
Submitted:
12 October 2023
Posted:
12 October 2023
You are already at the latest version
Abstract
Cabbage (B. oleracea var. capitata) contains three FLC homolog genes (BoFLC1, BoFLC2, and BoFLC3) and one pseudogene (BoFLC5) that responds to low temperatures necessary for flowering. We isolated three BoFLC-encoding genes from early-, mid-, and late-flowering cabbage plants. Leaf samples were collected for RNA extraction and expression analysis. Gene structure analysis and phylogenetic comparison were performed for intra- or interspecific relationships of the BoFLC homologs. Gene expression related to flowering regulators (BoGI, BoCOOLAIR, and BoVIN3) was analyzed. While BoFLC genes commonly have seven exons and six introns of 3,361–4.384 bp, variations in insertion or deletion were evident between the early- and late-flowering genotypes. Repressed expression of BoFLC 1, 2, and 3 genes under chilling temperatures appeared to occur from 8 weeks after vernalization. Higher expression levels of GI, COOLAIR, and VIN3 were initiated 7 weeks after chilling (WAT) treatment in the early flowering genotype. The vernalization trigger repressed the expression of BoFLC homologs. This study provides molecular insights into BoFLC homologs between early- and late-flowering cabbage genotypes, in which structural variations in BoFLC1 appeared to be important as a binding motif to flowering regulating factors such as BoGI, BoCOOLAIR, and BoVIN3 for which motif analysis is further implemented.
Keywords:
BoFLC homologs
; cabbage flowering
; flowering regulator
; structural variation
; vernalization
1. Introduction
In plants, the regulation of flowering time is one of the most important events that significantly affect the synchronization of reproduction under favorable environmental conditions. Floral transition, such as the vegetative-to-reproductive phase, is a vital developmental process that governs the timing of reproduction. Environmental (photoperiod and temperature) and endogenous factors (autonomous, gibberellins [GA], and aging), functionally already characterized in Arabidopsis, regulate this floral transition [1,2,3,4]. Cold temperature-induced floral transitions such as vernalization are widely present in long-day plants in biennial or winter forms. This vernalization pathway comprises many genes that act as transcriptional and epigenetic silencing mechanisms to form a complex regulatory network that controls flowering time.
Brassicaceae are some of the most economically important crop species in horticulture and are cultivated mainly for their edible seeds, stems, leaves, and flowers. In spring, crops in this family require prolonged vernalization to cause premature flowering, which results in bolting and impairing yield [5]. Therefore, breeding late-flowering cabbages that are resistant to cold temperatures and bolting is of considerable interest to breeders and researchers[6,7]. Important vernalization components, along with flowering-related genes (FLC, FRI, and VIN3), are evolutionarily conserved between the spring and winter forms of Arabidopsis and Brassica crops[8,9,10,11,12]. Natural variation in flowering time between spring and winter forms is mostly associated with allelic variations in paralogs of FLC locus, which has already been characterized in many Brassica species, including B. oleracea[13,14,15], B. rapa[16,17], B. napus [18,19].
The reference genome of B. oleracea contains four copies of BoFLC1/2/3 and a pseudogene of BoFLC5 orthologs[10,20]. In earlier studies, only BoFLC2 was the main functional gene responsible for floral transitions[10,14]. Supporting this, mutations in BoFLC2 cause a loss-of-function in annual cauliflower varieties[13]. A recent transformation study showed that three BoFLCs delayed flowering in Arabidopsis and suggested that these paralogs were functionally conserved as floral repressors in cabbage[21]. However, the investigated mRNA expression characteristics revealed that BoFLC1 declined less markedly than the highly repressed genes BoFLC2 or 3 under cold treatment[21]. In addition, an insertion mutation in intron 2 of BoFLC1 and its rapid downregulation under vernalization was strongly associated with the vernalization-dependent phenotype of an early flowering cabbage line. Allelic diversity and variations in FRI have also been closely linked to flowering or heading date variations in B. oleracea [22,23] and B. napus [24].
To date, variations in flowering time and vernalization sensitivity among the contrasting flowering time (early, mid-, and late) phenotypes of cabbage cultivars have been inadequately investigated. In this study, we characterized three BoFLC functional members, BoFLC1, BoFLC2/4, and BoFLC3, in the early-, mid-, and late-flowering cabbage cultivars. We found that allelic variations at the BoFLC locus could alter vernalization sensitivity between cabbage cultivars. This also suggests that these allelic nucleotide variations may influence the epigenetic silencing of FLC, as noted in Arabidopsis. Therefore, we propose that the identification of cis polymorphisms in BoFLC1, BoFLC2, and BoFLC3 provides beneficial information for breeding Brassica crops resistant to cold and late-flowering lines.
Arabidopsis contains a key gene for repressing floral transition called Flowering Locus C (FLC), which encodes a MADS-box transcription factor[25]. During vernalization, floral integrators, including Flowering Locus T (FT)[25], and transcription factors of MADS-box encoding proteins, such as Suppressor of Overexpression of Constans1 (SOC1) [26]and Leafy (LFY) [27,28] are activated, consequently promoting flowering by reducing the gene product of FLC. Gigantea (GI), a circadian clock-associated protein, is involved in various physiological processes, including the regulation of flowering time[29]. GI positively regulates Constans (CO) and FT, suggesting a role in floral transitions[30,31].
Extended vernalization induces genetic and epigenetic regulatory mechanisms that control the transcription of FLC by stabilizing histone proteins. Under non-vernalization conditions, FLC expression is increased by an activator called FRIGIDA (FRI), which recruits chromatin modification factors to the FLC chromatin, thereby repressing flowering time. Natural antisense transcripts (NATs) play pivotal roles in the epigenetic silencing of FLC [32]. Cold temperature induces the transcription of noncoding sense and antisense transcripts of COLDAIR and COOLAIR from the first intronic and multi-exonic or promoter adjacent 3’ regions in the FLC locus, respectively[33,34]. COLDAIR aids in the addition of repressive factors to FLC chromatin through interaction with VERNALIZATION INSENSITIVE 3 (VIN3) by recruiting the PRC2-like complex, a group of polycomb proteins, to the histones of FLC[34]. Independently from this, but before the decrease of FLC expression by the PRC2-like complex, antisense transcripts of COOLAIR are produced, repressing the sense transcripts of FLC [33].
2. Results
2.1. Molecular characterization of BoFLC-encoding genes from different cabbage bolting and flowering lines (variation in gene structures)
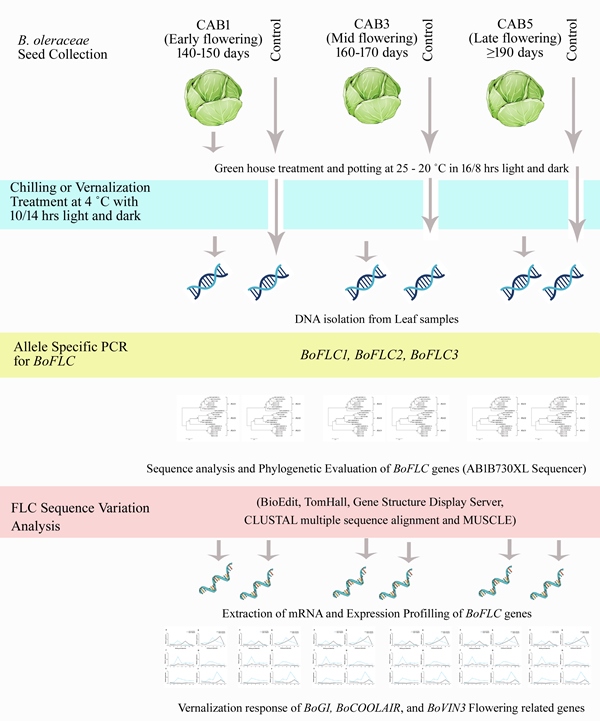
Experiments were conducted on cabbages with different flowering periods (CAB1: early-flowering at 140–150 days, CAB3: mid-flowering at 160–170 days, and CAB5: late-flowering at ≥ 190 days). Genomic DNA sequences ranging from approximately 3361 bp to 4384 bp for BoFLC1, BoFLC2, and BoFLC3, and CDS and untranslated regions of 594 bp were obtained (Figure 1). In the BoFLC1 gene sequence, the difference between CAB1 and CAB5 was in intron 1. Insertions of 23 and 335 bp were in intron 1 of CAB5 (Figure 1A). BoFLC2 also had a deletion in intron 1; however, there was no difference in BoFLC3 (Figure 1B-C). The reference and BoFLC gene sequences were compared (Figure S1). In the BoFLC1 gene sequence, 687 and 67 bp insertions were in introns 1 and 2, respectively, in early flowering type cabbage (CAB1), mid-flowering type cabbage (CAB3), and late-flowering type cabbage (CAB5), compared to the previously known AM231517.1 sequence. Unlike CAB1, CAB3,5 had 23, 613, and 5 bp deletions and 3, 9, 9, and 13 bp insertions in intron 1. CAB1 had 70, 40, 13, 29, and 78 bp deletions in introns 5 and 6, and CAB2 had 70, 70, 40, 5, 13, 29, and 13 bp deletions in introns 5 and 6. In CAB3, deletions of 70, 70, 40, 5, 13, 29, and 78 bp were observed in introns 5 and 6 (Figure S1A). In the BoFLC2 gene sequence, similar to the BoFLC1 gene, CAB3 and 5 had a 215 bp deletion in intron 1. In intron 6, CAB2 had 9, 2, and 1 bp insertions, 4, 2 bp deletions, and CAB3 had 9 bp insertions, 4, 2 bp deletions. CAB1 was 100% consistent with the previously known AY306124.1 sequence (Figure S1B). Finally, BoFLC3, CAB1, CAB3, and CAB5 all had 13 and 21 bp insertions and 2, 2, and 11 bp deletions in intron 1; specifically, it was confirmed that 15 bp insertions only existed in intron 5 in CAB3 (Figure S1C). The percent identity matrix was obtained using CLUSTAL multiple sequence alignment with ClustalOmega (http://www.ebi.ac.uk/Tools/msa/muscle/, version 3.8). BoFLC1, BoFLC2, and BoFLC3 were based on the sequences of Brassica species (Table S1). SNPs and Indels of the FLC were abundant in cabbage species. In genomic DNA, a significant In/Del in Introns was found in all FLC1, FLC2, and FLC3 genes (Figure S1), but in the CDS, it was confirmed that the similarity between each gene was high. In addition, the CDS of other Brassica species were highly similar (Table S3).
2.2. Phylogenetic relationships among B. oleraceae and other Brassica species
We investigated the relationship between the amino acid sequences (BoFLC1 (CAJ77613), BoFLC2 (AAQ76275), and BoFLC3 (CAJ77614), BrFLC1 (DQ866874.1), BrFLC2 (DQ866875.1), BrFLC3 (ABI30001.1), BnFLC1 (AAK70215.1), BnFLC2 (AAK70216.1), BnFLC3 (AAK70217.1), BniFLC (AIE38010.1), BcFLC (QCE32013.1), BjFLC (AHY82602.1)) using the neighbor-joining tree using protein levels (Figure 2). FLC homologs were evidently distinguishable from B. oleraceae (Bo), B. rapa (Br), B. napus (Bn), and B. nigra. BoFLC1, BoFLC2, and BoFLC3 proteins of CAB1 were closely related to the known cabbage proteins. CAB3 and CAB5 were highly correlated with FLC homology, indicating that most of the correlations between the proteins of the FLC homologs of B. oleraceae were close. However, it can be assumed that the complex intron In/Del caused a difference in flowering time.
2.3. Transcriptional expression patterns of BoFLC genes by vernalization
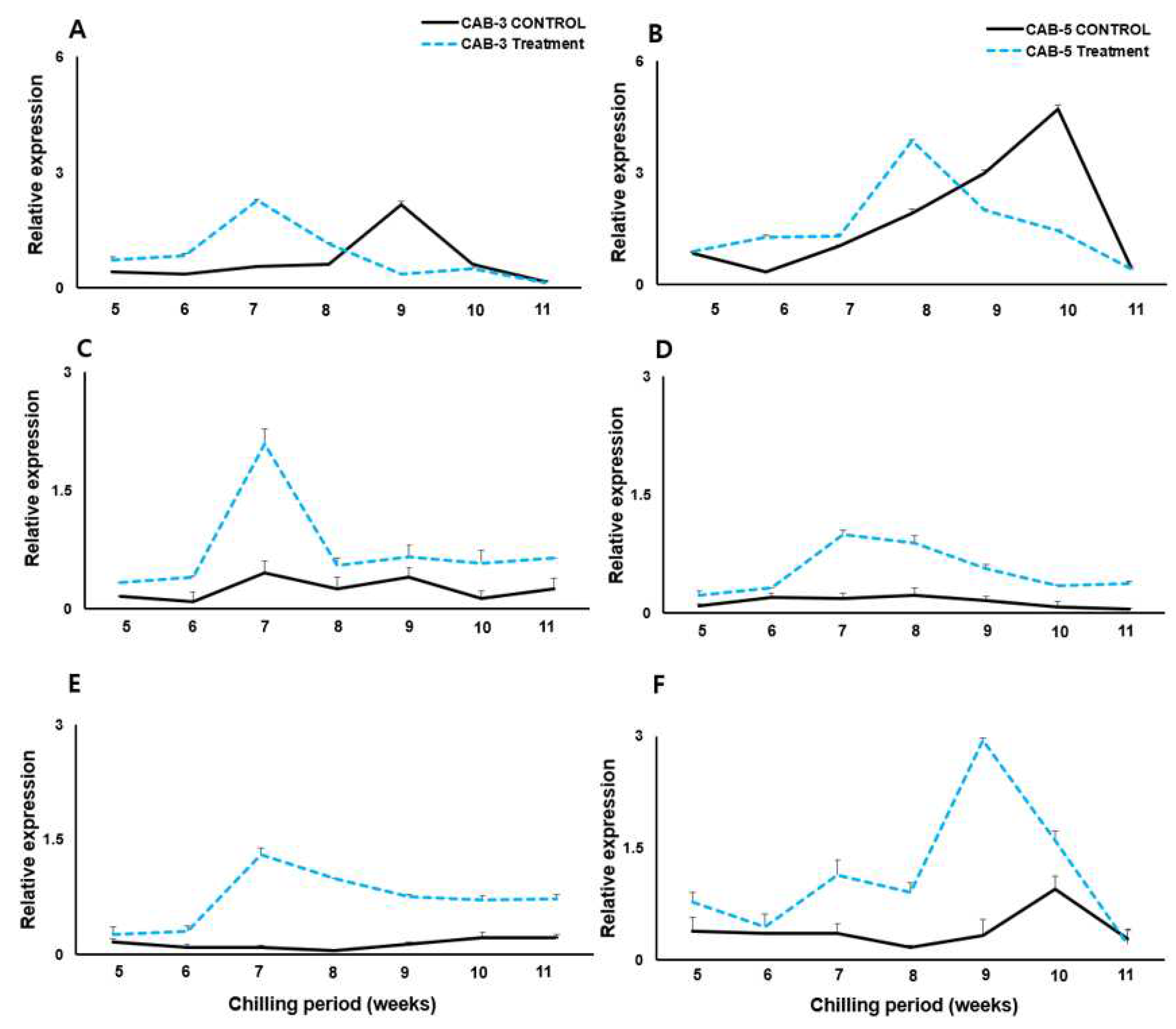
We performed transcript expression analysis using qRT-PCR to determine the effect of cold treatment on the BoFLC genes (Figure 3). Mid-flowering (CAB3) and late-flowering (CAB5) cabbages were sampled by low-temperature treatment from 0 to 11 weeks, and RNA was extracted and analyzed for expression from 5 to 11 weeks. Graphs were drawn with the CAB3 treatment, CAB5 control, and CAB5 treatment values as relative values based on the CAB3 control value. First, the controls were maintained without significant differences between the FLC homologs. In addition, the basic expression level in CAB5 was approximately two times higher than CAB3 in the control grown in a greenhouse. BoFLC1 plays a role in inhibiting flowering, and the late-flowering type undergoes significant low-temperature sensitization only after exposure to low temperatures for approximately 7 weeks[15]. Our experimental results confirmed that BoFLC1 expression decreased rapidly in CAB5 cells at low temperatures for approximately 8 weeks (Figure 3A). At low temperatures, CAB3 decreased BoFLC1 expression at 8 weeks. However, the expression did not continue to decrease, and BoFLC1 expression was maintained for 9 weeks in both cabbages under the low-temperature treatment. In the case of the FLC homologous gene BoFLC2, the expression of both CAB3 and 5 cabbages began to decrease after approximately 7 weeks of cold treatment, and the expression values were maintained until 9 weeks. BoFLC3 was similar to BoFLC1 in both CAB3 and the five cabbage varieties.
2.4. Vernalization response of BoGI, BoCOOLAIR, and BoVIN3 in cabbage
In addition to BoFLC, qRT-PCR analyses of BoGI, BoCOOLAIR, and BoVIN3 were also performed to investigate the expression of flowering-related genes. GIGANTEA (GI), which is related to the circadian clock and the control of flowering time, had a faster time to peak than the control. GI is controlled by temperature and light[37]. In the treatments we tested, the peak appeared faster for approximately 2 weeks compared to the control (Figure 4A). The expression of CAB3, which peaked at 9 weeks in the control group, was highest at 7 weeks after cold treatment. For CAB5, only the timing of expression was delayed, but the trend was the same; however, a difference existed in the amount of expression. In Arabidopsis thaliana, COOLAIR plays a role in regulating AtFLC expression as a long non-coding RNA[33,34]. We performed qRT-PCR to determine the trend of COOLAIR expression in cabbage plants (Figure 4B). Both CAB3 and CAB5, which continued the low-temperature treatment, showed the highest expression levels at 7 weeks. This coincided with the time when the expression of the BoFLC gene began to decrease (Figure 4). In CAB3, it increased rapidly at week 7 and then rapidly decreased again. In contrast, for CAB5, it increased more slowly and then gradually decreased. Among the vernalization pathway genes related to flowering, VIN3 inhibits the transcription of FLC by methylation through a series of processes that deacetylate specific regions of FLC chromatin, along with histone deacetylases (HDAC) and DNA recognition proteins in cold Arabidopsis thaliana[4]. The highest expression was observed at 7 weeks in the CAB3 low-temperature treatment and at 9 weeks in the late-flowering type low-temperature treatment. In CAB3, expression decreased after 8 weeks but was maintained after 9 weeks, whereas in CAB5, expression decreased continuously after 9 weeks.
3. Discussion
Various environmental and genetic factors affect flowering. Low temperatures are a major factor that essentially promotes flowering. Among the various genes that regulate flowering at such low temperatures, Arabidopsis contains a key gene that inhibits flower metastasis called FLOWERING LOCUS C (FLC), which encodes a MADS-box transcription factor[25]. Differences in FLC homolog genes were investigated in cabbage plants at different flowering times. BoFLC1, BoFLC2, and BoFLC3 were cloned using genomic DNA and compared to known genes. The sequences of the BoFLC1 gene (Bo9g173400) and CAB1 were similar, but those of CAB3 and CAB5 were different. BoFLC1 sequences were also similar in CAB3 and CAB5, but the difference between the two groups was an In/Del in intron 1. There were three deletions and four insertions in CAB3 and CAB5, respectively. This indicates that flowering may be delayed because the antisense function of COOLAIR and COLDAIR [34] by binding to FLC is lost, and they fail to inhibit FLC. Notably, In/Del occurred in intron1 of CAB3 (BoFLC2 and 3) and CAB5 (BoFLC2 and 3). Similarly, the possibility of the loss of COOLAIR and COLDAIR binding sites, the antisense of FLC, can be estimated. Variations in the insertion/deletion (In/Del) ratio in FLC introns, including other genes, may affect flowering. To compare the expression levels of FLC, experiments were conducted in a vernalized environment. To determine whether there were differences in the expression of FLC homologous genes, RNA was extracted. It was confirmed that the BoFLC1 gene is sensitive to low temperatures after exposure to low temperatures for approximately 7 weeks in the late flowering type[15]. Similarly, the expression of BoFLC1 began to decrease after 7 weeks in cold-treated CAB3 and 5 (Figure 3A). Expression values remained constant from weeks 9 to 11. In contrast, the expression of BoFLC1~3 in the control group remained constant in both CAB3 and 5 from weeks 5 to 11. The pattern of decreased expression after low-temperature treatment was similar. In the case of BoFLC2, the expression levels of CAB3 and 5 treated at low temperatures started to decrease after 7 weeks, and the expression remained constant from 9 to 11 weeks. BoFLC3 showed a trend similar to that of these two genes. Despite the distinct phenotype of the difference in flowering time between CAB3 and CAB5, the expression trends of BoFLC1, BoFLC2, and BoFLC3 between the two cabbages were similar during low-temperature treatment. This may be because the CDS sequences of the BoFLC1~3 genes between the two cabbages were similar, as were the mutations in the introns. In particular, because the expression values of the BoFLC1~3 genes were lowered in response to low temperatures, this suggests that the three genes may be related to the traits controlling flowering time. If a difference ensued between the two cabbages, the expression of the gene of BoFLC1~3 was approximately 1.5 to 2 times higher in CAB5 than in CAB3. This is considered as the amount of expression of the FLC homologous gene required to inhibit flowering, which varies among species. Furthermore, the expression values of the CAB5 cold-treated sample in BoFLC1 and the CAB3 control sample in week 11 were similar. This means that CAB5 needs to be cold-treated for a longer period. To confirm the FLC homologous similarity between Brassica oleracea species, the CDS of three homologous genes (BoFLC1, BoFLC2, and BoFLC3) of cabbage, kohlrabi, and broccoli with different flowering types were compared. When compared based on the BoFLC1 sequence of CAB1, the similarity was approximately ≥93% (Table S3). When comparing the sequences of different species based on CAB1, the BoFLC2 gene was also approximately 95% similar, and the BoFLC3 gene was approximately ≥94%. We compared the expression of BoGI, COOLAIR, and BoVIN3 to determine the relationship between expression differences and BoFLC according to the vernalization of genes related to flowering (Figure 4). This is because the expression patterns of the BoFLC homologous genes are similar. A difference existed in BoGI between the cold-treated and control groups. In CAB3, the peak appeared approximately two weeks earlier in the cold-treated group than in the control group. The same was true for CAB5. Therefore, BoGI is related to low temperatures because the moment when the low temperature of BoGI increased, the expression was accelerated (Figure 4A). When comparing the CAB3 and CAB5 controls, CAB3 peaked one week earlier, and CAB5 rapidly peaked at one week as well in the low-temperature treatment group. This suggests the possibility that flowering time may also be different because the expression time of BoGI differs between species, even at the same light time or temperature. Notably, the expression of COOLAIR, which binds to BoFLC and inhibits its expression, increased when the expression of BoFLC decreased under cold treatment (Figure 4B). The expression of COOLAIR increased rapidly at 7 weeks when BoFLC homologous expression began to decrease in cold-treated cabbages. The expression level, which peaked at week 7, began to decrease sharply after that and remained constant. This indicates that COOLAIR is related to BoFLC.
We analyzed the structural characteristics of FLC homologs between Brassica varieties at different flowering times using allele-specific polymerase chain reaction (AS-PCR) amplification and cloning. Mutations present in each sequence occur because of a cis polymorphism in the BoFLC homolog, suggesting that this may be important for FLC transcription and may contribute to flowering time variations between early- and late-flowering varieties. In particular, it has been suggested that the previously known BoFLC1, as well as BoFLC2 and BoFLC3 genes, may have roles and functions related to flowering. In future studies, it will be helpful to develop cabbage varieties that control flowering through target genome editing techniques, such as CRISPR-CAS9 for the BoFLC allele identified in this study, as well as Brassicaceae varieties. In addition, markers for selecting varieties with different flowering times could be developed.
4. Materials and Methods
4.1. Plant materials and growth conditions
In this study, we used genetically fixed cabbage genotypes to obtain uniform nucleotide sequences. Three inbred lines or commercial cultivars such as 20FL-CAB1 (early-flowering with a time of 140–150 days), 20FL-CAB3 (mid-flowering with time of 160–170 days), and 20FL-CAB5 (late-flowering with a time of ≥ 190 days) with distinct flowering times (varying by 40–45 days) were derived from a domestic seed company (Joeun Seeds Co., LTD, Goesan-gun, South Korea) in the Republic of Korea. Seeds of all the lines were sown and cultivated in a greenhouse (25 and 20 ºC under 16 and 8 h light and dark conditions, respectively) for 30 days in April 2020 and then transplanted to the field. Sixty days after field transplantation, the head portions of the cabbages were removed and transferred to plastic pots (30 × 25 cm) containing a mixture of a 1:1 ratio of cocopeat and soil. A set of pots without cold treatment (control) was allowed to grow under greenhouse conditions with 16 h of light at 25 °C and 8 h of dark at 18 °C. For the chilling or vernalization treatment, pots were transferred to incubators (TOGA clean system; model: TOGA UGSR01, Daejong, Korea) maintained at 4 °C with 10 h/14 h (light/dark) for 0–11 weeks. Leaf samples were collected from young leaves of plants grown under both the control and cold treatments each week (0–11 weeks of vernalization). The collected leaf samples were quickly frozen in liquid nitrogen and stored at − 80 °C for further use.
4.2. Genomic DNA extraction, PCR amplification, and sequencing of BoFLC homologs
Genomic DNA was extracted from young leaves using a WizPrep Plant DNA Mini Kit (WizBiosolutions, Seongnam, South Korea). According to the gene sequences of BoFLC1 (AM231517.1), BoFLC2 (AY306124.1), and BoFLC3 (AY306125.1) deposited in GenBank, allele-specific (AS) PCR primers were designed and used to amplify BoFLC- encoding genes in cabbage. The primer pairs used in this study are listed in Table S2. PCR amplifications were conducted in a 50 μL reaction volume containing 100 ng DNA, 0.5 μM of both forward and reverse oligos, 0.5 μM of dNTP, 10x Ex Taq buffer, 0.8 units of Ex Taq polymerase (Takara) and autoclaved water. PCR reactions were performed on a thermal cycler using the following program: one cycle of 30 s at 98 °C; 30 cycles of 30 s at 98 °C, 30 s at 60 °C, 3 min 30 s at 72 °C; one cycle of 15 min at 72 °C. The PCR products were resolved on a 1% agarose gel in 1x TBE buffer, then stained with ethidium bromide, and visualized under a UV transilluminator. The expected amplicons were excised from the gel and purified using a LaboPassTM Gel Extraction Kit (Cosmo Genetech, South Korea) as recommended by the manufacturer. To sequence the purified amplicons, 2000 bp contig primers flanking the entire gene were designed for each BoFLC locus using the Primer3Plus online tool. PCR amplicons were sequenced using an ABI3730XL sequencer (Macrogen Co., Seoul, South Korea).
4.3. Sequence analyses and Phylogenetic analysis of BoFLC homologs
The BoFLC1 genomic DNA sequences obtained in this study were deposited in the National Agricultural Biotechnology Information Center (NABIC; https://nabic.rda.go.kr/) database under the following numbers: NU-1420-000001 (BoFLC1 from CAB1) and NU-1425-000001 (BoFLC1 from CAB5). The sequencing results of nucleotide contigs were assembled, and comparative analysis among other BoFLC homologs to identify sequence variations (insertions and deletions) was conducted using BioEdit (version 7.2.5) sequence alignment software (Tom Hall, Ibis Biosciences, Carlsbad, CA, USA). The intron-exon organization of BoFLC homologs was predicted using the Gene Structure Display Server (GSDS2.0; http://gsds.cbi.pku.edu.cn/) by aligning the genomic sequences with their respective CDS sequences. Multiple sequence alignments of the deduced amino acid sequences of the identified BoFLC homologs were performed using CLUSTAL multiple sequence alignment in MUSCLE with the default parameters (http://www.ebi.ac.uk/Tools/msa/muscle/, version 3.8). To analyze the evolutionary relationships among FLC homologs from different Brassica species, a maximum likelihood tree and neighbor-joining tree (1000 replicates) were constructed using a dataset of FLC genes containing 31 deduced amino acid sequences, including those from cabbage in this study and those from other Brassica species (B. rapa, B. nigra, B. oleracea and B. napus) reported in previous studies[15,35]. The translated sequences of the whole coding regions of FLC were aligned using the MUSCLE method, and a tree was constructed using the neighbor-joining (NJ) method with MEGA version 10.0.5 and bootstrap analysis of 1000 replicates.
4.4. Extraction of mRNA and Expression profiling of Flowering genes
Total RNA from the leaf samples of both the control and treated plants was extracted using a GeneAll Hybrid RNA purification kit (GeneAll Biotechnology, Daejeon, Korea), as recommended by the manufacturer. cDNA was synthesized from 1.5 μg of RNA using the PrimeSrcript® RT reagent with a gDNA eraser kit (Takara Korea Biomedical, Seoul, Republic of Korea) according to the manufacturer’s instructions. Primers specific for BoFLC and Actin genes were designed using the Primer3Plus online tool and were used for qRT-PCR analysis (Table 1). A total of 13 μL of reaction mixture containing 2 μL of cDNA template, 6 μL 2x SYBR green Q master LaboPassTM (COSMOgenetech, South Korea), and 0.5 μM of gene-specific forward and reverse oligonucleotide primers. To estimate the relative mRNA expression of levels of transcripts, qRT-PCR reactions were performed on a CFX96 Real-Time PCR Detection System (Bio-Rad) with the following parameters: one cycle for initial denaturation at 95 °C for 3 min, 40 cycles of 95 °C for 15 s, 60 °C for 20 s, 72 °C for 15 s, a cycle at 65 °C for 5 s, and a final cycle of 95 °C for 2 s to detect primer specificity based on melt curve analysis. The relative expression levels of target gene transcripts were estimated using the 2-ΔΔCt method [36] from three independent replicates.
4.5. Statistical Analysis
To analyze statistically significant differences in the relative expression of genes, ANOVA with Duncan’s multiple range test was performed using SPSS version 21.0.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org and National Agricultural Biotechnology Information Center (NABIC; https://nabic.rda.go.kr/). Table S1: List of cabbage, kohlrabi, and broccoli materials used in this study with their flowering time. Table S2: List of primers used in this study. Table S3: Percent identity matrix of CDS multiple sequence alignment cabbage (FLC-CA1, -CA3, and -CA5), kohlrabi (FLC-KH7 and -KH8), and broccoli (FLC-BR10 and –BR11) (shown as in Table S1) having different flowering time. Figure S1: Comparisons of BoFLC1 (A), BoFLC2 (B), and BoFLC3 (C) gene structure, respectively, between reference (AM231617.1) and early- (CAB1), Mid- (CAB3) and late- (CAB5) flowering lines.
Author Contributions
Conceptualization, G. -J.L.; data curation, J.Y.A., S.S. F.Y. and J.Y.; formal analysis, J.Y.A. and S.S.; funding acquisition, G.-J.L.; investigation, J.Y.A., S.S. F.Y. and J.Y.; methodology, K.G.A. and G.-J.L.; project administration, G.-J.L.; resources, K.G.A. and G.-J.L.; software, J.Y.A. and S.S.; visualization, J.Y.A.; writing—original draft, J.Y.A., S.S. and C.A.; writing—review and editing, K.G.A. and G.-J.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the New Breeding Technologies Development Program (No. PJ01485802), Rural Development Administration and by the project of Development of New Varieties Breeding Technology with AI for Strengthening Food Sovereignty (No.RS-2023-00227464), Institute of Information & Communications Technology Planning & Evaluation (IITP), Ministry of Science and ICT (MSIT), Republic of Korea.
Data Availability Statement
Data is contained within the article or supplementary material.
Conflicts of Interest
All authors declare that they have no conflict of interest.
References
- Andrés, F.; Coupland, G. The Genetic Basis of Flowering Responses to Seasonal Cues. Nat Rev Genet 2012, 13, 627–639.
- Srikanth, A.; Schmid, M. Regulation of Flowering Time: All Roads Lead to Rome. Cellular and Molecular Life Sciences 2011, 68, 2013–2037.
- Fornara, F.; de Montaigu, A.; Coupland, G. SnapShot: Control of Flowering in Arabidopsis. Cell 2010, 141. [CrossRef]
- Ausín, I.; Alonso-Blanco, C.; Martínez-Zapater, J.M. Environmental Regulation of Flowering. International Journal of Developmental Biology 2005, 49, 689–705.
- Motoki, K.; Kinoshita, Y.; Hosokawa, M. Non-Vernalization Flowering and Seed Set of Cabbage Induced by Grafting onto Radish Rootstocks. Front Plant Sci 2019, 9. [CrossRef]
- Mao, Y.; Wu, F.; Yu, X.; Bai, J.; Zhong, W.; He, Y. MicroRNA319a-Targeted Brassica Rapa Ssp. Pekinensis TCP Genes Modulate Head Shape in Chinese Cabbage by Differential Cell Division Arrest in Leaf Regions. Plant Physiol 2014, 164, 710–720. [CrossRef]
- Kitamoto, N.; Nishikawa, K.; Tanimura, Y.; Urushibara, S.; Matsuura, T.; Yokoi, S.; Takahata, Y.; Yui, S. Development of Late-Bolting F1 Hybrids of Chinese Cabbage (Brassica Rapa L.) Allowing Early Spring Cultivation without Heating. Euphytica 2017, 213, 1–13. [CrossRef]
- Dittmar, E.L.; Oakley, C.G.; Ågren, J.; Schemske, D.W. Flowering Time QTL in Natural Populations of Arabidopsis Thaliana and Implications for Their Adaptive Value. Mol Ecol 2014, 23, 4291–4303. [CrossRef]
- Gazzani, S.; Gendall, A.R.; Lister, C.; Dean, C. Analysis of the Molecular Basis of Flowering Time Variation in Arabidopsis Accessions. Plant Physiol 2003, 132, 1107–1114. [CrossRef]
- Okazaki, K.; Sakamoto, K.; Kikuchi, R.; Saito, A.; Togashi, E.; Kuginuki, Y.; Matsumoto, S.; Hirai, M. Mapping and Characterization of FLC Homologs and QTL Analysis of Flowering Time in Brassica Oleracea. Theoretical and Applied Genetics 2007, 114, 595–608. [CrossRef]
- Osborn, T.C.; Kole, C.; Pa~-Kin, I.A.P.; Sharpe, ~ A G; Kuiper, M.; Lydiatet, D.J.; Trickt, M. Comparison of Flowering Time Genes in Brassica Rapa, B. Mpus and Arabidopsis Thulium. Genetics 1997, 146, 1129–1129.
- Camargo, L.E.A.; Osborn, T.C. Mapping Loci Controlling Flowering Time in Brassica Oleracea. Theor Appl Genet 1996, 92, 610–616.
- Ridge, S.; Brown, P.H.; Hecht, V.; Driessen, R.G.; Weller, J.L. The Role of BoFLC2 in Cauliflower (Brassica Oleracea Var. Botrytis L.) Reproductive Development. J Exp Bot 2015, 66, 125–135. [CrossRef]
- Razi, H.; Howell, E.C.; Newbury, H.J.; Kearsey, M.J. Does Sequence Polymorphism of FLC Paralogues Underlie Flowering Time QTL in Brassica Oleracea? Theoretical and Applied Genetics 2008, 116, 179–192. [CrossRef]
- Abuyusuf, M.; Nath, U.K.; Kim, H.T.; Islam, M.R.; Park, J.I.; Nou, I.S. Molecular Markers Based on Sequence Variation in BoFLC1.C9 for Characterizing Early- and Late-Flowering Cabbage Genotypes. BMC Genet 2019, 20. [CrossRef]
- Wu, J.; Wei, K.; Cheng, F.; Li, S.; Wang, Q.; Zhao, J.; Bonnema, G.; Wang, X. A Naturally Occurring InDel Variation in BraA.FLC.b (BrFLC2) Associated with Flowering Time Variation in Brassica Rapa. BMC Plant Biol 2012, 12. [CrossRef]
- Zhao, J.; Kulkarni, V.; Liu, N.; Pino Del Carpio, D.; Bucher, J.; Bonnema, G. BrFLC2 (FLOWERING LOCUS C) as a Candidate Gene for a Vernalization Response QTL in Brassica Rapa. J Exp Bot 2010, 61, 1817–1825. [CrossRef]
- Hou, J.; Long, Y.; Raman, H.; Zou, X.; Wang, J.; Dai, S.; Xiao, Q.; Li, C.; Fan, L.; Liu, B.; et al. A Tourist-like MITE Insertion in the Upstream Region of the BnFLC.A10 Gene Is Associated with Vernalization Requirement in Rapeseed (Brassica Napus L.). BMC Plant Biol 2012, 12. [CrossRef]
- Tadege, M.; Sheldon, C.C.; Helliwell, C.A.; Stoutjesdijk, P.; Dennis, E.S.; Peacock, W.J. Control of Flowering Time by FLC Orthologues in Brassica Napus. Plant Journal 2001, 28, 545–553. [CrossRef]
- Schranz, M.E.; Quijada, P.; Sung, S.-B.; Lukens, L.; Amasino, R.; Osborn, T.C. Characterization and Effects of the Replicated Flowering Time Gene FLC in Brassica Rapa. Genetics 2002, 162, 1457–1468.
- Itabashi, E.; Shea, D.J.; Fukino, N.; Fujimoto, R.; Okazaki, K.; Kakizaki, T.; Ohara, T. Comparison of Cold Responses for Orthologs of Cabbage Vernalizationrelated Genes. Horticulture Journal 2019, 88, 462–470. [CrossRef]
- Irwin, J.A.; Lister, C.; Soumpourou, E.; Zhang, Y.; Howell, E.C.; Teakle, G.; Dean, C. Functional Alleles of the Flowering Time Regulator FRIGIDA in the Brassica Oleracea Genome. BMC Plant Biol 2012, 12. [CrossRef]
- Fadina, O.A.; Pankin, A.A.; Khavkin, E.E. Molecular Characterization of the Flowering Time Gene FRIGIDA in Brassica Genomes A and C. Russian Journal of Plant Physiology 2013, 60, 279–289. [CrossRef]
- Wang, N.; Qian, W.; Suppanz, I.; Wei, L.; Mao, B.; Long, Y.; Meng, J.; Müller, A.E.; Jung, C. Flowering Time Variation in Oilseed Rape (Brassica Napus L.) Is Associated with Allelic Variation in the FRIGIDA Homologue BnaA.FRI.a. J Exp Bot 2011, 62, 5641–5658. [CrossRef]
- Sheldon, C.C.; Rouse, D.T.; Finnegan, E.J.; Peacock, W.J.; Dennis, E.S. The Molecular Basis of Vernalization: The Central Role of FLOWERING LOCUS C (FLC). PNAS 2000, 97, 3753–3758.
- Helliwell, C.A.; Wood, C.C.; Robertson, M.; James Peacock, W.; Dennis, E.S. The Arabidopsis FLC Protein Interacts Directly in Vivo with SOC1 and FT Chromatin and Is Part of a High-Molecular-Weight Protein Complex. Plant Journal 2006, 46, 183–192. [CrossRef]
- Nilsson, O.; Lee, I.; Blázquez, M.A.; Weigel, D. Flowering-Time Genes Modulate the Response to LEAFY Activity. Genetics 1998, 150, 403–410.
- Blázquez, M.A.; Green, R.; Nilsson, O.; Sussman, M.R.; Weigel, D. Gibberellins Promote Flowering of Arabidopsis by Activating the LEAFY Promoter. Plant Cell 1998; 10,791-800.
- Mizoguchi, T.; Wright, L.; Fujiwara, S.; Cremer, F.; Lee, K.; Onouchi, H.; Mouradov, A.; Fowler, S.; Kamada, H.; Putterill, J.; et al. Distinct Roles of GIGANTEA in Promoting Flowering and Regulating Circadian Rhythms in Arabidopsis. Plant Cell 2005, 17, 2255–2270. [CrossRef]
- Kardailsky, I.; Shukla, V.K.; Ahn, J.H.; Dagenais, N.; Christensen, S.K.; Nguyen, J.T.; Chory, J.; Harrison, M.J.; Weigel, D. Activation Tagging of the Floral Inducer FT. Science 1999, 286, 1962–1965.
- Jung, J.H.; Seo, Y.H.; Pil, J.S.; Reyes, J.L.; Yun, J.; Chua, N.H.; Park, C.M. The GIGANTEA-Regulated MicroRNA172 Mediates Photoperiodic Flowering Independent of CONSTANS in Arabidopsis. Plant Cell 2007, 19, 2736–2748. [CrossRef]
- Zhang, J.; Mujahid, H.; Hou, Y.; Nallamilli, B.R.; Peng, Z. Plant Long NcRNAs: A New Frontier for Gene Regulatory Control. Am J Plant Sci 2013, 04, 1038–1045. [CrossRef]
- Swiezewski, S.; Liu, F.; Magusin, A.; Dean, C. Cold-Induced Silencing by Long Antisense Transcripts of an Arabidopsis Polycomb Target. Nature 2009, 462, 799–802. [CrossRef]
- Heo, J.B.; Sung, S. Vernalization-Mediated Epigenetic Silencing by a Long Intronic Noncoding RNA. Science 2011, 331, 76–79. [CrossRef]
- Schiessl, S. V.; Quezada-Martinez, D.; Tebartz, E.; Snowdon, R.J.; Qian, L. The Vernalisation Regulator FLOWERING LOCUS C Is Differentially Expressed in Biennial and Annual Brassica Napus. Sci Rep 2019, 9. [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Tim e Quantitative PCR and the 2 C T Method. METHODS 2001, 2, 402–408.
- Paltiel, J.; Amin, R.; Gover, A.; Ori, N.; Samach, A. Novel Roles for GIGANTEA Revealed under Environmental Conditions That Modify Its Expression in Arabidopsis and Medicago Truncatula. Planta 2006, 224, 1255–1268. [CrossRef]
Figure 1.
Differences in gene structure of three homologs of BoFLC1, BoFLC2, and BoFLC3 between early- (CAB 1) and late- (CAB 5) flowering cabbages. The light-colored regions of the homologs indicate coding sequences and introns, respectively, with upstream and downstream regions in blue boxes. The deleted sequences of BoFLC1 and BoFLC2 (CAB 5) were noticed only in the late-flowering genotype (CAB 5).
Figure 1.
Differences in gene structure of three homologs of BoFLC1, BoFLC2, and BoFLC3 between early- (CAB 1) and late- (CAB 5) flowering cabbages. The light-colored regions of the homologs indicate coding sequences and introns, respectively, with upstream and downstream regions in blue boxes. The deleted sequences of BoFLC1 and BoFLC2 (CAB 5) were noticed only in the late-flowering genotype (CAB 5).
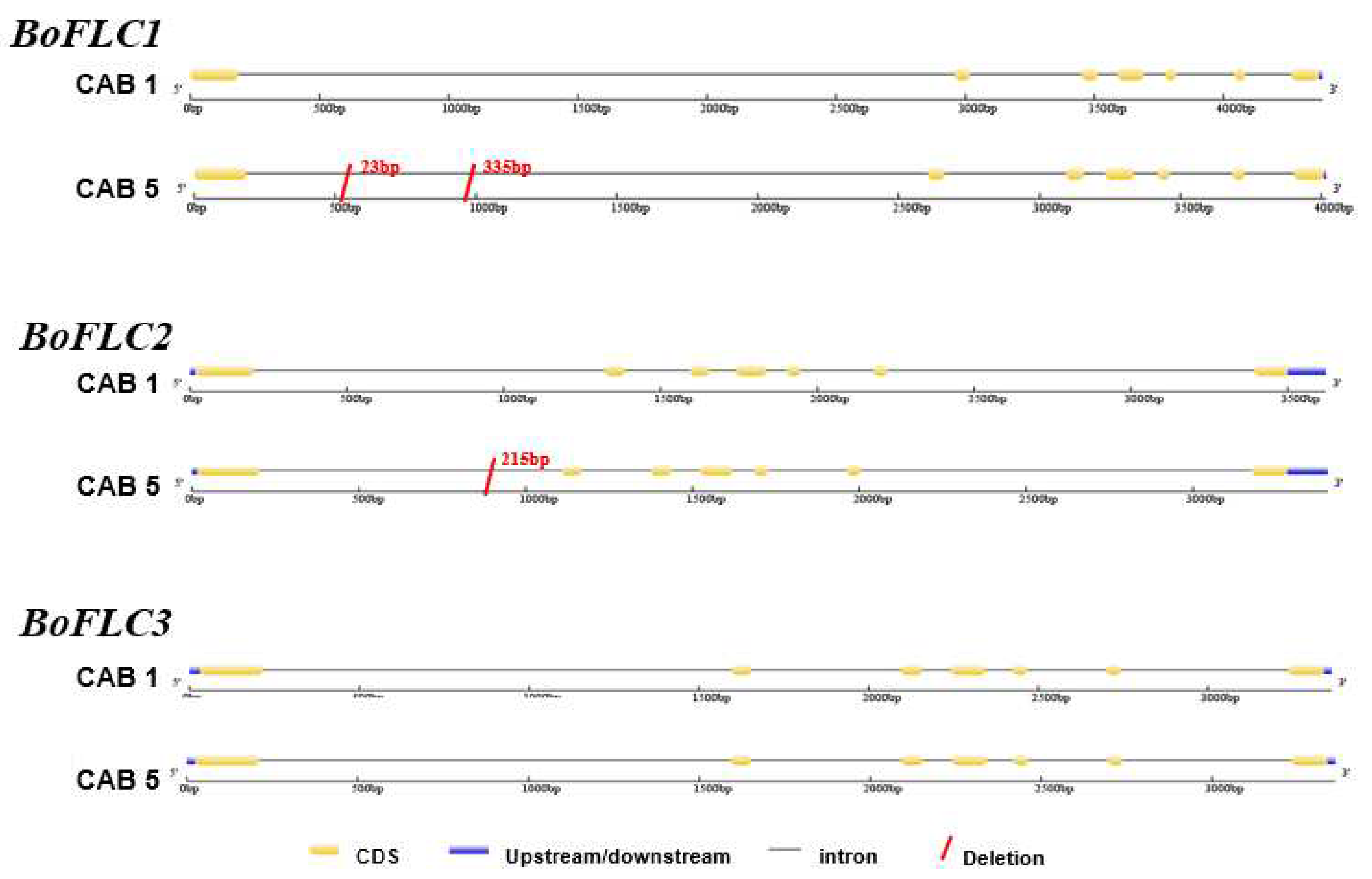
Figure 2.
Neighbor-joining tree showing phylogenetic relationships based on the decoded BoFLC amino acid sequences among early- (CAB 1), mid- (CAB 3), and late- (CAB 5) flowering cabbages. The GenBank accession number is shown in a parenthesis, and sequences were compared with three cabbage genotypes used in our study. Bootstrap values ˃ 50% are shown above the branches. Bc, Brassica carinata; Bj, B. juncea; Bn, B. napus; Bni, B. nigra; Bo, B. oleracea; Br, Brassica rapa.
Figure 2.
Neighbor-joining tree showing phylogenetic relationships based on the decoded BoFLC amino acid sequences among early- (CAB 1), mid- (CAB 3), and late- (CAB 5) flowering cabbages. The GenBank accession number is shown in a parenthesis, and sequences were compared with three cabbage genotypes used in our study. Bootstrap values ˃ 50% are shown above the branches. Bc, Brassica carinata; Bj, B. juncea; Bn, B. napus; Bni, B. nigra; Bo, B. oleracea; Br, Brassica rapa.
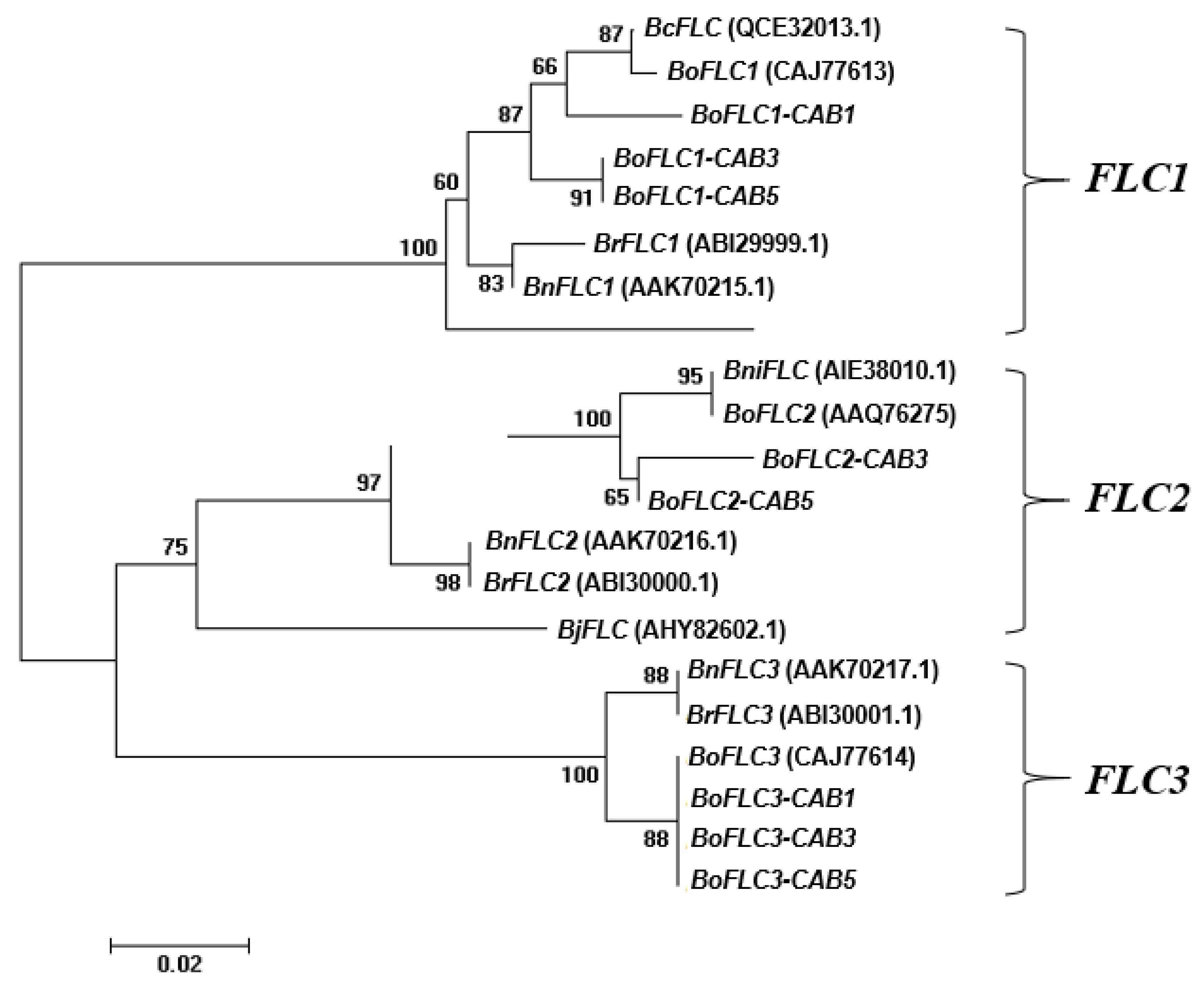
Figure 3.
mRNA expression patterns of three BoFLC homologs using cDNA from chilling (dotted line) and no-chilling (solid line) treatment between mid- (CAB 3) and late- (CAB 5) flowering cabbages. Relative expressions of BoFLC1 (top panels A and B), BoFLC2 (middle panels C and D), and BoFLC3 (bottom panels E and F) are shown for the mid-flowering CAB 3 (left panels A, C, and E, respectively) and late-flowering CAB 5 (right panels B, D, and F, respectively). Different letters indicate statistically significant differences in gene expression across chilling periods according to Duncan's multiple range test at p ≤ 0.05.
Figure 3.
mRNA expression patterns of three BoFLC homologs using cDNA from chilling (dotted line) and no-chilling (solid line) treatment between mid- (CAB 3) and late- (CAB 5) flowering cabbages. Relative expressions of BoFLC1 (top panels A and B), BoFLC2 (middle panels C and D), and BoFLC3 (bottom panels E and F) are shown for the mid-flowering CAB 3 (left panels A, C, and E, respectively) and late-flowering CAB 5 (right panels B, D, and F, respectively). Different letters indicate statistically significant differences in gene expression across chilling periods according to Duncan's multiple range test at p ≤ 0.05.
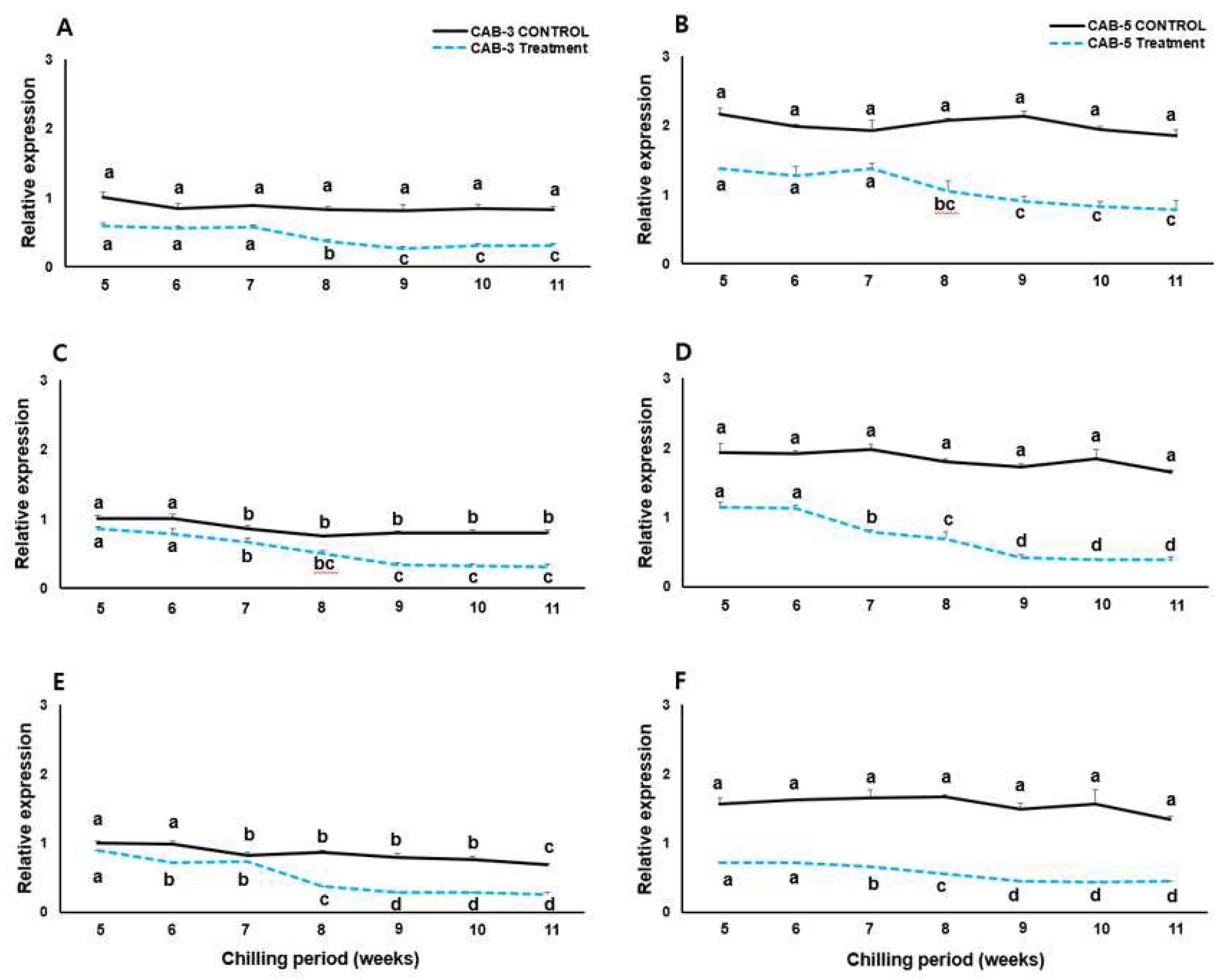
Figure 4.
mRNA expression patterns of the flowering-time regulator genes GIGANTEA (GI, top panel A and B), BoCOOLAIR (middle panel C and D), BoVIN3 (bottom panel E and F) between mid- (CAB 3) and late- (CAB 5) flowering cabbage lines under chilling (dotted line) and no-chilling treatment. The relative expressions of GI, BoCOOLAIR, and BoVIN3 were compared for the mid-flowering CAB-3 line (left A, C, E) and late-flowering CAB-5 (right B, D, F) line. The bars indicate standard errors of the means for each chilling week.
Figure 4.
mRNA expression patterns of the flowering-time regulator genes GIGANTEA (GI, top panel A and B), BoCOOLAIR (middle panel C and D), BoVIN3 (bottom panel E and F) between mid- (CAB 3) and late- (CAB 5) flowering cabbage lines under chilling (dotted line) and no-chilling treatment. The relative expressions of GI, BoCOOLAIR, and BoVIN3 were compared for the mid-flowering CAB-3 line (left A, C, E) and late-flowering CAB-5 (right B, D, F) line. The bars indicate standard errors of the means for each chilling week.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



