
Submitted:
10 October 2023
Posted:
12 October 2023
You are already at the latest version
Abstract
Whiteflies of Aleurocanthus spiniferus and A. woglumi (Hemiptera: Aleyrodidae) are serious pests of citrus and other important fruit crops. The problem in citrus has initiated the successful introductions of several natural enemies for biocontrol programs in Hawaii and many other countries. Here we summarized the history of infestation and biocontrol efforts of the two whiteflies in Hawaii for possible importation into Greece. Two Platygasteridae (Amitus hesperidum Silvestri, A. spiniferus (Brethes) and three Aphelinidae (Encarsia clypealis (Silvestri), E. smithi (Silvestri), E. perplexa Huang and Polaszek) were released in Hawaii for biocontrol of the citrus whiteflies during the period 1974-1999. The aphelinid, Cales noacki Howard, purposely released for Aleurothrixus flococcus (Maskell) in 1982 was also reported to attack other whiteflies including Aleurocanthus species on citrus. An additional aphelinid parasitoid, Encarsia nipponica Silvestri, native to Japan and China, was accidently introduced and found to attack both citrus whiteflies on the islands. Since the colonization of introduced parasitoids in infested fields on four Hawaiian Islands, no survey was done to evaluate their potential impact. We made two short surveys during September-November 2022 on Kauai, Hawaii, and Oahu Islands to introduce the dominant parasitoids to Greece for the biocontrol of A. spiniferus. Results showed that the infestation level was very low on Kauai, Hawaii, and Oahu Islands with mean infestation level ranged 1.4 - 3.1 on Hawaii and Oahu Islands mostly on pummelo and sweet orange, with no detection on Kauai Island. Only was E. perplexa the dominant parasitoid with parasitism rates ranged 0 -28 % on Hawaii Island and 11-65% on Oahu Island. Starter colony of the parasitoids has been colonized in Greece for evaluation and approval for release. This was the first field survey on Hawaii since the introduction and release of citrus whitefly natural enemies.
Keywords:
Hymenoptera
; natural enemies
; citrus
; citrus blackfly
; orange spiny whitefly
; Encarsia perplexa
; Amitus hesperidum
; Aleurocanthus woglumi
1. Introduction
The orange spiny whitefly, Aleurocanthus spiniferus (Quaintance) and citrus blackfly, A. woglumi Ashby (Hemiptera: Aleyrodidae) are two of the most important and hazardous citrus pests [1,2]. Both species are regulated as quarantine pests for European Union [3]. Aleurocanthus woglumi has not invaded Europe yet. However, climatic modelling predicts that areas in the Mediterranean basin are suitable for establishment [4]. Aleurocanthus spiniferus has already invaded Europe with the first record in Italy in 2008 [5]. Since then, its distribution in the European Union has expanded, including Albania [6], Croatia [7], Greece ([1], and Montenegro [8]. Both species cause similar symptoms to the infested host plants. In Hawaii during heavily infestation in 1998 infested trees were not fruiting with a range of 50 - >600 citrus blackfly pupae per infested leaf ([2], Ramadan, HDOA unpublished record). Main hosts are citrus species, but they are quite polyphagous with a wide host range that include mangoes and coffee [9]. Direct damage is caused by nymphs that suck the sap of the leaves causing lack of nutrients and water. Additionally, nymphs excrete honeydew drops that fall on the leaves where the growth of sooty mold is promoted which eventually covers the whole upper surface of the infested leaves. Both A. spiniferus and A. woglumi originate from Southeastern Asia [4,10]. In their native area they are not considered major pests, however, during the 20th century both species have invaded other parts of the world causing substantial damage, mainly on citrus crops [2,3,4]. Following their accidental introduction to new areas, biological control programs were initiated to mitigate the damage caused by the outbreaks of the two whiteflies [2,10,11,12,13,14].
In early seventies, A. spiniferus was introduced in Hawaii on rose foliage in Honolulu, Oahu Island in 1974. Subsequent surveys discovered it on navel orange, lime, tangerine, and pear, but infestations were reportedly low [15]. Two decades later, A. woglumi was also found being introduced on main Hawaiian Islands [15,16]. In both cases, the introduction of citrus whitefly species followed an introduction of biological control agents to compact their population outbreaks. Specifically, the parasitoid Encarsia smithi Silvestri and E. clypealis (Silvestri) (Hymenoptera: Aphelinidae) were introduced in the islands of Oahu and Hawaii [17] to control A. spiniferus and E. perplexa Huang & Polaszek (Hymenoptera, Aphelinidae) (misidentified at that time as Encarsia opulenta Silvestri) and Amitus hesperidum Silvestri (Hymenoptera: Platygastridae) were introduced to control A. woglumi [16], Table 1. Encarsia smithi was introduced in the island of Oahu, from Japan in 1974 and among three other Aphelinid species which were found to naturally parasitize A. spiniferus in Oahu. Encarsia smithi was the most important species, leading to the sufficient reduction of A. spiniferus population [17]. It was also reported on A. woglumi (Table 1). In 1998 exploratory investigations were carried out in Central America, in Guatemala, and two parasitoid species, E. perplexa and A. hesperidum were introduced to Hawaii in 1998. The wasps were mass reared on Oahu and released on other islands (Table 1). The two wasp species managed to disperse naturally and successfully controlled the population of A. woglumi in all releasing sites, Table 1 [18]. Another parasitoid, E. nipponica Silvestri, native to Japan and China, was discovered accidently as a parasitoid of both whiteflies with low parasitism ranged 0-11.5% (n= 103 infested leaves, Ramadan unpublished data). Similarly, the aphelind parasitoid, Cales noacki Howard, was released in 1981 for biological control of Aleurothrixus floccosus (Maskell), subsequently reported to attack Aleurocanthus species on citrus ([19], HDOA records). No field evaluation since the introduction and release of all the parasitoids but citrus growers on the islands were content, and less chemical control was used in their fields [2].
The recent invasion of A. spiniferus in the European Union and in Greece in particular, has caused the initiation of a new biological control program against this pest by introducing exotic natural enemies. Moreover, since the introduction of these parasitoids in Hawaii, no surveys have been conducted to determine the status of A. woglumi or A. spiniferus in the islands and about the presence and the abundance of their introduced parasitoids. Thus, an exploratory investigation on the Hawaiian Islands was performed to investigate the current parasitism rate of A. woglumi and to identify the parasitoid assembly species which were present in the populations of the pest. The results of those short surveys will give crucial information to examine the possibility of a successful introduction of those parasitoids either in Greece or in any other area where those serious pests have been established.
2. Materials and Methods
2.1. Collection and emergence of parasitoids.
Two surveys were performed in September and November 2022 on the islands of Hawaii, Kauai, and Oahu (Figure 1). A total of 62 sites located on the three Hawaiian Islands were surveyed. Host plants (mainly citrus trees) were macroscopically surveyed for citrus whitefly infestation and infested leaves were collected. Citrus whiteflies are very rare to be found in Hawaii in recent years. Sites were selected as orchards of citrus, trees in agriculture experimental stations, and local residential homes were examined by permission of landlords. Sites were selected randomly for the survey team to enter property, everywhere on the visited islands. Based on the presence of citrus trees, the survey team searched the trees for infestations and obtained permission to clip infested branches or leaves. Leaves with mature whitefly were picked, not those with only eggs or small nymphs that will not produce parasitoids.
The infested leaves were placed in paper envelopes which were sealed. (AJM paper grocery, lunch bags, USA, of different sizes as needed for holding infested leaves). Plastic bags are not suitable for life insects, https://www.gofoodservice.com/brand/ajm.
The leaves that were collected were picked according to the stage of nymphs of the citrus blackfly. Leaves that carried all the stages and older nymphs were preferred. Leaves with mature nymphs (3rd and 4th nymphs) were preferred for collection since these are the stages contain developed parasitoids. Leaves with eggs - 2nd nymphs were not collected since parasitoids from such leaves may not develop in the holding containers.
At the end of each collection day, all envelopes with leaf samples were transported to the insectary of the Hawaii Department of Agriculture, Plant Pest Control Branch in Honolulu. In the insectary, the leaves were carefully removed from the envelopes and placed in 70 mesh clear vinyl nylon screen collapsible lightweight aluminum cages (30x30x60 cm) appropriate for parasitic wasps’ emergence. Honey drops were placed in the inner surface of the mesh as food for adult wasps. We used smeared drops of SUE BEE® SPUN® honey on the inside cage sides and top for adult parasitoid feeding as indicated by HDOA insect rearing (https://siouxhoney.com/sue-bee-spun-honey/). Emerged adult parasitoids were aspirated and collected in falcon vials with honey drops (Falcon 50mL Conical Centrifuge Tubes; Fisher Scientific: A Thermo Fisher Scientific Brand (https://www.fishersci.com/shop/products/falcon-50ml-conical-centrifuge-tubes).
Emerged parasitoids and remaining infested leaves were shipped to Greece and placed in the containment facility in the biosecurity greenhouse at the Scientific Directorate of Entomology and Agricultural Zoology, in Benaki Phytopathological Institute (38°04'52.0"N 23°48'47.9"E). Emerged parasitoids were placed inside mesh cages with bitter orange, Citrus aurantium L. (Rutaceae), seedlings infested by A. spiniferus in the biosecurity greenhouse under controlled environmental conditions of 25°C ± 2°C, R.H. 60-70%, and natural day light photoperiod. Citrus aurantium seedlings were in mesh cages (45x45x45cm) which were put inside of larger ones (60x60x60 cm, 70 mesh) polyester white breathable fabric used to cover cages for rearing delicate encyrtid size parasitoids.
2.2. Identification of the parasitoids.
The identification of emerged parasitoids was based on morphological examination and molecular analyses. Morphological identification was performed according to appropriate keys and illustrations [20,21,22,23]. For distinguishing Amitus heperidum: The female is shiny black (0.75 mm long). The female's antenna is ten-segmented, with the last three segments forming a club. The male is like the female, with a filiform ten-segmented antenna and curved scape. A lateral plate-like process on the male fourth antennal segment is characteristic of the species, Figure 2 D, [20,21,23]. For identification of Encarsia perplexa, the mid lobe of mesoscutum dark, T1 and T2 of gaster largely pale (Figure 2 C). Male head is like that of female. Mesosoma orange yellow except pronotum, anterior half of mid lobe of mesoscutum, to propodeum and petiole are dark brown. Gaster brown to dark (Figure 2 C). Other characters for differentiation of closely related species are explained in [22]. The specimens were prepared for slide mounting as described as in slide preparation of chalcidoids by Noyes [24,25,26].
The insects that were destined for molecular analysis were stored in 1.5 mL microtubes with snap cap RNase & DNase free ClearLine®, filled with ethanol 98% (Analytical grade, Fisher Scientific). The specimens were stored and coded separately. In total, DNA was extracted from 45 parasitoids by using the DNeasy Blood & Tissue Kit (QIAGEN) according to manufacturer’s protocol. At the beginning of the procedure, every single insect was left on a filter paper until the ethanol was completely removed and the final DNA volume reached 20μL. Two sets of primers were used for species determination during the Polymerase Chain Reaction, targeting the genes of Cytochrome Oxidase I (COI) and 28S ribosomal RNA, which can investigate further the species’ determination [27,28]. For the amplification of the Barcoding gene, the primers LCO-1490 (5’-GGTCAACAAATCATAAAGATATTGG-3’) and HCO-2198 (5’-TAAACTTCAGGGTGACCAAAAAATCA-3’) were used [29], while the set of 28S-D2-F (5’-AGAGAGAGTTCAAGAGTACGTG-3’) and 28S-D2-R (5’-TTGGTCCGTGTTTCAAGACGGG-3’) was used for the amplification of the 28S gene [30].
Each PCR reaction mixture for both primers set contained 5µL of 10×PCR buffer, 1.5µL of MgCl2 (50 mM), 0.5µL of dNTPs (10 mM), 1µL of each primer (10µM), 5µL of template DNA (20-40ng), 0.5μL of the thermostable Taq DNA polymerase (Platinum, Invitrogen) and molecular grade water (up to 50µL). The thermocycling program included an initial denaturation step of 3 min at 94°C, followed by 35 cycles of 94°C for 30 sec, 51°C (LCO-HCO) or 58°C (28S) for 45 sec and 72°C for 1 min and a final step of extension at 72°C for 5 min.
The two template’s amplification was confirmed separately by using 5μL of the PCR products on 1.2% agarose gel electrophoresis, that finally resulted in the observation of an expected length product of 658bp and 550bp, correspondingly. The rest of the volume of 45μL was purified according to the supplier’s instructions of the NucleoFast 96 PCR Clean-up kit (Macherey-Nagel GmbH & Co. KG) and then forwarded to Macrogen Europe (Netherlands) for automated sequencing analysis. The obtained sequencing results were optimized, generated, and aligned through the software Geneious Prime 2023.0.1 (https://www.geneious.com/). The produced sequences were checked for their authenticity at genus or species level according to the BLAST public interface of the National Center for Biotechnology Information (NCBI - https://blast.ncbi.nlm.nih.gov).
2.3. Infestation and Parasitism Rate.
The parasitism rate of Amitus sp. and Encarsia sp. to A. woglumi from the collected leaves was estimated from the exit holes on the body of the nymphs. The exit hole of A. woglumi adults differs from the exit hole of the parasitoids by the shape of exit holes on the shells. The holes due to the exit of parasitoids are circular and placed at the back of the nymph, while those from the exit hole of A. woglumi adults is a T-shaped slit (Figure 2 A, E). To determine parasitism rate, all nymphs on each leaf were counted, under a Nikon SMZ-745 stereomicroscope in the laboratory. The nymphs were listed as “parasitized” (if the shells had parasitoid circular exit holes as in Figure 2 E). All eclosed parasitoids were Encarsia perplexa) and “not parasitized” (if the shells had the T-shape exit slit of the A. woglumi as in Figure 2 A), while broken shells were excluded from the results. Finally, the leaf samples were frozen at -20°C for 72 hours before discarded.
The level of infestation was determined by the population size of the citrus blackfly, which was categorized depending on the total number of nymphs of all developmental stages, per leaf and the total number of leaves collected per location. The infestation was scored as follows: 1 = 1 to 10 nymphs, 2 = 11 to 30 nymphs, 3= 31 to 99 nymphs, 4 = more than 100 nymphs per leaf. An average score was calculated for each location. Since we collected only infested leaves, there was no 0 score. The eggs of the citrus blackfly were excluded from the score.
The maps were made using the ArcGIS Pro, Version 3.0.3 (Redlands, CA: Environmental Systems Research Institute, Inc., https://www.esri.com), ESRI and Natural Earth (Free vector and raster map data @ naturalearthdata.com).

Figure 2.
A) Citrus blackflies Alerocanthus woglumi (arrow showing unparasitized pupa, and egg spirals); B) orange spiny whitefly, A. spiniferus and their parasitoid assembly established in Hawaii. The pupal stage of orange spiny whitefly with white wax fringe that surrounds their margins is twice as large as the orange spiny whitefly. C) Encaria perplexa male and female introduced from Guatemala 1998; D) Amitus hesperidum introduced from Guatemala 1998; E) Citrus blackfly parasitized pupae, arrows showing parasitoid’s circular exit holes. The oval-shaped pupa is black, convex, with dark dorsal spines. The marginal wax tubes produce a cottony fringe on the pupal margin. F) Encarsia nipponica adventive, G) Encarsia smithi introduced from Japan 1975 established on Oahu.
Figure 2.
A) Citrus blackflies Alerocanthus woglumi (arrow showing unparasitized pupa, and egg spirals); B) orange spiny whitefly, A. spiniferus and their parasitoid assembly established in Hawaii. The pupal stage of orange spiny whitefly with white wax fringe that surrounds their margins is twice as large as the orange spiny whitefly. C) Encaria perplexa male and female introduced from Guatemala 1998; D) Amitus hesperidum introduced from Guatemala 1998; E) Citrus blackfly parasitized pupae, arrows showing parasitoid’s circular exit holes. The oval-shaped pupa is black, convex, with dark dorsal spines. The marginal wax tubes produce a cottony fringe on the pupal margin. F) Encarsia nipponica adventive, G) Encarsia smithi introduced from Japan 1975 established on Oahu.
3. Results & Discussion
Survey results indicated that A. woglumi populations are rare on the Hawaiian Islands. Out of the 62 locations inspected for A. woglumi populations, its presence was found in only 11 sites (Figure 1). On Kauai Island no infestation was found and on Hawaii Island A. woglumi populations were found in just 3 sites out of the 23 inspected. Moreover, the infestation rate was quite low and only isolated citrus trees either on Oahu or in Hawaii Islands were found with large number of infested leaves (≥ 10 leaves). Throughout the survey, populations of other Aleyrodidae were very low and only a few isolated populations of Aleurothrixus floccosus (Maskell), and Aleurodicus dispersus Russell, were found.
Parasitism of A. woglumi nymphs was much higher on Oahu than Hawaii Island. Parasitism rates ranged 0 - 28 % on Hawaii Island and 11 - 65% on Oahu Island (Table 2 and Table 3).
Based on the morphological and molecular data, the emerged parasitoids were identified as E. perplexa. Only were a few of them identified as Amitus spiniferus Brethes, a parasitoid introduced for A. floccosus.
In specific, the molecular comparison via the NCBI database resulted in high similarity rates, suggesting that the analyzed parasitoids belong undoubtedly to the genus of Encarsia. Most of the produced consensus sequences were similar to E. perplexa at a range of 98.6 to 99.5%. The NCBI contained only limited data for the genus Amitus for both genes, a fact that limited the determination of some individuals to the genus level. Sequencing data provided from the USDA supported that these parasitoids are highly similar to the species Amitus spiniferus, much more than to the A. hesperidum.
Our survey results confirm that A. woglumi on the Hawaiian Islands is effectively controlled by the introduced natural enemies and that E. perplexa is the dominant parasitoid species. We were not able to find A. hesperidum either due to competition with E. perplexa or because its population is at very low levels not being detectable with our sampling effort during this short survey. In other locations where both E. perplexa and A. hesperidum have been released to control A. woglumi, soon after the reduction of the infestation, E. perplexa had become the dominant species. This is mainly due to its longer lifespan compared to A. hesperidum. The latter is considered an effective parasitoid at high densities of its host whereas E. perplexa is more efficient at low host density [11].
In 1997 the infestation by A. woglumi was so severe on Oahu Island that every citrus tree including oranges, lemon, and pummelo had a range of 50 - >600 citrus blackfly pupae/leaf (Figure 2 A). In 1998 all citrus trees on Oahu (urban trees and orchards) had reduced to no fruiting from the high infestation. There were complaints from citrus growers on the Hawaii and Kauai Islands requesting for parasitoid introductions. Recently, no reports of damage incurred by the pest or reports of growers or stakeholders having issues with citrus blackfly.
Non-citrus trees and ornamental shrubs (orange jasmine, Murraya paniculate (L.), Rutaceae) were also utilized for oviposition but were not normally affected by this whitefly. Mean number of egg mass/leaflet was 16.4 ± 2.8 (n= 25, during June 1997, Oahu Island) and 22.4 ± 3.1 eggs/mass (n=22). Citrus blackfly eggs hatched with no development beyond first nymph on non-citrus trees.
The infestation in 1997 was the worst on lemon trees (419.4 ± 32.4 pupae/leaf). Pummelo Citrus maxima (Burm.) Merr., infestation was even higher with 112.5 ± 14.0 nymphs/cm2 leaf, compared with Meyer lemon, Citrus meyeri Yu.Tanaka (45.8 ± 15.0 nymphs/cm2 leaf), pink tecoma, Tabebuia rosea DC., Bignoniaceae (13.8 ± 2.1 nymphs/cm2 leaf), and mango (5.5 ± 1.3 nymphs/cm2 leaf). These records demonstrated the potential of the citrus blackfly to infest the Hawaiian citrus and mango trees even faster than any other mainland States because of the favorite environmental conditions ([2], and unpublished data).
From the adult parasitoids that emerged either in Hawaii and shipped into Greece or emerged as adults in the quarantine facility of Benaki Phytopathological Institute in Athens >3000 adults were introduced into rearing cages with young Citrus aurantium plants infested with A. spiniferus. Parasitoids were introduced into the cages as they emerged from A. woglumi nymphs for a period of almost 20 days. Only E. perplexa adults emerged and they did not parasitize A. spiniferus in Greece. Parasitoids remain alive even after 30 days, without parasitizing any A. spiniferus nymph. Encarsia perplexa is known to parasitize A. woglumi, Aleuroclava kuwani (Takahashi) and Aleuroplatus pectiniferus Quaintance & Baker [22]. Amitus hesperidum was not recovered from the shipments. The parasitoid A. hesperidum was difficult to be recorded even soon after its initial release on the Hawaiian Islands due to extremely low population densities, which also confirmed from the current surveys. Encarsia smithi, a known parasitoid of A. spiniferus, has been found in the past on Oahu Island, [16] but was not recovered during the current surveys.
The current study confirms that A. woglumi is efficiently controlled by natural enemies on the Hawaiian Islands and that the dominant parasitoid is E. perplexa. Amitus hesperidum is either extinct or may have been present at extremely low populations due to the very low host density and scattered populations of A. woglumi. That parasitoid has a short adult life span, that limits its searching efficiency for new suitable hosts to parasitize. According to the results of the surveys and of the laboratory experiments we can conclude that the parasitoid E. perplexa which was recovered on the Hawaiian Islands is able to control A. woglumi but is not the appropriate species for A. spiniferus. Additional exploratory surveys are ongoing in Southeast Asia aiming to locate A. hesperidum or other Encarsia species that are reported to parasitize the orange spiny whitefly.
Acknowledgments
The current study has been partially funded by the Greek Ministry of Rural Development and Food. Mrs Iro Georgopoulou is thanked for creating the maps. We also thank Mr. Craig Kaneshige for helping in the transportation of the team members during the Kauai Island survey. Photos are credited to Walter Nagamine (Hawaii Department of Agriculture).
References
- Kapantaidaki, D., E., Antonatos, S., Kontodimas, D., Milonas, P., Papachristos, D., P. Presence of the invasive whitefly Aleurocanthus spiniferus (Hemiptera: Aleyrodidae) in Greece. Bulletin OEPP/EPPO Bulletin. 2019, 49 (1), 127–131.
- Ramadan, M., M., Kaufman, L., V., Wright, M., G. Insect and weed biological control in Hawaii: Recent case studies and trends. Biol. Control. 2023, 179, 105170. [CrossRef]
- EFSA Plant Health Panel (EFSA PLH Panel), Bragard, C., Dehnen-Schmutz, K., Di Serio, F., Gonthier, P., Jacques, M-A., Jaques Miret, J., A., Justesen, A., F., Magnusson, C., S., Milonas, P., Navas-Cortes, J., A., Parnell, S., Potting, R., Reignault, P., L., Thulke, H.,-H., Van der Werf, W., Vicent Civera, A., Yuen, J., Zappalà, L., Navarro, M., N., Kertesz, V., Czwienczek, E., MacLeod, A. 2018. Scientific Opinion on the pest categorisation of Aleurocanthus spp. EFSA Journal. 2018, 16 (10) :5436, 31 pp. [CrossRef]
- Akrivou, A., Georgopoulou, I., Papachristos, D., P., Milonas, P., G., Kriticos, D., J. Potential global distribution of Aleurocanthus woglumi considering climate change and irrigation. PLoS ONE, 2021, 16(12): e0261626. [CrossRef]
- Porcelli, F. First record of Aleurocanthus spiniferus (Homoptera: Aleyrodidae) in Puglia, Southern Italy. EPPO Bulletin, 2008, 38, 516– 518.
- Nugnes, F., Laudonia, S., Jesu, G., Jansen, M., G., M., Bernardo, U., Porcelli, F. Aleurocanthus spiniferus (Hemiptera: Aleyrodidae) in Some European Countries: Diffusion, Hosts, Molecular Characterization, and Natural Enemies. Insects, 2020, 11, 42. [CrossRef]
- Šimala, M., Masten Milek, T. First record of the orange spiny whitefly, Aleurocanthus spiniferus Quaintance, 1903 (Hemiptera: Aleyrodidae), in Croatia. Plant Zbornik predavanj in referatov 11. Slovenskega posvetovanja o varstvu rastlin z mednarodno udeležbo Bled, d. (Plant Protection Society of Slovenia). 2013, 5.–6.
- Radonjic, S., Hrncic, S., Malumphy, C. First Record of Aleurocanthus spiniferus (Quaintance) (Hemiptera: Aleyrodidae) in Montenegro. CRA-Research Centre for Agrobiology and Pedology, 2014, 97, 141– 154.
- EFSA (European Food Safety Authority), Schrader G, Camilleri M, Ciubotaru RM, Diakaki M and Vos S, Pest survey card on Aleurocanthus spiniferus and Aleurocanthus woglumi. EFSA supporting publication, 2019, EN-1565. 17pp. [CrossRef]
- Uesugi, R., Yara, K., Sato, Y. Changes in population density of Aleurocanthus camelliae (Hemiptera: Aleyrodidae) and parasitism rate of Encarsia smithi (Hymenoptera: Aphelinidae) during the early invasion stages. Appl. Entomol. Zool. 2016, 51, 581–588, . [CrossRef]
- Nguyen, R., Brasil, J., R., Poucher, C. Population density of the citrus blackfly, Aleurocanthus woglumi Ashby (Homoptera: Aleyrodidae), and its parasites in urban Florida in 1979-81. Environmental Entomology, 1983, 12: 878-884.
- Dowell, R.V. Factors affecting the field effectiveness of Encarsia opulenta (Hymenoptera: Aphelinidae), a parasitoid of citrus blackfly, Aleurocanthus woglumi (Homoptera: Aleyrodidae). Trop. Agr. 1989, 66: 110–112.
- Lopez, V., F., Kairo, M., T., Pollard, G., V., Pierre, C., Commodore, N., Dominique, D., 2009. Post-release survey to assess impact and potential host range expansion by Amitus hesperidum and Encarsia perplexa, two parasitoids introduced for the biological control of the citrus blackfly, Aleurocanthus woglumi in Dominica. BioControl, 2009, 54: 497–503. [CrossRef]
- Yamashita, K., Kasai, A., Suzuki, Y., Yoshiyasu, Y. Population dynamics of the camellia spiny whitefly, Aleurocanthus camelliae (Hemiptera: Aleyrodidae), in tea fields during the early phase of invasion into Kyoto, Japan. Applied Entomology and Zoology, 2016, 51, 117– 124.
- Anonymous, 1974. New States records orange spiny black fly, Aleurocanthus spiniferus (Quaintance). Hawaii Co-operative Economic Insect Report 24, 585 (R.A.E. Services A, 1974, (1975) 63 (781).
- Culliney, T., W., Nagamine, W., T., Teramoto, K., K. Introductions for Biological Control in Hawaii 1997-2001. Proc. Hawaii. Entomol. Soc. 2003, 36, 145-153. https://scholarspace.manoa.hawaii.edu/bitstream/10125/112/1/36_145-153.pdf.
- Nakao, H., K., Funasaki, G., Y. Introductions for biological control in Hawaii: 1975 and 1976. Proc. Hawaii. Entomol. Soc. 1979, 23:125–128.
- Heu, R., A., Nagamine, W., T. Citrus Blackfly Aleurocanthus woglumi Ashby (Homoptera: Aleyrodidae). New Pest Advisory, 2001, No. 99-03.
- Rossato, V. Ocorrência de parasitóides de Aleurocanthus woglumi Ashby, 1903 (Hemiptera: Aleyrodidae) e seu parasitismo por Cales noacki Howard, 1907 (Hymenoptera: Aphelinidae) nos municípios de Belém, Capitão Poço e Irituia no estado do Pará. Dissertação de Mestrado, Universidade Federal Rural da Amazônia, Belém/PA. 2007, 40 pp.
- MacGown, M., W., Nebeker, T., E. Taxonomic review of Amitus (Hymenoptera: Proctotrupoidae, Platygastridae). Can. Ent. 1978, 110: 275-283.
- Viggiani, G., Mazzone, P. The Amitus Hald. (Hym. Platygastridae) of Italy, with description of three new species. Bollettino de Laboratorio di entomologia agrarian “Filippo Silvestri” Portici. 1978, (39):59-69.
- Huang, J., Polaszek, A. A revision of the Chinese species of Encarsia Forster (Hymenoptera: Aphelinidae): parasitoids of whiteflies, scale insects and aphids (Hemiptera: Aleyrodidae, Diaspididae, Aphidoidea). Journal of Natural History, 1998, 32: 1825-1966.
- Nguyen, R. Amitus hesperidum (Hymenoptera: Platygasteridae), a parasite of the citrus blackfly (Aleurocanthus woglumi). Florida Department of Agricultural Consumer Services, Division Plant Industry, Entomolgy Circular.1988, 311:1–2. https://entnemdept.ufl.edu/creatures/beneficial/amitus_hesperidum.htm.
- Noyes, J.S. 2019. Universal Chalcidoidea Database. World Wide Web electronic publication. http://www.nhm.ac.uk/chalcidoids.
- Noyes, J., S. Collecting and preserving chalcid wasps (Hymenoptera: Chalcidoidea). Journal of Natural History, 1982, 16 (3): 315-334.
- Noyes, J., S. Chapter 2.7.2.5. Chalcid parasitoids. (In: Rosen, D. [editor] - The Armored Scale Insects. Their Biology, Naural Enemies and Control.) World Crop Pests, 1990, 4B:247-262.
- Hebert, P.D., Cywinska, A., Ball, S.L., deWaard, J.R. Biological identifications through DNA barcodes. Proc. Biol. Sci. 2003, 270, 313–321. [CrossRef]
- Pentinsaari, M., Salmela, H., Mutanen, M., Roslin, T. Molecular evolution of a widely-adopted taxonomic marker (COI) across the animal tree of life. Sci. Rep. 2016, 6, 35275. [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol Mar Biol Biotechnol, 3(5), 294-299. PMID: 1994, 7881515.
- Dowton, M., Austin. A.D., Simultaneous analysis of 16S, 28S, COI and morphology in the Hymenoptera: Apocrita–evolutionary transitions among parasitic wasps. Biol. J. Linn. Soc. 2001, 74, 87- 111. [CrossRef]
Figure 1.
Sampling locations for Aleurocanthus woglumi and its parasitoids in the Hawaiian Islands.
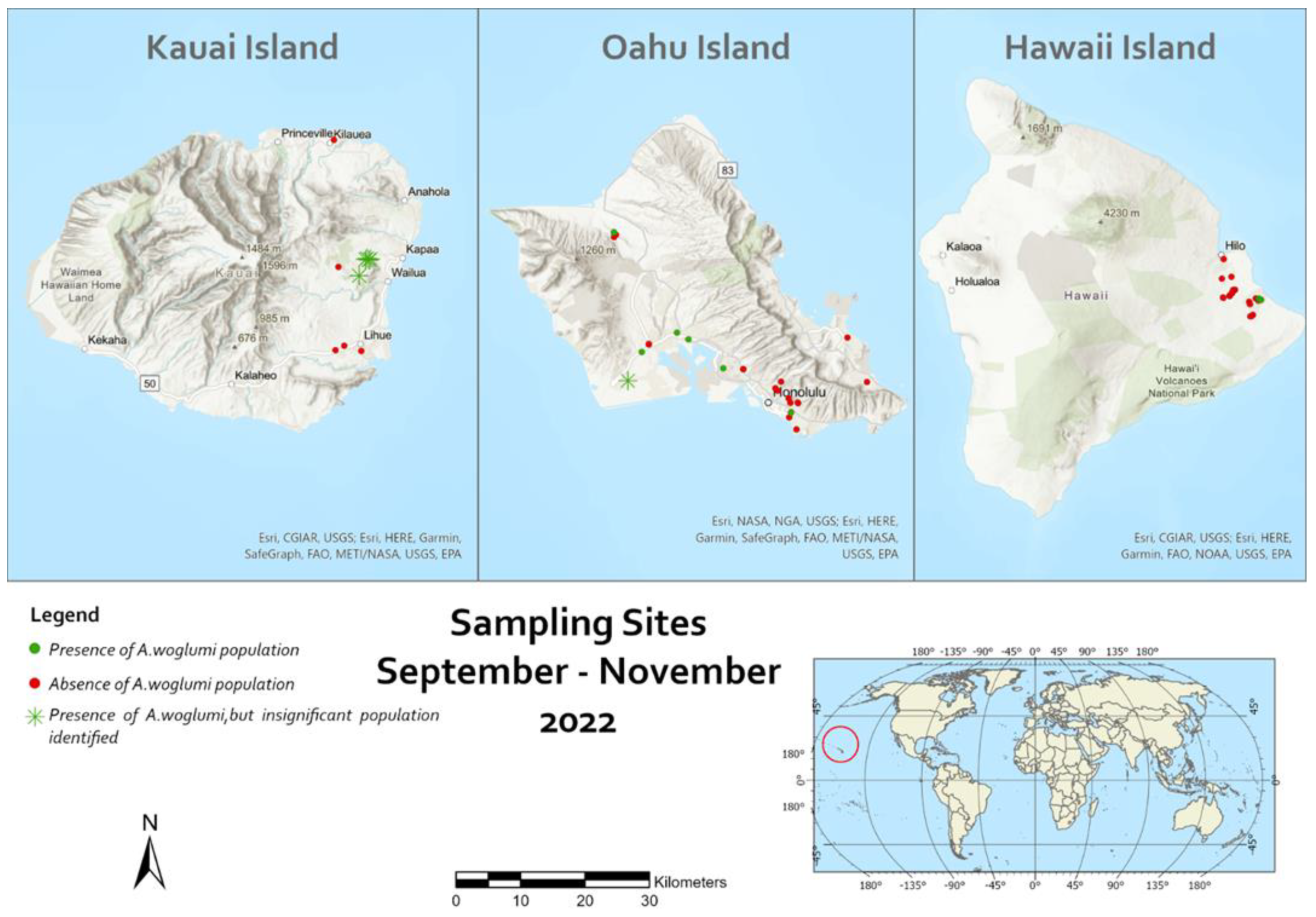

Table 1.
Hymenopterous parasitoid assembly of Aleurocanthus spiniferus and A. woglumi confirmed on the Hawaiian Islands, 1974 – 2002.
Table 1.
Hymenopterous parasitoid assembly of Aleurocanthus spiniferus and A. woglumi confirmed on the Hawaiian Islands, 1974 – 2002.
| Target whitefly (Hemiptera: Aleyrodidiae) |
Introduced and adventive natural enemies | ||||||
|---|---|---|---|---|---|---|---|
| Family | species | Origin of introduction |
Year of introduction and release period | Colonization records (total numbers released per island) |
Establishment records |
Release records |
|
|
Aleurocanthus spiniferus (Quaintance), citrus orange spiny whitefly |
Platygastridae |
Amitus hesperidum Silvestri |
Mexico | 1974 | Recorded on Oahu | Only Oahu Island | |
| Platygastridae |
Amitus spiniferus (Brethes) |
California | 1979-1982 | Oahu (200) | Introduced for Aleurothrixus floccosus (Maskell). Established on Oahu Island. |
Only on Oahu Island in 1982. |
|
| Aphelinidae |
Cales noacki Howard |
California | Feb. 1981 - Aug. 1982 | Lanai (275) Molokai (300) Oahu (12149) |
Introduced for Aleurothrixus floccosus (Maskell) Hawaii 1992 Kauai 1997 Oahu 1982 |
Hawaii 1992 Lanai 1982 Molokai 1982 Oahu 1982 |
|
| Aphelinidae |
Encarsia clypealis (Silvestri) |
Texas | 1975 | No recovery | Only on Oahu Island. |
||
| Aphelinidae |
Encarsia perplexa Huang & Polaszek |
Texas | 1975 | Oahu (165) | No recovery | Only on Oahu, released as Encarsia opulenta (Silvestri). |
|
| Aphelinidae |
Encarsia smithi (Silvestri) |
Japan (Nagasaki) and Guam |
September 1974 | Oahu (5330) | Established on Oahu Sep. 1975, reported 1981, 1984, 1999 on Oahu. |
Only on Oahu Island, some of the colony from Guam 1984 record (25 adults). |
|
|
Aleurocanthus woglumi Ashby, citrus blackfly |
Platygastridae |
Amitus hesperidum Silvestri |
Guatemala | Introduced 1998, released May 1999 -August 2000 |
Hawaii (20), Kauai (112), Maui (82), Oahu (8595) |
Established | Hawaii, Kauai, Maui, Molokai, Oahu Islands |
| Aphelinidae |
Cales noacki Howard |
California | Feb. 1981 - Aug. 1982 | Molokai (300) Oahu (12149) |
Introduced for Aleurothrixus floccosus (Maskell) |
Only on Oahu Island in 1982. Recorded established on Oahu Island. |
|
| Aphelinidae |
Encarsia perplexa Huang & Polaszek |
Guatemala | Introduced 1998, released April 1999 – June 2002 |
Hawaii (49190) Kauai (17090) Maui (47025) Molokai (27165) Oahu (5740) |
Established | Hawaii, Kauai, Maui, Molokai, Oahu Islands. First identified as Encarsia opulenta (Silvestri) |
|
| Aphelinidae | Encarsia nipponica Silvestri | Adventive, native to Japan and China |
- | - | Hawaii 2000, Kauai 2001 Oahu 1997, 1999 |
Fortuitous species | |
| Aphelinidae | Encarsia smithi (Silvestri) | Japan (Nagasaki) and Guam |
September 1974 | Reported on Oahu 1989, 1999 on citrus | Established on Oahu Sep. 1975 Reported in 1999 on Oahu |
Only on Oahu Island. |
|
Table 2.
Infestation rates and parasitization of A. woglumi infesting Citrus species on Oahu Island during September-November 2022. a residential house, roadside, highway service area, farmland, research station, community garden, cemetery, park, church.
Table 2.
Infestation rates and parasitization of A. woglumi infesting Citrus species on Oahu Island during September-November 2022. a residential house, roadside, highway service area, farmland, research station, community garden, cemetery, park, church.
| Survey area and habitata |
Citrus Plant | Coordinate and elevation |
Date | Total Number of leaves collected | Total Number of nymphs |
Level of infestation |
% parasitism |
|---|---|---|---|---|---|---|---|
| University of Hawaii at Manoa |
Sour orange | 21º 18’ 17.98” N 157º 48’ 51.87” W, 40 m |
26/9/2022 | 0 | 0 | 0 | - |
| University of Hawaii at Manoa |
Sweet lime | 21º 18’ 15.16” N 157º 48’ 48.99” W, 38 m |
“ | 0 | 0 | 0 | - |
| Ala Wai Community Garden |
lime | 21º 17’ 02.50” N 157º 49’ 37.15” W, 1.5 m |
“ | 0 | 0 | 0 | - |
| Waimanalo Research Station |
Citron, | 21º 20’ 07.91” N 157º 42’ 55.01” W, 23 m |
“ | 0 | 0 | 0 | - |
| Kapaakea Lane, Honolulu | tangerine | 21º 17’ 27.82” N 157º 49’ 25.95” W, 4 m |
“ | 0 | 0 | 0 | - |
| Kapaakea Lane, Honolulu | pummelo | 21º 17’ 25.55” N 157º 49’ 26.61” W, 3.6 m |
“ | 9 | 118 | 1.44 ± 0.49 | 17.8% |
| Pearl City Urban Botanical Garden | Sweet orange | 21º 23’ 38.76” N 157º 58’ 38.08” W, 8.5 m |
27/9/2022 | 71 | 1471 | 1.76 ± 0.78 | 56.6% |
| Diamond Head Community Garden |
tangelo | 21º 16’02.09” N 157º 58’ 59.55” W, 0.9 m |
“ | 0 | 0 | 0 | - |
| Moanalua Gardens | Grapefruit | 21º 20’ 52.68” N 157º 53’ 28.74” W, 6.4 m |
“ | 0 | 0 | 0 | - |
| North Shore. Poamoho Research Station |
Sweet orange | 21º 32’ 38.00” N 158º 05’ 16.28” W, 189 m |
28/9/2022 | 345 | 11981 | 2.17 ± 0.95 | 19.7% |
| Salt Lake Blvd | Sweet lime | 21º 21’ 09.13” N 157º 55’ 30.36” W, 29 m |
“ | 75 | 158 | 1.37 ± 0.48 | 53.8% |
| North Shore, Hawaii Queen bees, Hinshaw Farms |
Sour orange | 21º 32’ 11.10” N 158º 05’ 16.82” W, 226 m |
“ | 0 | 0 | 0 | - |
| North Shore, Hawaii Queen bees, Hinshaw Farms |
lemon | 21º 32’ 17.54” N 158º 05’ 14.90” W, 225 m |
“ | 0 | 0 | 0 | - |
| Hawaii Agriculture Research Center, Kunia | tangelo | 21º 23’ 09.02” N 158º 02’ 14.67” W, 82.6 m |
2/10/2022 | 0 | 0 | 0 | - |
| Hawaii Agriculture Research Center 2 |
lemon | 21º 23’ 07.28” N 158º 02’ 17.16” W, 84.4 m |
“ | 0 | 0 | 0 | - |
| Manoa valley community gardens |
citron | 21º 18’ 53.77” N 157º 48’ 25.49” W, 57 m |
“ | 0 | 0 | 0 | - |
| Nuuanu Mauna ala Royal mausoleum | pummelo | 21º 19’ 30.33” N 157º 50’ 49.81” W, 64 m |
26/10/2022 | 0 | 0 | 0 | - |
| Nuuanu old Pali drive |
lemon | 21º 21’ 14.24” N 157º 48’ 37.68” W, 308 m |
“ | 0 | 0 | 0 | - |
| Tantalus lookout | tangerine | 21º 19’ 06.92” N 157º 49’ 48.23” W, 277 m |
“ | 0 | 0 | 0 | - |
| Kailua | lemon | 21º 23’ 47.09” N 157º 44’ 44.49” W, 0.6 m |
“ | 0 | 0 | 0 | - |
| Mahiole street, Moanaloa | Sour orange | 21º 20’ 49.79” N 157º 53’ 21.81” W, 11 m |
31/10/2022 | 0 | 0 | 0 | - |
| North Shore Poamoho Research Station |
sweet orange, sweet lime | 21º 32’ 38.00” N 158º 05’ 16.28” W, 189 m |
“ | 573 | 23168 | 2.34 ± 0.81 | 24.9% |
| Waikele | tangerine | 21º 24’ 03.18” N 158º 00’ 13.75” W, 60.6 m |
1/11/2022 | 13 | 389 | 2.00 ± 1.03 | 34.4% |
| Aloun farm | pummelo | 21º 22’ 28.47” N 158º 02’ 42.94” W, 56.7 m |
“ | 14 | 212 | 1.78 ± 0.67 | 11.5% |
| Royal Mausoleum of Hawaii |
lime | 21º 19’ 30.33” N 157º 50’ 49.81” W, 64 m |
“ | 0 | 0 | 0 | - |
Table 3.
Infestation rates and parasitization of A. woglumi infesting Citrus species on Hawaii Island during October 2022. a residential house, roadside, highway service area, farmland, research station, community garden, cemetery, park, church.
Table 3.
Infestation rates and parasitization of A. woglumi infesting Citrus species on Hawaii Island during October 2022. a residential house, roadside, highway service area, farmland, research station, community garden, cemetery, park, church.
| Survey area and habitata |
Citrus Plant | GPS and elevation |
Date | Total Number of leaves collected |
Total Number of nymphs |
Level of infestation |
% parasitism |
|---|---|---|---|---|---|---|---|
| Waiakea | Sweet orange | 19º 38’ 30.98” N 155º 04’ 51.67” W, 194 m |
27/10/2022 | 0 | 0 | 0 | - |
| Waiakea Research station | lime | 19º 38’ 41.46” N 155º 04’ 37.82” W, 173 m |
“ | 0 | 0 | 0 | - |
| Hawaii Department of Agriculture, Hilo | lemon | 19º 42’ 23.03” N 155º 04’ 26.25” W, 11 m |
“ | 0 | 0 | 0 | - |
| Kurtistown 1 | Pummelo | 19º 35’ 35.98” N 155º 03’ 30.41” W, 206 m |
“ | 0 | 0 | 0 | - |
| Kurtistown 2 | Pummelo | 19º 35’ 30.36” N 155º 03’ 29.91” W, 199 m |
“ | 0 | 0 | 0 | - |
| Kurtistown 3 | Sweet orange | 19º 35’ 00.29” N 155º 03’ 36.53” W, 242 m |
“ | 0 | 0 | 0 | - |
| Kurtistown 4 | Sour orange | 19º 34’ 38.83” N 155º 03’ 56.19” W, 271 m |
“ | 0 | 0 | 0 | - |
| Hawaiian Paradise Park 1 | Grapefruit | 19º 36’ 12.88” N 154º 56’ 51.46” W, 13 m |
28/10/2022 | 173 | 17848 | 3.14 ± 0.76 | 2.7% |
| Hawaiian Paradise Park 2 | Pummelo | 19º 36’ 12.88” N 154º 56’ 51.46” W, 13 m |
“ | 7 | 423 | 3.00 ± 0.50 | 0.0% |
| Hawaiian Paradise Park 3 | Pummelo | 19º 34’ 17.70” N 154º 57’ 21.50” W, 50 m |
“ | 6 | 212 | 2.50 ± 0.80 | 28.3% |
| Hawaiian Paradise Park 4 | Sweet orange | 19º 34’ 35.47” N 154º 57’ 18.39” W, 9 m |
“ | 0 | 0 | 0 | - |
| Hawaiian Paradise Park 5 | citron | 19º 34’ 52.95” N 154º 57’ 39.69” W, 39 m |
29/10/2022 | 0 | 0 | 0 | - |
| Hawaiian Paradise Park 6 | lime | 19º 34’ 14.46” N 154º 57’ 45.16” W, 40 m |
30/10/2022 | 0 | 0 | 0 | - |
| Hawaiian Paradise Park 7 | lemon | 19º 35’ 39.96” N 154º 57’ 21.77” W, 28 m |
31/10/2022 | 0 | 0 | 0 | - |
| Keeau 1 | Sour orange | 19º 37’ 06.79” N 155º 02’ 37.40” W, 121 m |
29/10/2022 | 0 | 0 | 0 | - |
| Keeau 2 | lemon | 19º 36’ 52.38” N 155º 02’ 48.90” W, 149 m |
“ | 0 | 0 | 0 | - |
| Keeau 3 | tangerine | 19º 36’ 23.02” N 155º 02’ 59.69” W, 177 m |
“ | 0 | 0 | 0 | - |
| Keeau 4 | Sweet orange | 19º 37’ 11.96” N 155º 02’ 32.56” W, 117 m |
“ | 0 | 0 | 0 | - |
| Keeau 5 | Pummelo | 19º 37’ 42.04” N 155º 02’ 19.12” W, 123 m |
“ | 0 | 0 | 0 | - |
| Keeau 6 | tangelo | 19º 38’ 42.04” N 155º 02’ 32.56” W, 79 m |
“ | 0 | 0 | 0 | - |
| Ainaloa 1 | tangerine | 19º 31’ 40.76” N 154º 59’ 35.89” W, 217 m |
“ | 0 | 0 | 0 | - |
| Ainaloa 2 | lime | 19º 31’ 20.12” N 155º 00’ 05.43” W, 226 m |
“ | 0 | 0 | 0 | - |
| Ainaloa 3 | lemon | 19º 31’ 47.83” N 154º 59’ 49.05” W, 201 m |
“ | 0 | 0 | 0 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



