharmacology 2022, 102, 460-471.
- Nedergaard, J.; Bengtsson, T.; Cannon, B. Unexpected evidence for active brown adipose tissue in adult humans. Am. J. Physiol. 2007, 293, E444-E452. [CrossRef]
- Wu, J.; Boström, P.; Sparks, L.M.; Ye, L.; Choi, J.H.; Giang, A.-H.; Khandekar, M.; Virtanen, K.A.; Nuutila, P.; Schaart, G. Beige adipocytes are a distinct type of thermogenic fat cell in mouse and human. Cell 2012, 150, 366-376.
- Pilkington, A.-C.; Paz, H.A.; Wankhade, U.D. Beige adipose tissue identification and marker Specificity—Overview. Frontiers in endocrinology 2021, 12, 599134.
- Thyagarajan, B.; Foster, M.T. Beiging of white adipose tissue as a therapeutic strategy for weight loss in humans. Hormone molecular biology and clinical investigation 2017, 31, 20170016.
- Baskin, A.S.; Linderman, J.D.; Brychta, R.J.; McGehee, S.; Anflick-Chames, E.; Cero, C.; Johnson, J.W.; O’Mara, A.E.; Fletcher, L.A.; Leitner, B.P.; et al. Regulation of human adipose tissue activation, gallbladder size, and bile acid metabolism by a β3-adrenergic receptor agonist. Diabetes 2018, 67, 2113-2125. [CrossRef]
- Galitzky, J.; Carpene, C.; Bousquet-Mélou, A.; Berlan, M.; Lafontan, M. Differential activation of β1-, β2-and β3-adrenoceptors by catecholamines in white and brown adipocytes. Fundamental & clinical pharmacology 1995, 9, 324-331.
- Blondin, D.P.; Nielsen, S.; Kuipers, E.N.; Severinsen, M.C.; Jensen, V.H.; Miard, S.; Jespersen, N.Z.; Kooijman, S.; Boon, M.R.; Fortin, M. Human brown adipocyte thermogenesis is driven by β2-AR stimulation. Cell metabolism 2020, 32, 287-300. e287.
- Revelli, J.P.; Muzzin, P.; Giacobino, J.P. Modulation in vivo of ß-adrenergic-receptor subtypes in rat brown adipose tissue by the thermogenic agonist Ro 16-8714. Biochem. J. 1992, 286, 743-746.
- Evans, B.A.; Papaioannou, M.; Bonazzi, V.R.; Summers, R.J. Expression of ß3-adrenoceptor mRNA in rat tissues. Br. J. Pharmacol. 1996, 117, 210-216.
- Adli, H.; Bazin, R.; Perret, G.Y. Interaction of amiodarone and triiodothyronine on the expression of ß-adrenoceptors in brown adipose tissue of rat. Br. J. Pharmacol. 1999, 126, 1455-1461.
- Scarpace, P.J.; Matheny, M.; Thümer, N. Differential down-regulation of ß3-adrenergic receptor mRNA and signal transduction by cold exposure in brown adipose tissue of young and senescent rats. Pflügers Archiv: European Journal of Physiology 1999, 437, 479-483.
- Evans, B.A.; Papaioannou, M.; Anastasopoulos, F.; Summers, R.J. Differential regulation of β3-adrenoceptors in gut and adipose tissue of genetically obese (ob/ob) C57BL/6J-mice. Br. J. Pharmacol. 1998, 124, 763-771. [CrossRef]
- Evans, B.A.; Papaioannou, M.; Hamilton, S.; Summers, R.J. Alternative splicing generates two isoforms of the ß3-adrenoceptor which are differentially expressed in mouse tissues. Br. J. Pharmacol. 1999, 127, 1525-1531. [CrossRef]
- Hutchinson, D.S.; Evans, B.A.; Summers, R.J. ß3-Adrenoceptor regulation and relaxation responses in mouse ileum. Br. J. Pharmacol. 2000, 129, 1251-1259. [CrossRef]
- Susulic, V.S.; Frederich, R.C.; Lawitts, J.; Tozzo, E.; Kahn, B.B.; Harper, M.-E.; Himms-Hagen, J.; Flier, J.S.; Lowell, B.B. Targeted disruption of the β3-adrenergic receptor gene. Journal of Biological Chemistry 1995, 270, 29483-29492.
- Feve, B.; Elhadri, K.; Quignard-Boulange, A.; Pairault, J. Transcriptional down-regulation by insulin of the beta-3 adrenergic receptor expression in 3T3-F442A adipocytes: a mechanism for repressing the cAMP signaling pathway. Proc Natl Acad Sci 1994, 91, 5677-5681.
- Rohlfs, E.M.; Daniel, K.W.; Premont, R.T.; Kozak, L.P.; Collins, S. Regulation of the uncoupling gene (Ucp) by ß1, ß2 and ß3-adrenergic receptor subtypes in immortalized brown adipose cell lines. J. Biol. Chem. 1995, 270, 10723-10732. [CrossRef]
- Lönnqvist, F.; Krief, S.; Strosberg, A.D.; Nyberg, B.; Emorine, L.J.; Arner, P. Evidence for a functional ß3-adrenoceptor in man. Br. J. Pharmacol. 1993, 110, 929-936.
- Krief, S.; Lönnqvist, F.; Raimbault, S.; Baude, B.; van Spronsen, A.; Arner, P.; Strosberg, A.D.; Ricquier, D.; Emorine, L.J. Tissue distribution of beta 3-adrenergic receptor mRNA in man. J. Clin. Invest. 1993, 91, 344-349. [CrossRef]
- Berkowitz, D.E.; Nardone, N.A.; Smiley, R.M.; Price, D.T.; Kreutter, D.K.; Fremeau, R.T.; Schwinn, D.A. Distribution of ß3-adrenoceptor mRNA in human tissues. Eur. J. Pharmacol. 1995, 289, 223-228.
- Granneman, J.G.; Lahners, K.N. Analysis of human and rodent ß3-adrenergic receptor messenger ribonucleic acids. Endocrinology 1994, 135, 1025-1031.
- Baskin, A.S.; Linderman, J.D.; Brychta, R.J.; McGehee, S.; Anflick-Chames, E.; Cero, C.; Johnson, J.W.; O’Mara, A.E.; Fletcher, L.A.; Leitner, B.P.; et al. Regulation of human adipose tissue activation, gallbladder size, and bile acid metabolism by a β3-adrenergic receptor agonist. Diabetes 2018, 67, 2113-2125. [CrossRef]
- Cero, C.; Lea, H.J.; Zhu, K.Y.; Shamsi, F.; Tseng, Y.-H.; Cypess, A.M. β3-Adrenergic receptors regulate human brown/beige adipocyte lipolysis and thermogenesis. JCI Insight 2021, 6. [CrossRef]
- Thomas, R.F.; Liggett, S.B. Lack of ß3-adrenergic receptor mRNA expression in adipose and other metabolic tissues in the adult human. Mol. Pharmacol. 1993, 43, 343-348.
- Deng, C.; Paoloni-Giacobino, A.; Kuehne, F.; Boss, O.; Revelli, J.P.; Moinat, M.; Cawthorne, M.A.; Muzzin, P.; Giacobino, J.P. Respective degree of expression of ß1-, ß2- and ß3-adrenoceptors in human brown and white adipose tissue. Br. J. Pharmacol. 1996, 118, 929-934.
- Michel, M.C.; Gravas, S. Safety and tolerability of ß3-adrenoceptor agonists in the treatment of overactive bladder syndrome - insight from transcriptosome and experimental studies. Expert Opin Drug Safety 2016, 15, 647-657. [CrossRef]
- Riis-Vestergaard, M.J.; Richelsen, B.; Bruun, J.M.; Li, W.; Hansen, J.B.; Pedersen, S.B. Beta-1 and not beta-3 adrenergic receptors may be the primary regulator of human brown adipocyte metabolism. The Journal of Clinical Endocrinology & Metabolism 2020, 105, e994-e1005.
- Harms, H.H.; Zaagsma, J.; de Vente, J. Differentiation of ß-adrenoceptors in right atrium, diaphragm and adipose tissue of the rat, using stereoisomers of propranolol, alprenolol, nifenalol and practolol. Life Sci. 1977, 21, 123-128.
- Tan, S.; Curtis-Prior, P.B. Characterization of the beta-adrenoceptor of the adipose cells of the rat. Int. J. Obes. 1983, 7, 409-414.
- Wilson, C.; Wilson, S.; Piercy, V.; Sennitt, M.V.; Arch, J.R.S. The rat lipolytic β-adrenoceptor: studies using novel β-adrenoceptor agonists. Eur. J. Pharmacol. 1984, 100, 309-319. [CrossRef]
- Hollenga, C.; Zaagsma, J. Direct evidence for the atypical nature of functional beta-adrenoceptors in rat adipocytes. Br. J. Pharmacol. 1989, 98, 1420-1424. [CrossRef]
- Hollenga, C.; Haas, M.; Deinum, J.T.; Zaagsma, J. Discrepancies in lipolytic activities induced by beta-adrenoceptor agonists in human and rat adipocytes. Horm. Metab. Res. 1990, 22, 17-21. [CrossRef]
- Murphy, G.J.; Kirkham, D.M.; Cawthorne, M.A.; Young, P. Correlation of beta 3-adrenoceptor-induced activation of cyclic AMP-dependent protein kinase with activation of lipolysis in rat white adipocytes. Biochem. Pharmacol. 1993, 46, 575-581. [CrossRef]
- van Liefde, I.; van Witzenburg, A.; Vauquelin, G. Multiple beta adrenergic receptor subclasses mediate the l-isoproterenol-induced lipolytic response in rat adipocytes. J. Pharmacol. Exp. Ther. 1992, 262, 552-558.
- Germack, R.; Starzec, A.B.; Vassy, R.; Perret, G.Y. ß-Adrenoceptor subtype expression and function in rat white adipocytes. Br. J. Pharmacol. 1997, 120, 201-210.
- Moreno-Aliaga, M.; Alfredo, M.; Stanhope, K.; Otero, M.P.; Havel, P. Effects of TrecadrineR, a beta 3-adrenergic agonist, on leptin secretion, glucose and lipid metabolism in isolated rat adipocytes. International journal of obesity and related metabolic disorders : journal of the International Association for the Study of Obesity 2002, 26, 912-919. [CrossRef]
- Hatakeyama, Y.; Sakata, Y.; Takakura, S.; Manda, T.; Mutoh, S. Acute and chronic effects of FR-149175, a ß3-adrenergic receptor agonist, on energy expenditure in Zucker fatty rats. Am. J. Physiol. 2004, 287, R336-R341. [CrossRef]
- Rosenbaum, M.; Malbon, C.C.; Hirsch, J.; Leibel, R.L. Lack of ß3-adrenergic effect on lipolysis in human subcutaneous adipose tissue. J. Clin. Endocrinol. Metab. 1993, 77, 352-355. [CrossRef]
- Vicario, P.P.; Candelore, M.R.; Schaeffer, M.-T.; Kelly, L.; Thompson, G.M.; Brady, E.J.; Saperstein, R.; MacIntyre, D.E.; Tota, L.M.; Cascieri, M.A. Desensitization of β3-adrenergic receptor- stimulated adenylyl cyclase activity and lipolysis in rats. Life Sci. 1998, 62, 627-638. [CrossRef]
- Hollenga, C.; Brouwer, F.; Zaagsma, J. Differences in functional cyclic AMP compartments mediating lipolysis by isoprenaline and BRL 37344 in four adipocyte types. Eur. J. Pharmacol. 1991, 200, 325-330. [CrossRef]
- Hollenga, C.; Brouwer, F.; Zaagsma, J. Relationship between lipolysis and cyclic AMP generation mediated by atypical beta-adrenoceptors in rat adipocytes. Br. J. Pharmacol. 1991, 102, 577-580. [CrossRef]
- Oriowo, M.A.; Chapman, H.; Kirkham, D.M.; Sennitt, M.V.; Ruffolo, R.R., Jr.; Cawthorne, M.A. The selectivity in vitro of the stereoisomers of the beta-3 adrenoceptor agonist BRL 37344. J. Pharmacol. Exp. Ther. 1996, 277, 22-27.
- Tavernier, G.; Jimenez, M.; Giacobino, J.P.; Hulo, N.; Lafontan, M.; Muzzin, P.; Langin, D. Norepinephrine induces lipolysis in ß1/ß2/ß3-adrenoceptor knockout mice. Mol. Pharmacol. 2005, 68, 793-799. [CrossRef]
- Tavernier, G.; Galitzky, J.; Bousquet-Melou, A.; Montastruc, J.L.; Berlan, M. The positive chronotropic effect induced by BRL 37344 and CGP 12177, two beta-3 adrenergic agonists, does not involve cardiac beta adrenoceptors but baroreflex mechanisms. J. Pharmacol. Exp. Ther. 1992, 263, 1083-1090.
- Shen, Y.T.; Cervoni, P.; Claus, T.; Vatner, S.F. Differences in ß3-adrenergic receptor cardiovascular regulation in conscious primates, rats and dogs. J. Pharmacol. Exp. Ther. 1996, 278, 1435-1443.
- Bousquet-Melou, A.; Galitzky, J.; Carpene, C.; Lafontan, M.; Berlan, M. ß-Adrenergic control of lipolysis in primate white fat cells: a comparative study with nonprimate mammals. Am. J. Physiol. 1994, 267, R115-R123. [CrossRef]
- Forrest, M.J.; Hom, G.; Bach, T.; Candelore, M.R.; Cascieri, M.A.; Strader, C.; Tota, L.; Fisher, M.H.; Szumiloski, J.; Ok, H.O.; et al. L-750355, a human ß3-adrenoceptor agonist; in vitro pharmacology and profile of activity in vivo in the rhesus monkey. Eur. J. Pharmacol. 2000, 407, 175-181. [CrossRef]
- Lipworth, B.J. Clinical pharmacology of ß3-adrenoceptors. Br. J. Clin. Pharmacol. 1996, 42, 291-300. [CrossRef]
- Hoffstedt, J.; Shimizu, M.; Sjöstedt, S.; Lönnqvist, F. Determination of ß3-adrenoceptor mediated lipolysis in human fat cells. Obes. Res. 1995, 3, 447-457. [CrossRef]
- Hoffstedt, J.; Lönnqvist, F.; Shimizu, M.; Blaak, E.; Arner, P. Effects of several putative beta3-adrenoceptor agonists on lipolysis in human omental adipocytes. Int. J. Obes. Relat. Metab. Disord. 1996, 20, 428-434.
- Sennitt, M.V.; Kaumann, A.J.; Molenaar, P.; Beeley, L.J.; Young, P.W.; Kelly, J.; Chapman, H.; Henson, S.M.; Berge, J.M.; Dean, D.K.; et al. The contribution of classical (ß1/2-) and atypical ß-adrenoceptors to the stimulation of white adipocyte lipolysis and right atrial appendage contraction by novel ß3-adreoceptor agonists of differing selectivities. J. Pharmacol. Exp. Ther. 1998, 285, 1084-1095.
- Umekawa, T.; Yoshida, T.; Sakane, N.; Kogure, A.; Kondo, M.; Honjyo, H. Trp64Arg mutation of beta3-adrenoceptor gene deteriorates lipolysis induced by beta3-adrenoceptor agonist in human omental adipocytes. Diabetes 1999, 48, 117-120.
- El-Yazbi, A.F.; Elrewiny, M.A.; Habib, H.M.; Eid, A.H.; Elzahhar, P.A.; Belal, A.S. Thermogenic modulation of adipose depots: A perspective on possible therapeutic intervention with early cardiorenal complications of metabolic impairment. Molecular Pharmacology 2023.
- Astrup, A.; Toubro, S.; Cannon, S.; Hein, P.; Breum, L.; Madsen, J. Caffeine: a double-blind, placebo-controlled study of its thermogenic, metabolic, and cardiovascular effects in healthy volunteers. The American journal of clinical nutrition 1990, 51, 759-767.
- Van Schaik, L.; Kettle, C.; Green, R.; Sievers, W.; Hale, M.; Irving, H.; Whelan, D.; Rathner, J. Stimulatory, but not anxiogenic, doses of caffeine act centrally to activate interscapular brown adipose tissue thermogenesis in anesthetized male rats. Scientific Reports 2021, 11, 113.
- De Matteis, R.; Arch, J.; Petroni, M.; Ferrari, D.; Cinti, S.; Stock, M. Immunohistochemical identification of the β3-adrenoceptor in intact human adipocytes and ventricular myocardium: effect of obesity and treatment with ephedrine and caffeine. International journal of obesity 2002, 26, 1442-1450.
- Meyers, D.S.; Skwish, S.; Dickinson, K.E.; Kienzle, B.; Arbeeny, C.M. β3-Adrenergic receptor-mediated lipolysis and oxygen consumption in brown adipocytes from cynomolgus monkeys. The Journal of Clinical Endocrinology & Metabolism 1997, 82, 395-401.
- Rohlfs, E.M.; Daniel, K.W.; Premont, R.T.; Kozak, L.P.; Collins, S. Regulation of the uncoupling protein gene (Ucp) by β1, β2, and β3-adrenergic receptor subtypes in immortalized brown adipose cell lines. Journal of Biological Chemistry 1995, 270, 10723-10732.
- Puigserver, P.; Pico, C.; Stock, M.; Palou, A. Effect of selective β-adrenoceptor stimulation on UCP synthesis in primary cultures of brown adipocytes. Molecular and cellular endocrinology 1996, 117, 7-16.
- Yamakawa, A.; Tanaka, E.; Nakano, S. Effect of the adrenergic beta 3-agonist, BRL37344, on heat production by brown adipocytes in obese and in older rats. The Tokai journal of experimental and clinical medicine 1994, 19, 139-142.
- Hao, L.; Scott, S.; Abbasi, M.; Zu, Y.; Khan, M.S.H.; Yang, Y.; Wu, D.; Zhao, L.; Wang, S. Beneficial metabolic effects of mirabegron in vitro and in high-fat diet-induced obese mice. Journal of Pharmacology and Experimental Therapeutics 2019, 369, 419-427.
- Choe, S.S.; Huh, J.Y.; Hwang, I.J.; Kim, J.I.; Kim, J.B. Adipose tissue remodeling: its role in energy metabolism and metabolic disorders. Frontiers in endocrinology 2016, 7, 30.
- Tanaka, M.; Itoh, M.; Ogawa, Y.; Suganami, T. Molecular mechanism of obesity-induced ‘metabolic’tissue remodeling. Journal of Diabetes Investigation 2018, 9, 256-261.
- Auger, C.; Kajimura, S. Adipose tissue remodeling in pathophysiology. Annual Review of Pathology: Mechanisms of Disease 2023, 18, 71-93.
- Lee, M.-J.; Wu, Y.; Fried, S.K. Adipose tissue remodeling in pathophysiology of obesity. Current opinion in clinical nutrition and metabolic care 2010, 13, 371.
- Lee, Y.-H.; Mottillo, E.P.; Granneman, J.G. Adipose tissue plasticity from WAT to BAT and in between. Biochimica et Biophysica Acta (BBA)-Molecular Basis of Disease 2014, 1842, 358-369.
- Willows, J.W.; Blaszkiewicz, M.; Townsend, K.L. The sympathetic innervation of adipose tissues: regulation, functions, and plasticity. Comprehensive Physiology 2011, 13, 4985-5021.
- Zhao, Y.; Li, X.; Yang, L.; Eckel-Mahan, K.; Tong, Q.; Gu, X.; Kolonin, M.G.; Sun, K. Transient overexpression of vascular endothelial growth factor A in adipose tissue promotes energy expenditure via activation of the sympathetic nervous system. Molecular and Cellular Biology 2018, 38, e00242-00218.
- Jimenez, M.; Léger, B.; Canola, K.; Lehr, L.; Arboit, P.; Seydoux, J.; Russell, A.P.; Giacobino, J.-P.; Muzzin, P.; Preitner, F. β1/β2/β3-adrenoceptor knockout mice are obese and cold-sensitive but have normal lipolytic responses to fasting. FEBS letters 2002, 530, 37-40.
- van Marken Lichtenbelt, W.D.; Vanhommerig, J.W.; Smulders, N.M.; Drossaerts, J.M.; Kemerink, G.J.; Bouvy, N.D.; Schrauwen, P.; Teule, G.J. Cold-activated brown adipose tissue in healthy men. New England Journal of Medicine 2009, 360, 1500-1508.
- Finlin, B.S.; Memetimin, H.; Confides, A.L.; Kasza, I.; Zhu, B.; Vekaria, H.J.; Harfmann, B.; Jones, K.A.; Johnson, Z.R.; Westgate, P.M. Human adipose beiging in response to cold and mirabegron. JCI insight 2018, 3.
- Preite, N.Z.; do Nascimento, B.P.; Muller, C.R.; Américo, A.L.V.; Higa, T.S.; Evangelista, F.S.; Lancellotti, C.L.; dos Santos Henriques, F.; Batista Jr, M.L.; Bianco, A.C. Disruption of beta3 adrenergic receptor increases susceptibility to DIO in mouse. The Journal of endocrinology 2016, 231, 259.
- Granneman, J.; Burnazi, M.; Zhu, Z.; Schwamb, L. White adipose tissue contributes to UCP1-independent thermogenesis. American Journal of Physiology-Endocrinology and Metabolism 2003, 285, E1230-E1236.
- Warner, A.; Kjellstedt, A.; Carreras, A.; Böttcher, G.; Peng, X.-R.; Seale, P.; Oakes, N.; Lindén, D. Activation of β3-adrenoceptors increases in vivo free fatty acid uptake and utilization in brown but not white fat depots in high-fat-fed rats. American Journal of Physiology-Endocrinology and Metabolism 2016, 311, E901-E910.
- Van Schaik, L.; Kettle, C.; Green, R.; Irving, H.R.; Rathner, J.A. Effects of caffeine on brown adipose tissue thermogenesis and metabolic homeostasis: a review. Frontiers in neuroscience 2021, 15, 54.
- de Souza, C.J.; Burkey, B.F. Beta3-adrenoceptor agonists as anti-diabetic and anti-obesity drugs in humans. Current pharmaceutical design 2001, 7, 1433-1449.
- Arch, J.; Wilson, S. Prospects for beta 3-adrenoceptor agonists in the treatment of obesity and diabetes. International Journal of Obesity and Related Metabolic Disorders: Journal of the International Association for the Study of Obesity 1996, 20, 191-199.
- Liu, X.; Pérusse, F.; Bukowiecki, L.J. Mechanisms of the antidiabetic effects of the β3-adrenergic agonist CL-316243 in obese Zucker-ZDF rats. American Journal of Physiology-Regulatory, Integrative and Comparative Physiology 1998, 274, R1212-R1219.
- Himms-Hagen, J.; Cui, J.; Danforth Jr, E.; Taatjes, D.; Lang, S.; Waters, B.; Claus, T. Effect of CL-316,243, a thermogenic beta 3-agonist, on energy balance and brown and white adipose tissues in rats. American Journal of Physiology-Regulatory, Integrative and Comparative Physiology 1994, 266, R1371-R1382.
- Arbeeny, C.M.; Meyers, D.S.; Hillyer, D.E.; Bergquist, K.E. Metabolic alterations associated with the antidiabetic effect of beta 3-adrenergic receptor agonists in obese mice. American Journal of Physiology-Endocrinology and Metabolism 1995, 268, E678-E684.
- Hamann, A.; Flier, J.S.; Lowell, B.B. Decreased brown fat markedly enhances susceptibility to diet-induced obesity, diabetes, and hyperlipidemia. Endocrinology 1996, 137, 21-29.
- Lowell, B.B.; S-Susulic, V.; Hamann, A.; Lawitts, J.A.; Himms-Hagen, J.; Boyer, B.B.; Kozak, L.P.; Flier, J.S. Development of obesity in transgenic mice after genetic ablation of brown adipose tissue. Nature 1993, 366, 740-742.
- Viguerie-Bascands, N.; Bousquet-Mélou, A.; Galitzky, J.; Larrouy, D.; Ricquier, D.; Berlan, M.; Casteilla, L. Evidence for numerous brown adipocytes lacking functional beta 3-adrenoceptors in fat pads from nonhuman primates. The Journal of Clinical Endocrinology & Metabolism 1996, 81, 368-375.
- Vijgen, G.H.; Bouvy, N.D.; Teule, G.J.; Brans, B.; Schrauwen, P.; van Marken Lichtenbelt, W.D. Brown adipose tissue in morbidly obese subjects. PloS one 2011, 6, e17247.
- Chondronikola, M.; Volpi, E.; Børsheim, E.; Porter, C.; Annamalai, P.; Enerbäck, S.; Lidell, M.E.; Saraf, M.K.; Labbe, S.M.; Hurren, N.M. Brown adipose tissue improves whole-body glucose homeostasis and insulin sensitivity in humans. Diabetes 2014, 63, 4089-4099.
- Astrup, A.; Bülow, J.; Christensen, N.; Madsen, J. Ephedrine-induced thermogenesis in man: no role for interscapular brown adipose tissue. Clinical science (London, England: 1979) 1984, 66, 179-186.
- Widen, E.; Lehto, M.; Kanninen, T.; Walston, J.; Shuldiner, A.R.; Groop, L.C. Association of a polymorphism in the β3-adrenergic–receptor gene with features of the insulin resistance syndrome in Finns. New England Journal of Medicine 1995, 333, 348-352.
- Clément, K.; Vaisse, C.; Manning, B.S.J.; Basdevant, A.; Guy-Grand, B.; Ruiz, J.; Silver, K.D.; Shuldiner, A.R.; Froguel, P.; Strosberg, A.D. Genetic variation in the β3-adrenergic receptor and an increased capacity to gain weight in patients with morbid obesity. New England Journal of Medicine 1995, 333, 352-354.
- Arner, P.; Hoffstedt, J. Adrenoceptor genes in human obesity. Journal of Internal Medicine 1999, 245, 667-672.
- Malik, S.G.; Saraswati, M.R.; Suastika, K.; Trimarsanto, H.; Oktavianthi, S.; Sudoyo, H. Association of beta3-adrenergic receptor (ADRB3) Trp64Arg gene polymorphism with obesity and metabolic syndrome in the Balinese: a pilot study. BMC research notes 2011, 4, 1-7.
- Li, L.; Lnnqvist, F.; Luthman, H.; Arner, P. Phenotypic characterization of the Trp64Arg polymorphism in the beta3-adrenergic receptor gene in normal weight and obese subjects. Diabetologia 1996, 7, 857-860.
- Gagnon, J.; Mauriège, P.; Roy, S.; Sjöström, D.; Chagnon, Y.C.; Dionne, F.T.; Oppert, J.-M.; Pérusse, L.; Sjöström, L.; Bouchard, C. The Trp64Arg mutation of the beta3 adrenergic receptor gene has no effect on obesity phenotypes in the Québec Family Study and Swedish Obese Subjects cohorts. The Journal of clinical investigation 1996, 98, 2086-2093.
- Oksanen, L.; Mustajoki, P.; Kaprio, J.; Kainulainen, K.; Jänne, O.; Peltonen, L.; Kontula, K. Polymorphism of the beta 3-adrenergic receptor gene in morbid obesity. International Journal of Obesity and Related Metabolic Disorders: Journal of the International Association for the Study of Obesity 1996, 20, 1055-1061.
- Larson, C.J. Translational Pharmacology and Physiology of Brown Adipose Tissue in Human Disease and Treatment. Handb. Exp. Pharmacol. 2019, 251, 381-424. [CrossRef]
- Connacher, A.A.; Bennet, W.M.; Jung, R.T.; Rennie, M.J. Metabolic effects of three weeks administration of the beta-adrenoceptor agonist BRL 26830A. Int. J. Obes. Relat. Metab. Disord. 1992, 16, 685-694.
- Connacher, A.A.; Lakie, M.; Powers, N.; Elton, R.A.; Walsh, E.G.; Jung, R.T. Tremor and the anti-obesity drug BRL 26830A. Br. J. Clin. Pharmacol. 1990, 30, 613-615. [CrossRef]
- Wheeldon, N.M.; McDevitt, D.G.; Lipworth, B.J. Do beta 3-adrenoceptors mediate metabolic responses to isoprenaline. Q. J. Med. 1993, 86, 595-600.
- Cawthorne, M.A.; Sennitt, M.V.; Arch, J.R.; Smith, S.A. BRL 35135, a potent and selective atypical beta-adrenoceptor agonist. Am. J. Clin. Nutr. 1992, 55, 252s-257s. [CrossRef]
- Mitchell, T.H.; Ellis, R.D.; Smith, S.A.; Robb, G.; Cawthorne, M.A. Effects of BRL 35135, a beta-adrenoceptor agonist with novel selectivity, on glucose tolerance and insulin sensitivity in obese subjects. Int. J. Obes. 1989, 13, 757-766.
- Dow, R.L. β3-Adrenergic agonists: potential therapeutics for obesity. Expert Opinion on Investigational Drugs 1997, 6, 1811-1825. [CrossRef]
- Riis-Vestergaard, M.J.; Richelsen, B.; Bruun, J.M.; Li, W.; Hansen, J.B.; Pedersen, S.B. Beta-1 and not beta-3 adrenergic receptors may be the primary regulator of human brown adipocyte metabolism. J. Clin. Endocrinol. Metab. 2020, 105, e994-e1005. [CrossRef]
- Weyer, C.; Tataranni, P.A.; Snitker, S.; Danforth, E., Jr.; Ravussin, E. Increase in insulin action and fat oxidation after treatment with CL 316,243, a highly selective beta3-adrenoceptor agonist in humans. Diabetes 1998, 47, 1555-1561. [CrossRef]
- Muzzin, P.; Revelli, J.P.; Ricquier, D.; Meier, M.K.; Assimacopoulos-Jeannet, F.; Giacobino, J.P. The novel thermogenic ß-adrenergic agonist Ro 16-8714 increases the interscapular brown-fat ß-receptor-adenylate cyclase and the uncoupling-protein mRNA level in obese (fa/fa) Zucker rats. Biochem. J. 1989, 261, 721-724. [CrossRef]
- Henny, C.; Schutz, Y.; Buckert, A.; Meylan, M.; Jequier, E.; Felber, J.P. Thermogenic effect of the new beta-adrenoreceptor agonist Ro 16-8714 in healthy male volunteers. Int. J. Obes. 1987, 11, 473-483.
- Jéquier, E.; Munger, R.; Felber, J.P. Thermogenic effects of various beta-adrenoceptor agonists in humans: their potential usefulness in the treatment of obesity. Am. J. Clin. Nutr. 1992, 55, 249s-251s. [CrossRef]
- Toubro, S.; Astrup, A.; Hardmann, M. A double-blind randomized 14 day trials of the effect of the ß-3 agonist ICI D-7114 on 24 h energy expenditure and substract oxidation in adipose patients. Int. J. Obes. 1993, 17, S73.
- Kullmann, F.A.; Limberg, B.J.; Artim, D.E.; Shah, M.; Downs, T.R.; Contract, D.; Wos, J.; Rosenbaum, J.S.; De Groat, W.C. Effects of ß3-adrenergic receptor activation on rat urinary bladder hyperactivity induced by ovariectomy. J. Pharmacol. Exp. Ther. 2009, 330, 704-717. [CrossRef]
- Redman, L.M.; de Jonge, L.; Fang, X.; Gamlin, B.; Recker, D.; Greenway, F.L.; Smith, S.R.; Ravussin, E. Lack of an effect of a novel ß3-adrenoceptor agonist, TAK-677, on energy metabolism in obese individuals: a double-blind, placebo-controlled randomized study. J. Clin. Endocrinol. Metab. 2007, 92, 527-531. [CrossRef]
- van Baak, M.A.; Hul, G.B.J.; Toubro, S.; Astrup, A.; Gottesdiener, K.M.; DeSmet, M.; Saris, W.H.M. Acute effect of L-796568, a novel ß3-adrenergic receptor agonist, on energy expenditure in obese men. Clin. Pharmacol. Ther. 2002, 71, 272-279. [CrossRef]
- Larsen, T.M.; Toubro, S.; van Baak, M.A.; Larson, P.; Saris, W.H.; Astrup, A. Effect of a 28-d treatment with L-796568, a novel ß3-adrenergic receptor agonist, on energy expenditure and body composition in obese men. Am. J. Clin. Nutr. 2002, 76, 780-788.
- Michel, M.C.; Korstanje, C. ß3-Adrenoceptor agonists for overactive bladder syndrome: role of translational pharmacology in a re-positioning drug development project. Pharmacol. Ther. 2016, 159, 66-82. [CrossRef]
- Chapple, C.R.; Cardozo, L.; Nitti, V.W.; Siddiqui, E.; Michel, M.C. Mirabegron in overactive bladder: a review of efficacy, safety, and tolerability. Neurourol. Urodyn. 2014, 33, 17-30. [CrossRef]
- Cypess, A.M.; Weiner, L.S.; Roberts-Toler, C.; Franquet, E.; Kessler, S.H.; Kahn, P.A.; English, J.; Chatman, K.; Trauger, S.A.; Doria, A.; Kolodny, G.M. Activation of human brown adipose tissue by a ß3-adrenergic receptor agonist. Cell Metab. 2015, 21, 33-38. [CrossRef]
- Kennelly, M.J.; Rhodes, T.; Girman, C.J.; Thomas, E.; Shortino, D.; Mudd, P.N., Jr. Efficacy of vibegron and mirabegron for overactive bladder: a systematic literature review and indirect treatment comparison. Adv. Ther. 2021, 38, 5452-5464. [CrossRef]
- Okeke, K.; Angers, S.; Bouvier, M.; Michel, M.C. Agonist-induced desensitisation of β3-adrenoceptors: where, when and how? Br. J. Pharmacol. 2019, 176, 2539-2558. [CrossRef]
- Mo, W.; Michel, M.C.; Lee, X.W.; Kaumann, A.J.; Molenaar, P. The β3-adrenoceptor agonist mirabegron increases human atrial force through β1-adrenoceptors: an indirect mechanism? Br. J. Pharmacol. 2017, 174, 2706-2715. [CrossRef]
- Alexandre, E.C.; Kiguti, L.R.; Calmasini, F.B.; Silva, F.H.; da Silva, K.P.; Ferreira, R.; Ribeiro, C.A.; Monica, F.Z.; Pupo, A.S.; Antunes, E. Mirabegron relaxes urethral smooth muscle by a dual mechanism involving ß3-adrenoceptor activation and α1-adrenoceptor blockade. Br. J. Pharmacol. 2016, 173, 415-428. [CrossRef]
- Huang, R.; Liu, Y.; Ciotkowska, A.; Tamalunas, A.; Waidelich, R.; Strittmatter, F.; Stief, C.G.; Hennenberg, M. Concentration-dependent alpha1-adrenoceptor antagonism and inhibition of neurogenic smooth muscle contraction by mirabegron in the human prostate. Front. Pharmacol. 2021, 12. [CrossRef]
- Michel, M.C. α1-Adrenoceptor activity of β-adrenoceptor ligands – An expected drug property with limited clinical relevance. Eur. J. Pharmacol. 2020, 889, 173632. [CrossRef]
- Perrone, M.G.; Scilimati, A. β3-Adrenoceptor ligand development history through patent review. Expert Opin. Ther. Pat. 2011, 21, 505-536. [CrossRef]
- Grazia Perrone, M.; Scilimati, A. β3-Adrenoceptor agonists and (antagonists as) inverse agonists: history, perspective, constitutive activity, and stereospecific binding. In Methods Enzymol., Conn, P.M., Ed.; Academic Press: 2010; Volume 484, pp. 197-230.
Figure 1.
Differential metabolic impact of β3-adrenoceptor (AR) agonists across species. In rodents, β3-AR agonists, such as mirabegron, CL 316,243 and BRL 37,344, were found to promote positive metabolic implications by improving insulin release (indicated by the green plus sign), increasing glucose uptake in brown adipose tissue (BAT) and white adipose tissue (WAT) and lipolysis in the former only. In addition to upregulating uncoupling protein 1 (UCP1) expression and thermogenic capacity in BAT. On the other hand, data from human studies concerning insulin release, glucose uptake and lipolysis by WAT were inconsistent and inconclusive (indicated by the red question mark). Only glucose uptake human BAT was modulated consistently by β3-AR agonists. However, the data on β3-AR induced thermogenesis and UCP1 expression in human BAT were equivocal..
Figure 1.
Differential metabolic impact of β3-adrenoceptor (AR) agonists across species. In rodents, β3-AR agonists, such as mirabegron, CL 316,243 and BRL 37,344, were found to promote positive metabolic implications by improving insulin release (indicated by the green plus sign), increasing glucose uptake in brown adipose tissue (BAT) and white adipose tissue (WAT) and lipolysis in the former only. In addition to upregulating uncoupling protein 1 (UCP1) expression and thermogenic capacity in BAT. On the other hand, data from human studies concerning insulin release, glucose uptake and lipolysis by WAT were inconsistent and inconclusive (indicated by the red question mark). Only glucose uptake human BAT was modulated consistently by β3-AR agonists. However, the data on β3-AR induced thermogenesis and UCP1 expression in human BAT were equivocal..
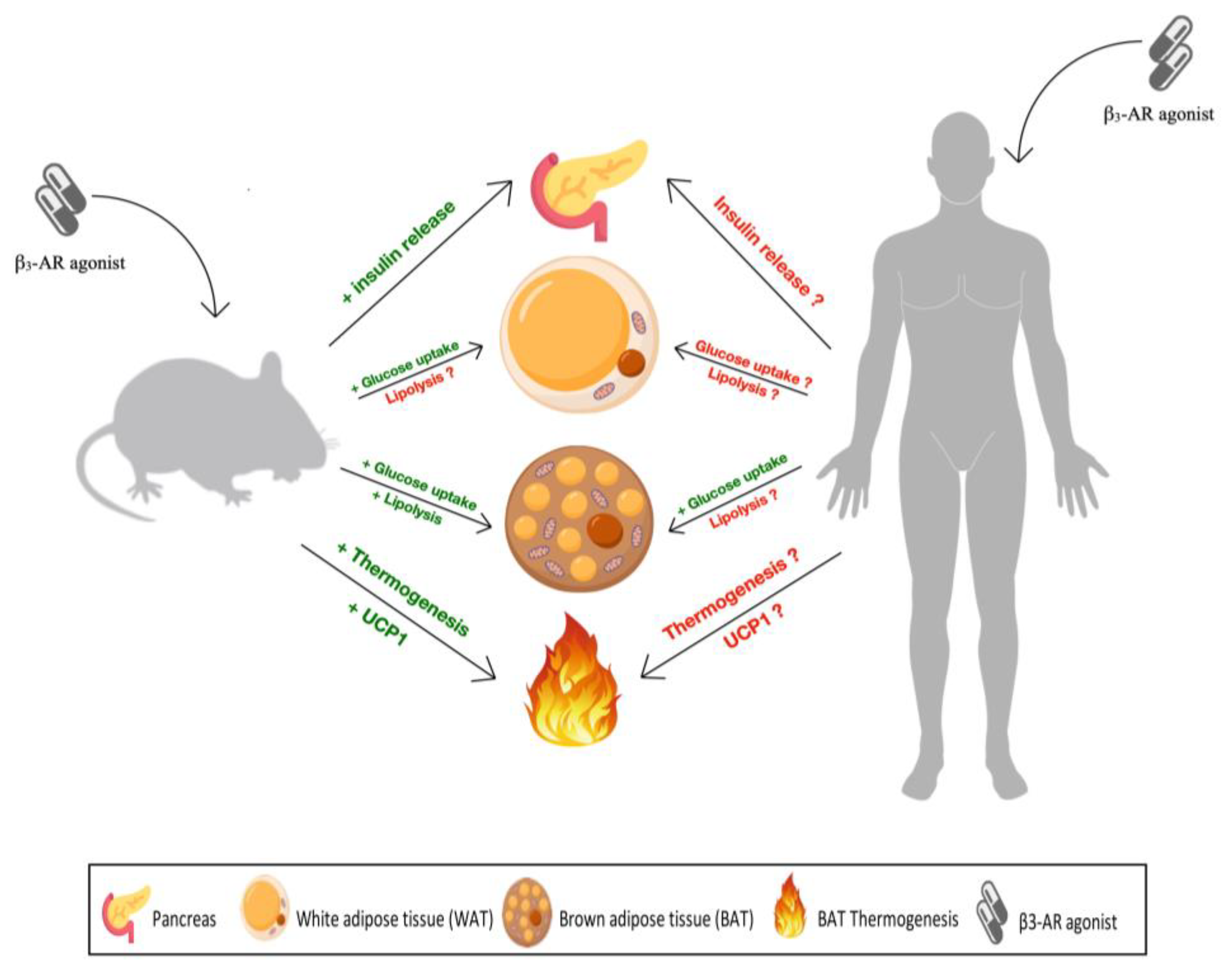

Table 1.
Quantification of β-AR subtype mRNA expression in human tissues. All data are shown as fragments per kilobase of transcript length per million mapped reads, a transcript abundance unit, in descending order of β3-AR expression and represent the median of samples from 3-7 patients.
Table 1.
Quantification of β-AR subtype mRNA expression in human tissues. All data are shown as fragments per kilobase of transcript length per million mapped reads, a transcript abundance unit, in descending order of β3-AR expression and represent the median of samples from 3-7 patients.
| β1-AR | β2-AR | β3-AR | |
|---|---|---|---|
| Ovary | 0.02 | 0.79 | 6.89 |
| Gall bladder | 0.10 | 4.43 | 2.57 |
| Placenta | 28.77 | 5.99 | 2.53 |
| Urinary bladder | 0.29 | 6.63 | 1.54 |
| Fallopian tube | 0.09 | 4.38 | 0.64 |
| Colon | 0.75 | 2.07 | 0.54 |
| Appendix | 0.38 | 2.34 | 0.41 |
| Prostate | 4.01 | 9.64 | 0.29 |
| Small intestine | 1.18 | 1.66 | 0.25 |
| Endometrium | 0.07 | 2.20 | 0.22 |
| Adipose tissue | 2.29 | 12.60 | 0.19 |
| Duodenum | 0.73 | 1.54 | 0.19 |
| Rectum | 0.87 | 2.63 | 0.16 |
| Brain | 4.58 | 2.02 | 0.12 |
| Myometrium | 0.13 | 3.62 | 0.12 |
| Stomach | 0.76 | 6.68 | 0.11 |
| Lung | 6.55 | 18.01 | 0.07 |
| Lymph nodes | 0.11 | 3.08 | 0.07 |
| Esophagus | 0.99 | 8.55 | 0.03 |
| Skin | 0.16 | 5.60 | 0.02 |
| Tonsil | 0.31 | 5.51 | 0.02 |
| Heart | 11.57 | 4.92 | 0.02 |
| Bone marrow | 0.32 | 7.82 | 0 |
| Spleen | 0.90 | 6.90 | 0 |
| Skeletal muscle | 0.06 | 4.19 | 0 |
| Liver | 1.03 | 4.12 | 0 |
| Salivary gland | 4.93 | 2.54 | 0 |
| Adrenal | 0.16 | 1.48 | 0 |
| Thyroid | 0.21 | 1.07 | 0 |
| Kidney | 0.99 | 0.68 | 0 |
| Pancreas | 0.65 | 0.58 | 0 |
| Testis | 0.26 | 0.52 | 0 |
1 Adapted with permission from [112].
Table 2.
Comparison of various characteristics of rodent (rat, mouse) vs. human and other primate species. For details see main text.
Table 2.
Comparison of various characteristics of rodent (rat, mouse) vs. human and other primate species. For details see main text.
| rodent | humans and other primates | |
|---|---|---|
| BAT presence in adults | abundant | sparse |
| Insulin release by β3-AR agonists | +++ | - |
| β3-AR expression in AT | +++ | + |
| Glucose uptake in BAT | +++ | + |
| Lipolysis/thermogenesis | +++ | + |
| Lipolysis in WAT | β3-AR | β1-AR |
| Weight loss | ++ | inconclusive |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



